Влияние способов обработки почвы с возвратом соломы на микробные сообщества
Влияние различных способов обработки почвы в сочетании с возвратом соломы на активность почвенных ферментов и структуру микробного сообщества, а также его разнообразие.
Аннотация
В связи с проблемами серьезного опустынивания почв, усиления эрозии почв и воды и снижения доступных питательных веществ в почве в агропастбищной экотонной зоне на северо-востоке Внутренней Монголии, данное исследование было проведено на экспериментальном поле в Аронг-Баннере с использованием кукурузного сорта A6565 в качестве культуры и анализа семи различных методов обработки почвы: глубокая вспашка, глубокое рыхление, мелкая обработка, фрезерование, тяжелое боронование, нулевая обработка и контроль, в сочетании с возвратом соломы. Результаты анализа следующие: активность уреазы и содержание микробной биомассы азота при обработке почвы с возвратом соломы были выше, чем при других методах. По сравнению с фазой всходов, альфа-индекс разнообразия бактерий увеличился на стадии уборки урожая, в то время как у грибов он снизился. Сравнение бета-разнообразия показало, что время отбора проб является основным фактором, влияющим на бактериальное сообщество и его состав. Было установлено, что доминирующими бактериями являются протеобактерии и актинобактерии, а доминирующим грибом — аскомицеты. Почвозащитная обработка в сочетании с возвратом соломы оказывает положительное влияние на разнообразие почвенных микроорганизмов, что более способствует улучшению доступных питательных веществ в почве и ее качества. Все
1. Введение
В последнее время из-за применения традиционных однообразных методов обработки почвы, интенсивного использования крупногабаритной техники и некоторых нерациональных практик земледелия моделирование эрозии почв постепенно усугубляется и, как следствие, приводит к снижению продуктивности почв в агропастбищных зонах [1,2]. Низкая продуктивность сельскохозяйственных культур, вызванная деградацией пахотных земель, оказывает огромное влияние на урожайность и устойчивость сельскохозяйственного развития [3]. Это стало одной из наиболее важных современных проблем, требующих решения, и необходимо найти наилучший метод обработки почвы для реализации устойчивого управления в агропастбищных зонах Внутренней Монголии. Исследования показывают, что почвозащитная обработка почвы, особенно метод возврата соломы, может улучшить плодородие почвы, содержание питательных веществ и активность ферментов за счет снижения воздействия техники на почву, поэтому она представляется весьма необходимой для повышения продуктивности земель, увеличения урожайности и устойчивого развития регионального сельского хозяйства [4,5,6].
В почвенной среде основными источниками ферментов являются остатки животных, растений, микроорганизмов и их секреции в процессе метаболизма [7]. Они могут катализировать все почвенные биохимические процессы, включая синтез и разложение гумуса, разложение органического вещества и остатков животных и растений, а также фиксацию и высвобождение почвенных питательных веществ [8]. Как правило, инвертаза почвы отражает обеспеченность почвы углеродом. В случае лучшего плодородия почвы и более богатого органического вещества активность инвертазы также будет увеличиваться [9]. Уреаза почвы принадлежит к гидролазам, которая может способствовать гидролизу мочевины в почве и поглощению азота сельскохозяйственными культурами [10]. Следовательно, её активность фактически указывает на способность почвы обеспечивать азотом. Кроме того, фосфатаза почвы может ускорять скорость дефосфорилирования органического фосфора в почве, что играет важную роль в цикле фосфора в почве, а её активность также может использоваться в качестве важного показателя для измерения плодородия почвы [11]. Также было показано, что активность ферментов тесно связана с изменениями содержания органического вещества и микробной популяции [12]. Интенсивность обработки при традиционной вспашке высока, что ускоряет минерализацию органических питательных веществ в агрегатах, влияя таким образом на накопление органического вещества в почве, что не способствует повышению активности ферментов [13]. Возврат соломы в поле не только приносит в почву обильные активные микроорганизмы, но и обеспечивает микроорганизмы экзогенными питательными веществами и энергией, тем самым значительно увеличивая численность микробных популяций [14]. Предыдущие исследования факторов, влияющих на активность почвенных ферментов, в основном были сосредоточены на физико-химических свойствах почвы, уровнях внесения азота, типах землепользования, других сельскохозяйственных методах и внешних факторах [15].
Обеспечивая микроорганизмы ресурсами питательных веществ, возврат соломы в поле может способствовать развитию водных микробов и увеличению их количества; таким образом, увеличивается количество бактерий и повышается почвенная биологическая активность [16,17]. Было выявлено, что микробная биомасса почвы состоит из углерода микробной биомассы, обеспечивающего энергией разложение органического вещества, и азота микробной биомассы, отражающего способность снабжения азотом. Его содержание может изменяться при использовании различных методов обработки почвы и анализе чувствительности к доступности почвенных питательных веществ и микроокружению, что является одним из показателей для оценки качества почвы [18].
Микроорганизмы в почве способствуют трансформации и круговороту почвенных питательных веществ, что может обеспечить устойчивое использование почвы и регулировать стабильность почвенного микроэкологического сообщества [19]. Различные почвенные микроорганизмы обладают специфическими функциональными и генетическими характеристиками, а другие структуры почвенного микробного сообщества формируют разнообразие почвенных микроорганизмов [20]. Исследования доказывают, что микробные сообщества не только реагируют на изменения микроокружения более прямо и надежно, чем такие параметры, как содержание органического вещества в агроэкосистемах, но и являются основным фактором изменения плодородия и продуктивности почвы [21]. Чем сложнее структура микробного сообщества почвы, тем стабильнее почвенная экосистема и тем сильнее буферный эффект на изменения внешней среды [22]. Поэтому изучение сообществ грибов и бактерий, а также исследование состава структуры микробного сообщества и обилия разнообразия сообществ могут отражать плодородие почвы и играть ключевую роль в защите качества пахотных земель [23,24]. Метод нулевой обработки почвы в основном relies on fungi to decompose органического вещества, в то время как в системе традиционной обработки почвы бактерии ответственны за разложение большей части органического вещества.
Было обнаружено, что традиционная обработка почвы нарушает почвенные слои и снижает микробное разнообразие, в то время как почвозащитная обработка улучшает микробное разнообразие почвы [25]. Нулевая обработка с мульчированием соломой увеличивала количество и разнообразие бактерий в поверхностном слое почвы. В то же время, глубокое рыхление и мульчирование соломой оказывали более значительное влияние на структуру сообщества почвенных грибов [26]. Возврат соломы увеличивал биомассу грибов в основном потому, что поступление экзогенного органического углерода улучшало среду обитания микроорганизмов, повышало экологическую буферную способность почвы и создавало благоприятные условия для размножения грибов [27]. С другой стороны, солома, внесенная на поверхность почвы, обеспечивает рост микроорганизмов в почве большим количеством питательных веществ и энергии через процессы окислительного разложения и гумификации, что увеличивает разнообразие и структуру сообщества почвенных микроорганизмов [28]. Некоторые исследования также предполагают, что вспашка увеличивает однородность верхнего слоя почвы и делает питательные вещества, необходимые для размножения микроорганизмов в почве, более равномерно распределенными [29]. В целом, традиционные методы обработки почвы нарушают почвенный слой и снижают почвенное микробное разнообразие, в то время как почвозащитная обработка способствует увеличению почвенного микробного разнообразия. В настоящее время исследования изменений состава и разнообразия почвенного микробного сообщества при возврате соломы с различными методами обработки почвы в основном сосредоточены на единственном сезонном изменении почвенного микробного сообщества.
Чтобы понять влияние различных методов почвозащитной обработки на биологию почвы и признаки урожайности в условиях возврата соломы в агропастбищной экотонной зоне вдоль хребта Большой Хинган во Внутренней Монголии, было установлено семь методов обработки почвы в различных условиях возврата соломы. Динамический закон изменения микробной биомассы и разнообразия, а также влияние на урожайность кукурузы на разных стадиях сравниваются и анализируются с помощью некоторых методов, включая высокопроизводительное секвенирование и фумигационное культивирование. Данное исследование направлено на отбор наиболее подходящих местных методов земледелия и предоставление теоретической основы для улучшения качества почвы и увеличения урожайности сельскохозяйственных культур в агропастбищной экотонной зоне вдоль хребта Большой Хинган во Внутренней Монголии.
2. Материалы и методы
2.1. Место проведения эксперимента
Место эксперимента расположено в Центре распространения сельскохозяйственных технологий Аронг-Баннера в городе Хулунбуир на северо-востоке Автономного района Внутренняя Монголия (47°56′54″–49°19′35″ с.ш., 122°02′30″–124°0′40″ в.д.).
Среднегодовая температура составляет 1,7 °C. Годовой эффективный accumulated temperature составляет 2394,1 °C. Годовое количество солнечных часов составляет 2750–2850 ч. Высота постепенно переходит от 1149 м до 198 м. Среднегодовое количество осадков составляет 458,4 мм, в основном с июня по август, а среднегодовое испарение составляет 1455,3 мм. Местно в середине и начале августа 2019 года прошли сильные дожди. Ранние заморозки произошли в большинстве районов в середине сентября, с безморозным периодом 90–130 дней.
Почвы в основном темно-коричневые и черноземы. Большинство основных культур — соя, кукуруза, картофель, рапс и свекла, система урожая — один урожай в год, в основном богарное земледелие. Основные химические свойства участка испытаний 0~60 см были: pH 8,50, органический углерод 3,95 г/кг, щелочно-гидролизуемый азот 30,73 мг/кг, доступный фосфор 2,21 мг/кг, доступный калий 4,37 мг/кг, общий азот 1,51 г/кг, общий фосфор 3,33 г/кг и общий калий 27,88 г/кг.
Сорт кукурузы A6565 был высеян в экспериментальной зоне в конце апреля 2019 года. Комплексное удобрение (N-P₂O₅-K₂O = 23-12-13) вносилось во время подготовки земли перед весенним посевом на экспериментальном участке, всего 600 кг/га.
2.2. Экспериментальные материалы
Сорт кукурузы A6565 в качестве экспериментальной культуры был предоставлен Зоной современного демонстрационного сельского хозяйства Аронг-Баннера. Материал удобрения, N-P-K (23-12-13), произведен компанией Arong Banner Xingnong Special Fertilizer Co., Ltd, Хух-Хото, Китай. Материалы препаратов включают фенолят натрия (Tianjin Jinbei Fine Chemical Co., Ltd., Тяньцзинь, Китай), гипохлорит натрия (Tianjin Jinbei Fine Chemical Co., Ltd., Тяньцзинь, Китай), перекись водорода (Tianjin Damao Chemical Reagent Factory, Тяньцзинь, Китай), перманганат калия (Tianjin Damao Chemical Reagent Factory, Тяньцзинь, Китай), 3-амино-5-нитрсалициловую кислоту (Guoyao Group Chemical Reagent Co., Ltd., Шанхай, Китай), фенилфосфат динатрия (Tianjin Zhiyuan Chemical Reagent Co., Ltd. Company, Тяньцзинь, Китай), Лин Фа Луолин (Tianjin Zhiyuan Chemical Reagent Co., Ltd., Тяньцзинь, Китай) и нингидрин (Guoyao Group Chemical Reagent Co., Ltd., Шанхай, Китай).
2.3. План эксперимента
Этот двухлетний эксперимент, с 2019 по 2020 год, включал посев 12 мая каждого года. Он включал семь участков, соответствующих семи обработкам почвы. В каждой делянке во время отбора проб было установлено три случайные точки отбора проб, а площадь каждой делянки составляла 750 м². Образцы почвы были собраны в период вегетации кукурузы в 2020 году для анализа.
Семь обработок: G1 (возврат соломы и глубокая вспашка), G2 (возврат соломы и глубокое рыхление с мелким оборотом пласта), G3 (возврат соломы и глубокое рыхление), G4 (возврат соломы и фрезерование), G5 (возврат соломы и боронование), G6 (возврат соломы и нулевая обработка), G7 (контроль: без возврата соломы и традиционная обработка); G1–G6 относятся к почвозащитной обработке, а G7 — к традиционной обработке. Кукурузная солома обрабатывалась непосредственно после уборки урожая предыдущего года, и солома возвращалась в поле в сочетании с различными методами обработки почвы. Количество соломы, возвращаемой в поле, составляло общее количество кукурузной соломы, возвращаемой в поле, и количество возврата составляло около 7000 кг/га. Возврат соломы в поле в сочетании с методом обработки почвы проводился после осеннего сбора урожая, а боронование проводилось перед посевом в следующем году. Подробные детали операций могут быть отражены в Таблице 1. В данном эксперименте применялась рандомизированная полная блочная схема.

Таблица 1. План эксперимента по заделке соломы в поле в сочетании с методом обработки почвы.
2.4. Измеряемые показатели и метод
На пяти стадиях (стадия всходов, стадия большого раструба, стадия вымётывания и силосования, стадия налива и послеуборочная стадия) образцы почвы отбирались почвенным буром (диаметр 2,5 см) на четырех различных глубинах: 0~10 см, 10~20 см, 20~40 см и 40~60 см соответственно. С каждого слоя почвы отбирались три случайные пробы для смешанного упаковывания. На поверхности пакета прикреплялась этикетка с указанием названия обработки, места отбора проб, времени и глубины. Собранные образцы почвы доставлялись в лабораторию, и часть из них естественным образом высушивалась в прохладном месте. После высушивания образцы почвы просеивались через сито 1 мм для определения активности почвенных ферментов. Другая часть сохранялась через сито 2 мм в низкотемпературном холодильнике при 4 °C, которая использовалась для определения микробной биомассы почвы.
Почва отбиралась маленькой лопаткой на стадии всходов, вымётывания, силосования и после уборки. Перед отбором почвы удалялись камни и растительные остатки на поверхности, и отбирались образцы почвы 0~20 см и пропускались через сито 2 мм. 5~10 г почвы после просеивания помещали в центрифужную пробирку на 1,5 мл (центрифужная пробирка была стерилизована перед отбором проб). Для каждой обработки отбирали три случайных образца почвы и просеивали после смешивания, и загружали соответственно три центрифужные пробирки. На центрифужной пробирке было написано название обработки, затем её помещали в жидкий азот для быстрой заморозки и загружали в пакет с клапаном. На поверхности пакета указывалось название обработки, место отбора проб, время и глубина. Собранные образцы почвы возвращались в лабораторию и хранились в ультранизкотемпературном холодильнике при -80 °C для экстракции микробной ДНК и высокопроизводительного секвенирования.
Активность уреазы почвы определялась индофеноловым синим колориметрическим методом, активность инвертазы почвы проверялась с помощью 3,5-динитросалициловой кислоты колориметрическим методом, активность каталазы почвы проверялась титрованием перманганатом калия, а активность щелочной фосфатазы почвы определялась колориметрическим методом с фенилфосфатом динатрия [30,31,32]. Углерод микробной биомассы почвы исследовался фумигационно-экстракционным объемным анализом, а азот микробной биомассы почвы определялся хлороформно-экстракционным нингидриновым колориметрическим методом [33].
ДНК из образцов почвы экстрагировалась стандартными наборами, а чистота и концентрация ДНК определялись электрофорезом в агарозном геле. Экспериментальный процесс был следующим: образец загружали в центрифужную пробирку и разбавляли до 1 нг/мкл. Используя разбавленную геномную ДНК в качестве матрицы, ПЦР проводили с использованием специфических праймеров и высокоточного фермента Takara EXTaq, и выбирали соответствующие области секвенирования. Продукты ПЦР детектировали электрофорезом и очищали магнитными шариками в качестве матрицы для ПЦР второго раунда для двухраундной ПЦР-амплификации. После амплификации вышеуказанные шаги повторяли, и конечный продукт количественно определяли с помощью Qubit7. Образцы смешивали в равных количествах в соответствии с концентрацией продукта ПЦР, а затем секвенировали онлайн. Впоследствии была использована библиотека платформы miseqpe300 (Illumina, Сан-Диего, Калифорния, США) для построения и секвенирования.
2.5. Анализ данных
Программное обеспечение Excel 2007 использовалось для обработки данных о почвенном микробном разнообразии, а программное обеспечение SPSS 21 (IBM, Армонк, Нью-Йорк, США) и Origin 2022 (OriginLab, Нортгемптон, Массачусетс, США) использовались для составления чертежей и обработки данных. Однофакторный дисперсионный анализ (ANOVA) проверял значимость различий между обработками, а метод Дункана использовался для множественных сравнений между парами.
Участки 16S и ITS секвенировались с помощью технологии высокопроизводительного секвенирования, включая экстракцию ДНК/РНК, ПЦР-амплификацию и секвенирование. Платформа Illumina miseq (тестовый набор v3-2x300 циклов, Illumina, Калифорния, США) использовалась для двустороннего секвенирования библиотеки амплификации генов 16s и ITS. Данные 16SrRNA и ITSrRNA анализировались с помощью программного обеспечения MOTHUR v.1.39.5 (Мичиганский университет, Анн-Арбор, Мичиган, США) в соответствии со стандартной операционной процедурой (SOP) MiSeq. Была получена кривая разрежения. Индексы разнообразия Шеннона и Симпсона и индекс богатства Чао были рассчитаны для измерения микробного разнообразия и обилия. Используя метод многомерного анализа на основе анализа главных компонент (PCA), сравнивали состав почвенных микроорганизмов при различных методах обработки почвы, различных периодах роста и различных слоях почвы, а также анализировали и сравнивали взаимосвязи между составом и разнообразием почвенных микроорганизмов при различных методах обработки почвы, различных периодах роста и различных слоях почвы.
Корреляционный анализ Пирсона проводился для четырех активностей ферментов, MBC и MBN, и индексов микробного разнообразия в слое почвы 0~20 см в периоды отбора проб в июне, августе и октябре, и графики были построены с помощью Origin 2022.
3. Результаты
3.1. Анализ активности почвенных ферментов
С ростом кукурузы тенденция активности каталазы и щелочной фосфатазы увеличивалась, а затем уменьшалась, в то время как активность уреазы и инвертазы увеличивалась, затем уменьшалась, а затем снова повышалась; с увеличением глубины почвы активность ферментов каждой обработки постепенно снижалась. Активность ферментов на стадии уборки урожая была выше, чем на стадии всходов (Рисунок 1).
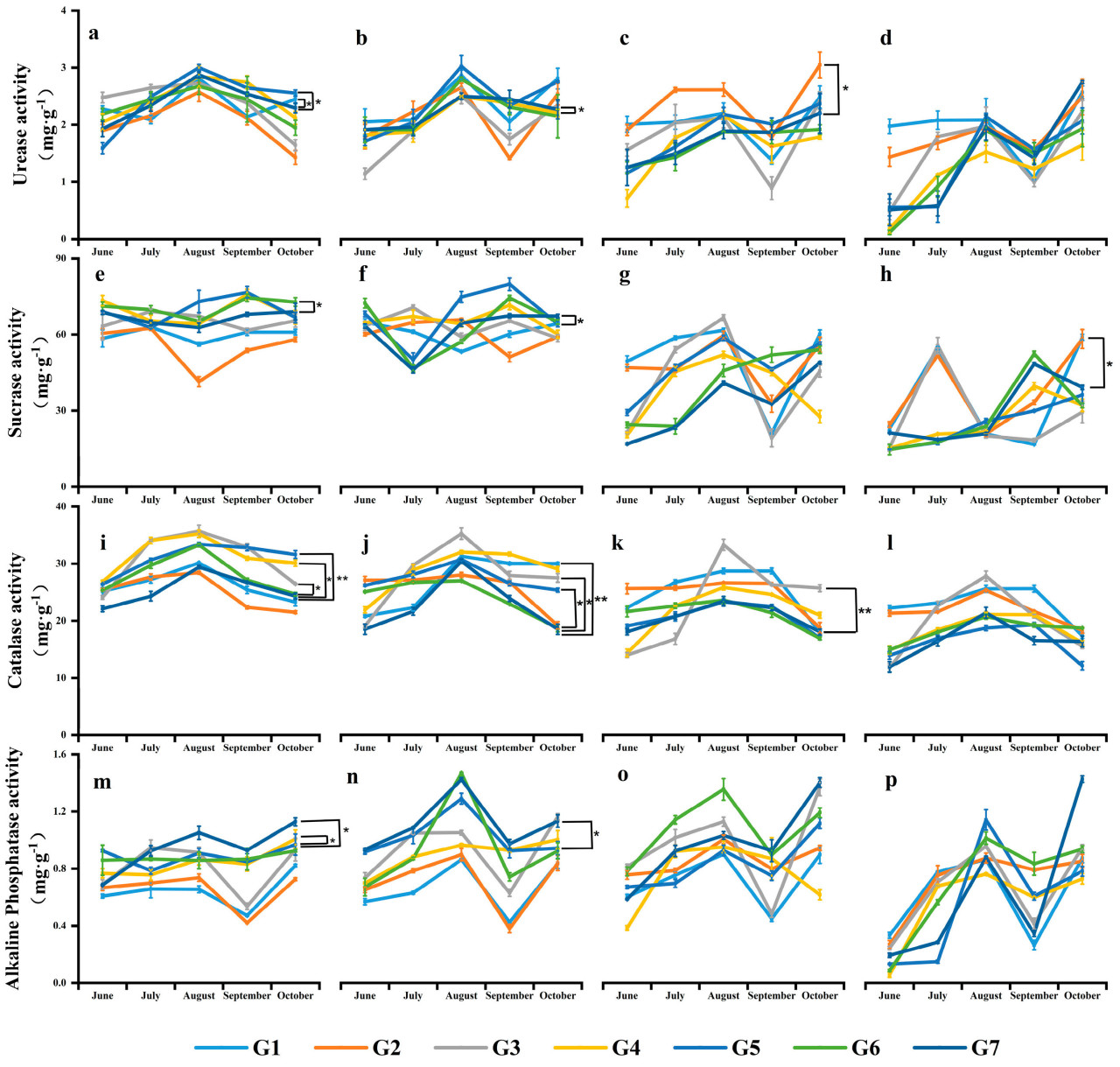
Рисунок 1. Влияние различных методов обработки почвы на активность уреазы почвы (мг·г⁻¹), активность инвертазы (мг·г⁻¹), активность каталазы (мг·г⁻¹) и активность щелочной фосфатазы (мг·г⁻¹). Панели (a–d) представляют активность уреазы в слоях почвы 0~10 см, 10~20 см, 20~40 см и 40~60 см соответственно; (e–h) — активность инвертазы; (i–l) — активность каталазы; (m–p) — активность щелочной фосфатазы. Июнь: стадия всходов, Июль: стадия большого раструба, Август: стадия вымётывания и силосования, Сентябрь: стадия налива, Октябрь: после уборки. По сравнению с традиционной обработкой (G7), *: p < 0,05, : p < 0,01. Столбцы ошибок построены с SE.
После периода уборки урожая 2020 года, в слое почвы 0~10 см, активность уреазы G1 и G5 была самой высокой, соответственно на 6,32% и 10,21%, значительно выше, чем у G7 (Рисунок 1a). Активность инвертазы G6 была на 5,19% значительно выше, чем у G7 (Рисунок 1e). В то время как активность каталазы G1 и G2 была ниже, чем у G7, другие обработки были на 1,39%~23,08% значительно выше, чем G7 (Рисунок 1i). За исключением того, что активность щелочной фосфатазы G1 и G2 была ниже, чем у G7, другие обработки были значительно выше, чем G7, на 0,93%~18,07% (Рисунок 1m). В слое почвы 10~20 см, за исключением того, что активность уреазы G4 и G6 была ниже, чем у G7, другие обработки были значительно выше, чем G7, на 3,13%~19,27% (Рисунок 1b). Активность инвертазы G1, G5 и G6 была выше на 0,28%, 4,72% и 1,34% соответственно (Рисунок 1f). Активность каталазы каждой обработки была на 1,04%~38,26% выше, чем у G7 (Рисунок 1j). Активность щелочной фосфатазы G1 и G2 также была ниже, чем у G7, но другие обработки были на 1,95%~20,56% выше, чем G7. G1 был значительно выше, чем G7, на 20,56% выше (Рисунок 1n). В слое почвы 20~40 см, за исключением G4 и G6, активность уреазы других обработок была выше, чем у G7, на 6,84%~27,68% выше. Среди них G4 был значительно выше, чем G7, на 27,68% выше (Рисунок 1c). За исключением G3 и G4, активность инвертазы была выше, чем у G7, от 9,38% до 18,67% (Рисунок 1g). Активность каталазы каждой обработки была выше, чем у G7, на 0,59%~34,24% выше, и G3 был значительно выше, чем G7, на 34,24% выше (Рисунок 1k). Только активность щелочной фосфатазы G3 и G6 была выше, чем у G7, на 12,96% и 15,04% выше, соответственно (Рисунок 1o). В слое почвы 40~60 см, за исключением G4, активность уреазы других обработок была выше, чем у G7, на 6,56%~29,67% выше (Рисунок 1d). За исключением обработки G3, активность инвертазы была выше, чем у G7, от 0,19% до 45,58%, и G2 был значительно выше, чем G7, на 45,58% выше (Рисунок 1h). Только активность каталазы G1, G2 и G6 была выше, чем у G7, на 5,73%, 10,51% и 12,48% выше, соответственно (Рисунок 1l). Активность щелочной фосфатазы G3 и G6 также была выше, чем у G7, на 1,16% и 34,38% выше (Рисунок 1p).
Реакция активности каталазы между различными слоями почвы на методы обработки почвы в течение двух лет была не очевидной. Кроме того, за исключением снижения активности почвенных ферментов под G7 в слое почвы 10~20 см, другие слои почвы показали значительный рост в течение двух лет (Рисунок 1). В течение всего вегетационного периода наблюдались различия между активностями четырех почвенных ферментов, которые обрабатывались различными методами обработки почвы при заделке соломы. Глубина почвы, метод обработки почвы и время отбора проб значительно влияли на активность почвенных ферментов. Активности уреазы, инвертазы, каталазы и щелочной фосфатазы в каждой обработке были выше, чем при традиционной обработке (G7), причем активности глубокого рыхления (G3) и нулевой обработки (G6) были выше (Рисунок 1). По сравнению с обработкой традиционной обработкой, активности уреазы и инвертазы при обработке глубоким рыхлением увеличились на 13,65% и 9,48% соответственно, в слое почвы 40~60 см (Рисунок 1a–h), в то время как активности каталазы и щелочной фосфатазы при нулевой обработке увеличились на 21,03% и 57,21% соответственно, в слое почвы 40~60 см (Рисунок 1i–p).
3.2. Анализ микробной биомассы почвы
Различные методы обработки почвы значительно влияли на обилие углерода микробной биомассы почвы (MBC) и азота микробной биомассы почвы (MBN). Содержание микробной биомассы углерода показало тенденцию сначала увеличиваться, а затем уменьшаться, в то время как содержание микробной биомассы азота показало тенденцию сначала увеличиваться, а затем уменьшаться. Глубокое рыхление и тяжелое боронование (G5) были выше, чем другие, а традиционная обработка была самой низкой (Рисунок 2). В слое почвы 0~10 см микробная биомасса каждого метода обработки была G3 > G6 > G5 > G2 > G4 > G1 > G7 (Рисунок 2a). Общая тенденция в слое почвы 10~20 см была аналогична таковой в слое почвы 0~10 см. В этом слое почвы микробная биомасса каждой обработки обработки почвы от высокой к низкой была G3 > G4 > G6 > G2 > G5 > G1 > G7 (Рисунок 2b). В слое почвы 20~40 см микробная биомасса каждой обработки обработки почвы была G3 > G4 > G5 > G6 > G2 > G1 > G7 (Рисунок 2c). Общая тенденция оставалась неизменной в слое почвы 40~60 см (Рисунок 2d), и обработка G7 (традиционная обработка) все еще была самой низкой среди всех обработок. В целом, обработка G3 (глубокое рыхление и возврат соломы) всегда была значительно выше, чем традиционный метод, во всем слое почвы (Рисунок 2).
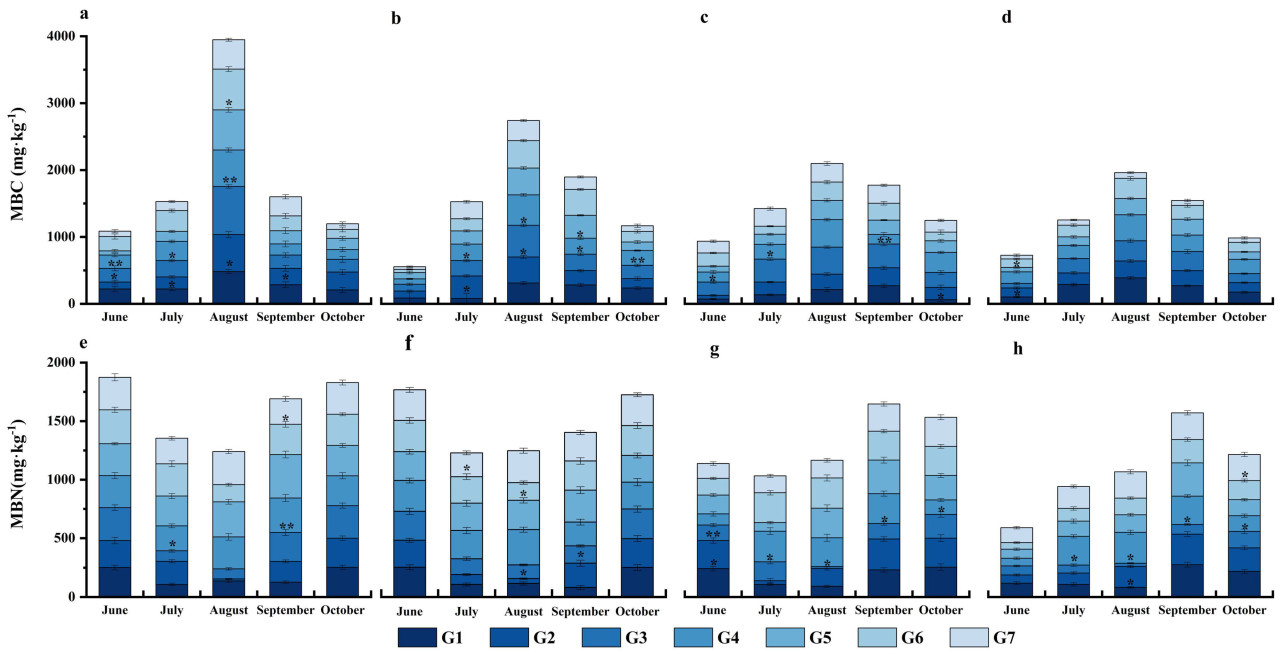
Рисунок 2. Влияние различных методов обработки почвы на MBC и MBN в разные периоды при заделке соломы (мг·кг⁻¹). Панели (a–d) представляют содержание MBC в слоях почвы 0~10, 10~20, 20~40 и 40~60 см соответственно; (e–h) представляют содержание MBN. Июнь: стадия всходов, Июль: стадия большого раструба, Август: стадия вымётывания и силосования, Сентябрь: стадия налива, Октябрь: после уборки. По сравнению с традиционной обработкой (G7), *: p < 0,05, : p < 0,01. Столбцы ошибок построены с SE.
После уборки урожая в 2020 году, в слое почвы 0~10 см, MBC каждой обработки был выше, чем у G7, на 36,16%~68,21% выше, в то время как он также был выше, чем у G7, от 33,45% до 64,51%, в слое почвы 10~20 см (Рисунок 2a,b). В слое почвы 20~40 см, MBC G2, G3 и G4 был выше, чем у G7, на 6,10%, 21,42% и 42,12% соответственно (Рисунок 2c). MBC каждой обработки был на 40,48%~69,14% выше, чем у G7, в слое почвы 40~60 см (Рисунок 2d). Не было большого изменения MBN в слое почвы 0~10 см от стадии всходов до послеуборочной стадии, и он был очень низким на стадии большого раструба и стадии вымётывания и силосования, что может быть связано со значительными дождями в экспериментальной зоне. MBN G3 и G6 был на 4,78% и 2,11% выше, чем у G7, соответственно. В слое почвы 10~20 см, содержание G3 после уборки также было выше, чем на стадии всходов (Рисунок 2e). В слое почвы 20~40 см, за исключением G1 и G2, MBN при других обработках увеличивался со временем. Содержание G1 и G6 было выше, чем у G7, на 2,71% и 0,22% соответственно. В слоях почвы 10~20 см и 40~60 см, MBN увеличивался при всех обработках, а изменения в других слоях почвы при каждой обработке были относительно незначительными (Рисунок 2).
MBN в слое почвы 0~20 см был выше, чем в слое почвы 20~60 см. MBN всех обработок в каждом слое почвы увеличивался в течение двух лет. Кроме того, в слое почвы 40~60 см, MBN в 40~60 см был ниже, чем в поверхностной почве, и каждая обработка в слое почвы увеличилась на 40,57%~88,13% после уборки урожая, из которых G6 увеличился на 88,13%, а G7 увеличился на 40,57%.
3.3. Обилие и состав сообщества бактерий и грибов
Не было значительной разницы в альфа-индексе разнообразия бактерий среди всех обработок. По сравнению с июнем, альфа-индекс разнообразия бактерий при каждой обработке в октябре значительно увеличился. В течение вегетационного периода не было значительной разницы в индексе Симпсона почвенных бактерий при различных методах обработки почвы. В октябре индекс Шеннона каждой обработки был выше, чем у G7, и G2 и G6 были на 9,31% и 5,95% выше, соответственно. Кроме того, по сравнению с июнем, индекс Шеннона других обработок увеличился в октябре, за исключением того, что индекс Шеннона G7 снизился на 3,27%. В июне индексы Чао G5 и G6 были выше, чем у G7, на 2,11% и 3,91% соответственно. В октябре каждая обработка была на 2,13%~38,57% выше, чем G7. Эти результаты также указывают на то, что почвозащитная обработка, особенно обработки G3 и G6, может улучшить разнообразие и богатство бактериального сообщества (Таблица 2).
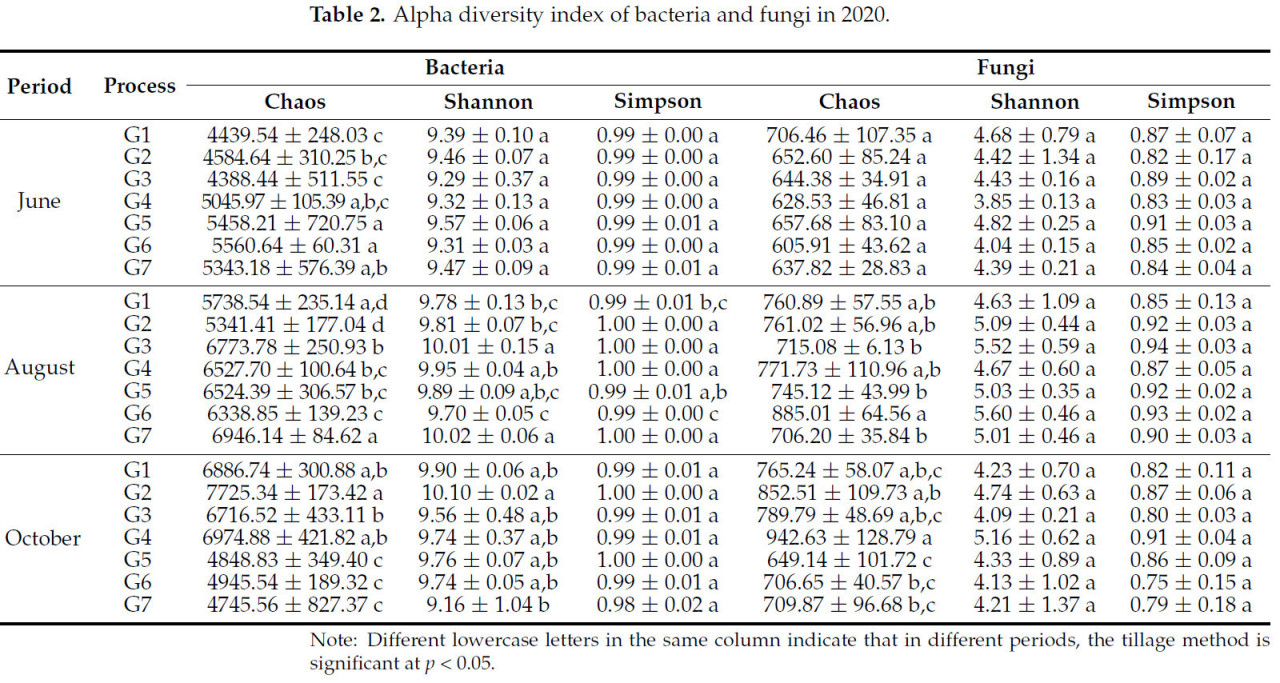
Таблица 2. Альфа-индекс разнообразия бактерий и грибов в 2020 году.
В октябре индекс Симпсона грибов в каждой обработке был ниже, чем в июне, и разница между обработками незначительна. В июне и октябре индекс Шеннона обработки G5 был на 8,92% и 2,77% выше, чем у G7, соответственно. В августе G6 был на 10,54% выше, чем G7. В июне и августе индекс Чао G1 был выше, на 9,72% и 7,19% соответственно. Кроме того, индекс Чао под G6 был на 20,20% выше, чем у G7 в августе. По сравнению с июнем, индекс каждой обработки в октябре увеличился на 7,68%~33,32%. Вышеуказанные результаты показывают, что тенденция изменения индекса разнообразия каждой обработки в течение всего вегетационного периода не была полностью последовательной, что указывает на то, что грибы более чувствительны к сезону, чем к обработке почвы (Таблица 2).
С точки зрения состава сообщества было установлено, что доминирующими бактериями являются Proteobacteria, Actinobacteria и Gemmatimonadetes (Рисунок 3a–c). С июня по август относительное содержание Bacteroidetes увеличилось, среди которых G3 увеличилось больше всего, на 42,66%; относительное содержание Acidobacteria и Gemmatimonadetes уменьшилось (Рисунок 3a,b). Доминирующими грибами были Ascomycota (Рисунок 3d–f). Грибы с относительной численностью >5% на уровне типа были Zygomycota и Basidiomycota (Рисунок 3a,c). По сравнению с июнем, доминирующие бактерии под G7 уменьшились в октябре, и относительное содержание Ascomycota уменьшилось на 33,18%. В каждый период Ascomycota, Zygomycota и Basidiomycota G6 были выше, чем другие обработки. В октябре относительное содержание Ascomycota G6 было на 28,66% выше, чем у G7 (Рисунок 3f). Анализ главных компонент показал, что основным фактором, влияющим на сообщество бактерий, было сезонное изменение, а основным фактором, влияющим на сообщество грибов, был метод обработки почвы.
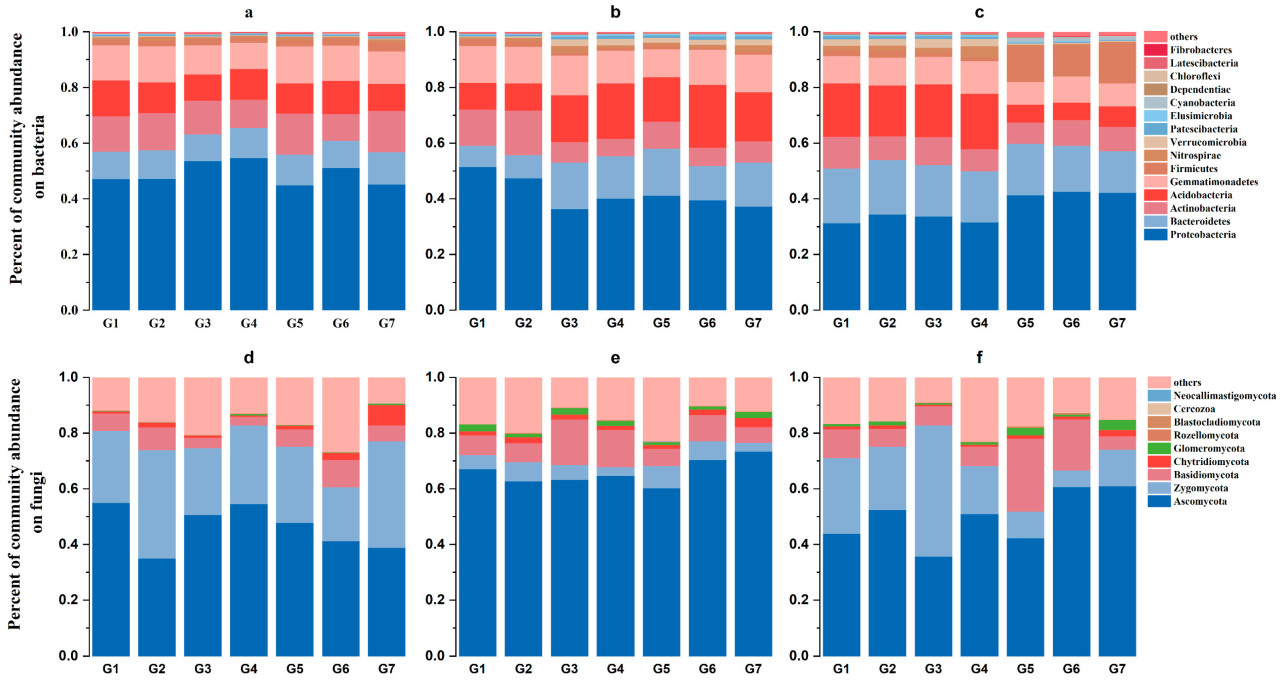
Рисунок 3. Диаграмма состава сообществ бактерий и грибов на уровне типов при различных методах обработки почвы в условиях заделки соломы. Панели (a–c) представляют сообщество бактерий на уровне типов в июне, августе и октябре. Панели (d–f) представляют сообщество грибов на уровне типов в июне, августе и октябре.
С помощью PCA-анализа бактериальных сообществ в различных обработках обработки почвы в разные периоды, G7 был наиболее дискретным в июне. Другие обработки были близки друг к другу, и бактериальные сообщества были схожи (Рисунок 4a). В августе G1 и G2 были похожи, а G3 был более дискретным. В октябре обработки G5, G6 и G7 значительно отличались от других обработок (Рисунок 4b). Результаты PCA для сообщества грибов показали, что обработка G7 была более дискретной в июне, что сильно отличалось от других обработок (Рисунок 4d). В сентябре не было значительной разницы между обработками, кроме G1 и G2 (Рисунок 4e). В октябре G7 был более дискретным, чем другие обработки (Рисунок 4f). Согласно диаграмме, сообщество грибов было менее подвержено сезонным колебаниям, чем сообщество бактерий.
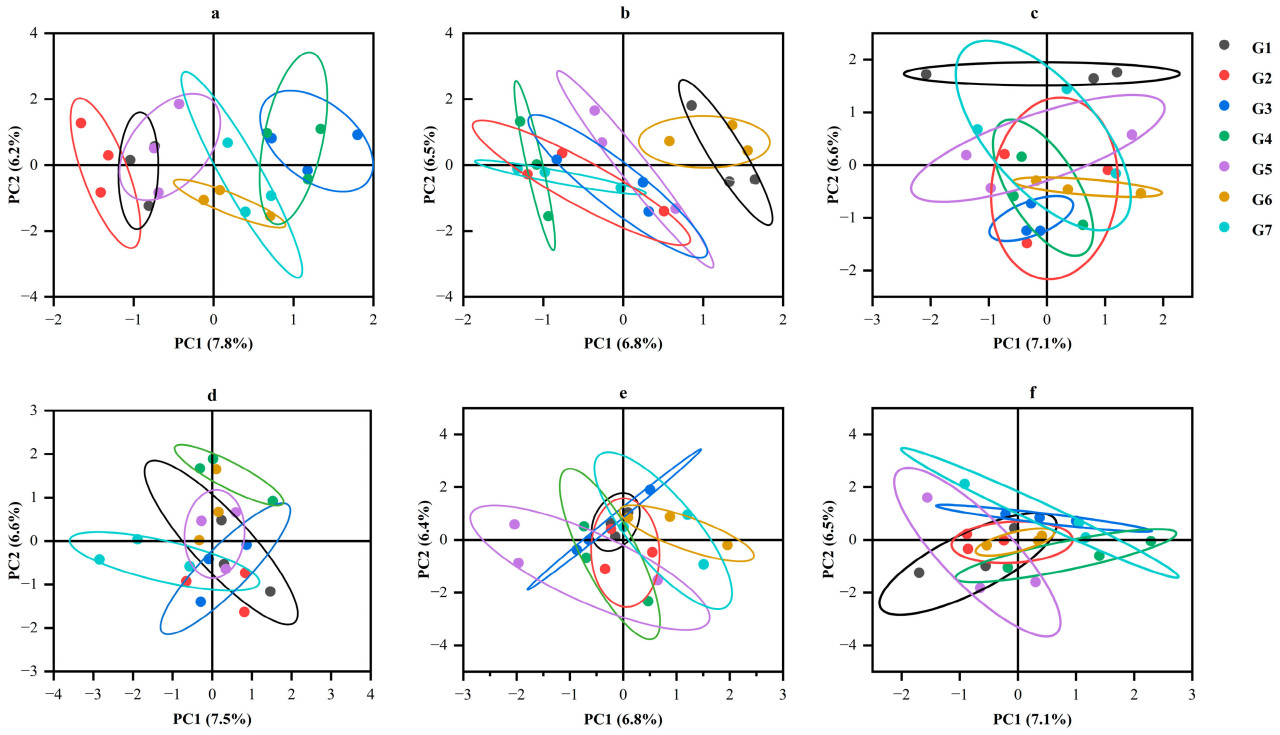
Рисунок 4. PCoA бактерий и грибов при различных методах обработки почвы. Панели (a–c) представляют PCoA бактерий в июне, августе и октябре при различных обработках; (d–f) представляют PCoA грибов.
3.4. Коэффициент корреляции Пирсона между активностью почвенных ферментов, микробной биомассой почвы и почвенным микробным разнообразием
Существовала значительная корреляция между большинством свойств почвы (p < 0,05). Различия в составе бактериального сообщества почвы были связаны с различиями в содержании уреазы, каталазы и углерода микробной биомассы (p < 0,01) (Рисунок 5). Существовала положительная корреляция между уреазой, каталазой и щелочной фосфатазой, и уреаза была значительно положительно коррелирована с каталазой (0,4 < r < 0,6, p < 0,01). Существовала значительная отрицательная корреляция между MBC и MBN (0,4 < r < 0,6, p < 0,05). Уреаза и каталаза были значительно положительно коррелированы с MBC (0,6 < r < 0,8, p < 0,05) и отрицательно коррелированы с MBN. Что касается бактерий, инвертаза была отрицательно коррелирована с индексами Чао, Шеннона и Симпсона. Щелочная фосфатаза была положительно коррелирована с индексами Шеннона и Симпсона и значительно положительно коррелирована с индексом Чао (0,4 < r < 0,6, p < 0,05). Два других фермента были значительно положительно коррелированы с индексами Чао и Шеннона (p < 0,01) и положительно коррелированы с индексом Симпсона. MBC был значительно положительно коррелирован с каждым индексом (p < 0,05), что было значительно положительно коррелировано с индексом Шеннона (p < 0,001), а MBN был отрицательно коррелирован с каждым индексом. В отношении грибов инвертаза была отрицательно коррелирована с индексом Чао, индексом Шеннона и индексом Симпсона, в то время как другие ферменты были положительно коррелированы с этими тремя индексами. Среди них уреаза была значительно положительно коррелирована с индексом Чао (0,4 < r < 0,6, p < 0,05), каталаза и индекс Шеннона были значительно положительно коррелированы с индексом Симпсона (p < 0,05), а щелочная фосфатаза была значительно положительно коррелирована с индексами Чао и Шеннона (p < 0,05). MBC был значительно положительно коррелирован с каждым индексом (p < 0,05) и был значительно положительно коррелирован с индексом Шеннона (p < 0,001). MBN был отрицательно коррелирован с каждым индексом и значительно отрицательно коррелирован с индексами Шеннона и Симпсона (p < 0,05).
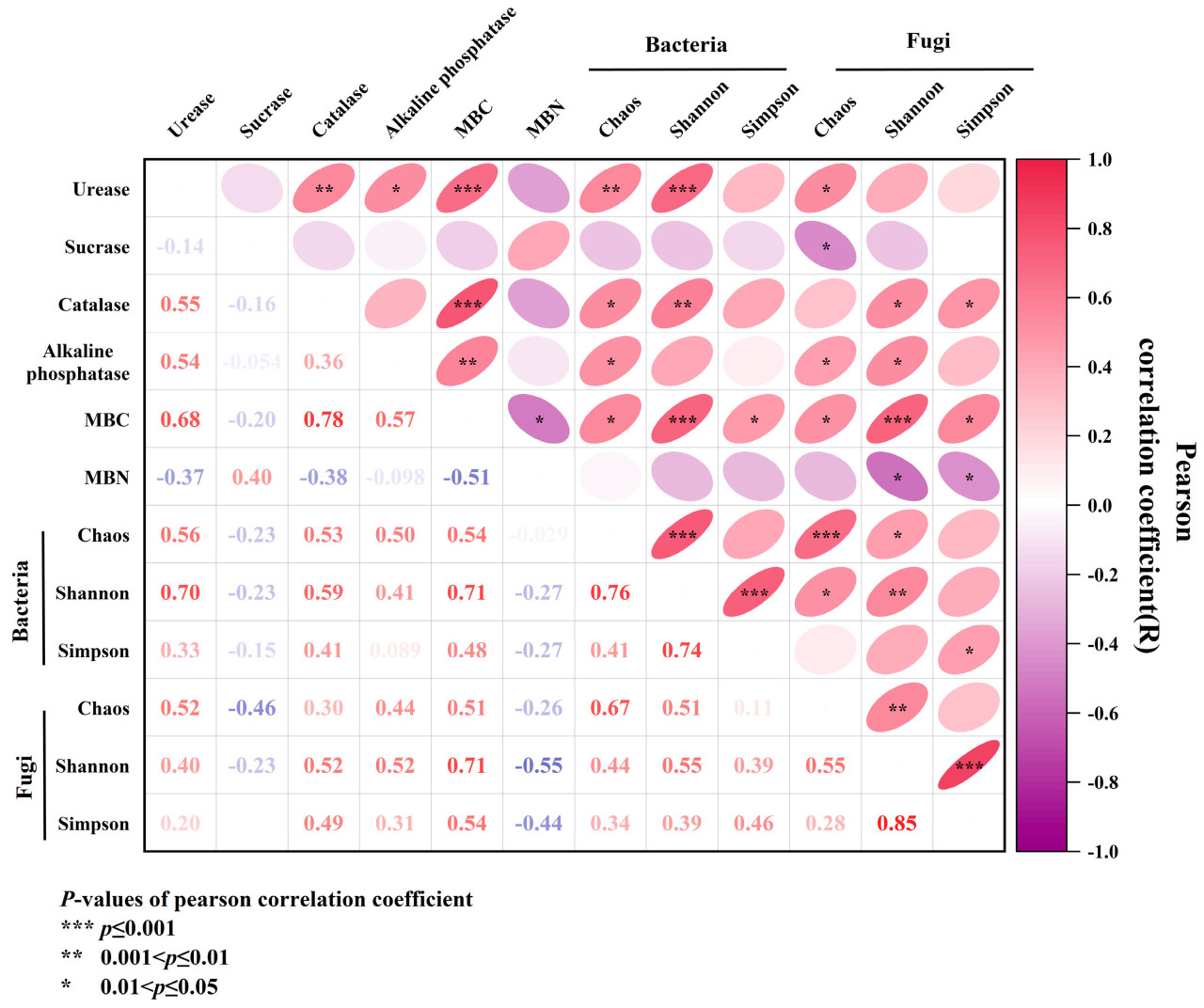
Рисунок 5. Корреляционный анализ Пирсона между активностью почвенных ферментов, микробной биомассой и почвенным микробным разнообразием.
Разнообразие Чао было сильно коррелировано с уреазой, щелочной фосфатазой и углеродом микробной биомассы (0,4 < r < 0,6, p < 0,05 или 0,01). Разнообразие Шеннона было сильно коррелировано с уреазой и углеродом микробной биомассы (0,5 < r < 0,8, p < 0,01). Разнообразие Симпсона было слабо коррелировано с углеродом микробной биомассы (0,4 < r < 0,6, p < 0,05). В то же время микробное разнообразие было отрицательно коррелировано с азотом микробной биомассы (0,2 < r < 0,4, p < 0,05) (Рисунок 5).
4. Обсуждение
4.1. Влияние различных методов обработки почвы на активность почвенных ферментов
По сравнению с традиционным методом обработки почвы, почвозащитная обработка имеет более высокую активность почвенных ферментов в верхнем слое почвы из-за меньшего вмешательства в почву [34]. Активность ферментов каждой обработки почвозащитной обработки была выше, чем у G7. Три обработки G4, G5 и G6 были выше, чем G7 в каждом слое почвы. Это связано с тем, что микроорганизмы могут разлагать солому после возврата в поле, и энергия, поставляемая процессом разложения и деградации, будет способствовать размножению почвенных микроорганизмов и стимулировать производство корневых экссудатов, таким образом помогая значительно улучшить активность ферментов в почве с возвратом соломы [35]. Однако активность ферментов G7 была относительно низкой из-за отсутствия возврата соломы.
Активность уреазы каждой обработки значительно увеличивалась с вегетационным периодом, и степень улучшения при обработке почвозащитной обработкой была выше, чем при обработке традиционной обработкой. Что не согласуется с результатами исследований Хамидо [36], так это то, что активность уреазы при вспашке была выше, чем при возврате соломы с фрезерованием в каждый период, возможно, из-за различных типов растений, типов почв, методов посадки и факторов окружающей среды. Активность инвертазы почвы показала волнообразные изменения в течение всего вегетационного периода сельскохозяйственных культур. С июня по август активность инвертазы почвы увеличивалась, а затем уменьшалась из-за явления, что микроорганизмы конкурируют с культурами за питательные вещества в этот период. После периода налива культуры поглощали меньше питательных веществ, и затем активность снова повышалась [37]. Подобно правилу изменения активности уреазы и инвертазы, активность каталазы почвы показала тенденцию к снижению с углублением почвы, что согласуется с результатами исследований Семанцовой [38]. Щелочная фосфатаза является одной из широко существующих гидролаз в почве и играет решающую роль в трансформации органического фосфора. Её активность непосредственно влияет на разложение, трансформацию и биодоступность органического фосфора в почве, отражая статус снабжения почвы фосфором [39]. В данном исследовании активность щелочной фосфатазы не имела значительной разницы между различными слоями почвы, что согласуется с результатами исследований Цзюньли [40].
В целом, активность ферментов каждой обработки показала общую тенденцию к росту. Глубокое рыхление и другие методы обработки почвы разрыхляют почву и превращают остатки разложения соломы в верхний слой почвы, увеличивая плодородие почвы. Кроме того, секреция корней растений также обеспечивает источник для почвенных ферментов [41], что положительно сказывается на увеличении почвенных ферментов.
4.2. Влияние различных методов обработки почвы на микробную биомассу почвы
Микробная биомасса углерода является важным этапом в трансформации почвенного углерода и регуляции цикла почвенных питательных веществ [42]. Традиционная обработка почвы часто нарушает почву, вызывая определенную степень повреждения почвенной среды, влияя на выживание микроорганизмов, тем самым снижая почвенную микробную биомассу [43,44]; наоборот, обработка глубоким рыхлением может значительно увеличить почвенную микробную биомассу [45]. Обработка фрезерованием (G4) превращает солому в почву, и микроорганизмы быстро разлагают солому, улучшают структуру пахотного слоя, увеличивают почвенные питательные вещества и обеспечивают хорошую материальную среду обитания для микроорганизмов. Метод почвозащитной обработки косвенно влияет на виды, количество и активность ферментов микроорганизмов при условии увеличения проницаемости почвы [46], что имеет большое значение для поддержания и улучшения биологического плодородия почвы и улучшения подземной среды роста сельскохозяйственных культур [47].
Данное исследование считает, что MBC в поверхностной почве выше, чем в глубокой почве. Это может быть связано с питательными веществами, небольшой активностью животных и растений и меньшим количеством остатков животных и растений с уменьшением корневых экссудатов в глубокой почве, что вызывает меньше доступных питательных веществ для почвенных микроорганизмов [48,49], тем самым снижая почвенную микробную биомассу, и, следовательно, снижая почвенную микробную биомассу. Этот результат аналогичен результату Антона [50], который считал, что MBC в 0~20 см был значительно выше, чем в 20~40 см. В течение роста и развития кукурузы MBC каждой обработки представляет собой вариацию: сначала рост, затем падение, а затем слабый подъем, что является максимальным значением на стадии вымётывания и силосования и минимальным на стадии налива. По мере непрерывного роста культур питательные вещества, необходимые им, будут постепенно обогащаться. Эти вещества необходимо получать из почвы [51]. В противном случае, начальная фаза вегетационного периода вызывает высокую температуру почвы. После того как большинство простых органических веществ разлагаются микроорганизмами, другие вещества в почве с трудом разлагаются, что будет влиять на пролиферацию и активность микроорганизмов, приводя к сезонным изменениям почвенной микрофлоры [52]. На стадии налива требуется масса питательных веществ, и разложение соломы в основном завершено в то же время; следовательно, MBC снижается [53]. В целом, G3 был самым высоким среди всех обработок. Это связано с тем, что глубокое рыхление в сезон кукурузы оказывало значительное влияние на MBC в слое почвы 0~10 см [54]. Глубокое рыхление может поддерживать доступность питательных веществ и значительно увеличивать MBC в каждом слое почвы [55].
MBN, как важный показатель количества и активности почвенных микроорганизмов, может эффективно отражать уровень плодородия почвы [56]. Влияние различных методов обработки почвы на азот микробной биомассы почвы показало, что G4 и G5 имели более высокий азот микробной биомассы, чем G7. MBN каждой обработки увеличивался со временем, в то время как во всем слое почвы он был ниже в июне, чем в октябре. Изменение среди всех обработок в каждом слое почвы было менее заметным из-за удлинения времени; постепенное усиление температуры почвы ускоряет разложение соломы и стерни, что делает остатки урожая, оставленные в почве в предыдущем году, производят больше доступных органических соединений. С отмиранием и опадением корней культур питательные вещества в почве также увеличиваются. Увеличение питательных веществ способствует росту и размножению органических трофических микроорганизмов в почве и увеличивает микробное количество [57]. Кроме того, почвенная микробная биомасса имеет явные сезонные колебания с методами обработки почвы и глубиной. Это тесно связано с влиянием многих факторов, таких как местные осадки и температура почвы [58].
4.3. Влияние различных методов обработки почвы на почвенное микробное разнообразие
Почвенные микроорганизмы играют ключевую роль в формировании и трансформации почвенных материалов и тесно связаны с плодородием и структурой почвы. Они являются ключевыми факторами, влияющими на качество, плодородие и продуктивность почвы [59]. Известно, что метод почвозащитной обработки может значительно влиять на состав микробных сообществ и оказывать различное влияние на относительное обилие различных категорий микроорганизмов [60]. В отношении бактериального сообщества, по сравнению с традиционной обработкой, нулевая обработка в сочетании с возвратом соломы увеличивала относительное обилие Acidobacteria и Bacteroidetes и уменьшала относительное обилие Actinobacteria [61,62]. Предыдущие исследования показали, что независимо от нулевой обработки или традиционной обработки, бактериальное сообщество состоит из Actinomycetes, Proteobacteria, Acidobacteria и Bacteroidetes [63]. Это совпадает с результатами, полученными в данном эксперименте. Поскольку почвенные бактериальные сообщества более чувствительны к изменениям окружающей среды, они могут по-разному реагировать на различные методы обработки почвы [64].
Напротив, было относительно мало исследований реакции сообществ грибов на почвозащитную обработку. Влияние почвозащитной обработки на состав сообщества почвенных грибов достигается за счет изменений физико-химических свойств почвы. По сравнению с обычной обработкой, нулевая обработка может эффективно улучшить состав и разнообразие сообществ грибов [65]. Мин [66] показал, что обработка нулевой обработкой в сочетании с мульчированием соломой увеличивала относительное обилие Ascomycota. Поскольку большинство Ascomycota играют важную роль в процессе разложения остатков урожая, данное исследование также показывает, что нулевая обработка может способствовать разложению и утилизации соломы грибными сообществами. Возврат соломы и глубокое рыхление могут обеспечить подходящую среду обитания для Ascomycota, таким образом сильно влияя на почвенные грибы Ascomycota [67].
Почвозащитная обработка может значительно увеличить разнообразие почвенного микробного сообщества. Нулевая обработка, глубокое рыхление и мульчирование соломой могут стимулировать индекс разнообразия, индекс богатства и индекс выравненности почвенных микроорганизмов при монокультуре пшеницы и режиме севооборота пшеница-кукуруза, особенно для грибов [68,69]. Кабири обнаружил, что [70] фрезерование более способствует улучшению структуры почвенного микробного сообщества, чем вспашка, потому что фрезерование имеет меньшее нарушение почвы, чем обработка вспашкой, и обеспечивает более стабильную среду обитания для микроорганизмов. Ичао [71] считал, что комбинация мульчирования соломой и уменьшенной обработки была полезна для увеличения общего количества бактерий в поверхностной почве и увеличения почвенного микробного разнообразия. Исследования показали, что почвозащитная обработка, особенно нулевая обработка с возвратом соломы, может значительно увеличить альфа-индекс грибов поверхностной почвы, и количество специфических OTUs почвенных грибов при нулевой обработке с возвратом соломы выше, чем при традиционной обработке [72]. Вышеуказанные исследования согласуются с результатами данного эксперимента.
4.4. Взаимосвязи между активностью почвенных ферментов, микробной биомассой почвы и почвенным микробным разнообразием
Чтобы лучше понять корреляцию между свойствами почвы и микробными сообществами, мы использовали корреляционный анализ Пирсона [73] для объяснения их взаимодействия. Активность почвенных ферментов и содержание микробной биомассы тесно связаны друг с другом и коррелируют с разнообразием бактериальных и грибных сообществ.
Активность ферментов была положительно коррелирована с бактериями и грибами, что указывает на то, что увеличение активности ферментов способствовало метаболической способности почвы и играло активную роль в биохимическом процессе в почве. В то же время, это могло удалять отрицательные микроорганизмы через метаболизм и накапливать обилие полезных бактерий. Напротив, увеличение микробного обилия также производило бы больше почвенных ферментов [74].
MBC представляет углерод в различных организмах в почве; микробная биомасса азота обеспечивает азот и питание для микроорганизмов, что способствует поддержанию уровня выживаемости и видового богатства микроорганизмов. Содержание почвенной микробной биомассы непосредственно отражает уровень почвенной активности. Высокое содержание микробной биомассы указывает на более высокое качество почвы [75]. Индекс Шеннона отражает богатство живых микроорганизмов, поэтому непрерывная вариация является справедливой. В заключение следует отметить, что существует мало исследований, посвященных корреляции между активностью ферментов, микробной биомассой и микробным разнообразием, которые еще необходимо изучать и улучшать.
Хотя порядок пропорций каждого типа при различных методах обработки почвы различен, общая разница не значительна. Бактериальное сообщество значительно коррелировало с уреазой почвы, каталазой, щелочной фосфатазой и углеродом микробной биомассы (p < 0,01), что согласуется с результатами Бхарати [76], который обнаружил, что активность почвенных ферментов является основным фактором, значительно влияющим на бактериальное сообщество. Исследуя влияние различных методов обработки почвы на почвенные микроорганизмы, было обнаружено, что существует также поразительная взаимосвязь между органическим веществом и почвенным бактериальным сообществом [77].
Функция почвозащитной обработки на сообщество почвенных грибов достигается через изменения физико-химических свойств почвы. По сравнению с обычной обработкой, нулевая обработка может эффективно исправлять сообщества грибов [78]. Метод нулевой обработки в сочетании с мульчированием соломой увеличивал относительное обилие аскомицетов [79]. Поскольку большинство Ascomycota играют решающую роль в процессе разложения остатков урожая, данное исследование также показало, что нулевая обработка может способствовать разложению и утилизации соломы грибными сообществами. Возврат соломы и глубокое рыхление могут обеспечить подходящую среду обитания для Ascomycota и, таким образом, сильно влиять на почвенные грибы аскомицеты [80].
Горизонтальное сравнение количества возвращаемой соломы в сочетании с различными методами обработки почвы исключило другие факторы, мешающие проведению теста. Результаты показывают, что бактериальное сообщество в вариантах G5 и G6 всегда было более многочисленным в варианте G7, а грибковое сообщество в варианте G7 было более многочисленным, чем в других вариантах. Анализ β-разнообразия показал, что основным фактором, влияющим на состав бактериального сообщества, было время отбора проб, а основным фактором, влияющим на грибковое сообщество, – способ обработки почвы.
5. Выводы
В заключение следует отметить, что методы консервативной обработки почвы, включая глубокое рыхление и нулевую обработку в сочетании с обработками возврата соломы, оказали положительное влияние на активность почвенных ферментов, микробную биомассу, микробное разнообразие и показатели урожайности в агропастбищных районах у подножия гор Дасинъань во Внутренней Монголии. Эти методы способствовали повышению плодородия почв, улучшению экологической обстановки в почве и эффективному смягчению проблем ветровой эрозии и опустынивания.
Ссылки
1. De Oliveira Jocenei, A.T.; Pires Luiz, F.; Cássaro Fabio, A.M.; Gaspareto José, V.; Posadas Adolfo, N.D.; Mooney Sacha, J. Soil pore system complexity and heterogeneity as affected by contrasting management practices. Soil Tillage Res. 2022, 224, 105497. [Google Scholar] [CrossRef]
2. Wenfeng, C.; Yuanyuan, Z.; Wenhui, K.; Tao, P.; Tu, B.; Jinshen, Z.; Liang, J.; Sisi, W. Impact of Cropland Evolution on Soil Wind Erosion in Inner Mongolia of China. Land 2021, 10, 583. [Google Scholar] [CrossRef]
3. Shankar, B.S.; Wim, D.V.; Roberto, P.-S. Soil carbon sequestration, greenhouse gas emissions, and water pollution under different tillage practices. Sci. Total Environ. 2022, 826, 154161. [Google Scholar]
4. Zhang, M.; Chen, S.; Ding, S.; Yao, X.; Wang, Z.; Sang, L. Effects of 7 Years of Warming and Straw Application on Soil Bacterial, Fungal, and Archaeal Community Compositions and Diversities in a Crop Field. J. Soil Sci. Plant Nutr. 2022, 22, 2266–2281. [Google Scholar] [CrossRef]
5. Xie, W.; Zhang, Y.; Li, J.; Wei, S.; Li, X.; Yu, H.; Guan, B. Straw application coupled with N and P supply enhanced microbial biomass, enzymatic activity, and carbon use efficiency in saline soil. Appl. Soil Ecol. 2021, 168, 104128. [Google Scholar] [CrossRef]
6. Zhang, M.; Song, D.; Pu, X.; Dang, P.; Qin, X.; Siddique, K.H. Effect of different straw returning measures on resource use efficiency and spring maize yield under a plastic film mulch system. Eur. J. Agron. 2022, 134, 126461. [Google Scholar] [CrossRef]
7. Wang, C.; Xue, L.; Dong, Y.; Jiao, R. Effects of stand density on soil microbial community composition and enzyme activities in subtropical Cunninghamia lanceolate (Lamb.) Hook plantations. For. Ecol. Manag. 2021, 479, 118559. [Google Scholar] [CrossRef]
8. Gupta, V.; Parmar, B.B.; Bhayal, D.; Kumar, K.; Yadav, B.; Choudhary, G.; Goyal, U. Assessment of Soil Enzyme Activity in Soybean Fields under Long-Term Fertilization and Manuring Practices. J. Adv. Biol. Biotechnol. 2024, 27, 1432–1439. [Google Scholar] [CrossRef]
9. Pang, X.; Jia, M.; Zhang, Y.; Chen, M.; Miao, P.; Cheng, W.; Zhou, Z.; Zhang, Q.; Ye, J.; Li, J.; et al. Dynamic interplay between soil microbial communities, enzyme activities, and pear quality across planting years. Front. Microbiomes 2024, 3, 1381270. [Google Scholar] [CrossRef]
10. Liu, S.; Chen, Y.; Li, X.; Lv, J.; Yang, X.; Li, J.; Bai, Y.; Zhang, S. Linking soil nutrients, microbial community composition, and enzyme activities to saponin content of Paris polyphylla after addition of biochar and organic fertiliser. Chemosphere 2024, 363, 142856. [Google Scholar] [CrossRef]
11. Hosseini, S.H.; Liang, X.; Niyungeko, C.; Zakeritabar, S.F. Soil nutrients, phosphatase activities and phosphorus species distribution in soils amended sheep manure biochar. J. Soils Sediments 2024, 24, 3297–3310. [Google Scholar] [CrossRef]
12. Zhao, X.; Zhang, X.; Li, Z.; Wang, B.; Zhang, T.; Wan, P. Development of root rot in Zanthoxylum bungeanum is closely linked to changes in soil microbial communities, enzyme activities, and physicochemical factors. Glob. Ecol. Conserv. 2024, 55, e03249. [Google Scholar] [CrossRef]
13. Szczepanek, M.; Siwik-Ziomek, A.; Lemańczyk, G.; Lamparski, R.; Graczyk, R. Effect of Reduced Tillage on Soil Enzyme Activity, Pests Pressure and Productivity of Organically Grown Spring Wheat Species. Agronomy 2023, 13, 287. [Google Scholar] [CrossRef]
14. Huang, W.; Wu, J.F.; Pan, X.H.; Tan, X.M.; Zeng, Y.J.; Shi, Q.H.; Liu, T.J.; Zeng, Y.H. Effects of long-term straw return on soil organic carbon fractions and enzyme activities in a double-cropped rice paddy in South China. J. Integr. Agric. 2021, 20, 236–247. [Google Scholar] [CrossRef]
15. Yuan, L.; Gao, Y.; Mei, Y.; Liu, J.; Kalkhajeh, Y.K.; Hu, H.; Huang, J. Effects of continuous straw returning on bacterial community structure and enzyme activities in rape-rice soil aggregates. Sci. Rep. 2023, 13, 2357. [Google Scholar] [CrossRef]
16. Alvey, S.; Yang, C.-H.; Buerkert, A.; Crowley, D.E. Cereal/legume rotation effects on rhizosphere bacterial community structure in west african soils. Biol. Fertil. Soils 2003, 37, 73–82. [Google Scholar] [CrossRef]
17. Yang, H.; Meng, Y.; Feng, J.; Li, Y.; Zhai, S.; Liu, J. Direct and indirect effects of long-term ditch-buried straw return on soil bacterial community in a rice–wheat rotation system. Land Degrad. Dev. 2020, 31, 851–867. [Google Scholar] [CrossRef]
18. Liu, D.; Wang, B.; Bhople, P.; Davlatbekov, F.; Yu, F. Land rehabilitation improves edaphic conditions and increases soil microbial biomass and abundance. Soil Ecol. Lett. 2020, 2, 145–156. [Google Scholar] [CrossRef]
19. Dong, M.; Zhou, H.; Wang, J. Responses of soil microbial metabolism, function and soil quality to long-term addition of organic materials with different carbon sources. Biochar 2024, 6, 80. [Google Scholar] [CrossRef]
20. Hoon, L.J.; Gyu, K.M. Simulative Study of Effects of LM Microorganism on Environment: Analyses of Metabolomes and Soil Microbial. Korean J. Environ. Agric. 2019, 38, 197–204. [Google Scholar]
21. Ling, N.; Deng, K.; Song, Y.; Wu, Y.; Zhao, J.; Raza, W.; Huang, Q.; Shen, Q. Variation of rhizosphere bacterial community in watermelon continuous mono-cropping soil by long-term application of a novel bioorganic fertilizer. Microbiol. Res. 2014, 169, 570–578. [Google Scholar] [CrossRef] [PubMed]
22. Yang, Y.; Zhang, Y.; Yu, X.; Jia, G. Soil microorganism regulated aggregate stability and rill erosion resistance under different land uses. Catena 2023, 228, 107176. [Google Scholar] [CrossRef]
23. Yang, J.; Liu, Q.; Liu, K.; Wang, S.; Gu, H. Chronosequence changes in soil microbes under Populus simonii × P. nigra stands: Implications for soil health restoration in Mollisols. Appl. Soil Ecol. 2025, 206, 105826. [Google Scholar] [CrossRef]
24. Liu, J.; Li, F.Y.; Liu, J.; Wang, S.; Liu, H.; Ding, Y.; Ji, L. Long-term straw return promotes soil phosphorus cycling by enhancing soil microbial functional genes responsible for phosphorus mobilization in the rice rhizosphere. Agric. Ecosyst. Environ. 2025, 381, 109422. [Google Scholar]
25. Aminollah, M.B.; Khosro, A.; Mohammad, F.; Reza, E.H. Effects of different tillage systems on soil properties, and yield and yield components of barley. Arch. Agron. Soil Sci. 2022, 68, 1784–1795. [Google Scholar]
26. Hannu, F.; Tero, T.; Luigi, O.; Loredana, C.; Jessica, C.; Margarita, R.; Antonio, P.J.; Raúl, Z.; Marcos, E.C.; Kristiina, L. Effect of no-till followed by crop diversification on the soil microbiome in a boreal short cereal rotation. Biol. Fertil. Soils 2024, 60, 357–374. [Google Scholar]
27. Liu, S.; Hao, C.; Xie, Z.; Wu, Y.; Liang, A.; Chang, L.; Wu, D.; Chen, T.W. Conservation tillage impacts on soil biodiversity: Additional insights from the Collembola-associated bacteria. Agric. Ecosyst. Environ. 2024, 362, 108827. [Google Scholar] [CrossRef]
28. Zhang, M.; Dang, P.; Haegeman, B.; Han, X.; Wang, X.; Pu, X.; Qin, X.; Siddique, K.H.M. The effects of straw return on soil bacterial diversity and functional profiles: A meta-analysis. Soil Biol. Biochem. 2024, 195, 109484. [Google Scholar] [CrossRef]
29. Sui, J.; Wang, C.; Hou, F.; Shang, X.; Zhao, Q.; Zhang, Y.; Hou, Y.; Hua, X.; Chu, P. Effects of Deep Tillage on Rhizosphere Soil and Microorganisms During Wheat Cultivation. Microorganisms 2024, 12, 2339. [Google Scholar] [CrossRef]
30. Guan, S.Y.; Zhang, D.; Zhang, Z. Soil Enzyme and Its Research Methods; Chinese Agricultural Press: Beijing, China, 1986; pp. 274–297. [Google Scholar]
31. Karaca, A.; Cetin, S.C.; Turgay, O.C.; Kizilkaya, R. Soil Enzymology; Springer: Heidelberg, Germany, 2011; pp. 119–148. [Google Scholar]
32. Franke-Whittle, I.H.; Manici, L.M.; Insam, H. Rhizosphere bacteria and fungi associated with plant growth in soils of three replanted apple orchards. Plant Soil 2015, 395, 317–333. [Google Scholar] [CrossRef]
33. Vance, E.D.; Brookes, P.O.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol 1987, 19, 703–707. [Google Scholar] [CrossRef]
34. Liu, W.S.; Wei, Y.X.; Deng, P.P.; Oladele, O.P.; N’Dri Bohoussou, Y.; Dang, Y.P.; Zhao, X.; Zhang, H.L. Conservation tillage increases surface soil organic carbon stock by altering fungal communities and enzyme activity. Environ. Sci. Pollut. Res. Int. 2023, 30, 80901–80915. [Google Scholar] [CrossRef] [PubMed]
35. Liu, L.; Cheng, M.; Jin, J.; Fu, M. Effects of different nitrogen applications and straw return depth on straw microbial and carbon and nitrogen cycles in paddy fields in the cool zone. Sci. Rep. 2024, 14, 6424. [Google Scholar] [CrossRef] [PubMed]
36. Hamido, S.A.; Kpomblekou, K. Cover crop and tillage effects on soil enzyme activities following tomato. Soil Tillage Res. 2009, 105, 269–274. [Google Scholar] [CrossRef]
37. Misha, K.; Kamaljit, K.; Gurjit, K. Effect of osmo priming on sucrose metabolism in spring maize, during the period of grain filling, under limited irrigation conditions. Chem. Chem. 2019, 25, 1367–1376. [Google Scholar]
38. Semancová, P.; Kotorová, D.; Fazekašová, D.; Michaeli, E.; Solár, V. The activity of soil catalase, depending on the soil granulometric fractions and soil depth for example dry polder besa(slovakia). Ecol. Saf. 2015, 9, 96–103. [Google Scholar]
39. Wang, Y.; Huang, R.; Xu, G.; Li, J.; Wang, Z.; Ci, E.; Gao, M. Soil alkaline phosphatase activity and bacterial phoD gene abundance and diversity under regimes of inorganic fertilizer reduction with straw. J. Soils Sediments 2020, 21, 388–402. [Google Scholar] [CrossRef]
40. Hu, J.; Yang, A.; Wang, J.; Zhu, A.; Dai, J.; Wong, M.H.; Lin, X. Arbuscular mycorrhizal fungal species composition, propagule density, and soil alkaline phosphatase activity in response to continuous and alternate no-tillage in Northern China. Catena 2015, 133, 215–220. [Google Scholar] [CrossRef]
41. Kumar, A.; Dorodnikov, M.; Splettstößer, T.; Kuzyakov, Y.; Pausch, J. Effects of maize roots on aggregate stability and enzyme activities in soil. Geoderma 2017, 306, 50–57. [Google Scholar] [CrossRef]
42. Liu, C.; Lu, M.; Cui, J.; Li, B.; Fang, C. Effects of straw carbon input on carbon dynamics in agricultural soils: A meta-analysis. Glob. Change Biol 2014, 20, 1366–1381. [Google Scholar] [CrossRef]
43. Lungmuana, L.; Singh, S.B.; Vanthawmliana, V.; Saurav Saha, S.S.; Dutta, S.K.; Rambuatsaiha, R.; Singh, A.R.; Boopathi, T. Impact of secondary forest fallow period on soil microbial biomass carbon and enzyme activity dynamics under shifting cultivation in North Eastern Hill region, India. Catena 2017, 156, 10–17. [Google Scholar] [CrossRef]
44. Zhang, X.; Ren, X.; Cai, L. Effects of Different Straw Incorporation Amounts on Soil Organic Carbon, Microbial Biomass, and Enzyme Activities in Dry-Crop Farmland. Sustainability 2024, 16, 10588. [Google Scholar] [CrossRef]
45. Liu, L.; Zhang, L.; Liu, J.; Fu, Q.; Xu, Q.; Idimesheva, O. Soil water and temperature characteristics under different straw mulching and tillage measures in the black soil region of China. Soil Water Conserv 2021, 76, 256–262. [Google Scholar] [CrossRef]
46. Zhang, H.; Tang, X.; Hou, Q. Combining conservation tillage with nitrogen fertilization promotes maize straw decomposition by regulating soil microbial community and enzyme activities. Pedosphere 2024, 34, 783–796. [Google Scholar] [CrossRef]
47. Douibi, M.; Carpio, J.M.; Cruz, R.S.M. Changes in Soil Microbial Parameters after Herbicide Application in Soils under Conventional Tillage and Non-Tillage. Processes 2024, 12, 827. [Google Scholar] [CrossRef]
48. Tang, H.; Li, C.; Shi, L.; Wen, L.; Li, W.; Cheng, K.; Xiao, X. Tillage with Crop Residue Returning Management Increases Soil Microbial Biomass Turnover in the Double-Cropping Rice Fields of Southern China. Agronomy 2024, 14, 265. [Google Scholar] [CrossRef]
49. Pinheiro Alves de Souza, Y.; Schloter, M.; Weisser, W.; Schulz, S. Deterministic Development of Soil Microbial Communities in Disturbed Soils Depends on Microbial Biomass of the Bioinoculum. Microb. Ecol. 2023, 86, 2882–2893. [Google Scholar] [CrossRef]
50. Anton, G.; Živa, P.; Klemen, E.; Rok, M.; Marjetka, S. Combined effects of long-term tillage and fertilisation regimes on soil organic carbon, microbial biomass, and abundance of the total microbial communities and N-functional guilds. Appl. Soil Ecol. 2023, 188, 104876. [Google Scholar]
51. Wang, P.; Xie, W.; Ding, L.; Zhuo, Y.; Gao, Y.; Li, J.; Zhao, L. Effects of Maize–Crop Rotation on Soil Physicochemical Properties, Enzyme Activities, Microbial Biomass and Microbial Community Structure in Southwest China. Microorganisms 2023, 11, 2621. [Google Scholar] [CrossRef]
52. Qu, R.; Liu, G.; Yue, M.; Wang, G.; Peng, C.; Wang, K.; Gao, X. Soil temperature, microbial biomass and enzyme activity are the critical factors affecting soil respiration in different soil layers in Ziwuling Mountains, China. Front. Microbiol. 2023, 14, 1105723. [Google Scholar] [CrossRef]
53. Wang, K.K.; Ren, T.; Yan, J.Y.; Zhu, D.D.; Fang, Y.T.; Cong, R.H.; Lu, Z.F.; Li, X.K.; Lu, J.W. Straw residue incorporation: Influence on soil microbial biomass and carbon–nitrogen dynamics in an oilseed rape–rice rotation. Soil Use Manag. 2023, 40, e12990. [Google Scholar] [CrossRef]
54. Yang, X.; Yue, L.; Wu, Y.; Wang, B.; Wu, L.; Liu, S.; Chen, H.; Chen, D. The validity of ecological hypotheses concerning aboveground organisms for soil microbial biomass and diversity across soil depths on the Mongolian Plateau. Appl. Soil Ecol. 2023, 181, 104679. [Google Scholar] [CrossRef]
55. Wang, K.; Ren, T.; Yan, J.; Zhu, D.; Liao, S.; Zhang, Y.; Lu, Z.; Cong, R.; Li, X.; Lu, J. Straw returning mediates soil microbial biomass carbon and phosphorus turnover to enhance soil phosphorus availability in a rice-oilseed rape rotation with different soil phosphorus levels. Agric. Ecosyst. Environ. 2022, 335, 107991. [Google Scholar] [CrossRef]
56. Sousa, R.S.D.; Nunes, L.A.P.L.; Antunes, J.E.L.; Araujo, A.S.F.D. Maize rhizosphere soil stimulates greater soil microbial biomass and enzyme activity leading to subsequent enhancement of cowpea growth. Environ. Sustain. 2019, 2, 89–94. [Google Scholar] [CrossRef]
57. Ilka, E.; Deborah, L.; Mignon, S.; Georg, J.R.; Catharina, M.; Martin, P. The Effects of Conservation Tillage on Chemical and Microbial Soil Parameters at Four Sites across Europe. Plants 2022, 11, 1747. [Google Scholar] [CrossRef]
58. Mang’erere, N.M.; Mugendi, N.E.; Monicah, M.M. Tillage, Mulching and Nitrogen Fertilization Differentially Affects Soil Microbial Biomass, Microbial Populations and Bacterial Diversity in a Maize Cropping System. Front. Sustain. Food Syst. 2021, 5, 614527. [Google Scholar]
59. Peng, Y.; Xu, H.; Shi, J.; Wang, Z.; Lv, J.; Li, L.; Wang, X. Soil microbial composition, diversity, and network stability in intercropping versus monoculture responded differently to drought. Agric. Ecosyst. Environ. 2024, 365, 108915. [Google Scholar] [CrossRef]
60. Sant’Anna, G.S.L.; Franzan, B.C.; Biassi, D.; dos Santos, C.M.; Schultz, N.; Zonta, E.; da Silva Coelho, I. Short-term effect of no-tillage management on the chemical attributes and bacterial diversity in soils in an organic production system. Soil Secur. 2024, 16, 100143. [Google Scholar] [CrossRef]
61. Zhang, Y.; Zhang, M.; Tang, L.; Che, R.; Chen, H.; Blumfield, T.; Boyd, S.; Nouansyvong, M.; Xu, Z. Long-Term Harvest Residue Retention Could Decrease Soil Bacterial Diversities Probably Due to Favouring Oligotrophic Lineages. Microb. Ecol. 2018, 76, 771–781. [Google Scholar] [CrossRef]
62. Guan, Y.; Wu, M.; Che, S.; Yuan, S.; Yang, X.; Li, S.; Tian, P.; Wu, L.; Yang, M.; Wu, Z. Effects of Continuous Straw Returning on Soil Functional Microorganisms and Microbial Communities. J. Microbiol. 2023, 61, 49–62. [Google Scholar] [CrossRef]
63. Guo, L.; Zheng, S.; Cao, C.; Li, C. Tillage practices and straw-returning methods affect topsoil bacterial community and organic C under a rice-wheat cropping system in central China. Sci. Rep. 2016, 6, 33155. [Google Scholar] [CrossRef] [PubMed]
64. Veloso, M.G.; Angers, D.A.; Chantigny, M.H.; Bayer, C. Carbon accumulation and aggregation are mediated by fungi in a subtropical soil under conservation agriculture. Geoderma 2020, 363, 114159. [Google Scholar] [CrossRef]
66. Gao, M.; Li, H.; Li, M. Effect of No Tillage System on Soil Fungal Community Structure of Cropland in Mollisol: A Case Study. Front. Microbiol. 2022, 13, 847691. [Google Scholar] [CrossRef]
67. Zhang, R.P.; Huang, Z.; Ashen, R.; Zhou, N.N.; Zhou, L.; Feng, T.Y.; Zhang, K.Y.; Liao, X.H.; Aer, L.; Ma, P. Phosphorus Application during Rapeseed Season Combined with Straw Return Improves Crop Productivity and Soil Bacterial Diversity in Rape-Rice Rotation. Agronomy 2023, 13, 506. [Google Scholar] [CrossRef]
68. Wang, Z.; Liu, L.; Chen, Q.; Wen, X.; Liao, Y. Conservation tillage increases soil bacterial diversity in the dryland of northern China. Agron. Sustain. Dev. 2016, 36, 28. [Google Scholar] [CrossRef]
69. Jackson, L.E.; Calderon, F.J.; Steenwerth, K.L.; Scow, K.M.; Rolston, D.E. Responses of soil microbial processes and community structure to tillage events and implications for soil quality. Geoderma 2003, 114, 305–317. [Google Scholar] [CrossRef]
70. Kabiri, V.; Raiesi, F.; Ghazavi, M.A. Tillage effects on soil microbial biomass, SOM mineralization and enzyme activity in a semi-arid Calcixerepts. Agric. Ecosyst. Environ. 2016, 232, 73–84. [Google Scholar] [CrossRef]
71. Yichao, S.; Roger, L.; Chantal, H.; Noura, Z. Winter effect on soil microorganisms under different tillage and phosphorus management practices in eastern Canada. Can. J. Microbiol. 2015, 61, 315–326. [Google Scholar]
72. Zhang, B.; Li, Y.; Ren, T.; Tian, Z.; Wang, G.; He, X.; Tian, C. Short-term effect of tillage and crop rotation on microbial community structure and enzyme activities of a clay loam soil. Biol. Fertil. Soils 2014, 50, 1077–1085. [Google Scholar] [CrossRef]
73. Qi, L.; Zhang, X.; Peng, Z.; Zhou, J. Canonical correlation analysis of soil nutrients, microorganisms and enzyme activities in vegetation restoration areas of degraded and eroded soils in northwestern Hunan Province, China. Front. For. China 2009, 4, 443–449. [Google Scholar] [CrossRef]
74. Han, C.; Zhou, W.; Gu, Y.; Wang, J.; Zhou, Y.; Xue, Y.; Shi, Z.; Siddique, K.H. Effects of tillage regime on soil aggregate-associated carbon, enzyme activity, and microbial community structure in a semiarid agroecosystem. Plant Soil 2024, 498, 543–559. [Google Scholar] [CrossRef]
75. Torabian, S.; Kim, E.; Qin, R.; Sathuvalli, V.; Gollany, H.T.; Kleber, M. Soil microbial biomass influenced by cover crop after fumigation of potato fields. Sci. Total Environ. 2024, 958, 177910. [Google Scholar] [CrossRef] [PubMed]
76. Bharathi, M.J.; Anbarasu, M.; Ragu, R.; Subramanian, E. Assessment of soil microbial diversity and soil enzyme activities under inorganic input sources on maize and rice ecosystems. Saudi J. Biol. Sci. 2024, 31, 103978. [Google Scholar] [CrossRef]
77. Cotrufo, M.F.; Haddix, M.L.; Kroeger, M.E.; Stewart, C.E. The role of plant input physical-chemical properties, and microbial and soil chemical diversity on the formation of particulate and mineral-associated organic matter. Soil Biol. Biochem. 2022, 168, 108648. [Google Scholar] [CrossRef]
78. Dos Santos Soares, D.; Ramos, M.L.G.; Marchão, R.L.; Maciel, G.A.; de Oliveira, A.D.; Malaquias, J.V.; de Carvalho, A.M. How diversity of crop residues in long-term no-tillage systems affect chemical and microbiological soil properties. Soil Tillage Res. 2019, 194, 104316. [Google Scholar] [CrossRef]
79. Zhu, X.C.; Sun, L.Y.; Song, F.B.; Liu, S.Q.; Liu, F.L.; Li, X.N. Soil microbial community and activity are affected by integrated agricultural practices in China. Eur. J. Soil Sci. 2018, 69, 924–935. [Google Scholar] [CrossRef]
80. Gajda, A.; Przewłoka, B. Soil biological activity as affected by tillage intensity. Int. Agrophys. 2012, 26, 15–23. [Google Scholar] [CrossRef]

Xiao S, Li B, Zhang T, Luo J, Wang J, Zhang X, Li J, Zhang D. Effects of Different Tillage Measures Combined with Straw Returning on Soil Enzyme Activity and Microbial Community Structure and Diversity. Agriculture. 2025; 15(1):56. https://doi.org/10.3390/agriculture15010056
Перевод статьи «Effects of Different Tillage Measures Combined with Straw Returning on Soil Enzyme Activity and Microbial Community Structure and Diversity» авторов Xiao S, Li B, Zhang T, Luo J, Wang J, Zhang X, Li J, Zhang D., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык

















































































































Комментарии (0)