Биотехнологический подход к разработке и характеристике белкового корма для пчел Melipona quadrifasciata
Бесжальные пчелы являются важнейшими опылителями в тропических и неотропических регионах, что подчеркивает необходимость сохранения этих видов. Однако дефицит ресурсов, особенно пыльцы, в определенные периоды негативно сказывается на здоровье пчел и эффективности опыления. Для решения этой проблемы мы разработали ферментированный белковый корм, используя микроорганизмы из пыльцы Melipona quadrifasciata — вида, широко распространенного в бразильском атлантическом лесу. Ферментированный корм представлял собой смесь белковых отрубей, сахарного сиропа и инокулянта, полученного из ферментированной пыльцы этого вида.
Для оценки качества корма были проведены питательный анализ и метаболомные исследования (методами UHPLC/DAD-HRMS/MS и GC-MS). Метаболомный анализ выявил 85 соединений в режиме положительной и 102 в режиме отрицательной ионизации. Общие метаболиты, такие как изокверцетин и палатиноза, подчеркнули сходство между кормом и естественной ферментированной пыльцой.
Лабораторные испытания на пчелах в садках показали сопоставимое потребление корма и ферментированной пыльцы без статистически значимой разницы (в среднем: 0,067 г против 0,060 г; p = 0,53). Корм продемонстрировал благоприятный переход pH (с 5,30 до ферментации до 4,41 после процесса ферментации) и оставался стабильным в течение шести месяцев при температуре ±2 °C.
Эти результаты демонстрируют биотехнологический прогресс в питании бесжальных пчел, предлагая жизнеспособное решение в период дефицита ресурсов и способствуя укреплению здоровья M. quadrifasciata.
1. Введение
В тропиках, в основном в неотропическом регионе, обитает более 605 видов безжальных пчел [ 1 ]. Эта группа играет важную роль в предоставлении экологических услуг, особенно опыления, и имеет преимущества перед другими видами пчел благодаря своей сильной адаптации к разнообразной флоре, встречающейся на всей территории их обитания [ 2 ]. Жизнь человека на Земле в значительной степени зависит от экосистемных услуг, таких как опыление [ 3 , 4 ]. Хотя безжальные пчелы традиционно известны производством меда и пыльцы, их роль в экосистемных услугах все больше признается, что способствует их использованию в устойчивых практиках, таких как управляемое опыление в сельском хозяйстве. Это делает пчеловодство с безжальными пчелами перспективным и экономически выгодным направлением [ 5 ].
Одна из ключевых стратегий борьбы с сокращением численности пчел — сохранение мест обитания опылителей [ 6 ], прежде всего для предотвращения пищевого стресса, вызванного нехваткой пищевых ресурсов. Нектар и цветочная пыльца, основные источники питательных веществ для пчел, обеспечивают их сахарами и белками соответственно. Перед употреблением безжальными пчелами они подвергаются процессам трансформации, таким как ферментация [ 7 ].
Помимо сохранения среды обитания, еще одной стратегией минимизации потерь пчелиных семей в периоды дефицита питательных веществ является разработка дополнительных кормов, особенно богатых белком. Добавление пыльцы, собранной с пчел Apis mellifera, в периоды дефицита пчел сопряжено со значительными рисками, поскольку такая пыльца может передавать такие заболевания, как европейский гнилец, вызываемый бактерией Melissococcus plutonius , что приводит к гибели расплода и потере пчелиных семей [ 8 ]. Кроме того, пыльца служит основным путем передачи вирусов между таксонами внутри сообществ опылителей, при этом РНК-вирусы в пыльце медоносных пчел распространяются на виды, не относящиеся к Apis , потенциально влияя на здоровье экосистемы и динамику заболеваний пчел [ 9 ].
Разработка специальной белковой добавки для безжальных пчел представляет собой ряд трудностей, поскольку многие рационы либо не принимаются пчелами, либо имеют низкую питательную ценность [ 10 ]. Исследования подчеркивают важность ферментированных белковых рационов для пчел, поскольку чем ближе рацион имитирует естественно ферментированную пыльцу, тем привлекательнее и питательнее он становится для них [ 11 ].
У Apis mellifera некоторые молочнокислые бактерии, такие как Lactobacillus kunkeei и Fructobacillus fructosus , играют активную роль в ферментации пыльцы внутри колонии, превращая сырую пыльцу (неферментированную) в пчелиный хлеб (ферментированный), который более обогащен питательными веществами [ 12 , 13 ]. В случае безжальных пчел существует значительный пробел в знаниях относительно микроорганизмов, участвующих в процессах в колонии, особенно ферментаторов, ответственных за трансформацию пыльцы [ 14 ]. Некоторые исследования сообщают о наличии бактерий рода Bacillus в пыльце безжальных пчел [ 15 ].
Помимо питательных и ферментативных аспектов корма, еще одним важным параметром является химический состав добавки после ферментации по сравнению с ферментированной пыльцой, обнаруженной в колониях M. quadrifasciata . Химический состав белковой добавки имеет решающее значение как для питательной ценности, так и для привлекательности, поскольку как полярные, так и неполярные летучие соединения в пыльце способствуют ее привлекательности во время кормления пчел [ 10 ]. Поэтому рационы, разработанные для имитации ферментированной пыльцы, также должны соответствовать ее химическому составу [ 16 ].
Целью данного исследования было разработать ферментированный белковый корм для M. quadrifasciata с использованием микроорганизмов из пыльцы, потребляемой самим видом пчел. Разработанный корм сравнивали с ферментированной пыльцой по содержанию сырого белка и химическому составу, а также предлагали пчелам, содержащимся в лабораторных клетках, для оценки потребления и, следовательно, вкусовых качеств ферментированного корма. Это исследование не только предлагает жизнеспособную альтернативу кормлению безжальных пчел, но и представляет собой значительный шаг вперед в изучении ферментированных кормов для пчел, способствуя совершенствованию технологий пчеловодства и повышению устойчивости и эффективности управления популяциями местных пчел.
2. Материалы и методы
2.1. Подготовка инокулята и разработка кормовой смеси
Кормовая добавка была произведена в марте 2023 года в Лаборатории прикладной микробиологии Координационного совета по лесной фауне штата Сан-Паулу. Для этой цели были проведены две ферментации: одна для получения инокулята ( рис. 1 А), а другая для разработки корма ( рис. 1 В). Закрытые емкости с ферментированной пыльцой, известной как «сабура», были собраны с колоний M. quadrifasciata в Meliponário Paraitinga, расположенном в Нативидаде-да-Серра, штат Сан-Паулу, Бразилия (S 23°19′26.400″ W 45°29′6.000″). Для обеспечения асептических условий во время сбора все материалы были предварительно стерилизованы автоклавированием при 121 °C в течение 20 мин. Закрытые контейнеры с пыльцой были доставлены в лабораторию, открыты в ламинарном боксе с помощью одноразовой петли и перенесены в стерильный контейнер для взвешивания. Затем в колбу Эрленмейера добавили 5 г ферментированной пыльцы и 150 мл раствора сахарозы и воды в соотношении 1:1 ( об / об ), предварительно прокипяченного и хранившегося в стерильном контейнере. Важно отметить, что использовалась только пыльца пастообразной консистенции, указывающая на ее зрелость в процессе естественной ферментации внутри колоний M. quadrifasciata . Затем смесь пыльцы и сиропа инкубировали в орбитальном шейкере (Tecnal, Бразилия) при 30 °C и 120 об/мин и накрывали предварительно стерилизованной ватной пробкой на шесть дней ( рис. 1 А). Эта процедура была выполнена в двух повторах для обеспечения воспроизводимости результатов. После инкубационного периода полученный раствор, называемый инокулятом, подвергался измерению pH с помощью цифрового pH-метра (Hanna Instruments pH21, Румыния) для оценки кислотности. Затем инокулят хранился в виде консорциума ферментирующих микроорганизмов из пчелиной колонии безжальных пчел для последующей ферментации корма.
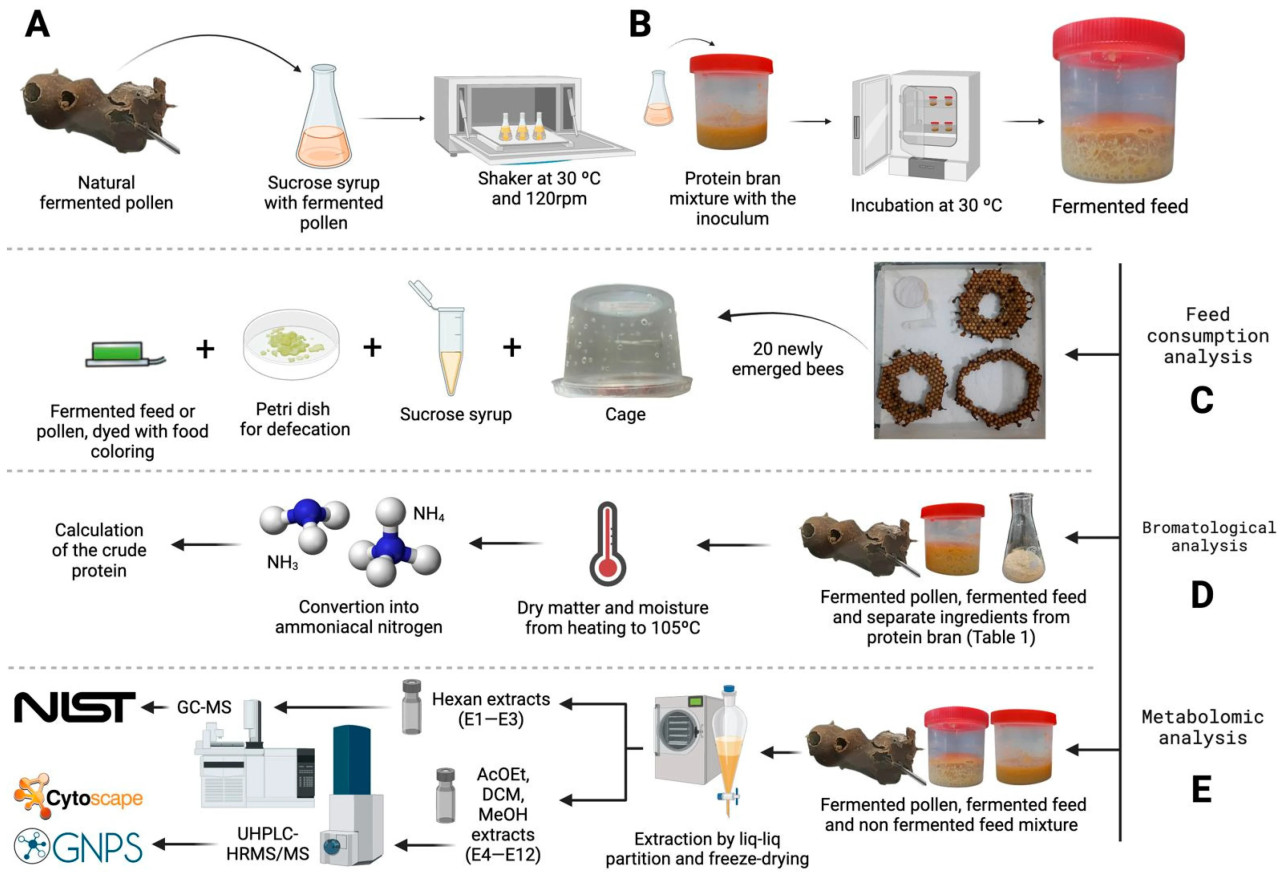
Рисунок 1. ( A ) Разработка инокулята для ферментации белковой муки, начатая со сбора естественно ферментированной пыльцы M. quadrifasciata ; ( B ) приготовленный инокулят, смешанный с белковой мукой и инкубированный в контролируемых условиях температуры и влажности, в результате чего был получен ферментированный корм, используемый в последующих анализах потребления; ( C ) диски расплода до вылупления, собранные из колоний M. quadrifasciata и использованные для проведения тестов на потребление в контролируемых клетках. После вылупления 20 рабочих пчел были переведены в контролируемые клетки, где им предоставляли сахарозный сироп и белковый корм с цветовой кодировкой в зависимости от их группы (ферментированный корм или ферментированная пыльца), а также чашку Петри для дефекации; ( D ) броматологический анализ проводился путем экстракции сухого вещества при 105 °C с последующим преобразованием азота из образца в аммонийный азот для расчета содержания сырого белка; ( E ) Метаболомный анализ проводился путем экстракции образцов различными органическими растворителями и введения экстрактов в системы ГХ-МС и ВЭЖХ-ВРМС/МС для анализа с использованием заданных библиотек.
2.2. Ферментация корма
В качестве белковой основы для ферментированного корма использовались ингредиенты белковых отрубей, перечисленные в Таблице 1 , рецептура которых была адаптирована по Пайве (2018) [ 14 ]. Перед ферментацией эти ингредиенты облучали в течение 3 дней на расстоянии 15 см от центра источника кобальта-60 для уничтожения внешних микроорганизмов и обеспечения стерильности белковой основы. После стерилизации ингредиенты взвешивали в соответствии с рецептурой корма и переносили в чашки Петри размером 90 × 15 мм, где они подвергались воздействию УФ-излучения в течение 2 минут в ламинарном боксе для предотвращения дополнительного загрязнения.
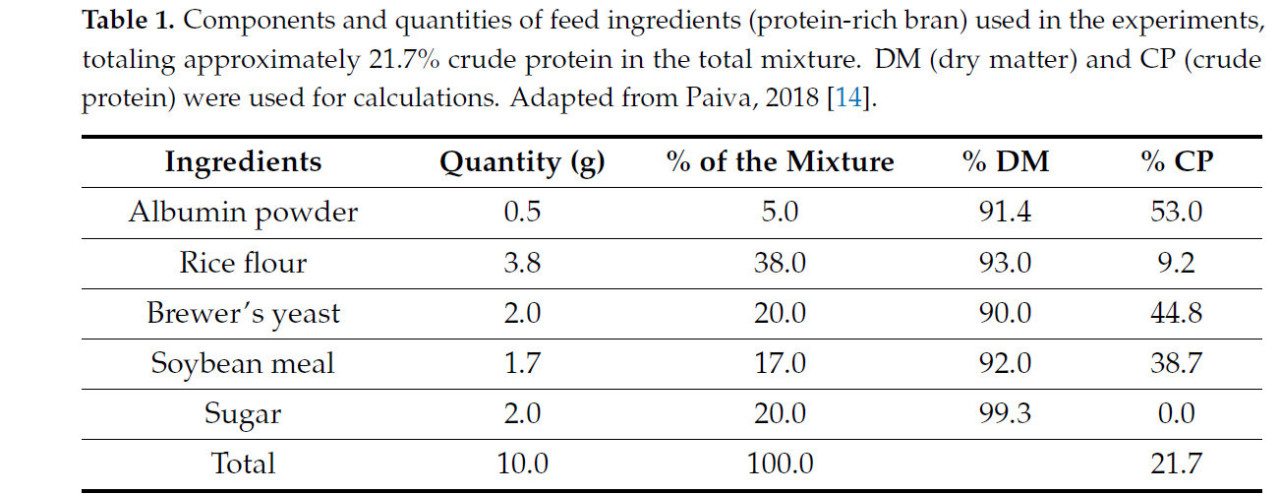
Таблица 1. Компоненты и количество кормовых ингредиентов (богатые белком отруби), использованных в экспериментах, в общей сложности приблизительно 21,7% сырого протеина в общей смеси. Для расчетов использовались СВ (сухое вещество) и СП (сырой протеин). Адаптировано из Пайва, 2018 [ 14 ].
Затем белковые отруби переносили в стерильный контейнер, куда добавляли 5 мл предварительно приготовленного инокулята и 5 мл сахарозного сиропа в соотношении 1:1 ( об / об ) на каждые 10 г смеси, оба компонента при комнатной температуре. Смесь гомогенизировали автоклавированным шпателем в ламинарном боксе до достижения однородной пастообразной консистенции. Гомогенизированный продукт затем переносили в стерильные контейнеры объемом 50 мл с полузавинчивающимися крышками и инкубировали при 30 °C в течение шести дней ( рис. 1 B). pH корма измеряли до и после инкубации с помощью цифрового pH-метра, разбавляя 1 г корма в 10 мл дистиллированной воды. После периода инкубации ферментированный корм хранили в холодильнике при температуре приблизительно ±4 °C. Весь процесс приготовления корма проводили в трех повторениях.
2.3. Потребление корма в контролируемых пчелиных клетках
Для оценки привлекательности и потребления ферментированного корма были проведены контролируемые эксперименты в клетках с только что вылупившимися пчелами M. quadrifasciata . Предвылупные расплодные диски были собраны из пяти разных семей и перенесены в контейнер («питомник») внутри инкубатора BOD, установленного на 28 °C. В период вылупления пчел-кормилиц собирали из семей, маркировали нетоксичными чернилами и помещали в питомник, чтобы помочь распечатать расплодные диски и облегчить вылупление рабочих пчел. Для поддержания влажности в инкубаторе на уровне 80% добавляли емкости с водой, влажность контролировали с помощью цифрового термогигрометра. Для обеспечения энергией только что вылупившихся пчел по мере необходимости предоставляли пробирку Эппендорфа с сахарозным сиропом в соотношении 1:1 ( об / об ), а также чашку Петри размером 35 × 10 мм, выстланную влажной бумагой, для использования рабочими пчелами в качестве места для дефекации.
Рабочие особи, появившиеся в течение 24 часов, были собраны вручную, подсчитаны (N = 20) и помещены в пластиковые клетки размером 120 × 78 мм (объемом 500 мл), как показано на рисунке 1 С. Клетки были предварительно вымыты моющим средством, продезинфицированы 70% спиртом и содержались в том же инкубаторе БПК в тех же условиях, что и питомник. Клетки были разделены на две группы: экспериментальная (ферментированный корм) и положительная контрольная (ферментированная пыльца). Обе группы получали сироп в соотношении 1:1 ( об / об ) в неограниченном количестве и чашки Петри размером 35 × 10 мм с влажной бумагой, которую меняли ежедневно. В экспериментальной группе рабочие пчелы получали приблизительно 0,145 г ферментированного корма, окрашенного зеленым пищевым красителем, тогда как в группе положительного контроля пчелы получали приблизительно 0,145 г ферментированной пыльцы, также окрашенной тем же красителем и полученной из закрытых пыльцевых горшков колоний M. quadrifasciata .
Для каждой группы (экспериментальной и контрольной) было установлено пять клеток, которые содержались в инкубаторе в течение восьми дней подряд. В течение этого периода клетки осматривались ежедневно, мертвых пчел удаляли и регистрировали. Белковый корм в обеих группах заменяли на второй, четвертый и шестой дни эксперимента, чтобы измерить потребление путем взвешивания остатков в кормушках с помощью цифровых весов (рассчитывая разницу между предложенным количеством и количеством, потребленным пчелами за два дня), а также для предотвращения высыхания или загрязнения корма. Кроме того, чашки Петри в каждой клетке проверяли на наличие следов зеленого красителя в помете пчел, чтобы контролировать потребление белкового корма как в экспериментальной, так и в контрольной группах. Методы, использованные для тестов на потребление, показаны на рисунке 1 С.
2.4. Броматологический анализ
Броматологические анализы проводились в Многопрофильной лаборатории питания животных и броматологии Университета Сан-Паулу (Факультет ветеринарной медицины и животноводства — кампус Пирассунунга). Анализы проводились (по одному образцу каждого) ферментированного корма, ферментированной пыльцы и отдельных ингредиентов, входящих в состав корма ( Таблица 1 ).
Для проведения этих анализов использовались стандартные рабочие процедуры [ 17 , 18 ]. Содержание сухого вещества и влаги в образцах определяли путем нагревания до 105 °C. На основе содержания сухого вещества в каждом образце определяли содержание сырого белка путем преобразования органического азота в аммонийный азот ( рис. 1 D).
2.5. Экстракция метаболитов
Для проверки молекулярного состава корма и сравнения его химического профиля с другими компонентами его рецептуры были использованы классические методы экстракции природных продуктов. Метаболиты, присутствующие в исследуемых образцах (ферментированная пыльца, ферментированный корм и неферментированный корм), были обнаружены с помощью процедур экстракции с использованием различных органических растворителей, охватывающих диапазоны от низкой до высокой полярности. Пять органических экстрактов, полученных для каждого образца, перечислены в таблице 2 .
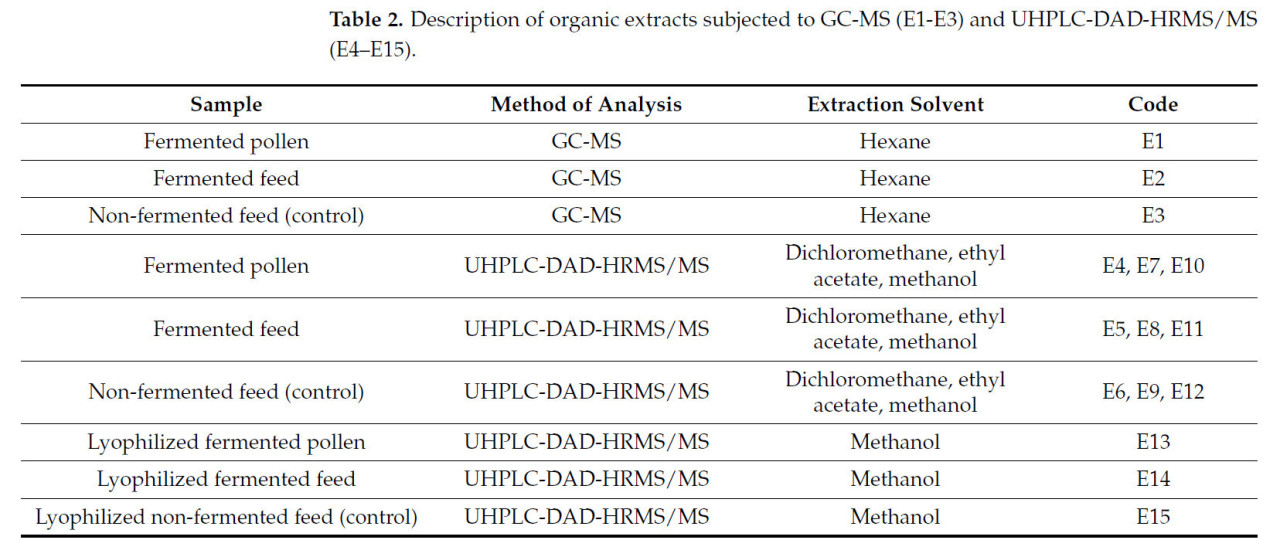
Таблица 2. Описание органических экстрактов, подвергнутых ГХ-МС (E1-E3) и УВЭЖХ-ДАД-ВРМС/МС (E4–E15).
Экстракты E1–E12 были получены методом жидкостно-жидкостной экстракции. В частности, 3,0 г каждого образца растворяли в сверхчистой воде и метаноле (1:9 об. / об. ), затем проводили экстракцию гексаном, а затем дихлорметаном, получая экстракты E1–E3 и E4–E6 соответственно. В результате была получена водно-спиртовая фаза E10–E12. Экстракты E7–E9 были получены из 3,0 г каждого образца, растворенного в 100 мл сверхчистой воды, с последующей экстракцией этилацетатом. Органические фазы высушивали с помощью роторного испарителя, а водно-спиртовые фазы — с помощью вакуумной сушильной системы.
Кроме того, 1,0 г каждого образца лиофилизировали, а затем экстрагировали примерно 1–2 мл метанола в течение 1 часа в ультразвуковой ванне. После центрифугирования при 13 000 об/мин надосадочную жидкость (E13–E15) собирали и высушивали.
Гексановые экстракты (E1–E3) растворяли в гексане для ВЭЖХ и вводили в систему ГХ-МС. Оставшиеся высушенные экстракты (E4–E15) повторно растворяли в метаноле для ВЭЖХ и вводили в систему УВЭЖХ-ДАД-ВРМС/МС. Перед анализом все образцы центрифугировали при 13 000 об/мин и анализировали при концентрации 200 мкг. Метод экстракции метаболитов представлен на рисунке 1 E. Каждый образец анализировали в трех повторениях.
2.6. Анализ метаболомных данных и сравнение химического состава.
Данные ГХ-МС были получены с использованием системы MS-QP2010 (Shimadzu, Киото, Япония) с электронной ионизацией при 70 эВ и введением образца в режиме разделения потока при 250 °C. Спектры полного сканирования регистрировались в диапазоне m / z 40–500 с интервалом сканирования 0,5 с. В качестве стационарной фазы использовалась хроматографическая колонка Rtx-5MS (30 м × 0,25 мм × 0,25 мкм, Agilent J&W GC Columns, Санта-Клара, Калифорния, США), поддерживаемая при температуре 80 °C, а подвижная фаза состояла из гелия под давлением 75,0 кПа с общей скоростью потока 60,2 мл/мин. Метод включал в себя повышение температуры от 80 °C до 250 °C в течение 0–10 мин, за которым следовал изократический режим в течение 10–50 мин при 250 °C.
Данные UHPLC-DAD-HRMS/MS были получены с использованием системы Nexera X2 (Shimadzu, Япония), оснащенной диодным детектором SPD M20A, соединенным с MicroTOF-QII (Bruker Daltonics, Биллерика, Массачусетс, США) с источником электрораспылительной ионизации (ESI), работающим как в положительном, так и в отрицательном режимах ионизации. Для хроматографического разделения в обращенной фазе использовалась аналитическая колонка Kinetex C18 (100 × 2,1 мм, 2,6 мкм, Phenomenex, Торранс, Калифорния, США) при температуре 50 °C и скорости потока 0,35 мл/мин. Подвижная фаза (A) состояла из деионизированной воды, а фаза (B) — из ацетонитрила, оба реактива хроматографической чистоты, с добавкой 20 мМ муравьиной кислоты. Метод включал градиент от 0 до 2 мин (15% B), от 2 до 12 мин (15–95% B) и от 12 до 17 мин (95% B). Для каждого анализа вводили образцы объемом 5 мкл.
Для масс-спектрометрического анализа в режиме AutoMS (DDA) был выбран диапазон масс m / z 50–1200, энергия столкновения варьировалась от 20 до 65 эВ для m / z 50–700 и фиксировалась на уровне 65 эВ для значений выше m / z 700. За цикл регистрировалось максимум пять ионов-предшественников. Рабочие параметры были следующими: напряжение капилляра 4500 В, газ распылителя (азот) 4 бар, осушающий газ (азот) 9,0 л/мин, температура источника 200 °C. Внутренняя калибровка проводилась с использованием 10 нМ раствора формиата натрия в изопропаноле/воде (1:1 об. / об. ).
Для молекулярной дедупликации аннотации химической структуры данных ГХ-МС были выполнены с использованием библиотеки NIST08.LIB, доступной на оборудовании. Для данных ЖХ-МС/МС получение молекулярных аннотаций осуществлялось в соответствии с методологией, описанной Де Карвалью и др. [ 19 ]. Процедура аннотирования проводилась на основе критериев, установленных Инициативой стандартов метаболомики (MSI) для уровней аннотации соединений [ 20 , 21 ], в сочетании с более специфичной системой для анализа на основе ВРМС [ 22 ].
2.7. Статистический анализ результатов теста на потребление корма
Сравнение потребления ферментированного корма и ферментированной пыльцы проводилось с использованием теста Шапиро-Уилка для оценки нормальности распределения независимых выборок на основе значения p ; данные с p > 0,05 считались нормально распределенными. Для оценки различий между двумя группами использовался парный t -тест, при этом p > 0,05 указывало на отсутствие статистически значимых различий. Распределение данных, включая медиану и стандартное отклонение, визуализировалось с помощью диаграммы размаха скрипичного типа. Все статистические анализы проводились с использованием программного обеспечения R (версия 4.3.2).
3. Результаты
3.1. Разработка кормовой базы
После периода ферментации корм сохранял однородный цвет, без видимых нитевидных грибов или бактериальных колоний, которые могли бы указывать на загрязнение. Внешний вид корма до и после процесса ферментации показан на рисунке 2. Результаты измерений pH образцов представлены в таблице 3. Образцы корма хранились в течение шести месяцев в холодильнике (±2 °C) и оставались стабильными, сохраняя однородный внешний вид.
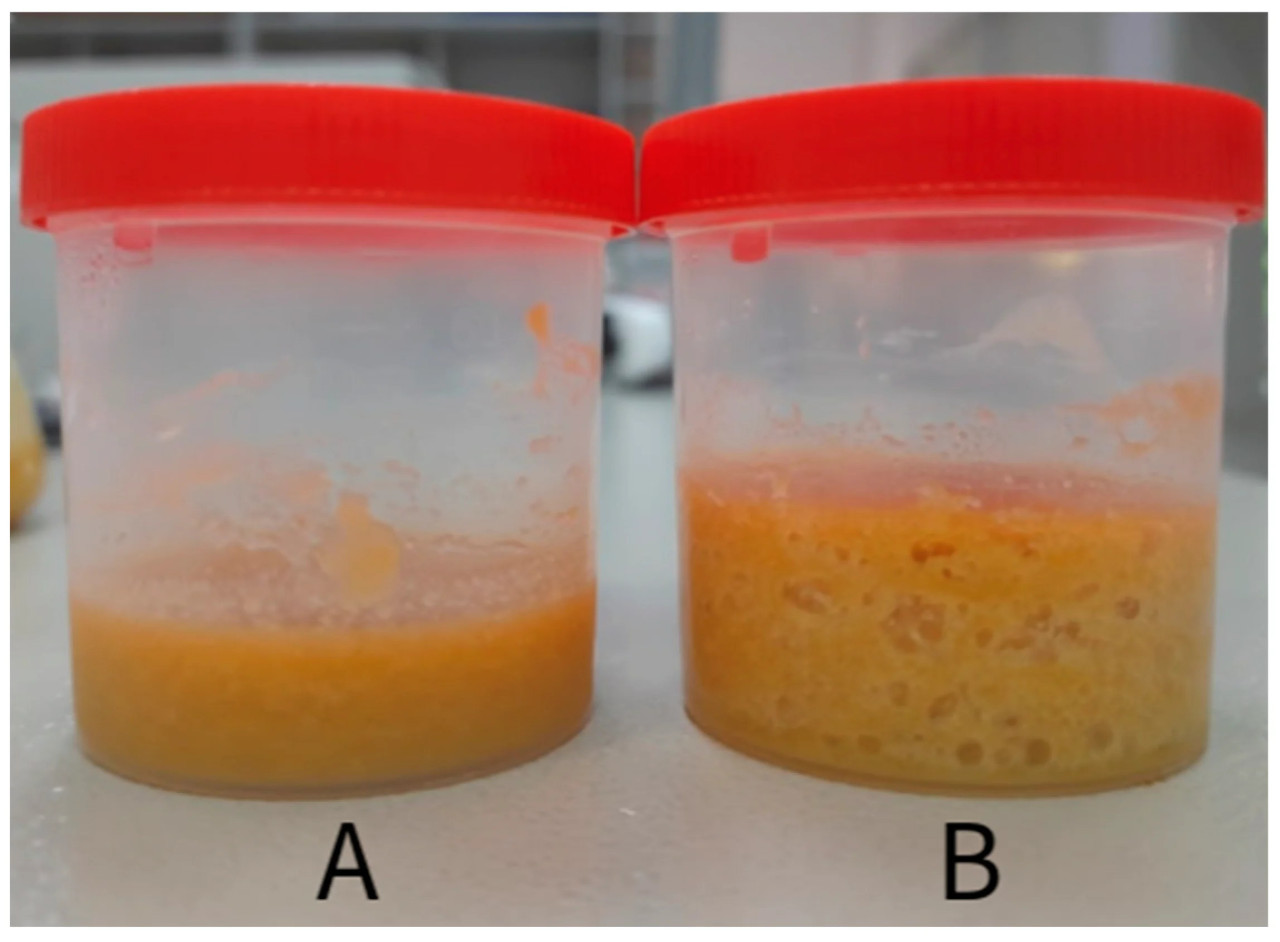
Рисунок 2. ( А ) Смесь белковых отрубей с инокулятом до инкубации для процесса ферментации; ( Б ) ферментированная смесь после инкубации, с выделением газа, указывающим на протекание ферментации.
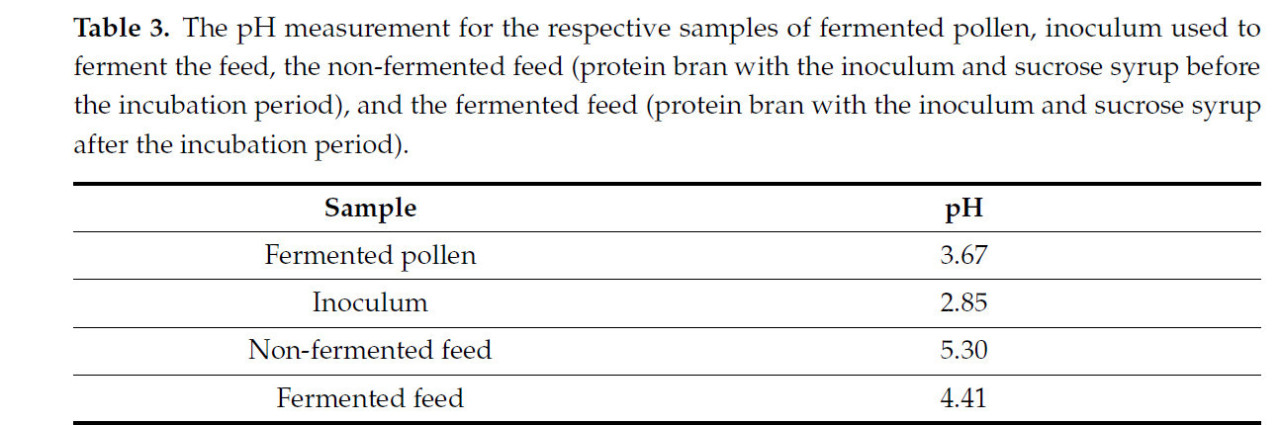
Таблица 3. Результаты измерения pH для соответствующих образцов ферментированной пыльцы, инокулята, использованного для ферментации корма, неферментированного корма (белковые отруби с инокулятом и сахарозным сиропом до инкубационного периода) и ферментированного корма (белковые отруби с инокулятом и сахарозным сиропом после инкубационного периода).
3.2. Броматологический анализ
Анализ питательной ценности ферментированного корма, ферментированной пыльцы и белковых отрубей ( таблица 1 ) проводился с помощью броматологического анализа для количественного определения сырого протеина в процентах от сухого вещества, что дало значения 15,93%, 45,32% и 21,7% соответственно.
3.3. Потребление корма в контролируемых пчелиных клетках
Кривая нормальности показала, что данные о потреблении соответствуют нормальному распределению (тест Шапиро-Уилка, p = 0,21). Статистически значимой разницы между группами, получавшими ферментированный корм (0,060 г) и ферментированную пыльцу (0,067 г), не было (парный t -тест, p = 0,53), что свидетельствует о схожем потреблении ферментированного корма и ферментированной пыльцы. Хотя медианы между вариантами обработки были сопоставимы, межквартильный размах группы, получавшей ферментированную пыльцу, был больше, чем у группы, получавшей ферментированный корм ( рисунок 3 ). В обоих вариантах обработки третий квартиль (Q3) был выше.
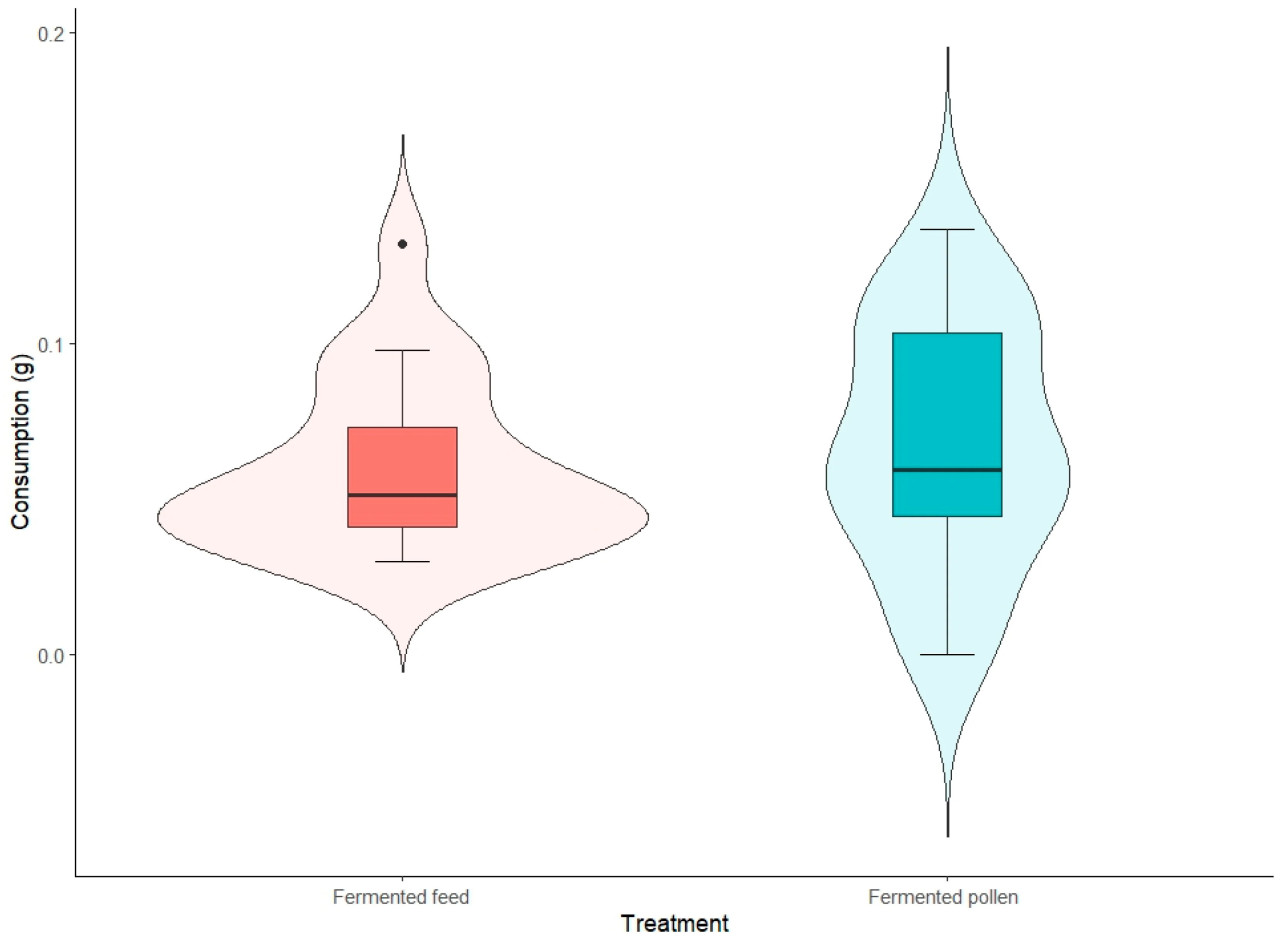
Рисунок 3. Распределение данных из каждой клетки в эксперименте по потреблению. Для каждой клетки в этом анализе использовалась сумма потребления за восемь дней эксперимента. Скрипичная диаграмма (на заднем плане) показывает плотность распределения данных, где более широкие области указывают на более высокую концентрацию значений, как это видно в образце ферментированного корма. Диаграмма размаха показывает медиану (центральная линия), которая одинакова для обоих образцов, первый и третий квартили (границы диаграммы размаха) и экстремальные значения (усы диаграммы размаха). Только образец ферментированного корма показал выброс. Значимой статистической разницы ( p = 0,53) между двумя образцами обнаружено не было.
3.4. Анализ данных GC-MS и UHPLC/DAD-HRMS/MS
Экстракция ферментированного корма и ферментированной пыльцы с использованием различных органических растворителей позволила проанализировать их химический состав с помощью ГХ-МС и ВЭЖХ/ДАД-ВРМС/МС, полученные хроматограммы представлены на рисунках 4 и 5 соответственно. В обоих случаях некоторые соединения были обнаружены только в одном из образцов, например, строфантидол, который был обнаружен в экстракте ферментированной пыльцы E1, но отсутствовал в экстракте ферментированного корма E2 ( рисунок 4 ) (одинаковый уровень полярности для обоих экстрактов). Аналогично, генистин был идентифицирован только в экстракте E11 ( рисунок 5 ). Основное различие между метаболитами ферментированного корма и пыльцы, по-видимому, заключается в относительной интенсивности соединений, присутствующих в каждом образце ( рисунок 4 и рисунок 5 ). Это химическое сходство дополнительно подтверждается соответствием времен удерживания и химических профилей обнаруженных метаболитов.
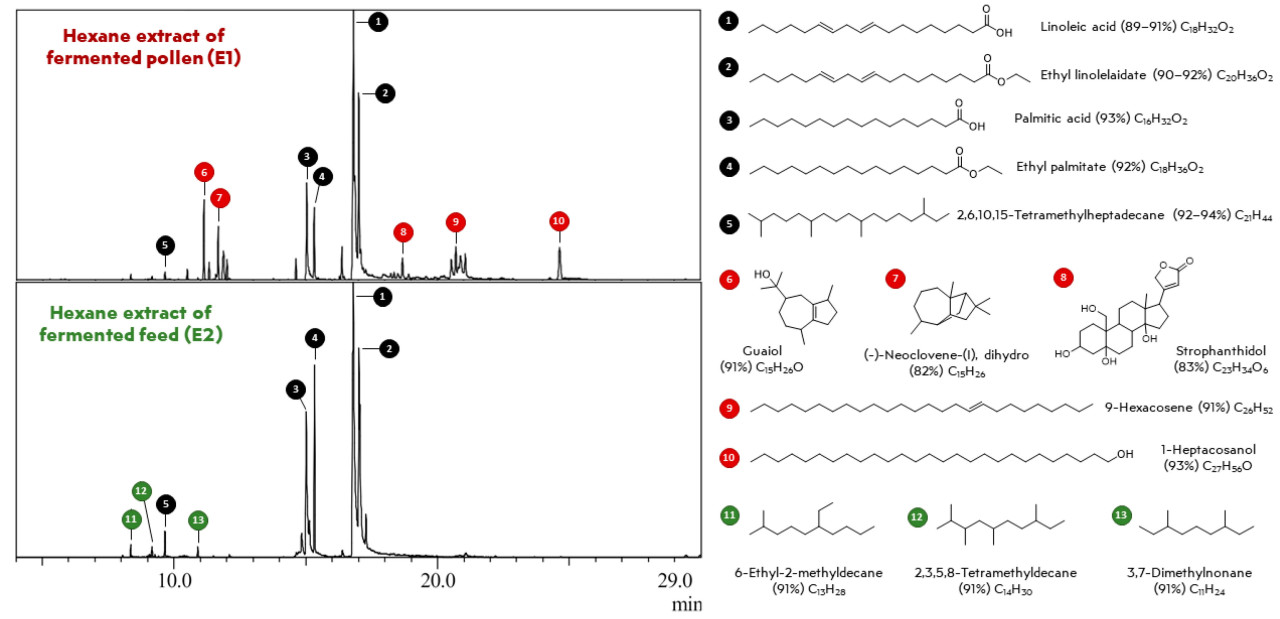
Рисунок 4. Хроматограммы ГХ-МС химических экстрактов ферментированной пыльцы и образцов корма, полученных методом жидкостно-жидкостной экстракции гексаном (E1 и E2). Красный, зеленый и черный цвета представляют соединения, идентифицированные в образцах ферментированной пыльцы, ферментированного корма или в обоих случаях, соответственно. Сбоку приведены химические структуры, предложенные библиотекой NIST08.LIB, с указанием процента сходства в скобках, основанного на профилях фрагментации.
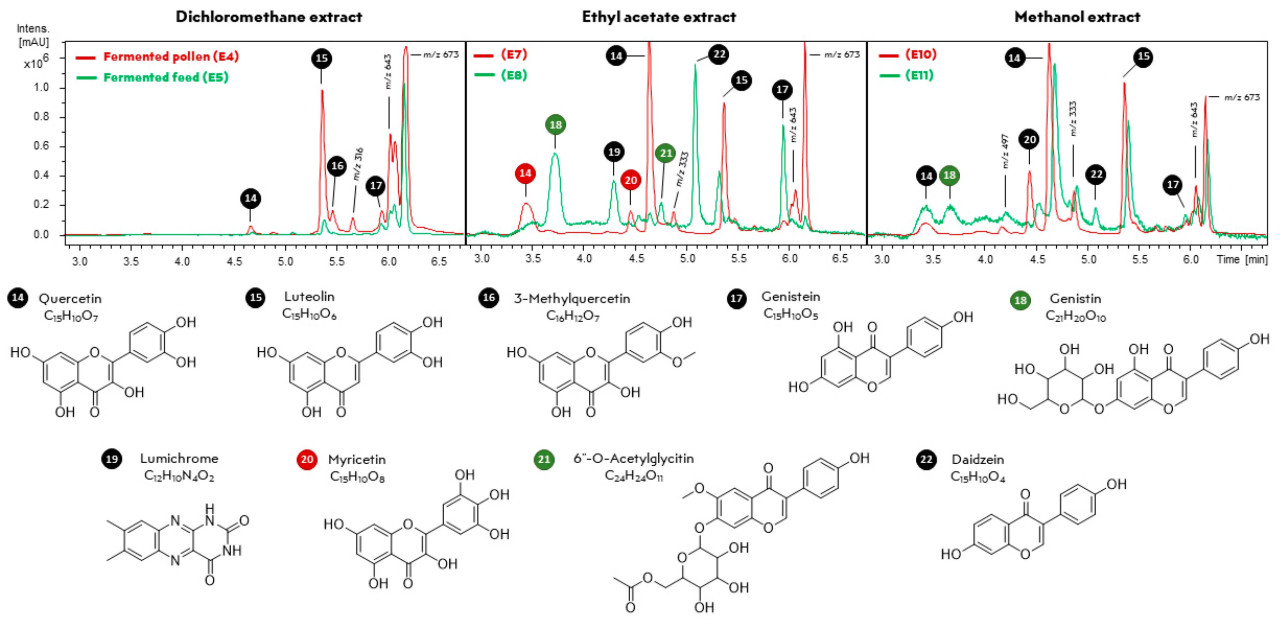
Рисунок 5. Хроматограммы UHPLC-DAD экстрактов ферментированной пыльцы и образцов корма, полученные методом жидкостно-жидкостной экстракции с использованием дихлорметана (E4 и E5), этилацетата (E7 и E8) и метанола (E10 и E11). Красный, зеленый и черный цвета представляют соединения, идентифицированные в образцах ферментированной пыльцы, ферментированного корма или в обоих случаях, соответственно. Хроматограммы E13 и E14 здесь не показаны из-за их сходства с E10 и E11, соответственно. Ниже приведены аннотации химических структур, основанные на совпадениях в библиотеке GNPS.
В соответствии с хроматографическими данными, автоматическая химическая дерепликация на основе данных MS/MS-спектрометрии подтвердила химическое сходство между ферментированным кормом и пыльцой. Молекулярные сети, полученные при положительной (ESI+) и отрицательной (ESI−) ионизации, представлены на рисунках 6 и 7 соответственно, обеспечивая визуальное представление кластеризации соединений, присущих образцам, с акцентом на более крупные кластеры. В режиме положительной ионизации было обнаружено в общей сложности 85 химических соединений, а в режиме отрицательной ионизации было предложено 102 метаболита на основе аннотации уровня 2 [ 21 , 22 ].
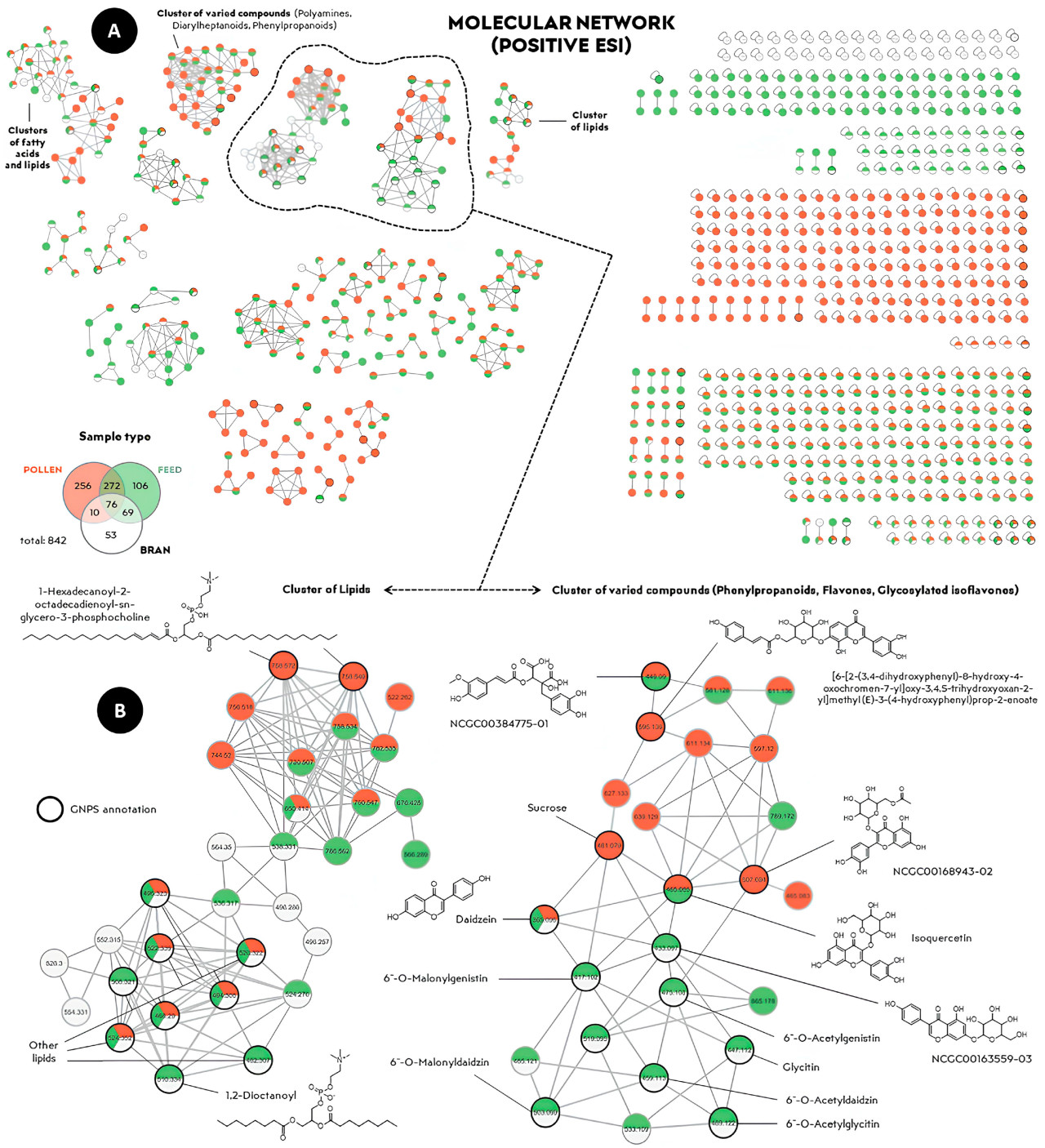
Рисунок 6. ( A ) Молекулярная сеть положительной ионизации органических экстрактов из образцов пыльцы (красным цветом), ферментированного корма (зеленым цветом) и смеси отрубей с сахарозным сиропом (белым цветом), сгенерированная с использованием параметров по умолчанию платформы GNPS. ( B ) Увеличенные кластеры семейств, выделенные на рисунке ( A ). Наличие определенного цвета указывает на обнаружение признака в соответствующем экстракте (типе образца).
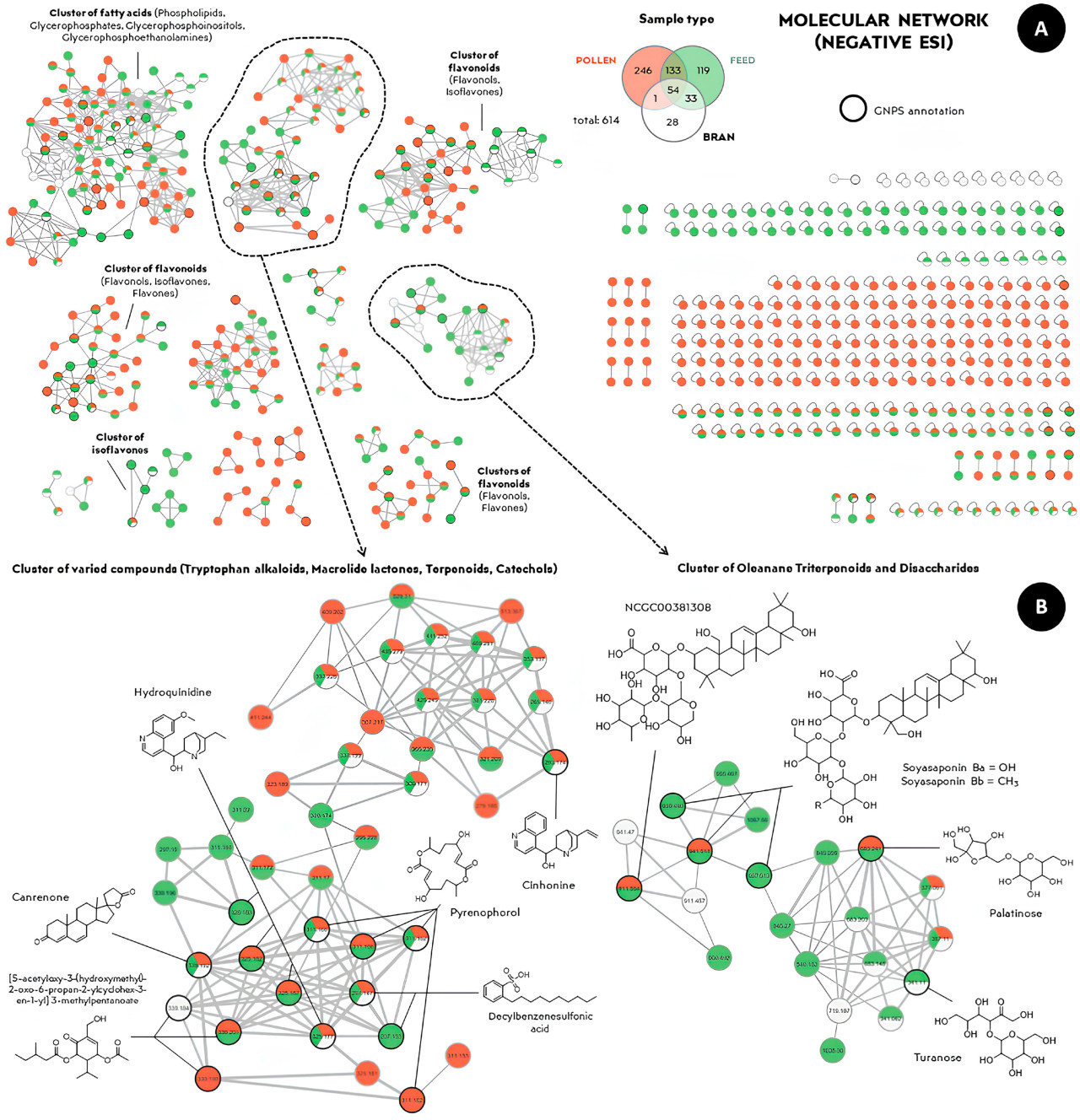
Рисунок 7. ( A ) Молекулярная сеть отрицательной ионизации органических экстрактов из образцов пыльцы (красным цветом), ферментированного корма (зеленым цветом) и смеси отрубей с сахарозным сиропом (белым цветом), сгенерированная с использованием параметров по умолчанию платформы GNPS. ( B ) Увеличенные кластеры семейств, выделенные на рисунке ( A ). Наличие определенного цвета указывает на обнаружение признака в соответствующем экстракте.
Были аннотированы молекулы различных структур растительного происхождения, в частности, из химических классов фенилпропаноидов (таких как флавоноиды) и терпеноидов (таких как сесквитерпены, дитерпены и тритерпены). Кроме того, были обнаружены химические соединения, присутствующие в обеих группах, такие как изокверцетин ( рис. 6 B) и палатиноза ( рис. 7 B), что еще раз подтверждает сходство химических профилей ферментированного корма и пыльцы. Наибольшие кластеры в основном включали липиды, жирные кислоты, флавонолы и флавоны, которые являются метаболитами, общими как для корма, так и для пыльцы ( рис. 6 и рис. 7 ). Данные о ежедневном потреблении ферментированного корма и ферментированной пыльцы приведены в дополнительных материалах для справки.
4. Обсуждение
Добавление ферментированного искусственного белка является жизнеспособной альтернативой для питания пчел, обеспечивая необходимые аминокислоты и другие питательные вещества в пропорциях, аналогичных естественно ферментированной пыльце, тем самым поддерживая здоровье и продуктивность колонии [ 23 , 24 ]. На данном этапе разработанный корм продемонстрировал показатели потребления, сопоставимые с показателями естественно ферментированной пыльцы, что позволяет предположить, что добавление в периоды дефицита является жизнеспособной стратегией. В дальнейшем следующие шаги включают идентификацию и количественное определение аминокислот, а также проведение биологических полевых испытаний для оценки развития колонии.
Кроме того, ферментация искусственной добавки повышает ее усвояемость и увеличивает биодоступность питательных веществ, имитируя естественный процесс хранения пыльцы в пчелиных колониях [ 25 ]. Поэтому крайне важно, чтобы добавка была разработана таким образом, чтобы питать пчел в критические периоды нехватки природных ресурсов, обладая при этом привлекательными для них характеристиками [ 16 ]. Корм, разработанный в этом исследовании, был создан с учетом этой проблемы, с использованием специфических микробных инокулянтов для обеспечения его привлекательности для пчел. pH ферментированной пыльцы безжальных пчел значительно варьируется: низкие значения, такие как 3,70, наблюдаются у Melipona seminigra [ 26 ], от 3,28 до 3,99 у Melipona scutellaris [ 27 ], а более высокие значения, такие как 5,5 у Melipona asilvai и 5,0 у M. quadrifasciata [ 28 ].
Важно подчеркнуть, что процесс ферментации пыльцы безжальных пчел остается малоизученным, что указывает на необходимость исследований, направленных на стандартизацию сбора и анализа пыльцы с учетом таких факторов, как сезон, географический регион и, в частности, стадия ферментации и созревания. Как отмечают Миранда-Пинто и др. [ 29 ], поведение пыльцы при хранении у M. quadrifasciata не следует какой-либо закономерности в отношении количества дней, в течение которых этот ресурс хранится в горшках до употребления, что указывает на различные стадии ферментации в разных горшках с пыльцой.
Наблюдаемое значение pH в ферментированном корме по сравнению с ферментированной пыльцой является многообещающим, поскольку оно указывает на то, что процесс ферментации с использованием инокулянтов из самого вида эффективен в обеспечении необходимой кислотности свежего корма, снижая pH, подобно естественно ферментированной пыльце. Этот процесс имеет решающее значение для привлекательности добавки, поскольку он не только изменяет текстуру, но и высвобождает ароматы и молочную кислоту [ 16 ]. Хотя процесс ферментации у безжальных пчел недостаточно хорошо описан, наблюдаемая кислотность предполагает наличие микроорганизмов, продуцирующих молочную кислоту, таких как бактерии и дрожжи [ 13 ]. Таким образом, наличие ферментативных микроорганизмов, способствующих высвобождению летучих соединений после ферментации, необходимо для привлечения рабочих пчел к дополнительному белковому корму и стимулирования его потребления.
Измерение сырого протеина, представляющего собой общее содержание белка в образце пищевого продукта, имеет важное значение для сравнения тестируемых продуктов, поскольку корм был разработан как источник белка, предназначенный для пчел M. quadrifasciata , чтобы служить заменителем пыльцы в периоды ее нехватки. Сырой протеин включает все белки в образце пищевого продукта путем количественного определения азота, присутствующего в виде истинных белков [ 30 ].
Учитывая дефицит исследований белковых кормов для безжальных пчел, мы использовали данные Коста и Вентурьери [ 31 ] по Melipona flavolineata в качестве эталона, которые предполагают идеальное содержание сырого протеина (СП) в 20%. Поскольку разработанный в этом исследовании белковый корм изначально не содержал ферментативных инокулянтов, мы установили идеальное значение СП приблизительно в 22%, предполагая, что процесс ферментации, вероятно, снизит общее содержание СП. Корм на основе отрубей, использованный в качестве отправной точки в этом исследовании, имел содержание СП 21,7%, а после ферментации содержание СП в ферментированном корме снизилось до 15,93%.
Важно отметить, что в процессе ферментации количество сырого протеина (СП) имеет тенденцию к снижению, как показано Пайвой [ 16 ]. Содержание СП в ферментированной пыльце в данном исследовании составило 45,32%, что значительно выше значений, указанных в литературе для других видов безжальных пчел рода Melipona . Также стоит отметить, что исследований, оценивающих этот пищевой индекс у безжальных пчел, немного. Как сообщает Ребело [ 26 ], содержание СП у Melipona seminigra составляло 37,63%, у Melipona interrupta — 24%, у Melipona subnitida — 27,02%, а у Melipona scutellaris — 20,81% [ 27 ], что демонстрирует значительную вариацию среди видов рода Melipona . Помимо содержания CP, другие факторы, такие как аминокислотный состав, имеют решающее значение для определения питательной ценности пыльцы, поскольку они дают более точный показатель ее питательной ценности [ 32 , 33 ].
Некоторые исследования, рассмотренные Ноордике и Эллисом [ 34 ], предполагают несколько причин, по которым пчелы могут потреблять такое же количество или даже больше заменителя корма, чем пыльцы. Одна из причин заключается в том, что заменитель корма может содержать больше сахара, чем природный ресурс, ингредиент, часто используемый для повышения его привлекательности для пчел [ 16 , 35 ]. Больший межквартильный размах, наблюдаемый для группы, получавшей пыльцу, по сравнению с группой, получавшей ферментированный корм ( рис. 3 ), может быть обусловлен тем, что пчелы потребляли большее количество пищи с разными интервалами в каждой группе. Как отмечает Пайва [ 16 ], важно учитывать, что поведение пчел, содержащихся в клетках, отличается от их поведения внутри колонии, в первую очередь из-за ограниченности пространства и отсутствия матки, которые могут влиять на то, как пчелы привлекаются к пище.
Обнаруженные соединения, показанные на рисунках 6 и 7, представляют особый интерес для понимания химических компонентов, которые способствуют повышению привлекательности корма для рабочих особей M. quadrifasciata по сравнению с ферментированной пыльцой. Следует отметить олеанановые тритерпеноиды и гликозилированные изофлавоны ( рисунки 6 и 7 ), которые могут быть результатом микробных взаимодействий в ферментированной пыльце с молочнокислыми бактериями, взаимосвязь, ранее описанная в других контекстах [ 36 ]. Кроме того, обогащение кормов за счет добавления соединений растительного происхождения имеет важное значение, поскольку эти химические классы распространены в растениях [ 37 ]. Идентифицированные молекулы растительного происхождения подтверждают предыдущие исследования, в которых были обнаружены флавоноиды, такие как кемпферол и кверцетин, в составе ферментированной пыльцы [ 25 , 38 ]. Обнаружение химических соединений, присутствующих в обеих экспериментальных группах ( рис. 6 и рис. 7 ), еще раз подтверждает сходство между ферментированным кормом и ферментированной пыльцой, что может частично объяснить сохранение химических характеристик корма после его приготовления, делая его узнаваемым и привлекательным для потребления M. quadrifasciata .
5. Выводы
Разработанный корм демонстрирует многообещающий биотехнологический потенциал для восполнения дефицита белка у пчел M. quadrifasciata в периоды нехватки ресурсов. В рецептуре подчеркивается важность ферментации белковых рационов для включения ключевых характеристик, таких как текстура, запах и кислотность, которые имеют решающее значение для привлечения пчел к пище. Сравнение химического состава разработанного корма и ферментированной пыльцы выявило значительное сходство соединений с пищей, которую пчелы естественным образом потребляют в периоды доступности ресурсов. Это указывает на то, что корм содержит молекулы, которые, вероятно, способствуют привлекательности пищи, что подтверждает благоприятные показатели потребления при сравнении ферментированного корма с ферментированной пыльцой. Подобные результаты имеют решающее значение для развития пчеловодства безжальных пчел в Бразилии, поскольку они подчеркивают малоизученную нишу искусственного кормления и здоровья безжальных пчел, способствуя значительному прогрессу в устойчивости и эффективности этой практики.
Дополнительные материалы
Следующие дополнительные материалы можно загрузить по адресу: https://www.mdpi.com/article/10.3390/agriculture15020168/s1 , Таблица S1: Потребление кормов и ферментированной пыльцы.
Ссылки
1. Engel, M.S.; Rasmussen, C.; Ayala, R.; de Oliveira, F.F. Stingless Bee Classification and Biology (Hymenoptera, Apidae): A Review, with an Updated Key to Genera and Subgenera. ZooKeys 2023, 1172, 239–312. [Google Scholar] [CrossRef] [PubMed]
2. Venturieri, G.C.; Pa, E. Criação de Abelhas Indígenas Sem Ferrão; Embrapa: Belém, Brazil, 2017; pp. 1–55. [Google Scholar]
3. Kevan, P.G.; Viana, B.F. The Global Decline of Pollination Services. Biodiversity 2003, 4, 3–8. [Google Scholar] [CrossRef]
4. Klein, A.M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of Pollinators in Changing Landscapes for World Crops. Proc. R. Soc. B Biol. Sci. 2006, 274, 303–313. [Google Scholar] [CrossRef] [PubMed]
5. Cortopassi-Laurino, M.; Imperatriz-Fonseca, V.L.; Roubik, D.W.; Dollin, A.; Heard, T.; Aguilar, I.; Venturieri, G.C.; Eardley, C.; Noguera-Neto, P. Global Meliponiculture: Challenges and Opportunities. Apidologie 2006, 37, 275–292. [Google Scholar] [CrossRef]
6. Naug, D. Nutritional Stress Due to Habitat Loss May Explain Recent Honeybee Colony Collapses. Biol. Conserv. 2009, 142, 2369–2372. [Google Scholar] [CrossRef]
7. Nogueira-Neto, P. Vida e Criação de Abelhas Indígenas Sem Ferrão; Nogueirapis: São Paulo, Brazil, 1997; 445p. [Google Scholar]
8. Teixeira, É.W.; Ferreira, E.A.; da Luz, C.F.P.; Martins, M.F.; Ramos, T.A.; Lourenço, A.P. European Foulbrood in Stingless Bees (Apidae: Meliponini) in Brazil: Old Disease, Renewed Threat. J. Invertebr. Pathol. 2020, 172, 107357. [Google Scholar] [CrossRef] [PubMed]
9. Singh, R.; Levitt, A.L.; Rajotte, E.G.; Holmes, E.C.; Ostiguy, N. RNA Viruses in Hymenopteran Pollinators: Evidence of Inter-Taxa Virus Transmission via Pollen and Potential Impact on Non-Apis Hymenopteran Species. PLoS ONE 2010, 5, 14357. [Google Scholar] [CrossRef] [PubMed]
10. Schmidt, J.O.; Hanna, A. Chemical Nature of Phagostimulants in Pollen Attractive to Honeybees. J. Insect Behav. 2006, 19, 521–532. [Google Scholar] [CrossRef]
11. Almeida-Dias, J.M.; Morais, M.M.; Francoy, T.M.; Pereira, R.A.; Turcatto, A.P.; De Jong, D. Fermentation of a Pollen Substitute Diet with Beebread Microorganisms Increases Diet Consumption and Hemolymph Protein Levels of Honey Bees (Hymenoptera, Apidae). Sociobiology 2018, 65, 760–765. [Google Scholar] [CrossRef]
12. Di Cagno, R.; Filannino, P.; Cantatore, V.; Gobbetti, M. Novel Solid-State Fermentation of Bee-Collected Pollen Emulating the Natural Fermentation Process of Bee Bread. Food Microbiol. 2019, 82, 218–230. [Google Scholar] [CrossRef] [PubMed]
13. Mohammad, S.M.; Mahmud-Ab-Rashid, N.K.; Zawawi, N. Stingless Bee-Collected Pollen (Bee Bread): Chemical and Microbiology Properties and Health Benefits. Molecules 2021, 26, 957. [Google Scholar] [CrossRef] [PubMed]
14. Vollet-Neto, A.; Maia-Silva, C.; Menezes, C.; Imperatriz-Fonseca, V.L. Newly Emerged Workers of the Stingless Bee Scaptotrigona aff. depilis Prefer Stored Pollen to Fresh Pollen. Apidologie 2017, 48, 204–210. [Google Scholar] [CrossRef]
15. Menezes, C.; Vollet-Neto, A.; Contrera, F.A.F.L.; Venturieri, G.C.; Imperatriz-Fonseca, V.L. The Role of Useful Microorganisms to Stingless Bees and Stingless Beekeeping. In Pot-Honey: A Legacy of Stingless Bees; Springer: New York, NY, USA, 2013; pp. 153–171. ISBN 978-1-4614-4960-7. [Google Scholar] [CrossRef]
16. Paiva, J.P.L.M. Avanços Biotecnológicos Sobre Os Efeitos Da Ensilagem Na Qualidade Da Ração Suplementar Proteica Para Abelhas Apis mellifera. Ph.D. Thesis, Universidade Federal de São Paulo, São José dos Campos, Brazil, 2018. [Google Scholar]
17. AOAC: Official Methods of Analysis, 1980: AOAC International: Free Download, Borrow, and Streaming: Internet Archive. Available online: https://archive.org/details/gov.law.aoac.methods.1980/page/n1/mode/1up (accessed on 27 October 2024).
18. Silva, D.J. Análise de Alimentos (Métodos Químicos e Biológicos). UFV: Viçosa, MG, Brazil, 1981; Volume 166.—References—Scientific Research Publishing. Available online: https://www.scirp.org/reference/referencespapers?referenceid=1534598 (accessed on 27 October 2024).
19. De Carvalho, A.C.; Rodrigues, L.D.C.; Ribeiro, A.I.; Da Silva, M.F.D.G.F.; De Medeiros, L.S.; Veiga, T.A.M. Integrated Analytical Tools for Accessing Acridones and Unrelated Phenylacrylamides from Swinglea Glutinosa. Molecules 2020, 25, 153. [Google Scholar] [CrossRef] [PubMed]
20. Fiehn, O.; Robertson, D.; Griffin, J.; vab der Werf, M.; Nikolau, B.; Morrison, N.; Sumner, L.W.; Goodacre, R.; Hardy, N.W.; Taylor, C.; et al. The Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 175–178. [Google Scholar] [CrossRef]
21. Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed Minimum Reporting Standards for Chemical Analysis Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [PubMed]
22. Schymanski, E.L.; Jeon, J.; Gulde, R.; Fenner, K.; Ruff, M.; Singer, H.P.; Hollender, J. Identifying Small Molecules via High Resolution Mass Spectrometry: Communicating Confidence. Environ. Sci. Technol. 2014, 48, 2097–2098. [Google Scholar] [CrossRef] [PubMed]
23. Morais, M.M.; Turcatto, A.P.; Francoy, T.M.; Gonçalves, L.S.; Cappelari, F.A.; De Jong, D. Evaluation of Inexpensive Pollen Substitute Diets through Quantification of Haemolymph Proteins. J. Apic. Res. 2013, 52, 119–121. [Google Scholar] [CrossRef]
24. Bonoan, R.E.; Gonzalez, J.; Starks, P.T. The Perils of Forcing a Generalist to Be a Specialist: Lack of Dietary Essential Amino Acids Impacts Honey Bee Pollen Foraging and Colony Growth. J. Apic. Res. 2020, 59, 95–103. [Google Scholar] [CrossRef]
25. Barta, D.G.; Cornea-Cipcigan, M.; Margaoan, R.; Vodnar, D.C. Biotechnological Processes Simulating the Natural Fermentation Process of Bee Bread and Therapeutic Properties—An Overview. Front. Nutr. 2022, 9, 871896. [Google Scholar] [CrossRef] [PubMed]
26. Rebelo, K.S.; Ferreira, A.G.; Carvalho-Zilse, G.A. Physicochemical Characteristics of Pollen Collected by Amazonian Stingless Bees. Ciência Rural 2016, 46, 927–932. [Google Scholar] [CrossRef]
27. Oliveira, D.d.J.; Rodrigues dos Santos, D.; Andrade, B.R.; Nascimento, A.S.d.; Oliveira da Silva, M.; da Cruz Mercês, C.; Lucas, C.I.S.; Cavalcante da Silva, S.M.P.; Dib de Carvalho, P.; Silva, F.d.L.; et al. Botanical Origin, Microbiological Quality and Physicochemical Composition of the Melipona scutellaris Pot-Pollen (“Samburá”) from Bahia (Brazil) Region. J. Apic. Res. 2021, 60, 457–469. [Google Scholar] [CrossRef]
28. Duarte, A.W.F.; Vasconcelos, M.R.D.S.; Oda-Souza, M.; de OLIVEIRA, F.F.; López, A.M.Q. Honey and Bee Pollen Produced by Meliponini (Apidae) in Alagoas, Brazil: Multivariate Analysis of Physicochemical and Antioxidant Profiles. Food Sci. Technol. 2018, 38, 493–503. [Google Scholar] [CrossRef]
29. Miranda-Pinto, P.; Cappelini, L.T.D.; Diogo, Y.R.; Francoy, T.M.; Morais, M.M. Scientific Note: Behavioral Dynamics of Pollen Storage in Melipona Quadrifasciata. Insectes Soc. 2023, 70, 487–491. [Google Scholar] [CrossRef]
30. Manual de Cálculo de Rações Para Os Animais Domésticos—N. Islabão—Google Books. Available online: https://books.google.co.uk/books/about/Manual_de_c%C3%A1lculo_de_ra%C3%A7%C3%B5es_para_os_a.html?id=8CTSZwEACAAJ&redir_esc=y (accessed on 27 October 2024).
31. Costa, L.; Venturieri, G.C. Diet Impacts on Melipona Flavolineata Workers (Apidae, Meliponini). J. Apic. Res. 2009, 48, 38–45. [Google Scholar] [CrossRef]
32. Mccaughey, W.F.; Gilliam, M.; Standifer, L.N. Amino Acids and Protein Adequacy for Honey Bees of Pollens from Desert Plants and Other Floral Sources. Apidologie 1980, 11, 75–86. [Google Scholar] [CrossRef]
33. Loper, G.M.; Cohen, A.C. Amino Acid Content of Dandelion Pollen, a Honey Bee (Hymenoptera: Apidae) Nutritional Evaluation. J. Econ. Entomol. 1987, 80, 14–17. [Google Scholar] [CrossRef]
34. Noordyke, E.R.; Ellis, J.D. Reviewing the Efficacy of Pollen Substitutes as a Management Tool for Improving the Health and Productivity of Western Honey Bee (Apis Mellifera) Colonies. Front. Sustain. Food Syst. 2021, 5, 772897. [Google Scholar] [CrossRef]
35. Somerville, D. FAT BEES SKINNY BEES—A Manual on Honey Bee Nutrition for Beekeepers—A Report for the Rural Industries Research and Development Corporation; AgriFutures Australia: Wagga Wagga, Australia, 2005. [Google Scholar]
36. Gaya, P.; Peirotén, Á.; Landete, J.M. Transformation of Plant Isoflavones into Bioactive Isoflavones by Lactic Acid Bacteria and Bifidobacteria. J. Funct. Foods 2017, 39, 198–205. [Google Scholar] [CrossRef]
37. Bag, B.G.; Majumdar, R. Self-Assembly of Renewable Nano-Sized Triterpenoids. Chem. Rec. 2017, 17, 841–873. [Google Scholar] [CrossRef]
38. Filannino, P.; Di Cagno, R.; Gambacorta, G.; Tlais, A.Z.A.; Cantatore, V.; Gobbetti, M. Volatilome and Bioaccessible Phenolics Profiles in Lab-Scale Fermented Bee Pollen. Foods 2021, 10, 286. [Google Scholar] [CrossRef] [PubMed]

Miranda-Pinto P, Soares JKC, Gonzalez IHL, Diogo YR, de Medeiros LS, Caperuto LC, Ramos PL, Francoy TM, Morais MM. Biotechnological Approach for Development and Characterization of Protein Feed for Melipona quadrifasciata. Agriculture. 2025; 15(2):168. https://doi.org/10.3390/agriculture15020168
Фото: wikipedia
Перевод статьи «Biotechnological Approach for Development and Characterization of Protein Feed forMelipona quadrifasciata» авторов Miranda-Pinto P, Soares JKC, Gonzalez IHL, Diogo YR, de Medeiros LS, Caperuto LC, Ramos PL, Francoy TM, Morais MM., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык

















































































































Комментарии (0)