Почвенная микробиота "голосует" за стерню. Что это меняет для урожая?
Обработка почвы и управление пожнивными остатками играют ключевую роль
в почвозащитном земледелии, оказывая существенное влияние на свойства почвы.
Цель данного исследования — восполнить пробелы в понимании того, как
взаимодействуют обработка почвы и управление пожнивными остатками, влияя на
таксономическую и функциональную структуру почвенного микробиома. Почвенные
образцы были взяты из долгосрочного полевого опыта, включающего нулевую
обработку (NT) и традиционную
обработку (CT) с удержанием
и удалением стерни.
Аннотация
Методом метагеномного секвенирования был собран генный каталог, содержащий 4,36 миллиарда нереduндантных генов. Управление пожнивными остатками значительно изменило как таксономический, так и функциональный состав прокариотного сообщества: добавление стерни привело к значительному увеличению доли протеобактерий, но уменьшению Chloroflexi по сравнению с вариантом без стерни. Были определены ключевые прокариотные виды и генетические функции, в наибольшей степени способствующие различию прокариотных сообществ между вариантами со стернёй и без неё, включая Rhodospirillum sp. Удержание стерни повысило доступность углеродных ресурсов в почве, что привело к увеличению доли генов, отвечающих за метаболическую активность и взаимодействие растений с патогенами. Однако система обработки почвы не повлияла на структуру или разнообразие сообщества почвенных прокариот. Полученные результаты определяют целевые виды микроорганизмов для будущей изоляции, что позволит разработать экологически безопасные биоудобрения для развития устойчивого сельского хозяйства.
1. Введение
Сельскохозяйственные практики играют ключевую роль в формировании почвенной микро-среды и микробных сообществ. Среди различных параметров, система обработки почвы и управление пожнивными остатками являются критически важными компонентами, которые значительно влияют на физические и химические свойства почвы, что приводит к воздействию на таксономическую и функциональную структуру почвенного микробиома [1]. Понимание взаимодействия обработки почвы и практики обращения со стерней на почвенную микробиоту имеет важнейшее значение для оптимизации управления сельским хозяйством и улучшения здоровья экосистемы.
Механизмы, с помощью которых обработка почвы и управление пожнивными остатками оказывают свое воздействие на свойства почвы, многогранны и различны. Обработка почвы предполагает механическое нарушение ее структуры с помощью таких действий, как вспашка, боронование, культивация, глубокое рыхление, чизелевание, дискование, окучивание, нарезка гребней и прикатывание [2], что тесно связано с увеличением плотности частиц, уменьшением инфильтрации воды и ростом рисков эрозии [3,4,5]. Оставляя растительные остатки на месте, удержание стерни формирует защитные слои на поверхности почвы [6], что приводит к улучшению структуры и пористости почвы, увеличению инфильтрации воды и уменьшению почвенной эрозии [7]. Эти изменения физических свойств почвы, вызванные управлением стерней и обработкой почвы, также влияют на химические свойства почвы, такие как содержание органического углерода (SOC), доступность питательных веществ, электропроводность (EC), pH и емкость катионного обмена (CEC) [8,9,10,11].
Изменения свойств почвы, особенно органического углерода, преобразуют таксономическую структуру почвенного микробиома [12,13,14]. Почвенный микробиом играет ключевую роль в поддержании здоровья и развития сельскохозяйственных культур за счет улучшения доступности питательных веществ [15,16] и подавления роста патогенов [17]. Практики удержания стерни и нулевой обработки (NT) могут сохранять структуру почвы стабильной и способствовать накоплению органического вещества почвы (SOM) [18]. Процесс разложения растительных остатков включает множество разнообразных микроорганизмов, таких как некоторые сапротрофные бактерии и грибы, которые производят ферменты для расщепления сложных органических молекул на основные C-соединения, тем самым увеличивая SOM [19,20,21]. Обработка почвы обычно изменяла активность [22] и структуру почвенного микробного сообщества [23,24], возможно, потому что почвы при традиционной обработке (CT) с низкой инфильтрацией были суше по сравнению с почвами при NT, поэтому органические остатки было труднее разлагать и они обеспечивали менее доступный источник C для микробов. По сравнению с удалением стерни, состав почвенного микробиома при удержании стерни был более сходен с таковым в соседних луговых экосистемах, что приводило к увеличению микробного разнообразия [25]. Удержание стерни влияло на структуру почвенного микробиома, увеличивая относительную обильность Acidobacteria, широко распространенного типа почвенных бактерий, способных использовать разнообразные источники углерода [26]. Кроме того, удержание стерни также взаимодействует с почвенным грибным сообществом [27,28], например, представители Ascomycota могут разлагать сложные C—субстраты, а микоризные грибы могут мобилизовать питательные вещества в стерне [29,30]. Однако другое исследование для полузасушливых систем земледелия показало, что стерня не влияла на бактериальное и грибное разнообразие в почве [31], возможно, из-за среды с низким количеством осадков, ограничивающей вклад растительных остатков в накопление SOM.
Влияние стерни, по-видимому, более выражено, чем влияние обработки почвы, в изменении разнообразия и состава почвенного микробиома. Например, было обнаружено, что органические добавки были гораздо важнее, чем обработка почвы, для изменения численности почвенных микробов, структуры сообщества и активности ферментов (инвертазы и уреазы) [23]. Сообщалось о стабильно схожих результатах: удержание стерни увеличивало как бактериальное, так и грибное разнообразие, независимо от системы обработки почвы, возможно, из-за изменений в доступности питательных веществ, вызванных стерней [32,33].
Изменения почвенной микро-среды, вызванные обработкой почвы и управлением стерней, по-разному влияют на сообщества прокариот и грибов [34]. Прокариоты обладают разнообразными метаболическими путями, с помощью которых они способны адаптироваться к различным абиотическим стрессам в почве, таким как засуха, переувлажнение, жара, а также дефицит кислорода и питательных веществ, вызванный сельскохозяйственными практиками [35,36]. Благодаря высокой скорости размножения и роста прокариоты могут акклиматизироваться к изменениям в почвенной микро-среде за короткий период времени [37]. Грибы, с другой стороны, получают питательные вещества в основном за счет разложения органического вещества [38]; следовательно, они могут быть более чувствительны к увеличению органических поступлений, например, от удержания стерни.
Однако предыдущие исследования, изучавшие взаимодействие обработки почвы и управления стерней на почвенные микробиомы, в основном концентрировались на таксономии микробов, часто упуская из виду функциональную структуру и ее корреляцию с таксономической структурой. Более того, несколько исследований были посвящены взаимодействию структур в разной степени среди различных царств микробиома. Это исследование направлено на восполнение этих пробелов путем изучения сложного взаимодействия между обработкой почвы и управлением стерней на таксономическую и функциональную структуру сообществ бактерий, архей и грибов как средства оценки устойчивого сельского хозяйства и управления экосистемами. Наша гипотеза заключается в том, что существует значительное взаимодействие между обработкой почвы и управлением стерней на почвенные микробиомы, наряду с сильной корреляцией между таксономической и функциональной структурами сообществ различных микробных царств.
2. Материалы и методы
2.1. Полевой опыт
Полевой опыт проводился в уезде Линьи провинции Шаньдун, в северном регионе Китая (координаты GPS 37.46709° с.ш., 116.94031° в.д.) с момента его закладки в 2015 году. На протяжении всего опыта регион получал среднегодовое количество осадков 536,9 мм, причем более 67% этих осадков выпадало между июнем и августом. Зарегистрированная среднегодовая температура составляла 12,6 °C. Почва в этом регионе классифицируется как камбисоли [39].
Этот полевой опыт проводился с чередующимся возделыванием пшеницы и кукурузы каждый год. Схема опыта следовала рандомизированному полному блочному плану с четырьмя повторностями, где каждая делянка обработки имела длину 20 м и ширину 4 м. Применялись как обработка почвы, так и практики удержания стерни, где обработка почвы включала нулевую обработку (NT) и традиционную обработку (CT), а обработка стерни включала удержание стерни (+stubble) и удаление стерни (−stubble). CT проводилась с использованием лемешного плуга на глубину 20 см. Удержание стерни осуществлялось путем прямого нанесения растительных остатков на поверхность почвы для обработки NT (NT +stubble) и путем смешивания растительных остатков с почвой для обработки CT (CT +stubble). В течение экспериментального периода озимый сорт пшеницы Jimai-22 высевался в октябре и убирался в июне следующего года, а сорт кукурузы Dika-1210 высевался после уборки пшеницы. Пшеничная культура удобрялась 225 кг N га⁻¹, 52,4 кг P га⁻¹ и 87,1 кг K га⁻¹, в то время как кукурузная культура удобрялась 240 кг N га⁻¹, 52,4 кг P га⁻¹ и 74,7 кг K га⁻¹. В марте каждого года для пшеничной культуры применялось орошение 130–160 мм воды в зависимости от осадков, и орошение не использовалось в остальное время сезона. Гербициды и пестициды применялись при появлении сорняков и вредителей.
Почвы отбирались после уборки кукурузы в 2021 году, что было через 6 лет после начала обработок. Это исследование было сосредоточено на почвенном микробиоме в верхнем слое почвы, где микробы были более активны и чувствительны к сельскохозяйственной практике. Почвы на глубине 0–10 см отбирались из восьми случайных точек на одной делянке и тщательно смешивались для формирования единого образца почвы, представляющего делянку. Портция в 10 г отбиралась из каждого образца почвы и немедленно переносилась в морозильник при −80 °C в лаборатории до экстракции ДНК. Остаток образца почвы высушивался на воздухе и обрабатывался для физического и химического анализа с использованием протокола Реймента и Лайонса [40]. Измеряемые характеристики почвы включали гранулометрический состав, pH, электропроводность (EC), а также содержание органического C, аммонийного N, нитратного N, общего N, доступного P (на основе теста Колвелла), общего P и общего K.
2.2. Метагеномное секвенирование и биоинформатика
ДНК из 16 образцов, состоящих из 4 повторностей каждой обработки (обработка почвы × стерня), экстрагировалась с использованием набора для выделения ДНК PowerSoil (Mo Bio, Карлсбад, Калифорния, США) в соответствии с инструкциями производителя. Секвенирование ДНК проводилось на основе рекомендаций, предоставленных Куинсом и др. [41]. Для подготовки библиотеки мы использовали набор Hieff NGS® MaxUp II DNA Library Prep Kit от Illumina® (Сан-Диего, Калифорния, США) в соответствии с инструкциями производителя. Адаптеры добавлялись для группировки различных последовательностей из одного образца. Мы количественно определяли и объединяли библиотеки, проводили парно-концевое секвенирование на секвенаторах NovaSeq 6000 от Illumina, США, а затем применяли определенные критерии для фильтрации ридов в процессе секвенирования. Риды отбрасывались, если они: (1) содержали ≥ 20% низкокачественных оснований, или (2) демонстрировали контаминацию адаптерами, что означало, что они имели не более 15 оснований перекрытия между ридами и адаптерами, допуская максимум 3 несовпадения оснований, (3) содержали "N", указывающее на низкое качество, или (4) демонстрировали низкую сложность, имея более 10 последовательных идентичных оснований.
Для выполнения de novo метагеномной сборки все чистые риды из 16 образцов были объединены. Изначально применялась смешанная сборка нескольких образцов с использованием метода графа де Брейна с Megahit версии 1.2.9 [42]. Затем чистые риды картировались обратно на собранные контиги с помощью bowtie2 версии 2.1.0 [43]. Некартированные риды извлекались и повторно собирались с использованием SPAdes версии 3.13 [44] для получения низко-обильных контигов. Мы определяли степень сборки каждого образца путем картирования всех чистых ридов на собранные контиги, используя порог идентичности 90%, с помощью программного обеспечения SoapAligner [45].
Для предсказания генов использовался Prodigal версии 2.60 [46] для идентификации открытых рамок считывания (ORF) из результатов сборки, где гены длиной 100 п.н. или более отбирались и конвертировались в аминокислотные последовательности. Предсказанные гены кластеризовались в нереduндантные гены с использованием CD-HIT версии 2.60 [47]. Для точного количественного определения обилия генов в каждом образце мы использовали двухфазный параллельный алгоритм вывода с Salmon версии 1.5.0 [48], учитывая как количество картированных последовательностей, так и длину гена.
Нереduндантные белковые последовательности аннотировались с использованием микробной базы данных NCBI NR, охватывающей бактерии, грибы, археи и вирусы, а также функциональных баз данных KEGG [49,50]. Эта аннотация проводилась с использованием программного обеспечения DIAMOND версии 0.8.20 [51] с порогом e-value ≤ 1 × 10⁻⁵. Таксономическая аннотация для каждого гена выполнялась с использованием метода наименьшего общего предка (LCA) с MEGAN [52]. Лучшее выравнивание на основе наименьшего e-value blast использовалось в качестве функциональной аннотации метагеномных генов в базе данных KEGG.
2.3. Статистика
Изначально низко-обильные гены отфильтровывались на основе критерия наличия как минимум в 4 образцах с более чем одним ридом. Преобразование Хеллингера [53] применялось для нормализации размера библиотеки в различных образцах. Весь генный каталог был разделен на три группы на основе таксономической аннотации на уровне царства, а именно бактерии, археи и грибы. Внутри каждой из этих групп гены классифицировались на таксономические виды или ортологи KEGG (KO); затем виды и KO использовались в качестве базовой единицы для анализа. Многомерный анализ проводился для видов и KO. Неметрическое многомерное шкалирование (NMDS) на основе расстояния Брея-Кертиса вычислялись функцией metaMDS из пакета Vegan в R. Дисперсионный анализ с перестановками (PERMANOVA) с максимум 999 перестановками с использованием функции adonis проводился для проверки эффектов обработки почвы и стерни на бета-разнообразие. Анализ процентного вклада в несходство (SIMPER) из пакета Vegan проводился для идентификации видов/KO, которые в наибольшей степени способствовали различию микробных сообществ между обработками + и − стерни. Идентифицированные виды/KO рассматривались как ключевые виды/KO. Относительная обильность секвенированных ридов, сгруппированных по видам и KO, сравнивалась в ответ на обработку почвы и стерни с использованием пакета STAMP и метода Бенджамини-Хохберга для коррекции p-значения.
Индекс Шеннона рассчитывался для обозначения альфа-разнообразия таксономического (на уровне видов) и функционального разнообразия (на уровне KO) в каждом из сообществ микробиома, включая бактерии, грибы и археи, с использованием пакета Vegan в R на основе рарифицированных таблиц видов и KO. Двухфакторный ANOVA использовался для проверки влияния обработки почвы и стерни на содержание песка и глины в почве, pH, EC, содержание органического C, аммонийного N, нитратного N, общего N, доступного P, общего P и общего K, а также индекса Шеннона в Minitab 15 (Minitab Inc., Стейт-Колледж, Пенсильвания, США). Перед анализом проверялось нормальное распределение данных. Анализ Мантела проводился для проверки связи между таксономическим и функциональным составом различных микробных царств. Корреляция Пирсона выполнялась среди значений индекса Шеннона таксономического и функционального разнообразия микробных царств. Многомерный дисперсионный анализ с перестановками использовался для проверки корреляции характеристик почвы с таксономическим и функциональным составом различных микробных царств.
3. Результаты
3.1. Физические и химические свойства почвы
Анализ физических и химических свойств почвы (Таблица 1) показал, что удержание стерни значительно увеличило SOC, общий N, P по Колвеллу, общий P и общий K на 14%, 12%, 45%, 25% и 4% соответственно, по сравнению с удалением стерни. Единственное значительное влияние обработки почвы (p < 0,05) было обнаружено на P по Колвеллу, где CT был на 22% выше, чем NT.
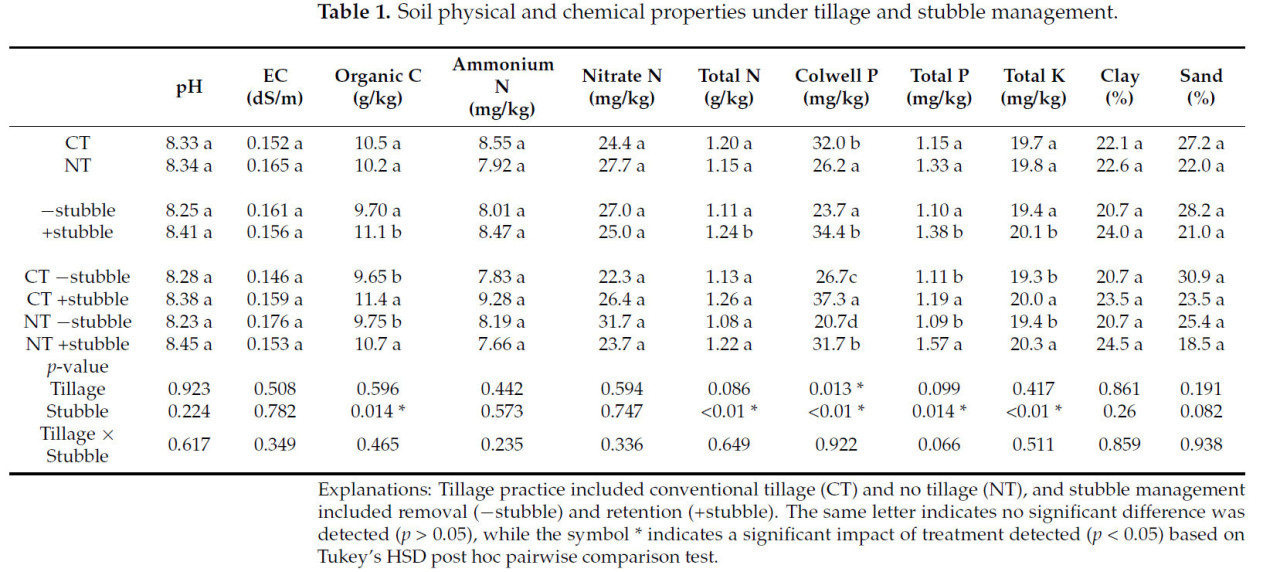
Таблица 1. Физические и химические свойства почвы при различных системах обработки почвы и управления стерней.
3.2. Разнообразие почвенного микробиома
Мы секвенировали 2,08 миллиарда сырых ридов (71,65–152,36 млн на образец) и получили 2,08 миллиарда чистых ридов (71,53–151,75 млн на образец) после контроля качества (Таблица S1). Путем объединения всех 16 образцов мы собрали генный каталог с 4,36 миллиардами нереduндантных генов (2,08–2,18 млн на образец). Генный каталог был разделен на три группы, включая бактерии, археи и грибы. Таксономическое и функциональное разнообразие бактериальных, архейных и грибных сообществ оценивалось при различных обработках обработки почвы и управления стерней.
Для альфа-разнообразия влияние стерни было обнаружено значительным (p < 0,05) только на функциональное разнообразие почвенного бактериального сообщества, где обработка удержания стерни вызывала более высокий индекс Шеннона (Таблица 2). Влияние обработки почвы как на видовое, так и на функциональное разнообразие почвенного грибного сообщества было значительным (p < 0,05), причем более высокое разнообразие наблюдалось при обработке NT (Таблица 2). Однако также наблюдалось значительное взаимодействие между обработками обработки почвы и стерни (p < 0,05) в отношении функционального разнообразия почвенных грибов.
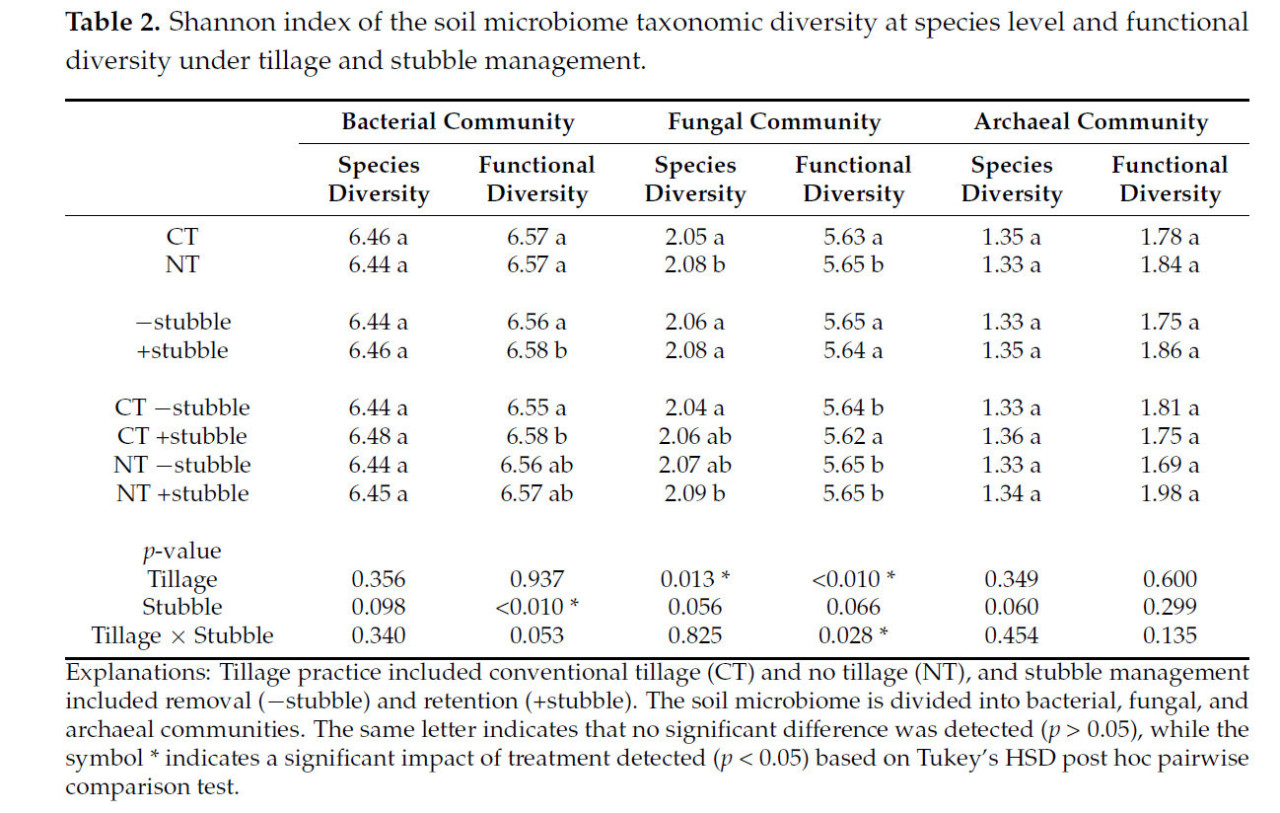
Таблица 2. Индекс Шеннона таксономического разнообразия почвенного микробиома на уровне видов и функционального разнообразия при различных системах обработки почвы и управления стерней.
Для бета-разнообразия влияние стерни было значительным (p < 0,05, Таблица 3) как на видовой состав, так и на функциональный состав бактериальных и архейных сообществ, что иллюстрируется зависимыми от стерни кластерами образцов на биплотах (Рисунок 1a,b,e,f). Влияние обработки почвы было значительным (p < 0,05, Таблица 3) только на функциональный состав архейного сообщества. Не было обнаружено значимости в реакции грибного сообщества на систему управления почвой.
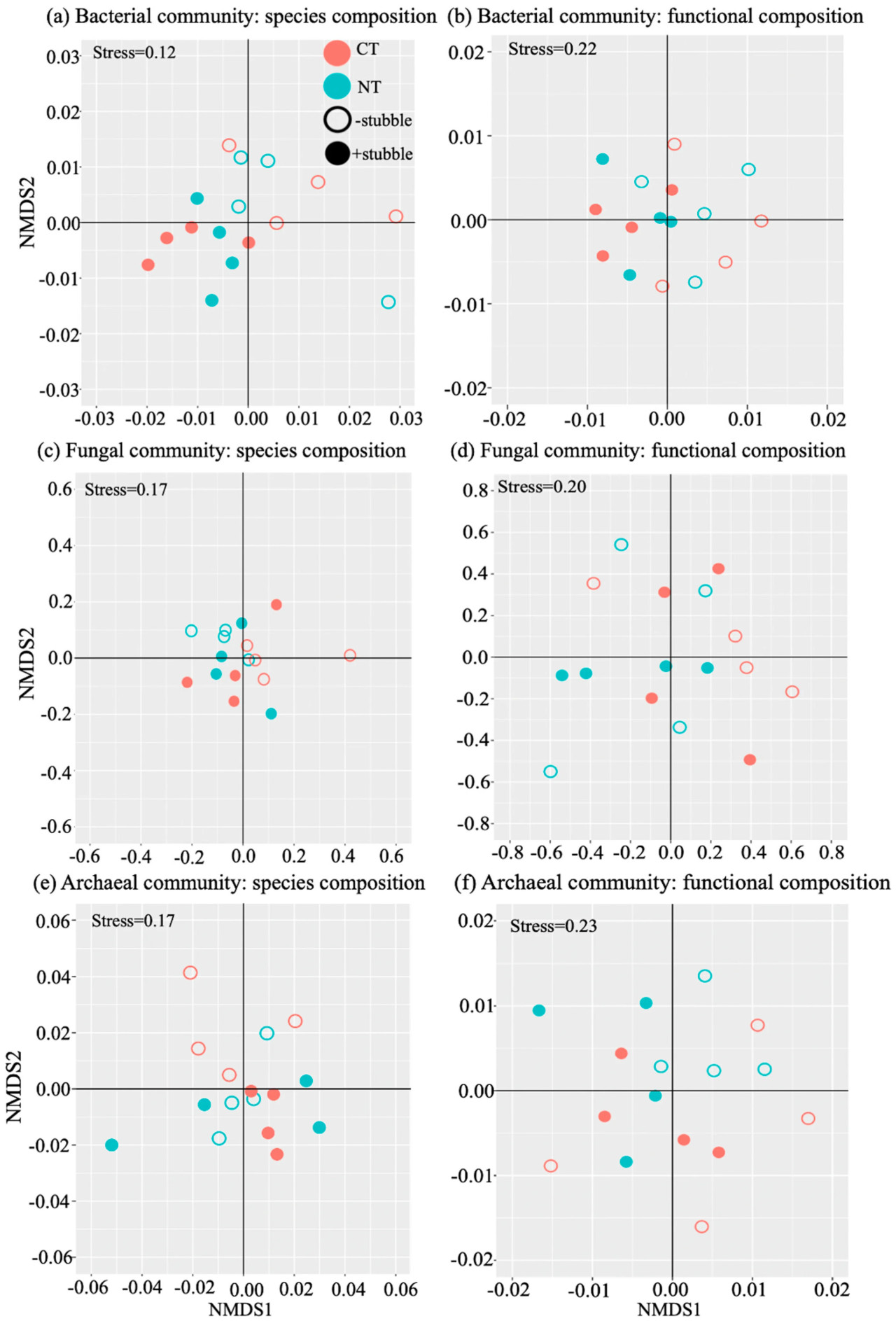
Рисунок 1. Анализ бета-разнообразия видового состава и функционального состава бактериальных (a,b), грибных (c,d) и архейных (e,f) сообществ. Каждая точка представляет отдельный образец обработок, включая традиционную обработку (CT), нулевую обработку (NT), без стерни (−stubble) и с удержанием стерни (+stubble).
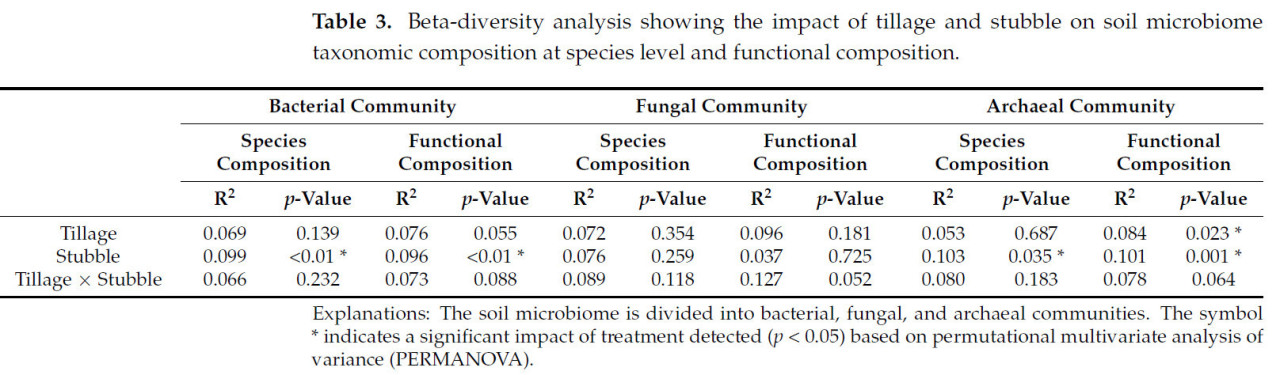
Таблица 3. Анализ бета-разнообразия, показывающий влияние обработки почвы и стерни на таксономический состав почвенного микробиома на уровне видов и функциональный состав.
Несмотря на обнаруженное значительное влияние стерни на уровне видов (p < 0,05, Таблица 3), только типы бактерий Proteobacteria и Chloroflexi были затронуты управлением стерней в различных почвенных микробиомах, включая бактериальные, архейные и грибные сообщества. Обработка удержания стерни вызывала значительное увеличение Proteobacteria (p < 0,05), но значительное уменьшение Chloroflexi (p < 0,05). Proteobacteria и Actinobacteria были наиболее обильными бактериальными типами, составляя одну треть и одну четверть сообщества соответственно, за ними следовали Acidobacteria, которые составляли только около 12% (Рисунок 2a). В грибном сообществе (Рисунок 2b) Ascomycota был наиболее обильным типом, составляя около 80% от общего числа, за ним следовали Mucoromycota (около 15%). Что касается архейного сообщества (Рисунок 2c), доминировали Thaumarchaeota, составляя около 98% от общего числа, причем более половины представителей Thaumarchaeota относились к классу Nitrososphaeria.
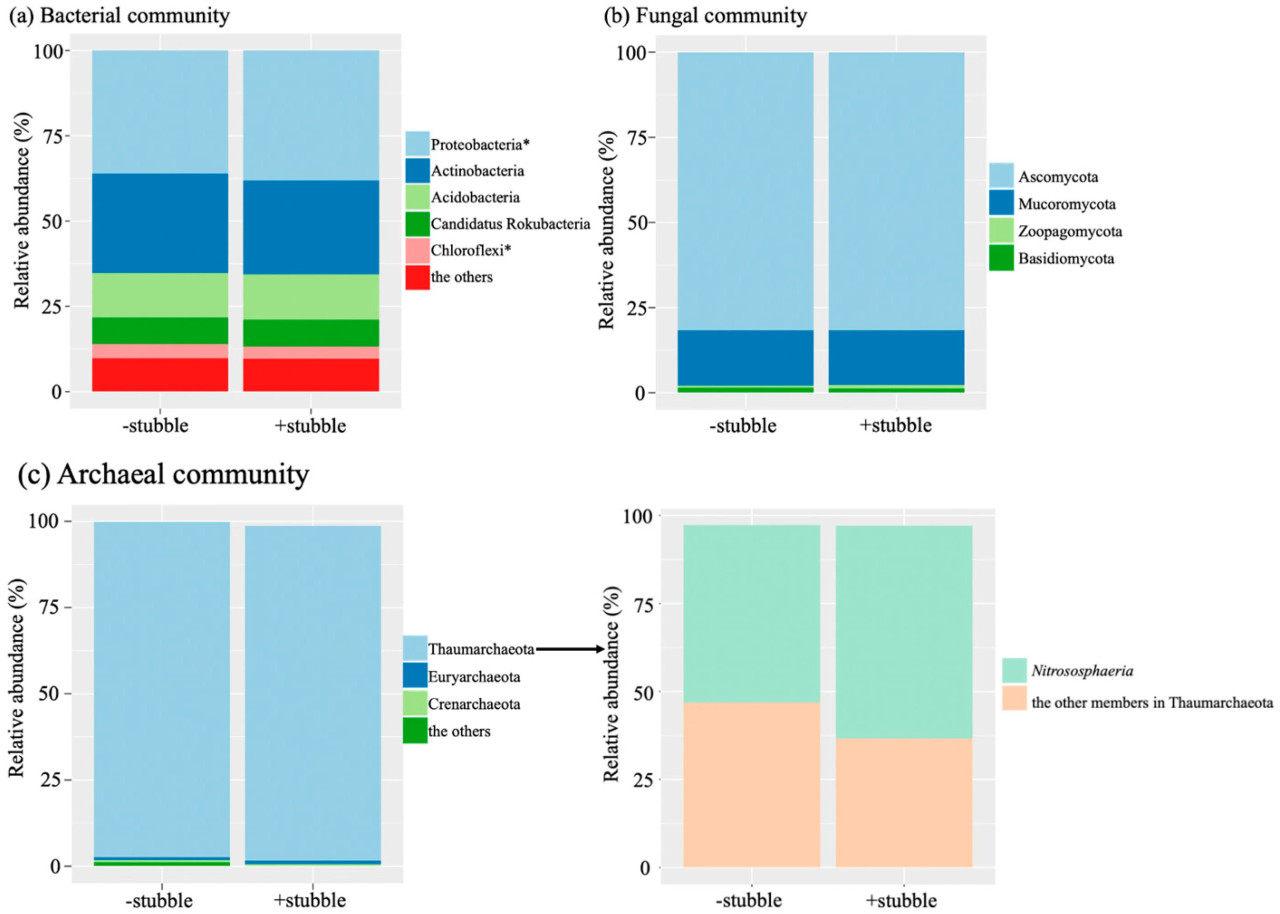
Рисунок 2. Последовательность относительной обильности таксономических групп на уровне типа в бактериальных (a), грибных (b) и архейных (c) сообществах в ответ на обработки −stubble и +stubble. Классовый состав доминирующего архейного типа Thaumarchaeota также показан (c). Символ указывает на значимость при p < 0,05 в ответ на обработку стерни на основе p-значения, скорректированного по методу Бенджамини-Хохберга.
3.3. Ключевые виды и ключевые функциональные единицы
Для дальнейшего изучения значительного влияния стерни (p < 0,05, Таблица 3) как на видовой состав, так и на функциональный состав, ключевые бактериальные и архейные виды, которые составляли более 1% вклада в дифференциацию почвенного микробиома между обработками удержания стерни и удаления стерни, были идентифицированы с использованием анализа процентного вклада в несходство (SIMPER) (Рисунок 3). Относительная обильность идентифицированных ключевых видов имела неоднозначную картину с вкладом в соответствующее сообщество; однако на каждый ключевой вид оказывалось значительное влияние стерни (p < 0,05), как показано на Рисунке 3a,b.
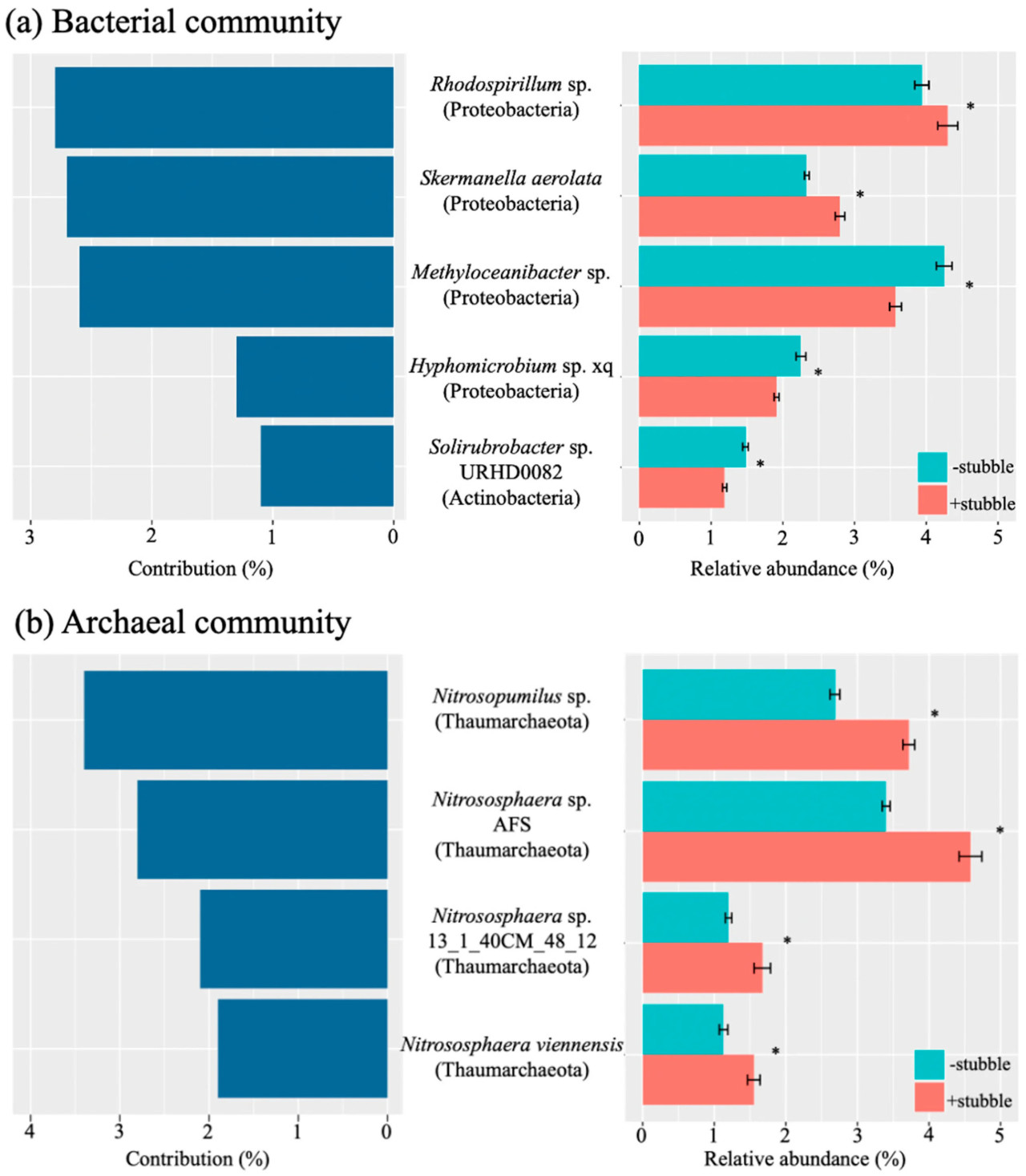
Рисунок 3. Ключевые виды, составляющие более 1% вклада в дифференциацию бактериального (a) и архейного (b) состава почвы под влиянием управления стерней, и относительная обильность ключевых видов при обработках −stubble и +stubble. Был проведен анализ процентного вклада в несходство (SIMPER) для определения вклада в дифференциацию состава микробиома. Идентифицированные микробы были аннотированы на уровне видов. Символ указывает на значимость при p < 0,05 в ответ на обработку стерни, на основе p-значения, скорректированного по методу Бенджамини-Хохберга.
Было идентифицировано пять ключевых бактериальных видов: Rhodospirillum sp., Skermanella aerolata, Methyloceanibacter sp., Hyphomicrobium sp. xq и Solirubrobacter sp. URHD0082, причем все четыре основных ключевых вида были представителями Proteobacteria. Для трех основных ключевых видов (т.е. Rhodospirillum sp., S. aerolata и Methyloceanibacter sp.) каждый составлял как минимум вдвое больший вклад (выше 2,6%), чем два других вида. Несмотря на второй по величине вклад, S. aerolata имела заметно более низкую относительную обильность, чем Methyloceanibacter sp. Rhodospirillum sp. и S. aerolata были единственными бактериальными видами, показавшими значительно более высокую относительную обильность при удержании стерни по сравнению с удалением стерни (Рисунок 3a).
В архейном сообществе (Рисунок 3b) четыре ключевых вида включали Nitrosopumilus sp., Nitrososphaera sp. AFS, Nitrososphaera sp. 131_40CM_48_12 и Nitrososphaera viennensis, и все были представителями Thaumarcheota. Несмотря на наибольший вклад (около 3,4%), относительная обильность Nitrosopumilus sp. была заметно ниже, чем у Nitrososphaera sp. AFS, который составлял наибольшую часть сообщества (Рисунок 3b). Значительно большая относительная обильность была обнаружена при обработке удержания стерни для всех видов (p < 0,05, Рисунок 3b).
Шесть ключевых аннотированных KO были идентифицированы в почвенном бактериальном сообществе (Рисунок 4a) и в почвенном архейном сообществе, соответственно (Рисунок 4b), причем две из шести KO были идентифицированы как имеющие одинаковую функцию. Наблюдалась неоднозначная картина для относительной обильности идентифицированных ключевых KO по сравнению с вкладом каждого в соответствующее сообщество, но влияние стерни было значительным (p < 0,05) на каждую из идентифицированных ключевых KO, независимо от сообщества. Наибольший вклад в почвенное бактериальное сообщество (Рисунок 4a) вносила функция "РНК-полимераза" (K03043), которая была примерно вдвое больше, чем функция "Взаимодействие растение-патоген" (K02358), но имела схожую относительную обильность. И "РНК-полимераза" (K03043), и "Взаимодействие растение-патоген" (K02358) имели относительную обильность, почти втрое превышающую таковую других ключевых функций в бактериальном сообществе, и обе получили значительно отрицательное влияние от обработки удержания стерни (p < 0,05, Рисунок 4a). Обработка удержания стерни оказала значительное положительное влияние (p < 0,05) только на "ABC-транспортеры" (K01996 и K01997) и "Окислительное фосфорилирование" (K00340).
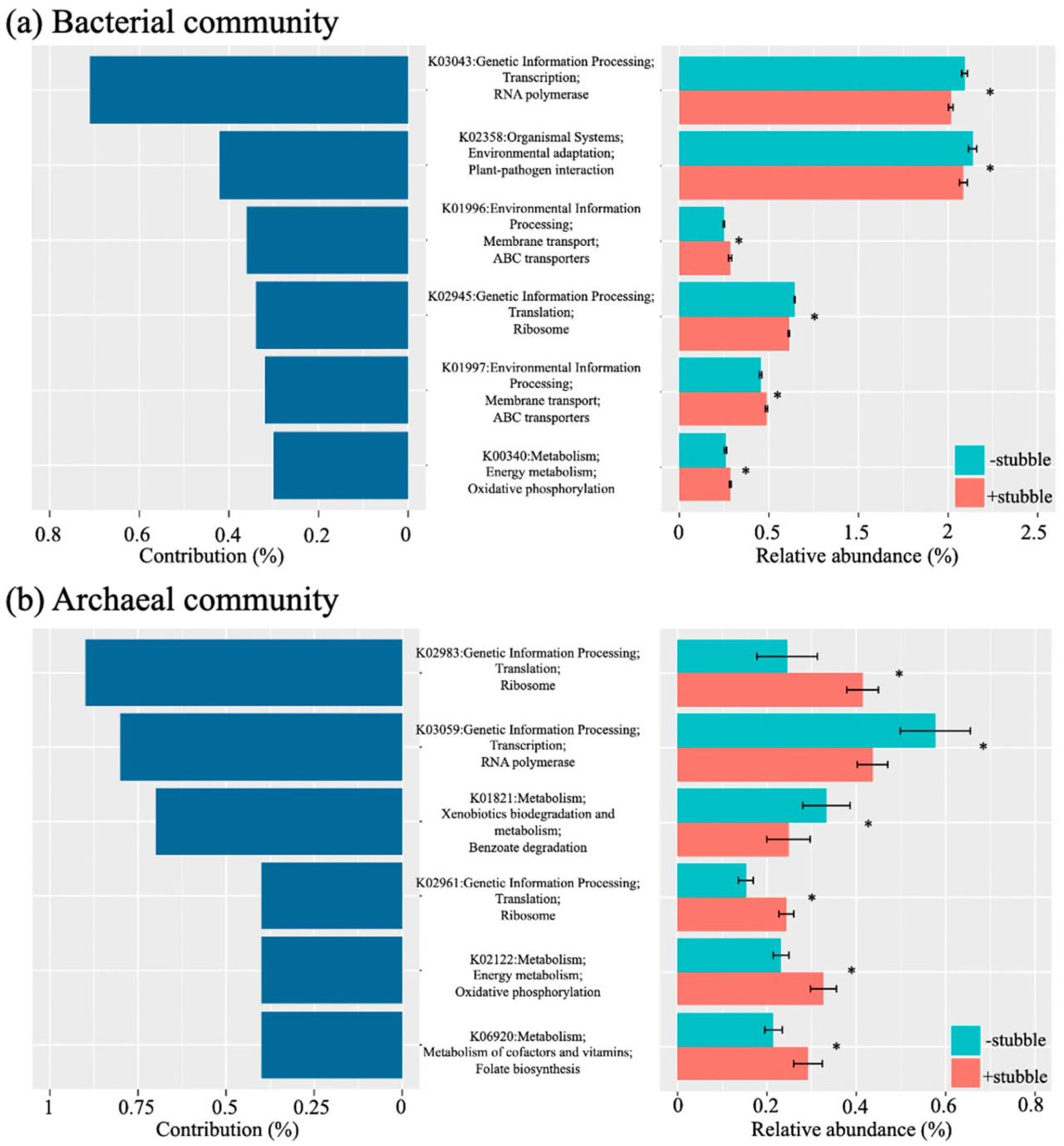
Рисунок 4. Ключевые функции, способствующие дифференциации состава бактериального (a) и архейного (b) микробиома почвы под влиянием управления стерней, и относительная обильность ключевых функций при обработках −stubble и +stubble. Был проведен анализ процентного вклада в несходство (SIMPER) для идентификации вклада в дифференциацию состава микробиома. Идентифицированные функции были аннотированы для ортологических групп KEGG на трех уровнях. Символ указывает на значимость при p < 0,05 в ответ на обработку стерни, на основе p-значения, скорректированного по методу Бенджамини-Хохберга.
Функция "Рибосома" (K02983) составляла наибольшую часть, почти 0,9% от общего вклада в почвенное архейное сообщество (Рисунок 4b), за ней следовали функции "РНК-полимераза" (K03059) и "Деградация бензоата" (K01821). Хотя относительная обильность "Рибосомы" (K02983) была естественно намного ниже, чем у "РНК-полимеразы" (K03059), она увеличилась до уровня, на который снизилась "РНК-полимераза" (K03059) из-за их противоположных реакций на обработку удержания стерни (Рисунок 4b). Функции "Рибосома" (K02961), "Окислительное фосфорилирование" (K02122) и "Биосинтез фолата" (K06920) также поддерживались обработкой удержания стерни; однако каждая вносила менее половины вклада "Рибосомы" (K02983).
Мы провели корреляционный анализ между таксономическим и функциональным разнообразием различных микробных царств. Таксономическое альфа-разнообразие коррелировало с функциональным альфа-разнообразием для архейного сообщества (r < 0) и грибного сообщества (r > 0, Рисунок 5a). Для бета-разнообразия таксономический состав бактерий коррелировал с их функциональным составом (Рисунок 5b). Дополнительно мы исследовали корреляцию физических и химических свойств почвы с видовым и функциональным разнообразием в различных сообществах. Для анализа использовались только свойства почвы, значительно затронутые управлением стерней и обработкой почвы. SOC не имел связи с разнообразием микробиома (данные не представлены). Грибное сообщество не подвергалось влиянию свойств почвы. Бактериальное таксономическое и функциональное разнообразие демонстрировало положительные корреляции с общим P и P по Колвеллу в почве, в то время как архейное разнообразие демонстрировало отрицательные корреляции с общим N и P по Колвеллу. Общий N и P по Колвеллу также значительно влияли на таксономическую и функциональную структуру бактериального микробиома, но только на функциональный состав архейного сообщества.
Рисунок 5. Коэффициент корреляции (r) между таксономическим и функциональным разнообразием различных микробных царств. (a) Корреляция Пирсона индекса Шеннона и (b) анализ Мантела между таксономическим и функциональным составом различных микробных царств. Символ указывает на значимость при p < 0,05.
4. Обсуждение
Это исследование собрало генный каталог почвенного метагенома, затем идентифицировало три группы бактериальных, архейных и грибных сообществ для таксономического и функционального анализа. Мы демонстрируем, что обработка почвы и управление стерней оказывали различное влияние на почвенный микробиом в разных микробных царствах. Не было взаимодействия между обработкой почвы и управлением стерней на видовой или функциональный состав любого из трех сообществ; индивидуальный эффект управления стерней был сильнее, чем эффект обработки почвы, в влиянии на состав и разнообразие прокариотного сообщества, в то время как реакция грибного сообщества была более стабильной к различным техникам возделывания. Кроме того, связь между таксономической и функциональной структурой была обнаружена только в бактериальном сообществе. Бактериальный и архейный микробиомы были тесно коррелированы в таксономическом и функциональном составе под влиянием обработки почвы и управления стерней, но отличались от грибного сообщества. Управление стерней модулировало абиотические свойства почвы, такие как SOC и доступный P, что, вероятно, объясняло его влияние на почвенный микробиом.
4.1. Эффекты управления стерней и обработки почвы на структуру и разнообразие микробного сообщества
В нашем исследовании не было обнаружено влияния обработки почвы на структуру и разнообразие почвенного прокариотного сообщества (т.е. бактерий и архей), но влияние стерни на таксономический и функциональный состав было значительным. Учитывая, что стерня служит дополнительным источником углерода и питательных веществ для почвенных прокариотических организмов в процессе разложения растительных остатков [54], удержание стерни увеличивает количество органического вещества в почве, обеспечивая микробную активность большей энергией и питательными веществами [55]. Сильное влияние управления стерней на таксономическую структуру почвенных прокариотных сообществ также было обнаружено в предыдущих исследованиях [28,56,57]. Согласно нашим результатам, изменения в составе типов изменили таксономическую структуру прокариотных сообществ. Мы обнаружили, что только два типа, Proteobacteria и Chloroflexi, были чувствительны к управлению стерней, но с противоположными реакциями, где более высокая обильность Proteobacteria наблюдалась, когда стерня удерживалась, и, что касается более высокой обильности Chloroflexi, когда стерня удалялась (Рисунок 2a). Интересно, что те же два типа также были идентифицированы как наиболее чувствительные бактериальные типы к удержанию стерни в другом исследовании [58]. Увеличение SOM, вызванное стерней, могло быть ключевым фактором увеличения Proteobacteria, потому что быстрорастущие копиотрофные представители Proteobacteria были благоприятны для микро-среды с высоким содержанием C [59]. Напротив, из-за своей чувствительности к олиготрофной среде обитания, Chloroflexi был менее конкурентоспособен в нише с высоким содержанием питательных веществ [55].
Дополнительно, это исследование выявило влияние удержания стерни на функциональную структуру почвенных прокариотных сообществ. Увеличение разложения SOM и высвобождения питательных веществ при обработке удержания стерни может усиливать микробные функции, связанные с круговоротом углерода и питательных веществ (т.е. азота и фосфора), чтобы облегчить адаптацию к различным почвенным условиям [35,36], таким как функции азотфиксации, нитрификации и денитрификации в цикле азота и окислительные функции в разложении SOM [11,60,61]. Наши результаты подчеркивают, что статус питательных веществ в почве, а не органический углерод, влиял на функциональную структуру почвенных прокариотных сообществ, хотя удержание стерни увеличивало как SOC, так и питательные вещества почвы N, P и K (Таблица 1). Возможная причина заключается в том, что деградация SOC является длительным и сложным процессом, который постепенно модулирует почвенный микробиом в длительной перспективе [62], тогда как почвенные питательные вещества могут напрямую обеспечивать краткосрочные пищевые ресурсы [63], оказывая, таким образом, более немедленное влияние на функцию и структуру микробиома.
В отличие от прокариотных сообществ, разнообразие и структура грибного сообщества были более стабильными и в меньшей степени модулировались управлением стерней и обработкой почвы; только альфа-разнообразие подвергалось влиянию системы обработки почвы. Подобно нашему наблюдению, более высокое альфа-разнообразие также обнаруживалось при применении NT или минимальной обработки почвы в различных системах возделывания [64].
4.2. Ключевые виды и функции, определяющие прокариотное сообщество
Ключевыми бактериальными видами, определяющими несходство в бактериальном сообществе между обработками удержания стерни и удаления стерни, были представители Proteobacteria, включая Rhodospirillum sp., S. aerolata и Methyloceanibacter sp. Учитывая, что Rhodospirillum sp. ассоциируется с возникновением болезней сельскохозяйственных культур [65], наш результат относительно его большой обильности в обработке удержания стерни, по-видимому, согласуется с предыдущим выводом о более высокой заболеваемости, когда стерня удерживалась [66]; однако Rhodospirillum известен как факультативный анаэроб и не известен как возбудитель болезней [67]. Rhodospirillum sp. известен своей способностью фиксировать азот; следовательно, он мог способствовать увеличению генов, связанных с азотфиксацией [68]. S. aerolata является денитрифицирующей бактерией, несущей ген nirS [69]. Обильность nirS была выше в почвах с органическим удобрением, чем в почвах с минеральным удобрением, что предполагает, что денитрифицирующие микроорганизмы обогащаются при добавлении органического вещества в почву [69], что согласуется с нашим выводом о том, что при удержании стерни относительная обильность S. aerolata была значительно выше по сравнению с таковой при удалении стерни. Methyloceanibacter sp. известен своей способностью к фиксации углерода [70] и потенциально может играть значительную роль в углеродном метаболизме почвенного микробиома.
Ключевыми архейными видами, определяющими несходство архейного сообщества между обработками удержания стерни и удаления стерни, были представители Thaumarcheota, включая Nitrosopumilus и Nitrososphaera. Эти два рода классифицируются как аммиак-окисляющие археи (AOA) [71]. AOA способны окислять аммиак, ключевой этап в превращении атмосферного азота в биологически доступную форму [71]. Внесение биоугля в почву способствовало росту AOA, увеличивая уровень органического углерода почвы, общего азота и аммонийного азота, в конечном итоге увеличивая доступность энергии [72]. Обработка удержания стерни в нашем исследовании создала обогащенное C и N состояние, что привело к увеличению обильности и разнообразия AOA, что могло способствовать азотному циклу и улучшению качества почвы.
Функция обработки генетической информации, в частности, транскрипция РНК-полимеразы, играла значительную роль в определении несходства в функциональной структуре бактерий и архей в ответ на обработки стерни. В частности, относительная обильность этих двух ключевых KO была ниже в образцах почвы из обработки удержания стерни по сравнению с удалением стерни. Удержание стерни увеличивает доступность углеродных ресурсов в почве, что приводит к более высокой доле генов, функциональных для метаболической активности [59], и может снижать относительную обильность функций обработки генетической информации.
В дополнение к упомянутым выше функциям обработки генетической информации, "взаимодействие растение-патоген" является еще одной значительной функциональной единицей, вызывающей изменения в бактериальном функциональном составе. Относительная обильность этой функциональной единицы была выше в обработке удаления стерни, чем в обработке удержания стерни, что указывает на сложные взаимодействия между патогенами, болезне-супрессивными микробами и растительными остатками в почве [13].
5. Выводы
Это исследование показывает сложное взаимодействие между обработкой почвы и управлением стерней на почвенный микробиом, включая таксономическое и функциональное разнообразие бактериальных, архейных и грибных сообществ. Оно подчеркивает, что системы обработки почвы не значительно изменили прокариотное сообщество в почве, в то время как управление стерней заметно влияло как на таксономический, так и на функциональный состав этих микробных сообществ. Интересно, что грибное сообщество реагировало иначе, показывая, что изменения альфа-разнообразия были связаны с методами обработки почвы. Дополнительные исследования будут направлены на выделение полезных микробных видов, идентифицированных с помощью метагеномного анализа в этом исследовании, скрининг микроорганизмов, способствующих росту растений, и разработку экологически безопасных биоудобрений и биопестицидов для продвижения устойчивых сельскохозяйственных практик.
Дополнительные материалы
Следующая вспомогательная информация может быть загружена по адресу: https://www.mdpi.com/article/10.3390/agriculture15020143/s1; Таблица S1: Количество секвенированных ридов и картированных генов 16 образцов в настоящем исследовании.
Ссылки
1. Shia, A.; Cavagnarob, T.R.; Sumbyb, K.M.; McDonaldb, G.; Dentonb, M.D.; Royb, S.J.; Schillinga, R.K. Impact of agronomic management on the soil microbiome: A southern Australian dryland broadacre perspective. Adv. Agron. 2024, 186, 113. [Google Scholar] [CrossRef]
2. Lal, R.; Eckert, D.; Fausey, N.; Edwards, W. Conservation tillage in sustainable agriculture. Sustain. Agric. Syst. 1990, 203–225. [Google Scholar]
3. Alam, M.K.; Islam, M.M.; Salahin, N.; Hasanuzzaman, M. Effect of tillage practices on soil properties and crop productivity in wheat-mungbean-rice cropping system under subtropical climatic conditions. Sci. World J. 2014, 2014, 437283. [Google Scholar] [CrossRef] [PubMed]
4. Blanco-Canqui, H.; Ruis, S.J. No-tillage and soil physical environment. Geoderma 2018, 326, 164–200. [Google Scholar] [CrossRef]
5. Sarkar, R.; Kar, S. Temporal changes in fertility and physical properties of soil under contrasting tillage-crop residue management for sustainable rice-wheat system on sandy-loam soil. J. Crop Improv. 2011, 25, 262–290. [Google Scholar] [CrossRef]
6. Roper, M.M.; Ward, P.R.; Keulen, A.F.; Hill, J.R. Under no-tillage and stubble retention, soil water content and crop growth are poorly related to soil water repellency. Soil Tillage Res. 2013, 126, 143–150. [Google Scholar] [CrossRef]
7. Busari, M.A.; Kukal, S.S.; Kaur, A.; Bhatt, R.; Dulazi, A.A. Conservation tillage impacts on soil, crop and the environment. Int. Soil Water Conserv. Res. 2015, 3, 119–129. [Google Scholar] [CrossRef]
8. Allen, D.E.; Singh, B.P.; Dalal, R.C. Soil health indicators under climate change: A review of current knowledge. In Soil Health and Climate Change; Springer: Berlin/Heidelberg, Germany, 2011; Volume 29. [Google Scholar] [CrossRef]
9. Chen, H.Q.; Hou, R.X.; Gong, Y.S.; Li, H.W.; Fan, M.S.; Kuzyakov, Y. Effects of 11 years of conservation tillage on soil organic matter fractions in wheat monoculture in Loess Plateau of China. Soil Tillage Res. 2009, 106, 85–94. [Google Scholar] [CrossRef]
10. Lv, L.; Gao, Z.; Liao, K.; Zhu, Q.; Zhu, J. Impact of conservation tillage on the distribution of soil nutrients with depth. Soil Tillage Res. 2023, 225, 105527. [Google Scholar] [CrossRef]
11. Zhang, L.; Su, X.; Meng, H.; Men, Y.; Liu, C.; Yan, X.; Mao, L. Cotton stubble return and subsoiling alter soil microbial community, carbon and nitrogen in coastal saline cotton fields. Soil Tillage Res. 2023, 226, 105585. [Google Scholar] [CrossRef]
12. Obayomi, O.; Seyoum, M.M.; Ghazaryan, L.; Tebbe, C.C.; Murase, J.; Bernstein, N.; Gillor, O. Soil texture and properties rather than irrigation water type shape the diversity and composition of soil microbial communities. Appl. Soil Ecol. 2021, 161, 103834. [Google Scholar] [CrossRef]
13. Pankhurst, C.E.; McDonald, H.J.; Hawke, B.G.; Kirkby, C.A. Effect of tillage and stubble management on chemical and microbiological properties and the development of suppression towards cereal root disease in soils from two sites in NSW, Australia. Soil Biol. Biochem. 2002, 34, 833–840. [Google Scholar] [CrossRef]
14. Wang, X.B.; Yao, J.; Zhang, H.Y.; Wang, X.G.; Li, K.H.; Lu, X.T.; Wang, Z.W.; Zhou, J.Z.; Han, X.G. Environmental and spatial variables determine the taxonomic but not functional structure patterns of microbial communities in alpine grasslands. Sci. Total Environ. 2019, 654, 960–968. [Google Scholar] [CrossRef] [PubMed]
15. Alemneh, A.A.; Zhou, Y.; Ryder, M.H.; Denton, M.D. Is phosphate solubilizing ability in plant growth-promoting rhizobacteria isolated from chickpea linked to their ability to produce ACC deaminase? J. Appl. Microbiol. 2021, 131, 2416–2432. [Google Scholar] [CrossRef]
16. Zhou, Y.; Lambrides, C.J.; Li, J.; Xu, Q.; Toh, R.; Tian, S.; Yang, P.; Yang, H.; Ryder, M.; Denton, M.D. Nitrifying microbes in the rhizosphere of perennial grasses are modified by biological nitrification inhibition. Microorganisms 2020, 8, 1687. [Google Scholar] [CrossRef]
17. Sui, L.; Li, J.; Philp, J.; Yang, K.; Wei, Y.; Li, H.; Li, J.; Li, L.; Ryder, M.; Toh, R.; et al. Trichoderma atroviride seed dressing influenced the fungal community and pathogenic fungi in the wheat rhizosphere. Sci. Rep. 2022, 12, 9677. [Google Scholar] [CrossRef]
18. Anderson, G. The Impact of Tillage Practices and Crop Residue (Stubble) Retention in the Cropping System of Western Australia; Department of Agriculture and Food: Perth, WA, Australia, 2009.
19. Ayilara, M.S.; Olanrewaju, O.S.; Babalola, O.O.; Odeyemi, O. Waste management through composting: Challenges and potentials. Sustainability 2020, 12, 4456. [Google Scholar] [CrossRef]
20. Gupta, A.; Singh, U.B.; Sahu, P.K.; Paul, S.; Kumar, A.; Malviya, D.; Singh, S.; Kuppusamy, P.; Singh, P.; Paul, D.; et al. Linking soil microbial diversity to modern agriculture practices: Review. Int. J. Environ. Res. Public Health 2022, 19, 3141. [Google Scholar] [CrossRef]
21. Hubbe, M.; Nazhad, M.; Sanchez, C. Composting as a way to convert cellulosic biomass and organic waste into high-value soil amendments: A Review. Bioresource 2010, 5, 2808–2854. [Google Scholar] [CrossRef]
22. Zuber, S.M.; Villamil, M.B. Meta-analysis approach to assess effect of tillage on microbial biomass and enzyme activities. Soil Biol. Biochem. 2016, 97, 176–187. [Google Scholar] [CrossRef]
23. Chen, X.; Henriksen, T.M.; Svensson, K.; Korsaeth, A. Long-term effects of agricultural production systems on structure and function of the soil microbial community. Appl. Soil Ecol. 2020, 147, 103387. [Google Scholar] [CrossRef]
24. Jackson, L.E.; Calderon, F.J.; Steenwerth, K.L.; Scow, K.M.; Rolston, D.E. Responses of soil microbial processes and community structure to tillage events and implications for soil quality. Geoderma 2003, 114, 305–317. [Google Scholar] [CrossRef]
25. Wang, Y.; Li, C.; Tu, C.; Hoyt, G.D.; DeForest, J.L.; Hu, S. Long-term no-tillage and organic input management enhanced the diversity and stability of Soil Microbial Community. Sci. Total Environ. 2017, 609, 341–347. [Google Scholar] [CrossRef] [PubMed]
26. Ward, N.L.; Challacombe, J.F.; Janssen, P.H.; Henrissat, B.; Coutinho, P.M.; Wu, M.; Kuske, C.R. Three genomes from the phylum Acidobacteria provide insight into the lifestyles of these microorganisms in soils. Appl. Environ. Microbiol. 2009, 75, 2046–2056. [Google Scholar] [CrossRef]
27. Fang, Y.Y.; Van Zwieten, L.; Rose, M.T.; Vasileiadis, S.; Donner, E.; Vancov, T.; Rigg, J.L.; Weng, Z.; Lombi, E.; Drigo, B.; et al. Unraveling microbiomes and functions associated with strategic tillage, stubble, and fertilizer management. Agric. Ecosyst. Environ. 2022, 323, 107686. [Google Scholar] [CrossRef]
28. Wakelin, S.A.; Colloff, M.J.; Harvey, P.R.; Marschner, P.; Gregg, A.L.; Rogers, S.L. The effects of stubble retention and nitrogen application on soil microbial community structure and functional gene abundance under irrigated maize. FEMS Microbiol. Ecol. 2007, 59, 661–670. [Google Scholar] [CrossRef]
29. Hobbie, E.A.; Horton, T.R. Evidence that saprotrophic fungi mobilise carbon and mycorrhizal fungi mobilise nitrogen during litter decomposition. New Phytol. 2007, 173, 447–449. [Google Scholar] [CrossRef]
30. Ma, A.; Zhuang, X.; Wu, J.; Cui, M.; Lv, D.; Liu, C.; Zhuang, G. Ascomycota members dominate fungal communities during straw residue decomposition in arable soil. PLoS ONE 2013, 8, e66146. [Google Scholar] [CrossRef]
31. Hoyle, F.C.; Murphy, D.V. Seasonal changes in microbial function and diversity associated with stubble retention versus burning. Soil Res. 2006, 44, 407. [Google Scholar] [CrossRef]
32. Sarker, J.R.; Singh, B.P.; Cowie, A.L.; Fang, Y.; Collins, D.; Badgery, W.; Dalal, R.C. Agricultural management practices impacted carbon and nutrient concentrations in soil aggregates, with minimal influence on aggregate stability and total carbon and nutrient stocks in contrasting soils. Soil Tillage Res. 2018, 178, 209–223. [Google Scholar] [CrossRef]
33. Nazaries, L.; Singh, B.P.; Sarker, J.R.; Fang, Y.; Klein, M.; Singh, B.K. The response of soil multi-functionality to agricultural management practices can be predicted by key soil abiotic and biotic properties. Agric. Ecosyst. Environ. 2021, 307, 107206. [Google Scholar] [CrossRef]
34. Yang, T.; Lupwayi, N.; Marc, S.A.; Siddique, K.H.; Bainard, L.D. Anthropogenic drivers of soil microbial communities and impacts on soil biological functions in agroecosystems. Glob. Ecol. Conserv. 2021, 27, e01521. [Google Scholar] [CrossRef]
35. Hayatsu, M.; Tago, K.; Saito, M. Various players in the nitrogen cycle: Diversity and functions of the microorganisms involved in nitrification and denitrification. J. Soil Sci. Plant Nutr. 2008, 54, 33–45. [Google Scholar] [CrossRef]
36. Kuypers, M.M.M.; Marchant, H.K.; Kartal, B. The microbial nitrogen-cycling network. Nat. Rev. Microbiol. 2018, 16, 263–276. [Google Scholar] [CrossRef] [PubMed]
37. Paul, E.; Frey, S. (Eds.) Soil Microbiology, Ecology and Biochemistry; Elsevier: Amsterdam, The Netherlands, 2023. [Google Scholar]
38. Mayer, M.; Rewald, B.; Matthews, B.; Sanden, H.; Rosinger, C.; Katzensteiner, K.; Godbold, D.L. Soil fertility relates to fungal-mediated decomposition and organic matter turnover in a temperate mountain forest. New Phytol. 2021, 231, 777–790. [Google Scholar] [CrossRef]
39. Deckers, J.A.; Nachtergaele, F.; Spaargaren, O.C. (Eds.) World Reference Base for Soil Resources Introduction; Acco: Louven, Belgium, 1998. [Google Scholar]
40. Rayment, G.E.; Lyons, D.J. Soil Chemical Methods Australasia; CSIRO: Canberra, Australia, 2011; Volume 3.
41. Quince, C.; Walker, A.W.; Simpson, J.T.; Loman, N.J.; Segata, N. Shotgun metagenomics, from sampling to analysis. Nat. Biotechnol. 2017, 35, 833–844. [Google Scholar] [CrossRef]
42. Li, D.; Liu, C.M.; Luo, R.; Sadakane, K.; Lam, T.W. MEGAHIT: An ultra-fast single-node solution for large and complex metagenomics assembly via succinct de Bruijn graph. Bioinformatics 2015, 31, 1674–1676. [Google Scholar] [CrossRef]
43. Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
44. Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
45. Li, R.; Li, Y.; Kristiansen, K.; Wang, J. SOAP: Short oligonucleotide alignment program. Bioinformatics 2008, 24, 713–714. [Google Scholar] [CrossRef]
46. Liu, Y.; Guo, J.; Hu, G.; Zhu, H. Gene prediction in metagenomic fragments based on the SVM algorithm. BMC Bioinform. 2013, 14, S12. [Google Scholar] [CrossRef] [PubMed]
47. Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [PubMed]
48. Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef] [PubMed]
49. Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
50. Kanehisa, M. From genomics to chemical genomics: New developments in KEGG. Nucleic Acids Res. 2006, 34, D354–D357. [Google Scholar] [CrossRef] [PubMed]
51. Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
52. Huson, D.H.; Auch, A.F.; Qi, J.; Schuster, S.C. MEGAN analysis of metagenomic data. Genome Res. 2007, 17, 377–386. [Google Scholar] [CrossRef]
53. Bahram, M.; Hildebrand, F.; Forslund, S.K.; Anderson, J.L.; Soudzilovskaia, N.A.; Bodegom, P.M.; Bengtsson-Palme, J.; Anslan, S.; Coelho, L.P.; Harend, H.; et al. Structure and function of the global topsoil microbiome. Nature 2018, 560, 233–237. [Google Scholar] [CrossRef]
54. Wang, L.; Lu, P.; Feng, S.; Hamel, C.; Sun, D.; Siddique, K.H.M.; Gan, G.Y. Strategies to improve soil health by optimizing the plant–soil–microbe–anthropogenic activity nexus. Agric. Ecosyst. Environ. 2024, 359, 108750. [Google Scholar] [CrossRef]
55. Tian, W.; Wang, L.; Li, Y.; Zhuang, K.; Li, G.; Zhang, J.; Xiao, X.; Xi, Y. Responses of microbial activity, abundance, and community in wheat soil after three years of heavy fertilization with manure-based compost and inorganic nitrogen. Agric. Ecosyst. Environ. 2015, 213, 219–227. [Google Scholar] [CrossRef]
56. Arunrat, N.; Sereenonchai, S.; Sansupa, C.; Kongsurakan, P.; Hatano, R. Effect of rice straw and stubble burning on soil physicochemical properties and bacterial communities in central thailand. Biology 2023, 12, 501. [Google Scholar] [CrossRef] [PubMed]
57. Marsch, R.; Verhulst, N.; Govaerts, B.; Dendooven, L. Bacterial indicator taxa in soils under different long-term agricultural management. J. Appl. Microbiol. 2016, 120, 921–933. [Google Scholar] [CrossRef]
58. Yu, D.; Wen, Z.; Li, X.; Song, X.; Wu, H.; Yang, P. Effects of straw return on bacterial communities in a wheat-maize rotation system in the North China Plain. PLoS ONE 2018, 13, e0198087. [Google Scholar] [CrossRef] [PubMed]
59. Liu, H.; Xu, W.; Li, J.; Yu, Z.; Zeng, Q.; Tan, W.; Mi, W. Short-term effect of manure and straw application on bacterial and fungal community compositions and abundances in an acidic paddy soil. J. Soils Sediments 2021, 21, 3057–3071. [Google Scholar] [CrossRef]
60. Lai, H.L.; Gao, F.Y.; Su, H.; Zheng, P.; Li, Y.Y.; Yao, H.Y. Nitrogen distribution and soil microbial community characteristics in a legume-cereal intercropping system: A review. Agronomy 2022, 12, 1900. [Google Scholar] [CrossRef]
61. Zhang, H.Y.; Shi, Y.C.; Dong, Y.X.; Lapen, D.R.; Liu, J.H.; Chen, W. Subsoiling and conversion to conservation tillage enriched nitrogen cycling bacterial communities in sandy soils under long-term maize monoculture. Soil Tillage Res. 2022, 215, 105197. [Google Scholar] [CrossRef]
62. Liu, C.; Lu, M.; Cui, J.; Li, B.; Fang, C. Effects of straw carbon input on carbon dynamics in agricultural soils: A meta-analysis. Glob. Change Biol. 2014, 20, 1366–1381. [Google Scholar] [CrossRef]
63. Huang, T.; Yang, N.; Lu, C.; Qin, X.; Siddique, K.H.M. Soil Organic Carbon, total nitrogen, available nutrients, and yield under different straw returning methods. Soil Tillage Res. 2021, 214, 105171. [Google Scholar] [CrossRef]
64. Finn, D.R.; Lee, S.; Lanzén, A.; Bertrand, M.; Nicol, G.W.; Hazard, C. Cropping systems impact changes in soil fungal, but not prokaryote, alpha-diversity and community composition stability over a growing season in a long-term field trial. FEMS Microbiol. Ecol. 2021, 97, fiab136. [Google Scholar] [CrossRef]
65. Donn, S.; Almario, J.; Muller, D.; Moënne-Loccoz, Y.; Gupta, V.V.; Kirkegaard, J.A.; Richardson, A.E. Rhizosphere microbial communities associated with Rhizoctonia damage at the field and disease patch scale. Appl. Soil Ecol. 2014, 78, 37–47. [Google Scholar] [CrossRef]
66. De Boer, R.F.; Steed, G.R.; Kollmorgen, J.F.; Macauley, B.J. Effects of rotation, stubble retention and cultivation on take-all and eyespot of wheat in northeastern Victoria, Australia. Soil Tillage Res. 1993, 25, 263–280. [Google Scholar] [CrossRef]
67. Campbell, A.R.; Titus, B.R.; Kuenzi, M.R.; Rodriguez-Perez, F.; Brunsch, A.D.; Schroll, M.M.; Shepherd, J.N. Investigation of candidate genes involved in the rhodoquinone biosynthetic pathway in Rhodospirillum rubrum. PLoS ONE 2019, 14, e0217281. [Google Scholar] [CrossRef] [PubMed]
68. Guo, K.; Yang, J.; Yu, N.; Luo, L.; Wang, E. Biological nitrogen fixation in cereal crops: Progress, strategies, and Perspectives. Plant Commun. 2023, 4, 100499. [Google Scholar] [CrossRef] [PubMed]
69. Han, H.; Chen, C.; Bai, M.; Xu, T.; Yang, H.; Shi, A.; Li, J. Abundance and diversity of denitrifying bacterial communities associated with N2O emission under long-term organic farming. Eur. J. Soil Biol. 2020, 97, 103153. [Google Scholar] [CrossRef]
70. Garritano, A.N.; Song, W.; Thomas, T. Carbon fixation pathways across the bacterial and archaeal tree of life. PNAS Nexus 2022, 1, pgac226. [Google Scholar] [CrossRef]
71. Klein, T.; Poghosyan, L.; Barclay, J.E.; Murrell, J.C.; Hutchings, M.I.; Lehtovirta-Morley, L.E. Cultivation of ammonia-oxidising archaea on solid medium. FEMS Microbiol. Lett. 2022, 369, fnac029. [Google Scholar] [CrossRef]
72. Zheng, J.; Luan, L.; Luo, Y.; Fan, J.B.; Xu, Q.S.; Sun, B.; Jiang, Y.J. Biochar and lime amendments promote soil nitrification and nitrogen use efficiency by differentially mediating ammonia-oxidizer community in an acidic soil. Appl. Soil Ecol. 2022, 180, 104619. [Google Scholar] [CrossRef]

Xu C, Toh R, Li J, Zhou Y, Denton MD. Stubble Management Plays a Greater Role than Tillage Practice in Shaping Soil Microbiome Structure and Function. Agriculture. 2025; 15(2):143. https://doi.org/10.3390/agriculture15020143
Перевод статьи «Stubble Management Plays a Greater Role than Tillage Practice in Shaping Soil Microbiome Structure and Function» авторов Xu C, Toh R, Li J, Zhou Y, Denton MD., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык

















































































































Комментарии (0)