Кариотип верблюдов
Основная задача частной цитогенетики сельскохозяйственных животных заключается в изучении связи количественной и качественной изменчивости наследственных структур клеток с биологическими и хозяйственно-полезными признаками животных.
Хромосомы (от греческого chroma – цвет, краска, и греческого soma – тело) – органоиды клеточного ядра, являющиеся носителями генов и определяющие наследственные свойства клеток и организмов. Способны к самовоспроизведению, обладают структурной и функциональной индивидуальностью и сохраняют ее в ряду поколений. Каждый вид организмов обладает характерным и постоянным набором хромосом в клетке, закрепленным в эволюции данного вида, и его изменения происходят только в результате мутаций.
В кариотипе различают половые хромосомы, аутосомы, ядрышко - образующие хромосомы; у некоторых видов сельскохозяйственных животных и птиц могут существовать добавочные хромосомы, число которых непостоянно и которые не содержат генов, свойственных данному виду.
Кариотип (от греческого karyon – орех, ядро ореха и греческого typos – образец, форма), совокупность признаков хромосомного набора (число, размер, форма хромосом), характерных для того или иного вида. Постоянство кариотипа каждого вида поддерживается закономерностями митоза и мейоза. Описание хромосомного набора проводится на стадии метафазы или поздней профазы и сопровождается подсчетом числа хромосом, морфометрией, идентификацией центромеры (первичной перетяжки), ядрышкового организатора (вторичной перетяжки), спутника и т.д. Особенности строения хромосом выявляется дифференциальным окрашиванием.
Задачей цитогенетического мониторинга в верблюдоводстве является изучение хромосомного полиморфизма, оценка и прогнозирование распространения хромосомных мутаций с последующим наблюдением за фенотипом и его изменчивостью у животных основных пород используемых в племенном деле.
Задачей феногенетического мониторинга является обследование здорового поголовья, как правило, используемых в племенных репродукторах для дальнейшего воспроизводства стада, разработки общезоотехнических и феногенетических параметров животных конкретной породы.
В настоящее время в связи с ухудшением состояния окружающей среды цитогенетическим исследованиям млекопитающих придается пристальное внимание при оценке мутагенной опасности различных регионов.
При цитогенетическом мониторинге различных пород верблюдов анализ частоты геномных мутаций представляется очень важным, поскольку этот тип мутаций может возникать иными путями в сравнении с генными и хромосомными нарушениями.
Мутациями (от латинского mutatio – изменение, перемена), называют внезапные естественные или вызванные искусственно наследуемые изменения генетического материала, приводящие к изменению тех или иных признаков организма. От характера изменений в генетическом материале различают мутации: точковые, инверсии, хромосомные перестройки (аберрации) и мутации, заключающиеся в изменении числа хромосом.
Спонтанные мутации возникают как ошибки при воспроизведении генетического материала, поскольку редупликация не происходит с абсолютной точностью, а процессы репарации не обладают абсолютной эффективностью. Частота спонтанного мутирования у каждого вида верблюдов генетически обусловлена и поддерживается на оптимальном уровне.
Вопрос о вкладе различных мутаций в генетическую изменчивость различных пород верблюдов окончательно не решен. Основными типами мутаций являются изменения в числе или структуре хромосом, хромосомные мутации, изменения в структуре ДНК – генные мутации.
Хромосомные и генные мутации вызывают либо нарушение жизнеспособности и плодовитости, либо снижает устойчивость к болезням и продуктивность. Это связано с тем, что они приводят к нарушению деления клеток, нормального распределения хромосом между ними, изменяют ход синтеза белков, ферментов.
Мутации обычно разделяют на три типа
1) генные или точковые, связанные с изменениями в определенном локусе хромосомы, в результате чего из имеющего аллеля образуется новый.
2) структурные перестройки (хромосомные аберрации) когда затрагивается структура одной хромосомы.
3) Геномные связанные с изменениями числа хромосом (полиплоидия, анеуплоидия).
Хромосомные перестройки делятся на два основных типа:
1. Внутрихромосомные – изменения внутри одной хромосомы.
2. Межхромосомные – обмен участками между негомологичными хромосомами.
Внутрихромосомные перестройки представлены:
1) Нехватками – делеции, дефишенси. Делеция – потеря внутреннего участка хромосомы. Продуктом нехваток может быть образование колец. Большие нехватки вызывают эмбриональную смерть, малые могут вызвать фенотипический эффект, сходный с генными мутациями.
2) Дупликация – умножение отдельных участков хромосом несущие одни и те же гены. По фенотипическому эффекту дупликации во многих случаях сходны с точечными мутациями и являются одним из важных механизмом эволюции.
3) Инверсия – переворачивания внутреннего участка хромосом на 1800. Может быть парацетрическая – в переворачиваемый участок не входит центромера; перицентрическая – участвует центромера и близлежащие участки. Инверсии отводится важная эволюционная роль в дивергенции видов.
4) Внутрихромомсомные транслокации – перемещение участка хромосом из одного района в другой район той же хромосом.
Реципрокные транслокации – обмен участками хромосом.
Инверсия – внутрихромосомное перемещение хромосом материала без обмена и tr может быть в масштабе одного плеча, так и обоих.
К межхромосомным перестройкам относятся центрические (робертоновские) слияния (tr), реципрокные tr.
Центрическими слияниями называют слияние двух акроцентрических хромосом и имеет важное значение в эволюции животных.
Идентификация и классификация хромосомных нарушений непосредственно связаны с проблемой кариотипичесокй нормы. Анализ результатов многочисленных исследований хромосом человека показывает, что имеется большое число случаев отклонений от общепринятого кариологического стандарта, которые не приводят к фенотипическим отклонениям и к аномалиям. Изучение линейной дифференцированности хромосом выявило широкую вариабельность кариотипического полиморфизма у самых различных видов животных. Поэтому в настоящее время возникает необходимость пересмотра понятия кариологического стандарта. Однако, это требует получения надежной информации об особенностях распространения типов полиморфизма хромосом среди отдельных видов, пород и популяций животных, о наследственных их характеристиках, о связи кариотипического полиморфизма с фенотипическими аномалиями, особенно, с воспроизводительными качествами животных.
Исследование гетероплоидии у эмбрионов млекопитающих показывает, что излишек или недостаток отдельных аутосом неодинаково влияет на течение раннего эмбриогенеза. Гаплоидия считается непреодолимым барьером для прохождения эмбриогенеза у млекопитающих Смерть партеногенетических диплоидных эмбрионов у млекопитающих, возможно, объясняется нарушением взаимоотношения процессов, протекающих в матке в связи с подготовкой к имплантации, и характером развития самого зародыша.
Полиплоидия, в частности, тетраплоидия существенно замедляют пролиферативные процессы, что, вероятно, сказывается на течении морфогенеза. Поэтому, несмотря на то, что развитие плода может иногда достигать, относительно поздних стадий, смерть эмбриона является логическим концом таких случаев.
Трисомия аутосом – приводит, как правило, к раннему прекращению беременности. Более того, плодные пузыри вообще могут не содержать эмбриона, а иметь только зародышевые оболочки.
Моносомия аутосом вызывает нарушение процесса имплантации эмбриона и поэтому препятствует дальнейшему развитию.
Значительную долю всех хромосомных нарушений составляют числовые аномалии хромосом.
Мозаицизм – наличие клеток в организме с различным набором хромосом при условии, что все они ведут начало от одной зиготы.
Химеризм – наличие клеток с различным набором хромосом происходящие от двух и более зигот.
По нашим данным частота полиплоидии, анеуплоидии, являются устойчивым признаком и зависит как от продуктивности, так и используемых методов совершенствования продуктивных качеств животных.
Изучение числа и формы хромосом важно при гибридизации животных для разработки многих теоретических вопросов частной генетики и селекции.
Наибольший успех при гибридизации обычно сопутствует видам, схожим по числу и морфологии хромосом, например, зубр х бизон → F1 плодовитое. При гибридизации як х домашний скот → F ♂ бесплодны 2n=60, т.е. наблюдается ограниченная полом стерильность. Обычно стерильные гетерогаметные животные. По данным И.К. Шарипова при межвидовой гибридизации уриала (2n=58) х муфлона (2n=54) и домашних овец все гибриды плодовиты]. Мул (2 лошадь х ♂ осел), лошак (♀ осел х ♂ конь). Породы выведены межвидовой гибридизацией архаромеринос ( 56 – архар х 56 – овцы). Кидус гибрид соболя и куницы (2n=38). Среднеазиатская черная пестрая порода свиней (дикий кабан х свиноматка).
Межродовая гибридизация проводится в лабораторных условиях, и ее результаты скромны в сравнении с межвидовой гибридизацией.
Различают
полиморфизм:
— по числу хромосом, связанные с Rtr – рабертсоновскими трансклокациями (NF у всех
особей постоянно), по числу вариации добавочных хромосом.
— по хромосомным перестройкам, не изменяющимся числом хромосом (перицентрическая инверсия и др.), изменчивость гетерохроматина.
Установлены породные различия у крупного рогатого скота по особенностям распределения ЯОР в хромосомах. То есть полиморфизм является широко распространенным явлением у сельскохозяйственных животных.
Данные отечественных и зарубежных ученых показывают, возможность выбраковки животных в раннем возрасте по конституциональному кариотипическому статусу (ККС), конституциональной кариотипической изменчивости (ККИ), а также прогнозировать продуктивность по локусам хромосом. Племенная ценность сельскохозяйственных животных обычно определяется по способности передавать ценные качества потомству. Хотя есть данные о связи определенных продуктивных показателей с частотой хромосомных и геномных нарушений кариотипа, имеются данные о связи с фенотипом. В настоящее время прогнозирование продуктивности по кариотипу не нашло широкого применения, несмотря на положительные стороны отбора по кариотипу и его изменчивости.
Основные направления исследований в цитогенетике верблюдов и ряда видов сельскохозяйственных животных связаны с выявлением конститутивных цитогенетических аномалий, из которых широкое распространение получила Rtr – робертсоновские транслокации. Имеются три основные гипотезы образования Rtr – робертсоновские транслокации.
1. «Двойного разрыва и транслокации» в негомологичных акроцентрических хромосомах – образуются два разрыва, которые локализуются в одной хромосоме проксимально, а в другой дистально от центромеры. Затем в результате реципрокной транслокации из двух акроцентриков образуется одна метацентрическая хромосома и небольшой центрический фрагмент, который вскоре утрачивается.
2. «Прямого соединения акроцентриков» своими короткими плечами, при этом центромеры обеих хромосом сливаются в один общий участок, функционирующий в дальнейшем как единый кинетокор (Matthey R., l963, 1965) здесь указано и на центрическое разделение хромосом.
3. «Притяжения между гомологичными участками негомологичных хромосом – причем центрическое слияние можно рассматривать как крайнюю степень хромосомной ассоциации. Это прочное соединение возникает, если притяжение гетерохроматиновых районов коротких плеч акроцентриков, составляющих общую хромосомную ассоциацию, достигает максимума и преодолевает силы отталкивания, существующие между теломерами негомологичных аутосом.
Притяжение гетерохроматиновых районов негомологичных хромосом обусловлено локализацией в них генов, контролирующих синтез рибосомной ДНК (Ohno S., 1969).
У носителей транслокации в мейозе возможны 3 типа расхождения перестроенных хромосом 1-й – обе гаметы получают сбалан-
сированный набор хромосом (одна полностью сбалансированный, другая – условно сбалансированный с транслокацией); 2-й – несбалансированный набор (одна гамета с дупликацией хромосом, другая – с делецией по одной из хромосом, вовлеченная в транслокацию); 3-й – несбалансированный набор (одна гамета с делецией хромосом, другая – с дупликацией). Поэтому от животных с транслокацией наряду с нормальным потомством, время от времени (и от потомства с условно сбалансированным кариотипом) можно ожидать особей с генетическим дефектом. Метод контроля производителей по качеству потомства в этом случае мало эффективен, т.к. вредное действие несбалансированного набора может проявляться не сразу, а через поколение.
Геномные и хромосомные мутации в сравнении с генными мутациями встречаются на порядок чаще и могут быть выявлены с меньшей затратой времени, сил и средств. Случаи конституциональной кариотипической изменчивости обнаруживаются достаточно редко. В то же время у любого животного с нормальным кариотипом и фенотипом можно выявить клетки с различными аномалиями числа и структуры хромосом. Количество таких клеток с нарушениями у отдельных особей колеблется в довольно широких пределах.
В связи с тем, что такие аномалии числа и структуры хромосом различны у одного и того же животного, нельзя утверждать об общей природе происхождении таких клеток. Такая кариотипическая изменчивость называется неконституциональной. Ее источником является спонтанный прижизненный мутагенез в соматических тканях. Смысл изучения неконституциональной кариотипической изменчивости хромосом в соматических тканях состоит, во-первых, в контроле спонтанного мутагенеза, которому подвергаются сами изучаемые животные, что дает возможность своевременно выявлять и освобождать селекционное стадо от носителей высокого уровня хромосомных нарушений.
Анализ спонтанной кариотипической нестабильности необходимо начинать с изучения анеуплоидии в различных популяциях чистопородных верблюдов, разводимые в различных областях Республики Казахстан.
Анеуплоидия относится к довольно распространенному типу кариотипических изменений. Она образуется вследствие нерасхождения хромосом или хроматид во время митоза или мейоза, а также элиминации поврежденных хромосом. Качественными производными этих нарушений являются образовавшиеся гиподиплоидные и гипердиплоидные клетки. Частота образования анеуплоидных клеток, как правило, находится под генетическим контролем. Повышенная частота анеуплоидии имеет связь с ухудшением воспроизводительных функций и с различными заболеваниями.
Превышение частоты гиподиплоидных клеток над частотой гипердиплоидных клеток у верблюдов трудно объяснить только естественными причинами. Большая часть гиподиплоидных клеток однозначно имеет артефактное происхождение и связана с техническими приемами при обработке культур и приготовлением препаратов хромосом. Действительно, манипуляции, применяемые при приготовлении препаратов хромосом, например, центрифугирование и особенно гипотонизация, могут повышать частоту гиподиплоидных клеток. Интенсивное разбрасывание хромосом по предметному стеклу культивированных клеток лейкоцитов крови, предварительно набухших в гипотонической среде, также может приводить к утере части хромосомного набора. Довольно часто при микроскопировании кариотипа некоторые хромосомы обнаруживаются недалеко от основной метафазной клетки. Однако, установить принадлежность отсутствующих или дополнительных хромосом к какой-либо паре затруднительно вследствие сложности их идентификации. Потеря хромосом может иметь и другую природу, как результат элиминации поврежденных хромосом. В случае гипердиплоидных клеток абсолютно исключена вероятность механического проникновения добавочных хромосом, утерянных из других клеток. В связи с этим, необходимо отметить, что основным механизмом образования анеуплоидии является нерасхождение хромосом в мейозе или митозе. Тогда число возникших вследствие этого гиподиплоидных клеток должно быть равным числу гипердиплоидных, так как, если одна дочерняя клетка получила лишние хромосомы, то другая, естественно, останется с нехваткой этих хромосом. Следовательно, за критерий истинной анеуплоидии можно принять число гиперплоидных клеток, умноженное на два.
Как генетический феномен, полиплоидия состоит в кратном увеличении гаплоидного числа хромосом в ядрах клеток. Есть все основания считать, что появление полиплоидных клеток связано с восстановительными процессами, регенерацией, функциональной активностью органов и тканей. При исследовании голштинского скота и его помесей наблюдается общая закономерность – влияние генотипа производителя на частоту полиплоидии у потомства. Поэтому изучение влияния генотипа быков на частоту полиплоидии у дочерей является необходимым при кариотипировании клеток культивированных лимфоцитов крови.
Помимо вышеперечисленных спонтанных хромосомных нарушений, нередко в кариотипе животных обнаруживаются такие аномалии, как хроматидные и изохроматидные пробелы, разрывы, делеции и образующиеся в результате этого различные фрагменты генетического материала. У изученных пород верблюдов при анализе кариотипа культивированных лимфоцитов крови довольно часто регистрировались пробелы, хроматидные и изохроматидные разрывы, фрагменты, делеции и множественные разрывы.
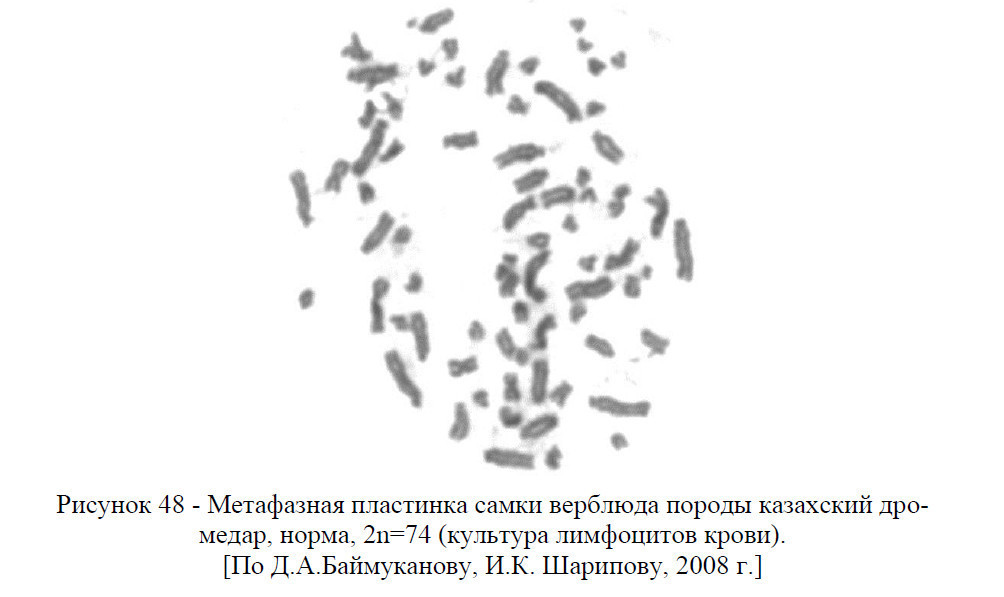
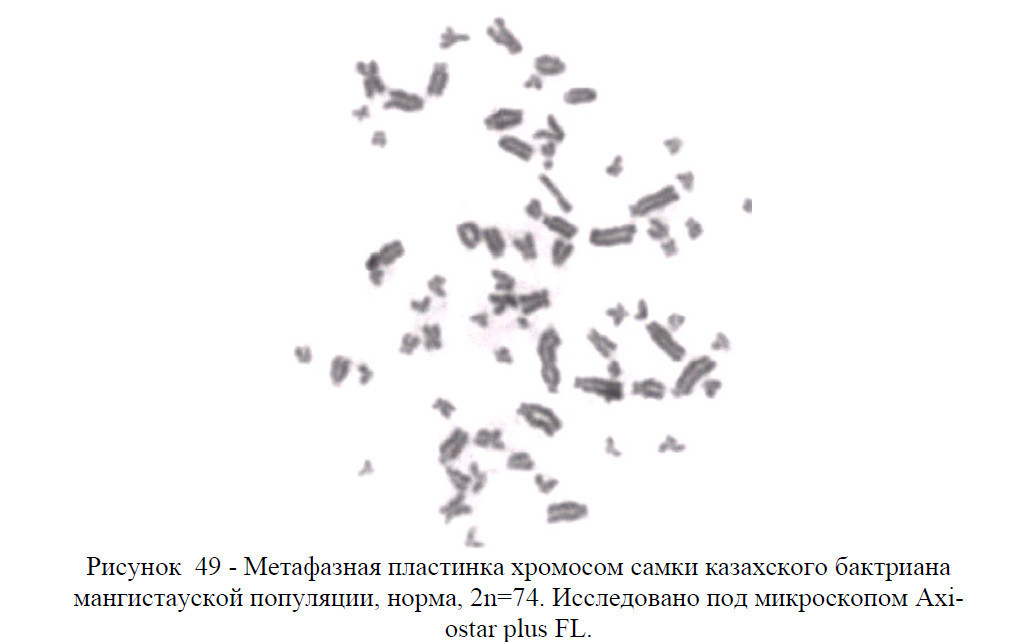
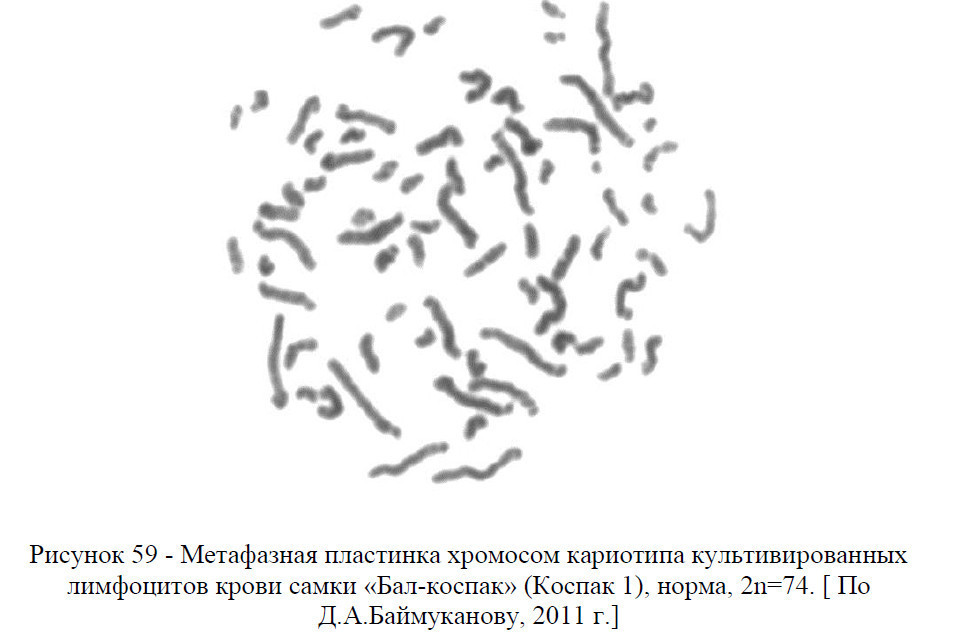
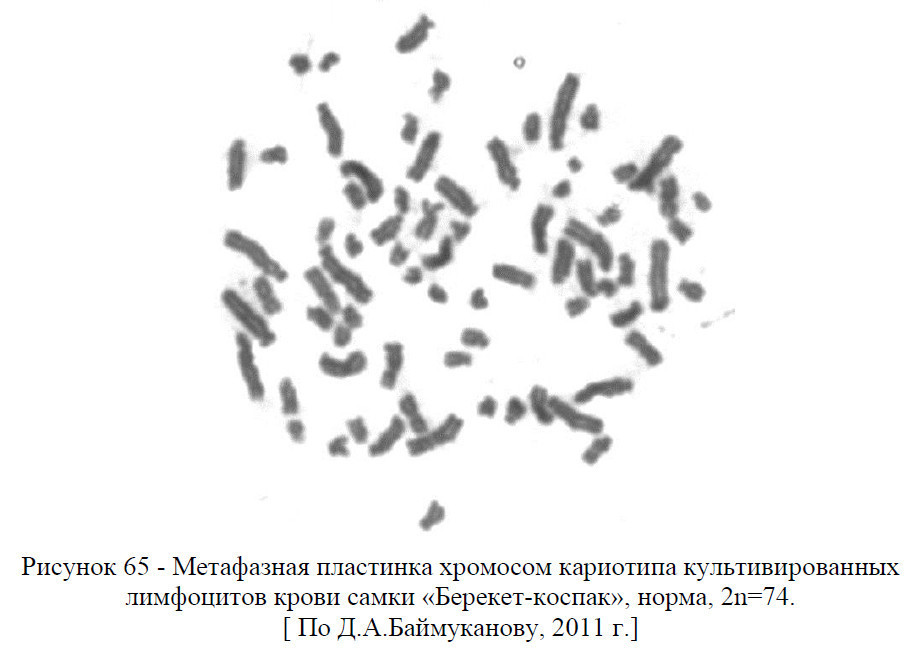
У верблюдов кариотип представлен 74 хромосомами, из них 12 метацентрические аутосомы, 60 акроцентрические аутосомы, ХХ (у самок) и ХУ (у самцов) половые хромосомы – гоносомы. То есть, кариотип – это набор хромосом соматической клетки свойственный тому или иному виду животных или растении.
В кариотипе верблюдов на основании размеров хромосом и положения центромер четко выделяются две группы хромосом: 30 пар аутосом представляют постепенно убывающий по размерам ряд акроцентриков разной величины и 6 пар аутосом являются небольшими метацентрическими хромосомами.
В кариотипе самок самая крупная пара метацентриков опознавалась как половая Х-хромосома, а у самцов самая крупная непарная метацентрическая хромосома также является Х-хромосомой, а самая мелкая (по видимому, метацентрик) – У-хромосомой.
Формулу кариотипа домашнего верблюда можно представить следующим образом:
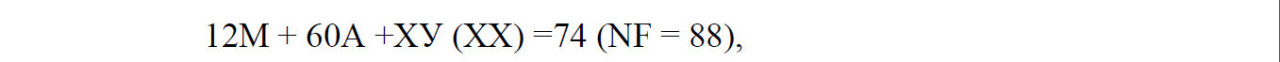
У некоторых крупных акроцентриков были хорошо выражены короткие плечи, но у большинства аутосом этого типа центромеры расположены почти терминально. Акроцентрические хромосомы по своим размерам образуют ряд постепенно убывающих величин, в связи с чем, их индивидуальная идентификация при использовании обычных методов окраски не всегда возможна (рисунки 41-65).
С учетом распределения хромосом по размерам и положения центромеры, нами предлагается следующая классификация хромосом верблюдов:
Группа «А» – Крупные акроцентрики – 6 пар. Хорошо выражены у всех пар короткие плечи. Относительные размеры 6,324,0%.
Группа «В» – Крупно-средние акроцентрики – 9 пар. Короткие плечи заметны только у некоторых крупных акроцентриков. Относительные размеры – 4,16-2,40%.


















Группа «С» – Средне-малые акроцентрики – 15 пар. Короткие плечи хромосом выявлены не у всех пар. Относительные размеры – 2,43-0,54%.
Группа «М» – Метацентрики – 6 пар. Относительные размеры – 3,07-1,01%.
Группа половых хромосом – Х и У
Половые хромосомы верблюдов идентифицируются у самцов: две их хромосомы не имеют гомологов, при этом одна из них идентична двум гомологам хромосом самок.
Индивидуальные особенности дифференциально g-окрашенных хромосом кариотипа верблюдов
При идентификации хромосом кариотипа верблюдов практикуется - С и - G дифференциальное окрашивание хромосом. В результате дифференциальной окраски хромосом образуются - С и - G – полосы. Механизм дифференциальной окраски заключается в том, что компоненты краски связываются исключительно с ДНК хромосомы. Если удалить ДНК, эффект исчезает. Это связывание ступенчато: сначала с ДНК реагирует циклическая молекула метиленового синего, затем молекула эозина с последующим взаимодействием этих молекул между собой и формированием красящего компонента (комплекса). Разная степень или прочность связи красителя с ДНК зависит и от особенностей конфигурации и взаиморасположения ДНК в хромосоме.
При дифференциальной окраске каждая хромосома приобретает свой специфический рисунок - чередование светлых и темных полос, отражающих функциональную различную активность отдельных районов хромосом.
Окрашенные участки хромосом - это низкоактивные в генетическом отношении гетерохроматиновые районы хромосом, а активные неокрашенные эухроматиновые районы.
Таким образом, следует отметить, что G-полосы выявляются после специальных обработок. При этом в хромосомах видны поперечные оптически более плотные (темные) и менее плотные жизненно важные транскрипционные гены с умеренными повторами
С-полосы – это околоцентромерный гетерохроматин, не содержит транскрипционные гены, и представляет собой участки хромосом с высокой повторностью нуклеотидов.
ЯОР – это участки хромосом, в которых сосредоточены кластеры генов кодирующих синтез рРНК.
Ядрышко – представляет собой сближенные участки хромосом, содержащие большое число кластеров рибосомных генов, на которых путем транскрипции образуются молекулы рРНК, потом поступают в цитоплазму и входят в рибосомы.
Микрофотографирование позволяет детально изучить морфологию, подсчитать число хромосом в метафазной пластинке, измерить каждую из них.
Парижская конференция (1989) рекомендовала следующие основные типы дифференциальной окраски при микрофотографировании и последующего анализа кариотипа:
Q - окраска и полосы, выявляемые флуоресцентными красителями (акрихин);
G - окраска и полосы. Окраска хромосом красителем Гимзы после воздействия (трипсин, раствор солей и t°С). Наиболее информативный метод;
С - окраска и полосы выявляются в районе центромеры. Показано, что в этих областях хромосомы находятся гетерохроматин, содержит ДНК;
N - метод, выявляющий ЯОР в хромосомах ядрышка в интерфазном ядре.
При Q и G - окраске затрагиваются одни и те же участки хромосом, ярко флуоресцирующий сегмент при Q окраске соответствует темноокрашенные G-полосы.
Изучение числа и формы хромосом важно при гибридизации животных для разработки многих теоретических вопросов частной генетики и селекции.
В цитогенетических исследованиях хромосомных мутации соматических клеток млекопитающих необходимо дальнейшее применение новейших методов дифференциальной окраски, в частности, выявления ядрышкообразующих районов (ЯОР) хромосом и структурного гетерохроматина (S-бэндинг).
Дифференциальная окраска хромосом на Гимза-полосы существенно расширила возможности индивидуальной идентификации хромосом. Установлено, что каждая хромосомная пара обнаруживает свой специфический рисунок поперечной исчерченности по Гимза-полосам, что позволило подбирать пары хромосом с достаточно высокой достоверностью.
В качестве диагностических признаков при опознавании хромосом использовали расположение, число и степень интенсивности окраски полос, причем по этим признакам видовых и межпородных отличий у изученных верблюдов не обнаружено.
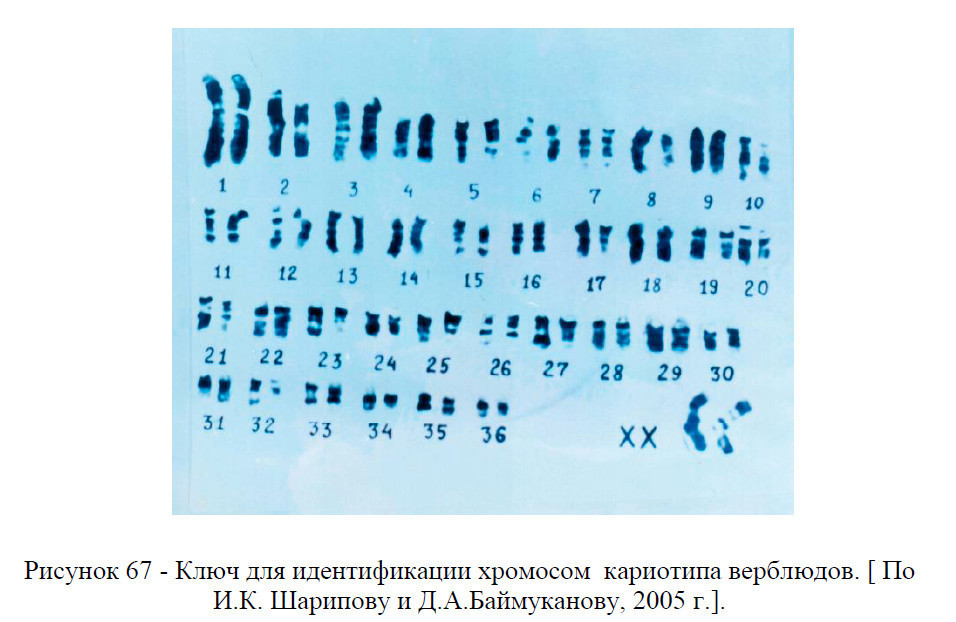
В приводимом ниже описании хромосом азур-положительные полосы фигурируют как темные, а азур-отрицательные – как светлые (негативные) участки хромосом (рисунок 66, 67).
Хромосомы № 1-3. Крупные хромосомы со сходным рисунком узких положительных полос, равномерно чередующихся по всей длине хромосом. Хромосома №2 выглядит более однородно окрашенной по сравнению с другими хромосомами. В этой хромосоме в ее проксимальной и дистальной частях полосы более сближены, чем в хромосомах №1 и №3 и образуют блоки положительных полос. Хромосома №3 имеет четкую интенсивно окрашенную полосу в прицентромерном районе. Отличать хромосомы этой группы от остальных хромосом кариотипа можно по их крупным размерам и наличию коротких вторых плеч, но индивидуально хромосомы идентифицируются с трудом.
Хромосома № 4. По рисунку полос схожи с хромосомой №5, отличаясь от нее лишь более тесной сближенностью позитивных полос.
Хромосома №5. Прицентромерный район слабо окрашен. В проксимальной и дистальной части хромосомы расположены по четыре положительные интенсивно окрашенные полосы, разделенные по середине хромосомы светлым участком.
Хромосома № 6. Прицентромерный район положительно окрашен. Далее следует светлый участок и слабо окрашенная положительная полоса. За ней расположены, пять темных полос средней степени окрашенности. Теломерный район светлый.
Хромосома № 7. В прицентромерном районе находится узкая темная полоса. Далее по всей длине хромосомы расположены четыре симметричных блока положительных полос, в каждом блоке можно различить три-четыре узких темных полос.
Хромосома №8. Отделенным сравнительно широким светлым промежутком, в проксимальной и дистальной части хромосомы расположены два блока интенсивно окрашенных полос: в проксимальной части из 4 -5 узких полос, в дистальной – из 2-3-х полос. Теломерный участок окрашен.
Хромосома № 9. Хромосома в целом выглядит положительно окрашенной. На общем темном фоне слабо еле заметны 10-12 узких полос. Прицентромерная светлая полоса сходна с таковой в хромосоме №10. Трудная для идентификации хромосома.
Хромосома № 10. Хромосома отличается от сходной по рисунку полос хромосомы №11 наличием более широкой светлой полосы в прицентромерном районе, но это различие не всегда очевидно.
Хромосома № 11. По всей длине хромосомы примерно на равном расстоянии друг от друга расположены 8-9 полос средней степени окрашенности.

Хромосома № 12. Прицентромерный район слабо окрашен. Далее по направлению к дистальной части хромосомы видны три блока положительных полос средней степени окраски: в проксимальной, срединной и дистальной части хромосомы.
Хромосома № 13. Начиная с центромеры и кончая срединной частью хромосомы, хорошо видны пять темных полос одинаковой ширины. Далее видны две близко расположенные темные полосы, после которых располагается протяженный светлый район, включающий и теломеру.
Хромосома № 14. Прицентромерный район темно окрашен. Далее следует неширокий светлый участок, сменяемый отчетливой темной полосой, за которой после узкого светлого промежутка расположен блок из 3-4 узких темных полос. За ним следует заметный светло окрашенный участок и темная узкая полоса. Теломерный район светлый.
Хромосома № 15. Прицентромерная темная полоса сменяется широким светлым участком. Далее находится блок из двух-трех темных тесно сближенных полос, за которым снова следует широкая светлая полоса. В дистальной части видны две узкие темные полосы, сменяемые светлым теломером.
Хромосома №16. В хромосоме хорошо видны две группы положительных интенсивно окрашенных полос – в проксимальной и дистальной части хромосомы. В каждой группе различимы три крупные полосы. Группы полос разделены между собой хорошо заметным светлым участком.
Хромосома № 17. В прицентромерном районе находится положительная полоса средней степени окраски. Далее следует сравнительно широкий светлый участок и четыре неширокие положительные полосы, разделенные такими же по ширине светлыми участками. Теломерный район слабо окрашен.
Хромосома № 18. Для хромосомы характерна серия из 6-7 положительных полос средней степени окрашенности, расположенных примерно на равном расстоянии друг от друга.
Хромосома № 19. В прицентромерном районе видна интенсивно окрашенная положительная полоса. Далее следует неширокий светлый участок и три положительные полосы средней степени окрашенности, расположенные примерно на равном расстоянии друг от друга. Теломерный район светлый.
Хромосома № 20. Хромосома в целом выглядит темноокра- шенной. На общем положительном фоне выделяются более интенсивно окрашенные полосы в срединной, дистальной части и в при- центромерном районе. Хромосома идентифицируется с трудом.
Хромосома № 21. В проксимальном районе заметны три- четыре узкие полосы разной степени окрашенности. В дистальном районе виден блок из трех-пяти сближенных полос, которые не всегда различимы. Проксимальная и дистальная части хромосомы разделены ясно заметным светлым промежутком. Трудная для идентификации хромосома.
Хромосома № 22. В прицентромерном районе видна темная полоса средней степени окрашенности. Далее следует сравнительно широкая светлая полоса, сменяемая тремя узкими положительными полосами средней степени окраски. Теломерный участок короткий, положительно окрашен.
Хромосома № 23. Прицентромерная узкая полоса интенсивно окрашена. Далее следует сравнительно широкая полоса, сменяемая нешироким положительно окрашенным бэндом средней интенсивности. За ним следует довольно широкий светлый участок, сменяемый тесно сближенными полосами, которые часто сливаются в одну широкую полосу. Теломерный участок однородно слабо окрашен.
Хромосома № 24. Рисунок полос в хромосоме сходен с таковым в хромосоме №25, за исключением более широкого однородно светло окрашенного района. Хромосома идентифицируется не очень легко.
Хромосома № 25. Темная узкая прицентромерная полоса сменяется светлым участком такой же ширины. Далее в большинстве случаев видна узкая положительная полоса средней или слабой интенсивности, за которой следует более широкая негативная полоса. За ней располагаются три полосы примерно одинаковой ширины: темная, негативная и снова темная, но меньшей интенсивности. Прителомерный район светло окрашен.
Хромосома № 26. Прицентромерная полоса интенсивно окрашена. За ней следует положительная полоса меньшей интенсивности, расположенная дистально. Далее находится негативная полоса, сменяемая наиболее широким темным бэндом, отделенным от дистального конца довольно большим негативным прителомерным районом.
Хромосома № 27. В прицентромерном районе видна интенсивно окрашенная темная узкая полоса, далее за нешироким негативным участком расположены две интенсивно окрашенные узкие темные полосы (часто сливающиеся в одну). За ними следует широкая светлая полоса, в которой иногда различима положительная полоса слабой интенсивности окраски, расположенная в середине хромосомы. В дистальной части находится неширокая темная полоса. Теломерный район слабо окрашен.
Хромосома № 28. Хромосома отличается от сходной по рисунку полос хромосомы №29 тем, что прицентромерный район окрашен в виде четкой темной полосы.
Хромосома № 29. В прицентромерной области расположена четкая темная полоса. Далее следует сравнительно широкий светлый участок, прерываемый двумя близко расположенными полосами средней степени окрашенности. Теломерный район слабо окрашен.
Хромосома № 30. Прицентромерная полоса темная или средней интенсивности окраски, за которой следует не всегда различимая очень узкая полоса. Далее располагается светлый участок и примерно в середине плеча хорошо заметная темная полоса. Дистальная половина хромосомы однородно негативная, кроме неширокого слабо окрашенного прителомерного района.
Хромосома № 31. Прицентромерная узкая полоса интенсивно окрашена. Далее следует четкая негативная широкая полоса и примерно в средней части хромосомы два узких тесно сближенных бэнда средней интенсивности окраски. В прителомерном районе видна узкая слабо окрашенная полоса.
Хромосома № 32. В прицентромерном районе видны две четкие интенсивно окрашенные темные полосы. Далее при внимательном анализе в дистальной части хромосомы заметны две узкие слабо окрашенные полосы. Теломер короткий и негативный.
Хромосома № 33. Для хромосомы характерны две положительные полосы средней интенсивности окраски, расположенные вблизи центромеры и в ее дистальной части. Между ними находится заметный бледно окрашенный район.
Хромосома № 34. В хромосоме видна одна темная полоса, расположенная в проксимальной части плеча. Теломерный район слабо окрашен.
Хромосома № 35. В хромосоме видны две положительные полосы средней интенсивности окраски, расположенные вблизи центромеры и в ее дистальной части. Между ними находится отчетливо негативный район. Теломерный район негативно окрашен.
Хромосома № 36. Положительная средней интенсивности окраски полоса расположена примерно в середине хромосомы. Теломерный участок негативный, короткий, похож на центромеру.
Половые хромосомы. Х-хромосома – крупная непарная хромосома, по размерам сходная с шестой или седьмой хромосомами. Характерной чертой хромосомы является наличие срединной темной полосы, ограниченной от проксимального и дистального блока положительных полос отчетливым светлым участком. У-хромосома – очень мелкая хромосома овальной формы. В У-хромосоме при очень внимательном рассмотрении можно различить две слабо окрашенные узкие позитивные полосы – в проксимальной и дистальной частях хромосомы. Трудно идентифицируемая хромосома. В целом, в хромосомном наборе верблюдов достаточно уверенно распознаются 20-25 пар гомологичных хромосом (в зависимости от качества полученных препаратов и степени длины хромосом).
С целью облегчения идентификации хромосом верблюдов на практике нами разработан специальный определитель хромосом – ключ (рисунок 67) для идентификации хромосом верблюдов (нумерация хромосом дана в убывающем по их размерам порядке).
1. Крупные акроцентрические
хромосомы со сходным рисунком узких положительно окрашенных полос, равномерно
расположенных по всей длине хромосомы. В хромосоме №2 полосы в ее дистальной
части и проксимальной области более сближены по сравнению с
хромосомами №1 и №3. Часто заметны вторые короткие
плечи………1-3
(цифры
после ряда точек указывают на номера хромосом).
2. Крупные хромосомы, в проксимальной и дистальной частях которых расположено по 4 темноокрашенных полосы. Одна из хромосом (№4) отличается от другой более тесной сближенностью позитивных полос, что видно при внимательном рассмотрении………4-5
3. Хромосома легко распознается
по четко окрашенной полосе в прицентромерной
области ………6
4. Легко идентифицируемая хромосома по наличию 4-х симметричных блоков положительных полос ………7
5. Довольно крупная хромосома распознается по наличию широкого светлого участка в срединной части………8
6. В темноокрашенной хромосоме при внимательном рассмотрении можно заметить 10-12 узких полос………9
7. В растянутых хромосомах по всей длине выделяются 8-9 полос, в более спирализованных хромосомы объединены в 4 крупных блока, причем в хромосоме №11 два срединных блока более тесно сближены, чем в хромосоме №10………10-11
8. Три хромосомы средней величины. В первой из них видны три блока полос, во второй – пять и в третьей хромосоме, в ее срединной части 3-4 узкие темные полосы ……… 12-13-14
9. Распознавание хромосом возможно методом исключения и с помощью схемы полос ………15-30
10. Самая крупная из метацентрических аутосом. В средней части хромосомы видны две узкие сближенные полосы ………31
11. В прицентромерном районе метацентрической хромосомы видны две четко окрашенные темные полосы………32
12. В хромосоме имеются две положительные полосы: вблизи центромеры и в ее дистальной части ………33
13. В проксимальной части плеча видна одна темная полоса ………34
14. Хромосома сходна по рисунку полос с хромосомой №33, но меньше ее по размерам ………35
15. Примерно в средней части хромосомы расположена одна темная полоса………36
Половые хромосомы:
Крупная непарная
субметацентрическая хромосома. В срединной части имеется темная полоса,
ограниченная от проксимального и дистального блока отчетливым светлым
промежутком………Х
Очень мелкая непарная хромосома овальной формы………У
Как показали наши исследование ЯОР (Ag NOR) в кариотипе верблюдов локализованы в тех хромосомах, которые являются гомологичными по картине G-полос с соответствующими хромосомами, то есть наблюдается видовой консерватизм по числу ЯОР и их локализации. В 50 изученных метафазных пластинках хромосом верблюдов казахского бактриан 2 (4%) имели 5 ЯОР на клетку, 6 (12%) - 6 ЯОР на клетку, 19 (38%) - 7 ЯОР на клетку, 17 (34%) - 8 ЯОР на клетку, 2 (4%) - 9 ЯОР на клетку и 4 (8%) - 10 ЯОР на клетку.
Морфометрическая характеристика хромосом кариотипа верблюдов
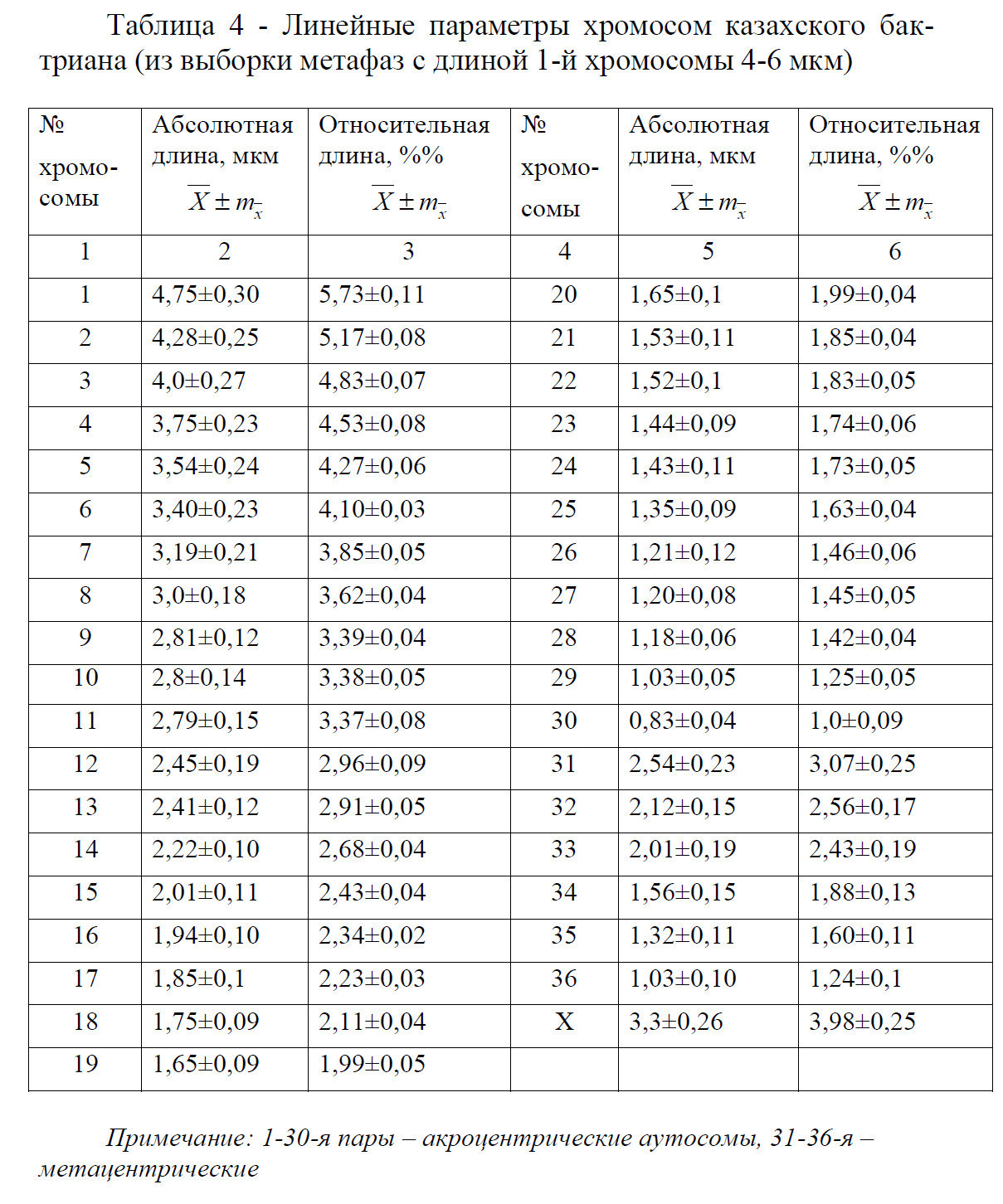
При определении линейных параметров хромосом верблюдов разных пород выявлена общая закономерность, указывающая на внутри - индивидуальную изменчивость абсолютных размеров хромосом, обусловленная различной степенью их спирализации. В кариотипе верблюдов казахского бактриана при сильной спирализа- ции больше всего укорачиваются аутосомы группы А и В сравнении С и М, связанная с так называемым эффектом дифференциальной спирализации хромосом (таблица 4).
В метафазных пластинках с длиной 1-й хромосомы менее 4 мкм размеры 1-й, 2-й и 3-й хромосом почти одинаковые ввиду проявления эффекта дифференциальной спирализации, а пластинки с длиной 1-й хромосомы более 6 мкм довольно редко встречаются.
По абсолютным размерам четко отличаются хромосомы в группе «А» (1-6). В группе «В» (7-15) прослеживается равномерное уменьшение хромосом. В группе «С» (15-30) хромосомы не четко отличаются по абсолютным размерам, то есть при сильной дифференциальной спирализации морфометрический анализ их затруднен. Хромосомы группы «М» (31-36) также как и в группе «В» равномерно уменьшаются.
Определены абсолютная и относительная длина хромосом верблюдов разных пород с учетом принятой нами классификации хромосом кариотипа по группам (А, В, С, М, Х).
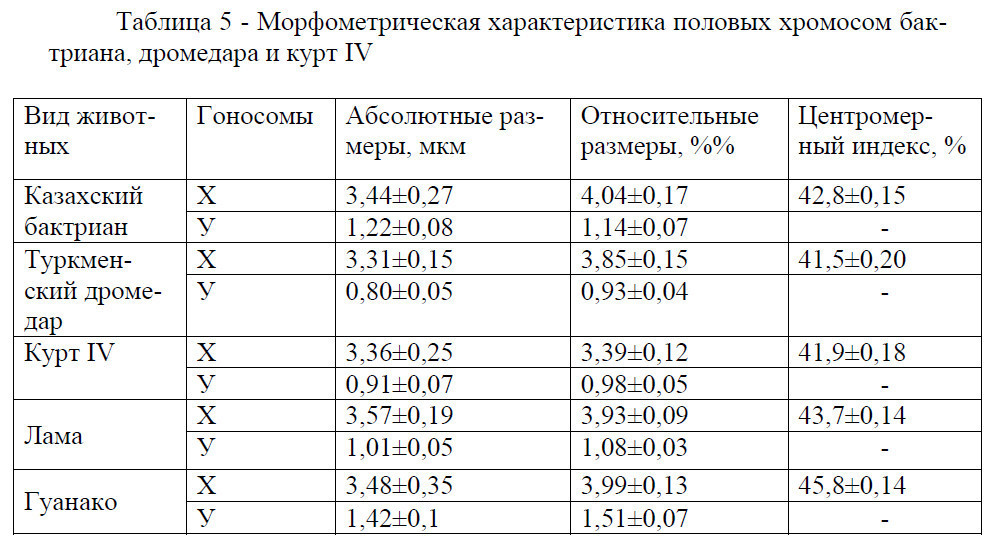
Различия по длине между Х-хромосомами между туркменским дромедаром и казахским бактрианом достигает 4%, между курт IV и бактрианом почти 10% (таблица 5).
Центромерный индекс Х-хромосомы составляет у бактриана казахской породы 42,8±0,15%, дромедара туркменской породы 41,5±0,20% и курт IV 41,9±0,18%. То есть по абсолютной и относительной длине, а также центромерному индексу Х-хромосом у изученных животных существенной разницы не установлено (Р<0,90).
Проведенный морфометрический анализ хромосом верблюдов разных пород (к гаплоидному женскому набору) не позволили выявить полиморфизм по длине хромосом как внутри породы, так и между сравниваемыми группами.
Cпектр хромосомных аберрации и геномных нарушении кариотипа культивированных лимфоцитов крови верблюдов
Вопрос о вкладе различных мутаций в генетическую изменчивость верблюдов еще окончательно не решен. Изучение хромосомных аномалий имеет большое значение в селекционно-племенной работе с целью профилактики распространения нежелательных мутаций.
Некоторые мутаций, возникшие у верблюда в единичном случае, распространяются в некоторых популяциях в результате эффекта родоначальника. Мутантные гены распространяются в популяциях в течении длительного времени. Появление особей с мутантными генами обусловлено сегрегацией генов в соответствии с законами Г.Менделя. Поэтому часть генетического груза, обусловленная передающимися и расщепляющимися в потомстве генами, называются сегрегационным грузом. Часть носителей появляются в результате вновь возникающих в каждом новом поколении мутаций. Мутации генов, возникающие заново в каждом поколении, составляют мутационный груз.
У верблюдов выявлены некоторые мутаций, которые существенно не влияют на их приспособленность, и, поэтому не исключено широкое распространение в популяциях. Одним из ценных свойств мутаций является случайный характер их возникновения. Не следует путать случайность с беспричинностью.
В массовых цитогенетических исследованиях рекомендуем определять частоту гетероплоидных клеток. В воспроизводстве животных необходимо использовать взрослые особи, имеющие низкую частоту гетероплоидии в соматических клетках.
При изучении спонтанных хромосомных аберрации в соматических клетках необходимо определить: тип и частоту спонтанных хромосомных аберраций, генетический риск образования аномальных клеток по методике Д.А.Баймуканова и др.; частоту клеток с хромосомными аберрациями хромосомного и хроматидного типа в сравнительном аспекте с общей частотой; преобладающие хромосомных аберраций с частотой более 5%.
Для цитогенетических исследований необходимо отбирать ме- тафазные пластики с числом хромосом соответствующие видовой норме с погрешностью в сторону уменьшения 2 и увеличения 1, что позволит достоверно определить конституциональный кариотипический статус и изменчивость кариотипа.
Анеуплоидия – изменение числа хромосом, некратное гаплоидному набору. Анеуплоидия представляет собой добавление или потерю одной или двух хромосом диплоидного набора (рисунки 6871).
Основной механизм возникновения анеуплоидии нерасхождение и потери отдельных хромосом в митозе и мейозе. Анеуплоидия приводит к понижению жизнестойкости и нередко к гибели анеуплоидов, особенно у животных (анеуплоидия лежит в основе ряда хромосомных болезней).
Относительно числа гиподиплоидных клеток мы считаем, что большинство из них являются артефактами, вызванными техническими манипуляциями. То есть, истинным показателем анеуплоидии служит число гипердиплоидных клеток, которые мы рекомендуем учитывать при определении показателя генетической анеуплоидии. У сельскохозяйственных животных обычно частота гиподиплоидных клеток выше гипердиплоидных.
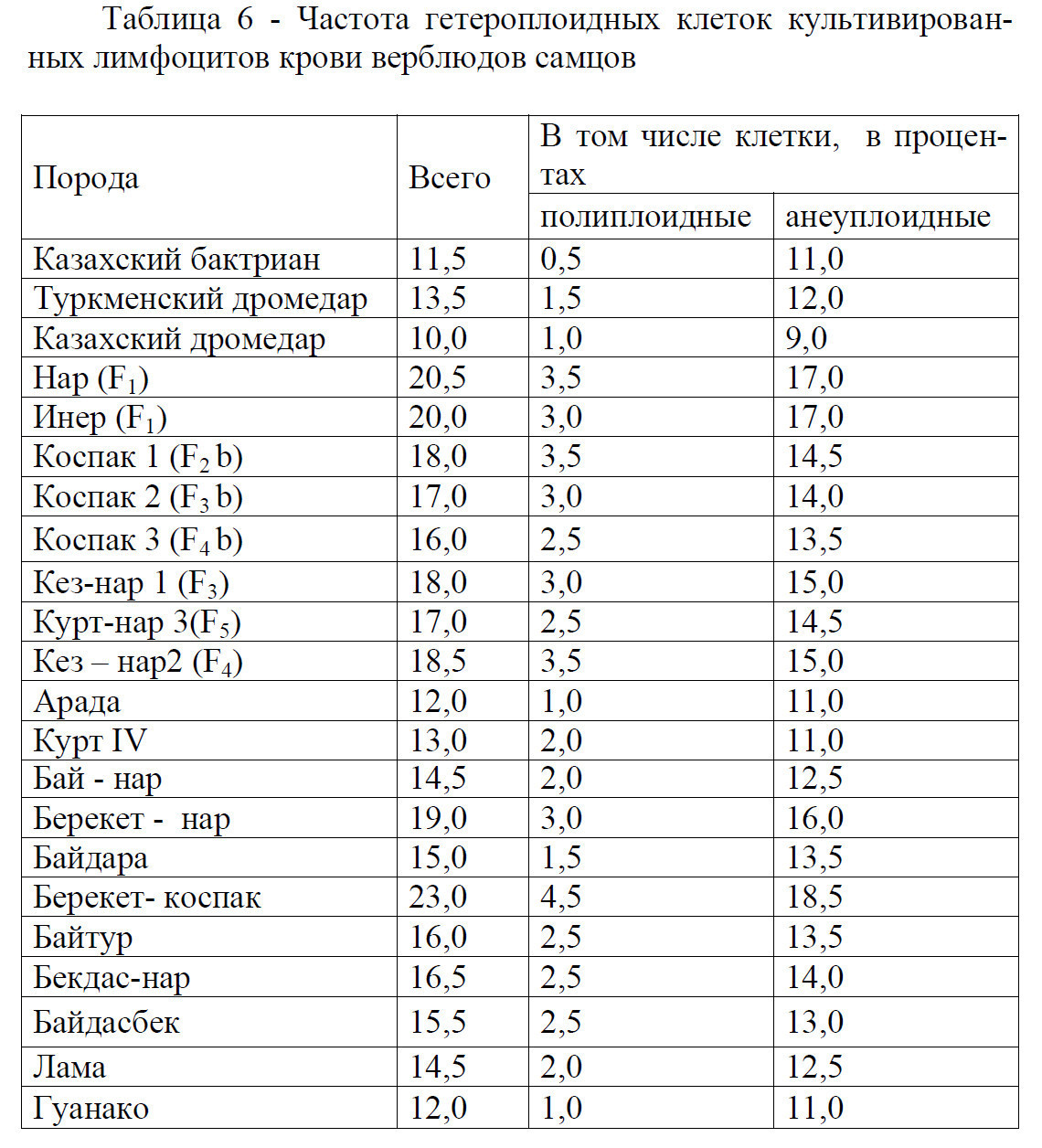
В исследовании Д.А.Баймуканова было установлено, что частота образования анеуплоидных клеток у гибридных самцов выше в сравнении с чистопородными казахскими бактрианами (таблица 6).
Полиплоидия - это геномная мутация, заключающаяся в увеличении числа хромосом, кратного к гаплоидному набору.
Полиплоидия – увеличение числа полных хромосомных наборов в четное и нечетное число раз. У верблюдов зарегистрированы триплоидия (3n) и тетраплоидия (4n) (рисунки 72, 73).
Имеются данные о том, что у верблюдов казахской породы бактрианов с увеличением удоя наблюдается повышение образования анеуплоидных клеток, в частности гипердиплоидных клеток.
У верблюдов выявлена обратная зависимость уровня хромосомных аберрации у матерей и их верблюжат. Причины этого до сих пор не выявлены.
Можно предположить, что причиной обратной зависимости уровня хромосомных аберрации у матерей и их верблюжат являются: во-первых, презиготическая селекция на уровне гамет, направленная на отбор клеток с низким уровнем мутаций; во-вторых, с возрастом в результате давления мутагенов число клеток с аберрациями возрастает; в третьих возможность разной чувствительности организма матери и плода к действию мутагенных факторов.






При межвидовой гибридизации достоверно увеличивается частота анеуплоидных клеток на 70% в сравнении с казахскими бактрианами и на 30% в сравнении с туркменскими дромедарами. Общая частота полиплоидных клеток у нар-мая выше по сравнению с казахскими бактрианами в 1,7 раза, с туркменскими дромедарами в 1,2 раза.
Частота гетероплоидных клеток культивированных лимфоцитов крови оказалась выше у межвидовых гибридных самцов (14,523,0%) в сравнении с казахскими бактрианами (11,5%), туркменскими дромедарами (13,5%), казахскими дромедарами (10,0%) и гибридными самцами арада (12,0%).
В таблице 7 приведены сведения о динамике генетического риска образования аномальных клеток в культуре лимфоцитов крови верблюдов разных генотипов. Генетический риск образования аномальных клеток (ГРОАК) у казахских бактрианов составил - 7,6±0,01%, туркменских дромедаров -7,7±0,09%, нар - мая F1 - 12,0±0,44%, коспак F2-11,0±0,19%, кез-нар F3-10,0±0,36%.
Транслокации хромосом. Основные направления исследования кариотипа млекопитающих связаны с выявлением цитогенетических аномалий, из которых широкое распространение получили Rtr-робертсоновские транслокации.
Транслокации – взаимные обмены между негомологичными хромосомами (рисунок 74). При образовании обычной транслокации происходит разрыв хромосомы в двух местах и обмен участками.
Робертсоновские транслокации – особый тип транслокаций, который приводит к изменению числа хромосом. Робертсоновские транслокации могут приводить как к слиянию акроцентрических хромосом в метацентрические, так и к разделению метацентрических хромосом на акроцентрические в области центромеры Центрические слияния (Робертсоновские транслокации) представляют собой слияние двух негомологичных акроцентрических хромосом с образованием одной субметацентрической хромосомы. При разделении, наоборот, одна субметацентрическая хромосома делится на две акроцентрические хромосомы. При этом должна образоваться новая центромера, иначе хромосома без центромеры будет потеряна при митозе.
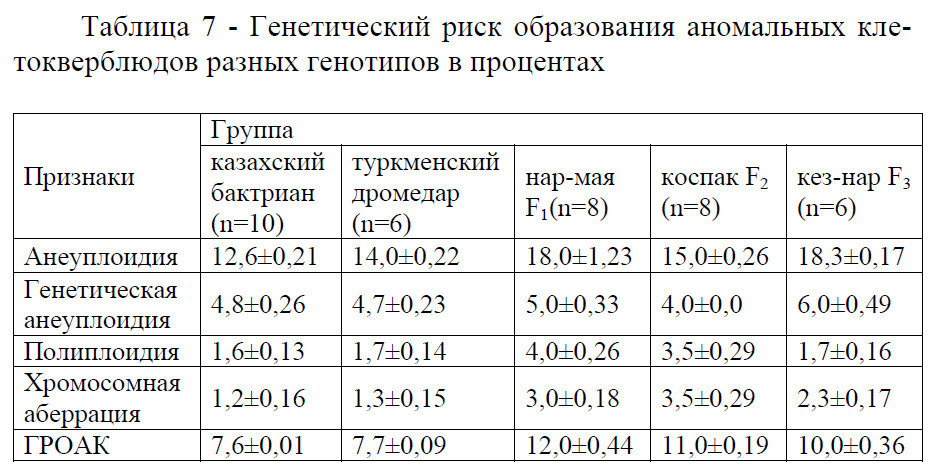
Робертсоновские перестройки приводят к изменению числа хромосом в кариотипе, не влияя на общее количество генетического материала в клетке. Робертсоновские транслокации в природе встречаются часто, и поэтому являются одним из основных путей эволюции кариотипа.
Установлено, что у носителей транслокации в мейозе возможны три типа расхождения перестроенных хромосом. Первый – обе гаметы получают сбалансированный набор хромосом (одна полностью сбалансированный, другая – условно сбалансированный с транслокацией). Второй – несбалансированный набор (одна гамета с дупликацией хромосом, другая – с делецией по одной из хромосом, вовлеченная в транслокацию). Третьи – несбалансированный набор (одна гамета с делецией хромосом, другая – с дупликацией). Поэтому от животных с транслокацией наряду с нормальным потомством, время от времени (и от потомства с условно сбалансированным кариотипом) можно ожидать особей с генетическим дефектом. Метод контроля производителей по качеству потомства в этом случае мало эффективен, т.к. вредное действие несбалансированного набора может проявляться не сразу, а через поколение.
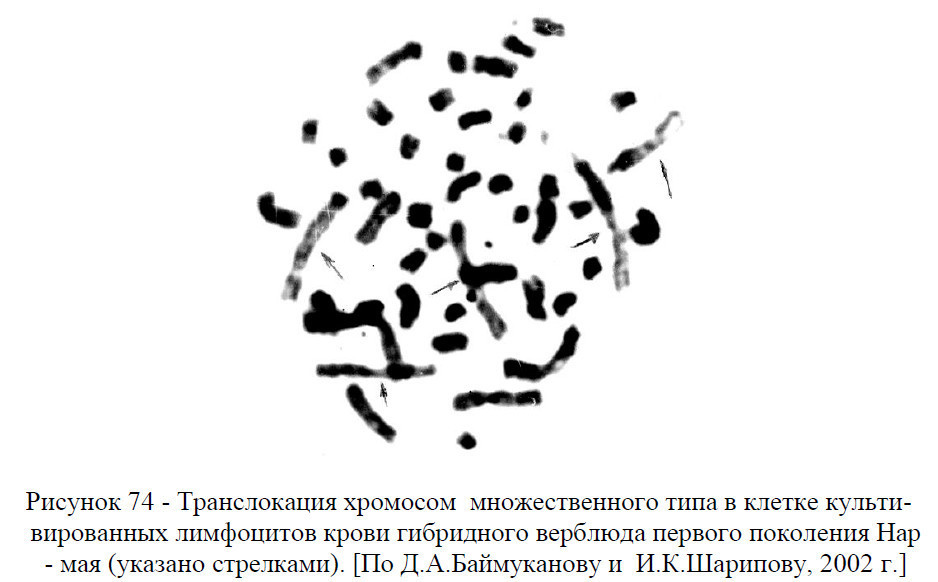
Частота и типы хромосомных аберрации. Индивидуальный учет частоты и типа хромосомных аберраций, культивированных клеток лейкоцитов крови верблюдов и ламы (таблица 8, рисунок 75-76) позволил достоверно идентифицировать одиночные и парные фрагменты, ацентрические кольца и разрывы в центромере (рисунки 77-82). Из всех выявленных хромосомных аберраций лишь разрыв в центромере влияет на плодовитость верблюдов. Влияние других типов хромосомных мутаций на продуктивность, воспроизводительную способность верблюдов не установлено.



В практическом плане цитогенетические исследования хромосомных мутаций в соматических клетках млекопитающих позволяет, во-первых, своевременно проводить выбраковку животных по результатам комплексного цитогенетического контроля, во-вторых, идентифицировать отцовские и материнские хромосомы у межвидовых гибридов, в третьих проводить раннюю цитогенетическую оценку кариотипа молодняка выращиваемые для племенного использования.
В таблице 8 нами приводятся данные по частоте клеток с хромосомными аберрациями у изученных верблюдов самцов.

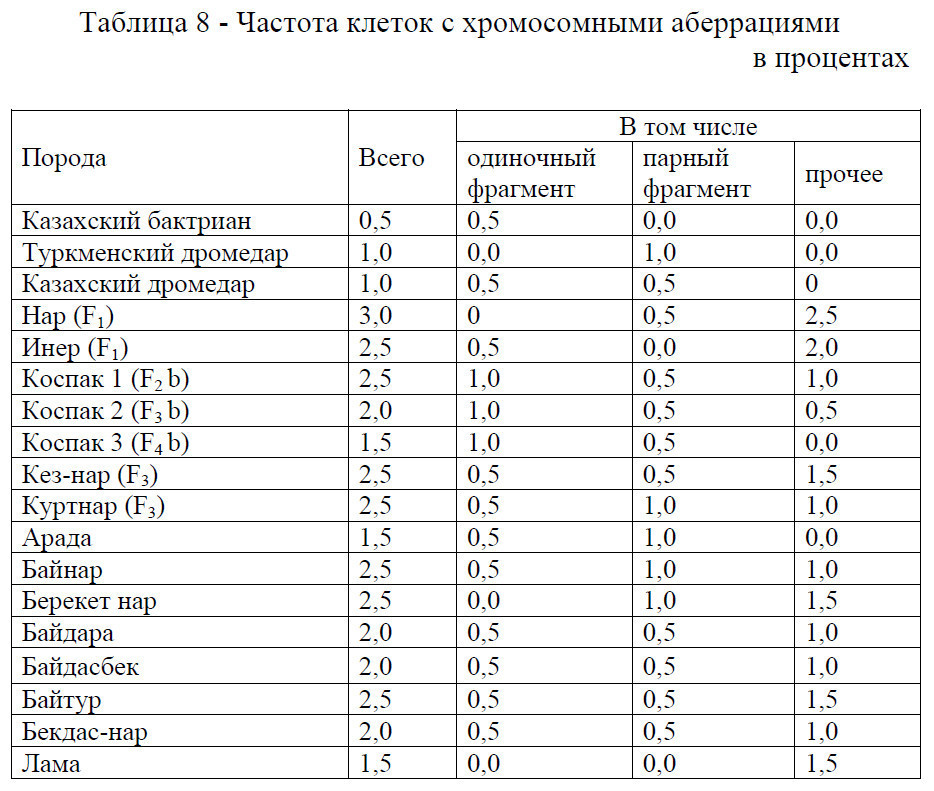

Установлено, что видовой особенностью является наличие у казахских бактрианов одиночного фрагмента (0,5%), туркменских дромедаров парного фрагмента (1,0%). Общее количество аберраций хромосом, дано нами на основании прямого микроскопического анализа пораженных клеток лейкоцитов периферической крови.

Послесловие
На основании, многолетних, цитогенетических исследований установлено, что в культивированных клетках лимфоцитов крови общая частота клеток с хромосомными аберрациями для всех групп верблюдов в среднем составляет 1,52±0,4%. В общем спектре аберраций перестройки хромосомного типа уступали хроматидным (36,9% и 63,1%).
Выявлена положительная корреляция по числу анеуплоидных клеток у верблюдоматок и их верблюжат: r=0,53 при tr=1,36. Частота клеток с хромосомными аберрациями составляет у верблюдома- ток от 0,5% до 2,0% (в среднем 1,6%), верблюжат от 0% до 4,0% (в среднем 2,8%).
Коэффициент корреляции по частоте хромосомных аберраций между верблюдоматками и верблюжатами отрицательный r=-0,50 при tr=1,36. У верблюдов казахской породы бактрианов хромосомные аберрации в клетках культивированных лимфоцитов крови преимущественно формируются за счет нестабильных перестроек, такие как парные и одиночные фрагменты, обмены хромосомного и хроматидного типов, которые при последующих делениях элиминируются.
У верблюдоматок частота образования анеуплоидных клеток составляет от 6,0 до 12,0% (в среднем 10,0%), а их верблюжат – от 6,0% до 10,0% (в среднем 8,8%).
Установлено, что чем выше молочная продуктивность, тем выше физиологическая гиподиплоидия. У молодых животных физиологическая гиподиплоидия достоверно ниже, чем у возрастных лактирующих верблюдоматок (P<0,05). Частота образования генетически анеуплоидных клеток выше у низкопродуктивных верблю- доматок (8,0%) в сравнении с высокопродуктивными (4,0%) и в среднем по стаду. Однако, у высокоудойных верблюдоматок выше частота образования физиологически гиподиплоидных клеток (8,0%), в то время как средние показатели составляют 6,8%, а у низкопродуктивных верблюдоматок – 6,0% и ниже.
Генетический риск образования аномальных клеток у верблю- доматок в среднем составляет 8,0%, а у верблюжат – 11,2%. Численно генетический риск образования аномальных клеток у верблюдов достоверно выше, чем выход физиологически гиподиплоидных клеток.
Результаты анализа влияния различных вариантов подбора чистопородных казахских бактрианов, туркменских дромедаров породы Арвана на цитогенетические показатели показывает эффективность использования верблюдов-производителей, с низкой и средней частотой хромосомных и геномных нарушении кариотипа в спаривании с верблюдоматками для получения потомства с оптимальной кариологической нормой (частота %): анеуплоидные клетки (10-12); полиплоидные клетки (1-2); клетки с хромосомными аберрациями (1-2); генетический риск образования аномальных клеток (6-10).
На основании проведенного цитогенетического мониторинга верблюдов казахского бактриана разработан способ отбора верблюдов включающий дополнительную цитогенетическую оценку и отбор по оптимальной кариологической норме (Предварительный патент РК на изобретение №16357). Доказана эффективность отбора и подбора родительских пар по цитогенетическому статусу. Потомство, полученное от цитогенетически отобранных верблюдов, имеют удой молока 1700 кг с жирностью 5,2%, настриг шерсти 7,0 кг и 100%-ную оплодотворяемость, а сверстницы полученные традиционным способом селекции соответственно 1200 кг - 5,3% - 6,0 кг - 70%.
AgNOR в кариотипе верблюдов локализованы в тех хромосомах, которые являются гомологичными по картине G-полос с соответствующими хромосомами, то есть наблюдается видовой консерватизм по числу ЯОР и их локализации, причем чаще всего встречаются пластинки с 7-8 ЯОР (ядрышкообразующиеся районы).
Генетические методы исследований позволяют своевременно выявлять как генетические аномалии, так и большое число генетических маркеров с различными функциями. Поэтому развитие так называемой маркер-опосредованной селекции (MAS-селекция – marker-assisted selection) в верблюдоводстве является перспективным направлением. Анализируя молекулы ДНК, определяя их полиморфизм, изучая гены и локусы селекционер может отбирать высокоценных особей не только по фенотипу, но и по генотипу. Секвенирование геномов отечественных пород верблюдов позволяет надеяться, что генетический мониторинг обогатится методами относительно нового направления генетики – геномики, изучающей геном, индивидуальные гены и их экспрессию.
Генетический мониторинг позволяет сделать обоснованный выбор и определить оптимум и пределы допустимых изменений. В дальнейшем накопленные знания по структурным генам, полило- кусным спектрам ампликонов ДНК, мутационной изменчивости у малочисленных популяции отечественных пород верблюдов могут стать незаменимыми источниками информации при фундаментальных геногеографических исследованиях и выборе генетических стратегий для программ по сохранению пород и дальнейшей селекции новых генотипов верблюдов Казахстана с заданными признаками.
При сохранении пород in situ преследуется задача сохранить сбалансированную систему генов обуславливающих развитие уникальных экстерьерных и продуктивно-биологических особенностей, которые обусловливают фенотипические породные характеристики. Именно перечисленные в данной монографии особенности, отличающие чистопородные, помесные и гибридные генотипы верблюдов показывают необходимость их сохранения и увеличения численности в генофондных хозяйствах.
Без соответствующего контроля, систематического наблюдения, разработки методов прогноза, четких критериев и эффективных средств оценки состояния генофонда пород верблюдов невозможно смоделировать процесс разведения и выбрать оптимальную программу сохранения in situ редких исчезающих и малочисленных генотипов. Генетический мониторинг может носить локальный (стадо, популяция в породе) или глобальный характер (контролируется все породное разнообразие как внутри республике, так и в сопредельных государствах).
Литература
1. Боголюбский С.Н. Происхождение верблюдов // Верблюдоводство. - Алматы - Москва: Казахское краевое изд-во,1934.-С. 57-71.
2. Лакоза И.И. Верблюдоводство. Москва, 1953.- 312 с.
3. Кугенев П.В. Верблюдоводство.- Москва, 1982. – 87 c.
4. Fowler M.E. Evolutionary histоry and defferences between camelids and ruminants // Journal of Camel practice and research, 1997. -V 4. - Рр.99-105
5. Баймуканов Д.А. Эволюция и систематическое положение верблюдов //Верблюдоводство Казахстана XXI века. – Алматы: Ба- стау,2009. –С.12-13.
6. Баймуканов Д.А., Баймуканов А. Генетика, эволюция и систематика верблюдов (полное издание) //Монография. – Шымкент: Полиграф, 2011. -117с.
7. Тимофеев - Ресовский Н.В., Яблоков А.В., Глотов Н.В. Очерк учения о популяции. - Москва,1973. -277с.
8. Тимофеев - Ресовский Н.В., Воронцов Н.Н.. Яблоков А.В. Краткий очерк теории эволюции. – Москва: Наука, 1977. -301с.
9. Шмальгаузен И.И. Факторы эволюции (теория стабилизирующего отбора). - Москва: АН СССР,1946. -396с.
10. Оно С. Генетические механизмы прогрессивной эволюции. – Москва: Мир, 1973. -227с.
11. Наумов Н.П. Экология животных. - Москва: Наука, 1963. - 618с.
12. Грант В. Эволюция организмов. – Москва: Мир, 1980. - 407с.
13. Патент РК на изобретение 13740 // Способ отбора верблюдов казахского дромедара для селекции. Опубл. 15.12.2006, бюл.№12 (Баймуканов А., Турумбетов Б.С., Баймуканов Д.А.).
14. Айала Ф. Введение в популяционную и эволюционную генетику. – Москва: Мир, 1984. -230с.
15. Баймуканов Д.А., Баймуканов А. Цитогенетика верблюдов (альбом). – Алматы: Светоч, 2011. – 97.
16. Баймуканов Д.А., Баймуканов А. Генетика, эволюция и систематика верблюдов (полное издание) //Монография. – Шымкент: Полиграф, 2011. -117с.
17. Баймуканов Д.А. Цитогенетика и селекция двугорбых, одногорбых верблюдов и их гибридов. – Алматы: Бастау, 2002. -160 с.
18. Леватин Р.К. Генетические основы эволюции. – Москва: Мир, 1978. -351с.
19. Рэфф Р., Кофмен Т. Эмбрионы, гены и эволюция, -Москва: Мир, 1986.-402с.
20. Шмальгаузен И.И. Значение корреляций и эволюция животных // Сб. тр. Ин-та эволюц. Морфологии животных посв. Памяти академика А.Н.Северцова. – Москва, 1939. – Т.1. –С.175. – 230.
21. Шмальгаузен И.И. Естественный отбор и информация //Известия АН СССР, - Москва, 1960. Серия биологическая. - №1. – С.19-38.
22. Шишкин М.А. Фенотипические реакции и эволюционный прогресс //Экология и эволюционная теория. – Ленинград: Наука, 1984. –С.196-216.
23. Способ селекции чистопородных туркменских дромедаров //Предварительный патент РК №14891. - Опубл. 15.10.2004, бюл. №10.
24. Способ отбора верблюдов казахского дромедара // Патент РК на изобретение №13740. - Опубл. 15.12.2006, бюл. №12.
25. Способ получения гибридных верблюдов Коспак // Патент РК на изобретение №14890. - Опубл. 15.10.2008, бюл. №10.
26. Способ выведения гибридных верблюдов «Арада» // Патент РК на изобретение №15452. - Опубл. 15.07.2009,бюл. №7.
27. Способ выведения гибридных верблюдов «Байдара» // Патент РК №15884. - Опубл. 15.07.2009, бюл. №7.
28. Способ выведения гибридных верблюдов «Бай-нар» //Предварительный патент РК №15885. - Опубл. 12.07.2005, бюл. №7.
29. Способ выведения гибридных верблюдов «Берекет-нар» //Патент РК № 16748. Опубл. 15.01.2010, бюл. №1.
30. Способ гибридных верблюдов «Кез-нар» //Патент РК №14148. - Опубл. 15.08.2008, бюл. №8.
31. Способ получения гибридных верблюдов «Курт-нар» // Патент РК №14147. - Опубл. 15.07.2009, бюл. №7.
32. Способ селекции гибридных верблюдов мясо-молочного направления // Патент РК №14246. - Опубл. 15.08.2008, бюл. №8.
33. Баймуканов Д.А. Селекция верблюдов породы казахский бактриан и методы их совершенствования. – Алматы: Бастау, 2009. -280с.
34. Способ выведения гибридных верблюдов «Байдасбек» //Патент РК №23600.. – Опубл. 15.12.2010, бюл №12.
35. Способ выведения гибридных верблюдов «Байтур» // Патент РК №23602. –Опубл. 15.12.2010, бюл №12.
36. Способ выведения гибридных верблюдов «Бекдас - нар» //Патент РК №23601. – Опубл. 15.12.2010, бюл №12.
37. Баймуканов Д.А. Изучение хромосомных наборов верблюдов Казахстана //Ж.Вестник сельскохозяйственной науки Казахстана. - Алматы: Бастау, 1996. -№5. -С.124-128.
38. Баймуканов Д.А. Изучение возможности использования цитогенетических методов для раннего прогнозирования племенных и продуктивных качеств верблюдов и их воспроизводительных способностей (Шифр 84.95.ФН) //Отчет о НИР за 1995г заключительный /ГР №0196РК00207. Инв.№0296РК00357. –Шымкент: РФПиВВ «Camel», 1996. -12с.
39. Баймуканов Д.А. Цитогенетические методы прогнозирования племенных и продуктивных качеств верблюдов //Ж.Новости науки Казахстана: научно-технический сборник. – Алматы: КазГо- сИНТИ, 1997. –Вып.2. –С.73-74.
40. Баймуканов Д.А., Шарипов И.К. Цитогенетические исследования верблюдов Казахстана (Шифр 269.96.ФН) //Отчет о НИР за 1996-1997гг заключительный /ГР №0196РК00207. Инв.№0297РК00575. – Шымкент: РФПиВВ «Camel», 1997. -65с.
41. Баймуканов Д.А. Түйенің кариотипі туралы //Ж.Жаршы. - Алматы: Бастау, 1998. -№3. -С.23-25.
42. Баймуканов Д.А., Шарипов И.К. Кариологические нарушения хромосом казахского бактриана созакской популяции и влияние их на фенотип //Роль молодых ученых в развитии пустынного животноводства и аридного кормопроизводства /Матер. межд. научн.- практ. конф. молодых ученых аграриев, посв.10-летию независимости РК. -Шымкент, 2001. -С.71-74.
43. Баймуканов А., Баймуканов Д.А., Шарипов И.К. Разработка и внедрение прогрессивных биотехнологических приемов повышения воспроизводительной способности верблюдов (Шифр 08.05.07) //Отчет о НИР за 1996-2000гг заключительный /ГР №0198РК00220. –Шымкент: НАЦАИ (КазНИИК), 2001. -28с.
44. Баймуканов Д.А. Эволюция, экология распространения и систематическое положение рода Camelus (аналитический обзор) //Ж.Поиск. Серия естественных и технических наук. - Алматы: Высшая школа Казахстана, 2002. -№1. -С.108-119.
45. Шарипов И.К., Ахметова Ж.Ш., Беккулов Х.Б., Баймуканов Д.А., Баймуканов А., Зайтбеков Е. Цитогенетическое исследование гуанако //Ж.Вестник сельскохозяйственной науки Казахстана. - Алматы: Бастау, 2002. -№11. -С.39-41.
46. Баймуканов Д.А. Цитогенетика и селекция двугорбых, одногорбых верблюдов и их гибридов //Монография (ISBN9965-13- 382-4). -Алматы: Бастау, 2002. -160с.
47. Баймуканов Д.А. Семейный анализ частоты хромосом аберраций у верблюдов //Каракулеводство, верблюдоводство и аридное кормопроизводство /Сб.науч.трудов КазНИИК. - Алматы: Бастау, 2003. -Т.24. -С.171-174.
48. Шарипов И.К., Графодатский А.С., Баймуканов Д.А., Бай- муканов А., Зайтбеков Е. Идентификация хромосом кариотипа верблюдов //Ж.Поиск. Серия естественных и технических наук. - Алматы: Высшая школа Казахстана, 2003. -№3 (2). -С.108-111.
49. Баймуканов Д.А., Зайтбеков Е.Д., Шарипов И.К., Баймұқанов А. Қазақ дромедары боталарының цитогенетикалық ерекшеліктері //Ж.Жаршы. - Алматы: Бастау, 2003. -№12. -С.16-18.
50. Баймуканов Д.А., Баймуканов А., Алиханов О., Татибеков А., Есбай С., Зайтбеков Е., Шарипов И.К. Изменчивость молочной продуктивности верблюдоматок казахской породы бактрианов и цитогенетическая аттестация ремонтного молодняка //Ж.Поиск. Серия естественных и технических наук. - Алматы: Высшая школа Казахстана, 2004. -№1 (2). -С.124-132.
51. Баймуканов Д.А., Шарипов И.К., Баймуканов А., Зайтбеков Е. Взаимосвязь цитогенетических показателей верблюдов казахского бактриана с их продуктивными особенностями //Ж.Поиск. Серия естественных и технических наук. - Алматы: Высшая школа Казахстана, 2004. - №1 (2). -С.116-124.
52. Баймуканов Д.А., Зайтбеков Е.Д., Баймұқанов А., Шарипов И.К. Қазақ бактрианы тұқымы түйелерінің кариологиялық көрсеткіштері және өнімділігі //Ж. Жаршы. - Алматы: Бастау, 2004. -№2. - С.28-30.
53. Баймұқанов Д.А., Шарипов И.К., Баймұқанов А., Зайтбеков Е. Camelidae тұқымдарының цитогенетикалық ерекшеліктері. //Ж.Жаршы. - Алматы: Бастау, 2004. - №7. - С.17-19.
54. Баймұқанов Д.А., Шарипов И.К., Баймұқанов А., Зайтбеков Е.Д. Түйе кариотипіндегі AgNOR //Ж. Жаршы. - Алматы: Бастау, 2005. -№5. - Б.12-15.
55. Баймуканов Д.А., Алибаев Н.Н., Шарипов И.К., Баймуканов А., Зайтбеков Е.Д. Способ приготовления культуры лейкоцитов для препаратов хромосом верблюдов //Описание изобретения №13840 /Промышленная собственность Казахстана. - Опубл. 15.08.2006, бюл. №8. -4с.
56. Баймуканов Д.А. Түйелер селекциясындағы цитогенетика. //Ж.Поиск: Серия естественных и технических наук. - Алматы: Высшая школа Казахстана, 2006. -№4. -С.112-130.
57. Баймуканов Д.А. Цитогенетическая структура кариотипа верблюдов Казахстана //Современное состояние и перспективы развития зоотехнической науки и практики животноводства: Матер. межд. науч.-практ. конф. (Шымкент, 23-24 ноября 2007 г.). – Шымкент: Жебе 2007. – С.225-230.
58. Баймуканов Д.А., Зайтбеков Е.Д. Қазақ бактрианы тұқымы түйелерінің селекциясында кариологиялық көрсеткіштерді пайдалану //Современное состояние и перспективы развития зоотехнической науки и практики животноводства: Матер. межд. науч.-практ. конф. (Шымкент, 23-24 ноября 2007 г.). – Шымкент: Жебе 2007. – С.230-231.
59. Баймуканов Д.А. Селекция верблюдов породы казахский бактриан южно-казахстанского типа молочной продуктивности //Автореферат диссертации на соискание ученой степени доктора сельскохозяйственных наук (06.02.01). –Шымкент: ЮЗНПЦСХ, 2007. - 46с.
60. Баймұқанов Д.А, Зайытбеков Е.Д. Түйе кариотипінің хромосомалық және геномдық бұзылуы //История и перспективы развития каракулеводства в Казахстане: Матер. междун. науч.– практ. конф. (г. Шымкент, 5-6 декабря 2008 г). – Шымкент: Жебе, 2008. С. 81-83.
61. Баймұқанов Д.А, Зайытбеков Е.Д. Түрлі генотиптегі түйелерде аномальды жасушалар генетикалық қауіптің пайда болуы //История и перспективы развития каракулеводства в Казахстане: Матер. междун. науч.-практ. конф. (г. Шымкент, 5-6 декабря 2008 г). – Шымкент: Жебе, 2008. С. 84-85.
62. Баймуканов Д.А., Зайытбеков Е.Д. Түраралық будан туйелердің кариотиптерінің цитогенетикалық құрылымы // Проблемы экологии, аридного кормопроизводства и животноводства в Казахстане: Матер. межд. науч.-практ. конф.(г. Шымкент, 26 марта 2009г.) – Шымкент: Жебе, 2009. –С.214-216.
63. Баймуканов Д.А., Алибаев Н.Н., Шарипов И.К. Гетероплоидия и хромосомные аберрации кариотипа чистопородных и гибридных верблюдов //Аграрная наука – сельскохозяйственному производству Казахстана, Сибири и Монголии: Труды ХII-ой межд.науч. – практ. конф.(г. Шымкент, 16-17 апреля 2009г.). - Алматы: Бастау, 2009. –Т.2. (Животноводство). – С.237239.
64. Баймуканов Д.А., Зайытбеков Е.Д. Таза тұқымды қазақ бактрианы түйелерінің цитогенетикалых картасы //Аграрная наука – сельскохозяйственному производству Казахстана, Сибири и Монголии: Труды ХII-ой межд. науч. – практ. конф. (г. Шымкент, 16-17 апреля 2009г.). - Алматы: Бастау, 2009. –Т.2. (Животноводство). – С.239-242.
65. Баймуканов Д.А., Баймуканов А., Турумбетов Б.С. Генефонд верблюдов Казахстана //Верблюдоводство Казахстана ХХI века (к 70-летию профессора Асылбека Баймуканова). – Алматы: Бастау, 2009. –С.20-34.
66. Баймуканов Д.А. Селекция верблюдов породы казахский бактриан и методы их совершенствования //Монография (ISBN9965-413-90-8). –Алматы: Бастау, 2009.-280 с.
67. Баймуканов Д.А. Актуальные проблемы изучения хромомомных мутации в соматических клетках млекопитающих (аналитический обзор) //Ж.Поиск:Серия естественных и технических наук.-Алматы:ВШК, 2010.-№3. –С.22-30.
68. Баймуканов Д.А., Баймуканов А. Генетические исследования верблюдов //Монография. – Шымкент, 2011. - 108с.
69. Баймуканов Д.А., Баймуканов А. Эволюция и систематика верблюдов //Монография. – Шымкент: Полиграф, 2011. -70с.
70. Баймуканов Д.А., Баймуканов А. Микроэволюция верблюдов //Актуальные вопросы животноводства и растениеводства: Материалы международной научно – практической конференции. – Алматы: Бастау, 2011. – С. 178 – 182.
71. Баймуканов Д.А. Внутривидовая дифференциация верблюдов в локальных условиях существования // Традиционны отрасли животноводства (коневодство и верблюдоводство): Четвертая Международная научно-практическая конференция. - Костанай, 2013. – С. 134- 139.
72. Alibaev N.N., Baimukanov D.A. Systematic position of the genus Camelus // Селекционно-технологические аспекты развития продуктивного верблюдоводства, каракулеводства и аридного кормопроизводства в Казахстане: Матер. междун. науч.-практ. конф.(Шымкент, 25-26 ноября 2012г.). –Шымкент, 2012. –С. 191194.
73. Baimukanov D.A. Citogenetics of Camels // Селекционнотехнологические аспекты развития продуктивного верблюдоводства, каракулеводства и аридного кормопроизводства в Казахстане: Матер. междун. науч.-практ. конф.(Шымкент, 25-26 ноября 2012г.). –Шымкент, 2012. –С. 200-204.
74. Дошанов Д.А., Баймуканов Д.А., Юлдашбаев Ю.А. Казахстанские «корабли пустыни» // Ж. Агробизнес.Опубл 17.03.2015г.
75. Баймуканов А., Баймуканов Д.А., Дошанов Д. Воспроизводительная способность верблюдов породы калмыцкий и казахский бактриан /Материалы 4-ой конференции ISOCARD «Верблюды шелкового пути: исследования камелидов для устойчивого развития //Ж.Ветеринария, №2, 2015. – С. 364-365.
76. Омбаев А.М., Баймуканов Д.А., Тоханов М. Молочная продуктивность верблюдов разных генотипов и физико – химические свойства верблюжьего молока /Материалы 4-ой конференции ISOCARD «Верблюды шелкового пути: исследования камелидов для устойчивого развития //Ж. Ветеринария, №2, 2015. – С. 411-412.
Глоссарий
Аберрация хромосомная (или хромосомная аномалия) — обобщенное название любого из типов хромосомных мутаций: де- леций, транслокаций, инверсий, дупликаций. Иногда также обозначают и геномные мутации (анеуплодии, трисомии и т. д.).
Аберрации хромосом (синоним - структурные мутации) – изменения структуры одной или группы хромосом.
Аллель — одна из двух или более альтернативных форм гена, каждая из которых характеризуется уникальной последовательностью нуклеотидов; аллели, как правило, отличаются последовательностями нуклеотидов.
Аллель дикого типа (нормальный) — нуклеотидная последовательность гена, обеспечивающая его нормальную работу.
Аллель доминантный - аллель, наличие которого проявляется в фенотипе.
Аллель мутантный - мутация, приводящая к изменению последовательности аллеля дикого типа.
Аллель рецессивный - аллель, фенотипически проявляющийся только в гомозиготном состоянии и маскирующийся в присутствии доминантного аллеля.
Аллельные серии - моногенные наследственные заболевания, вызванные различными мутациями в одном и том же гене, но относящиеся к разным нозоологическим группам по своим клиническим проявлениям.
Ампликон- внехромосомная единица амплификации.
Амплификатор ДНК (термоциклер) – прибор, необходимый для проведения полимеразной цепной реакции (ПЦР); позволяет задавать нужное количество циклов и выбирать оптимальные временные и температурные параметры для каждой процедуры цикла.
Амплификация – увеличение числа копий генов (количества ДНК).
Амплификация ДНК - выборочное копирование определённого участка ДНК.
Амфидиплоиды-— эукариотические клетки, содержащие два двойных набора хромосом в результате объединения двух геномов.
Анеуплоидия – увеличение числа хромосом в клетке на одну, более одной (гипердиплоидный набор хромосом) или уменьшение на одну, две (гиподиплоидный набор хромосом) в кариотипе.
Анеуплоидия (от греч. аn. – отрицательная частица, eu – хорошо, вполне, ploos – кратный, eidos – вида). Гетероплоидия, явление при котором клетки организма содержат измененное число хромосом, не- кратное гаплоидному набору. Основной механизм возникновения анеуплоидии – не расхождение и потери отдельных хромосом в митозе и мейозе. Анеуплоидия приводит к понижению жизнестойкости и нередко к гибели анеуплоидов, особенно у животных (анеуплоидия лежит в основе ряда хромосомных болезней). В генетическом анализе с помощью анеуплоидии (скрещивая мутантов с анеуплоидами по определенным хромосомам) определяют, в какой группе сцепления находится исследуемый ген. (Биология. БЭС. М.: БРЭ, 1999. –С.27).
Антикодон – последовательность из трёх нуклеотидов в молекуле транспортной РНК, комплементарная кодирующему триплету в молекуле мРНК.
Антимутагенез – процесс предотвращения закрепления (становления) мутации, т. е. возврат первично повреждённой хромосомы или гена в исходное состояние.
Аутбридинг (англ. dut – вне, breeding – разведение), скрещивание или система скрещиваний неродственных форм одного вида. «Неродственность» подразумевает отсутствие общих предков в ближайших 4-6 поколениях. Аутбридинг используют для повышения или сохранения определенной степени гетерозиготности особей (гетерозиготы часто превосходят по многим биологическим параметрам гомозиготные формы). (Биология. БЭС. М.: БРЭ, 1999. - С.44.).
Аутосомы (от греч. autos – сам, soma – тело), все хромосомы в клетках раздельнополых животных, растений и грибов, за исключением половых хромосом. (Биология. БЭС. М.: БРЭ, 1999. -С.44.
Аутосома – любая неполовая хромосома. У человека имеется 22 пары аутосом.
Аутосомно-доминантное наследование – тип наследования, при котором одного мутантного аллеля, локализованного в аутосоме, достаточно, чтобы болезнь (или признак) могла быть выражена.
Аутосомно-рецессивное наследование – тип наследования признака или болезни, при котором мутантный аллель, локализованный в аутосоме, должен быть унаследован от обоих родителей.
Бактериофаг – вирус бактерий: состоит из ДНК или РНК, упакованной в белковую оболочку.
Банк (библиотека) генов – полный набор генов данного организма, полученный в составе рекомбинантных ДНК.
Белковая инженерия – создание искусственных белков с заданными свойствами путём направленных изменений (мутаций) в генах или путём обмена локусами между гетерологичными генами.
Биопсия хориона – процедура, осуществляемая на 7-11-й неделе беременности, с целью получения клеток для пренатальной диагностики.
Биогенез (от греч. bios – жизнь, genesis – происхождение, возникновение) – образование органических соединений живыми организмами. (Биология. БЭС. М.: БРЭ, 1999. - С.60.).
Биогенетический закон, обобщение в области взаимоотношений, онтогенеза и филогенеза организмов, установленное Ф.Мюллером (1864) и сформулированное Э.Геккелем (1866). Онтогенез всякого организма есть краткое и сжатое повторение (рекапитуляция) филогенеза данного вида.
Биогеоценоз (bios – жизнь, rpeч. ge – Земля, koinos – общий), однородный участок земной поверхности с определенным составом живых (биоценоз) и косных (приземный слой атмосферы, солнечная энергия, почва и др.) компонентов, объединенных обменом вещества и энергии в единый природный комплекс. (Биология. БЭС. М.: БРЭ, 1999. - С.62).
Биологическое долголетие – это длительность жизни животного, прерываемая естественной смертью.
Биологическая номенклатура, система научных названий в биологии для групп организмов, связанных той или иной степенью родства – таксонов. Биологическая номенклатура обеспечивает единство и стабильность научных названий животных.
Биология развития, раздел биологии, изучающий причинные механизмы и движущие силы индивидуального развития (онтогенеза) животных и растений. (Биология. БЭС. М.: БРЭ, 1999. - С.67).
БИОМ (англ. – biome, от греч. bios – жизнь и лат. оmа – окончание, обозначающее совокупность), совокупность различных групп организмов и среды их обитания в определенной ландшафтно-географической зоне. (Биология. БЭС. М.: БРЭ, 1999. - С.68).
Биометрия (от bios – жизнь и греч. metreo – измеряю) раздел вариационной статистики с помощью методов которого производят обработку экспериментальных данных и наблюдений, а также планирование количественных экспериментов в биологических исследованиях.
Биосинтез (от bios – жизнь и греч. synthesis – соединение), образование органических веществ из более простых соединений, происходящее в живых организмах под действием биокатализаторов – ферментов.
Биосфера (от био... и греч. sphaira – шар), оболочка Земли, состав, структура и энергетика которой определяются совокупной деятельностью живых организмов.
Биотехнология (от био..., греч. techne – искусство, мастерство и ...логия...) использование живых организмов и биологических процессов в производстве.
Биотип (от био... и тип) совокупность особей в составе популяции, имеющих сходный генотип, мельчайшая таксономическая категория из которой складывается вид. (Биология. БЭС. М.: БРЭ, 1999. - С.70).
Биохимия, биологическая химия, наука о химическом составе живой материи и о химических процессах, происходящих в живых организмах и лежащих в основе их жизнедеятельности.
Биоценоз (от био... и ценоз) совокупность животных, растений, грибов и микроорганизмов, совместно населяющих участок суши или водоема.
Биоцикл (от био... и греч. kyklos – круг) закономерная смена фаз или стадии развития организма.
Биоэлектрические потенциалы, электрические потенциалы, возникающие в тканях и отдельных клетках живых организмов, важнейшие компоненты процессов возбуждения и торможения.
Биоэнергетика совокупность процессов преобразования энергии в биологических системах, а также раздел биологии, изучающий эти процессы.
Борьба за существование, одно из основных понятий в теории эволюции Ч.Дарвина, которое он употреблял для обозначения всей совокупности отношений между особями и различными факторами внешней среды. Эти отношения определяют успех или неудачу данной особи в выживании и оставлении потомства и включают внутривидовую и межвидовую конкуренцию, а также отношения хищник-жертва, взаимодействие организмов с абиотическими факторами внешней среды.
Бонитировка – определение уровня племенной ценности животных путем оценки их по комплексу признаков (породность, продуктивные качества, экстерьерно-конституциональные особенности) с присвоением соответствующего класса.
Блотинг - перенос молекул ДНК, РНК или белка из геля, в котором шёл электрофорез, на фильтр (мембрану).
Саузерн блоттинг - метод идентификации участков ДНК, содержащих комплементарные ДНК-зонду последовательности, среди электрофоретически разделенных фрагментов ДНК, фиксированных на твердом матриксе (нитроцеллюлозных или нейлоновых фильтрах).
Вакцина - препарат ослабленного или убитого инфекционного агента (вируса, бактерии и т. п.) или его отдельных компонентов, несущих антигенные детерминанты, способный вызывать образование иммунитета к данной инфекции у животных (человека). Кроме того, в последнее время появились вакцины, произведенные методами генной инженерии (примером такой вакцины может служить вакцина против гепатита B).
Везикулы - мембранные пузырьки. Кроме того, везикулами в медицине называют любые элементы сыпи, представляющие собой пузырьки.
Вектор - молекула ДНК, способная к включению чужеродной ДНК и к автономной репликации, служащая инструментом для введения генетической информации в клетку.
Ветеринария (область ветеринарии) – область специальных научных знаний и практической деятельности, направленная на изучение болезней и пищевых отравлений (поражений) животных, их профилактику, диагностику, лечение и ликвидацию, обеспечение соответствия объектов государственного ветеринарного надзора требованиям законодательства Республики Казахстан в области ветеринарии, а также защиту населения от болезней, общих для животных и человека.
Ветеринарно-санитарная безопасность - состояние объектов государственного ветеринарного надзора, не представляющее опасности для здоровья животных и человека при обычных (установленных) условиях их использования.
Ветеринарно-санитарная экспертиза — проверка соответствия животных, продуктов и сырья животного происхождения, ветеринарным нормативам комплекса органолептических, биохимических, микробиологических, паразитологических, токсикологических и радиологических исследований в порядке, установленном уполномоченным государственным органом в области ветеринарий.
Ветеринарные мероприятия — комплекс противоэпизоотоло- гических, ветеринарно-санитарных процедур, направленных на предотвращение возникновения, распространения или ликвидацию болезней животных, включая их профилактику, лечение или диагностику; обезвреживание (обеззараживание), изъятие и уничтожение животных; обеспечение безопасности продуктов и сырья животного происхождения, включая процедуры идентификации, в целях зашиты здоровья животных человека от заразных болезней, в том числе, общих для животных и человека.
Вид (Species), основная структурная единица в системе живых организмов, качественный этап их эволюции. Основная таксономическая категория в биологической систематике. Обычно под видом понимается совокупность популяции особей, способных к скрещиванию с образованием плодовитого потомства, населяющих определенный ареал, обладающих рядом общих морфологических признаков и типов взаимоотношений с абиотической и биотической средой и отделенных от других таких же групп особей практически полным отсутствием гибридных форм. (Биология. БЭС. М.: БРЭ, 1999. – С.94-95).
Виды это группы скрещивающихся естественных популяций, репродуктивно изолированные от других таких групп (определение по Э.Майру 1974 г. так называемая биологическая концепция вида).
Видообразование, процесс возникновения новых видов посредством разветвления предковой филетической линии на несколько новых, постепенное превращение (во времени) одного вида в другой (так называемое филетическое видообразование происходящее без увеличения числа видов), а также образование новых видов путем гибридизации.
Вектор для клонирования – любая небольшая плазмида, фаг или ДНК-содержащий вирус животных, в которые может быть встроена чужеродная ДНК.
Виды гистонов – существуют пять видов гистонов: Н1 (очень богатый лизином), Н2А и Н2В (богатые лизином), Н3 (богатый аргинином) и Н4 (богатый глицином и аргинином).
Вирусы – инфекционные агенты неклеточной природы, способные в процессе реализации генетической информации, закодированной в их геноме, перестроить метаболизм клетки, направив его в сторону синтеза вирусных частиц. Вирусы могут иметь белковую оболочку, а могут и состоять только из ДНК или РНК.
Водородная связь – образуется между электроотрицательным атомом молекулы (кислород, азот) и электроположительным ядром водорода (протоном), который, в свою очередь, ковалентно связан с другим электроотрицательным атомом той же или соседней молекулы.
Внутрипопуляционный хромосомный полиморфизм у млекопитающих - в книге В.Н.Орлова (1974) «Кариосистематика млекопитающих» есть интересный пример у хлопоковой крысы 30 хромосом, большинство акроцентрики. Но в маленькой популяции на юге-западе штата Нью-Мексико (США) у некоторых особей обнаружено 29 хромосом, в т.ч. новый метацентрик. Исследование мей- оза показало, что этот метацентрик гомологичен двум акроцентри- кам (Hsu, Mead, 1969), т.е. образовался в результате робертсоновской транслокации.
Врождённые болезни - болезни, имеющиеся при рождении, могут быть как наследственными, так и дефектами индивидуального развития организма.
β-Галактозидаза - фермент, гидролизующий – β-галактозиды, в частности лактозу, с образованием свободной галактозы.
Габитус, хабитус (от лат.habitus - внешность, наружность) внешний облик организма, совокупность признаков, характеризующих общий тип телосложения.
Гамета - зрелая половая клетка.
Гамета (от rpeч. gamete – жена, gametes – муж) половая клетка, репродуктивная клетка животных и растений. Гаметы обеспечивают передачу наследственной информации от родителей потом-
кам. Гаметы обладают гаплоидным набором хромосом, что обеспечивается сложным процессом гаметогенеза.
Гаметогенез (от гамета и ... генез), развитие половых клеток (гамет). Гаметогенез у большинства животных бывает локализованный (гаметы развиваются в половых железах – ганадах).
Гамия (от греч. gamos – брак), часть сложных слоев, означающая отношение между полами, половой процесс, оплодотворение.
Гаплоид – клетка, содержащая одинарный набор генов или хромосом.
Гаплоид (от греч. haploos – одиночный, простой и eidos – вид), организм (клетка, ядро) с одинарным (гаплоидным) набором хромосом, который обозначается латинской буквой n. У млекопитающих гаплоидны только половые клетки.
Гемизигота (от греч. hemi – полу и зигота) диплоидный организм, у которого имеется только одна доза определенных генов. Гемизиготное состояние может возникнуть вследствие анеуплоидии и делеций. В норме оно характерно для генов, локализующихся в половых хромосомах у особей гетерогаметного пола. Рецессивные аллели (мутации) в гемизиготном состоянии проявляются фенотипически, что используют, например, при оценке мутагенности анализируемых факторов. У человека гемизиготными по генам Х- хромосоме являются мужчины, поэтому рецессивные наследственные заболевания обусловленные такими генами (гемофилия, цветовая слепота, мышечная дистрофия и др.), встречаются чаще у мужчин, чем у женщин. (Биология. БЭС. М.: БРЭ, 1999. - С.120).
Гемизиготность – состояние организма, при котором какой-то ген представлен в одной хромосоме.
Ген - последовательность нуклеотидов в ДНК, которая кодирует определённую РНК.
Генез (от греч. genesis – происхождение, возникновение), происхождение, процесс, образование, часть сложных слов, например, онтогенез.
Генетика (от rpeч. genesis – происхождение), наука о наследственности и изменчивости живых организмов и методах управления ими.
Генетическая карта - схема расположения структурных генов и регуляторных элементов в хромосоме.
Генетический код - соответствие между триплетами в ДНК (или РНК) и аминокислотами белков.
Генетика поведения – раздел общей генетики, изучающий наследственную детерминацию поведения животных. Поведенческие реакции животных, как и многие их другие признаки и свойства, обусловлены наследственностью и влиянием факторов внешней среды. Генотип служит наследственной информацией, которая реализуется в процессе индивидуального развития в виде того или иного поведения животного. Контролируемое генотипом поведение животного совершенствуется под действием условий среды адаптации к данным животным.
Генная инженерия - совокупность приемов, методов и технологий получения рекомбинантных РНК и ДНК, выделения генов из организма (клеток), осуществления манипуляций с генами и введения их в другие организмы.
Генетическая инженерия – это новое направление в современной биологии, ставящее своей задачей моделирование желательных для практики и науки формы генетических программ и затем воплощать их в жизнь. В 1934 г. Н.П.Дубинин с помощью рентгеновских лучей создал у дрозофилы измененный кариотип по заранее предсказанной модели. В 1971 г. В.А.Струнников используя генетические манипуляции на генном и хромосомном уровне получил особей тутового шелкопряда, пол которых был мечен окраской грены, что достигалось транслокацией между аутосомой и половой хромосомой.
Генетическая инженерия, генная инженерия, раздел молекулярной генетики, связанный с целенаправленным созданием in vitro новых комбинаций генетического материала, способного размножаться в клетке-хозяине и синтезировать конечные продукты обмена.
Генетической инженерией называют прикладную молекулярную и клеточную генетику, разрабатывающую приемы экспериментального вмешательства, позволяющего по заранее намеченному плану перестраивать геном организмов, изменяя содержащуюся в нем генетическую информацию (О.М.Гершензон //Основы современной генетик. –Киев: Науковадумка, 1983. -558 с.
Генетическая инженерия – прикладная молекулярная генетика, имеющая дело с элементарными генетическими системами –
молекулами ДНК и их отдельными фрагментами. В основе генетической инженерии лежит технология рекомбинантной ДНК.
Генетическая инженерия сельскохозяйственных животных – прикладной раздел молекулярной генетики, разрабатывающий технологию гибридной (рекомбинантной) ДНК в целях повышения комбинативной наследственной изменчивости и создания животных с новыми генетическими признаками.
Генетическая информация, информация о свойствах организма, которая передается по наследству. Генетическая информация записана последовательностью нуклеотидов молекул нуклеиновых кислот (ДНК, у некоторых вирусов также РНК). У многоклеточных организмов при половом размножении. Генетическая информация передается из поколения в поколение посредством половых клеток.
Генетическая карта хромосомы, схема взаимного расположения генов, находящихся в одной группе сцепления. Расстояние между генами на генетической карте хромосом определяют по частоте кроссинговера между ними.
Генная терапия - введение генетического материала (ДНК или РНК) в клетку для восстановления нормальной функции.
Геном - общая генетическая информация, содержащаяся в генах организма, или генетический состав клетки.
Генотип 1) вся генетическая информация организма; 2) генетическая характеристика организма по одному или нескольким изучаемым локусам.
Ген-регулятор - ген, кодирующий регуляторный белок активирующий или подавляющий транскрипцию других генов.
Ген-репортер - ген, чей продукт определяется с помощью простых и чувствительных методов и чья активность в тестируемых клетках в норме отсутствует. Используется в генно-инженерных конструкциях для подтверждения наличия вектора.
Ген-усилитель (энхансер) - короткий сегмент ДНК, который влияет на уровень проявления (экспрессии) определённых генов, увеличивая частоту инициации и транскрипции Генетический анализ, совокупность методов исследования наследственных свойств организма (его генотипа). К основным методам генетического анализа относятся: селекционный метод, с помощью которого осуществляют подбор или создание исходного материала, подвергающегося дальнейшему анализу: гибридологический метод, представ-
ляющий собой систему специальных скрещиваний и учета результатов; цитогенетический метод, заключающийся в цитологическом анализе генетических структур и явлений на основе гибридологического анализа с целью сопоставления генетических явлений со структурой и поведением хромосом и их участков (анализ хромосомных и геномных мутаций, построение цитологических карт хромосом, цитохимическое изучение активности генов и т.п.). (Биология. БЭС. М: БРЭ, 1999. - С.124).
Генетическая анеуплоидия - показатель доли генетически анеу-плоидных клеток, определяется как удвоенное число доли ги- перди- плоидных клеток.
Генетическая аномалия (синоним генетический риск образования аномальных клеток) – определяет долю клеток, ставших аномальными вследствие численных или структурных изменений отдельных хромосом или всего генома. Генетическая анеуплоидия определяется суммированием числа полиплоидных клеток, клеток с хромосомными аберрациями и генетически анеуплоидных клеток.
Генетический груз, часть наследственной изменчивости популяции, которая определяет появление менее приспособленных особей, подвергающихся избирательной гибели в процессе естественного отбора.
Генетический груз в популяции верблюдов – это распространение в популяции скрытых рецессивных генов, которая обуславливает в дальнейшем генетическую изменчивость и ухудшение приспособленности к среде в результате действия вредных аллелей, либо снижение жизнеспособности и плодовитости особей. В верблюдоводстве генетический груз, являясь источником генетической изменчивости, имеет большое значение при искусственном отборе высокоценных генотипов верблюдов более приспособленных к условиям резкоконтинентального климата Казахстана и соответствующих селекционного процесса.
Генетический код, свойственная живым организмам единая система записи наследственной информации в молекулах нуклеиновых кислот в виде последовательности нуклеотидов: определяет последовательность включения аминокислот в синтезирующуюся полипептидную цепь в соответствии с последовательностью нуклеотидов ДНК гена.
Генетический материал. Компоненты клетки, структурнофункциональное единство, которых обеспечивает хранение, реализацию и передачу наследственной информации при вегетативном и половом размножении.
Генокопия (от ген и лат. сорiа – множество, запас), одинаковые изменения фенотипа, обусловленные аллелями различных генов.
Геном (нем. – Genom). Совокупность генов, характерных для гаплоидного набора хромосом данного вида организмов; основной гаплоидный набор хромосом.
Геномный анализ, метод цитогенетического анализа, заключающийся в определении геномного состава аллополиплоидов и общности геномов в пределах родственных систематических групп организмов (видов, родов и др.). Геномный анализ основан на анализе поведения хромосом в мейозе у гибридных форм. Конъюгация между хромосомами, полученными гибридом от разных родителей, свидетельствует о наличии у родительских форм общих геномов, а обнаружение унивалентов об отсутствии общности. Окончательные выводы делают после количественного учета числа хромосом, уни – и бивалентов у гибрида. Генетический анализ позволяет делать предположения о происхождении и степени родства между изучаемыми видами. (Биология. БЭС. М.: БРЭ, 1999. –С.126).
Геномные мутации (синоним – числовые мутации хромосом) – изменения числа хромосом в кариотипе, подразделяются на полиплоидию и анеуплоидию.
Генотип (от ген и греч. typos – отпечаток), генетическая (наследственная) конституция организма, совокупность всех наследственных зачатков данной клетки или организма, включая аллели генов, характер физического сцепления в хромосомах и наличие хромосомных перестроек. Генотип контролирует развитие, строение и жизнедеятельность организма, то есть совокупность всех признаков организма – его фенотип. Особи с разным генотипом могут иметь одинаковый фенотип, поэтому для определения генотипа организма необходимо проводить его генетический анализ, например анализирующее скрещивание. Особи с одинаковым генотипом могут отличаться друг от друга по фенотипу. Поэтому в генетике используют понятие о норме реакции – возможном размахе фенотипической изменчивости без изменения генотипа под влиянием внешних условий (генотип определяет пределы нормы реакции).
(Инге-Вечтонов С.Г. Система генотипа //Физиологическая генетика, Л., 1976, С.57-114).
Гетеро… (от греч. heteros – иной, другой), часть сложных слов, означающая разнородность, чужеродность (противоположное гомо... или гомео...) например гетерогамия, гетерокарпия.
Гетерогаметность (от гетеро... и гаметы), характеристика организма или группы организмов, имеющих в своем хромосомном наборе одну половую хромосому (тип ХО) или пару различающихся половых хромосом (X и У) и вследствие этого образующих разные гаметы. Пол, представленный особями с такими наборами половых хромосом называют гетерогаметным.
Гетерогенез (от гетеро... и ...генез) внезапное появление особей, резко отличающихся по ряду признаков от родительских форм.
Гетерозигота (от гетеро... и зигота) организм (клетка) у которого гомологичные хромосомы несут различные аллели (альтернативные формы) того или иного гена. Гетерозиготность, как правило, обуславливает высокую жизнеспособность организмов, хорошую приспособляемость их к изменяющимся условиям среды и поэтому широко распространено в природных популяциях. В экспериментах гетерозигот получают скрещиванием между собой гомозигот по различным аллелям. Термин гетерозигота используют и для хромосомных перестроек (говорят о гетерозиготе по инверсии, транслокации и т.п.). (Биология. БЭС. М.: БРЭ, 1999. - С.129-130).
Гетерозигота – клетка (или организм), содержащая два различных аллеля в конкретном локусе гомологичных хромосом.
Гетерозиготность – наличие разных аллелей в диплоидной клетке.
Гетерозиготный организм – организм, имеющий две различные формы данного гена (разные аллели) в гомологичных хромосомах.
Гетерохроматин - область хромосомы (иногда целая хромосома), имеющая плотную компактную структуру в интерфазе из-за отсутствия транскрипции.
Гибридизация in situ – гибридизация между денатурированной ДНК клеток на предметном стекле и меченной радиоактивными изотопами или иммунофлюоресцентными соединениями одноцепочечной РНК или ДНК.
Гибридизация ДНК – образование в опыте двуцепочечной ДНК или дуплексов ДНК:РНК в результате взаимодействия комплементарных нуклеотидов.
Гибридизация соматических клеток - слияние неполовых клеток, способ получения соматических гибридов.
Гибридомы - гибридные лимфоидные клетки, полученные путём слияния опухолевой миеломной клетки с нормальными лимфоидными клетками иммунизированного животного или человека.
Гистоны – это хромосомные основные белки с высоким содержанием аминокислот аргинина и лизина. Гистоны прочно соединяются с молекулами ДНК, чем препятствуют считыванию заключенной в ней биологической информации. В этом состоит их регуляторная роль. Кроме того, эти белки выполняют структурную функцию, обеспечивая пространственную организацию ДНК в хромосомах.
Гликозилирование – присоединение к белку углеводного остатка.
Голандрическое наследование – наследование, сцепленное с Y- хромосомой.
Гомозигота – клетка (или организм), содержащая два одинаковых аллеля в конкретном локусе гомологичных хромосом.
Гомозиготность – наличие одинаковых аллелей в диплоидной клетке.
Гомозиготный организм – организм, имеющий две идентичные копии данного гена в гомологичных хромосомах.
Гомологичные хромосомы – хромосомы, одинаковые по набору составляющих их генов.
Группа сцепления – все гены, локализованные в одной хромосоме.
Гетерозис (от греч. heterosis – изменение, превращение), «гибридная мощность», превосходство гибридов по ряду признаков и свойств над родительскими формами. Термин гетерозис предложен Дж.Шеллом в 1914. Как правило, гетерозис характерен для гибридов первого поколения, полученных при скрещивании неродственных форм: различных линий, пород, видов. В дальнейших поколениях (скрещивание гибридов между собой) его эффект ослабляется и исчезает. В животноводстве гетерозис у животных нередко приводит к значительному повышению продуктивности. Однако, его
использование часто недостаточно эффективно, так как до сих пор не решена проблема закрепления гетерозиса в ряду поколений. (Биология. БЭС. М.: БРЭ, 1999. - С.130).
Гетерохроматин (от гетеро... и хроматин) участки хроматина, находящиеся в конденсированной (плотно упакованном) состоянии в течение всего клеточного цикла. Интенсивно окрашиваются ядер- ными красителями и хорошо видны в световой микроскоп даже во время интерфазы. Различают факультативный и конститутивный (структур-ный) гетерохроматин. Факультативный гетерохроматин присутствует только в одной из гомологичных хромосом. Пример гетерохроматина такого типа - вторая Х-хромосома у женской особи млекопитающих, которая в ходе раннего эмбриогенеза инактивируется вследствие ее необратимой конденсации. Структурный гетерохроматин содержится в обеих гомологичных хромосомах, локализован преимущественно в экспонированных участках хромосомы – в центромере, теломере, ядрышковом организаторе (во время интерфазы он располагается неподалеку от ядерной оболочки), обеднен генами. (Биология. БЭС. М.: БРЭ, 1999. - С.131).
Гибрид (от лат. hibrida, hybrida - помесь), организм (клетка), полученный в результате объединения материала генотипически разных организмов (клеток), то есть гибридизаций. Отдаленные гибриды (разных таксонов видов и выше) в природе встречаются довольно редко и, как правило, бесплодны. (Биология. БЭС. М.: БРЭ, 1999. - С. 132).
Гибридизация, процесс образования или получение гибридов, в основе которого лежит объединение генетического материала разных клеток в одной клетке. Может осуществляться в пределах одного вида (внутривидовая гибридизация, гибриды характеризуются гетерозиготностью по многим или анализируемому гену) и между разными систематическими группами (отдаленная гибридизация, при которой происходит объединение разных геномов). Для первого поколения гибридов часто характерен гетерозис, выражающийся в лучшей приспособляемости, большей плодовитости и жизнеспособности организма. При отдаленной гибридизации гибриды, как правило, неплодовиты. (Биология. БЭС. М.: БРЭ, 1999. -С. 132).
Гибридологический анализ, анализ характера наследования признаков с помощью системы скрещиваний. Гибридологический анализ заключается в получении гибридов и дальнейшем их сравни-
тельном анализе в ряду поколений (анализ расщепления). Информация, полученная при гибридологическом анализе, необходима для получения организмов с заданными генетическими свойствами. ( Биология. БЭС. М.: БРЭ, 1999. - С. 132- 133).
Гиподиплоидные клетки (синоним гипоплоидия) - уменьшение числа хромосом в кариотипе, у верблюдов 2n<74 (70,72,73).
Гипердиплоидные клетки (синоним гиперплоидия) - увеличение числа хромосом в кариотипе, у верблюдов 2n>74 (75,76,78).
Гомогаметность (от гомо... и гамета), характеристика организма имеющего в хромосомном наборе пару или несколько пар гомологичных половых хромосом и вследствие этого образующих одинаковые по набору хромосом гаметы. Пол, представленный такими особями называют гомогаметными. Для млекопитающих го- могаметность характерна для женского пола (ХХ) у птиц гомога- метны самцы (ZZ).
Гомозигота (от гомо... и зигота), диплоидная или полиплоидная клетка (особь), гомологичные хромосомы которой несут идентичные аллели того или иного гена. Получают гомозигот, как правило, с помощью инбридинга той или иной степени.
Гомологических рядов наследственной изменчивости закон, устанавливает параллелизм в наследственной изменчивости организмов со сходным набором генов. Закон объясняет полиморфность видов и, таким образом, обосновывает целостность вида, несмотря на существование в его пределах морфологически четко различающихся форм. С другой стороны, закон вносит ясность в явление фенотипической однородности множества видов, которая может быть связана с их L гетерозиготностью и явлением доминирования, что и выявляется при инбридинге. Закон гомологических рядов отражая общую закономерность мутационного процесса и формообразования организмов является биологической основой методов целенаправленного получения нужных наследственных изменений. (Биология. БЭС. М: БРЭ, 1999. -С.152-153).
Гомологичные хромосомы содержат одинаковый набор генов, сходных по морфологическим признакам, конъюгируют в профазе мейоза. В диплоидном наборе хромосом каждая пара хромосом представлена двумя гомологичными хромосомами, которые могут различаться аллелями, содержащихся в них генов и обмениваться участками в процессе кроссинговера.
Гомология (от греч. homologia - соответствие, согласие) соответствие органов у организмов разных видов, обусловленные их филогенетическим родством.
Группа крови, иммуногенетический признак крови, обуслов- лен-ные специфическими антигенами (изоантигенами) и позволяющие делить кровь особей одного вида на группы.
Дактилоскопия генная - выявление вариаций в числе и длине тандемных повторов ДНК.
Дегенерация (от лат. degenero - вырождаюсь), упрощение структуры органов и тканей в процессе онтогенеза организмов. Редукция отдельных органов и целых систем в процессе филогенеза.
Дезоксирибонуклеиновые кислоты, ДНК, нуклеиновые кислоты, содержащие в качестве углеводного компонента дезоксирибозу, а в качестве азотистых оснований аденин (А), гуанин (Г), цитозин (Ц), тинин (Т). Присутствует в клетках любого организма, а также входит в состав многих вирусов.
Дезоксирибонуклеотиды, нуклеотиды, содержащие углевод дезоксирибозу, пуриновое (аденин или гуанин) или пиринидиновое (цитозин или тимин) основание и остатки фосфорной кислоты; мономеры, из которых построены ДНК. ( Биология. БЭС. М.: БРЭ, 1999. - С.171).
Деление, форма размножения некоторых организмов и многих клеток, входящих в состав клеток многоклеточных.
Делеция (от лат. deletion – уничтожение), тип хромосомной перестройки, в результате которой выпадает участок генетического материала. Размер делеции от нескольких нуклеотидных пар до фрагментов, содержащих ряд генов.
Делеция - тип хромосомной мутации, при которой утрачивается участок хромосомы; тип генной мутации, при которой выпадает участок молекулы ДНК.
Денатурация - нарушение пространственной структуры молекулы в результате разрыва внутри- или межмолекулярных нековалентных связей.
ДНК-полимераза - фермент, ведущий матричный синтез ДНК.
Диплоид (от греч. diploos – двойной и eidos – вид), организм, клетки которого несут два гомологичных набора хромосом.
Дифференциальная окраска хромосом - При дифференциальной окраске каждая хромосома приобретает свой специфический
рисунок – чередование светлых и темных полос, отражающих различную функциональную активность отдельных районов хромосом. Окрашенные участки – это низкоактивные в генетическом отношении гетерохромативные районы хромосом, а неокрашенные сильноактивные эухроматиновые районы. Гетерохроматин, как показывает дифференциальное окрашивание, существует в двух формах: 1) констутивной – постоянно действующей в хромосоме и 2) факультативной, края выявляются лишь в части клеточного цикла или в одной из пар хромосом.
Домашние гены (Housekeeping gene) - это гены, которые транскрибируются с относительным постоянством и используются в качестве нормализатора (стандарта) в PCR (полимеразной цепной реакции), поскольку предполагается, что на их экспрессию не влияют условия эксперимента.
Доминантность - преимущественное проявление только одного аллеля в формировании признака у гетерозиготной клетки.
Доминантность, участие только одного аллеля в определении признаков у гетерозиготных особей. Когда нет доминирования раз- ли-чают следующие варианты фенотипа: промежуточный (непольное доминирование), более функциональный по данному признаку (сверхдоминирование) и фенотип, обусловленный обоими аллелями (кодоминантность).
Доминантный - признак или соответствующий аллель, проявляющийся у гетерозигот.
Дрейф генов, генетико-автоматические процессы изменения частоты генов в популяции в ряду поколений под действием случайных (стохастических) факторов, приводящее, как правило, к снижению наследственной изменчивости популяции. В генотипической структуре популяции под действием дрейфа генов происходит усиление процесса гомозиготизации, которая нарастает с уменьшением численности популяции. Связано это с тем, что в популяциях ограниченного размера увеличивается частота близкородственных скрещиваний, и в результате заметных случайных колебаний частот отдельных генов происходит закрепление от них аллелей при одновременной утрате других. (Биология. БЭС. М.: БРЭ, 1999. - С. 185).
Дрейф генов - изменение частот генов в ряду поколений, обусловленное случайными событиями митоза, оплодотворения и размножения.
Дупликация - тип хромосомной мутации, при которой удвоен какой-либо участок хромосомы; тип генной мутации, при которой удвоен какой-либо участок ДНК.
Задачи сельскохозяйственной биотехнологии – выведение трансгенных животных с улучшенной продуктивностью и более высоким качеством продукции, резистентностью к болезням, а также создание так называемых животных – биореакторов – продуцентов биологически активных веществ.
Зонд генетический - короткий отрезок ДНК или РНК известной структуры или функции, меченный каким-либо радиоактивным или флуоресцентным соединением.
Идиограмма – графическое изображение хромосом с учетом их морфологических деталей: длины, расположения центромеры, вторичных перетяжек и при дифференциальной окраске расположения положительно и негативно окрашенных полос. Идиограмма может быть построена по обобщенным данным или для конкретного кариотипа. Число хромосом в ядрах клеток всех особей одного вида постоянно и представляет собой один из его признаков. У верблюдов кариотип представлен 74 хромосомами, из них 12 метацентрические аутосомы, 60 акроцентрические аутосомы, ХХ (у самок) и ХУ (у самцов) половые хромосомы – гоносомы. То есть, кариотип – это набор хромосом соматической клетки свойственный тому или иному виду животных или растении.
Изменчивость - вариабельность (разнообразие) признаков среди представителей данного вида.
Иммунитет - механизм борьбы организма с инфекционным агентами типа вирусов и микробов.
Иммунотоксин - комплекс между антителом и каталитической субъединицей какого-либо белкового яда (дифтерийного токсина, рицина, абрина и др.).
Инбридинг в современной селекции - Ин - эндинбридинг - родственное разведение в нескольких поколениях. Клозебридинг – кровосмешение (близкородственное разведение). Инбредлайнкроссинг – спаривание животных из разных инбредных линий одной породы. Страинкроссинг – спаривание животных из близкородственных линий. Топкроссинг – спаривание инбредных самцов с неинбредными самками. Боттомкроссинг – спаривание инбредных самок с аут- бредными самцами. Топкроссбридинг – скрещивание инбредных
самцов одной породы с неинбредными самками другой породы. Инкроссбридинг – скрещивание инбредных самцов одной породы с инбредными самками другой породы. Боттомкроссбридинг – скрещивание инбредных самок одной породы с аутбредными самцами другой породы.
Инбредная депрессия в верблюдоводстве – Инбредная депрессия – снижение продуктивности и жизнеспособности животных в результате использования инбридинга. Сила проявления инбредной депрессии зависит от индивидуальных особенностей инбридируемых животных, степени их исходной гетерозиготности, конституциональной крепости, пола (самцы более подвержены инбредной депрессии в сравнении с самками), возраста (при спаривании особей в раннем и старости инбредная депрессия проявляется сильнее), природы признака, скорости падения гетерозиготности и числа инбридированных поколений (чем теснее инбридинг, тем сильнее проявление инбредной депрессии), условий среды (оптимальное условие среды способствует ослаблению инбредной депрессии, повышению продуктивности животных, то есть лучшей реализации их генетических возможностей).
Индекс спирализации - о пределение дано В.М.Гиндилисом в 1966 г. – это отношение суммарной длины 2-х хромосом человека из группы F (19 и 20 пары) к суммарной длине 2-х хромосом из группы А (1-й и 2-я пары) в процентах.
Индуктор – фактор (вещество, свет, теплота), вызывающий транскрипцию генов, находящихся в неактивном состоянии.
Индукция профага – инициирование вегетативного развития фага в лизогенных клетках.
Интеграза – фермент, осуществляющий внедрение какого-либо генетического элемента в геном через специфический сайт.
Интегроны – генетические элементы, которые содержат в себе ген интегразы, специфический сайт и рядом с ним промотор, что придает им способность интегрировать в себя мобильные генные кассеты и экспрессировать присутствующие в них беспромоторные гены.
Интерфероны – белки, синтезируемые клетками позвоночных в ответ на вирусную инфекцию и подавляющие их развитие.
Интрон – некодирующий участок гена, который транскрибируется, а затем удаляется из предшественника мРНК при её редактировании сплайсинге.
Интронированный ген – ген, содержащий интроны.
Интроны – повторяющиеся последовательности нуклеотидных остатков в ДНК.
Каллус – масса недифференцированных клеток, образующаяся при повреждении растения. Может образовываться из единичных клеток при их культивировании на искусственных средах.
Капсид – белковая оболочка вируса.
Кассета экспрессионная – фрагмент ДНК, содержащий все необходимые генетические элементы для экспрессии внедренного в него гена.
Кариотип – совокупность признаков хромосомного набора (число, размер, форма), характерных для того или иного вида, устанавливается путем определения постоянного диплоидного набора хромосом в клетке. Диплоидный набор хромосом верблюдов 2n=74.
Кариограмма – микрофотографии хромосом индивидуума, систематизированного по группам в зависимости от морфологического строения.
Кариосистематика – раздел систематики, изучающий структуру клеточного ядра у разных групп организмов. Кариосистемати- ка развивалось на стыке систематики с цитологией и генетикой, обычно изучает строение и эволюцию хромосомного набора- кариотипа. Этот раздел биологической науки очень важен при изучении верблюдов разных видов, пород, помесных и гибридных верблюдов.
кДНК - однонитевая ДНК, синтезируемая in vivo по матрице РНК с помощью обратной транскриптазы.
Клон - группа генетически идентичных клеток, возникших неполовым путём от общего предка.
Клонирование ДНК - процесс получения рекомбинантных молекул ДНК путем встраивания чужеродной ДНК в векторную молекулу ДНК или РНК и введение этой конструкции в фаговые, бактериальные или эукариотические клетки хозяина.
Клонирование клеток - их разделение путём рассева в питательной среде и получение колоний, содержащих потомство от изолированной клетки.
Кодон - тройка расположенных подряд нуклеотидных остатков в ДНК или РНК, кодирующая определённую аминокислоту или являющаяся сигналом окончания трансляции.
Компартментализация - ограничение процесса (продукта) определённой областью клетки.
Компетентность - способность клеток к трансформации.
Комплементарность (в генетике) - свойство азотистых оснований образовывать с помощью водородных связей парные комплексы аденин—тимин (или урацил) и гуанин—цитозин при взаимодействии цепей нуклеиновых кислот.
Конкатемерная ДНК - линейная ДНК, в которой некоторый элемент (например, фаговый геном) повторен несколько раз.
Контиг - в секвенировании группа из нескольких последовательно соединенных участков ДНК.
Конъюгат - комплекс из нескольких ковалентно связанных молекул.
Конъюгация - способ обмена генетической информацией у бактерий, при котором вследствие физического контакта между клетками происходит перенос клеточной, плазмидной или транспозонной ДНК от донорной клетки в реципиентную.
Космида - вектор, содержащий cos-сайт ДНК фага λ.
Коэффициент отбора – это количественное выражение давления отбора на конкретный аллель локуса.
Кроссинговер - явление обмена участками гомологичных хромосом во время конъюгации при мейозе.
Лектины - белки, связывающие углеводы.
Лигаза - фермент, образующий фосфодиэфирную связь между двумя полинуклеотидами.
Лиганд - молекула, распознаваемая специфической структурой, например, клеточным рецептором.
Лидерная последовательность - N-концевая последовательность секретируемых белков, обеспечиваюшая их транспорт через мембрану и отщепляющаяся при этом.
Лизис - распад клетки, вызванный разрушением её оболочки.
Лизогения - явление носительства бактериальными клетками фага в виде профага.
Линия клеток - генетически однородные клетки животных или растений, которые можно выращивать in vitro в течение неограниченно долгого времени.
Линкер - короткий синтетический олигонуклеотид, применяемый для соединения фрагментов ДНК in vitro; обычно содержит участок узнавания определённой рестриктазой.
Липкие концы - комплементарные однонитевые участки ДНК, расположенные на концах молекул ДНК.
Липосомы - капельки жидкости, окруженные искусственной мембраной; искусственные липидные везикулы.
Литическое развитие фага - фаза жизненного цикла фага, начинающаяся инфекцией клетки и завершающаяся её лизисом.
Локус - участок ДНК (хромосомы), где расположена определённая генетическая детерминанта.
Маркерный ген - ген в рекомбинантной ДНК, кодирующий селективный признак.
Материнского эффекта гены - гены, проявляющиеся в яйцеклетке и определяющие фенотип потомства вне зависимости от генотипа самца.
Межвидовые гибриды - гибриды, полученные от слияния клеток, принадлежащих к разным видам.
Метаболизм - совокупность ферментативных процессов, обеспечивающих существование и воспроизведение клетки.
Метаболит - вещество, образующееся в химических реакциях живой клетки.
Метилазы - ферменты, присоединяющие метильную группу к определённым азотистым основаниям в ДНК.
Методы исследований в генетической инженерии. 1) Рестри- кация – расщепление ДНК, необходимо для выделения генов и манипуляции с ними. 2) Гибридизация нуклеиновых кислот – используется для выявления специфической последовательности ДНК и РНК, а также совмещению различных генетических элементов. Используется в полимеразной цепной реакции для амплификации ДНК in vitro. 3) Клонирование ДНК – осуществляется путем введения фрагмента ДНК или их групп в быстрореплицирующиеся генетические элементы (плазмиды или вирусы), что дает возможность размножать гены в клетках бактерии, дрожжей или эукариот. 4) Секвенирование – определение нуклеотидных последовательностей в
клонируемом фрагменте ДНК. Позволяет определить структуру генов и аминокислотную последовательность кодируемых ими белков. 5) Химико-ферментативный синтез полинуклеотидов – часто необходим для целенаправленной модификации генов и облегчения манипуляции с ними.
Механизм действия колхицина - колхицин препятствует «досо- биранию» элементов веретена, если колхицин начал действовать в процессе уже начавшейся сборки веретено (микротубули). Доказательством служит то, что колхицин не действует на уже сформировавшееся веретено.
Механизм дифференциальной окраски еще окончательно не выяснены, различные гипотезы изложены в обзорах А.Ф.Захарова, 1975, А.А.Прокофьева-Бельговской, 1977. Считается, что компоненты краски связываются исключительно с ДНК хромосомы. Предложены две гипотезы: первая так называемая ДНК-вая гипотеза. ДНК-вая исходит из того, что в дифференциальную окраску вовлекается ДНК хромосомы. Если удалить ДНК из хромосомы, то она теряет способность к окраске. То есть, если удалить ДНК, эффект исчезает. Это связывание ступенчато: сначала с ДНК реагирует циклическая молекула метиленового синего, затем молекула эозина с последующим взаимодействием этих молекул между собой и формированием красящего компонента (комплекса). Разная степень или прочность связи красителя с ДНК будет зависеть и от особенностей конфигурации и взаиморасположения ДНК в хромосоме.
Вторая белковая гипотеза исходит из того, что после воздействия трипсином (обработка протеолетическими ферментами) из хромосомы вымываются кислые белки и эти участки не окрашиваются и получается полосатость. Но так как ДНК связана в хромосомах с разными белками, то можно в целом полагать, что рисунок сегментации хромосом зависит от особенностей организации целостного комплекса ДНК в белок.
Механизм образования анеуплоидии - основным механизмом образования анеуплоидии является нерасхождения хромосом в митозе и мейозе, а также отставание хромосом при расхождении в анафазе (Синдром Тернера 45х0), частичное артефактное происхождение. Изучение анеуплоидии представляется очень важным, так как некоторые химические вещества вызывают исключительно геномные мутации. Геномные и структурные мутации изучались
попутно, с точки зрения оценки качества, методики. Специальных исследований по качественному составу анеуплоидии не проводили, так как необходимо изучать много клеток с применением дифференциальной окраски, чтобы знать одни и те же хромосомы теряются каждый раз или разные. Это задача наших дальнейших исследований.
Механизм образования полиплоидии - полиплоидия возникает в результате удвоения хромосом без их расхождения в результате: эндоредупликации, объединения двух наборов хромосом при гибридизации аллоплоидия в результате искусственного воздействия колхицином – К митозы.
Механизм центрических слиянии хромосом Rtr - соединение двух акроцентриков в одну хромосому может происходить 3 способами:
1. В результате реципрокных tr-транслокации с последующей утерей одной из 2-х центромер с небывалым количеством околоцентромерного гетерохроматина.
2. Вследствие разрыва в центромерах и слияния центромер.
3. Вследствие разрыва в коротких плечах акроцентриков и соединения обеих центромер.
Механизм цитогенетического действия фитогемагглютинина (ФГА) - в основе механизма цитогенетического действия ФГА лежит иммунологический механизм, который возникает как реакция «антиген-антитело».
Микросателлит – микросателлитный локус (STR – от английского Short Tandem Repeats): участок ДНК с определённой геномной локализацией, содержащий короткие тандемные повторы.
Миниклетки – клетки, не содержащие хромосомной ДНК. Модификация биополимера — изменение его структуры.
Микрофотографирование хромосом кариотипа позволяет детально изучить морфологию, подсчитать число хромосом в метафазной пластинке, измерить каждую из них. Парижская конференция (1989) рекомендовала следующие основные типы дифференциальной окраски при микрофотографировании и последующего анализа кариотипа: Q – окраска и полосы, выявляемые флуоресцентными красителями (акрихин). G – окраска и полосы. Окраска хромосом красителем Гимзы после воздействия (трипсин, р-р солей и t0С). Наиболее информативный метод. G – окраска и полосы выявляются в районе центромеры. Показано, что в этих областях хромосомы находятся гетерохроматин, содержащий ДНК. N – метод, выявляющий ЯОР в хромосомах ядрышка в интерфазном ядре. При Q и G – окраске затрагиваются одни и те же участки хромосом, ярко флуоресцирующий сегмент при Q окраске соответствует темноокрашенные G-полосы.
Мобильные элементы генома – последовательности ДНК, способные перемещаться внутри генома живых организмов.
Моногибридное скрещивание – скрещивание форм, отличающихся друг от друга по одной паре альтернативных признаков.
Мониторинг (от англ. monitoring) – представляет собой постоянное наблюдение за каким-либо процессом для выявления его соответствия желаемым параметрам или первоначальным предположениям.
Мониторинг – комплексная система наблюдений, оценки и прогноза изменений состояния биосферы или отдельных элементов под влиянием антропогенных воздействий с целью контроля ее качества и изменений.
Морфологическое строение хромосом наиболее четко выражено в стадии метафазы. В этот период хромосома состоит из двух нитей – хроматид, интенсивно окрашивающихся основными красителями. В определении формы хромосом большое значение имеет положение ее обязательного структурного элемента – первичной перетяжки, в районе которой расположена центромера. Центромера делит хромосому на две части (называемые плечами) равной или различной длины. Объективным критерием для отнесения хромосом к той или иной группе служит центромерный индекс – отношение длины короткого плеча к длине хромосомы в процентах. К акроцентрическим хромосомам принято относить хромосомы с центромерным индексом 12,5%, к субметацентрическим от 12,6% до 37,0%, к метацентрическим от 37,1% до 50%.
Морфозы – это резкие изменения в строении органов и проявлении признаков в результате нарушения процесса органогенеза в эмбриональный период онтогенеза, они не наследуются и чаще всего имеют явную патологию. У животных отмечается образование дополнительных конечностей симбиоз (срастание) близнецов, развитие органов в непредназначенном месте.
Мутация – изменение типа, числа или порядка расположения нуклеотидов в генетическом материале.
Моноклональные антитела – антитела со специфичностью к определённому антигену, синтезируемые гибридомами.
Морфогенез – осуществление генетической программы развития организма.
Мутагенез – процесс индукции мутаций.
Мутагены – физические, химические или биологические агенты, увеличивающие частоту возникновения мутаций.
Мутация – изменение генетического материала, часто приводящее к изменению свойств организма.
Мутон – элементарная единица мутирования, т. е. наименьший участок генетического материала, изменение которого представляет собой улавливаемую фенотипически мутацию и приводит к нарушению функции к.-л. гена.
Наследственность – свойство организмов обеспечивать материальную и функциональную преемственность между поколениями, а также повторять определённый тип индивидуального развития.
Наследуемость – доля фенотипической изменчивости в популяции, обусловленная генетической изменчивостью (в отношении к определённому качественному или количественному признаку).
Нитрогеназа – фермент, осуществляющий фиксацию атмосферного азота.
Норма кариотипа (синоним конституциональный кариотипический статус) – особенности кариотипа, являющиеся обычными, то есть нормальными для клеток определенного типа или для всего организма. Норма кариотипа верблюдов: модальное число хромосом 74, в том числе 60 акроцентрических аутосом, 12 метацентрических аутосом и 2 половые хромосомы-гоносомы (XX - у самок, XY - у самцов).
Нуклеазы – общее название ферментов, расщепляющих молекулы нуклеиновых кислот.
Обратная транскриптаза – фермент, катализирующий реакцию синтеза ДНК на матрице РНК.
Олигонуклеотид – цепь ДНК, состоящая из нескольких (от 2 до 20) нуклеотидных остатков.–
Онкогены – гены, чьи продукты обладают способностью трансформировать эукариотические клетки так, что они приобретают свойства опухолевых клеток.
Онкорнавирус – РНК-содержащий вирус, вызывающий перерождение нормальных клеток в раковые; содержит в своем составе обратную транскриптазу.
Оператор – регуляторный участок гена (оперона), с которым специфически связывается репрессор, предотвращая тем самым начало транскрипции.
Оперон – совокупность совместно транскрибируемых генов, обычно контролирующих родственные биохимические функции.
Отбор – это сохранение более приспособленных к определенным жизненным условиям и технологии производства особей или выбор человеком соответствующих его требованиям и устранение менее приспособленных, худших экземпляров.
Относительная длина хромосом – определяется путем отношения длины хромосомы к длине гаплоидного набора, включающего Х-хромосому. Он необходим для построения кариограммы и идиограммы.
Пенетрантность генов – явление, когда один и тот же признак проявляется или не проявляется у особей родственных групп. Пенетрантность определяется по проценту особей в популяции, у которых данный ген проявился. Пенетрантность бывает полная (когда проявляется у 100% особей) и неполная (у определенной части особей).
Плазмида – кольцевая или линейная молекула ДНК, реплицирующаяся автономно от клеточной хромосомы.
Племенное животное – чистопородное, высококлассное, высокопродуктивное животное, отвечающее типу, направлению и уровню продуктивности, стандарту породы, имеющие документально подтвержденное происхождение.
Племенная продукция (материал) – племенное животное, его семя, эмбрионы.
Племенная ценность – уровень генетического потенциала племенного животного, влияющий на хозяйственно-полезные признаки потомства.
Плейотропным действием гена – называют влияние одного гена не на один, а одновременно на несколько признаков. Гены плейотропного действия контролируют синтез ферментов, участвующих в разных обменных процессах в клетке и в организме в целом и оказывающих одновременно влияние на проявление и развитие других признаков.
Полиплоидия – увеличение числа полных хромосомных наборов в четное и нечетное число раз. У верблюдов зарегистрированы триплоидия (3n) и тетраплоидия (4n).
Полилинкер – синтетический олигонуклеотид, содержащий участки узнавания для нескольких рестриктаз (см. рестриктаза).
Полимеразы – ферменты, ведущие матричный синтез нуклеиновых кислот.
Полипептид – белок, полимер, состоящий из аминокислотных остатков, связанных пептидными связями.
Праймер – короткая олиго- или полинуклеотидная последовательность со свободной З’ОН-группой, комплементарно связанная с однонитевой ДНК или РНК; с его 3’-конца ДНК-полимераза начинает наращивать полидезоксирибонуклеотидную цепь.
Прокариоты – организмы, у которых нет клеточного ядра.
Промотор – регуляторный участок гена (оперона), к которому присоединяется РНК-полимераза с тем, чтобы начать транскрипцию.
Протоонкогены – нормальные хромосомные гены, мутации которых могут привести к злокачественному перерождению клетки.
Протопласт – растительная или микробная клетка, лишённая клеточной стенки.
Профаг – внутриклеточное состояние фага в условиях, когда его литические функции подавлены.
Процессинг - частный случай модификации, когда в биополимере уменьшается число звеньев.
Регулон – система генов, разбросанных по всему геному, но подчиняющихся общему регуляторному белку.
Регуляция экспрессии генов – контроль над клеточной структурой и функцией, а также основа дифференцировки клеток, морфогенеза и адаптации.
Рекомбинантная молекула ДНК (в генетической инженерии) – получается в результате ковалентного объединения вектора и чужеродного фрагмента ДНК.
Рекомбинантная плазмида – плазмида, содержащая фрагмент(ы) чужеродной ДНК.
Рекомбинантный белок – белок, полученный в результате экспрессии с рекомбинантной молекулы ДНК, часто получаемый в кишечной палочке.
Рекомбинация in vitro – операции in vitro, приводящие к созданию рекомбинантных молекул ДНК.
Рекомбинация гомологическая – обмен генетическим материалом между двумя гомологичными молекулами ДНК.
Рекомбинация сайт-специфическая – объединение путём разрыва и слияния двух молекул ДНК или участков одной молекулы, происходящее по определённым сайтам.
Рекон – элементарная единица генетической рекомбинации, т. е. минимального участка генетического материала, в пределах которого возможна рекомбинация.
Ренатурация - восстановление исходной пространственной структуры молекул.
Репарация ДНК - исправление повреждений молекулы ДНК, восстанавливающее её первоначальную структуру.
Репликатор – участок ДНК, ответственный за инициацию репликации.
Репликация – процесс удвоения молекул нуклеиновых кислот.
Репликон – молекула ДНК или её участок, находящиеся под контролем репликатора.
Репрессия – подавление активности генов, чаще всего путём блокирования их транскрипции.
Репрессор – белок или антисмысловая РНК, подавляющие активность генов.
Рестриктазы – группа бактериальных сайт-специфических эндонуклеаз, которые узнают определённые участки ДНК длиной от четырёх и более пар нуклеотидов и расщепляют нуклеотидную цепь внутри участка узнавания или вне его, образуя "липкие" или "тупые" концы.
Рестрикты – фрагменты ДНК, образовавшиеся после её гидролиза рестриктазой.
Рестрикционная карта – схема молекулы ДНК, на которой указаны места разрезания её различными рестриктазами.
Рестрикционный анализ – установление мест расщепления ДНК рестриктазами.
Ретровирусы – РНК-содержащие вирусы животных, кодирующие обратную транскриптазу и образующие провирус с хромосомной локализацией.
Рецессивность – неучастие аллеля в формировании признака у гетерозиготной клетки.
Рибонуклеазы (РНКазы) – ферменты расщепляющие РНК.
Сайт – участок молекулы ДНК, белка и т. п.
Секвенирование – установление последовательности звеньев в молекулах нуклеиновых кислот или белков (полипептидов).
Селективные среды – питательные среды, на которых могут расти лишь клетки с определёнными свойствами.
Септум – структура образующаяся в центре бактериальной клетки в конце цикла деления и разделяющая её на две дочерние клетки.
Синдромами называют комплекс патологических изменений фенотипа обусловленная отрицательным действием мутантного гена. То есть плейотропное действие гена может быть как отрицательным, так и положительным.
Скрининг – поиск в рассевах клеток или фагов тех колоний, которые содержат рекомбинантные молекулы ДНК.
Слитый белок (полипептид) – белок, образованный слиянием двух различных полипептидов.
Соматические гибриды – продукт слияния неполовых клеток.
Соматические клетки – клетки тканей многоклеточных организмов, не относящиеся к половым.
Спейсер – в ДНК или РНК – некодирующая последовательность нуклеотидов между генами; в белках - аминокислотная последовательность, связывающая соседние глобулярные домены.
С-полосы — это по существу структурный гетерохроматин. Известно, что хромосома делится на районы эухроматина и гетерохроматина, причем последний подразделяется на собственно гетеро- оматин (ГР) и гетерохроматизированный материал (факультативный гетерохроматин). Эурхроматин и гетерохроматин различаются по плотности конденсации и это различие может иметь значение в восприятии красителя Гимза. Классическая точка зрения гетерохроматин – это более поздно редуплицированные участки хромосом. ГР образует центромеры, теломеры, большую часть половых хромосом. Функция ГР структурная, защитная роль, роль в клеточном метаболизме, транскрипции, эволюции кариотипов. Так, многочисленные повторы нуклеотидов ГР внутри эухроматических районов могут выполнять роль остановок в транскрипции или быть участками инициации для полимераз.
Сплайсинг – процесс формирования зрелой мРНК или функционального белка путём удаления внутренних частей молекул – интронов РНК или интеинов у белков.
Способ картирования G-полос – Дретсом и Сеунером (1974,1975) предложен способ картирования G-полос, основанный на количественной оценке относительной удаленности каждой полосы от центромеры.
Структурные мутации – это группа мутаций связана с изменениями формы, размеров хромосом, порядка расположения генов (изменения групп сцепления), утратой или добавкой отдельных фрагментов.
Суперпродуцент – микробный штамм, нацеленный на синтез определённого продукта в высокой концентрации.
Сущность генетической инженерии в животноводстве состоит в целенаправленном конструировании особых гибридных молекул вне организма с последующим их введением в организм животных. При этом гибридные молекулы (рекомбинантные ДНК) становятся составной частью генетического аппарата данного организма. В результате наследственная программа организма изменяется, ему сообщаются новые генетические, биохимические и физиологические свойства.
Сущность методики Раджабли (Цитология, Т.XV, №2, 1973, 1527-1538) – Это модификация методики Сибрайт (Sеabright, 1971) и Ивенса с соавт. (1971 г.). Препараты на 15-20 сек. помещали в 0,25% раствор трипсина, нагретый до 28-300С. Затем отмывали в буфере (0,6 м NaC1 – 0,06 м цитрат натрия, рН 7,0) в течение 5 мин и затем инкубировали в течение 1 часа в свежей порции того же буфера при t-620С.
Трансген – ген, интегрированный в ген реципиента, называют трансгеном. Благодаря переносу генов у трансгенных животных возникают новые признаки, которые при селекции закрепляются в потомстве. Трансгенных животных получают путем микроинъекции рекомбинантной ДНК в извлеченные из донорских организмов эмбрионы и дальнейшей пересадки инъецированных эмбрионов в яйцеводы или после культивирования в матку синхронизированных реципиентов.
Трансгенные животные – животных, несущих в своем геноме рекомбинантный (чужеродный) ген.
Трансдукция – перенос фрагментов ДНК с помощью бактериофага.
Транскрипция – синтез РНК на ДНК-матрице; осуществляется РНК-полимеразой.
Транскрипт – продукт транскрипции, т. е. РНК, синтезированная на данном участке ДНК как на матрице и комплементарная одной из его нитей.
Транскриптаза обратная – фермент, синтезирующий по РНК как по матрице комплементарную ей однонитевую ДНК.
Трансляция – синтез полипептидной цепи белков, осуществляемый в рибосомах.
Транспозон – генетический элемент, реплицируемый в составе репликона и способный к самостоятельным перемещениям (транспозиции) и интеграции в разные участки хромосомной или внехромосомной ДНК.
Трансфекция – трансформация клеток с помощью изолированной ДНК.
Трансформация – изменение наследственных свойств клетки, вызванное поглощенной ДНК.
Трансформация (в молекулярной генетике) – перенос генетической информации посредством изолированной ДНК.
Трансформация (онкотрансформация) – частичная или полная дедифференцировка клеток, вызванная нарушением регуляции роста клеток.
Умеренный фаг – бактериофаг, способный лизогенизовать клетку и в виде профага находиться внутри бактериальной хромосомы или в плазмидном состоянии.
Фактор F (фактор фертильности, половой фактор) – коньюгативная F-плазмида найденная в клетках Е. coli .
Фенотип – внешнее проявление свойств организма, зависящих от его генотипа и факторов окружающей среды.
Фенокопия – это изменение признака под влиянием действия внешней среды, как и под влиянием действия генов, но возникшие особенности не являются наследственными. Разнообразные фенокопии могут возникнуть после перенесенных заболеваний во время беременности, нарушения баланса микро-макроэлементов и витаминов. У фенокопии нормальному действию не мутантного гена (аллеля) препятствует факторы внешней среды.
Физиологическая гипоплоидия – показатель доли физиологически гиподиплоидных клеток, определяется как разница между долей гиподиплоидных клеток и гипердиплоидных клеток.
Физиологическая гипоплоидия возникает за счет физиологического явления ослабления осморезистентности клеток, не выдержавших гипотонизации. Мембраны клеток разрываются и хромосомы теряются.
Физиологическая гипоплоидия рассчитывается по формуле:
ФГ = Гпо — Гпр
Гиподиплоидные – гипердиплоидные клетки
Например:
ФГ = 10 – 2,4 = 7,6%
У сельскохозяйственных животных обычно частота гиподи- плоидных клеток выше гипердиплоидных.
Химеры – лабораторные гибриды (рекомбинанты).
Хозяйственное долголетие – это длительность использования животного и способность его сохранять экономически выгодный уровень продуктивности и давать качественное потомство, то есть не утратившего способность к воспроизводству. Например, продолжительность жизни лошади составляет 67 лет, а хозяйственная 20.
Хроматин – нитчатые комплексные молекулы дезоксирибонуклеопротеида (ДНП), которые состоят из ДНК, связанной с гистонами.
Хроматиновая нить – это гистон и ДНК, объединенные в структуру, представляющая собой двойную спираль ДНК, окружающую гистоновый стержень. Хроматиновая нить образует спираль диаметром около 25 мкм, что находится на грани разрешающей способности современных световых микроскопов. По способности окрашиваться ядерными красителями хроматиновые нити подразделяют на две группы: эухроматин и гетерохроматин. Последний окрашивается более интенсивно.
Хромосомы – органоиды клеточного ядра, являющиеся носителями генов и определяющие наследственные свойства клеток и организмов.
Хромосомы – представляют собой нитевидные нуклеопротеидные структуры, способные к саморепродукции и сохранению своих морфологических особенностей на протяжении ряда поколений. Они удваиваются в результате идентичной репродукции перед каждым клеточным делением, а затем распределяются поровну между дочерними клетками. Хромосомы состоят из хроматина, который содержит ДНК (40%), гистоны (40%), негистоновые хромосомные белки (20%) и небольшое количество РНК.
Цитогенетический мониторинг – комплексная система наблюдений, оценки и прогноза изменений кариотипа животных в разрезе вида, породы, возраста, половой принадлежности, условий содержания и кормления животных с целью контроля их качества и изменений.
Цель генетической инженерии – создание рекомбинантных ДНК, которые придавали бы организму новые, полезные для человека свойства.
Центромера – локус на хромосоме, физически необходимый для распределения гомологичных хромосом по дочерним клеткам.
Шайн-Далгарно последовательность – участок прокариотической мРНК, необходимый для посадки на неё рибосом и её правильной трансляции. Содержит последовательность нуклеотидов, комплементарную 3’-концу 16S рибосомной РНК.
Шаффлинг ДНК – рекомбинация фрагментов генов двух и более гомологичных белков. Трехступенчатый процесс, включающий разрушение родительских молекул ДНК и два раунда амплификации (без праймеров и со специально подобранными), с целью получения восстановленных по длине, но измененных по составу (с перетасованными последовательностями) химерных молекул ДНК, с существенно улучшенными или новыми свойствами кодируемых ими белков
Штамм – линия клеток, бактерий (или вирусов), ведущая начало от одной клетки (или вируса).
Экзон – сохраняющаяся при сплайсинге часть интронированного гена.
Экзонуклеаза – фермент, гидролизующий фосфодиэфирные связи с концов ДНК.
Эксплантат – выделенный из организма материал какой-либо ткани.
Экспрессия гена – процесс реализации информации, закодированной в гене. Состоит из двух основных стадий – транскрипции и трансляции.
Экспрессивность генов – это степень фенотипического проявления гена как мера силы его действия, определяемая по степени развития самого признака. В системе взаимосвязанных генов развитие одного признака может зависеть от взаимодействия многих генов и один ген может влиять на развитие и проявление нескольких признаков.
Электрофорез – разделение электрически заряженных полимеров в электрическом поле. Обычно ведется в гелях (гель-электрофорез), чтобы зоны разделяемых молекул не размывались тепловым движением.
Эндонуклеаза – фермент, гидролизующий фосфодиэфирные связи внутри нити ДНК.
Энхансер – регуляторный участок ДНК, усиливающий транскрипцию с ближайшего к нему промотора.
Эпизоотический мониторинг – система сбора количественных данных о распространении болезней животных, включая эпизоотологическое обследование и информацию о закономерностях развития конкретной болезни животных, природно-географических и экономических (хозяйственных) условиях территорий их обитания (содержания, разведения), проводимых ветеринарно-санитарных мероприятиях, и последующая их статистическая обработка для анализа эффективности ветеринарно-санитарных мероприятий и прогнозирования возникновения, развития и ликвидации эпизоотий или панзоотий.
Эукариоты – организмы, клетки которых содержат ядра.

















































































































Комментарии (0)