Влияние различных доз биоугля в субстрате на транспорт макроэлементов у подвоев цитранжа каризо и сопутствующий анализ экспрессии генов
Биоуголь может представлять собой ценный компонент субстрата для производства саженцев цитрусовых.
Аннотация
Цитрусовые питомники значительно увеличивают затраты на производство из-за применения строгих технических и санитарных протоколов. Используемые субстраты для выращивания обычно основаны на торфе, ограниченном ресурсе, который становится все более дефицитным и, следовательно, более дорогим. Среди альтернатив торфу — биоугля, который может представлять собой ценный компонент субстрата для производства саженцев цитрусовых. Было сравнено три субстрата для выращивания, каждый из которых содержал 50% песчаной вулканической почвы, а оставшиеся 50% составляли: (i) биоугля 50%; (ii) черный торф 25% + биоугля 25%; и (iii) черный торф 25% + лапилус 25% в качестве контроля. Была оценена влияние на агрономические показатели саженцев цитрусовых, а также участие конкретных генов в поглощении макроэлементов. Разрушающие и молекулярные анализы проводились на листьях и корнях в два разных периода года: февраль и апрель. На основе физико-химических параметров и роста саженцев можно предположить, что торф может быть частично заменен биоугля из древесины хвойных пород в общем количестве 25 или 50%. Общее сравнение средних значений из проб и различных анализируемых субстратов показало, что в феврале оцениваемые гены, участвующие в поглощении и транспорте питательных веществ, дифференциально экспрессировались как в листьях, так и в корнях, в то время как в апреле экспрессия была нестабильной. Кроме того, общее сравнение между анализируемыми тканями показало, что в большинстве случаев экспрессия была выше в корнях, чем в листьях. В целом, сравнение между растениями, выращенными в разных субстратах, указало, что субстрат с 50% биоугля демонстрировал наивысшие уровни экспрессии.
1. Введение
Индустрия цитрусовых питомников имеет высокое значение в Средиземноморском бассейне, где Италия занимает третье место по производству. Примерно 1000 ферм производят цитрусовые растения на 16 000 гектарах, нанимая 80 000 работников и генерируя около 600 миллионов евро, из которых 50% предназначены для экспорта. В среднем цикл от посева до товарного горшечного привитого растения занимает 36 месяцев. В этот период основные затраты для производителей возникают из-за экономической амортизации защитных конструкций, покупки семян и горшков, субстратов для выращивания, питания (как твердого, так и через фертигацию) и управления формированием в течение трех лет. Что касается субстратов для выращивания и питания, торф признан основным органическим материалом, в то время как питание растений в основном полагается на минеральные удобрения. Хотя эти технические средства обеспечивают высокую дренажность и аэрацию, тем самым улучшая подходящую среду для развития корней, питомники сталкиваются с двумя фундаментальными вызовами: (1) необходимость замены торфа, поскольку он является дорогим, дефицитным и невозобновляемым ресурсом; и (2) необходимость улучшения эффективности питания для снижения затрат на производство и минимизации загрязнения грунтовых вод. Более того, в контексте изменения климата растет глобальный интерес к устойчивому сельскому хозяйству, которое стремится смягчить выбросы СО2 от традиционных практик путем увеличения ввода углерода в почву [1] и замены минеральных удобрений органическими альтернативами [2]. Среди потенциальных альтернативных субстратов биоугля показывает многообещающие результаты как высокопроизводительный субстрат для выращивания. Биоугля — это твердый материал, получаемый из термохимической конверсии различных биомасс в отсутствие кислорода [3,4].
Роль биоугля в качестве почвенной добавки или компонента субстрата для выращивания в питомниках была широко исследована [5,6]. Его влияние на питание растений считается косвенным эффектом благодаря содержанию углерода, способности снижать выщелачивание, способности мобилизовать питательные вещества и модификации физических и химических свойств почвы и/или субстрата для выращивания [7]. Однако, на основе его состава, также возможно предположить, что он играет прямую роль в питании растений, влияя на системы транспорта питательных веществ под генетическим контролем. Для достижения адекватного роста растениям требуется вклад шестнадцати основных минеральных элементов, которые делятся на три класса: макроэлементы (N, Р, К), поглощаемые в относительно высоких концентрациях, и 11 мезо- и микроэлементов (С, Н, О, S, Са, Mg, CI, Ее, Мп, Zn, Си, В, Мо), необходимые в гораздо меньших количествах. За последние двадцать лет были достигнуты значительные успехи в идентификации генов-транспортеров минеральных элементов в различных видах растений, особенно в регуляции и использовании в модельных растениях, таких как Arabidopsis и рис [8]. Транспортеры растений предназначены для перемещения минеральных элементов в разные органы, и были идентифицированы многочисленные типы транспортеров и систем транспорта в различных видах растений [9].
Хотя быстрый прогресс в секвенировании геномов предоставил много информации о транспортерах через поиски гомологии, роль большинства транспортеров в растениях не была тщательно исследована. Азот, фосфор и калий — самые важные макроэлементы и тесно связаны с продуктивностью культур. Несколько исследований сосредоточились особенно на модельных растениях [9,10,11,12,13,14,15]. В цитрусовых некоторые исследования сосредоточились на транспортерах азота [16,17,18,19,20,21,22], в то время как несколько исследований касались фосфора и калия [23,24,25,26].
Это исследование направлено на изучение влияния биоугля на агрономические показатели саженцев подвоя Carrizo citrange (Citrus sinensis (L.) Osbeck x Poncirus trifoliata (L.) Raf.), выращенных в горшках с использованием субстратов, содержащих разные количества биоугля, а также уровни экспрессии некоторых генов, участвующих в транспорте макроэлементов.
Это исследование направлено на оценку уровней экспрессии некоторых генов, участвующих в транспорте макроэлементов, а также влияния биоугля на агрономические показатели саженцев подвоя Carrizo citrange (Citrus sinensis (L.) Osbeck x Poncirus trifoliata (L.) Raf.), выращенных в горшках с использованием субстратов, содержащих разные количества биоугля.
2. Материалы и методы
Это исследование проводилось в Маскали (Катания), южная Италия, в коммерческом питомнике, производящем сертифицированные цитрусовые деревья. Саженцы подвоя Carrizo citrange использовались и управлялись в питомнике, как описано Ферлито и соавторами [1]. После пересадки саженцы удобрялись каждые шесть месяцев удобрением с медленным высвобождением, содержащим азот (N), фосфор (Р), калий (К) и магний (Мд) в соотношениях 16:8:10:2. Удобрение также содержало основные микроэлементы. Отбор проб листьев и корней проводился через 14-16 месяцев после пересадки (в феврале и апреле соответственно), что соответствует 24-26 месяцам после посева.
Было подготовлено три субстрата для выращивания, каждый из которых содержал 50% песчаной вулканической почвы, а оставшиеся 50%: биоугля 50% (850), черный торф 25% + биоугля 25% (ВР25В25) и черный торф 25% + лапилус 25% (BP25L25). BP25L25 был субстратом, обычно используемым в питомнике, где проводилось испытание, и служил контролем. Биоугля получали из древесины хвойных лесов и производили путем пиролиза при 400 °C. Эксперимент проводился в рандомизированном полном блоковом дизайне с тремя повторениями, где каждый повтор включал 20 растений, всего 180 растений (3 субстрата х 3 повтора х 20 растений на повтор).
Основные физико-химические характеристики песчаной вулканической почвы, черного торфа и биоугля, используемых в качестве компонентов для изученных субстратов для выращивания, показаны в Таблице 1.
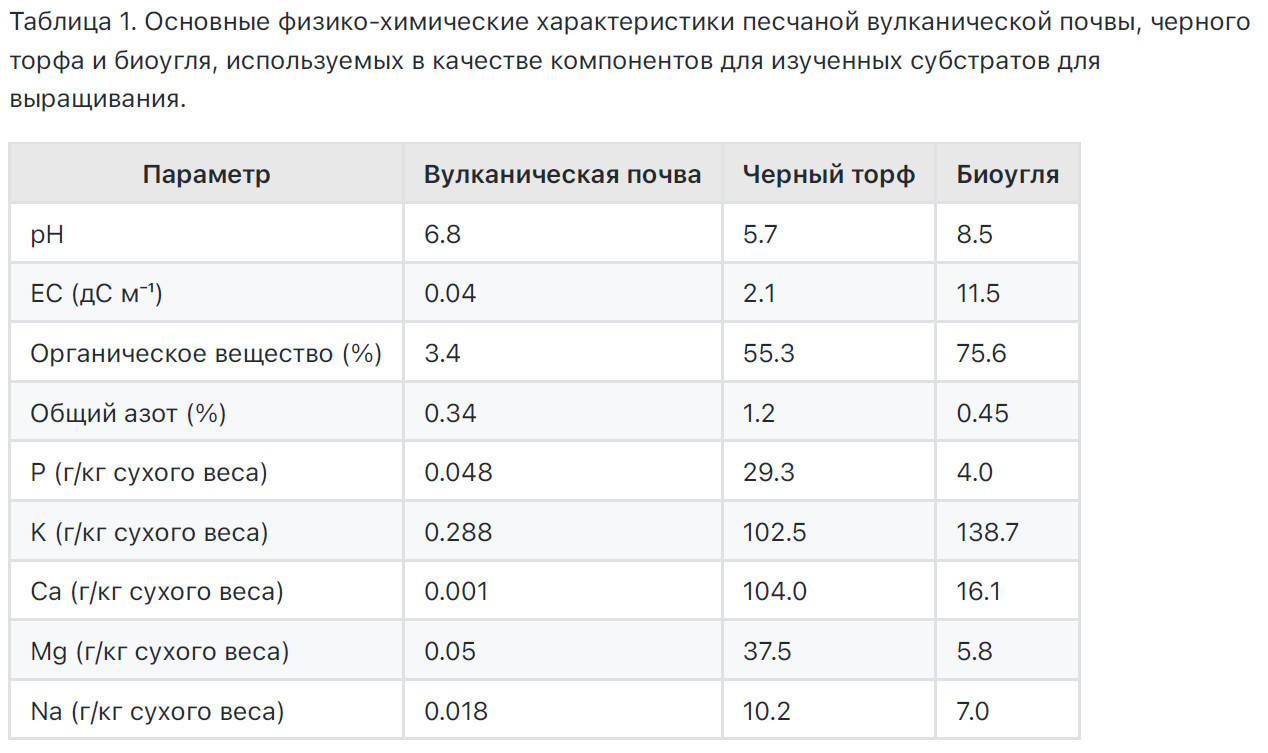
Для каждого субстрата для выращивания, использованного в этой работе, определяли общую доступную воду (TAW), воду при полевом насыщении (WFS), содержание воды при полевой влагоемкости (WCFC) и при точке постоянного увядания (WCPWP), порозность, заполненную водой (WFP), общую порозность (ТР) и воздушное пространство смесей (AS). Все примененные методы для вышеуказанных анализов описаны Ферлито и соавторами [1]. Химические параметры, оцениваемые, включали азот (г кг1), органическое вещество (ОМ), общий органический углерод (ТОО, мг кг1), pH и ЕС [27].
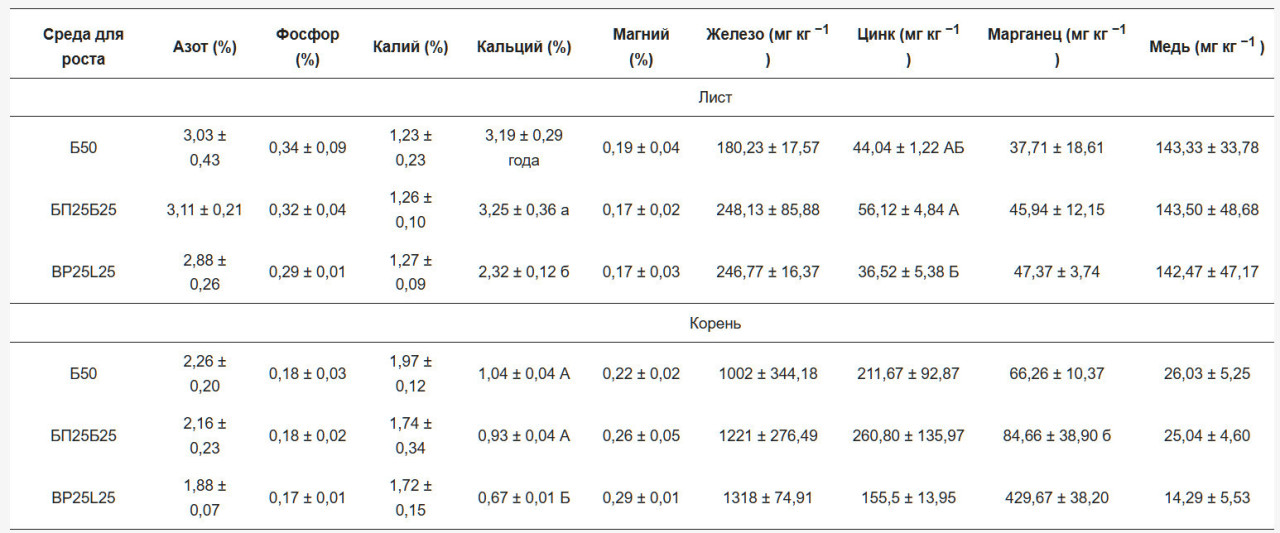
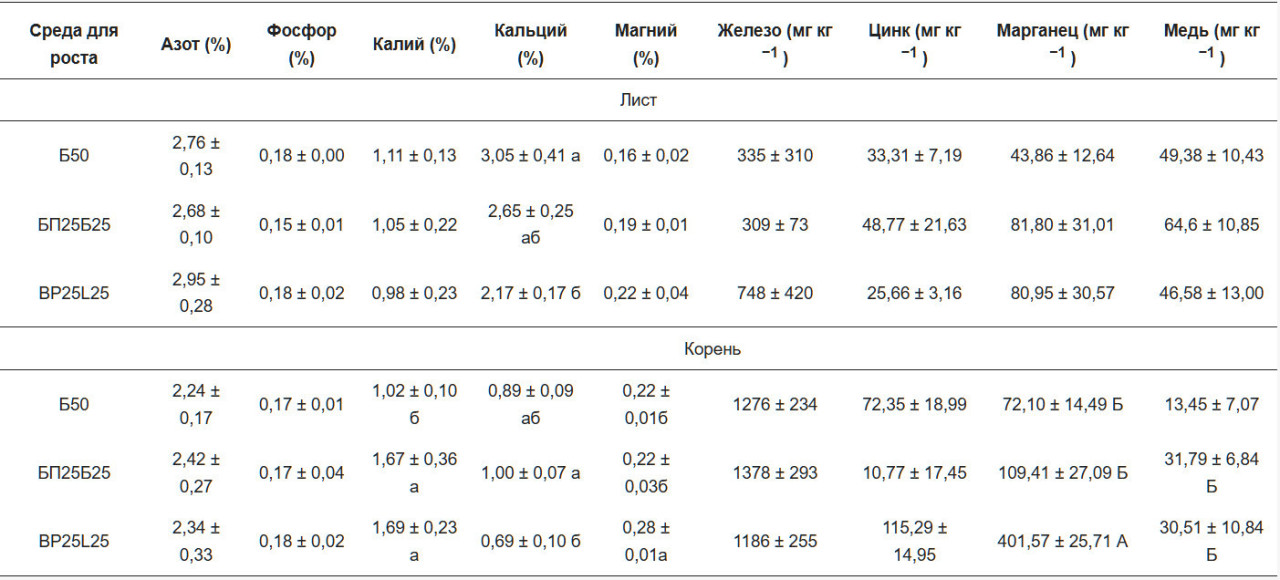
Листья и корни, собранные в феврале и апреле, были химически охарактеризованы для определения концентраций макроэлементов (N, Р, К и Са) и микроэлементов (Fe, Zn, Мп и Си). Следуя экспериментальному протоколу, образцы листьев и корней высушивались в сушилке при 60 °C и измельчались в мелкий порошок (Ika-M 20 Universal mill, 20 000 об/мин). Annquot 0.5 г порошка анализировался на общий азот (%N) с использованием микро-Кьельдаля [28] с BCichi Digest Automat К-438, BCichi Scrubber В-414 и BCichi Distillation Unit K370 (BCichi, Швейцария). Для концентраций P, К, Ca, Mg, Fe, Zn, Мп и Си отдельный aliquot 0.5 г сжигался в муфельной печи при 600 °C в течение 12 ч. Полученная зола растворялась в 0.5 мл 69.5% азотной кислоты (Superpure; Merck, Дармштадт, Германия), переносилась в 50 мл мерную колбу и разбавлялась деионизованной водой для достижения конечной концентрации азотной кислоты 1/100 об/об. Раствор анализировался с использованием спектрометрии с индуктивно-связанной плазмой- оптической эмиссионной спектрометрии (ICP-OES, Optima 2000 DV, PerkinElmer).
Рост саженцев наблюдался каждые 60 дней, начиная с одного месяца до пересадки (декабрь) до апреля следующего года, путем измерения высоты стебля и базального диаметра растений. Рост саженцев определялся разрушающими измерениями, проводимыми на шести саженцах на субстрат, для определения распределения сухого вещества в корневой системе и листьях. Измерения проводились два раза, зимой (февраль) и весной (апрель), соответствующие 14-му и 16-му месяцу после пересадки саженца в горшок, соответственно.
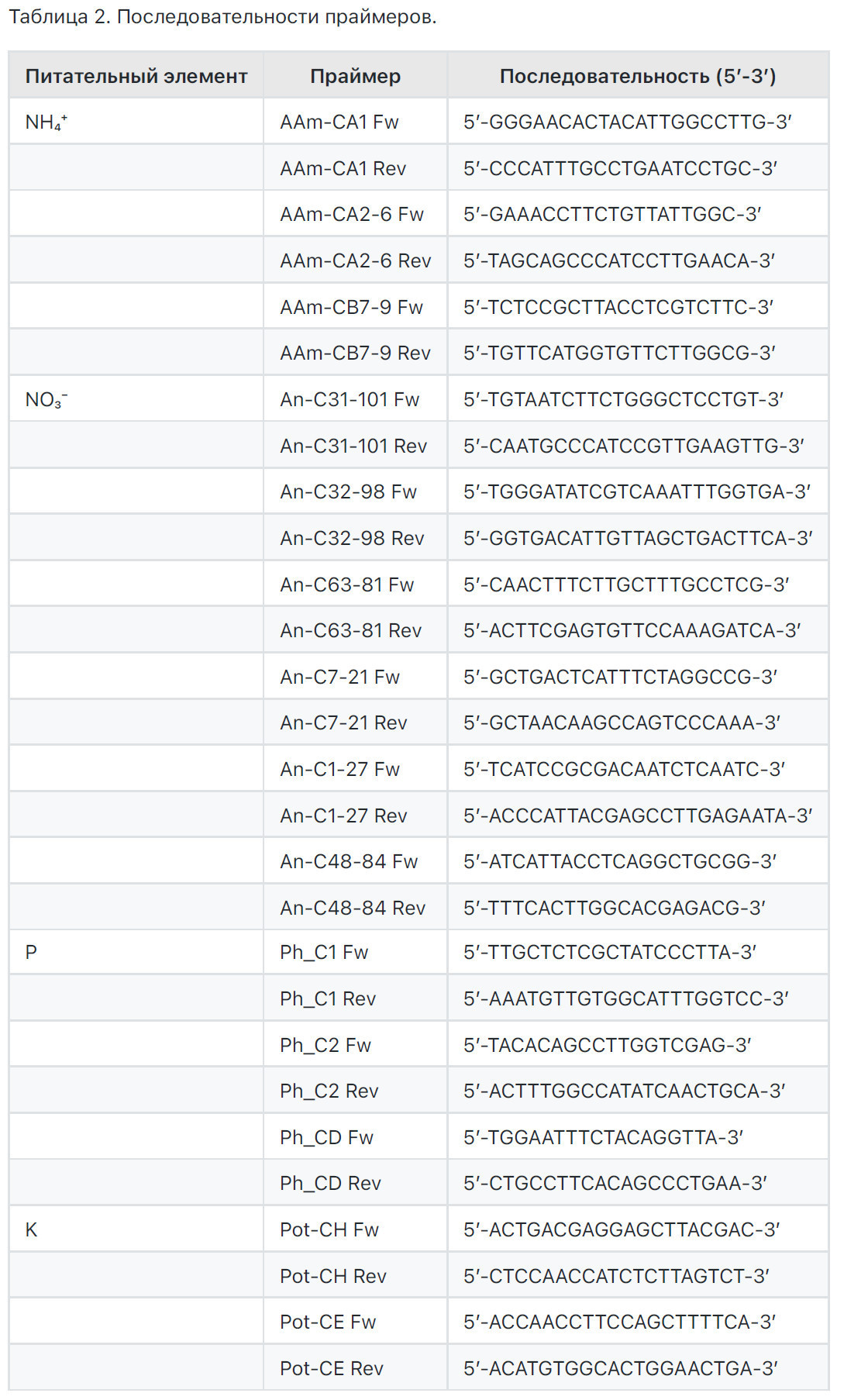
Все гены, участвующие в транспорте N, Р и К, были выбраны из баз данных NCBI [29] и С. sinensis Annotation Project (CAP) [30], включая название "транспортер" в качестве ключевого слова. Мы извлекли 24 последовательности, кодирующие семейство генов Ammonium Transporter (АМТ), 77 для нитрата, принадлежащих к семействам nitrate transporter (NRT) и nitrate transporter/peptide transporter (NRT/PTR), 38 для фосфора, принадлежащих к семейству Phosphate Transporter genes (PHTs) и семейству PHO (ранее идентифицированному в Arabidopsis как белок, участвующий в загрузке неорганического фосфата (Pi) в ксилему корней), и 36 для калия, принадлежащих к К+ uptake permeases КТ (К+ transporter)/HAK (high-affinity K+)/KUP (К+ uptake) family (HAK/KUP/KT).
Мы выровняли последовательности каждого семейства генов с использованием Clustal Omega [31], и соответствующая кладограмма была построена с использованием программного обеспечения Molecular Evolutionary Genetics Analysis (MEGA X) версии 10.0,6 [32,33] с использованием метода Maximum Likelihood и модели Tamura-Nei [34] (Рисунок 81). Это выравнивание позволило выбрать уникальные последовательности для каждого семейства генов, производя консенсусную последовательность, полезную для дизайна прямых и обратных праймеров для количественной ПЦР в реальном времени (qRT-PCR). Мы разработали три (AAm-GAI, AAm-CA2, AAm-CB7-9) и шесть (Ап-С31-101, Ап-С32-98, Ап-С63-81, Ап-С7-21, Ап-С1-27, Ап-С48-84) пар праймеров для последовательностей транспортеров аммония (NH4+) и нитрата азота (NO3’), соответственно; три (Ph_C1, Ph_C2, Ph_CD) пары праймеров были разработаны для фосфора (Р) и две (Pot-CH, Pot-CE) для калия (К) (Таблица 2).
Примерная последовательность для каждого кластера была бластирована против самого недавнего генома С. sinensis (Citrus Genome Database: https://www.citrusgenomedb.org/, доступ 4 ноября 2024 г.) для подтверждения различий между кластерами. Анализ BLAST проводился для идентификации ортологичных последовательностей к С. sinensis в Arabidopsis thaliana (At) и Populus trichocarpa (Pt) для вывода предполагаемой подобной функции (Таблица S1).
Общая РНК экстрагировалась из листьев и корней в два разных периода года, февраль и апрель, с использованием набора SV Total RNA Isolation System (Promega, Мэдисон, Wl, США). Количество общей РНК измерялось с использованием спектрофотометра Nanodrop 1000 (Thermo Fisher Scientific, Уолтем, MA, США), и качество проверялось электрофорезом в геле (а га раза 0.8% в ТАЕ 1х). кДНК синтезировалась с использованием 1 мкг общей РНК без ДНКазы, согласно инструкциям производителя (High-Capacity cDNA Reverse Transcription Kit; Applied Biosystems, Фостер-Сити, CA, США). Три биологических повторения для каждой ткани (листья и корни) из растений в горшках, выращенных в трех разных субстратах, использовались для выполнения qRT-PCR с 100 нг кДНК. Анализы проводились с использованием системы 7500 real-time PCR System (Applied Biosystems, Фостер-Сити, CA, США) c SYBR® Master Mix (Applied Biosystems, Фостер-Сити, CA, США), согласно протоколу производителя. Фактор элонгации 1а (EF; AY498567) использовался в качестве референсного гена [35]. Относительная квантификация нормализовалась к референсному гену ЕЕ, и кратное увеличение рассчитывалось с использованием метода стандартной кривой.
Статистический анализ использовал приложение StatSoft 6.0 (Statistic for Windows. StatSoft, Талса, OK, США, 2001) для анализа дисперсии (ANOVA), и разделение средних по тесту Tukey HSD проводилось при р < 0.05, р < 0.01 и р < 0.001.
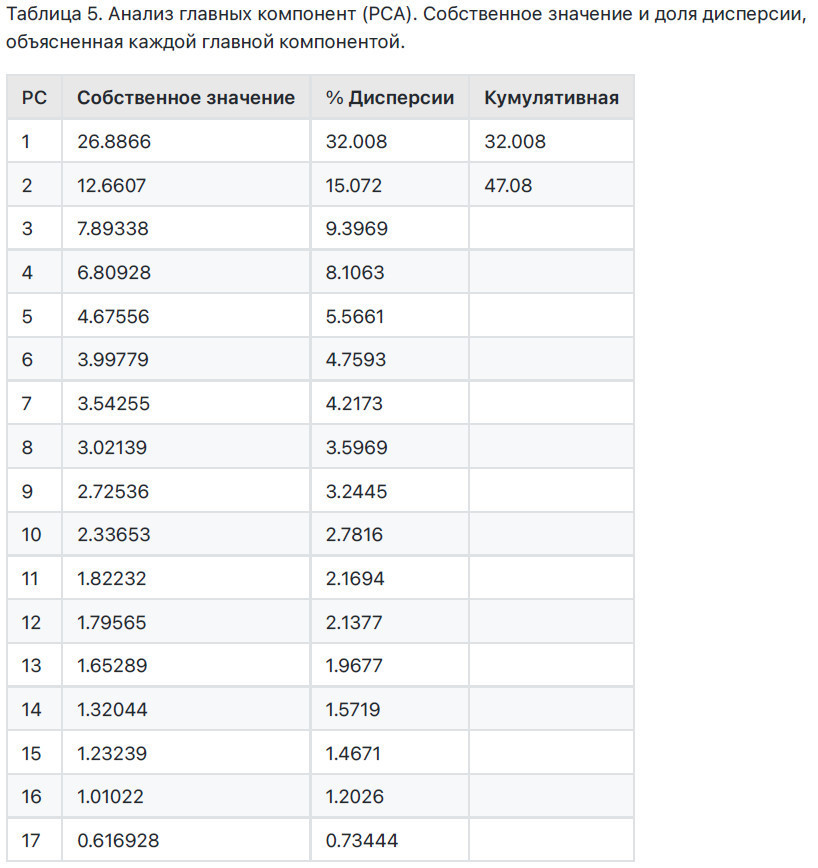
Анализ главных компонент (РСА) проводился с использованием данных, собранных в феврале и апреле, состоящих из физических и химических характеристик трех субстратов для выращивания (850, ВР25В25 и BP25L25), физико-химических свойств, распределения биомассы саженцев Carrizo citrange и значений экспрессии различных анализируемых генов. РСА проводился с использованием программного пакета PAIeontological Statistics (PAST), версия 2011 [36].
3. Результаты и обсуждение
Это исследование оценивало эффекты субстратов для выращивания, содержащих разные количества биоугля, с использованием многодисциплинарного подхода. Конкретно, учитывались как агрономическое, так и генетическое поведение саженцев Carrizo. Что касается агрономических эффектов биоугля, мониторинг роста саженцев, генетические анализы проводились для лучшего понимания участия генов, принадлежащих разным семействам, в транспорте и поглощении макроэлементов.
Основной сезон роста для растений обычно простирается от апреля до октября. Однако февраль и апрель были выбраны для отбора проб, потому что вегетативный рост начинается весной на широте, где проводилось исследование. Примерно 30-40 дней до начала этого роста уже происходит активация генов, связанных с этим процессом, используя первые доступные ресурсы питательных веществ. В этот период реакция генов, ответственных за поглощение и транспорт питательных веществ, вероятно, более немедленная, и их экспрессия более выраженная.
В течение первых шести месяцев не наблюдалось значительных различий в высоте и диаметре стебля саженцев; на 8-м месяце (июнь) значительное увеличение обоих параметров было зарегистрировано для саженцев, растущих в субстрате BP25L25, в то время как не наблюдалось различий в саженцах, выращенных в субстратах, содержащих биоугля (В50, ВР25В25). После восьмого месяца поведение саженцев было похожим во всех трех субстратах для выращивания, а также во время сезона прививки (апрель) (Рисунок 1А,В). Разрушающие анализы зимой (февраль) и весной (апрель) (Рисунок 2A-D) показали, что распределение сухого вещества, накопленного в листьях и корнях, было похожим для всех субстратов для выращивания. Однако между двумя периодами отбора проб (февраль и апрель) и во время первого периода вегетативного возобновления роста было выявлено разное поведение среди разных субстратов для выращивания. Конкретно, для листьев, собранных с саженцев, выращенных в BP25L25, мы наблюдали снижение (-25%) в распределении биомассы, в то время как в В50 мы зарегистрировали увеличение (+22%). Эта тенденция также наблюдалась в корнях. В частности, саженцы, выращенные в BP25L25, показали снижение (-20%) между февралем и апрелем, в то время как те, собранные из ВР25В25 и В50, зарегистрировали увеличение +27% и +8%, соответственно.
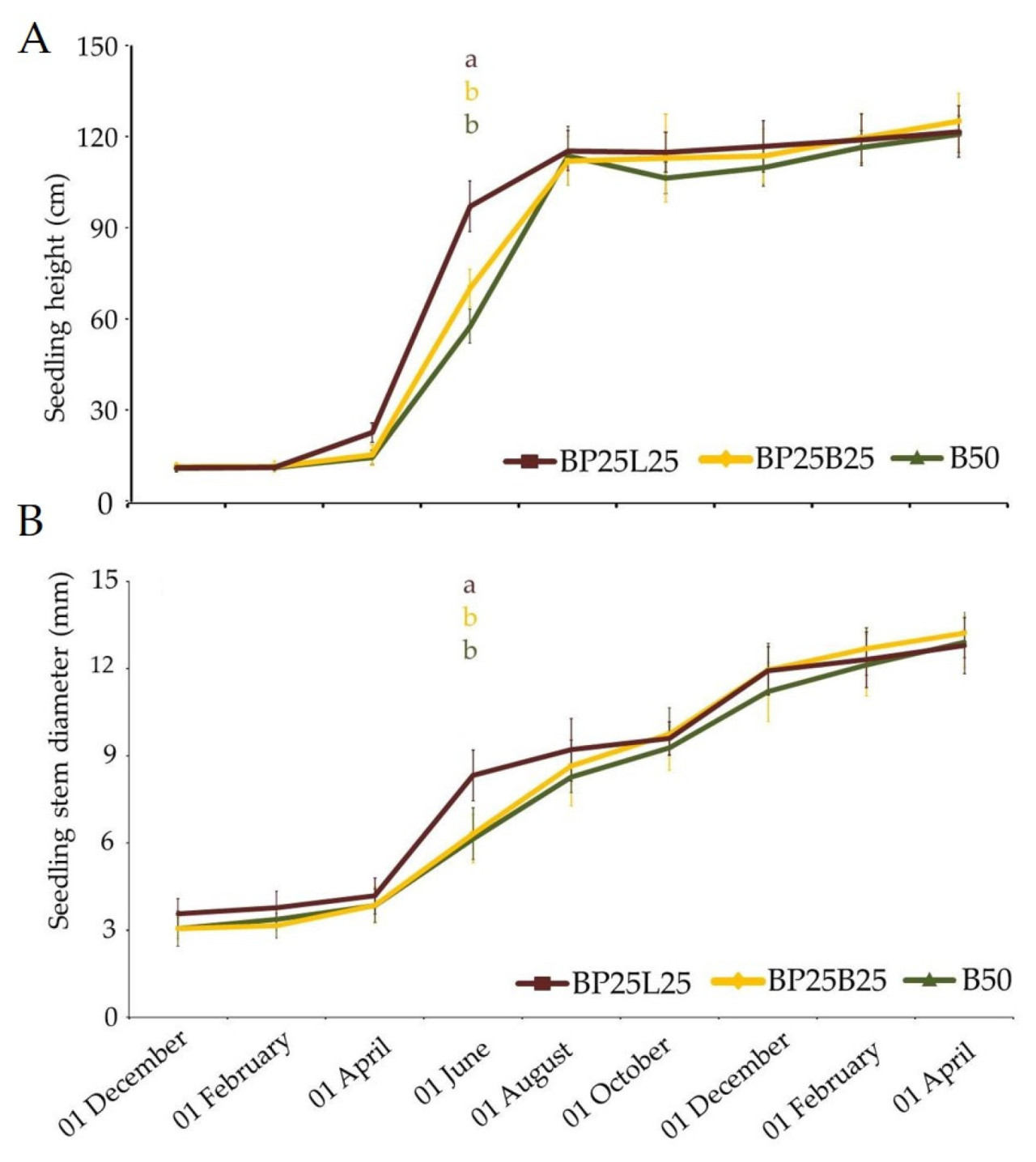
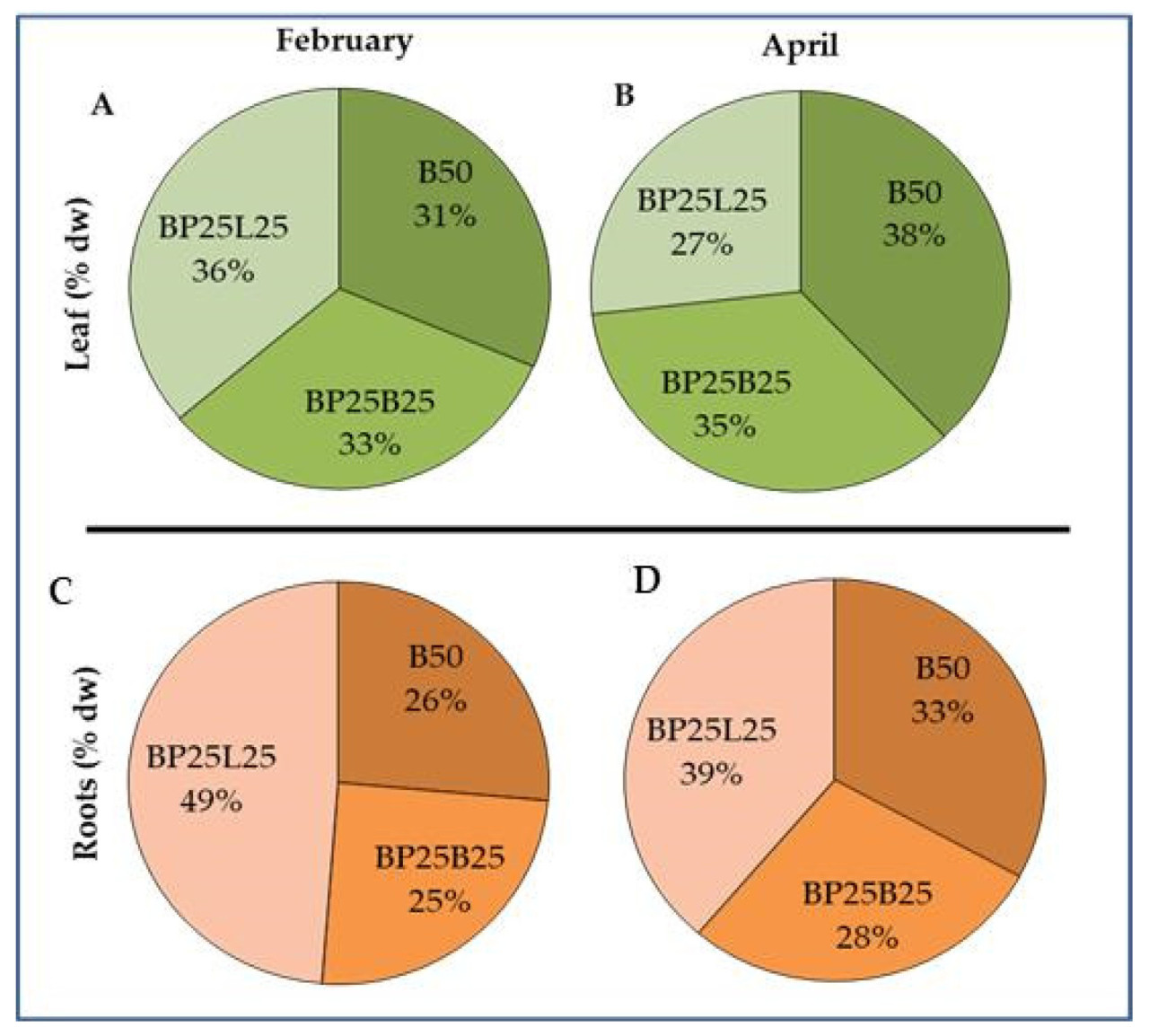
Данные по морфологии саженцев (увеличение высоты саженца, диаметра и распределения сухого вещества) в листьях и корневых системах показали, что накопление сухого вещества было наиболее очевидным во время первых стадий управления саженцами; конкретно, те, выращенные в BP25L25, были наиболее реактивными только во время средней фазы (июнь). Однако отсутствие роста биомассы над- и подпочвенной для листьев и корней как в феврале, так и в апреле указывало, что эффективность корневой системы была удовлетворительной. Самое важное, саженцы в субстратах для выращивания, содержащих биоугля, в основном в В50, показали лучшие показатели во время первого периода вегетативного роста (интервал февраль-апрель). Более того, во время разрушающих измерений наблюдалось снижение роста корней в нижних слоях контейнера, указывая, что качество субстрата было скомпрометировано на этом уровне. Это приписывалось накоплению мелких частиц на уровне дна контейнера, что снижало порозность (не показано). Было сообщено, что когда объемная плотность увеличивается, количество больших пор уменьшается, делая более трудным для корней деформировать и смещать частицы субстрата, тем самым ограничивая скорости удлинения корней [37] и влияя на развитие и морфологию корней. Конкретно, когда субстрат был более компактным, корни концентрировались в верхней части горшка для обеспечения развития надпочвенной биомассы и адекватной аэрации корней.
Не было найдено значительных различий в содержании азота в листьях и корнях в обе даты отбора проб. Статистически значимые различия в листьях субстрата В50 и корнях ВР25В25 были найдены для Са в феврале. В апреле содержание Са в обоих субстратах для выращивания было значительно выше, чем в BP25L25 в листьях и корнях. В феврале субстрат BP25L25 показал значительно более высокие значения магния и марганца в корнях по сравнению с другими двумя субстратами (Таблица 3 и Таблица 4).
Из выравнивания последовательностей, идентифицированных для каждого семейства генов, была получена кладограмма, из которой было возможно выделить разное количество кластеров для каждого исследуемого питательного элемента (3 для NH4+, 6 для NO3’, 3 для Р и 2 для К) (Рисунок 81).
Генетический анализ генов, участвующих в транспорте и поглощении питательных веществ, проводился для сравнения уровней экспрессии в листьях и корнях саженцев, выращенных на трех разных субстратах (В50, ВР25В25 и BP25L25) в два разных времени отбора проб, февраль и апрель.
Данные показывают экспрессию генов, участвующих в транспорте питательных веществ: (1) сравнивая их поведение в феврале и апреле (Рисунок 3), (2) оценивая различия в зависимости от ткани (листья и корни) (Рисунок 4) и (3) учитывая их эффекты в зависимости от субстратов для выращивания (Рисунок 5).
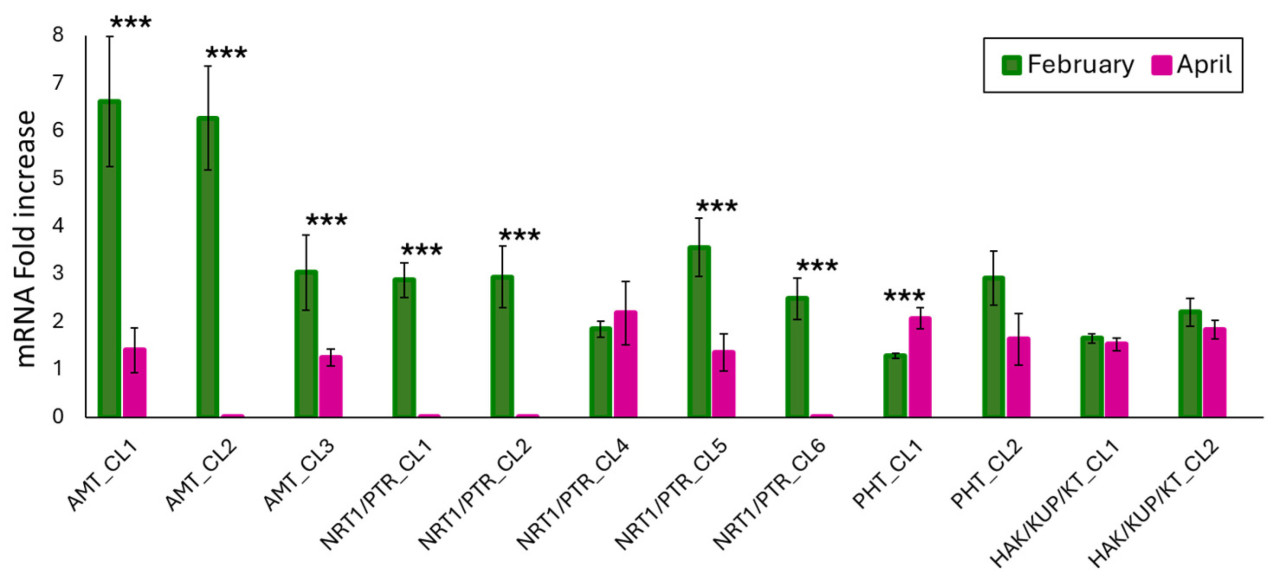
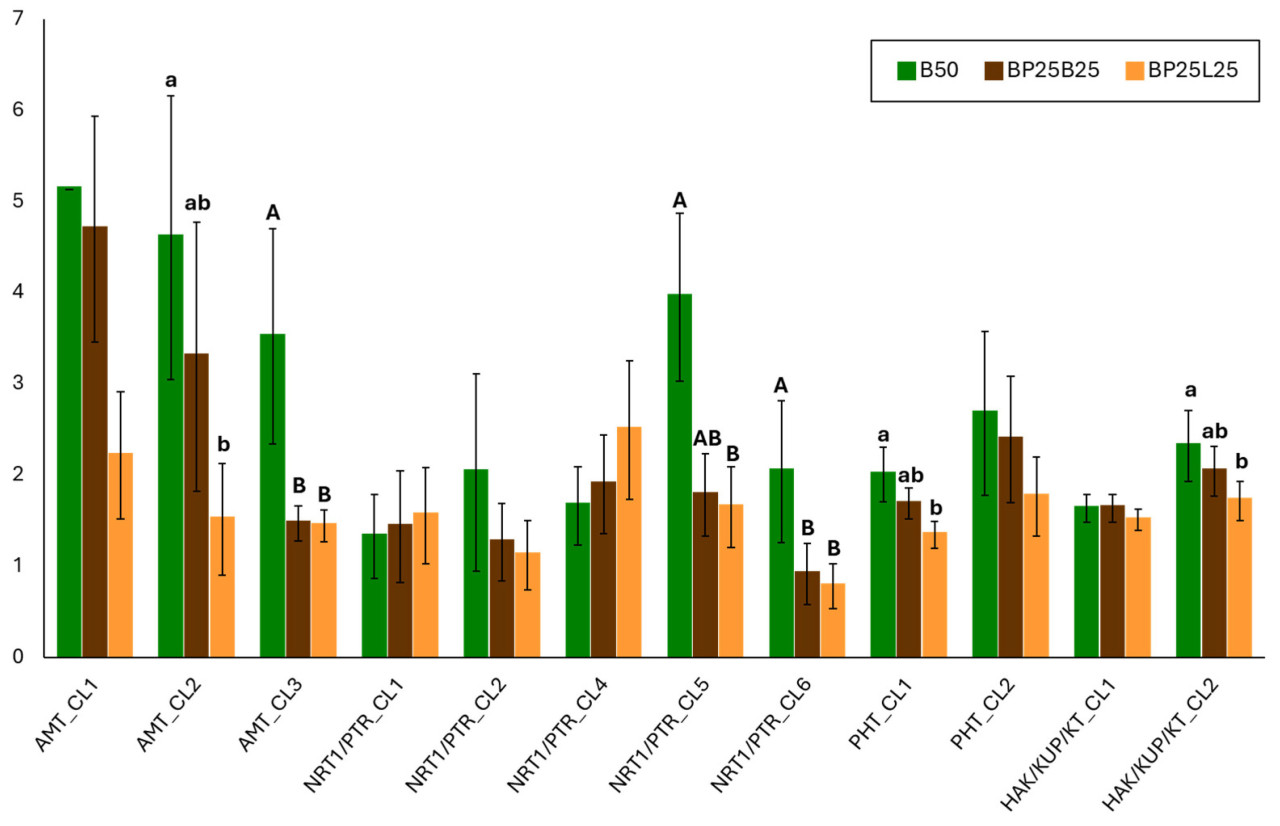
В феврале все анализируемые гены, включенные в ранее описанные кластеры, экспрессировались как в листьях, так и в корнях. Уровень многих транскриптов был статистически разным по сравнению с апрелем, за исключением генов NRT1/PTR_CL4, PHT_CL2, HAK/KUP/KT_CL2 и HAK/KUP/KT_CL1. В апреле, напротив, мы не наблюдали экспрессии AMT_CL2, NRT1/PTR_CL1, NRT1/PTR_CL2 и NRT1/PTR_CL6 в обеих тканях (Рисунок 3).
Средние значения экспрессии генов для каждого отбора проб согласуются, по крайней мере, с тем, что уже известно о поглощении азота во многих видах растений, включая цитрусовые.
Фактически, поглощение N варьируется во время жизненного цикла растений, обычно снижаясь во время цветения [38]. Более высокая экспрессия почти всех генов, участвующих в поглощении нитрата в феврале, может быть обусловлена меньшей доступностью этого питательного вещества. Кроме того, для лучшего понимания этой динамики важно учитывать роли нитрат- и нитрит- редуктазы, которые также влияют на присутствие калия [39].
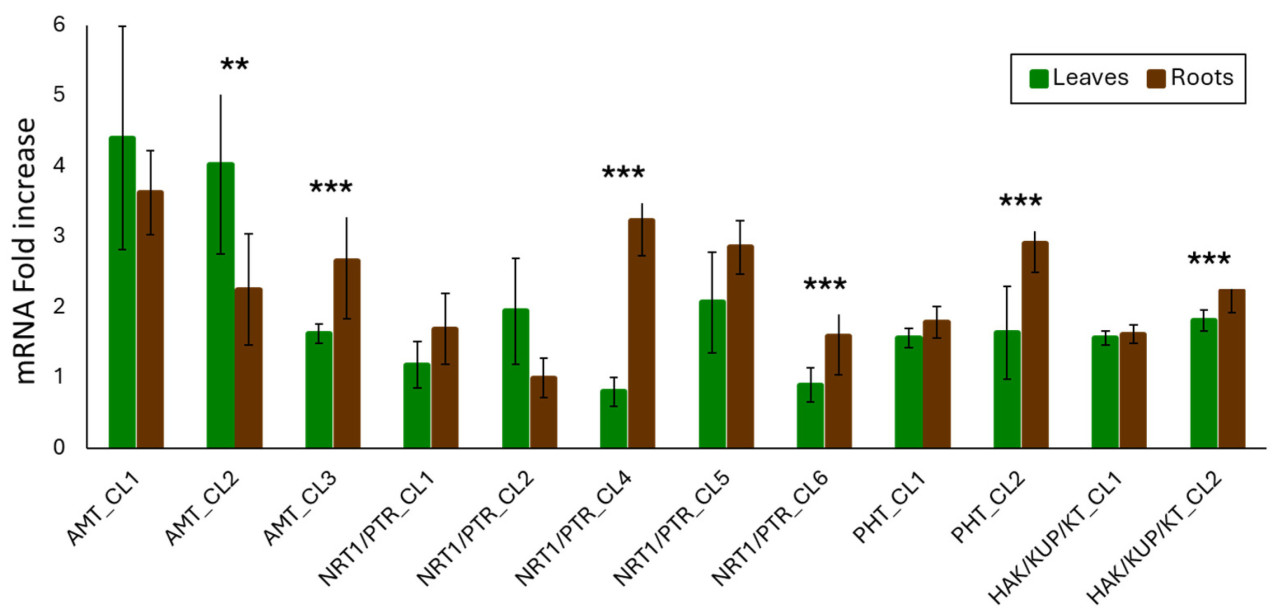
Когда мы сосредоточились на двух тканях, мы наблюдали, что экспрессия генов, кодирующих AMT_CL3, NRT1/PTR_CL4, NRT1/PTR_CL6, PHT_CL2 и HAK/KUP/KT_CL2, была статистически значимой в корнях по сравнению с листьями (Рисунок 4). Напротив, гены, кодирующие AMT_CL2 и NRT1/PTR_CL2, были высоко экспрессированы в листьях по сравнению с корнями (Рисунок 4). Это замечание также было подчеркнуто Ванг и соавторами [10], которые выделили важность корней в физиологическом поведении растения. Это могло быть обусловлено модификацией и доступностью аммония в почве по сравнению с листьями. Однако эти результаты могли быть из-за влияния компонента биоугля на субстрат.
Когда мы оценивали экспрессию генов в тестируемых субстратах для выращивания, наиболее очевидные различия наблюдались для генов, участвующих в транспорте аммония (АМТ_С1_1, AMT_CL2 и AMT_CL3) между В50 и BP25L25 (Рисунок 5). Действительно, наивысшее количество транскриптов для субстрата В50 согласуется с результатами предыдущих исследований, полученных с биоугля [40], где сообщалось о индуцированных изменениях в почве и их влиянии на рост растений с учетом доступности азота. Даже в цикле Р и К присутствие биоугля определяет увеличение этих двух питательных веществ в почве и усиление концентрации К в растительной ткани (корнях и листьях) [40,41,42]. Более того, можно предположить, что содержание NH4+ в субстратах для выращивания увеличилось из-за снижения потери питательных веществ выщелачиванием [43,44]. Этот результат подтверждает роль биоугля в улучшении поглощения NH4+ растениями; этот компонент субстрата для выращивания снижал бы испарение аммиака, важного атмосферного загрязнителя [45]. Все это могло бы улучшить эффективность питания в питомнике путем снижения использования N-удобрений и объема горшка.
Результаты, похожие на те, что выявлены для генов, участвующих в транспорте аммония, наблюдались для генов NRT1/PTR_CL5, NRT1/PTR_CL6 (нитрат), РНТ_С1_1 (фосфор) и HAK/KUP/KT_CL2 (калий). Вместо этого ген NRT1/PTR_CL4 показал повышенную экспрессию в BP25L25 по сравнению с субстратом В50, в то время как ген PHT_CL2 демонстрировал более высокую экспрессию в ВР25В25 по сравнению с BP25L25 (Рисунок 5).
Основными источниками азота для растений являются NH4+ и NO3’. В почве NO3’ обычно присутствует в более высоких концентрациях и также более мобильна и доступна растениям, чем NH4+ [12]. Растения развили различные механизмы приобретения азота, которые включают системы транспорта с разными affinities для нитрата и аммония (HATS и LATS) [18,20,46].
Что касается генов, участвующих в транспорте NH4+, генетические последовательности С. sinensis из кластера 1 были ортологичными к Pt неохарактеризованному белку. Кластер 2 включал члены С. sinensis ортологов члена семейства АМТ2 At и членов С. sinensis ортологов семейства АМТ2 и АМТЗ Pt. Кластер 3 включал членов семейства АМТ1 из С. sinensis, Poncirus trifoliata и Carrizo citrange (Таблица SI, Рисунок S1A).
Что касается NH4+, почти все изученные гены демонстрировали высокий уровень экспрессии в феврале, как в листьях, так и в корнях (Рисунок 6А). В апреле ген AMT_CL3 экспрессировался в обеих тканях; напротив, АМТ_С1_1 экспрессировался только в корнях, в то время как не было выявлено экспрессии для AMT_CL2 (Рисунок 6В). Конкретно, наибольшая экспрессия среди субстратов для выращивания была найдена в тех, содержащих 50% (В50) и 25% (ВР25В25) биоугля, что было статистически значимым только в корнях (Рисунок 6А). Недавно Ферлито и соавторы [1] наблюдали снижение роста как для воздушной, так и для подземной частей саженцев, в основном с субстратом, содержащим высокий процент биоугля (В50).
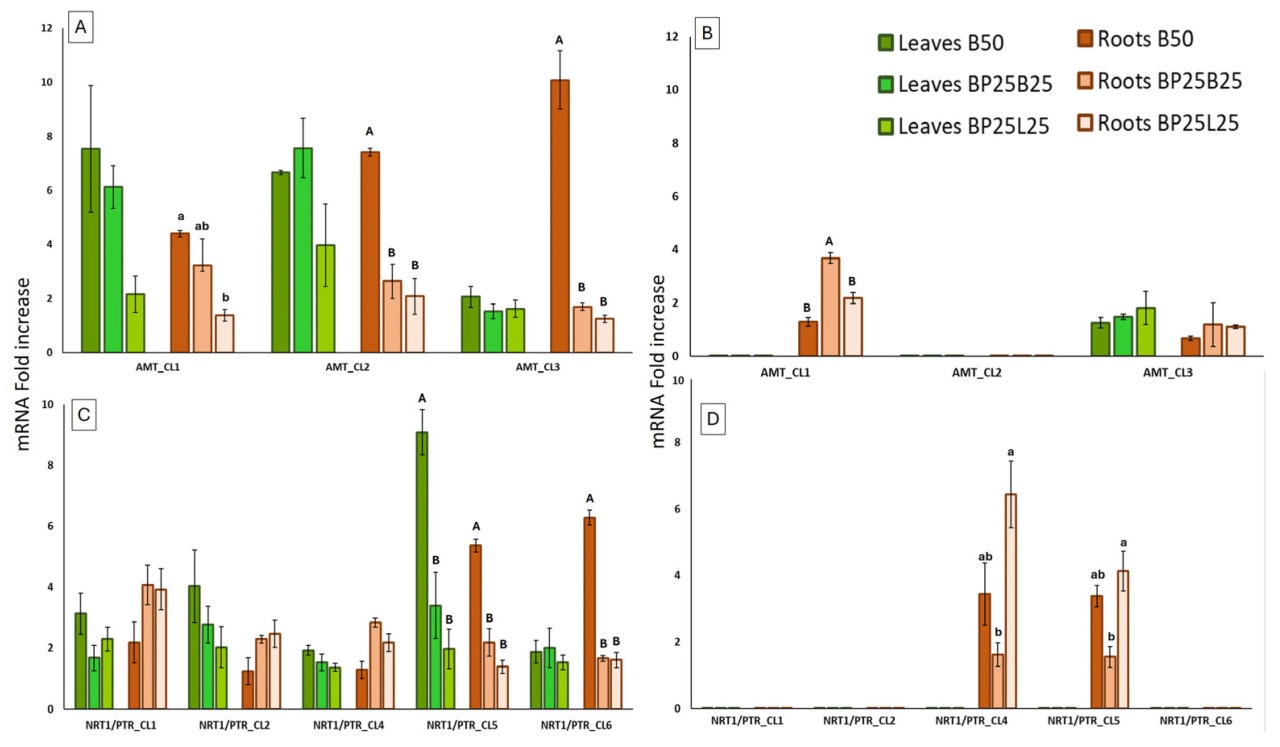
Последовательности, выбранные для нитрата азота, были найдены ортологичными к семействам NRT и NRT/PTR как в At, так и в Pt. Кластеры 1-6 в основном были ортологичными к членам семейства NRT и NRT/PTR 8, 5, 4, 6, 2 и 1, соответственно (Таблица S1, Рисунок S1B). Наблюдалось разное поведение для изученных генов между зимой и весной. В феврале транскрипты получались как в корнях, так и в листьях, в то время как в апреле гены NRT1/PTR_CL1, NRT1/PTR_CL2 и NRT1/PTR_CL6 не показывали экспрессии, и гены NRT1/PTR_CL5 и NRT1/PTR_CL4 экспрессировались только в корнях (Рисунок 6C,D). В феврале уровень транскриптов NO3’ в среднем был ниже, чем у транскриптов NH4+ (Рисунок 6А,С); более того, для каждого гена и ткани, исследованных, всегда в NO3’, значимые различия между субстратами для выращивания были похожи на те, что наблюдались для NH4+. В феврале ген NRT1/PTR_CL5 показал статистически значимый уровень экспрессии в субстрате В50 по сравнению с другими двумя субстратами для выращивания как в листьях, так и в корнях. Напротив, ген NRT1/PTR_CL6 производил уровень транскрипта, статистически значимый среди изученных субстратов, который наблюдался только в корнях (Рисунок 60). В отборе проб апреля только NRT1/PTR_CL4 и NRT1/PTR_CL5 были аналогично экспрессированы, исключительно в корнях для всех субстратов для выращивания; более того, экспрессия, найденная в В50, была большей по сравнению с наблюдаемой в ВР25В25 (Рисунок 6D).
Зимой низкие температуры ответственны за снижение ферментативной активности нитрат- и нитрит-редуктазы, снижая доступность нитрического азота [47]. Этот дефицит может вызвать увеличение экспрессии генов транспортеров NH4+ и N03’. Мы предполагаем, что саженцы, выращенные в субстратах, содержащих биоугля, благодаря способности этого добавки мобилизовать питательные вещества, могут способствовать более ранней активации генов и накапливать более высокие уровни азота, чем контроль в феврале. Отсутствие транскриптов других анализируемых генов в апреле могло быть ожидаемым, потому что в этот сезон растение уже должно было накопить количество азота, необходимое для его развития (Рисунок 6D).
Фосфор — один из основных ограничивающих факторов для роста растений. В почве этот питательный элемент обычно присутствует в формах, недоступных растениям, и имеет очень низкую растворимость [48]. Только неорганическая форма (Pi) действительно доступна для поглощения корнями. Более того, его содержание в почвах относительно низкое, с только маргинальной частью, присутствующей в растворе, такой как ион фосфата (Н2Р04’; НР042’). Это количество очень низкое по сравнению с оптимальными потребностями растений [48].
Подобно азоту, фосфор имеет две разные системы транспорта с разными affinities для Pi [49]. Транспортер высокой affinity активируется, когда Pi в почве находится в низком диапазоне в терминах микромолей (цМ) [50], в то время как система транспорта низкой affinity запускается, когда диапазон концентрации Pi в почве находится в порядке миллимолей (тМ) [51]. Кроме того, некоторые авторы предполагают, что система транспорта низкой affinity конститутивно активна в растениях [52].
Данные по экспрессии генов фосфора показаны на Рисунке 7.
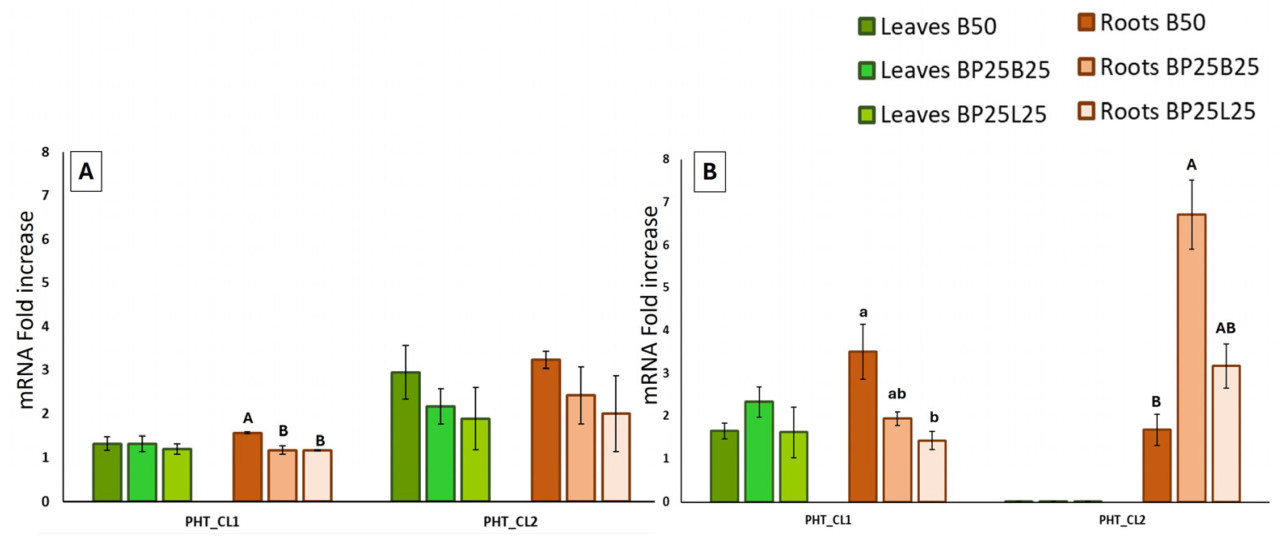
Не сообщалось об экспрессии для гена РНО1_С1_3. Не наблюдалось значимых различий среди субстратов для выращивания в обеих датах отбора проб в листьях для гена РНТ_С1_1. Напротив, статистически значимое различие было выявлено в корнях между В50 и другими двумя субстратами для выращивания в феврале и между В50 и BP25L25 в апреле (Рисунок 7А,В). Ген РНТ_С1_1 включен среди последовательностей кластера 1, среди которых ортолог At и Pt — РНТ 1-4, описанный в основном экспрессируемым в корнях [51] (Таблица 81, Рисунок S1C).
В отличие от гена РНТ_С1_1, транскрипты PHT_CL2 производились в обеих тканях только в феврале и только в корнях в апреле (Рисунок 7А,В). Уровень экспрессии PHT_CL2 был, в общем, выше, чем у РНТ_С1_1. В апреле обилие транскриптов в корнях достигло пика в субстрате для выращивания ВР25В25, показывая статистически значимое различие по сравнению с В50.
Наша работа выделила определенную экспрессию гена РНТ_С1_1 как в анализируемых тканях, так и в данных отбора проб (февраль и апрель). Отсутствие различий в обеих датах отбора проб на уровне листьев предполагает, что активация генов в этом кластере начинается в корневой системе, возможно, потому что отбор проб и анализ проводились слишком рано к пику вегетативного роста листьев.
Экспрессия гена РНТ_С1_1 могла быть объяснена тем, что кластер 1 включает семейство генов РНТ1 (1;1 и 1;4). Как описано для генов AtPHT1;1 и AtPHT1;4 в Arabidopsis [53,54], мы предполагаем, что эти могли играть главную роль в поглощении фосфата, особенно поскольку многие члены этого семейства обычно модулируются низким содержанием Pi [55,56]. Это согласуется с Деб и соавторами [57], которые сообщили, как биоугля увеличивал производство органической кислоты, solubilizing Pi, в почвах, бедных питательными веществами, таких как вулканическая почва, использованная в этом исследовании [58]. Предыдущие исследования выделили, что в почвах, дефицитных Р, биоугля активно влияет на рост и урожайность некоторых видов путем увеличения микробной биомассы [59]. Поэтому мы можем предположить, что наши выводы, связанные с экспрессией генов, ответственных за поглощение и транспорт Р, также влияют биоугля.
Кроме того, кластер 1 включает членов семейства РНТ2, которые обычно имеют транспортеры Pi низкой affinity. Семейство генов РНТ2 преимущественно экспрессируется в зеленой ткани [60,61], хотя Гуо и соавторы [62] описали присутствие транскриптов в корнях, как показано на Рисунке 7А,В. Слабое несоответствие в транскриптах корней, выделенное в двух датах отбора проб (высокое значение в апреле по сравнению с февралем; Рисунок 7А,В), могло быть обусловлено поддержанием Р в его двух формах, Pi и РО, после поглощения. Когда фосфат транслокируется в ксилему, только фракция Pi инкорпорируется, в то время как фракция РО высвобождается обратно как Pi, в основном ограниченная ксилемой корней, из которой она перемещается к вновь образованным органам (листьям) весной [12]. В феврале Pi и его эстерифицированная фракция (такая как РО) еще не мобилизованы к листьям, что могло бы оправдать экспрессию гена в этой ткани (Рисунок 7А). По мере того, как РО начинает мобилизоваться к листьям, корни становятся дефицитными в Р; это могло бы объяснить экспрессию гена в апреле в корнях, наблюдаемую для обоих генов РНТ_С1_1 и РНТ_С1_1 (Рисунок 7А,В).
Калий необходим для всех растительных активностей, требующих транспорта воды, включая открытие и закрытие устьиц [63]. Кроме того, калий обеспечивает стабильность и качество растения и регулирует многие другие процессы [64,65]. В почве калий обычно делится на четыре категории: водорастворимый, обменный, медленно доступный и структурный калий [66]. Последний составляет наибольшее количество в окружающей среде, хотя его трудно использовать напрямую растениями [67].
Транспортеры калия могут быть приписаны четырем семействам мембранных белков: К+ uptake permeates КТ (К+ transporter)/HAK (high-affinity K+)/KUP (К+ uptake) family, K+ transporter Trk/Ktr/HKT family, KEA (K+ efflux antiporter) family и CHX (cation/hydrogen exchanger) family [68]. Семейство HAK/KUP/KT — самое большое семейство транспортеров К+ в растениях. Оно содержит носители К+ высокой affinity и в основном участвует в росте и развитии растений [69].
Из анализа BLAST мы нашли, что последовательности С. sinensis ортологичны членам семейства HAK/KUP/KT и соответствуют HAK/KUP/KT 11, 3, 2 и 6 At и Pt, которые участвуют в развитии и морфологии корней и листьев [68,70,71]. Кроме того, шесть последовательностей С. sinensis были найдены как "Probable potassium transporter 11-like isoform XI" в At.
Экспрессия гена HAK/KUP/KT_CL1 не показала никаких статистически значимых различий ни между анализируемыми тканями, ни между периодами отбора проб (февраль и апрель) (Рисунок 8А,В).
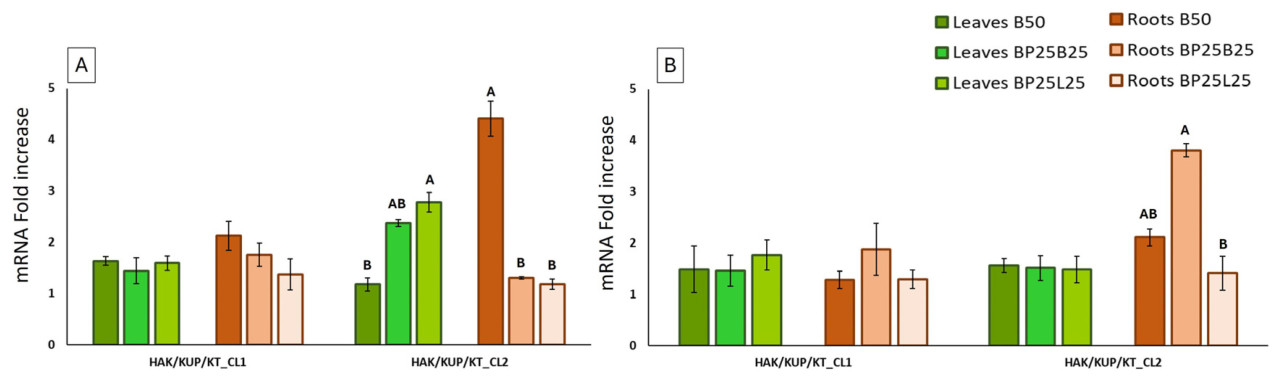
Напротив, экспрессия HAK/KUP/KT_CL2 показала статистически значимые различия между корнями (Рисунок 8А,В) в феврале и апреле. В феврале корни показали более высокие значения с субстратом для выращивания, содержащим 50% биоугля (В50), в то время как в апреле более высокая экспрессия была выявлена в субстрате ВР25В25. В феврале листья показали статистически значимые различия между В50 и другими двумя субстратами для выращивания (Рисунок 8А).
Можно предположить, что поскольку корневая система начинает свой биологический и морфологический рост на месяц раньше, чем воздушная часть растения, рост в субстрате В50 позволяет для выгодного поглощения К, которое способствует значительному росту корневой системы уже в феврале. Результаты апреля согласуются с предыдущими исследованиями морфологии корневой системы с использованием тех же композиций субстратов, оцениваемых в этой работе [1], которые показали значительное снижение корневой системы растений, выращенных в субстрате ВР25В25 по сравнению с другими субстратами (Рисунок 9). Биоугля кажется эффективным материалом, который может улучшить доступность калия в почве [26]; поэтому субстрат с большим количеством биоугля (850) облегчает большее поглощение К по сравнению с ВР25В25. В апреле экспрессия гена HAK/KUP/KT_CL2 была менее выраженной в субстрате В50 в корнях, потому что растения уже поглотили необходимое количество К. Напротив, растения, выращенные в субстрате ВР25В25, который облегчает поглощение в меньшей степени, показали повышенную экспрессию в апреле, вероятно, потому что им все еще нужен питательный элемент, поглощенный в относительном количестве в феврале [72,73]. Можно спекулировать, что в апреле растение, выращенное в ВР25В25, пытается поглотить как можно больше калия, чтобы восстановить правильную морфологию корневой системы, как уже описано; некоторые члены семейства генов HAK/KUP/KT влияют не только на приобретение и транспорт К+, но также на морфологию и удлинение корней и побегов [68,74,75].

4. Статистический анализ
Для оценки эффекта всех анализов, выполненных в этой работе, и вывода предполагаемой связи между ними проводился РСА, который показал, что первые две компоненты учитывали 50.1% общей объясненной дисперсии (РС1 - 33.2%; РС2 - 16.9%) (Таблица 5). Субстраты для выращивания В50, ВР25В25 и BP25L25 были четко различимы в многомерном пространстве (Рисунок 10).
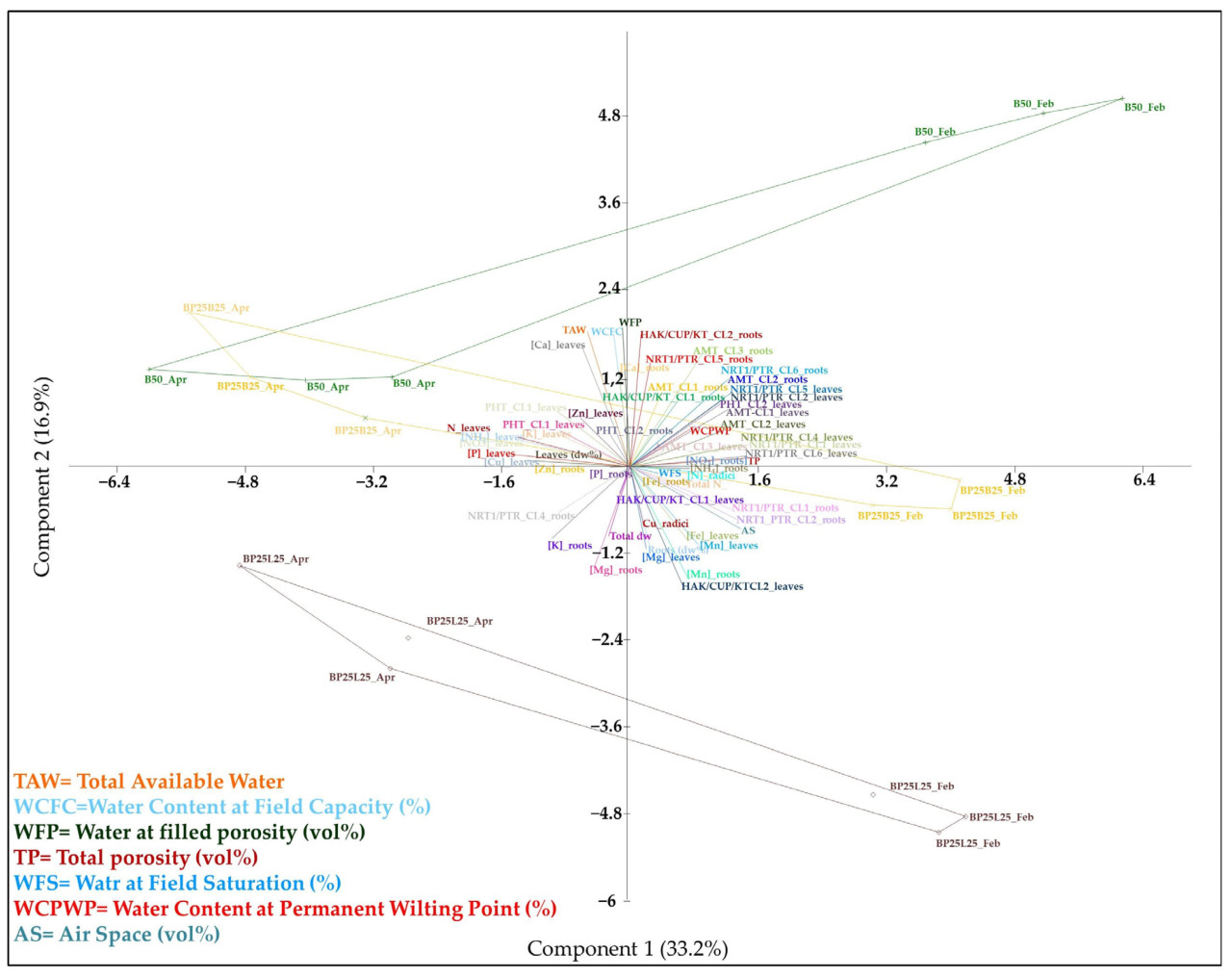
Транскрипты NRT1/PTR_CL1, NRT1/PTR_CL2, NRT1/PTR_CL4, NRT1/PTR_CL5, NRT1/PTR_CL6, AMT_CL1, AMT_CL2 и PHT_CL2 в листьях, NRT1/PTR_CL1, NRT1/PTR_CL2, NRT1/PTR_CL6 и AMT_CL2 в корнях, наряду с общей порозностью и воздушным пространством субстратов для выращивания, показали сильную положительную корреляцию (>70.0%) с РС1 (Таблица 82). Все образцы, собранные в феврале, расположенные в положительной части РС1, характеризовались высокими значениями для этих параметров. Напротив, Р, N, Си, NO3’, NH4+ и N_minerale_tot_mg/Kg в листьях, наряду с Zn в корнях, показали сильную отрицательную корреляцию (>-70.0%), представляя образцы, собранные в апреле, расположенные в отрицательной секции РС1 (таблица S2,рисунок 10).
Емкость катионного обмена (CIC), доступная водная емкость AWC и порозность, заполненная водой (WFS), а также HAK/KUP/KT_CL2 в корнях и Са в листьях, показали сильную положительную корреляцию с РС2, которая представляла субстрат В50 для выращивания как в феврале, так и в апреле, наряду с ВР25В25, собранным в апреле, все расположенные в положительной части РС2. Напротив, сильная отрицательная корреляция с РС2 была отмечена для HAK/KUP/KT_CL2 в листьях, которая в основном характеризовала субстрат BP25L25 для выращивания, расположенный в отрицательной части РС2 (Таблица 82, Рисунок 10).
5. Выводы
На основе физико-химических параметров и роста саженцев мы заключаем, что торф может быть частично заменен биоугля из древесины хвойных пород в общем количестве 25-50%. Новый субстрат для выращивания для цитрусовых питомников позволяет темпы роста быть сравнимыми с теми, что зарегистрированы для растений, выращенных в традиционных торфяных смесях.
Далее, мы наблюдали, что сравнение между разными данными отбора проб (февраль и апрель) и тканями выделило, что все анализируемые гены были высоко экспрессированы в листьях и корнях в феврале. Напротив, в апреле некоторые гены не экспрессировались в обеих тканях. В общем, уровни транскрипции были большими в корнях, чем в листьях. Кроме того, сравнивая растения, выращенные на разных субстратах, очевидно, что субстрат В50 для выращивания имеет наивысшую экспрессию.
Более высокая экспрессия генов транспортеров аммиачного и нитрического азота предполагает, что субстраты, содержащие биоугля, компенсируют дефицит азота во время холодных периодов благодаря более ранней активации генов в феврале.
Дополнительные материалы
Следующая дополнительная информация может быть загружена по адресу https://www.mdpi.com/article/10.3390/agriculture15010113/s1, Рисунок 81: Филогенетический анализ на основе множественного выравнивания последовательностей. Эволюционная история была выведена с использованием метода Maximum Likelihood и модели Tamura-Nei (56). Анализы проводились с использованием MEGA X (24). Для каждого полученного дерева (одно для каждого макроэлемента) было возможно разделить анализируемые последовательности на кластеры. Вид показан рядом с его номером доступа. Дерево нарисовано в масштабе, с длинами ветвей, измеряемыми в количестве замен на сайт. Показано дерево с наивысшей логарифмической вероятностью. (А) Двадцать три последовательности транспортеров аммония (NH4+); (В) Сорок последовательностей транспортеров нитрата (NO3’); (С) Двадцать шесть последовательностей транспортеров фосфора (Р); (D) Двадцать последовательностей транспортеров калия (К). Таблица 81: Название питательного элемента, номера доступа, кластер, Семейство генов, Ортологи из OrthoDB, функция, ID С. sinensis DHSO v3.0 геном CDS всех генных последовательностей, использованных в этом исследовании. Таблица 82: Загрузки анализа главных компонент (РСА) значений экспрессии различных анализируемых генов и физических и химических характеристик трех субстратов для выращивания, каждый содержащий 50% песчаной вулканической почвы и оставшиеся 50%: биоугля 50% (В50); черный торф 25% + биоугля 25% (ВР25В25); черный торф 25% + лапилус 25% (BP25L25), физические и химические свойства, распределение биомассы саженцев Carrizo citrange и экспрессия генов, участвующих в транспорте азота, фосфора и калия в листьях и корнях. Данные были собраны в феврале (зима) и апреле (весна).
Вклад авторов
Концептуализация, надзор, написание — черновик, П.К.; исследование, написание — рецензия и редактирование, кураторство данных, М.П.Р.; формальный анализ, написание — рецензия и редактирование, М.А.; формальный анализ, написание — рецензия и редактирование, Б.Т.; ресурсы, финансирование, написание — рецензия и редактирование, Дж.Ф.; Исследование, написание — рецензия и редактирование, Дж.Л.К.; Написание — Рецензия и Редактирование, К.Л.; Написание — Рецензия и Редактирование М.К.; визуализация, А.К.; Концептуализация, надзор, написание — черновик, Ф.Ф. Все авторы прочитали и согласились с опубликованной версией рукописи.
Финансирование
Это исследование не получило внешнего финансирования.
Заявление о этике институционального совета
Не применимо.
Заявление о доступности данных
Авторы заявляют о своей готовности поделиться существующими наборами данных или сырыми данными по запросу.
Конфликты интересов
Авторы заявляют об отсутствии конфликтов интересов.
Ссылки
1. Ferlito, F.; Torrisi, B.; Allegra, M.; Stagno, F.; Caruso, P.; Fascella, G. Evaluation of conifer wood biochar as growing media component for citrus nursery. Appl. Sci. 2020, 10, 1618. [Google Scholar] [CrossRef]
2. Torrisi, B.; Allegra, M.; Amenta, M.; Gentile, F.; Rapisarda, P.; Fabroni, S.; Ferlito, F. Physico-chemical and multielemental traits of anaerobic digestate from Mediterranean agro-industrial wastes and assessment as fertiliser for citrus nurseries. Waste Manag. 2021, 131, 201–213. [Google Scholar] [CrossRef] [PubMed]
3. Bridgwater, A.V.; Meier, D.; Radlein, D. An overview of fast pyrolysis of biomass. Org. Geochem. 1999, 30, 1479–1493. [Google Scholar] [CrossRef]
4. Fascella, G.; Mammano, M.M.; D’Angiolillo, F.; Rouphael, Y. Effects of conifers wood biochar as substrate component on ornamental performance, photosynthetic activity and mineral composition of potted Rosa rugosa. J. Hort. Sci. Biotech. 2018, 93, 519–528. [Google Scholar] [CrossRef]
5. Carrubba, A.; Lazzara, S.; Fascella, G.; Marceddu, R.; Napoli, E.; Sarno, M. Biochar enhances root development and aloin content of mature leaves in containerized Aloe arborescens Mill. S. Afr. J. Bot. 2023, 163, 703–714. [Google Scholar] [CrossRef]
6. Kozioł, A.; Paliwoda, D.; Mikiciuk, G.; Benhadji, N. Biochar as a multi-action substance used to improve soil properties in horticultural and agricultural crops—A review. Agriculture 2024, 14, 2165. [Google Scholar] [CrossRef]
7. Fascella, G.; D’Angiolillo, F.; Ruberto, G.; Napoli, E. Agronomic performance, essential oils and hydrodistillation wastewaters of Lavandula angustifolia grown on biochar-based substrates. Ind. Crops Prod. 2020, 154, 112733. [Google Scholar] [CrossRef]
8. Ma, J.F.; Tsay, Y.F. Transport systems of mineral elements in plants: Transporters, regulation and utilization. Plant Cell Physiol. 2021, 62, 539–540. [Google Scholar] [CrossRef] [PubMed]
9. Yadav, B.; Jogawat, A.; Rahman, M.S.; Narayan, O.P. Secondary metabolites in the drought stress tolerance of crop plants: A review. Gene Rep. 2021, 23, 101040. [Google Scholar] [CrossRef]
10. Wang, P.; Yamaji, N.; Inoue, K.; Mochida, K.; Ma, J.F. Plastic transport systems of rice for mineral elements in response to diverse soil environmental changes. New Phytol. 2020, 226, 156–169. [Google Scholar] [CrossRef]
11. Wang, W.; Li, A.; Zhang, Z.; Chu, C. Post-translational modifications: Regulation of nitrogen utilization and signaling. Plant Cell Physiol. 2021, 62, 543–552. [Google Scholar] [CrossRef] [PubMed]
12. Hawkesford, M.J.; Cakmak, I.; Coskun, D.; De Kok, L.J.; Lambers, H.; Schjoerring, J.K.; White, P.J. Functions of macronutrients. In Marschner’s Mineral Nutrition of Plants; Rengel, Z., Cakmak, I., White, P.J., Eds.; Academic Press; Elsevier Ltd.: Amsterdam, The Netherlands, 2023; pp. 201–281. [Google Scholar] [CrossRef]
13. Sasaki, A.; Yamaji, N.; Ma, J.F. Transporters involved in mineral nutrient uptake in rice. J. Exp. Bot. 2016, 67, 3645–3653. [Google Scholar] [CrossRef] [PubMed]
14. Couturier, J.; Montanini, B.; Martin, F.; Brun, A.; Blaudez, D.; Chalot, M. The expanded family of ammonium transporters in the perennial poplar plant. New Phytol. 2007, 174, 137–150. [Google Scholar] [CrossRef] [PubMed]
15. Castro-Rodríguez, V.; Cañas, R.A.; de la Torre, F.N.; Pascual, M.B.; Avila, C.; Cánovas, F.M. Molecular fundamentals of nitrogen uptake and transport in trees. J. Exp. Bot. 2017, 68, 2489–2500. [Google Scholar] [CrossRef]
16. Lea-Cox, J.D.; Syvertsen, J.P. Nitrogen uptake by Citrus leaves. J. Amer. Soc. Hort. Sci. 1995, 120, 505–509. [Google Scholar] [CrossRef]
17. Camañes, G.; Cerezo, M.; Primo-Millo, E.; Gojon, A.; García-Agustín, P. Ammonium transport and CitAMT1 expression are regulated by light and sucrose in Citrus plants. J. Exp. Bot. 2007, 58, 2811–2825. [Google Scholar] [CrossRef]
18. Camañes, G.; Cerezo, M.; Primo-Millo, E.; Gojon, A.; García-Agustín, P. Ammonium transport and CitAMT1 expression are regulated by N in Citrus plants. Planta 2009, 229, 331–342. [Google Scholar] [CrossRef]
19. Cerezo, M.; Tillard, P.; Gojon, A.; Primo-Millo, E.; Garcia-Agustin, P. Characterization and regulation of ammonium transport systems in Citrus plants. Planta 2001, 214, 97–105. [Google Scholar] [CrossRef]
20. Cerezo, M.; Camañes, G.; Flors, V.; Primo-Millo, E.; García-Agustín, P. Regulation of nitrate transport in Citrus rootstocks depending of nitrogen availability. Plant Signal Behav. 2007, 2, 337–342. [Google Scholar] [CrossRef][Green Version]
21. Wickert, E.; Marcondes, J.; Lemos, M.V.; Lemos, E.G.M. Nitrogen assimilation in Citrus based on CitEST data mining. Genet. Mol. Biol. 2007, 30, 810–818. [Google Scholar] [CrossRef]
22. Sorgonà, A.; Cacco, G.; di Dio, L.; Schmidt, W.; Perry, P.J.; Abenavoli, M.R. Spatial and temporal patterns of net nitrate uptake regulation and kinetics along the tap root of Citrus aurantium. Acta Physiol Plant. 2010, 32, 683–693. [Google Scholar] [CrossRef]
23. Shu, B.; Xia, R.X.; Wang, P. Differential regulation of Pht1 phosphate transporters from trifoliate orange (Poncirus trifoliata L. Raf) seedlings. Sci. Hortic. 2012, 146, 115–123. [Google Scholar] [CrossRef]
24. Caballero, F.; García-Sánchez, F.; Gimeno, V.; Syvertsen, J.P.; Martínez, V.; Rubio, F. High-affinity potassium uptake in seedlings of two citrus rootstocks Carrizo citrange (Citrus sinensis [L.] Osb. × Poncirus trifoliata [L.] Raf.) and Cleopatra mandarin (Citrus reshni Hort. ex Tanaka). AJCS 2013, 7, 538–542. [Google Scholar]
25. Meng, X.; Chen, W.W.; Wang, Y.Y.; Huang, Z.R.; Ye, X.; Chen, L.S. Effects of phosphorus deficiency on the absorption of mineral nutrients, photosynthetic system performance and antioxidant metabolism in Citrus grandis. PLoS ONE 2021, 16, e0246944. [Google Scholar] [CrossRef]
26. Zhang, M.; Riaz, M.; Xia, H.; Li, Y.; Wang, X.; Jiang, C. Four-year biochar study: Positive response of acidic soil microenvironment and citrus growth to biochar under potassium deficiency conditions. Sci. Total Environ. 2022, 813, 152515. [Google Scholar] [CrossRef] [PubMed]
27. Vanella, D.; Ferlito, F.; Torrisi, B.; Giuffrida, A.; Pappapardo, S.; Saitta, D.; Longo-Minnolo, G.; Consoli, S. Long-term monitoring of deficit irrigation regimes on citrus orchards in Sicily. J. Agric. Eng. 2021, 52, 1193. [Google Scholar] [CrossRef]
28. Bremner, J.M.; Mulvaney, C.S. Nitrogen-total. In Methods of Soil Analysis, Part 2, Chemical and Microbiological Properties, 2nd ed.; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; John Wiley & Sons: Madison, WI, USA, 1982; Volume 9, pp. 595–624. [Google Scholar]
29. Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: A Better Web Interface. Nucleic Acids Res. 2008, 36, W5–W9. [Google Scholar] [CrossRef]
30. Xu, Q.; Chen, L.L.; Ruan, X.; Chen, D.; Zhu, A.; Chen, C.; Bertrand, D.; Jiao, W.B.; Hao, B.H.; Lyon, M.P. The Draft Genome of Sweet Orange (Citrus sinensis). Nat. Genet. 2012, 45, 59–68. [Google Scholar] [CrossRef] [PubMed]
31. Sievers, F.; Wilm, A.; Dineen, D.G.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
32. Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000; p. 352. [Google Scholar]
33. Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
34. Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar]
35. Mahe, A.; Grisvard, J.; Dron, M. Fungal-and plant-specific gene markers to follow the bean anthracnose infection process and normalize a bean chitinase mRNA induction. Mol. Plant Microbe Interact. 1992, 5, 242–248. [Google Scholar] [CrossRef]
36. Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
37. Taylor, H.M.; Ratliff, L.F. Root elongation rates of cotton and peanuts as a function of soil strength and soil water content. Soil Sci. 1969, 108, 113–119. [Google Scholar] [CrossRef]
38. Malagoli, P.; Lainé, P.; le Deunff, E.; Rossato, L.; Ney, B.; Ourry, A. Modelling nitrogen uptake in oilseed rape cv. Capitol during a growth cycle using influx kinetics of root nitrate transport systems and field experimental data. Plant Physiol. 2004, 134, 388–400. [Google Scholar] [CrossRef]
39. Xu, X.; Du, X.; Wang, F.; Sha, J.; Chen, Q.; Tian, G.; Zhu, Z.; Ge, S.; Jiang, Y. Effects of potassium levels on plant growth, accumulatio and distribution of carbon, and nitrate metabolism in apple dwarf rootstock seedlings. Front. Plant Sci. 2020, 11, 904. [Google Scholar] [CrossRef] [PubMed]
40. Taghizadeh-Toosi, A.; Clough, T.J.; Sherlock, R.R.; Condron, L.M. Biochar adsorbed ammonia is bioavailable. Plant Soil 2012, 350, 57–69. [Google Scholar] [CrossRef]
41. Dempster, N.; Gleeson, B.; Solaiman, M.; Jones, L.; Murphy, V. Decreased soil microbial biomass and nitrogen mineralisation with eucalyptus biochar addition to a coarse textured soil. Plant Soil 2012, 354, 311–324. [Google Scholar] [CrossRef]
42. Dempster, N.; Jones, L.; Murphy, V. Organic nitrogen mineralisation in two contrasting agro-ecosystems is unchanged by biochar addition. Soil Biol. Biochem. 2012, 48, 47–50. [Google Scholar] [CrossRef]
43. Laird, D.; Fleming, P.; Wang, B.; Horton, R.; Karlen, D. Biochar impact on nutrient leaching from a Midwestern agricultural soil. Geoderma 2010, 158, 436–442. [Google Scholar] [CrossRef]
44. Biederman, L.A.; Harpole, W.S. Biochar and its effects on plant productivity and nutrient cycling: A meta-analysis. GCB Bioenergy 2013, 5, 202–214. [Google Scholar] [CrossRef]
45. Beusen, A.H.W.; Bouwman, A.F.; Heuberger, P.S.C.; van Drecht, G.; van der Hoek, K.W. Bottom-up uncertainty estimates of global ammonia emissions from global agricultural production systems. Atmos. Env. 2008, 42, 6067–6077. [Google Scholar] [CrossRef]
46. Noguero, M.; Lacombe, B. Transporters involved in root nitrate uptake and sensing by Arabidopsis. Front. Plant Sci. 2016, 7, 1391. [Google Scholar] [CrossRef]
47. Zhou, J.; Ding, L.; Cui, C.; Lindeboom, R.E.F. High nitrite accumulation in hydrogenotrophic denitrification at low temperature: Transcriptional regulation and microbial community succession. Water Res. 2024, 263, 122144. [Google Scholar] [CrossRef]
48. Bieleski, R.L. Phosphate pools, phosphate transport and phosphate availability. Annu. Rev. Plant Physiol. 1973, 24, 225–252. [Google Scholar] [CrossRef]
49. Smith, F.W. The phosphate uptake mechanism. In Food Security in Nutrient-Stressed Environments: Exploiting Plants’ Genetic Capabilities. Developments in Plant and Soil Sciences; Adu-Gyamfi, J.J., Ed.; Springer: Dordrecht, Germany, 2002; Volume 245, pp. 235–244. [Google Scholar]
50. Poirier, Y.; Bucher, M. Phosphate transport and homeostasis in Arabidopsis. Arab. Book 2002, 1, e0024. [Google Scholar] [CrossRef]
51. Karthikeyan, A.S.; Varadarajan, D.K.; Mukatira, U.T.; D’Urzo, M.P.; Damsz, B.; Raghothama, K.G. Regulated expression of Arabidopsis phosphate transporters. Plant Physiol. 2002, 130, 221–233. [Google Scholar] [CrossRef] [PubMed]
52. Raghothama, K.G. Phosphate transport and signaling. Curr. Opin. Plant Biol. 2000, 3, 182–187. [Google Scholar] [CrossRef]
53. Shin, H.; Shin, H.S.; Dewbre, G.R.; Harrison, M.J. Phosphate transport in Arabidopsis: Pht1;1 and Pht1;4 play a major role in phosphate acquisition from both low- and high-phosphate environments. Plant J. 2004, 39, 629–642. [Google Scholar] [CrossRef]
54. Yang, J.; Zhou, J.; Zhou, H.J.; Wang, M.M.; Liu, M.M.; Ke, Y.Z. Global survey and expressions of the phosphate transporter gene families in Brassica napus and their roles in phosphorus response. Int. J. Mol. Sci. 2020, 21, 1752. [Google Scholar] [CrossRef] [PubMed]
55. Sun, S.; Gu, M.; Cao, Y.; Huang, X.; Zhang, X.; Ai, P. A constitutive expressed phosphate trans-porter OsPht11 modulates phosphate uptake and translocation in phosphate-replete rice. Plant Physiol. 2012, 159, 1571–1581. [Google Scholar] [CrossRef]
56. Remy, E.; Cabrito, T.R.; Batista, R.A.; Teixeira, M.C.; Sá-Correia, I.; Duque, P. The Pht19 and Pht18 transporters mediate inorganic phosphate acquisition by the Arabidopsis thaliana root during phosphorus starvation. New Phytol. 2012, 195, 356–371. [Google Scholar] [CrossRef] [PubMed]
57. Deb, D.; Kloft, M.; Lässig, J.; Walsh, S. Variable effects of biochar and P solubilizing microbes on crop productivity in different soil conditions. Agroecol. Sustain. Food Syst. 2016, 40, 145–168. [Google Scholar] [CrossRef]
58. Nicolosi, E.; Ferlito, F.; Allegra, M.; Cicala, A.; Trovato, F.; la Malfa, S. Influences of aspect and tillage on two winegrape cultivars on Mount Etna. N. Z. J. Crop Hort. Sci. 2016, 44, 83–102. [Google Scholar] [CrossRef][Green Version]
59. Masto, R.E.; Kumar, S.; Rout, T.K.; Sarkar, P.; George, J.; Ram, L.C. Biochar from water hyacinth (Eichornia crassipes) and its impact on soil biological activity. Catena 2013, 111, 64–71. [Google Scholar] [CrossRef]
60. Velasco, V.M.; Mansbridge, J.; Bremner, S.; Carruthers, K.; Summers, P.S.; Sung, W.W.; Champigny, M.J.; Weretilnyk, E.A. Acclimation of the crucifer Eutrema salsugineum to phosphate limitation is associated with constitutively high expression of phosphate-starvation genes. Plant Cell Environ. 2016, 39, 1818–1834. [Google Scholar] [CrossRef]
61. Versaw, W.K.; Harrison, M.J. A chloroplast phosphate transporter PHT2;1 influences allocation of phosphate within the plant and phosphate-starvation responses. Plant Cell 2002, 14, 1751–1766. [Google Scholar] [CrossRef] [PubMed]
62. Guo, C.; Zhao, X.; Liu, X.; Zhang, L.; Gu, J.; Li, X.; Lu, W.; Xiao, K. Function of wheat phosphate transporter gene TaPHT2;1 in Pi translocation and plant growth regulation under replete and limited Pi supply conditions. Planta 2013, 237, 1163–1178. [Google Scholar] [CrossRef] [PubMed]
63. Hsiao, T.C.; Läuchli, A. Role of potassium in plant–water relations. In Advances in Plant Nutrition, Volume 2; Tinker, B., Läuchli, A., Eds.; Praeger Scientific Publication: New York, NY, USA, 1986; pp. 281–312. [Google Scholar]
64. Almeida, H.J.; Pancelli, M.A.; Prado, R.M.; Cavalcante, V.S.; Cruz, F.J.R. Effect of potassium on nutritional status and productivity of peanuts in succession with sugarcane. J. Soil Sci. Plant Nutr. 2015, 15, 1–10. [Google Scholar] [CrossRef]
65. Cecilio, L.A.B.; Feltrim, A.L.; Cortez, J.W.M.; Gonsalves, M.V.; Pavani, L.C.; Barbosa, J.C. Nitrogen and potassium application by fertigation at different watermelon planting densities. J. Soil Sci. Plant Nutr. 2015, 15, 928–937. [Google Scholar] [CrossRef]
66. Oborn, I.; Andrist-Rangel, Y.; Askekaard, M.; Grant, C.A.; Watson, C.A.; Edwards, A.C. Critical aspects of potassium management in agricultural systems. Soil Use Manag. 2005, 21, 102–112. [Google Scholar] [CrossRef]
67. Simonsson, M.; Hillier, S.; Oborn, I. Changes in clay minerals and potassium fixation capacity as a result of release and fixation of potassium in long-term field experiments. Geoderma 2009, 151, 109–120. [Google Scholar] [CrossRef]
68. Yang, X.; Zhang, J.; Wu, A.; Wei, H.; Fu, X.; Tian, M.; Ma, L.; Lu, J.; Wang, H.; Yu, S. Genome-wide identification and expression pattern analysis of the HAK/KUP/KT gene family of cotton in fiber development and under stresses. Front. Genet. 2020, 11, 566469. [Google Scholar] [CrossRef]
69. Yang, T.; Lu, X.; Wang, Y.; Xie, Y.; Ma, Y.; Cheng, X.; Xia, E.; Wan, X.; Zhang, Z. HAK/KUP/KT family potassium transporter genes are involved in potassium deficiency and stress responses in tea plants (Camellia sinensis L.): Expression and functional analysis. BMC Genom. 2020, 21, 556. [Google Scholar] [CrossRef]
70. Grabov, A. Plant KT/KUP/HAK Potassium Transporters: Single Family—Multiple Functions. Ann. Bot. 2007, 99, 1035–1041. [Google Scholar] [CrossRef]
71. Ragel, P.; Raddatz, N.; Leidi, E.O.; Quintero, F.J.; Pardo, J.M. Regulation of K+ Nutrition in Plants. Front. Plant Sci. 2019, 10, 281. [Google Scholar] [CrossRef] [PubMed]
72. Al-Wabel, M.I.; Al-Omran, A.; El-Naggar, A.H.; Nadeem, M.; Usman, A.R. Pyrolysis temperature induced changes in characteristics and chemical composition of biochar produced from conocarpus wastes. Bioresour. Technol. 2013, 131, 374–379. [Google Scholar] [CrossRef]
73. Zhang, Z.H.; Hu, B.; Chu, C.C. Towards understanding the hierarchical nitrogen signalling network in plants. Curr. Opin. Plant Biol. 2020, 55, 60–65. [Google Scholar] [CrossRef] [PubMed]
74. Osakabe, Y.; Arinaga, N.; Umezawa, T.; Katsura, S.; Nagamachi, K.; Tanaka, H.; Ohiraki, H.; Yamada, K.; Seo, S.U.; Abo, M. Osmotic stress responses and plant growth controlled by potassium transporters in Arabidopsis. Plant Cell 2013, 25, 609–624. [Google Scholar] [CrossRef]
75. Elumalai, R.P.; Nagpal, P.; Reed, J.W. A mutation in the Arabidopsis KT2/KUP2 potassium transporter gene affects shoot cell expansion. Plant Cell 2002, 14, 119–131. [Google Scholar] [CrossRef]
Impact of Different Amounts of Biochar as Growth Media on Macronutrient Transport Systems of Carrizo Citrange Rootstocks and Related Expression Analysis
by Paola Caruso 1*,Maria Patrizia Russo 1, Maria Allegra 1, Biagio Torrisi 1, Giancarlo Fascella 2*, Giuseppina Las Casas 1, Concetta Licciardello 1, Marco Caruso 1, Alessandra Caruso 3 and Filippo Ferlito 1
2 CREA, Research Centre for Plant Protection and Certification, 90128 Palermo, Italy
3 Department of Physiology and Pharmacology “V. Erspamer”, University Sapienza of Rome, 00185 Rome, Italy
* Authors to whom correspondence should be addressed.
Перевод статьи «Research on the Control System for the Conveying and Separation Experimental Platform of Tiger Nut Harvester Based on Sensing Technology and Control Algorithms» авторов Sirui Chen, Jiangtao Qi, Jianping Gao, Wenhui Chen, Jiaming Fei, Hewei Meng, Zhen Ma, оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык

















































































































Комментарии (0)