Изучение переваримости аминокислот сорго без танинов (США) в подвздошном отделе кишечника бройлеров
Цель данного исследования — оценить стандартную илеальную переваримость аминокислот (SIAD) у бройлеров (в возрасте 3 недель) для образцов бестанинного сорго, отобранных в юго-восточном регионе США, а также провести общее сравнение между сорго и кукурузой.
В работе использовалась неполная блочная схема с применением прямого метода для оценки SIAD восьми образцов сорго и одного образца кукурузы в качестве стандарта. Три образца сорго были получены из Северной Каролины (NC-PL, NC-T2, NC-LW), четыре — из Южной Каролины (SC-Flo, SC-Pei, SC-Tub, SC-Experience) и один — из Джорджии (GA-Gf). Каждое зерно скармливалось в восьми клетках, по 13 птиц в клетке.
Птицы содержались в течение 3 недель, при этом последние 4 дня составили экспериментальную фазу, во время которой собирали илеальный химус для оценки SIAD. Переваримость каждого образца сорго сравнивали с кукурузой.
В целом, сорта сорго из Южной Каролины показали наименьшие значения переваримости АК, за исключением SC-Flo, у которого была зафиксирована наивысшая переваримость, превзошедшая кукурузу по лизину, треонину, глицину и аспарагиновой кислоте (p < 0,05).
Далее значения SIAD всех образцов сорго были объединены для общего сравнения с кукурузой. В целом, переваримость АК сорго была сопоставима с кукурузой (p > 0,05), однако между отдельными аминокислотами были выявлены различия. Сорго показало лучшую переваримость по лизину, треонину и аспарагиновой кислоте, тогда как кукуруза — по метионину, лейцину, пролину и фенилаланину (p < 0,05).
Интегрировав данные о содержании сырого протеина с показателями SIAD, это исследование представляет обновленную оценку качества белка бестанинного сорго. Полученные результаты позволяют предположить, что бестанинное сорго может служить практичной альтернативой кукурузе в комбикормах для птицы, обеспечивая сопоставимый питательный профиль.
1. Введение
Кукуруза является основным источником энергии в рационах птицы благодаря своей рентабельности и хорошо изученному питательному профилю [1]. Однако колебания цен и доступности кукурузы делают целесообразным изучение альтернативных зерновых культур для поддержания конкурентоспособности в птицеводстве. Безтанинные сорта сорго представляют собой перспективную альтернативу, предлагая сопоставимые значения видимой метаболизируемой энергии (ВМЭ) около 3600-3700 ккал/кг сухого вещества (СВ) и сходный питательный профиль [2,3]. Исследования продемонстрировали отсутствие негативных эффектов на продуктивность бройлеров при полной замене кукурузы сорго в рационах на основе кукурузносоевого шрота [3,4].
Способность сорго успешно расти на почвах низкого качества и переносить продолжительные засушливые периоды делает эту культуру рентабельной для производителей по сравнению с другими культурами [5,6]. Ее устойчивость позиционирует сорго как жизнеспособную культуру в сельском хозяйстве США, особенно в юго-восточном регионе, где сосредоточена значительная часть птицеводческой отрасли. Увеличение объемов уборки сорго в юго-восточном регионе может принести пользу производителям птицы, поскольку увеличение предложения сорго в «американском бройлерном поясе» может снизить зависимость от кукурузы и уменьшить затраты на транспортировку зерна в эти производственные зоны [7].
Несмотря на свои преимущества, сорго исторически имело стигму «защищенного от птиц зерна» — термин, который относится к ее естественной устойчивости к повреждению птицами due to ее антипитательным и вкусовым характеристикам, в частности танинам [8]. Эти антинутриенты, которые включают танины, нетанниновые фенольные соединения, фитаты и кафирины, негативно влияют на ее питательное качество при использовании в кормах для животных, так как они могут связывать белки и минералы, снижая переваримость и усвоение питательных веществ в рационах птицы [8,9]. Как следствие, исследования привели к разработке безтанинных гибридов и стратегий применения диетических ферментов для смягчения эффектов этих антинутриентов, повышая жизнеспособность сорго как альтернативы кукурузе в комбикормах для птицы [8,10]. В настоящее время Совет по зерну США сообщает об отсутствии танинов в американской сорго в своем ежегодном отчете о качестве сорго [11].
Зерновки сорго демонстрируют широкий спектр цветов, от жемчужно-белого до темно-красного, в основном due to пигментов в семенной кожуре (тесте) и околоплоднике [12,13]. Эта вариация во внешнем виде, в частности у зерновок с пигментированной тестой, часто указывает на более высокие уровни фенольных соединений, которые связаны с потенциально более высокими антипитательными факторами для птицы [14,15]. Широкая генетическая изменчивость сорго также способствует несоответствиям в его питательном составе, осложняя составление рационов и их надежность [8,16]. По этим причинам диетологи часто неохотно включают сорго в рационы птицы, что подкрепляется устаревшими данными о питательной ценности и восприятием сорго как «защищенного от птиц».
Ограниченная и устаревшая информация о переваримости аминокислот (АК) сортов сорго США подчеркивает необходимость проведения обновленных исследований [8,12]. Обзор использования сорго в качестве корма для бройлеров выявил положительную корреляцию между средней подвздошной переваримостью АК сорго, убранного в 2004 и 2005 годах, и выходом грудного мяса (г - 0,672) [8]. Кроме того, были обнаружены значительные различия в переваримости между годами урожая, что подчеркивает влияние генотипа, агротехнических практик и условий окружающей среды на белковый состав сорго [17]. Хотя и сообщалось, что эти факторы влияют на питательный состав зерна, то, как они влияют на переваримость, еще предстоит выяснить. Настоящее исследование представляет обновленные значения стандартной подвздошной переваримости аминокислот (СПП АК) для современных сортов сорго, выращиваемых в США, и устанавливает базовый уровень для будущих исследований. В последующем исследовании авторы оценили влияние агрономических практик и условий окружающей среды на переваримость сорго [Ссылка: рукопись представлена совместно с данной рукописью].
Точные значения переваримости имеют решающее значение для оптимизации рационов, продуктивности и выхода мяса птицы. Учитывая, что современный средний бройлер на 10% тяжелее и на семь пунктов эффективнее, чем 10 лет назад [18], существует острая необходимость в обновленной матрице питательности для сорго, в частности в отношении метаболизируемой энергии и переваримости АК.
В сфере переваримости значения СПП АК дают более точные оценки доступности АК по сравнению с значениями видимой переваримости, поскольку они корректируют на эндогенные потери АК, происходящие из пищеварительных секретов и спущенных клеток. Эта корректировка особенно важна для кормовых средств с низким содержанием протеина, таких как кукуруза и сорго, где относительная доля эндогенных АК может приводить к занижению значений переваримости [19].
Настоящее исследование оценило СПП у 3-недельных бройлеров для зерна безтанинной сорго, убранного в юго-восточной части США, с использованием кукурузы в качестве контроля. Мы выдвинули гипотезу, что не будет существенных различий между образцами сорго по переваримости у бройлеров. Выясняя СПП сортов сорго, мы стремились предоставить обновленную информацию производителям птицы и формуляторам комбикормов, тем самым повышая эффективность и устойчивость этой отрасли.
2. Материалы и методы
Этот эксперимент был проведен в соответствии с принципами и конкретными руководствами, одобренными Комитетом по уходу и использованию животных Университета Клемсона (IACUC), AUP 2023-0191.
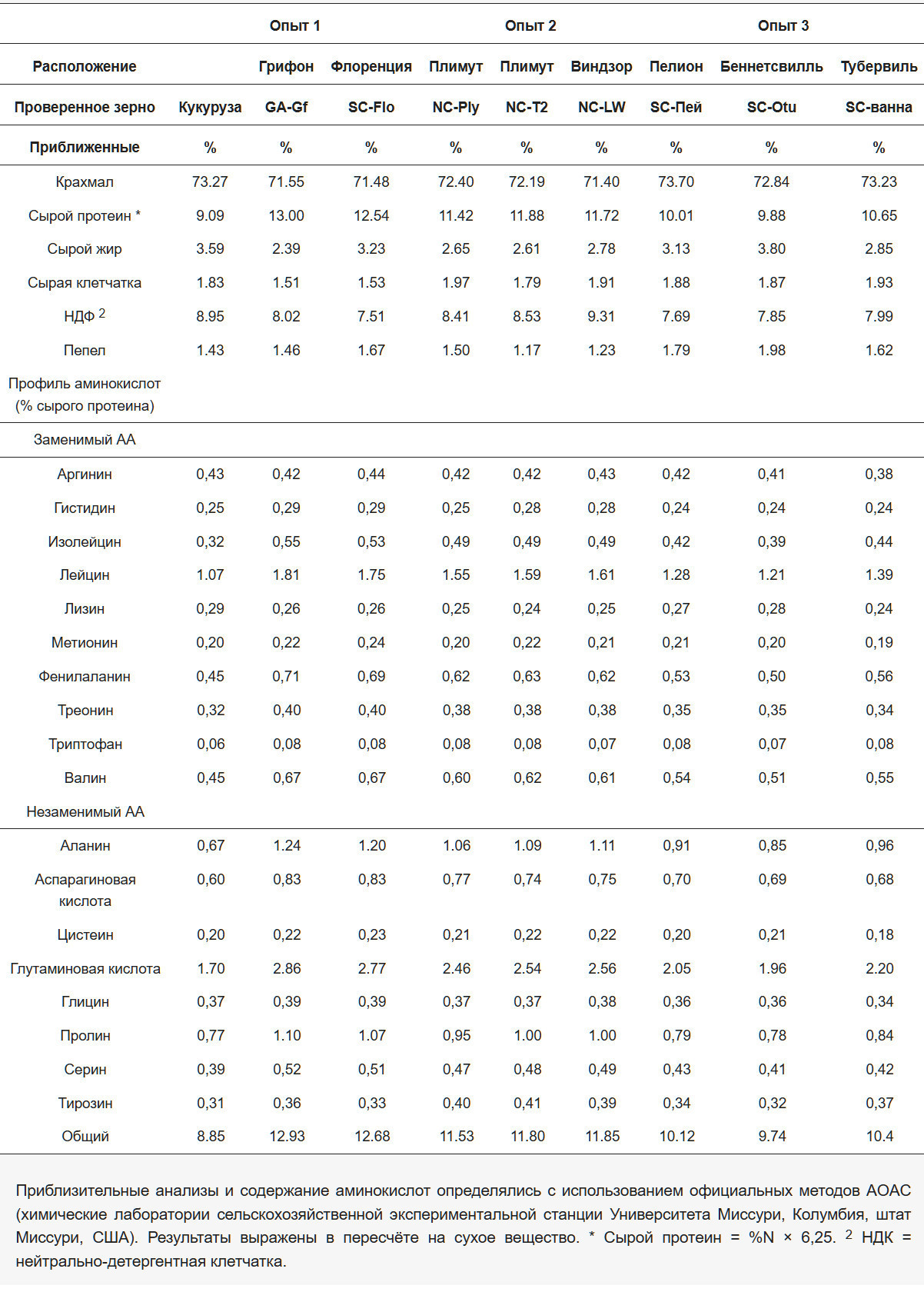
Для оценки СПП восьми образцов зерна безтанинной сорго (Рисунок 1) был проведен эксперимент по переваримости in vivo в Центре птицеводства Моргана в Клемсоне, Южная Каролина, США, осенью/зимой 2023/2024 года. Эти образцы, убранные в сезоне 2023 года, были получены от различных производителей и исследовательских станций на юго-востоке США. В частности, три образца поступили из Северной Каролины: два с экспериментальных станций в Плимуте (NC-PL, NC-T2) и один из Виндзора (NC-LW). Четыре дополнительных образца были собраны в локациях Южной Каролины, включая Флоренс (SC-Flo), Пелион (SC-Pei), Табервилл (SC- Tub) и Беннетсвилл (SC-OTU). Наконец, один образец был получен с экспериментальной станции в Гриффине, Джорджия (GA-Gf). Каждый образец состоял из нескольких гибридов сорго, и их идентичность сохранялась на протяжении всего эксперимента.

Наблюдаемые различия между изображениями отражают внутренние вариации в составе сортов семян, а не артефакты процесса съемки.
Всего в исследовании использовали 936 самцов бройлеров (Cobb 500 х Hubbard) из местного коммерческого инкубатория. Из-за ограниченности пространства исследование проводилось в трех отдельных опытах, в каждом из которых оценивали три вида зерна. Все птицы во всех опытах в этом исследовании следовали одной и той же программе кормления, состоящей из стартового рациона и экспериментального рациона. Стартовый рацион состоял из стандартного рациона на основе кукурузно-соевого шрота (23,53% сырого протеина), полученного от птицеводческого интегратора (Таблица 1). Что касается экспериментального рациона, то в опыте 1 оценивали СПП кукурузы, SC-Flo и GA-Gf; в опыте 2 оценивали NC-PL, NC-T2 и NC-LW; а в опыте 3 оценивали SC- Pei, SC-Tub и SC-OTU.

В каждом опыте размещали 312 суточных цыплят, которых случайным образом распределяли по 24 термостатизированным брудерным батареям для птицы (Модель 0540, GQF Manufacturing Inc., Саванна, Джорджия, США). Брудеры (клетки) были оборудованы приподнятыми сетчатыми полами и размещены в климат-контролируемом помещении, где тщательно регулировались температура, влажность и вентиляция. В каждом опыте использовали шесть брудерных батарей, занимающих верхние четыре из пяти клеток/батареи, с тринадцатью птицами в клетке. Клетки имели размеры 81 см в ширину х 91 см в длину х 25 см в высоту. Птицы содержались с 0 до 21 или 22 дней, при этом последние 3 дня получали экспериментальный рацион. В частности, в опытах 1 и 2 предэкспериментальная фаза длилась с 0 до 19-дневного возраста, за которой следовала экспериментальная фаза с 19 до 22-дневного возраста, а в опыте 3 предэкспериментальная фаза продлилась с 0 до 18-дневного возраста, а экспериментальная фаза проходила с 18 до 21- дневного возраста.
После этого периода всех птиц подвергали эвтаназии для сбора подвздошного содержимого и последующей оценки переваримости. После подтверждения смерти подвздошную кишку удаляли путем энтеротомии, на 1 дюйм дистальнее дивертикула Меккеля и на 1 дюйм проксимальнее илеоцекального соединения. Подвздошный дигест затем промывали дистиллированной водой в пластиковый контейнер. Весь дигест от птиц объединяли в одном контейнере и немедленно отправляли в лабораторию для лиофилизации в рамках подготовки к последующему анализу переваримости.
Репрезентативные образцы стартового рациона, изучаемых зерен и экспериментальных рационов отбирали для химического анализа для обеспечения согласованности. Питательный состав зерен, использованных в опытах по переваримости, приведен в Таблице 2, а состав всех экспериментальных рационов показан в Таблице 3.
Был проведен дополнительный опыт с предоставлением безазотистого рациона (БАР) для определения эндогенных потерь АК (ЭП АК), необходимых для расчета СПП (Таблица 3). Применялась та же методология, что и в других исследованиях, с той разницей, что в общей сложности было задействовано 234 цыпленка. НППР был разработан на основе исследования, проведенного Конгом и Адеолой и соавторами (2013) [20].

Химические анализы проводились на образцах зерна, стартового рациона, полуочищенного рациона (экспериментальный рацион) и подвздошного содержимого. Образцы отправляли в Сельскохозяйственную экспериментальную станцию, Химические лаборатории (ESCL) при Университете Миссури-Колумбия (лаборатория, одобренная AOCS) для анализа [22].
Для определения питательной ценности каждого образца зерна, а также стартового рациона, был проведен приблизительный анализ. Анализ следующих компонентов проводился с использованием методов АОАС: содержание сырого протеина методом сжигания на анализаторе LECO (метод 990.03 A), сырого жира методом эфирной экстракции (метод 920.39) и сырой клетчатки (метод 978.10). Содержание влаги определялось с помощью вакуумной печи (метод 934.01), а содержание золы – с помощью печи для озоления при температуре 600 °C (метод 942.05) [23] .
Анализ аминокислот Cys, Met и Lys + 9 (CML+9) проводился для количественной оценки уровней этих аминокислот в образцах стартового корма, использованных во всех испытаниях, и образцах пищеварительного тракта, полученных при оценке EAL (метод 994.12) [23]. Для определения EAL значения аминокислот Ser, Tyr, Phe, His, Arg и Trp, которые не удалось определить с помощью анализа CML+9, были получены из бухгалтерских данных, предоставленных Ravindran (2021) [24].
Общий профиль АХ и содержание титана определяли для всех экспериментальных рационов и образцов дигеста. Профиль АХ включал оценку концентраций различных АХ, включая Asp, Thr, Ser, Glu, Pro, Gly, Ala, Cys, Vai, Met, lie, Tyr, Phe, Leu, His, Lys, Arg (метод 982.30 E:a,b,c) и Тгр путем щелочного гидролиза (метод 988.15) [23]. Содержание титана определяли в образцах полуочищенного рациона и подвздошного содержимого с использованием методологии, изложенной Myers et al. (2004) [25].
Вес птицы в клетке, а также количество предложенного и отвергнутого корма измеряли в первый и последний день каждой фазы, а падеж птиц собирали и взвешивали дважды в день. Полученные данные использовались для расчета индивидуального веса птицы (BW), прироста веса (BWG), потребления корма (FI) и коэффициента конверсии корма с поправкой на падеж (AdjFCR). Все параметры определялись для подтверждения обеспечения однородных условий содержания, обеспечивающих равномерный рост птиц.
Потребление корма определялось в течение экспериментального периода для точного расчета SIAD. В последний день эксперимента всех птиц взвешивали в клетке и гуманно умерщвляли с помощью CO2 . После подтверждения смерти проводили вскрытие кишечника для отбора проб его содержимого. Подвздошную кишку осторожно удаляли на 2,5 см кзади от дивертикула Меккеля и на 2,5 см кпереди от илеоцекального соединения.
Образцы содержимого подвздошной кишки собирали путём промывания дистиллированной водой в пластиковый контейнер с помощью промывалки. Осторожно вводили кончик промывалки в подвздошную кишку, обеспечивая тщательное промывание и минимизируя риск повреждения просвета кишечника и возможного загрязнения образца.
Образцы Digesta были объединены по клеткам для обеспечения достаточного размера выборки, хранились на сухом льду для сохранения их целостности, а затем транспортировались в лабораторию, где они были заморожены в морозильнике со сверхнизкой температурой -80 °C (Eppendorf, Enfield, CT, США). После этого образцы были лиофилизированы с использованием лотковой сушилки (Labconco, Kansas, MO, США). После лиофилизации образцы были измельчены с помощью коммерческой мельницы (Waring Commercial, Torrington, CT, США) и отправлены в лабораторию для анализа содержания аминокислот и TiO2 для определения SIAD. После того, как были собраны данные химического анализа и эксплуатационных характеристик, были определены стандартизированные коэффициенты усвояемости путем первоначального расчета EAL с использованием уравнения (1) Могана и др. (1992) [ 26 ]:
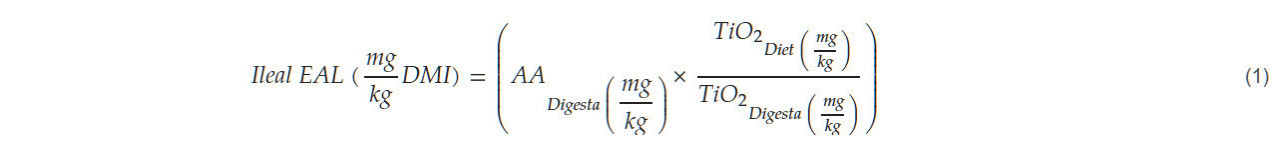
После определения содержания эндогенных аминокислот в подвздошной кишке (EAL) была рассчитана стандартная илеальная переваримость аминокислот (SIAD) с использованием уравнения (2) по Лемме и соавт. (2004) [27].
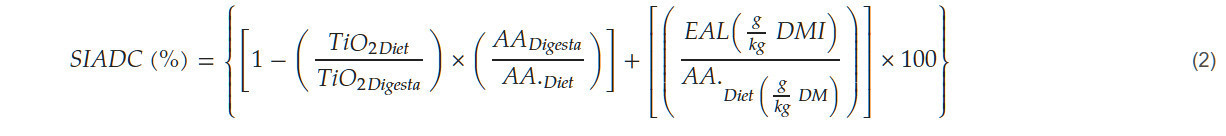
Этот метод широко применяется в исследованиях питания птицы для оценки усвояемости аминокислот в кормах [28]. Уровни диоксида титана как в рационе, так и в пищеварительном тракте, ввиду его неперевариваемости, служат контрольными маркерами. Эти уравнения позволили нам рассчитать долю аминокислот, усвоенных в подвздошной кишке, с учётом эндогенных аминокислот, чтобы избежать занижения результатов.
Всего в эксперименте было девять вариантов рациона, которые распределялись по схеме рандомизированного неполного блока; экспериментальной единицей считалась одна клетка с птицами. Каждый рацион скармливали в восьми клетках-повторностях. Отдельная экспериментальная серия (партия) служила фактором, формирующим неполный блок.
Для анализа показателей продуктивности применяли описательную статистику с расчетом среднего значения и стандартного отклонения.
Статистический анализ включал три этапа:
Данные по восьми образцам сорго были объединены и сопоставлены с данными по кукурузе (n = 64 и n = 8 соответственно) с использованием описательной статистики: среднего значения, стандартного отклонения и доверительного интервала.
Для выявления значимых различий между вариантами рациона был проведен однофакторный дисперсионный анализ (ANOVA) с последующим пост-хок тестом Тьюки для сравнения средних. Анализ выполняли в программе JMP Pro v.16 [29].
С помощью линейного контраста провели сравнение объединенных данных по стандартной илеальной переваримости аминокислот (SIAD) сорго с аналогичным показателем для кукурузы.
Статистически значимыми считали различия при p < 0,05. Значения p в интервале от 0,05 до 0,10 расценивали как тенденцию.
3. Результаты
Результаты химического анализа кукурузы и восьми образцов сорго, изученных в данном исследовании, представлены в таблице 2. Образцы сорго в среднем содержали примерно на 20% больше сырого протеина, чем кукуруза, но имели схожее содержание крахмала (в среднем около 72,81% на сухое вещество). Содержание сырого жира в кукурузе было примерно на 22% выше, чем в сорго, за исключением зерна сорго SC-Otu, которое содержало наибольшее количество жира - на 5,52% больше, чем кукуруза. Содержание клетчатки и золы было сходным среди образцов, за исключением GA-Gf и SC-Flo, где их было примерно на 20% меньше, чем в кукурузе.
Что касается аминокислотного состава, наибольшее общее количество аминокислот было зафиксировано в образцах GA-Gf и SC-Flo, тогда как кукуруза показала наименьшие значения, отставая в среднем на 20% от средних показателей образцов сорго. Аминокислотный профиль был схожим across different grains, за исключением аланина, разветвленных аминокислот (BCAA), глутаминовой кислоты и пролина, содержание которых было выше во всех образцах сорга по сравнению с кукурузой.
Результаты по продуктивности птицы представлены в таблице 4. При анализе данных влияние экспериментальной серии (партии) учитывалось в качестве блокирующего фактора, что позволило эффективно минимизировать потенциальное влияние мешающих переменных.Полученные результаты выявили существенное влияние экспериментальной серии на показатели продуктивности. При этом внутри каждой отдельной серии не было обнаружено статистически значимых различий в продуктивности между группами птиц, получавшими разные виды зерна (p > 0,05). Это свидетельствует о высокой однородности условий содержания птицы в начале экспериментальной фазы в рамках каждой серии.
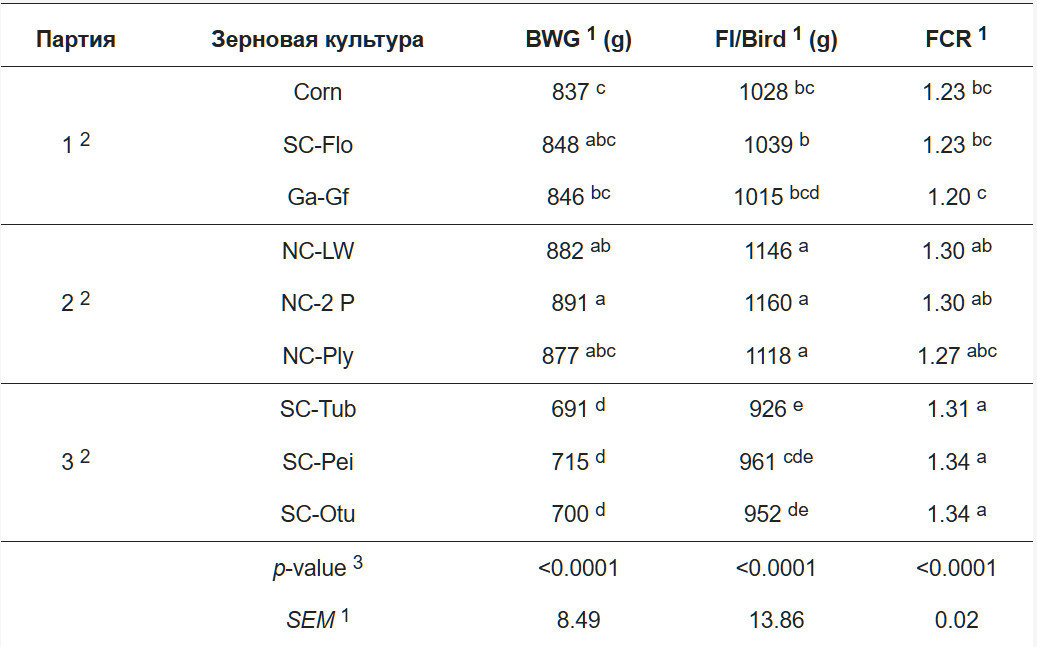
1 - Примечания: Trail -> Партия (экспериментальная серия). Grain -> Зерновая культура. BWG -> Привес (общепринятый зоотехнический термин). FI/Bird -> Потребление корма на птицу. FCR -> Конверсия корма. SEM -> среднее стандартное отклонение для n = 5.
2 Продолжительность предэкспериментального периода: для партий 1 и 2 = 19 дней;
для партии 3 = 18 дней.
3 p-значение получено в результате однофакторного дисперсионного анализа (ANOVA)..
a–e Статистические значимости: значения в одном столбце, не имеющие общего верхнего индекса, статистически значимо различаются при p<0,05.
В таблице 5 представлено сравнение объединённых образцов сорго и кукурузы. Этот анализ был проведён с целью получения более широкого представления о сравнении современного сорго, выращиваемого в США, и кукурузы с учётом изменчивости внутри образцов сорго.
Таблица 5. Описательная статистика сравнения стандартной илеальной переваримости аминокислот (%) кукурузы и зерна сорго юго-восточного региона США у бройлеров в возрасте 3 недель.
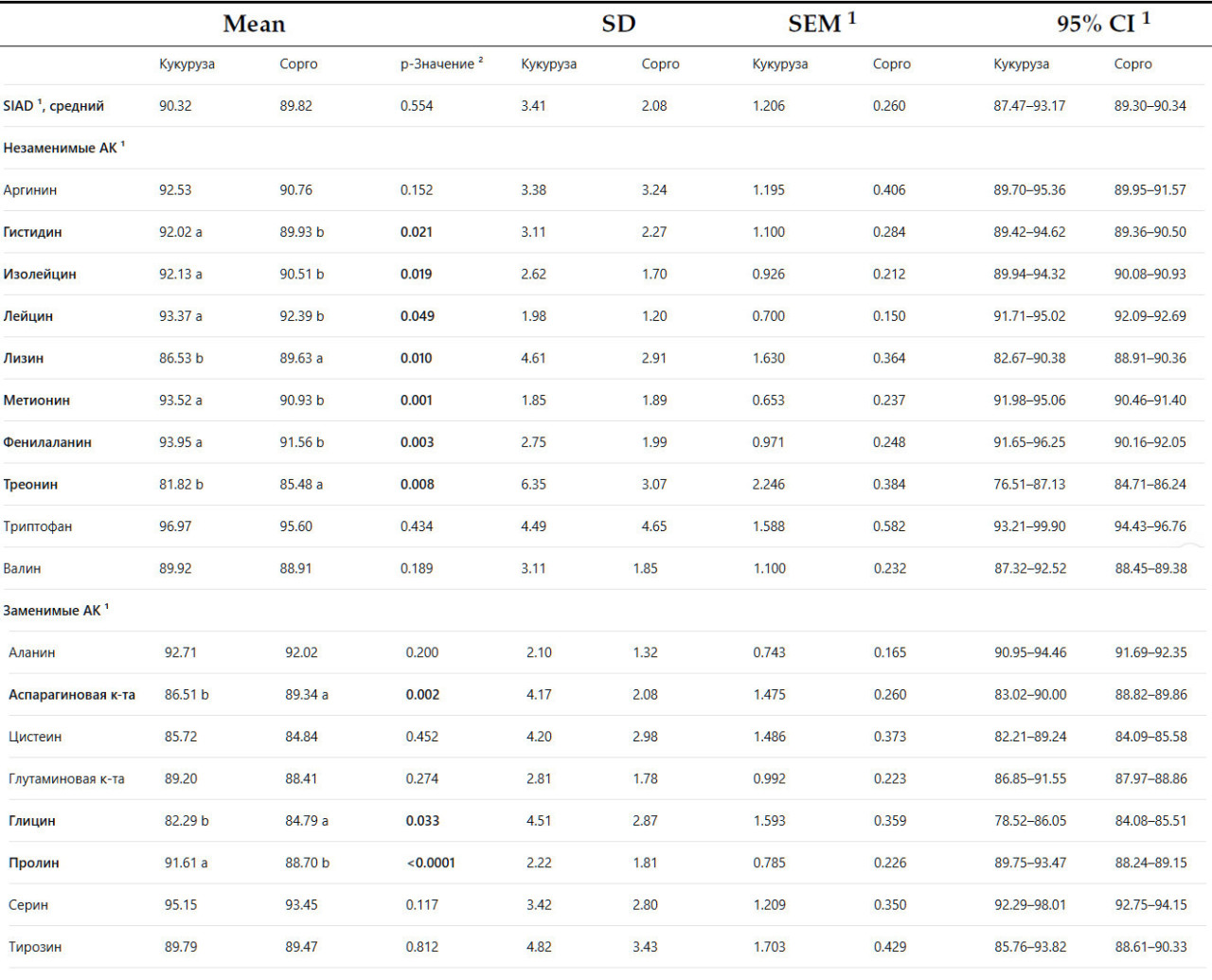
Объединение образцов сорго помогло снизить вариабельность показателей усвояемости, обеспечив более ёмкое сравнение. Результаты не выявили значительной разницы в среднем показателе SIAD между сорго (89,82%) и кукурузой (90,32%) (p> 0,05). Однако были выявлены значительные различия в показателе SIAD 10 из 18 аминокислот (p< 0,05). Метионин, лейцин, илеин, фенилфенил, гистидин и пролин были более усвояемыми в кукурузе, чем в сорго (p< 0,05), в то время как лизин, треонин, глицин и аспарагин были более усвояемыми в сорго (p< 0,05).
Объединение образцов сорго помогло снизить вариабельность показателей усвояемости, обеспечив более ёмкое сравнение. Результаты не выявили значительной разницы в среднем показателе SIAD между сорго (89,82%) и кукурузой (90,32%) ( p > 0,05). Однако были выявлены значительные различия в показателе SIAD 10 из 18 аминокислот ( p < 0,05). Метионин, лейцин, илеин, фенилфенил, гистидин и пролин были более усвояемыми в кукурузе, чем в сорго ( p < 0,05), в то время как лизин, треонин, глицин и аспарагин были более усвояемыми в сорго ( p < 0,05).
3.4. Сравнение средних групп между SIAD оцениваемых образцов зерна
3.4.1 Профили усвояемости образцов сорго
В таблице 6 представлено подробное сравнение индивидуальных значений SIAD для образцов сорго и кукурузы. Во всех наблюдениях наблюдались значительные различия при сравнении исследуемых зерен ( p < 0,05). Образец SC-Flo показал самую высокую среднюю усвояемость – 92,6%, за ним следовали NC-Ply, NC-T2 и GA-Gf с промежуточными значениями от 90,3% до 89,8% соответственно. Напротив, SC-Otu, NC-LW, SC-Pei и SC-Tub продемонстрировали значительно более низкую среднюю усвояемость, чем SC-Flo ( p < 0,05), зафиксировав значения в диапазоне от 89,1% до 88,7% соответственно.
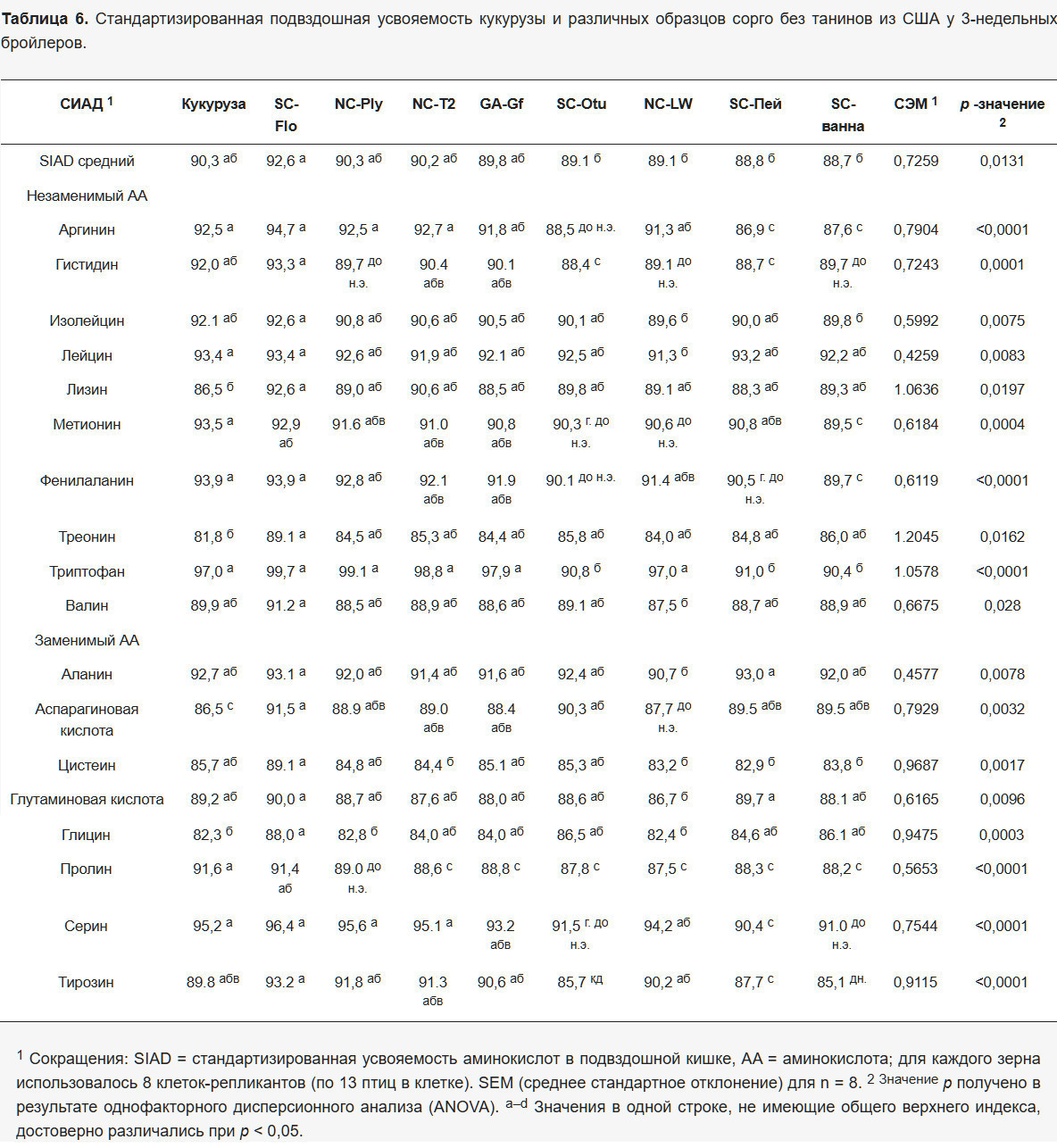
3.4.2. Сравнение отдельных аминокислот
Сравнение SC-Flo и кукурузы выявило схожую усвояемость большинства аминокислот, за исключением Lys, Thr, Gly и Asp, которые были более усвояемы в SC-Flo ( p < 0,05). Примечательно, что Lys и Thr из сорго оказались более усвояемыми, чем из кукурузы, в то время как Pro, по-видимому, был более усвояем в кукурузе, чем в сорго ( p < 0,05), за исключением SC-Flo.
В целом, как показано в Таблице 6 , Trp оказался наиболее усвояемой аминокислотой. Образцы SC-Otu, SC-Pei и SC-Tub, оцененные в последнем испытании, продемонстрировали более низкую усвояемость по сравнению с зерновыми, протестированными в испытаниях 1 (SC-Flo, Corn, GA-Gf) и 2 (NC-Ply, NC-T2, NC-LW; p < 0,05). Кроме того, Met, Arg и Phe показали более низкую усвояемость в образцах SC-Otu, SC-Pei и SC-Tub по сравнению с кукурузой ( p < 0,05), в то время как Leu из NC-LW и Tyr из SC-Tub показали самую низкую усвояемость среди всех оцененных образцов ( p < 0,05). SC-Pei также продемонстрировал более низкую усвояемость для Ser по сравнению с SC-Flo, Corn, NC-Ply и NC-T2 ( p < 0,05). В результате сорт SC-Flo продемонстрировал самую высокую усвояемость среди зерен сорго, при этом значения SIAD были сопоставимы или превышали показатели кукурузы.
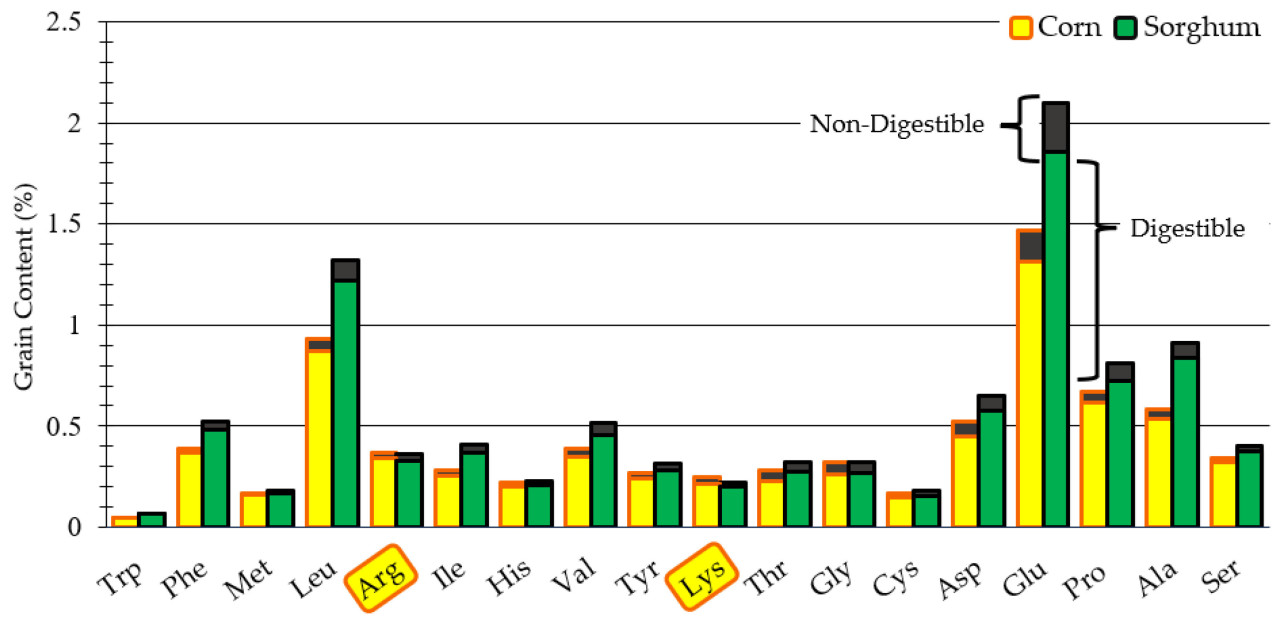
На рисунке 2 объединены данные из таблиц 2 и 5, что позволяет наглядно представить качество белка в зерне, отражая долю усвояемых аминокислот. Сорго, обладающее более высоким содержанием белка и примерно такой же усвояемостью по сравнению с кукурузой, привело к большему количеству легкоусвояемых аминокислот, за исключением аргинина и лизина.
3.4.3. Потери эндогенных аминокислот
Кроме того, подробная информация о SIAD представлена в таблице 7 , где EAL выражена в мг/кг потребления сухого вещества (DMI) в крайнем правом столбце. Ser, Asp и Glu в совокупности составили приблизительно 30% от общего EAL, за ними следуют Thr, Phe, Arg, Lys, Leu, Tyr, Val, Gly, Pro, Ala, Ile и His, каждый из которых вносит примерно 5–10%. Наконец, Cys, Trp и Met в совокупности составили оставшиеся 5–10%.
Таблица 7. Потери эндогенных аминокислот у бройлеров, питающихся безазотистыми рационами.

4. Обсуждение
Авторы признают, что проведение исследования по схеме с неполными блоками в трёх отдельных испытаниях, а не по схеме с полными блоками, где все варианты лечения представлены в каждом испытании, наложило определённые ограничения с точки зрения контроля ошибок и вариабельности. Поскольку в каждом испытании тестировалась только подгруппа образцов сорго, было невозможно полностью контролировать потенциальную межэкспериментальную изменчивость, возникающую из-за небольших различий в условиях окружающей среды, темпах роста птиц или потреблении корма в разных испытаниях. Это ограничение схемы могло повлиять на точность сравнений между всеми вариантами питания, особенно при интерпретации результатов однофакторного дисперсионного анализа (ANOVA) по всем вариантам лечения. Кроме того, логистические ограничения потребовали, чтобы последний эксперимент (испытание 3) завершился на 21-й день, на день раньше, чем испытания 1 и 2, которые завершились на 22-й день.
Эти изменения были необходимы из-за задержек с поставкой некоторых образцов зерна и ограниченного пространства в исследовательских помещениях, что не позволило реализовать полноблочный дизайн. Несмотря на эти ограничения, были предприняты усилия по обеспечению надежности результатов путем объединения данных по генетически разнообразным образцам сорго для получения более широкого диапазона значений усвояемости. Этот подход был направлен на получение практической, отраслевой информации о характеристиках усвояемости сорго, хотя и с некоторыми ограничениями, присущими дизайну. Читателям рекомендуется учитывать эти логистические ограничения при интерпретации результатов, хотя представленные здесь данные представляют собой ценную основу для понимания усвояемости сорго в рационах птицы.
Небольшие различия, наблюдаемые между профилями АА и значениями SIAD в образцах сорго, вероятно, были обусловлены генетическим разнообразием в каждом объединенном образце в сочетании с воздействием агрономических методов, применяемых во время выращивания [ 30 ]. Хотя это и не было статистически значимым, SIAD объединенных образцов сорго был незначительно ниже, чем у кукурузы (89,82% против 90,32%). Это согласуется с литературными данными о том, что сорго, как правило, имеет меньше усвояемого протеина, чем кукуруза, в первую очередь из-за более низкой питательной ценности кафирина, его основного белка проламина [ 8 , 31 ]. Однако более низкая усвояемость сорго определенных аминокислот может быть компенсирована его более высокими концентрациями аминокислот, которые обеспечивают сопоставимое снабжение усвояемых аминокислот кукурузой ( Рисунок 2 ).
Кафирин служит резервом аминокислот во время прорастания семян и составляет 50–70 % от общего содержания белка в сорго [ 32 ]. Его профиль аминокислот богат Glu, Pro и BCAA, но содержит низкие уровни незаменимых аминокислот, таких как Lys [ 33 , 34 ]. Напротив, глютелины, второй по величине компонент белка в сорго, представляют собой высокомолекулярные белки, богатые Lys, в основном расположенные в белковой матрице эндосперма [ 8 , 35 ]. Глютелины демонстрируют более высокую усвояемость, чем кафирины, и исследования показывают обратную корреляцию между содержанием кафирина и общей усвояемостью белка [ 36 ]. Таким образом, сорта сорго с более низким уровнем белка, потенциально указывающие на более низкое соотношение кафирина к глютелину, могут демонстрировать более высокую усвояемость для птицы [ 37 ]. Для сравнения, проламины (зеины) кукурузы составляют меньшую долю её белка (40–50%), оставляя большую долю перевариваемых белков, таких как глютелины и альбумины, для балансировки её аминокислотного профиля [ 38 ]. Наши результаты не могут быть полностью объяснены только содержанием сырого протеина. Исследование значительно выиграло бы от доступа к конкретным измерениям содержания проламинов и глютелинов в исследованных образцах сорго. Такие данные позволили бы более чётко понять взаимосвязь между общим составом белка и наблюдаемыми значениями усвояемости, поскольку полученные результаты не полностью согласуются с данными существующей литературы.
Уровни диетического белка и клетчатки также играют важную роль в модуляции процесса пищеварения и активности ферментов, влияя как на значения эндогенных потерь, так и на SIAD у птицы [ 24 ]. Среди образцов SC-Flo, который имел самые низкие уровни сырой клетчатки (1,53%) и НДК (7,51%), продемонстрировал самую высокую усвояемость. Напротив, известно, что более высокие уровни клетчатки, особенно НДК, содержат некрахмальные полисахариды, такие как гемицеллюлоза и лигнин, которые неперевариваются для птицы. Эти НСП усиливают перистальтику кишечника и ускоряют прохождение питательных веществ, что ограничивает время взаимодействия фермента с субстратом и снижает эффективность усвоения питательных веществ, в конечном итоге снижая SIAD [ 39 , 40 ].
Эта взаимосвязь между содержанием клетчатки и усвояемостью частично объясняет вариабельность SIAD между образцами сорго. Как правило, образцы с более высоким содержанием клетчатки и НДК демонстрировали более низкую усвояемость. Однако сорт NC-Ply, несмотря на относительно высокое содержание клетчатки (1,97%) и НДК (8,41%), достиг значений SIAD, сопоставимых с кукурузой и SC-Flo. Это говорит о том, что, хотя содержание клетчатки является влиятельным фактором, другие переменные, такие как генетика зерна, аминокислотный состав или более низкий уровень антипитательных факторов, также могли способствовать вариабельности SIAD между сортами сорго.
В целом образец сорго SC-Flo показал превосходную усвояемость по сравнению с другими образцами сорго, даже превзойдя кукурузу по усвояемости Lys, Thr, Gly и Asp. Эта повышенная усвояемость может быть приписана специфическим гибридам, включающим SC-Flo, которые показали более светлые цвета зерна (желтый, красный, бронзовый) по сравнению с преимущественно красными зернами в других образцах ( рисунок 1 ). Цвет зерна сорго определяется сложным взаимодействием генетических факторов, включая цвет перикарпия, толщину перикарпия, наличие или отсутствие пигментированной семенной кожуры и цвет эндосперма. Эти характеристики влияют не только на внешний вид зерна, но и на его питательные свойства и усвояемость [ 15 , 41 , 42 ]. Интенсивность цвета зерна часто связана с более высоким содержанием фенольных соединений, которые, обеспечивая антиоксидантные свойства, могут отрицательно влиять на использование питательных веществ и энергии [ 9 , 17 ]. Присутствие более светлых зерен в SC-Flo, вероятно, коррелирует с более низким уровнем этих антипитательных факторов, что потенциально приводит к более высокой усвояемости и AME.
Экологические и агрономические методы управления также играют решающую роль в определении усвояемости зерна. Хотя известно, что эти методы влияют на состав питательных веществ, их влияние на усвояемость питательных веществ требует дальнейшего изучения [ 30 ]. Только одно исследование было проведено по этому вопросу, в котором было показано, что удобрение азотом влияет на усвояемость аминокислот среди сортов тритикале; в то время как оно увеличивает концентрацию аминокислот, оно также снижает усвояемость некоторых аминокислот, таких как Ala, Ile, Lys, Met и Val [ 43 ]. Дополнительные исследования в нашей лаборатории изучили корреляции между факторами окружающей среды и управления урожаем с усвояемостью и составом питательных веществ, хотя важно отметить, что ассоциация не подразумевает причинно-следственную связь. Схожие местоположения и агрономические методы в этом исследовании могли ограничить обнаружение сильных экологических корреляций, оставив генетические компоненты в качестве главного действующего лица в различиях SIAD среди сортов сорго.
Кроме того, важно учитывать, что птицы в опыте 3 начали и закончили экспериментальную фазу на один день раньше, что могло способствовать более низким значениям SIAD, наблюдаемым у SC-Pei, SC-Otu и SC-Tub (в среднем 88,86%) по сравнению с SC-Flo и GA-Gf (в среднем 91,20%) в первом опыте. Исследования показывают, что эффективность пищеварения и развитие ферментов у бройлеров постепенно увеличиваются, достигая примерно 92% мощности к 21 дню [ 44 ]. Учитывая, что у молодых птиц пищеварительная способность ниже, эта небольшая разница в возрасте могла повлиять на значения SIAD, зарегистрированные в опыте 3.
Наконец, общепризнанно, что сорго служит в первую очередь источником энергии в рационе птицы, а не ингредиентом с высоким содержанием белка. Однако максимизация его питательной ценности требует более глубокого понимания усвояемости крахмала и его взаимодействия с белковой матрицей, особенно с фракциями кафирина и глютелина. Усвояемость крахмала сорго составляет в среднем около 83%, что ниже, чем у кукурузы (90%) и пшеницы (88%) [ 45 ]. Эта сниженная усвояемость может быть связана с тесной химической и физической связью крахмала с белковой матрицей, состоящей в основном из кафирина и глютелинов, что ограничивает доступ амилазы [ 8 , 15 ]. В будущих исследованиях можно было бы изучить обработку ферментами, особенно протеазами, для расщепления этой белковой «клетки», потенциально повышая доступность крахмала и использование питательных веществ в корме для птицы. Более глубокое понимание этих взаимодействий может привести к улучшению стратегий питания и составов кормов, которые максимизируют энергетическую эффективность и усвоение белка в рационах бройлеров.
5. Выводы
Было обнаружено, что американское сорго без танинов является ценным ингредиентом с качеством белка, сопоставимым с кукурузным, хотя и немного уступает по количеству легкодоступных лизинов и аргининов. Вариации SIAD наблюдались между различными образцами сорго, что, вероятно, обусловлено генетическим разнообразием внутри каждого образца, включавшего несколько гибридов сорго.
Результаты показали, что, хотя значения SIAD для некоторых аминокислот (например, Met, Leu, Ile, Phe, His, Pro) были ниже в сорго, чем в кукурузе, другие аминокислоты (например, Lys, Thr, Gly, Asp) показали лучшую усвояемость в сорго. Эти результаты предоставляют специалистам по питанию птицы ценные данные для обновления матриц питательных веществ сорго в кормовых составах. Внедряя эти обновления, специалисты по питанию могут помочь обеспечить сбалансированный рацион, потенциально снижая зависимость от кукурузы и оптимизируя затраты на корма за счет более точного удовлетворения потребности бройлеров в аминокислотах.
Вклад авторов: Концептуализация, MA-R. и SS; Методология, MA-R., SS и WB; Формальный анализ, SS и WB; Исследование, MA-R. и SS; Ресурсы, MA-R.; Курирование данных, SS; Написание — подготовка первоначального черновика, SS; Написание — рецензирование и редактирование, MA-R. и WB; Руководство, MA-R.; Администрирование проекта, MA-R. и SS; Получение финансирования, MA-R. Все авторы прочитали и согласились с опубликованной версией рукописи.
Финансирование: Данное исследование финансировалось программой United Sorghum Checkoff Program, грант № RG001-21. Материал основан на работе, поддержанной Национальным институтом продовольствия и сельского хозяйства/Министерством сельского хозяйства США (Вашингтон, округ Колумбия) в рамках проекта № SC-1700565. Это технический вклад № 7349 Экспериментальной станции Университета Клемсона.
Заявление институционального контрольного совета по этике: Протокол исследования на животных был одобрен Комитетом по уходу и использованию животных Университета Клемсона (протокол 2023-0191, одобренный 14 июня 2023 года).
Информированное согласие: Не применимо.
Заявление о доступности данных: Данные, представленные в этом исследовании, доступны по запросу у соответствующего автора.
Благодарности
Авторы выражают благодарность Роберту Бурешу, Майклу Блэру и Ричарду Бойлсу за внесение изменений в данную рукопись. Авторы выражают искреннюю благодарность сотрудникам Центра птицеводства Моргана за их неоценимую помощь, а также студентам бакалавриата кафедры зоотехнии и ветеринарии Университета Клемсона, которые внесли активный вклад в это исследование. Авторы благодарны им за преданность делу и энтузиазм, проявленные на протяжении всего проекта.
Конфликты интересов
Мирей Аргуэльес-Рамос сообщает о сотрудничестве с программой United Sorghum Checkoff Program, включающей оплату лекций. Остальные авторы заявляют об отсутствии конфликта интересов. Финансирующие организации не принимали участия в разработке исследования, сборе, анализе или интерпретации данных, написании рукописи или принятии решения о публикации результатов.
Список литературы
1. Ravindran, V. Poultry feed availability and nutrition in developing countries: Main ingredients used in poultry feed formulation. Poult. Dev. Rev. 2013, 2, 60–63. [Google Scholar]
2. INRAE. Sorghum: Sorghum Grain (Sorghum bicolor (L.) Moench). The Nutritional Values for Monogastric Animals Correspond to Low-Tannin Varieties. 2021. Available online: https://www.feedtables.com/content/sorghum (accessed on 1 March 2024).
3. Moritz, A.H.; Krombeen, S.K.; Presgraves, B.; Blair, M.E.; Buresh, R.E.; Kaminski, R.M.; Bridges, W.C.; Arguelles-Ramos, M.; Wilmoth, T.A. Apparent metabolizable energy and performance of broilers fed selected grain sorghum varieties. Appl. Anim. Sci. 2022, 38, 268–278. [Google Scholar] [CrossRef]
4. Ciurescu, G.; Vasilachi, A.; Idriceanu, L.; Dumitru, M. Effects of corn replacement by sorghum in broiler chickens diets on performance, blood chemistry, and meat quality. Ital. J. Anim. Sci. 2023, 22, 537–547. [Google Scholar] [CrossRef]
5. Beyene, A.; Hussien, S.; Pangirayi, T.; Mark, L. Physiological mechanisms of drought tolerance in sorghum, genetic basis and breeding methods: A review. Afr. J. Agric. Res. 2015, 10, 3029–3040. [Google Scholar] [CrossRef]
6. Naim, A.M.E.l.; Jabereldar, A.A.; El Naim, A.M.; Abdalla, A.A.; Dagash, Y.M. Effect of Water Stress on Yield and Water Use Efficiency of Sorghum (Sorghum bicolor L. Moench) in Semi-Arid Environment. Int. J. Agric. For. 2017, 7. [Google Scholar] [CrossRef]
7. Bean, B. Why Sorghum? 2019. Available online: https://www.sorghumcheckoff.com/agronomy-insights/why-sorghum/ (accessed on 1 March 2024).
8. Selle, P.H.; Cadogan, D.J.; Li, X.; Bryden, W.L. Implications of sorghum in broiler chicken nutrition. Anim. Feed. Sci. Technol. 2010, 156, 57–74. [Google Scholar] [CrossRef]
9. Truong, H.H.; Neilson, K.A.; McInerney, B.V.; Khoddami, A.; Roberts, T.H.; Cadogan, D.J.; Liu, S.Y.; Selle, P.H. Comparative performance of broiler chickens offered nutritionally equivalent diets based on six diverse, “tannin-free” sorghum varieties with quantified concentrations of phenolic compounds, kafirin, and phytate. Anim. Prod. Sci. 2017, 57, 828–838. [Google Scholar] [CrossRef]
10. Liu, S.Y.; Selle, P.H.; Cowieson, A.J. Strategies to enhance the performance of pigs and poultry on sorghum-based diets. Anim. Feed. Sci. Technol. 2013, 181, 1–14. [Google Scholar] [CrossRef]
11. Grain Council. 2023. Sorghum Quality Report. Available online: https://grains.org/wp-content/uploads/2023/03/2022-2023-Sorghum-Quality-Report-Compressed.pdf (accessed on 6 November 2024).
12. Rooney, L.W. Genetics and cytogenetics. In Sorghum: Origin, History, Technology, and Production; Smith, C.W., Frederiksen, R.A., Eds.; Wiley: New York, NY, USA, 2000; Volume 2, pp. 261–308. [Google Scholar]
13. Waniska, R.D.; Rooney, L.W. Structure and chemistry of the sorghum caryopsis. In Sorghum: Origin, History, Technology, and Production; Smith, C.W., Frederiksen, R.A., Eds.; Wiley: New York, NY, USA, 2000; Volume 2, pp. 649–668. [Google Scholar]
14. Boren, B.; Waniska, R.D. Sorghum seed color as an indicator of tannin content. J. Appl. Poult. Res. 1992, 1, 117–121. [Google Scholar] [CrossRef]
15. Serna-Saldivar, S.O.; Espinosa-Ramírez, J. Grain structure and grain chemical composition. In Sorghum and Millets: Chemistry, Technology, and Nutritional Attributes; Woodhead Publishing: Cambridge, UK, 2018. [Google Scholar] [CrossRef]
16. Kemmerer, A.R.; Heywang, B.W. A comparison of various varieties of sorghum as substitute for corn in practical chick diets. Poult. Sci. 1965, 44, 260–264. [Google Scholar] [CrossRef] [PubMed]
17. Chisi, M.; Peterson, G. Breeding and agronomy. In Sorghum and Millets: Chemistry, Technology, and Nutritional Attributes; Woodhead Publishing: Cambridge, UK, 2018. [Google Scholar] [CrossRef]
18. National Chicken Council. U.S. Broiler Performance. 2024. Available online: https://www.nationalchickencouncil.org/statistic/us-broiler-performance/ (accessed on 9 March 2024).
19. Parsons, C.M. Unresolved issues for amino acid digestibility in poultry nutrition. J. Appl. Poult. Res. 2020, 29, 1–10. [Google Scholar] [CrossRef]
20. Kong, C.; Adeola, O. Additivity of amino acid digestibility in corn and soybean meal for broiler chickens and White Pekin ducks. Poult. Sci. 2013, 92, 2381–2388. [Google Scholar] [CrossRef] [PubMed]
21. Adeola, O.; Xue, P.C.; Cowieson, A.J.; Ajuwon, K.M. Basal endogenous losses of amino acids in protein nutrition research for swine and poultry. Anim. Feed Sci. Technol. 2016, 221, 274–283. [Google Scholar] [CrossRef]
22. ESCL. Agricultural Experiment Station Chemical Laboratories University of Missouri-Columbia: Method References. 2024. Available online: https://aescl.missouri.edu/MethRefs.html (accessed on 3 July 2024).
23. Gaithersburg, M.D. International A of OAC. Official Methods of Analysis, 18th ed.; AOAC International: Rockville, MD, USA, 2007. [Google Scholar]
24. Ravindran, V. Progress in ileal endogenous amino acid flow research in poultry. J. Anim. Sci. Biotechnol. 2021, 12, 1–11. [Google Scholar] [CrossRef]
25. Myers, W.D.; Ludden, P.A.; Nayigihugu, V.; Hess, B.W. Technical Note: A procedure for the preparation and quantitative analysis of samples for titanium dioxide. J. Anim. Sci. 2004, 82, 179–183. [Google Scholar] [CrossRef]
26. Moughan, P.J.; Marlies Leenaars, G.S. Endogenous amino acid flow in the stomach and small intestine of the young growing pig. J. Sci. Food Agric. 1992, 60, 437–442. [Google Scholar] [CrossRef]
27. Lemme, A.; Ravindran, V.; Bryden, W.L. Ileal digestibility of amino acids in feed ingredients for broilers. World’s Poult. Sci. J. 2004, 60, 423–438. [Google Scholar] [CrossRef]
28. Adedokun, S.A.; Adeola, O.; Parsons, C.M.; Lilburn, M.S.; Applegate, T.J. Standardized ileal amino acid digestibility of plant feedstuffs in broiler chickens and turkey poults using a nitrogen-free or casein diet. Poult Sci. 2008, 87, 2535–2548. [Google Scholar] [CrossRef] [PubMed]
29. SAS Institute Inc. JMP® 2021; Pro 16. Available online: https://www.jmp.com/en_is/events/mastering/topics/new-in-jmp16-and-jmp-pro16.html (accessed on 15 February 2024).
30. Ajakaiye, C.O. Amino acid composition of sorghum grains as influenced by grain maturity, genotype, and nitrogen fertilization. J. Agric. Food Chem. 1984, 32, 47–50. [Google Scholar] [CrossRef]
31. Taylor, J.; Bean, S.R.; Ioerger, B.P.; Taylor, J.R.N. Preferential binding of sorghum tannins with γ-kafirin and the influence of tannin binding on kafirin digestibility and biodegradation. J. Cereal Sci. 2007, 46, 22–31. [Google Scholar] [CrossRef]
32. Bean, S.R.; Ioerger, B.P.; Smith, B.M.; Blackwell, D.L. Sorghum Protein Structure and Chemistry: Implications for Nutrition and Functionality; ACS Symposium Series: Washington, DC, USA, 2011; Volume 1089. [Google Scholar] [CrossRef]
33. Shull, J.M.; Watterson, J.J.; Kirleis, A.W. Purification and immunocytochemical localization of kafirins in Sorghum bicolor (L. Moench) endosperm. Protoplasma 1992, 171, 64–74. [Google Scholar] [CrossRef]
34. Watterson, J.; Shull, J.M.; Mohamed, A.A.; Reddy, V.; Kirleis, A.W. Isolation of a high-cysteine kafirin protein and its cross-reactivity with gamma-zein antiserum. J. Cereal Sci. 1990, 12, 137–144. [Google Scholar] [CrossRef]
35. Wong, J.H.; Lau, T.; Cai, N.; Singh, J.; Pedersen, J.F.; Vensel, W.H.; Hurkman, W.J.; Wilson, J.D.; Lemaux, P.G.; Buchanan, B.B. Digestibility of protein and starch from sorghum (Sorghum bicolor) is linked to biochemical and structural features of grain endosperm. J. Cereal Sci. 2009, 49, 73–82. [Google Scholar] [CrossRef]
36. Taylor, J.R.N.; Lottelore Schüssler, L.; van der Walt, W.H. Fractionation of proteins from low-tannin sorghum grain. J. Agric. Food Chem. 1984, 32, 149–154. [Google Scholar] [CrossRef] [PubMed]
37. McCuistion, K.C.; Selle, P.H.; Liu, S.Y.; Goodband, R.D. Sorghum as a feed grain for animal production. In Sorghum and Millets: Chemistry, Technology, and Nutritional Attributes; Woodhead Publishing: Cambridge, UK, 2018. [Google Scholar] [CrossRef]
38. Larkins, B.A.; Pedersen, K.; Marks, M.D.; Wilson, D.R. The zein proteins of maize endosperm. Trends. Biochem. Sci. 1984, 9, 306–308. [Google Scholar] [CrossRef]
39. Wu, G. Principles of Animal Nutrition; Taylor & Francis: London, UK, 2017. [Google Scholar] [CrossRef]
40. Kluth, H.; Rodehutscord, M. Effect of inclusion of cellulose in the diet on the inevitable endogenous amino acid losses in the ileum of broiler chicken. Poult. Sci. 2009, 88, 1199–1205. [Google Scholar] [CrossRef]
41. Rooney, L.W. Sorghum and pearl millet lipids. Cereal Chem. 1978, 55, 584–590. [Google Scholar]
42. Diatta-Holgate, E.; Hugghis, E.; Weil, C.; Faye, J.M.; Danquah, A.; Diatta, C.; Tongoona, P.; Danquah, E.Y.; Cisse, N.; Tuinstra, M.R. Natural variability for protein digestibility and grain quality traits in a West African Sorghum Association Panel. J. Cereal Sci. 2022, 107, 103504. [Google Scholar] [CrossRef]
43. Siegert, W.; Boguhn, J.; Maurer, H.P.; Weiss, J.; Zuber, T.; Möhring, J.; Rodehutscord, M. Effect of nitrogen fertilisation on the amino acid digestibility of different triticale genotypes in caecectomised laying hens. J. Sci. Food Agric. 2017, 97, 144–150. [Google Scholar] [CrossRef] [PubMed]
44. Noy, Y.; Sklan, D. Digestion and absorption in the young chick. Poult. Sci. 1995, 74, 366–373. [Google Scholar] [CrossRef] [PubMed]
45. Weurding, R.E.; Veldman, A.; Veen, W.A.G.; Van Der Aar, P.J.; Verstegen, M.W.A. Starch digestion rate in the small intestine of broiler chickens differs among feedstuffs. J. Nutr. 2001, 131, 2329–2335. [Google Scholar] [CrossRef]
Determination of the Standardized Ileal Amino Acid Digestibility of U.S. Tannin-Free Sorghum in Broilers
by Santiago Sasia1, William Bridges2 and Mireille Arguelles-Ramos1
1 Department of Animal and Veterinary Sciences, Clemson University, Clemson, SC 29634, USA
2 Department of Mathematical and Statistical Sciences, Clemson University, Clemson, SC 29634, USA
Перевод статьи «Determination of the Standardized Ileal Amino Acid Digestibility of U.S. Tannin-Free Sorghum in Broilers» авторов Sasia, Santiago, William Bridges, and Mireille Arguelles-Ramos, оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык

















































































































Комментарии (0)