Trioza erytreae (Del Guercio, 1918) и взаимодействие с хозяевами
Выращивание цитрусовых в Средиземноморском регионе имеет важное экономическое значение. Жизнеспособность этой отрасли зависит от ряда факторов, среди которых особое значение имеет адекватный фитосанитарный контроль. За последнее десятилетие ареал инвазивной листоблошки Trioza erytreae (Del Guercio, 1918) расширился до материковой части Португалии и Испании. Trioza erytreae является переносчиком болезни Хуанлунбин (ГЛБ).
Аннотация
В данном обзоре представлены современные знания о хозяевах листоблошки, их привлекательности и пригодности. Приведена классификация хозяев в зависимости от их пригодности, согласно данным литературы. Описаны характеристики хозяев и методы оценки их пригодности, а также климатические факторы, влияющие на взаимодействие листоблошки с хозяином. В обзоре подчеркивается важность комплексной оценки взаимодействий между листоблошками и их хозяевами для разработки и внедрения более эффективных стратегий борьбы с T. erytreae .
1. Введение
Цитрусовая индустрия является важнейшим сектором фруктов в мире, с годовым производством более 166 миллионов тонн [ 1 ]. В настоящее время отрасль сталкивается с невероятно разрушительным бактериальным заболеванием, которое быстро распространяется по всему миру: болезнью Хуанлунбина (HLB). HLB вызывается бактериями Candidatus Liberibacter spp., которые закупоривают флоэму и ограничивают поток питательных веществ в дереве, тем самым влияя на его развитие, производство плодов и качество. Двумя функциональными векторами, которые передают C. Liberibacter, являются листоблошки Diaphorina citri (Kuwayama, 1908) (Hemiptera: Liviidae), в основном присутствующие на азиатском и американском континентах, и Trioza erytreae (Del Guercio, 1918) (Hemiptera: Triozidae), в основном присутствующие на африканском континенте. Два вектора недавно достигли европейского континента, представляя угрозу для средиземноморской цитрусоводства. Diaphorina citri распространяется с востока, будучи обнаруженной в Израиле и на Кипре [ 2 , 3 ], в то время как T. erytreae распространяется с запада, наблюдения на Пиренейском полуострове датируются 2014 годом в северо-западном регионе вблизи береговой линии [ 4 , 5 ].
Средиземноморский бассейн является одним из немногих регионов производства цитрусовых, которые не пострадали от HLB. На сегодняшний день не было получено положительных результатов от тестов HLB, проведенных в Европе для обоих векторов [ 3 , 6 , 7 ]. Потенциальное распространение этих вредных насекомых в Средиземноморском регионе представляет серьезную проблему для производителей цитрусовых, поскольку их присутствие может способствовать быстрому распространению заболевания HLB. Распространение D. citri, переносящего C. Liberibacter, было быстрым в регионах производства цитрусовых на обоих американских континентах в течение 15-летнего периода с момента первого обнаружения HLB. Это привело к сокращению производства цитрусовых на 74% и сокращению числа производителей цитрусовых на 62% во Флориде [ 8 ], что служит демонстрацией разрушительной способности этого заболевания. Управление HLB требует устранения зараженных растений [ 9 ] и существенного изменения агротехнических приемов [ 10 ]. Было испытано несколько стратегий борьбы с этим заболеванием, таких как инъекция антибиотиков в стебель, термотерапия и применение эндофитов. Однако ни один из методов лечения не показал эффективности в борьбе с ГЛБ [ 11 , 12 , 13 ].
Контроль переносчиков листоблошки представляет собой первостепенную задачу для цитрусовой промышленности. Айдо [ 14 ] подробно изложил основные стратегии управления. Эти стратегии включают использование химического контроля, применение инсектицидов в периоды пикового промывания [ 15 ] и биологического контроля с выпуском паразитоидов Tamarixia dryi (Waterston, 1922) [ 16 , 17 ] и T. radiata [ 14 ]. Выпуск Tamarixia dryi был проведен во Франции, Испании и Португалии, охватывая все территории, которые были заражены, и зависел от соответствующих государственных органов [ 4 , 7 , 18 , 19 , 20 , 21 , 22 ]. Кроме того, в настоящее время изучаются новые стратегии, основанные на энтомопатогенах [ 23 ] и применении каолина [ 24 ].
Присутствие T. erytreae на Пиренейском полуострове представляет собой серьезную проблему для защиты основных европейских производителей цитрусовых. Множество факторов, включая климатические условия [ 25 ], наличие естественных врагов [ 26 ], химическую обработку и растения-хозяева, оказывают существенное влияние на выживание и развитие T. erytreae . Насекомое проявляет высокую степень зависимости от своих хозяев, поскольку они играют ключевую роль в его укоренении, развитии и распространении. Более того, взрослая листоблошка имеет ограниченную продолжительность жизни (85 часов), когда лишена своих хозяев [ 27 , 28 , 29 ]. При отсутствии бактерий прямой ущерб, наносимый листоблошкой цитрусовым хозяевам, считается незначительным [ 30 , 31 ]. Тем не менее, специалисты по питомникам сообщают о значительных последствиях, поскольку симптомы ямчатого галла, образующиеся на листьях цитрусовых во время развития нимф, влияют на пригодность растений для коммерциализации [ 30 , 31 ]. Более глубокое понимание круга хозяев T. erytreae может способствовать разработке более эффективных стратегий контроля и продвижению эпидемиологических исследований листоблошек, учитывая ключевую роль взаимодействий между переносчиком, растением и патогеном. В этом обзоре представлены обновленные данные о современных знаниях о взаимодействии T. erytreae с ее хозяевами, с особым акцентом на факторы, влияющие на ее укоренение, откладку яиц и развитие.
2. Trioza erytreae Хозяева
Trioza erytreae питается и развивается в основном на растениях семейства рутовых [ 30 , 31 ]. Trioza erytreae откладывает яйца на верхушки молодых побегов. Нимфы вылупляются и поселяются на нижней стороне развивающихся листьев, где они завершают свое пятивозрастное развитие до появления новой летающей взрослой особи. Во время своего развития нимфы образуют «ямочные галлы», которые представляют собой круглые или овальные углубления на нижней стороне листа. Ямочные галлы идеально подходят к нимфе, и на верхней стороне листа они видны как выпуклые вздутия [ 30 , 32 ].
Подсемейство Aurantioideae, которое является частью семейства Rutaceae, включает 33 рода. Род Citrus , который является частью этого подсемейства, включает предпочтительных хозяев T. erytreae [ 32 ]. Три вида из Rutaceae, а именно два Aurantioideae, Clausena anisata [(Willd.) Hook.fil., De Wild. & Staner] и Citrus × limon (L.) Burm, наряду с Vepris lanceolata [(Lam.) G.Don] (не Aurantioideae), были исторически связаны с T. erytreae , поскольку они были первыми хозяевами листоблошек, которые были задокументированы в литературе [ 21 , 30 , 31 , 33 , 34 , 35 ]. Первым зарегистрированным хозяином был Citrus × limon , который был официально описан в 1918 году в Эритрее [ 36 ]. Почти все виды и разновидности рода Citrus служат хозяевами, включая те, которые используются в качестве подвоев [ 37 ]. Термин «пригодность» используется для обозначения способности хозяина поддерживать все стадии развития листоблошки до появления нового поколения. Пригодность некоторых видов цитрусовых остается неубедительной, как в случае C. australasica (F.Muell) [ 21 ], или даже противоречивой, как наблюдалось у C. trifoliata (L.) [ 21 , 37 ] и C. japonica (Thunb.) [ 21 , 25 ].
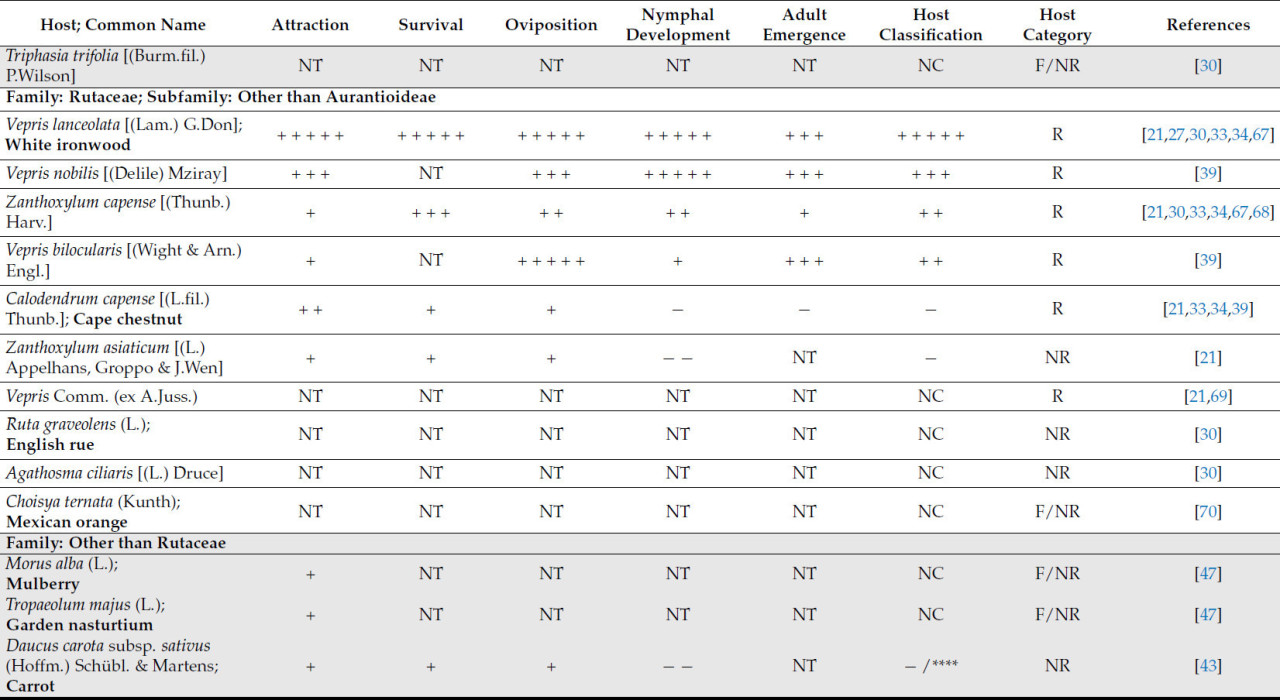
Растения из других родов подсемейства Aurantioideae за пределами рода Citrus были описаны как подходящие хозяева. К ним относятся Murraya paniculata [(L.) Jacq.] и Glycosmis pentaphylla [(Retz.) Corrêa], которые обычно используются в качестве декоративных растений [ 38 ]. Отсутствует единое мнение относительно пригодности определенных хозяев Aurantioideae для роста листоблошки. Например, C. anisata была указана как подходящая хозяйка в некоторых исследованиях [ 21 , 30 , 39 , 40 ], в то время как в других она была определена как неподходящая из-за отсутствия яйцекладки [ 33 ]. Аналогичным образом, Calodendrum capense [(L.fil.) Thunb.], который был описан как подходящая хозяйка [ 39 ], был признан неподходящей в других исследованиях [ 21 , 30 , 34 ]. Подходящими хозяевами Rutaceae, выявленными за пределами подсемейства Aurantioideae, были V. lanceolata и Zanthoxylum capense [(Thunb.) Harv] [ 21 , 30 , 34 ].
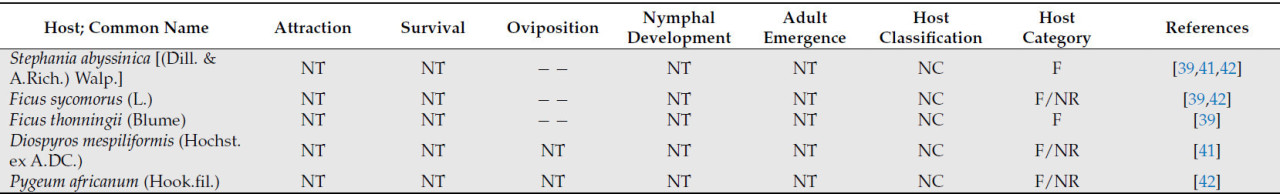
Возможные виды хозяев за пределами семейства Rutaceae — это Ficus spp. (L.) [ 41 ], включая Ficus sycomorus (L.) (семейство Moraceae) [ 42 ], Pygeum africanum (Hook.fil.) (семейство Rosaceae) [ 42 ], Stephania abyssinica [(Dill. & A.Rich.) Walp.] (семейство Menispermaceae) [ 41 , 42 ] и Diospyros mespiliformis (Hochst. ex A.DC.) (семейство Ebenaceae) [ 41 ]. Trioza erytreae, как было задокументировано, питается только этими хозяевами и является вероятной причиной симптомов галлов на листьях. Однако нет никаких доказательств, позволяющих предположить, что произошло развитие нимф или появление нового поколения листоблошек [ 41 , 42 ]. Таким образом, в свете современных знаний эти виды можно рассматривать только как нерепродуктивных или питающихся хозяев ( таблица 1 ).
Растения моркови [ Daucus carota subsp. sativus (Hoffm.) Schübl. & Martens] также рассматривались в качестве возможных хозяев для передачи Candidatus Liberibacter solanacearum растениям апельсина [ C. aurantium (L.)]. Однако, несмотря на откладку яиц, T. erytreae не смогла завершить свой жизненный цикл [ 43 ].
Как отмечалось в предыдущих параграфах, имеющиеся данные подтверждают классификацию T. erytreae как олигофага. Собранные данные указывают на то, что хозяева, не относящиеся к семейству рутовых , являются нежизнеспособными. Некоторые из этих нежизнеспособных хозяев могут быть потенциально использованы для борьбы с листоблошками. Например, исследователи предложили использовать Nicotiana tabacum , нежизнеспособного хозяина, вблизи цитрусовых садов в качестве средства борьбы с D. citri [ 44 ].
В таблице 1 представлен краткий обзор данных о пригодности T. erytreae в качестве хозяев, изученных к настоящему времени. Хозяева были классифицированы в соответствии с их привлекательностью для листоблошки и пригодностью в качестве хозяев на основе оценки результатов, описанных в литературе.
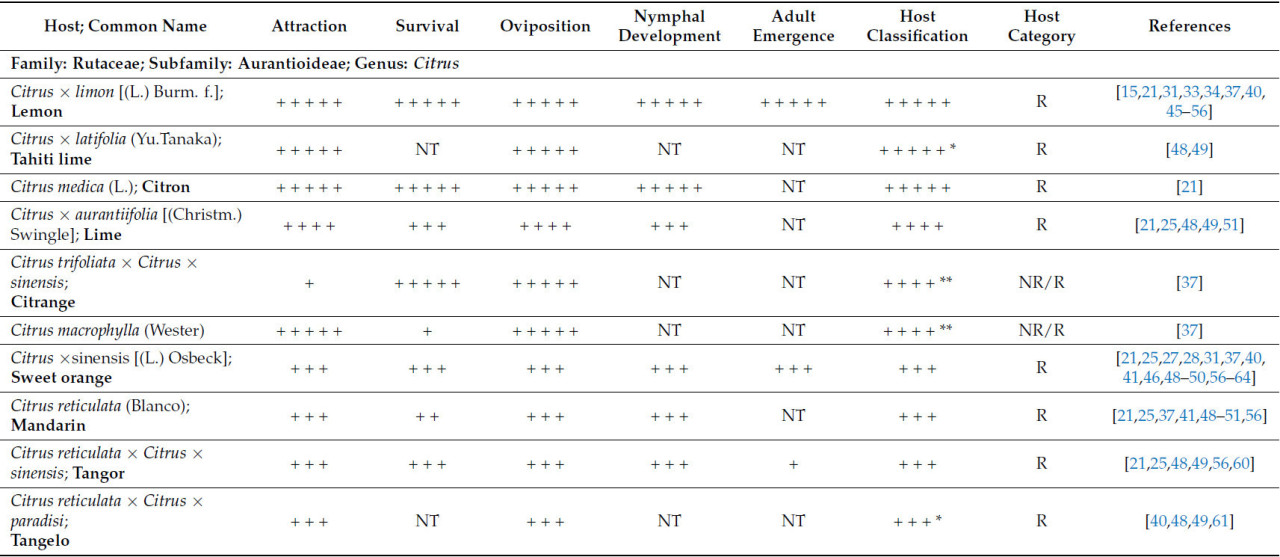
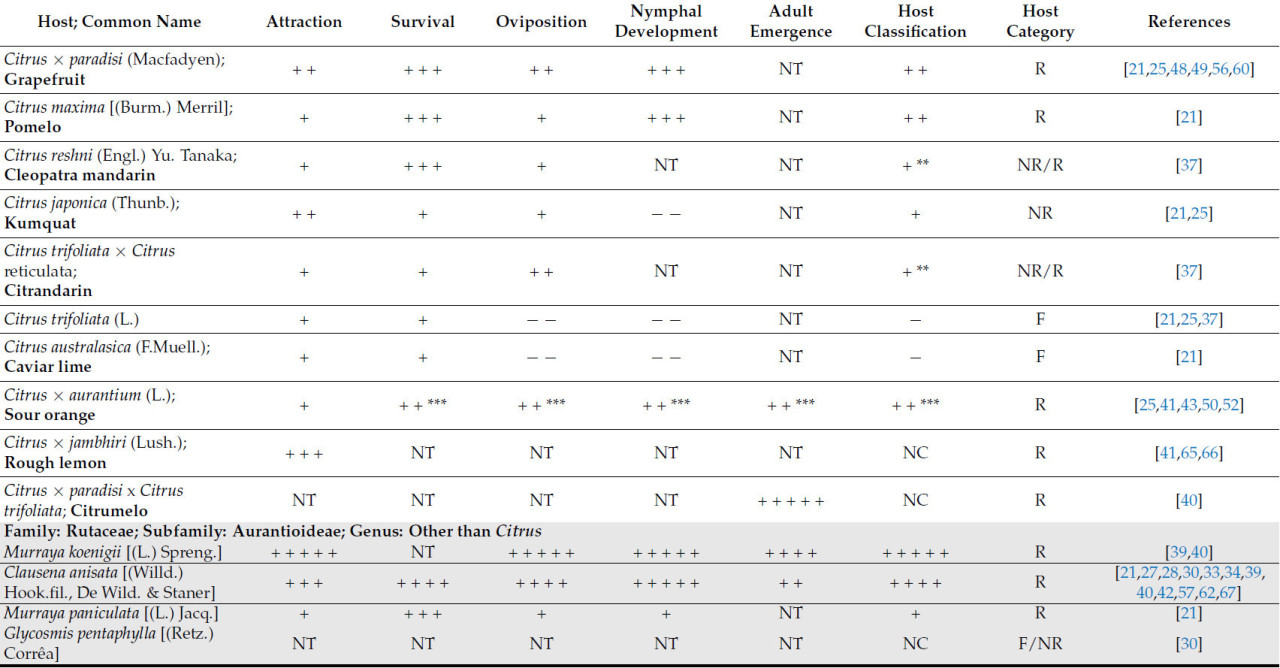
Что касается рода Citrus , который включает в себя более 30 видов, только 17 были оценены как потенциальные хозяева для T. erytreae . Десять видов были классифицированы как имеющие высокую или наивысшую пригодность для T. erytreae . Наибольшая пригодность была выявлена для лимона ( C. × limon ), цитрона ( C. medica ) и лайма ( C. × aurantiifolia ) ( Таблица 1 ). Два наиболее широко культивируемых цитрусовых дерева [ 1 ], сладкий апельсин ( C. × sinensis) и мандарин ( C. reticulata ), были классифицированы как имеющие хорошую пригодность в качестве хозяина ( Таблица 1 ). Кроме того, Clausena anisata и M. koenigii (L.) Spreng., два дополнительных вида из подсемейства Aurantioideae, были классифицированы как имеющие высокую и наивысшую пригодность соответственно ( Таблица 1 ).
Видимо, виды семейства Rutaceae, не входящие в подсемейство Aurantioideae, менее пригодны для T. erytreae . Только два представителя рода Vepris продемонстрировали классификацию пригодности выше среднего уровня ( таблица 1 ).
Учитывая скудность исследований растений-хозяев, не относящихся к семейству рутовых, пока невозможно сделать вывод об их непригодности. Однако имеющиеся данные свидетельствуют о том, что изученные на данный момент растения-хозяева непригодны ( таблица 1 ).
В то время как большинство цитрусовых растений-хозяев продемонстрировали сопоставимые уровни привлекательности и пригодности для откладки яиц ( таблица 1 ), два растения-хозяина, а именно цитранж ( C. trifoliata × C. × sinensis ) [ 37 ] и Vepris bilocularis [(Wight & Arn.) Engl.] [ 39 ], продемонстрировали низкий уровень привлекательности и высокую скорость откладки яиц. Это говорит о том, что эти растения-хозяева обладают свойствами, позволяющими им оптимально откладывать яйца, не привлекая T. erytreae ( таблица 1 ).
3. Характеристики хозяина и их влияние на развитие Trioza erytreae
Пригодность хозяина для T. erytreae зависит от внутренних и внешних факторов. Следующие аспекты, связанные с молодыми волнами, особенно важны для определения пригодности хозяина: интенсивность волн, время волн, длина листа, длина побега и мягкость/сочность тканей. Кроме того, большое значение имеют пищевой статус, возраст и фитосанитарное состояние растения-хозяина. Значимость этих характеристик хозяина заключается в потребности в молодых листьях и побегах для успешной откладки яиц и развития нимф [ 28 , 57 , 58 , 64 , 69 , 71 ] ( Рисунок 1 ).
![<b>Рисунок 1.</b> Различные стадии развития листьев и симптомы заражения <i>Trioza erytreae</i> , а также стадии развития листоблошки. Верхняя половина рисунка изображает внешний вид листьев и фенологические стадии <i>Citrus</i> × <i>limon</i> согласно шкале Biologische Bundesanstalt, Bundessortenamt und Chemische Industrie (BBCH) [ <a href="https://www.mdpi.com/2077-0472/15/1/101#B72-agriculture-15-00101"><b>72</b></a> ] при заражении стадиями <i>T. erytreae</i> . Нижняя часть рисунка изображает стадии развития <i>Trioza erytreae</i> : стадия 0 — яйца; стадия 1 — нимфа первой стадии; стадия 2 — нимфа второй стадии; стадия 3 — нимфа третьей стадии; стадия 4 — нимфа четвертой стадии; стадия 5 — нимфа пятой стадии; и стадия A — взрослая особь.](https://storage.agriexpert.ru/storage/images/articles/5537/block/67436/model/xl/DFvE9n5VPJP8.jpg)
Фитосанитарный статус растения-хозяина оказывает влияние на ритм промывания и состояние питания побегов и листьев, что, в свою очередь, влияет на рост и развитие псиллид. Например, снижение уровня азота в листьях существенно затруднило развитие псиллид [ 59 ]. Кроме того, в саду C. × sinensis наличие хлоротичных молодых побегов вызвало высокий уровень смертности T. erytreae. Более того, было замечено, что выжившие нимфы демонстрировали уменьшенный размер и уплощенную морфологию, а также более длительный период стадий развития [ 59 ], в отличие от более быстрого распространения T. erytreae , наблюдаемого на молодых, здоровых и сильных деревьях [ 61 ].
Trioza erytreae демонстрирует тенденцию к перемещению между подходящими хозяевами вблизи цитрусовых садов в зависимости от наличия свежего прилива. Распространенность листоблошек выше, когда цитрусовые сады подвергаются чередованию прилива с внесезонным приливом у хозяев, расположенных за пределами садов [ 58 , 62 , 68 ]. Таким образом, развитие T. erytreae облегчается, если растение-хозяин дает молодые приливы в течение всего года [ 69 ]. Citrus × limon является весьма привлекательным хозяином для листоблошек из-за непрерывного образования молодых приливов в течение года, что обеспечивает постоянные возможности для заселения и роста [ 58 , 73 ].
Нимфы T. erytreae могут перемещаться на расстояние до 300 мм в поисках оптимальных мест питания, которые характеризуются наличием молодых побегов и мягких тканей [ 57 ]. Чем больше твердость ткани, тем больше времени нимфы будут искать, тем самым увеличивая вероятность обезвоживания, нападения или паразитирования [ 57 ]. По сравнению с нимфами, развивающимися по обе стороны листьев (более мягкие ткани), меньше нимф завершают свое развитие на ветвях (более жесткие ткани), что действует как сдерживающий фактор для откладки яиц [ 53 , 57 ]. Было замечено, что более длинные побеги, которые характерны для лимонных деревьев и мандаринов сатсума [ 73 ], как правило, привлекают большее количество псиллид [ 60 ]. По мере созревания и роста листа наблюдается снижение скорости вылупления яиц и выживаемости нимф. Следовательно, зрелые, более длинные и крупные листья пагубно влияют на развитие T. erytreae [ 59 , 60 ]. Это может быть связано с твёрдостью листьев, которая ухудшает способность яиц поглощать воду, что является жизненно важным условием их выживания [ 59 , 74 ].
Trioza erytreae способна продлевать свою жизнь и период до откладки яиц при отсутствии молодых взлётов. Однако это явление наблюдалось лишь в течение ограниченного периода времени [ 58 ]. В цитрусовых садах Южной Африки высокая интенсивность взлётов сделала растение весьма привлекательным для T. erytreae , в то время как низкое количество молодых взлётов привело к высокой смертности листоблошки. Это указывает на то, что количество взлётов является важным фактором, влияющим на привлекательность листоблошки к растению-хозяину [ 61 , 64 ].
Время промывки имеет решающее значение, поскольку, когда оно совпадает с оптимальными климатическими условиями для развития T. erytreae , наблюдаются значительные пики численности популяции [ 58 ]. Эффективность паразитоидов в борьбе с T. erytreae повышается при высокой численности популяции листоблошки [ 75 ].
Предыдущие исследования показали, что растения, зараженные HLB, распространяются в разные периоды по сравнению со здоровыми растениями [ 76 ]. Наличие как здоровых, так и зараженных HLB растений с асинхронными периодами роста создает более благоприятные условия для размножения листоблошки [ 58 ]. Этот аспект следует учитывать при управлении садами, пораженными HLB.
Чтобы обеспечить собственное выживание и снизить конкуренцию за ресурсы со стороны своего вида, T. erytreae избегает зон интенсивного заражения. Такое поведение способствует расселению насекомых, что приводит к поиску новых, неколонизированных побегов. В своём исследовании Ван ден Берг и др. [ 77 ] наблюдали положительную корреляцию между количеством яиц, нимф и взрослых особей на хозяине и скоростью расселения.
Обрезка — это культурная практика, которая регулирует рост новых побегов [ 73 ]. Срезы верхушки стимулируют развитие новых побегов [ 78 ] и делают цитрусовые деревья более привлекательными для псиллид. Верхушечная обрезка — это операция, которая состоит из нескольких срезов верхушки, применяемых к верхушкам деревьев [ 73 ], тем самым вызывая развитие многочисленных новых побегов и привлекая псиллид [ 79 ]. Эта операция уже была опробована для борьбы с D. citri в сочетании с применением инсектицидов к обрезанным деревьям [ 79 ]. Кроме того, было показано, что дефицитное орошение продлевает период, в течение которого цитрусовые растения не показывают приливов. Это было подтверждено в лимонных садах, подвергавшихся дефицитному орошению в течение 12-недельного периода зимой, хотя та же процедура не повлияла на сладкие апельсиновые деревья [ 71 ]. Можно сделать обоснованный вывод, что выращивание сладких апельсинов и лимонов в одном саду или в соседних садах не рекомендуется, учитывая, что лимонные деревья демонстрируют непрерывный отток плодов [ 73 ]. В период оттока плодов у сладких апельсинов наблюдается приток насекомых, мигрирующих с лимонных деревьев на апельсиновые, что приводит к увеличению популяции листоблошки [ 58 ]. Для внедрения агротехнических приемов, снижающих численность популяций T. erytreae , необходимо всестороннее понимание циклов оттока плодов у цитрусовых и эффективных методов управления.
4. Влияние климатических условий на Trioza erytreae и ее хозяев
Климатические переменные оказывают влияние на продолжительность развития насекомых [ 30 , 32 , 69 ], а также на интенсивность и сроки вымывания хозяина [ 80 ]. Два основных исследования, описанных в этом разделе, были проведены в климатически контролируемых условиях, и в обоих в качестве хозяина T. erytreae использовался C. × limon [ 5 , 55 ]. В других исследованиях хозяин-цитрус не указывается [ 30 , 32 , 54 ] или они основаны на полевых исследованиях на садах сладкого апельсина ( C. × sinensis ) [ 63 , 64 ]. Установлено, что продолжительность процесса развития от стадии яйца до стадии взрослой особи существенно зависит от температуры [ 5 , 55 ]. Оптимальный диапазон температур для роста популяции T. erytreae составляет от 18 °C до 24 °C [ 5 , 55 ]. Это соответствует диапазону среднесуточных температур весной от 12 °C до 20 °C в субтропических регионах, где цитрусовые деревья дают значительное количество коротких побегов [ 80 ].
Температуры выше 27 °C или ниже 10 °C серьезно задерживают или предотвращают завершение жизненного цикла T. erytreae , и если температура постоянна на уровне 10 °C, 27 °C или 30 °C, жизненный цикл не завершается [ 5 ]. Хорошо известно, что цитрусовые хозяева впадают в состояние покоя, когда среднесуточная температура опускается ниже 12 °C [ 80 ]. Таким образом, температура ниже 10 °C будет ограничивать развитие как хозяина, так и листоблошки. В то время как температуры выше 30 °C, по-видимому, оказывают пагубное воздействие на листоблошку, этого не происходит в отношении смывания хозяина, учитывая, что цитрусовые хозяева производят длинные новые побеги при среднесуточной температуре в диапазоне от 25 °C до 35 °C [ 80 ]. Конкретные условия, необходимые для каждой стадии развития, были предметом значительных исследований. Период, предшествующий откладке яиц, при температуре 25 °C длится от 3,4 до 10,5 дней [ 5 ]. Жизнеспособность яиц снижается при температуре ниже 8 °C и выше 33 °C [ 5 ]. Успешное развитие яиц и нимф зависит от диапазона температур от 15 °C до 24 °C [ 5 , 55 ]. Айду и др. [ 55 ] наблюдали, что уровень смертности нимф первой стадии был самым высоким при 15 °C и самым низким при 18 °C, тогда как третья стадия демонстрировала самые высокие показатели выживаемости при 20 °C.
Продолжительность жизненного цикла T. erytreae также зависит от влияния влажности. Недавние исследования, проведенные в контролируемых условиях, показали, что при относительной влажности 65% и температуре 15 °C период развития составляет 56,23 дня, тогда как при 24 °C он сокращается до 19,95 дня. Аналогично, при температуре 15 °C и относительной влажности 70% период развития составляет 46,7 дня, тогда как при 25 °C он сокращается до 23,9 дня [ 5 , 55 ]. Что касается развития нимф, то при постоянной температуре 25 °C, когда поддерживались как 40% ОВ, так и 90% ОВ, никакого развития не наблюдалось. В этих условиях листоблошка достигала только стадии третьей стадии, а время, необходимое для предоткладки яиц и вылупления яиц, удлинялось. Однако при относительной влажности 70% весь жизненный цикл листоблошки завершился за 23,9 дня [ 5 ].
Влияние климатических условий на время развития насекомых также косвенно влияет на общую численность популяции T. erytreae [ 49 , 63 , 70 ]. Высокие показатели смертности были приписаны жарким и сухим летним дням [ 30 ]. 100% смертность яиц и нимф первой стадии наблюдалась, когда параметры температуры и влажности, представленные как индекс дефицита насыщения (SD), составляли 45 мбар или выше. При 35 мбар уровень смертности составлял 70%, а при 15 мбар он снижался до 10% [ 21 ]. В результате Кэтлинг [ 64 ] ввел термин «летальные дни» для описания периодов, когда значения превышали 34,6 мбар, что оказывало значительное влияние на жизнеспособность яиц и стадию развития первой стадии [ 55 , 64 ].
Исследования, проведенные до 1970 года с использованием веток цитрусовых (вид цитрусовых не указан), показали, что необходимая продолжительность вылупления яиц составляет 7–9 дней, с диапазоном 5–17 дней в летних и зимних условиях соответственно. Стадия нимфы длилась в среднем 20–27 дней, причем в летних условиях требовалось 18 дней, а в зимних — 34 дня [ 30 , 32 , 54 ]. Кроме того, продолжительность жизни взрослого насекомого в периоды теплой погоды составляет от 26 до 36 дней [ 32 ]. Более поздние исследования, проведенные в контролируемых условиях, показали аналогичную тенденцию к вышеупомянутым исследованиям веток, с небольшими различиями. Яйца вылуплялись в среднем через 7,2–13,5 дней после откладки яиц, в то время как развитие нимф длилось от 16,4 до 33,4 дней [ 5 ]. Указанные временные рамки относятся к постоянным температурам 15 °C и 25 °C соответственно. Перес-Отеро и др. [ 5 ] дополнительно наблюдали, что самки переживают самцов при всех протестированных температурах. Средняя выживаемость самок составила 44,2 дня при 15 °C, тогда как средняя выживаемость самцов составила 17,2 дня при 25 °C. У цитрусовых растений, выращиваемых в умеренном климате, продолжительность от распускания почек до полного развития листьев составляет 60 дней, когда среднесуточные температуры превышают 13 °C [ 81 ]. Этот временной интервал позволяет T. erytreae генерировать по крайней мере одно поколение. В оптимальных климатических условиях за сезон промывки могут появиться два поколения [ 58 ].
Для определения географических регионов, где T. erytreae может процветать, необходимо оценить климатические условия в зависимости от орографии региона, а также наличие или отсутствие подходящих хозяев. Этот систематический подход позволяет определить регионы, где T. erytreae может обосноваться и размножиться, что способствует оперативному принятию защитных мер [ 16 , 82 , 83 ].
5. Методы изучения привлекательности и пригодности хозяина для T. erytreae и их применение
Для комплексного изучения взаимодействия насекомого с хозяином и пригодности хозяина применялось несколько методологий. К ним относятся популяционные исследования, тесты выбора, тесты без выбора, исследования развития насекомых и морфометрические исследования, а также исследования химических и молекулярных взаимодействий. Некоторые исследования фокусируются на факторах, направляющих T. erytreae к хозяину, в то время как другие – на факторах, влияющих на выживание и рост насекомого после выбора хозяина ( рис. 2 ).
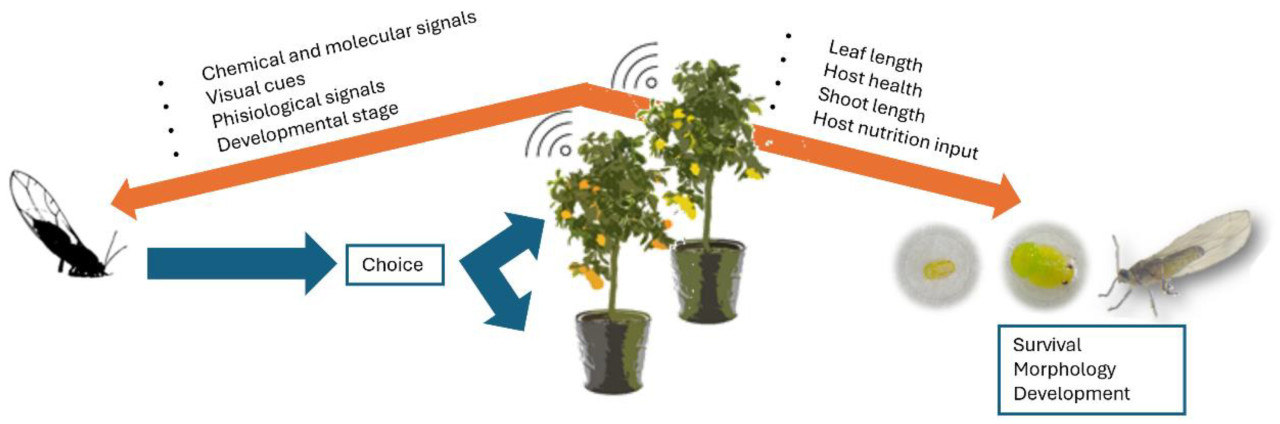
В ходе популяционных обследований отбираются растения-хозяева в определённом географическом районе и в определённый временной интервал для регистрации наличия и/или симптомов T. erytreae. Этот подход позволяет определить естественные предпочтения листоблошки. Такие обследования могут проводиться на национальном уровне или на уровне отдельных садов.
Обширные обследования популяций представляют собой ценный начальный шаг в определении хозяев псиллид и различении их предпочтений [ 4 , 41 , 84 ]. Для иллюстрации, обследование на национальном уровне, проведенное в Уганде, задокументировало долю каждого вида хозяина в общем количестве выявленных зараженных растений. Результаты показали, что мандарины ( C. reticulata ) были предпочтительными хозяевами (66,7%), за ними следовали кислый апельсин ( C. aurantium ) и шершавый лимон C. × jambhiri (Lush.) (оба с 13,3%), в дополнение к наименее привлекательному сорту сладкого апельсина «Washington Navel» (6,7%) ( C. sinensis ) [ 41 ]. В том же исследовании три растения, не относящиеся к семейству рутовых, были идентифицированы как имеющие ямчатые галлы и взрослых особей T. erytreae , а именно Stephania abyssinica (Dill. & A. Rich) Walp. var. tomentella (Oliv.) Deils (семейство Menispermaceae), Diospyros mespiliformis (семейство Ebenaceae) и Ficus spp. (семейство Moraceae). Однако отсутствие наблюдений нимф [ 41 ] указывает на необходимость дальнейших исследований для определения пригодности этих хозяев для T. erytreae .
Изучение популяций псиллид на уровне сада дает значительные преимущества с точки зрения информации, которую они предоставляют относительно привлекательности хозяина и их пригодности для T. erytreae. Это в первую очередь связано с уменьшенной эдафоклиматической и орографической изменчивостью, наблюдаемой в таких исследованиях. Результаты исследований, проведенных в многовидовых садах, предоставили ценную информацию об привлекательности хозяина для T. erytreae . Выводы Сэмвейса и Мэникома [ 60 ] показали, что сорт 'Valencia' ( C . × sinensis ) продемонстрировал более высокое среднее количество ветвей с яйцами и взрослыми особями T. erytreae по сравнению с апельсином 'Navel' ( C . × sinensis ), тангором 'Ortanique' ( C . reticulata × C . × sinensis ) и грейпфрутом ( C . × paradisi (Macfadyen)). Напротив, сопоставимое исследование, проведенное Ван ден Бергом и соавторами [ 25 ], показало, что апельсин сорта «Навел» был более привлекательным, чем апельсин сорта «Валенсия», что демонстрирует, как переменные, присущие экспериментальной области и методологии, могут влиять на результаты.
Значительное количество сортов мандарина ( C. reticulata ) , а также их гибридов, тангора ( C. reticulata × C. × sinensis ) и танжело ( C. reticulata × C. × paradisi ), были оценены на предмет их привлекательности для T. erytreae в различных многовидовых садах [48 , 51 , 77 ] . Результаты этих исследований указывают на то, что мандарины имеют высокую степень внутривидовой изменчивости с точки зрения привлекательности для T. erytreae . Подгруппа сортов «Сацума», включая «Овари», «Сайгон» и «Васэ», оказалась весьма привлекательной для листоблошки. Напротив, сорта «Dancy», «Fortune» (гибрид «Dancy») и гибридные сорта танжело, такие как «Page» и «Osceola», демонстрируют низкую привлекательность для T. erytreae ( таблица 2 ) [ 48 , 51 , 77 ].
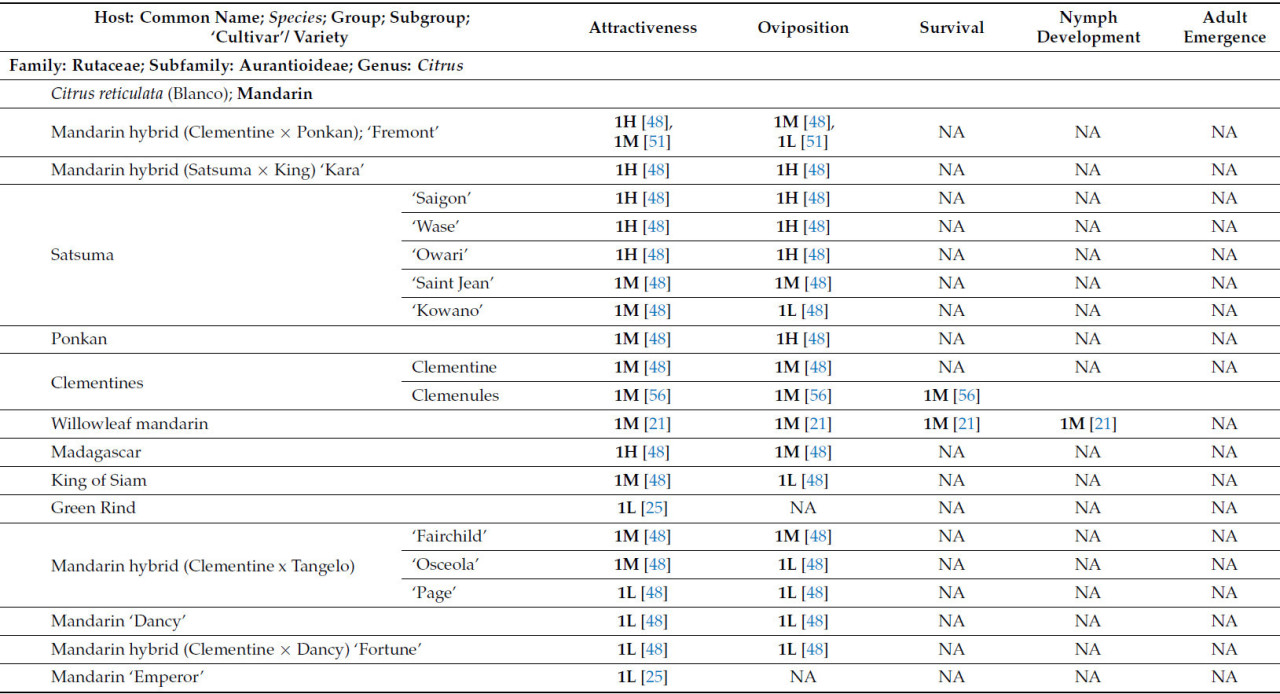
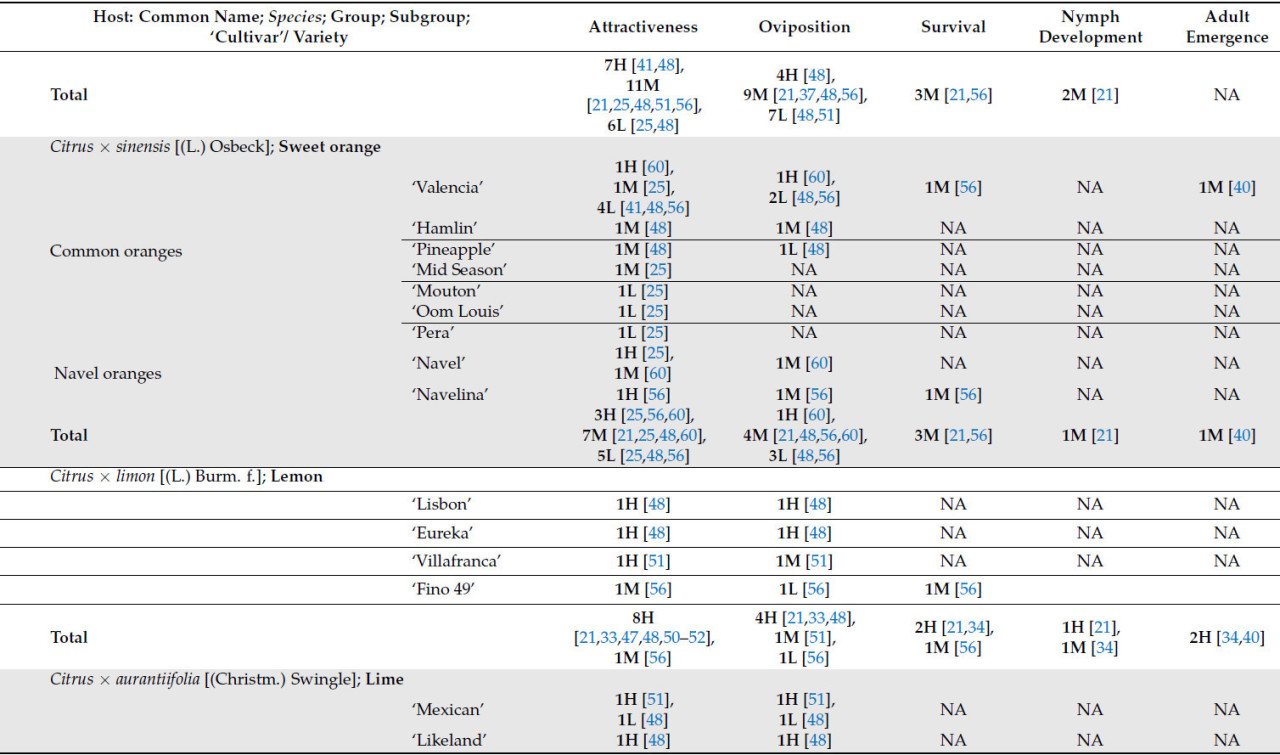
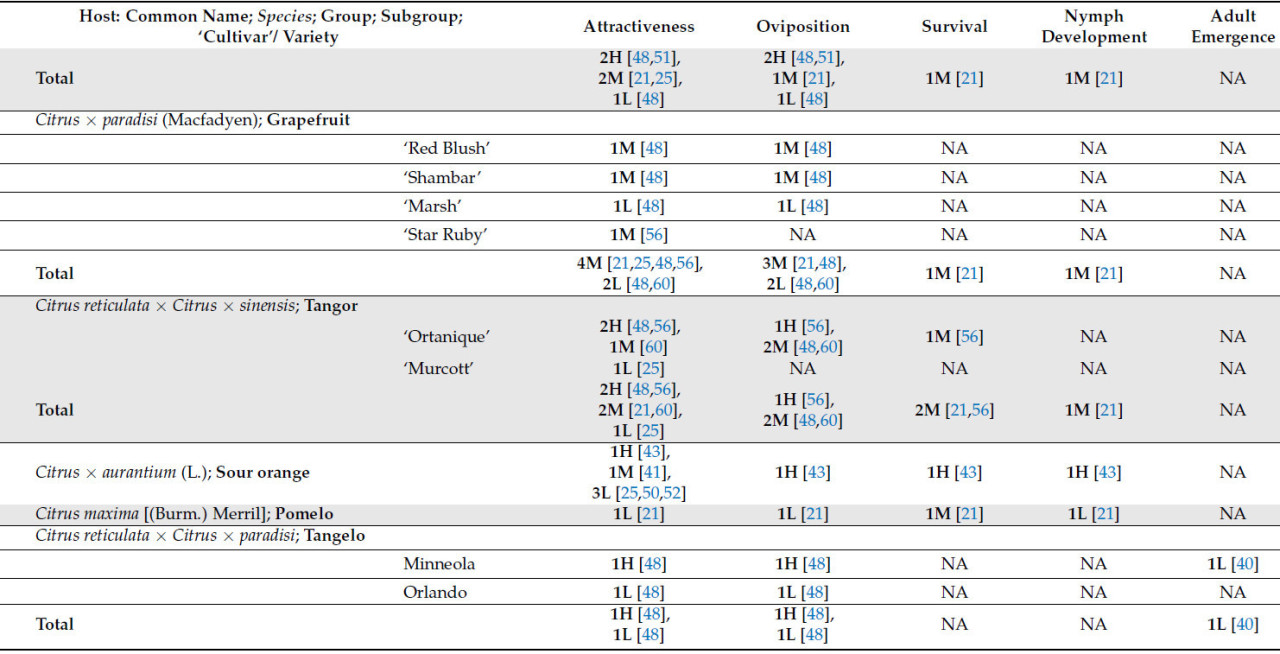
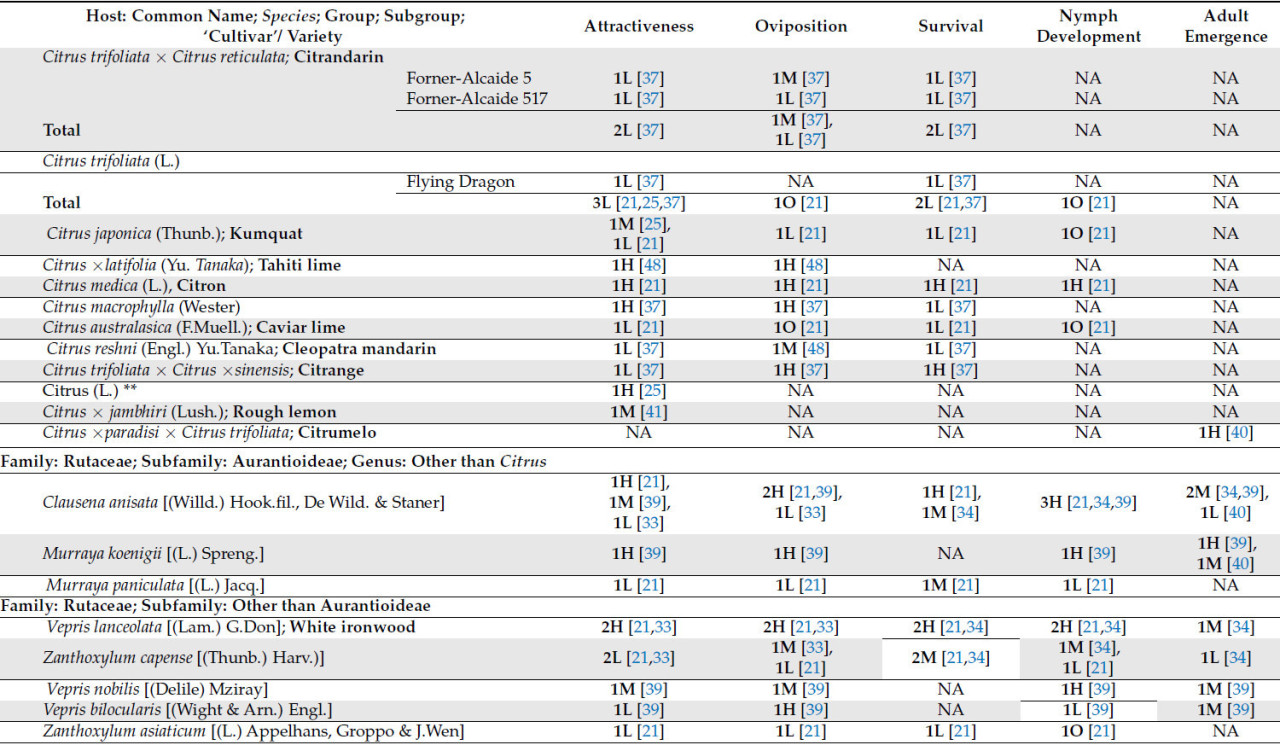
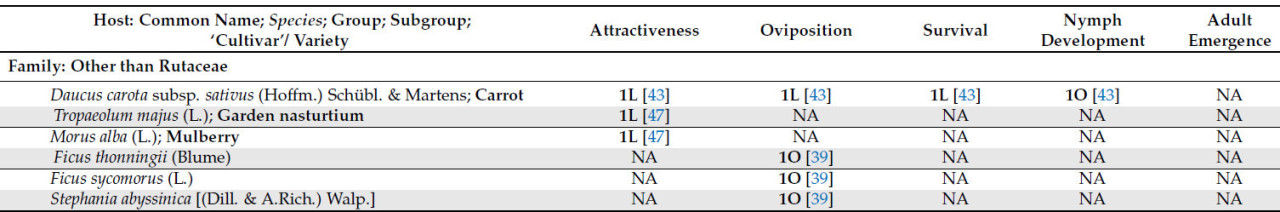
«Total» представляет собой сумму всех сравнений, описанных для каждого хозяина; следует отметить, что количество сравнений может превышать указанное в вышеупомянутых подгруппах, поскольку в некоторых исследованиях не указывается исследуемый сорт или разновидность; NA означает «Not Plaiden»; номенклатура OLMH: «O» представляет собой нулевое значение, «L» представляет собой низкие сравнительные значения, «M» представляет собой промежуточные сравнительные значения, а «H» представляет собой высокие сравнительные значения. Число, предшествующее номенклатуре OLMH, представляет собой количество классифицированных хозяев. Это число иногда превышает количество ссылок, поскольку в некоторых исследованиях сравнивалось более одного сорта одного и того же вида. ** « Citrus » представляет собой исследования, в которых хозяин упоминался как «citrus micellaneous crosss».
Привлекательность T. erytreae к Citrus × limon была хорошо задокументирована [ 85 ]. В саду кислых апельсинов ( C. aurantium ) на одном дереве C. × limon было обнаружено в два раза больше псиллид в желтых липких ловушках по сравнению с другими ловушками, размещенными рядом с кислыми апельсинами [ 50 ] . В отличие от сортов C. × sinensis и C. reticulata , различные сорта C. × limon , как было замечено, постоянно демонстрируют высокие уровни привлекательности и скорости откладки яиц [ 48 , 51 ]. Единственным исключением был лимон «Fino 49», поскольку, несмотря на высокую скорость откладки яиц при прививке на подвой цитранжа Карризо ( C. trifoliata × C. × sinensis ), при прививке на другие протестированные подвои наблюдалась низкая скорость откладки яиц [ 56 ] ( таблица 2 ). Можно сделать вывод, что для некоторых видов цитрусовых привлечение и откладка яиц листоблошкой в меньшей степени зависят от сорта, вероятно, из-за меньшей генетической изменчивости этих видов [ 86 ].
Характеристики хозяина и сигналы, которые он подаёт для привлечения или отпугивания листоблошки, также можно изучать с помощью экспериментов с выбором. Результаты исследований с использованием тестов выбора, проведённых в контролируемой среде, позволили понять характеристики листоблошек, влияющие на их предпочтения и привлекательность. Исследование показало, что пол листоблошки может оказывать влияние на её привлекательность к хозяину. В тесте с двойным выбором вероятность выбора кислого апельсина составила 39% для самцов и 19% для самок [ 52 ].
Экспериментальное исследование показало, что мягкость листьев влияет на скорость откладки яиц T. erytreae ; однако она не повлияла на поведение листоблошки при оседании [ 33 , 53 ]. Кроме того, экспериментальное исследование показало, что откладка яиц не происходит на листьях с твёрдостью, превышающей 90 г/мм [ 53 ]. Значения твёрдости представляют собой вес, необходимый для того, чтобы булавка с плоским концом диаметром 0,254 мм проколола 1 мм листовой ткани [ 53 , 87 ].
Тест выбора и тест без выбора использовались для сравнения привлекательности и яйцекладки T. erytreae на непривитых подвоях. Результаты показали, что C. macrophylla является наиболее привлекательным хозяином, в то время как цитранж 'Carrizo' ( C. trifoliata × C. × sinensis ) продемонстрировал самую высокую скорость яйцекладки [ 37 ]. Самая низкая частота яйцекладки наблюдалась у C. trifoliata , который был определен как наименее привлекательный хозяин [ 37 ]. Имеющиеся данные указывают на то, что подвои цитрусовых могут влиять на летучие профили привоя-хозяина, что, в свою очередь, может влиять на привлекательность для них вредителей листоблошек [ 88 ]. Это также может влиять на пригодность привоя для T. erytreae , влияя как на привлекательность, так и на скорость яйцекладки. Некоторые сорта, по-видимому, подвержены большему влиянию, как это видно на примере сорта «Fino 49» C. × limon [ 56 ].
Использование экспериментов без выбора, в которых насекомому предоставляется только один вариант хозяина, позволяет изучить пригодность хозяина, а также выживаемость и особенности поведения насекомых после заселения. Этот подход даёт ценные данные для эпидемиологических исследований, включая количество взрослых особей, формирующих новое поколение [ 34 ], и количество поколений, формируемых в течение года [ 58 ].
В эксперименте без выбора Айду и др. [ 39 ] наблюдали различия в откладке яиц среди восьми нецитрусовых хозяев из семейства рутовых. У Clausena anisata был самый высокий процент откладывания яиц (52%), в то время как у C. capense были самые низкие значения (24%). У Ficus thonningii, F. sycomorus и S. abyssinica откладка яиц не наблюдалась. Морковь ( D. carota subsp. sativus ) оказалась непригодной, поскольку ни одна нимфа не смогла достичь стадии взрослой особи [ 43 ].
Исследования морфометрии T. erytreae , развивающихся в разных хозяевах, дали представление о влиянии видов хозяев на развитие листоблошки. Сравнительный анализ был проведен на пяти нецитрусовых хозяевах из семейства Rutaceae. Результаты показали, что C. capense дал наименьшее количество и самых маленьких имаго T. erytreae , Clausena anisata - второе по количеству и самых крупных имаго, а Murraya koenigii - наибольшее количество вылупившихся имаго с размерами, аналогичными тем, которые сформировались у Clausena anisata [ 39 ]. Кроме того, морфология и размер крыльев T. erytreae [ 40 ] также различались у разных видов хозяев, что потенциально могло влиять на способность листоблошки к полету и расселению.
Привлечение псиллид к хозяевам зависит от летучих веществ растений. Вальтерова и соавторы [ 89 ] провели исследование псиллид Dyspersa apicalis Foerster, которая принадлежит к семейству Triozidae, среди разнообразных видов хозяев. В исследовании изучались предпочтения псиллид в питании и откладке яиц в зависимости от летучих веществ, получаемых от хозяев, и пришли к выводу, что наименее привлекательные виды хозяев демонстрировали более высокое содержание лимонена. У цитрусовых концентрация летучих веществ в молодых листьях была выше, чем в зрелых листьях, несмотря на отсутствие каких-либо изменений в их привлекательности при оценке с помощью теста выбора [ 66 ]. Тем не менее, эти результаты свидетельствуют о том, что более высокая концентрация летучих веществ в молодых листьях может увеличить вероятность того, что T. erytreae обнаружит их в полевых условиях. Среди летучих веществ, вырабатываемых хозяевами, терпены, по-видимому, играют роль в их привлекательности для T. erytreae [ 66 ]. Профиль летучих веществ как молодых, так и зрелых листьев C. × jambhiri , в частности (S)-(−)-лимонен, сабинен и β-оцимен, использовался в виде синтетических смесей, изолированно и в различных соотношениях, в тестах на выбор. Эти летучие вещества были более эффективны в привлечении T. erytreae в сочетании с другими из профиля летучих веществ растения, чем при использовании по отдельности [ 66 ]. Листья Vepris lanceolata [(Lam.) G. Don] имеют лимонный запах, похожий на запах C. × limon [(L.) Burm. f.], что может быть связано с высокой привлекательностью и высокой скоростью откладки яиц T. erytreae , наблюдаемой у этого вида растений [ 21 , 33 ] ( таблица 1 ). В свете этих многообещающих результатов, в состав липких ловушки желтого цвета для привлечения T. erytreae были включены летучие вещества растительного происхождения (уксусная кислота, (R)-(+)-лимонен, сабинен, смесь изомеров оцимена, включающая цис-оцимен и ß-, мирцен, этилбутират, метилсалицилат и п-цимен) . Однако в полевых условиях эта добавка оказалась неэффективной [ 90 ].
Анализ летучих веществ растений, не являющихся хозяевами, показал, что T. erytreae избегает летучих веществ гуавы ( Psidium guajava L.), чеснока ( Allium sativum L.) и лимонника ( Cymbopogon citratus (DC.) Stapf) [ 65 ]. Исследования показали, что практика посадки цитрусовых деревьев с гуавой ( P. guajava ) на открытых полях снижает популяции D. citri в садах. Однако в контролируемых условиях теплицы эффекты не были очевидны [ 91 , 92 ]. Было бы интересно изучить влияние посадки гуавы, чеснока и/или лимонника в цитрусовых садах на T. erytreae , чтобы выяснить отпугивающий эффект в полевых условиях.
Большинство исследований взаимодействия T. erytreae с растениями-хозяевами были сосредоточены на анализе летучих веществ растений. Тем не менее, недавнее исследование, проведенное нашей исследовательской группой с использованием экспериментального дизайна без выбора и протеомного анализа, показало, что протеомная реакция растений лимона и сладкого апельсина на T. erytreae была различной. Протеомная реакция растений сладкого апельсина на листоблошку была более выраженной и обширной [ 46 ]. Данное исследование предполагает, что растения-хозяева цитрусовых корректируют свой протеом в ответ на заражение T. erytreae , что может быть связано с пригодностью хозяина.
6. Заключительные замечания и перспективы на будущее
Целью данного обзора было представить синтез существующих знаний о хозяевах T. erytreae и их взаимодействии с листоблошкой. Несмотря на обширные исследования, проведенные по T. erytreae , необходимы дальнейшие исследования для выяснения влияния хозяина на развитие нимф и появление имаго. Это включает в себя анализ количества и соотношения вылупляющихся яиц и нимф, времени развития нимф, появления имаго и морфометрии вылупившихся имаго. Понимание взаимодействия насекомого с хозяином на молекулярном уровне необходимо для разработки эффективных стратегий контроля над листоблошкой. Разработка эффективных искусственных диет для T. erytreae и улучшение уже протестированных диет, таких как предложенная Расселом и Пельц-Стелинском [ 93 ] для D. citri, может облегчить изучение влияния изолированных соединений рациона на листоблошек. Это может потенциально привести к разработке эффективной стратегии контроля популяций листоблошек.
Основная проблема, выявленная в этом обзоре, заключалась в пробеле в знаниях о молекулярном аспекте этого специфического взаимодействия насекомого и хозяина. Подходы, основанные на омиксах, обеспечивают всестороннее понимание взаимодействий между насекомыми и растениями [ 94 ]. Применение подходов на основе омиксов к изучению D. citri способствовало более глубокому пониманию реакции хозяина на заражение псиллидами [ 95 , 96 ]. Кроме того, стало возможным выявление характерных белков, связанных с фенотипами псиллид [ 97 ] и стадиями развития [ 98 ]. Мета-анализ омиксов по молекулярным профилям цитрусовых хозяев также был бы полезен для выявления потенциальных молекулярных корреляций с предпочтениями T. erytreae . Этот подход был использован для изучения цитрусовых хозяев, толерантных к HLB [ 99 ].
Еще одной серьезной проблемой, выявленной в этом обзоре, была разрозненность данных и различные типы отчетности о взаимодействии T. erytreae с ее хозяевами. Следовательно, создание точной базы данных о потенциальных хозяевах T. erytreae имеет важное значение, поскольку это способствует принятию обоснованных решений относительно стратегий управления цитрусовыми, включая политику контроля, программы разведения, исследовательские направления и управление садами [ 14 ]. Кроме того, внедрение стандартизированной методологии для отчетности о популяциях T. erytreae в централизованном репозитории будет способствовать более полной и доступной базе знаний о популяциях, поведении и хозяевах листоблошки, повышая точность моделей прогнозирования, как это уже было разработано для D. citri и HLB [ 16 , 100 ].
Финансирование
Это исследование было проведено при финансовой поддержке проекта «Горизонт Европейского Союза 2020» в рамках соглашения о гранте исследовательского проекта № 817526 (Pre-HLB «Предотвращение эпидемий HLB для обеспечения выживания цитрусовых в Европе»). Томас Магальяйнс с благодарностью признает финансовую поддержку «Fundação para a Ciência e Tecnologia» (FCT — Португалия) в виде стипендии PhD 2020.07798.BD («Понимание предпочтений хозяев Trioza erytreae (Del Guercio) с помощью мультиомного подхода») ( https://doi.org/10.54499/2020.07798.BD ). Дополнительные средства были получены из программ, поддерживаемых FCT/MCTES (PIDDAC) для MED UIDB/05183/2020 (doi: 10.54499/UIDB/05183/2020) и UIDP/05183/2020 (doi: 10.54499/UIDP/05183/2020), для CHANGE LA/P/0121/2020 (doi: 10.54499/LA/P/0121/2020), для CIMO UIDB/00690/2020 (doi: 10.54499/UIDB/00690/2020) и UIDP/00690/2020 (doi: 10.54499/UIDP/00690/2020), а также для SusTEC LA/P/0007/2020 (doi: 10.54499/LA/P/0007/2020).
Заявление о доступности данных
Непригодный.
Благодарности
Мнения, выводы, заключения и рекомендации, высказанные в данном материале, принадлежат исключительно авторам.
Конфликты интересов
Авторы заявляют об отсутствии конфликта интересов.
Заявление об исправлении
Статья переиздана с небольшим исправлением читабельности Таблицы 2. Это изменение не влияет на научное содержание статьи.
Ссылки
References
1. FAO. Citrus Fruit Statistical Compendium 2020; FAO: Rome, Italy, 2021. [Google Scholar]
2. EPPO. First report of Diaphorina citri in Israel. Available online: https://gd.eppo.int/reporting/article-7262 (accessed on 26 April 2024).
3. EPPO. First Report of Diaphorina citri in Cyprus. Available online: https://gd.eppo.int/reporting/article-7660 (accessed on 26 April 2024).
4. Duarte, B.; Poeira, R.; Magalhães, T.; Paiva, P.; Soares, C.; Neto, L.; Marques, N.T.; Duarte, A. Current distribution of the African citrus psyllid Trioza erytreae in Portugal: Relation to climatic conditions. Acta Hortic. 2024, 1399, 423–428. [Google Scholar] [CrossRef]
5. Pérez-Otero, R.; Pérez-Turco, R.; Neto, J.; Fereres, A. The African psyllid Trioza erytreae Del Guercio (1918) is very sensitive to low relative humidity and high temperatures. Insects 2024, 15, 62. [Google Scholar] [CrossRef]
6. Alquézar, B.; Carmona, L.; Bennici, S.; Miranda, M.P.; Bassanezi, R.B.; Peña, L. Cultural management of huanglongbing: Current status and ongoing research. Phytopathology 2022, 112, 11–25. [Google Scholar] [CrossRef] [PubMed]
7. EPPO. Update on the Situation of Trioza erytreae in Portugal. Available online: https://gd.eppo.int/reporting/article-7215 (accessed on 15 May 2024).
8. Singerman, A.; Rogers, M.E. The economic challenges of dealing with citrus greening: The case of Florida. J. Integr. Pest. Manag. 2020, 11, 3–4. [Google Scholar] [CrossRef]
9. Ayres, A.J.; Belasque, J., Jr.; Bové, J.M. The experience with huanglongbing management in Brazil. Acta Hortic. 2015, 1065, 55–61. [Google Scholar] [CrossRef]
10. Bassanezi, R.B.; Lopes, S.A.; de Miranda, M.P.; Wulff, N.A.; Volpe, H.X.L.; Ayres, A.J. Overview of citrus huanglongbing spread and management strategies in Brazil. Trop. Plant Pathol. 2020, 45, 251–264. [Google Scholar] [CrossRef]
11. Li, M.; Nangong, Z. Precision trunk injection technology for treatment of huanglongbing (HLB)-affected citrus trees—A review. J. Plant Dis. Prot. 2022, 129, 15–34. [Google Scholar] [CrossRef]
12. Munir, S.; Ahmed, A.; Li, Y.; He, P.; Singh, B.K.; He, P.; Li, X.; Asad, S.; Wu, Y.; He, Y. The hidden treasures of citrus: Finding huanglongbing cure where it was lost. Crit. Rev. Biotechnol. 2021, 42, 634–649. [Google Scholar] [CrossRef] [PubMed]
13. Limayem, A.; Martin, E.M.; Shankar, S. Study on the citrus greening disease: Current challenges and novel therapies. Microb. Pathog. 2024, 192, 106688. [Google Scholar] [CrossRef]
14. Aidoo, O.F. The African citrus psyllid Trioza erytreae (Hemiptera: Triozidae): Biology, management, and its role as a vector of huanglongbing. Crop Prot. 2023, 172, 106348. [Google Scholar] [CrossRef]
15. Urbaneja-Bernat, P.; Hernández-Suárez, E.; Tena, A.; Urbaneja, A. Preventive measures to limit the spread of Trioza erytreae (Del Guercio) (Hemiptera: Triozidae) in mainland Europe. J. Appl. Entomol. 2020, 144, 553–559. [Google Scholar] [CrossRef]
16. Benhadi-Marín, J.; Fereres, A.; Pereira, J.A. Potential areas of spread of Trioza erytreae over mainland Portugal and Spain. J. Pest. Sci. 2022, 95, 67–78. [Google Scholar] [CrossRef]
17. Urbaneja-Bernat, P.; Pérez-Rodríguez, J.; Krüger, K.; Catalán, J.; Rizza, R.; Hernández-Suárez, E.; Urbaneja, A.; Tena, A. Host range testing of Tamarixia dryi (Hymenoptera: Eulophidae) sourced from South Africa for classical biological control of Trioza erytreae (Hemiptera: Psyllidae) in Europe. Biol. Control. 2019, 135, 110–116. [Google Scholar] [CrossRef]
18. EPPO. Trioza erytreae Occurs in Mainland Portugal. Available online: https://gd.eppo.int/reporting/article-5151 (accessed on 28 June 2024).
19. Reynaud, B.; Turpin, P.; Molinari, F.M.; Grondin, M.; Roque, S.; Chiroleu, F.; Fereres, A.; Delatte, H. The African citrus psyllid Trioza erytreae: An efficient vector of Candidatus Liberibacter asiaticus. Front. Plant Sci. 2022, 13, 1089762. [Google Scholar] [CrossRef]
20. Etienne, J.; Aubert, B. Biological control of psyllid vectors of greening disease on Reunion Island. IOCV Conf. Proc. 1980, 8, 118–121. [Google Scholar] [CrossRef]
21. Aubert, B. Trioza erytreae Del Guercio and Diaphorina citri Kuwayama (Homoptera: Psylloidea), the two vectors of citrus greening disease: Biological aspects and possible control strategies. Fruits 1987, 42, 149–162. [Google Scholar]
22. Aubert, B.; Quilici, S. Biological control of the African and Asian citrus psyllids (Homoptera: Psylloidea), through eulophid and encyrtid parasites (Hymenoptera: Chalcidoidea) in Reunion Island. IOCV Conf. Proc. 1984, 9, 100–108. [Google Scholar] [CrossRef]
23. Aidoo, O.F.; Tanga, C.M.; Mohamed, S.A.; Khamis, F.M.; Opisa, S.; Rasowo, B.A.; Kimemia, J.W.; Ambajo, J.; Sétamou, M.; Ekesi, S.; et al. The African citrus triozid Trioza erytreae Del Guercio (Hemiptera: Triozidae): Temporal dynamics and susceptibility to entomopathogenic fungi in East Africa. Int. J. Trop. Insect. Sci. 2021, 41, 563–573. [Google Scholar] [CrossRef]
24. Oliveira, D.F.; Benhadi-Marín, J.; Neto, J.; Sanz, L.; Garzo, E.; Aguiar, A.; Fereres, A.; Pereira, J.A. Kaolin particle films disrupt landing, settling behavior and feeding of Trioza erytrae on lemon plants. Pest. Manag. Sci. 2022, 78, 4753–4763. [Google Scholar] [CrossRef]
25. Van den Berg, M.A.; Anderson, S.H.; Deacon, V.E. Population studies of the citrus psylla, Trioza erytreae: Factors influencing population size. Phytoparasitica 1991, 19, 183–193. [Google Scholar] [CrossRef]
26. Catling, H.D. The bionomics of the south African citrus psylla Trioza erytreae (Del Guercio) (Homoptera: Psyllidae) 4. The influence of predators. J. Entomol. Soc. South. Afr. 1970, 33, 342–348. [Google Scholar]
27. Van den Berg, M.; Deacon, V.E. Dispersal of the citrus psylla, Trioza erytreae (Hemiptera: Triozidae) in the absence of its host plants. Phytophylactica 1988, 20, 361–368. [Google Scholar]
28. Van den Berg, M.A.; Deacon, V.E.; De Jager, K. Ecology of the citrus psylla, Trioza erytreae (Hemiptera: Triozidae) 1. Daily activities and habits of adults. Phytophylactica 1990, 22, 323–328. [Google Scholar]
29. Gottwald, T.R. Current epidemiological understanding of citrus huanglongbing. Annu. Rev. Phytopathol. 2010, 48, 119–139. [Google Scholar] [CrossRef]
30. Van Der Merwe, C.P. The citrus psylla (Trioza merwei, Pettey.). J. Dep. Agric. 1923, 7, 135–141. [Google Scholar]
31. de Carvalho, J.P.; Aguiar, A.M.F. Pragas dos Citrinos na Ilha da Madeira; Secretaria Regional da Agricultura Florestas e Pescas: Funchal, Portugal, 1997; ISBN 972-648-117.1. [Google Scholar]
32. Annecke, D.P.; Cilliers, C.J. The citrus psylla, Trioza erytreae (Del Guercio), and its parasites in South Africa. South Afr. J. Agric. Sci. 1963, 6, 187–192. [Google Scholar]
33. Moran, V.C. Preliminary observations on the choice of host plants by adults of the citrus psylla, Trioza erytreae (Del Guercio) (Homoptera: Triozidae). J. Entomol. Soc. South. Afr. 1968, 31, 404–410. [Google Scholar]
34. Moran, V.C. The development of the citrus psylla, Trioza erytreae (Del Guercio) (Homoptera: Psyllidae), on Citrus limon and four indigenous host plants. J. Entomol. Soc. South. Afr. 1968, 31, 391–402. [Google Scholar]
35. Bové, J. Keynote address: Heat-tolerant Asian HLB meets heat-sensitive African HLB in the Arabian Peninsula! Why? J. Citrus Pathol. 2014, 1, 1–78. [Google Scholar] [CrossRef]
36. Del Guercio, G. Note ed osservazioni di entomologia agraria. il cecidio delle foglie del limone ed il suo cecidozoo in eritrea. Agric. Colon. 1918, 12, 167–169. [Google Scholar]
37. Hernández-Suárez, E.; Suárez-Méndez, L.; Parrilla, M.; Arjona-López, J.M.; Hervalejo, A.; Arenas-Arenas, F.J. Feeding and oviposition behaviour of Trioza erytreae (Hemiptera: Triozidae) on different citrus rootstock material available in Europe. Insects 2021, 12, 623. [Google Scholar] [CrossRef]
38. Barkley, P.B.; Beattie, G.A.C. Contingency plans for HLB (huanglongbing) and his vectors in Australia. In Proceedings of the First International Workshop on Citrus Huanglongbing, Hermosillo, Mexico, 6 May 2008. [Google Scholar]
39. Aidoo, O.F.; Tanga, C.M.; Khamis, F.M.; Rasowo, B.A.; Mohamed, S.A.; Badii, B.K.; Salifu, D.; Sétamou, M.; Ekesi, S.; Borgemeister, C. Host suitability and feeding preference of the african citrus triozid Trioza erytreae (Del Guercio) (Hemiptera: Triozidae), natural vector of “Candidatus Liberibacter africanus”. J. Appl. Entomol. 2019, 143, 262–270. [Google Scholar] [CrossRef]
40. Aidoo, O.F.; Tanga, C.M.; Paris, T.M.; Allan, S.A.; Mohamed, S.A.; Khamis, F.M.; Sétamou, M.; Borgemeister, C.; Ekesi, S. Size and shape analysis of Trioza erytreae Del Guercio (Hemiptera: Triozidae), vector of citrus huanglongbing disease. Pest. Manag. Sci. 2019, 75, 760–771. [Google Scholar] [CrossRef] [PubMed]
41. Kalyebi, A.; Aisu, G.; Ramathani, I.; Ogwang, J.; McOwen, N.; Russell, P. Detection and identification of etiological agents (Liberibacter spp.) associated with citrus greening disease in Uganda. Uganda J. Agric. Sci. 2016, 16, 43–54. [Google Scholar] [CrossRef][Green Version]
42. Abate, T. The identity and bionomics of insect vectors of tristeza and greening diseases of citrus in Ethiopia. Trop. Pest Manag. 1988, 34, 19–23. [Google Scholar] [CrossRef]
43. Quintana-González De Chaves, M.; Teresani, G.R.; Hernández-Suárez, E.; Bertolini, E.; Moreno, A.; Fereres, A.; Cambra, M.; Siverio, F. “Candidatus Liberibacter solanacearum” is unlikely to be transmitted spontaneously from infected carrot plants to citrus plants by Trioza erytreae. Insects 2020, 11, 514. [Google Scholar] [CrossRef] [PubMed]
44. Zheng, L.; Xu, Q.; Gong, G.; Liao, Y.; Yu, M.; Shabala, S.; Chen, W.; Wu, W. Nicotiana tabacum as a dead-end trap for adult diaphorina citri: A potential biological tactic for protecting citrus orchards. Front. Plant Sci. 2023, 13, 1081663. [Google Scholar] [CrossRef]
45. Ameline, A.; Karkach, A.; Denoirjean, T.; Grondin, M.; Molinari, F.; Turpin, P.; Delatte, H.; Reynaud, B. Bacterial plant pathogens affect the locomotor behavior of the insect vector: A case study of Citrus volkameriana–Triozae erytreae–Candidatus Liberibacter asiaticus system. Insect Sci. 2023, 31, 901–910. [Google Scholar] [CrossRef] [PubMed]
46. Magalhães, T.; Dandlen, S.A.; Anjos, L.; Power, D.M.; Pereira, J.A.; Duarte, A.; Marques, N.T. Comparing the response of Citrus × limon and Citrus × sinensis to Trioza erytreae infestation using a proteomic approach. Acta Hortic. 2024, 1399, 379–386. [Google Scholar] [CrossRef]
47. Moran, V.C.; Brown, R.P. The antennae, host plant chemoreception and probing activity of the citrus psylla, Trioza erytreae (Del Guercio) (Homoptera: Psyllidae). J. Entomol. Soc. South Afr. 1973, 36, 191–202. [Google Scholar]
48. Tamesse, J.L.; Messi, J. Réceptivité à Trioza erytreae (Del Guercio) de variétés d’ agrumes au Cameroun. Fruits 2000, 55, 389–400. [Google Scholar]
49. Tamesse, J.L.; Messi, J. Facteurs influençant la dynamique des populations du psylle Africain des agrumes Trioza erytreae Del Guercio (Hemiptera: Triozidae) au Cameroun. Int. J. Trop. Insect Sci. 2004, 24, 213–227. [Google Scholar] [CrossRef]
50. Cook, G.; Maqutu, V.Z.; Van Vuuren, S.P. Population dynamics and seasonal fluctuation in the percentage infection of Trioza erytreae with “Candidatus” Liberibacter africanus, the african citrus greening pathogen, in an orchard severely infected with African greening and transmission by field-collected Trioza erytreae. Afr. Entomol. 2014, 22, 127–135. [Google Scholar] [CrossRef]
51. Tamesse, J.L.; Messi, J. Incidence de Trioza erytreae (Del Guercio) (Homoptera: Triozidae), psylle vecteur du greening sur la sensibilité des plantules d’agrumes dans une pépinière au Cameroun. Insect Sci. Appl. 2002, 22, 97–103. [Google Scholar] [CrossRef]
52. Benhadi-Marín, J.; Garzo, E.; Moreno, A.; Pereira, J.A.; Fereres, A. Host plant preference of Trioza erytreae on lemon and bitter orange plants. Arthropod Plant Interact. 2021, 15, 887–896. [Google Scholar] [CrossRef]
53. Moran, V.C.; Buchan, P.R. Oviposition by the citrus psylla, Trioza erytreae (Homoptera: Psyllidae), in relation to leaf hardiness. Entomol. Exp. Appl. 1975, 18, 96–104. [Google Scholar] [CrossRef]
54. Moran, V.C.; Blowers, J.R. On the biology of the South African citrus psylla, Trioza erytreae (Del Guercio) (Homoptera: Psyllidae). J. Entomol. Soc. South. Afr. 1967, 30, 96–106. [Google Scholar]
55. Aidoo, O.F.; Tanga, C.M.; Azrag, A.G.A.; Mohamed, S.A.; Khamis, F.M.; Rasowo, B.A.; Ambajo, J.; Sétamou, M.; Ekesi, S.; Borgemeister, C. Temperature-based phenology model of African citrus triozid (Trioza erytreae Del Guercio): Vector of citrus greening disease. J. Appl. Entomol. 2022, 146, 88–97. [Google Scholar] [CrossRef]
56. Quintana-González de Chaves, M.; Montero-Gomez, N.; Álvarez-Acosta, C.; Hernández-Suárez, E.; Hervalejo, A.; Arjona-López, J.M.; Arenas-Arenas, F.J. The combination of citrus rootstock and scion cultivar influences Trioza erytreae (Hemiptera: Triozidae) survival, preference choice and oviposition. Insects 2024, 15, 363. [Google Scholar] [CrossRef]
57. Van den Berg, M.A.; Deacon, V.E.; Thomas, C.D. Ecology of the citrus psylla, Trioza erytreae (Hemiptera: Triozidae). 4. settling and general behaviour of nymphs. Phytophylactica 1991, 23, 201–206. [Google Scholar]
58. Catling, H.D. The bionomics of the South African citrus psylla, Trioza erytreae (Del Guercio) (Homoptera: Psyllidae) I. The influence of the flushing rhythm of citrus and factors which regulate flushing. J. Entomol. Soc. South. Afr. 1969, 32, 191–208. [Google Scholar]
59. Catling, H.D. The bionomics of the South African citrus psylla, Trioza erytreae (Del Guercio) (Homoptera: Psyllidae) 5. The influence of host plant quality. J. Entomol. Soc. South. Afr. 1971, 34, 381–391. [Google Scholar]
60. Samways, M.J.; Manicom, B.Q. Immigration, frequency distributions and dispersion patterns of the psyllid Trioza erytreae (Del Guercio) in a citrus orchard. J. Appl. Ecol. 1983, 20, 463. [Google Scholar] [CrossRef]
61. Catling, H.D.; Atkinson, P.R. Spread of greening by Trioza erytreae (Del Guercio) in Swaziland. IOCV Conf. Proc. 1974, 6, 6. [Google Scholar] [CrossRef]
62. Van den Berg, M. Developmental biology and population studies on the citrus psylla Trioza erytreae (Del Guercio) (Hemiptera: Triozidae). Fruits 1992, 47, 583–589. [Google Scholar]
63. Catling, H.D.; Annecke, D.P. Ecology of citrus psylla in the Letaba district of Northern Transvaal. S. Afr. Citrus J. 1968, 410, 8–17. [Google Scholar]
64. Catling, H.D. The bionomics of the South African citrus psylla, Trioza erytreae (Del Guercio) (Homoptera: Psyllidae). 6. Final population studies and discussion of population dynamics. J. Entomol. Soc. South. Afr. 1972, 35, 235–251. [Google Scholar]
65. Antwi-Agyakwa, A.K.; Yusuf, A.A.; Pirk, C.W.W.; Mohamed, S.A.; Ekesi, S.; Torto, B. Exploring non-host plant-based management strategy with lemongrass, garlic and guava volatiles for the African citrus triozid. J. Appl. Entomol. 2021, 145, 757–766. [Google Scholar] [CrossRef]
66. Antwi-Agyakwa, A.K.; Fombong, A.T.; Deletre, E.; Ekesi, S.; Yusuf, A.A.; Pirk, C.; Torto, B. Lemon terpenes influence behavior of the African citrus triozid (Hemiptera: Triozidae). J. Chem. Ecol. 2019, 45, 934–945. [Google Scholar] [CrossRef]
67. Hollis, D. Afrotropical jumping plant lice of the family Triozidae (Homoptera: Psylloidea). Bull. Br. Mus. Nat. Hist. Entomol. 1984, 49, 1–102. [Google Scholar]
68. Van den Berg, M.A.; Deacon, V.E.; Steenekamp, P.J. Dispersal within and between citrus orchards and native hosts, and nymphal mortality of citrus psylla, Trioza erytreae (Hemiptera: Triozidae). Agric. Ecosyst. Environ. 1991, 35, 297–309. [Google Scholar] [CrossRef]
69. Van den Berg, M.A. The citrus pyslla, Trioza erytreae (Del Guercio) (Hemiptera: Triozidae): A review. Agric. Ecosyst. Environ. 1990, 30, 171–194. [Google Scholar] [CrossRef]
70. Cocuzza, G.E.M.; Alberto, U.; Hernández-Suárez, E.; Siverio, F.; Di Silvestro, S.; Tena, A.; Carmelo, R. A review on Trioza erytreae (African citrus psyllid), now in mainland Europe, and its potential risk as vector of huanglongbing (HLB) in citrus. J. Pest. Sci. 2017, 90, 1–17. [Google Scholar] [CrossRef]
71. Van den Berg, M.A. Effects of citrus cultivars and reduced irrigation on availability of new growth for citrus psylla breeding. Fruits 1986, 41, 597–604. [Google Scholar]
72. Agustí, M.; Zaragoza, S.; Bleiholder, H.; Buhr, L.; Hack, H.; Klose, R.; Staub, R. Escala BBCH para la Descripción de los Estadios Fenológicos del Desarrollo de los Agrios (Gén. Citrus). Available online: https://redivia.gva.es/handle/20.500.11939/7875 (accessed on 18 July 2022).
73. Matias, P.; Barrote, I.; Azinheira, G.; Continella, A.; Duarte, A. Citrus pruning in the Mediterranean climate: A review. Plants 2023, 12, 3360. [Google Scholar] [CrossRef] [PubMed]
74. White, T.C.R. Uptake of water by eggs of Cardiaspina densitexta (Homoptera: Psyllidae) from leaf of host plant. J. Insect. Physiol. 1968, 14, 1669–1683. [Google Scholar] [CrossRef]
75. Mc Daniel, J.R.; Moran, V.C. The parasitoid complex of the citrus psylla Trioza erytreae (Del Guercio) [Homoptera: Psyllidae]. Entomophaga 1972, 17, 297–317. [Google Scholar] [CrossRef]
76. Cifuentes-Arenas, J.C.; de Oliveira, H.T.; Raiol-Júnior, L.L.; de Carvalho, E.V.; Kharfan, D.; Creste, A.L.; Gastaminza, G.; Salas, H.; Bassanezi, R.B.; Ayres, A.J.; et al. Impacts of huanglongbing on fruit yield and quality and on flushing dynamics of Sicilian lemon trees. Front. Plant. Sci. 2022, 13, 1005557. [Google Scholar] [CrossRef] [PubMed]
77. Van den Berg, M.A.; Anderson, S.H.; Deacon, V.E. Population studies of the citrus psylla, Trioza erytreae: Factors influencing dispersal. Phytoparasitica 1991, 19, 283–289. [Google Scholar] [CrossRef]
78. Jacinto, C.; Matias, P.; Oliveira, C.; Duarte, A. Effect of heading cuts on branch growth of ‘Encore’ mandarin. Acta Hortic. 2024, 1399, 241–246. [Google Scholar] [CrossRef]
79. de Carvalho, D.U.; Girardi, E.A.; Pacheco, C.d.A.; Primiano, I.V.; Kharfan, D.; Moreira, A.S.; Laranjeira, F.F.; Bassanezi, R.B. Topping sweet orange trees as Diaphorina citri bait on the farm edge for huanglongbing management: Opportunities and limitations. Sci. Hortic. 2024, 338, 113612. [Google Scholar] [CrossRef]
80. Primo-Millo, E.; Agustí, M. Vegetative growth. In The Genus Citrus; Elsevier: Amsterdam, The Netherlands, 2020; pp. 193–217. [Google Scholar]
81. Micheloud, N.G.; Castro, D.C.; Buyatti, M.A.; Gabriel, P.M.; Gariglio, N.F. Factors affecting phenology of different citrus varieties under the temperate climate conditions of Santa Fe, Argentina. Rev. Bras. Frutic. 2018, 40, e315. [Google Scholar] [CrossRef]
82. Paiva, P.E.B.; Cota, T.; Neto, L.; Soares, C.; Tomás, J.C.; Duarte, A. Water vapor pressure deficit in Portugal and implications for the development of the invasive African citrus psyllid Trioza erytreae. Insects 2020, 11, 229. [Google Scholar] [CrossRef]
83. Benhadi-Marín, J.; Fereres, A.; Pereira, J.A. A model to predict the expansion of Trioza erytreae throughout the Iberian Peninsula using a pest risk analysis approach. Insects 2020, 11, 576. [Google Scholar] [CrossRef]
84. Aidoo, O.F.; Tanga, C.M.; Mohamed, S.A.; Rasowo, B.A.; Khamis, F.M.; Rwomushana, I.; Kimani, J.; Agyakwa, A.K.; Daisy, S.; Sétamou, M.; et al. Distribution, degree of damage and risk of spread of Trioza erytreae (Hemiptera: Triozidae) in Kenya. J. Appl. Entomol. 2019, 143, 822–833. [Google Scholar] [CrossRef]
85. Bové, J. Huanglongbing: A destructive, newly-emerging, century-old disease of citrus. J. Plant Pathol. 2006, 1, 7–37. [Google Scholar]
86. Uzun, A.; Yesiloglu, T. Genetic diversity in citrus. In Genetic Diversity in Plants; InTech: Rijeka, Croatia, 2012. [Google Scholar] [CrossRef]
87. Pollard, D.G. The use of polyporus for the investigation of stylet behaviour in the Hemiptera. Entomol. Exp. Appl. 1971, 14, 283–296. [Google Scholar] [CrossRef]
88. Jones, S.E.; Killiny, N. Influence of rootstock on the leaf volatile organic compounds of citrus scion is more pronounced after the infestation with Diaphorina citri. Plants 2021, 10, 2422. [Google Scholar] [CrossRef] [PubMed]
89. Valterová, I.; Nehlin, G.; Borg-Karlson, A.K. Host plant chemistry and preferences in egg-laying Trioza apicalis (Homoptera, Psylloidea). Biochem. Syst. Ecol. 1997, 25, 477–491. [Google Scholar] [CrossRef]
90. Pullock, D.A.; Krüger, K.; Manrakhan, A.; Yusuf, A.A.; Weldon, C.W. Addition of selected plant-derived semiochemicals to yellow sticky traps does not improve citrus psyllid captures. J. Chem. Ecol. 2024, 50, 701–713. [Google Scholar] [CrossRef] [PubMed]
91. Hall, D.G.; Gottwald, T.R.; Nguyen, N.C.; Ichinose, K.; Le, Q.D.; Beattie, G.A.C.; Stover, E. Greenhouse investigations on the effect of guava on infestations of Asian citrus psyllid in grapefruit. Proc. Fla. State Hort. Soc. 2008, 121, 104–109. [Google Scholar]
92. Gottwald, T.R.; Hall, D.G.; Beattie, G.A.C.; Ichinose, K.; Nguyen, M.C.; Le, Q.D.; Bar-Joseph, M.; Lapointe, S.; Stover, E.; Parker, P.E.; et al. Investigations of the effect of guava as a possible tool in the control/management of huanglongbing. IOCV Conf. Proc. 2010, 17, 2313–5123. [Google Scholar] [CrossRef]
93. Russell, C.W.; Pelz-Stelinski, K.S. Development of an artificial diet and feeding system for juvenile stages of the Asian citrus psyllid, Diaphorina citri. Entomol. Exp. Appl. 2015, 154, 171–176. [Google Scholar] [CrossRef]
94. Barah, P.; Bones, A.M. Multidimensional approaches for studying plant defence against insects: From ecology to omics and synthetic biology. J. Exp. Bot. 2015, 66, 479–493. [Google Scholar] [CrossRef] [PubMed]
95. Nehela, Y.; Hijaz, F.; Elzaawely, A.A.; El-Zahaby, H.M.; Killiny, N. Citrus phytohormonal response to Candidatus Liberibacter asiaticus and its vector Diaphorina citri. Physiol. Mol. Plant. Pathol. 2018, 102, 24–35. [Google Scholar] [CrossRef]
96. Sun, X.; Yu, T.; Bin, M.; Hu, C.; Bi, F.; Peng, X.; Yi, G.; Zhang, X. Transcriptomic analysis reveals the defense mechanisms of citrus infested with Diaphorina citri. Hortic. Plant J. 2022, 9, 450–462. [Google Scholar] [CrossRef]
97. Hosseinzadeh, S.; Higgins, S.A.; Ramsey, J.; Howe, K.; Griggs, M.; Castrillo, L.; Heck, M. Proteomic polyphenism in color morphotypes of Diaphorina citri, insect vector of citrus greening disease. J. Proteome. Res. 2021, 20, 2851–2866. [Google Scholar] [CrossRef]
98. El-Shesheny, I.; El-Hawary, I.; Mesbah, I.; Killiny, N. Comparative proteomic analysis between fifth-instar nymphs and adults of Asian citrus psyllid Diaphorina citri. Physiol. Entomol. 2016, 41, 162–184. [Google Scholar] [CrossRef]
99. Rawat, N.; Kiran, S.P.; Du, D.; Gmitter, F.G.; Deng, Z. Comprehensive meta-analysis, co-expression, and MiRNA nested network analysis identifies gene candidates in citrus against huanglongbing disease. BMC Plant Biol. 2015, 15, 184. [Google Scholar] [CrossRef] [PubMed]
100. Galvañ, A.; Bassanezi, R.B.; Luo, W.; Vanaclocha, P.; Vicent, A.; Lázaro, E. Risk-based regionalization approach for area-wide management of HLB vectors in the Mediterranean basin. Front. Plant Sci. 2023, 14, 1256935. [Google Scholar] [CrossRef]
Trioza erytreae (Del Guercio, 1918) и взаимодействие с
хозяевами: обзор
Томас Магальес1,2, Амилкар Дуарте1, Хосе
Альберто Перейра2 и Наталия Т. Маркес1
1 MED — Средиземноморский институт сельского хозяйства, окружающей среды и развития и CHANGE — Институт глобальных изменений и устойчивого развития, Университет Алгарве, кампус Гамбелас, 8005-139 Фару, Португалия
2 CIMO, LA SusTEC, Политехнический институт Брагансы, Кампус Санта-Аполония, 5300-253 Браганса, Португалия
Magalhães, T.; Duarte, A.; Pereira, J.A.; Marques, N.T. Trioza erytreae (Del Guercio, 1918) and the Interaction with Its Hosts: A Review. Agriculture2025, 15, 101.
Перевод статьи «Trioza erytreae (Del Guercio, 1918) and the Interaction with Its Hosts: A Review» авторов Magalhães, T.; Duarte, A.; Pereira, J.A.; Marques, N.T., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык



















































































































Комментарии (0)