Влияние органических удобрений на рост покровных культур, поглощение азота и содержание азота в почве
Покровные культуры выполняют одновременно множество функций, повышая устойчивость сельского хозяйства по сравнению с оставлением земель под паром в пахотных системах. В климате с обильными осадками важная роль покровных культур заключается в улавливании питательных веществ, предотвращая их попадание в окружающую среду. Во многих регионах с интенсивным животноводством осенью вносят органические удобрения, оставляя землю под паром на зиму, а весной высаживают товарную культуру. Такая практика длительного парования в сочетании с недостаточным согласованием внесения питательных веществ с потребностями товарных культур в питательных веществах ещё больше увеличивает вероятность значительных потерь питательных веществ, в частности азота (N), в окружающую среду посредством вымывания и улетучивания.
Аннотация
Следовательно, если бы покровные культуры могли реагировать на эти питательные вещества за счёт ускоренного роста биомассы и усвоения питательных веществ, они могли бы снизить эти потери. Поэтому важно выбрать правильный вид, который будет расти достаточно хорошо, чтобы обеспечить эти потенциальные преимущества. В регионе проведения данного исследования, Северной Ирландии, было проведено мало исследований, посвящённых не только изучению оптимальных видов покровных культур, но и тому, какие виды могут реагировать на дополнительные питательные вещества в навозной жиже. Это исследование включает два эксперимента, проведенных в последовательные годы, включая комбинацию трех факторов: виды покровных культур (пять видов; смесь и контроль, представляющий собой пар); свиной навоз. Следовательно, было обнаружено, что при хороших условиях выращивания, в опытном году 2018/19, такие виды, как кормовой рапс и пропашной редис, могли значительно (p< 0,05) увеличить количество азота, содержащегося в их биомассе и почве (до 15 см) на 70 и 63% соответственно, в ответ на навозную жижу. Альтернативно, когда навозную жижу вносили на паровые земли (традиционная практика), низкие количества (14 и 0% в два года испытаний соответственно) азота из навозной жижи были обнаружены в биомассе сорняков или сохранены в почве. Это свидетельствует о большой потере азота из системы. Таким образом, интеграция отзывчивых покровных культур с жидким навозом является лучшим методом сокращения потерь азота, чем традиционная практика, если внесение жидкого навоза нельзя отложить.
1. Введение
В умеренных регионах земля, оставленная под паром на длительное время после сбора урожая коммерческой культуры, подвержена потере вымываемых питательных веществ из-за избыточных осадков, смывающих их ниже ризосферы [ 1 ]. Аллингем и др. [ 2 ] обнаружили в Англии, что в среднем 65 кг/га нитрата (NO3− ) вымывается ежегодно из пахотных земель, что составляет 25% от поступления N. Кроме того, органические удобрения, вносимые осенью, увеличивают этот риск [ 3 , 4 ] из-за плохой синхронизации поставки и спроса на питательные вещества. Эта потеря питательных веществ представляет собой не только экологическую проблему, но и экономические потери для фермера [ 5 ]. Покровные культуры иммобилизуют питательные вещества в своей биомассе и, таким образом, защищают их [ 6 ]. Это может снизить вымывание N в среднем на 43% [ 7 ]. В случаях, когда органические удобрения вносились осенью, покровные культуры могли бы стать стратегией для снижения плохой синхронизации спроса и предложения питательных веществ. Исследования показали, что интеграция покровных культур и органических удобрений может увеличить биомассу покровных культур [ 6 , 8 ]. В свою очередь, это уменьшило выщелачивание нитратов по сравнению с внесением навоза в чистый пар [ 3 , 8 ]. Однако после того, как биомасса была возвращена в почву, небобовые виды не смогли в достаточной степени минерализовать N и поставлять его товарной культуре по сравнению с бобовыми [ 6 , 9 ]. В результате это не компенсировало потребность товарной культуры в N. Эверетт и др. [ 3 ] заявили, что региональные климатические различия влияют на то, как покровные культуры растут и реагируют, а это означает, что для максимизации их эффекта необходимы региональные исследования этой практики [ 10 ].
Основным ограничением для усвоения покровных культур и их прибыльной интеграции в севообороты является неопределенность относительно влияния на последующую урожайность культур и того, можно ли снизить потребность в неорганических питательных веществах, что может помочь сократить затраты. Абдалла и др. [ 1 ] обнаружили в метаанализе, что покровные культуры снизили урожайность основных культур на 4%. Это было в первую очередь из-за иммобилизации N из-за недостаточного уровня минерализации N из остатков покровной культуры для снабжения коммерческой культуры [ 11 , 12 ]. Однако, когда небобовые и бобовые виды покровных культур выращивались вместе, было обнаружено, что они повышали урожайность на 13% [ 1 ] из-за улучшения круговорота N [ 13 ]. Это привело к большему количеству азота из-за снижения потерь N в течение зимы и более высокой скорости высвобождения N [ 13 ]. В качестве альтернативы, бобовые могут повысить уровень азота в почве посредством биологической фиксации [ 14 ]. Количество фиксированного N пропорционально произведенной биомассе [ 15 ] и выбору видов [ 14 ]. Однако единственная покровная культура из бобовых не снижает риск вымывания, а, как было обнаружено, даже увеличивает вымывание [ 7 ]. Кроме того, было обнаружено, что их рост значительно варьируется в зависимости от региона, при этом исследования показывают, что они хорошо растут в таких климатических регионах, как Швейцария [ 14 , 16 ], Норвегия, Швеция, Дания [ 7 ] и Франция [ 17 , 18 ]. Однако в климатических условиях Ирландии Роарти и др. [ 19 ] обнаружили, что покровная культура из гороха ( Pisum sativum L.) продемонстрировала ограниченный рост, при этом за один год из четырех лет испытаний было зафиксировано только более 1 т/га сухого вещества.
Уровень услуг, предоставляемых покровными культурами, такой как сокращение вымывания N, круговорот питательных веществ, влияние на биологию почвы и подавление сорняков, в значительной степени зависит от произведенной биомассы [ 20 , 21 , 22 , 23 , 24 ]. Поэтому высокое производство биомассы должно быть целью покровных культур. Хотя, Хантер и др. [ 23 ] обнаружили, что покровные культуры с высокой биомассой (<2,5 т/га) могут быть пагубны для урожайности последующей коммерческой культуры. Эти авторы обнаружили, что урожайность кукурузы отрицательно коррелирует с соотношением C:N в остатках покровной культуры, где соотношение менее 25 не снижает урожайность. Аналогичным образом, исследование показало, что соотношение C:N менее 24 имеет решающее значение для покровных культур для обеспечения N следующей культуры [ 25 ]. Соотношение C:N можно регулировать путем выбора видов [ 26 ], даты окончания [ 27 ] и даты посадки [ 12 ]. В контролируемых условиях было показано, что использование жидкого навоза может не только улучшить рост покровных культур и общее усвоение питательных веществ, но и улучшить качество остатков за счёт снижения соотношения C:N [ 28 ]. Однако это не было исследовано в ходе полевых экспериментов в регионе Великобритании.
Кей и соавторы [ 21 ] обнаружили, что ограниченное число отдельных видов эффективно как для снижения вымывания азота, так и для увеличения его поступления в последующую культуру. Крайне важно, чтобы минерализация азота, накопленного покровными культурами, синхронизировалась с потребностями в азоте товарных культур, что способствует снижению уровня неорганического азота в товарной культуре [ 10 ]. Поэтому целесообразно проводить больше исследований видов, их взаимодействия с органическими удобрениями и их влияния на потребность в азотных удобрениях для последующей товарной культуры.
Цель
Целью данного исследования является изучение влияния свиного навоза на рост и поглощение азота пятью видами покровных культур и их смесью. Для сравнения с типичной практикой будет включен контрольный пар. У покровных культур будут измерены индекс листовой поверхности (LAI), биомасса и поглощение азота в сравнении с контрольным паром.
2. Материалы и методы
2.1 Экспериментальный дизайн
Экспериментальный план с разделенным участком был создан с использованием процедуры AGHIERARCHICAL [ 29 ] с 4 блоками и 3 примененными обработками. Обработки включали навозную жижу (весь участок), виды (подучасток) и азот (подучасток), и каждая имела 2, 7 и 2 уровня соответственно. 2 уровня для навозной жижи либо применялись, либо не применялись и далее сообщаются как S и NS соответственно. Было 5 видов покровных культур, смесь всех и контроль, который представлял собой пар. Неорганический азот вносился в последующую культуру, посеянную после покровных культур, которой был яровой ячмень, в двух нормах 0 кг/га или 70 кг/га. В этой статье исследовались только рост и усвоение питательных веществ во время роста покровной культуры, в соответствии с чем подучастки, которые должны были получить обработку, действовали как дополнительные реплики покровных культур. Таким образом, весь участок был повторен 4 раза, а подучасток — 8 раз.
2.2 Местоположение
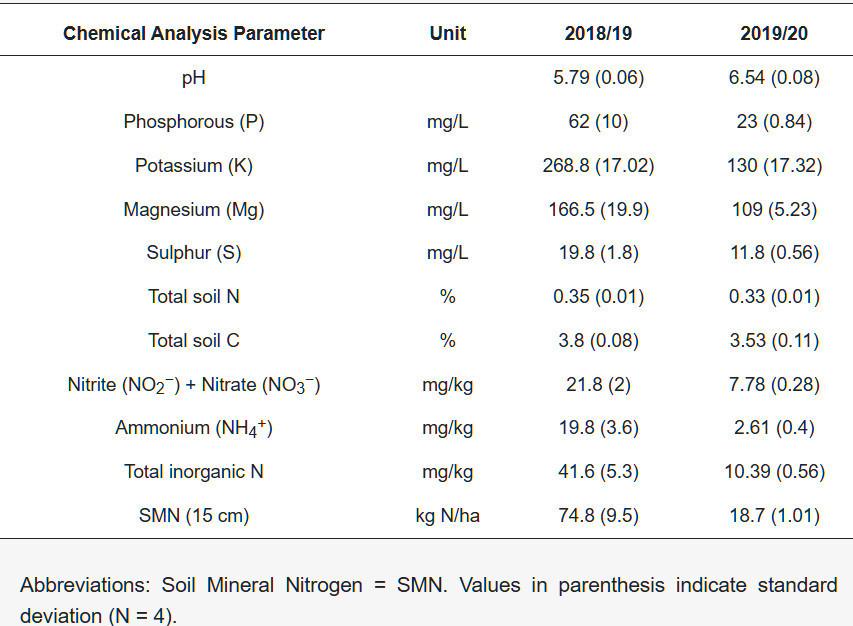
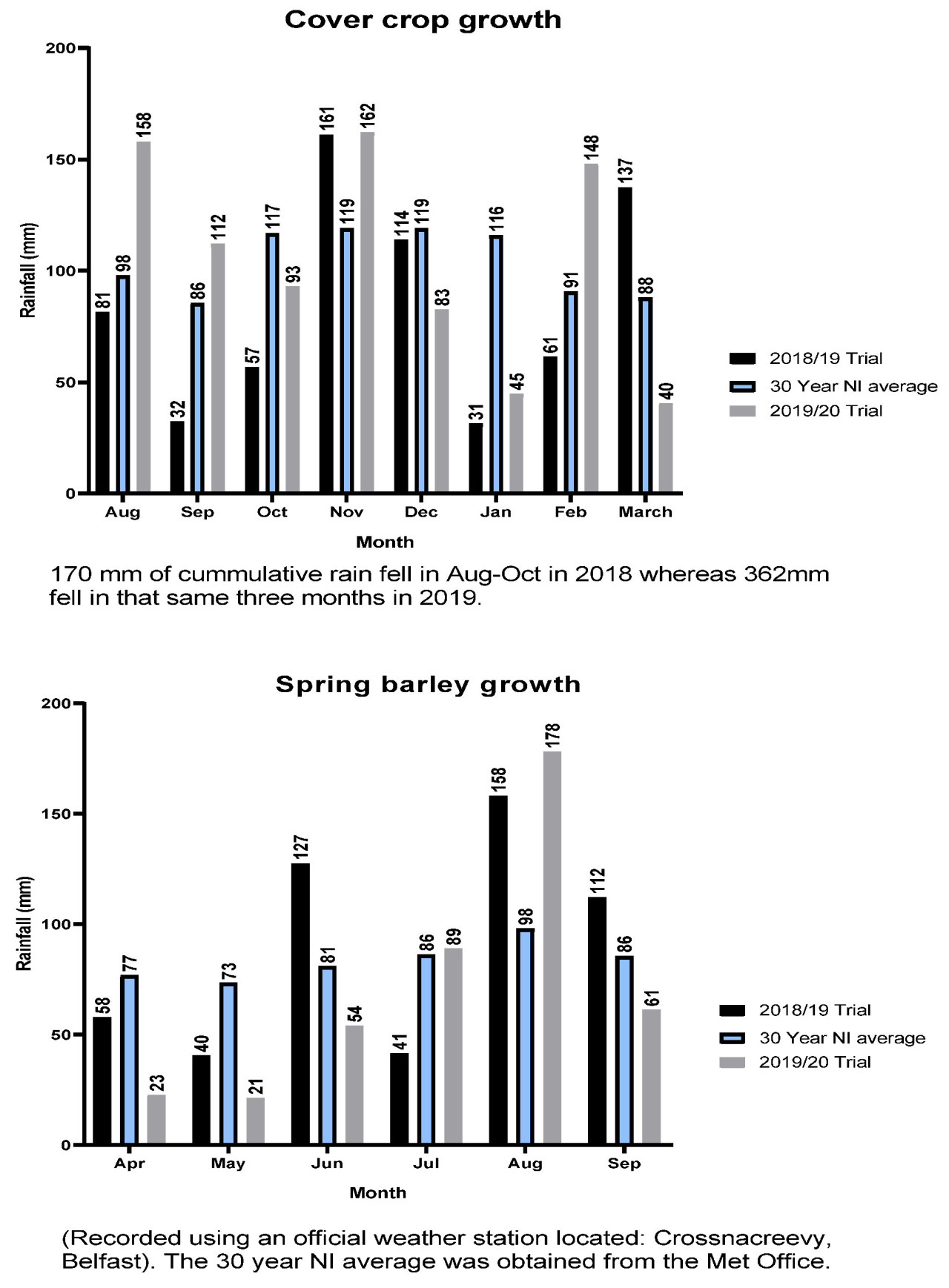
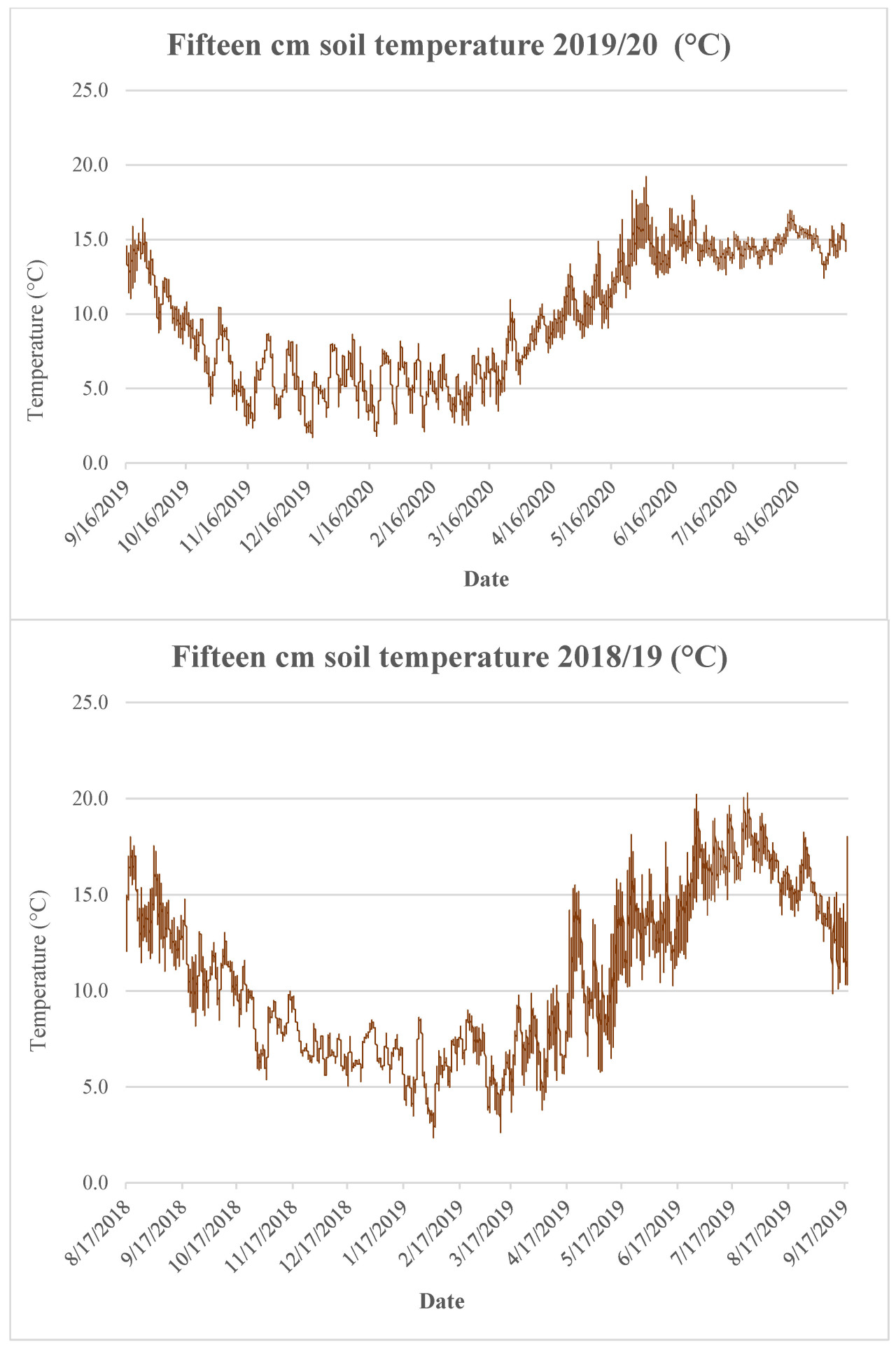
Испытания проводились в Хиллсборо, графство Даун, Северная Ирландия (широта 54.445117, долгота −6.096430) на разных полях каждый год, но в непосредственной близости. Данные об осадках показаны на рисунке A1 , температура почвы — на рисунке A2 , а суточные минимальные/максимальные температуры — на рисунке A3 . Почва представляла собой суглинок до 30 см с распределением частиц 44,5% песка, 29,9% ила и 25,7% глины в 2018/19 году, с глубокой глиной впоследствии. Участок, используемый в 2019/20 году, имел среднее распределение 44,9% песка, 32,1% ила и 23,0% глины, с глубокой глиной впоследствии. Водоудерживающая способность почв составляла 70% и 71% для участков 1 и 2 соответственно. Химический анализ почвы, проведенный до начала испытаний с каждого блока 2 августа 2018 года и 10 августа 2019 года, представлен в таблице 1 .
2.3 Выбор вида
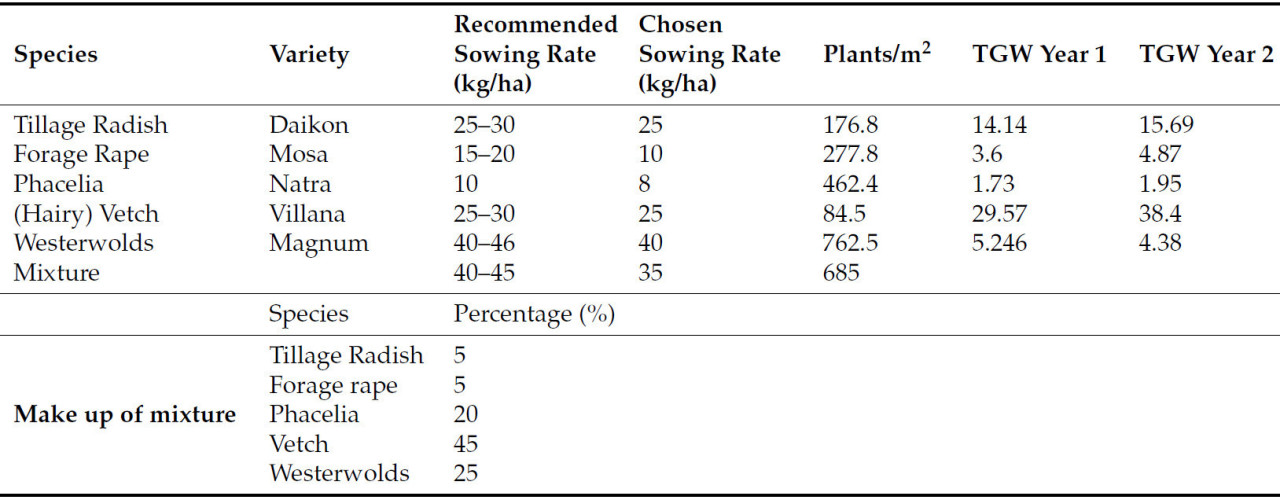
Рапс кормовой ( B. napus L.), редис посевной ( R. sativus L.), вика ( Vicia villosa L.), фацелия ( Phacelia tanacetifolia L.) и райграс многоцветковый ( Lolium multiflorum L.) высевали в соответствии с рекомендациями RAGT Seeds (Уилсон, 2018, личное сообщение). Плотность посева растений поддерживалась путём расчёта количества растений на м² и использования веса тысячи зёрен ( таблица A1 ). Доля каждого вида в смеси представлена в таблице A1 .
2.4. Выращивание
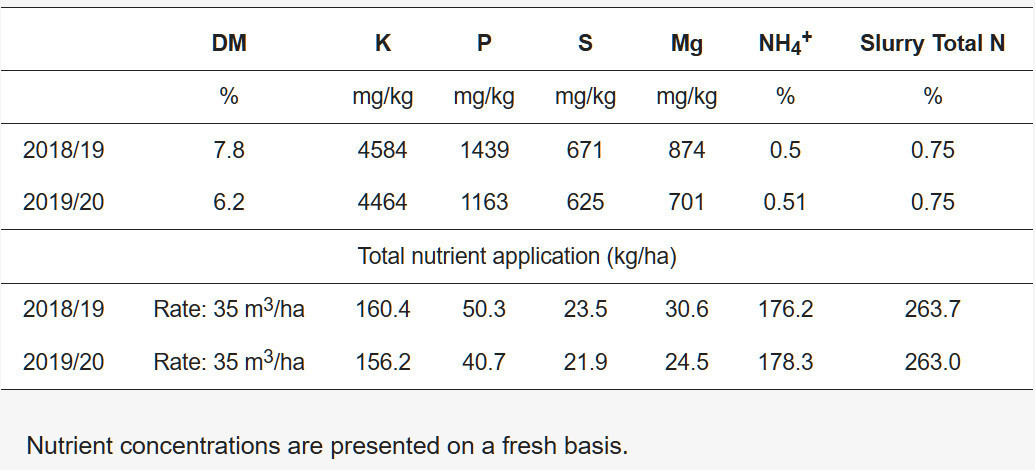
Испытание проводилось после посева цельнозернового ярового ячменя в 2018/19 году и после уборки озимого ячменя в 2019/20 году. Навозную жижу откорма свиней вносили по стерне в количестве 35 м 3 /га ( таблица 2 ) с помощью цистерны для навоза и капельной планки, а расход измеряли с помощью датчика расхода. Во второй год испытаний (2019/20 год) навозную жижу вносили позже из-за влажных условий. Опытный участок был обработан дисковым культиватором Lemken Heliodor (Alpen, Германия) перед посевом 14 августа 2019 года. 10 сентября 2019 года в связи с влажными условиями опытный участок был обработан отвальным плугом (Kverneland, Мерсисайд, Великобритания), после чего проведена дополнительная обработка почвы пружинной бороной (Porter, Лисберн, Великобритания) для создания семенного ложа. Покровные культуры были посеяны сразу после культивации с помощью сеялки Wintersteiger (Эссекс, Великобритания). Площадь заложенной делянки составила 1,68 × 16 м.
2.5. Отбор проб
2.5.1. Измерения септометра
Показания септометра снимались ежемесячно с помощью AccuPAR LP-80 (METER Group, Decagon Devices Inc., Пуллман, штат Вашингтон, США) для определения индекса листовой поверхности (LAI). Септометр использует два датчика для измерения интенсивности фотосинтетически активной радиации (ФАР) в диапазоне волн 400 нм и 700 нм, падающей на растение, и количества, падающего на землю. Это позволяет автоматически рассчитывать LAI, который представляет собой площадь зеленой листовой поверхности на м² , по уравнениям, задокументированным в руководстве по цептометру. На каждую дату отбора проб регистрировали четыре показания с каждого участка. В 2019/20 году первое ежемесячное показание септометра было отложено до 17 ноября по сравнению с 17 октября предыдущего года, чтобы учесть более поздний посев.
2.5.2 Отбор проб биомассы покровных культур
Биомасса покровных культур отбиралась с площади 0,5 м2 10 декабря 2018 г. и 9 декабря 2019 г., а окончательный сбор биомассы перед прекращением выращивания покровных культур производился 4 февраля 2019 г. и 11 марта 2020 г. Корни крестоцветных (редька пахотная и рапс кормовой) извлекались и промывались водой, так как масса их стержневых корней составляет большую долю от общей биомассы [ 17 , 30 ]. Сорняки отделялись от биомассы покровных культур. Различные фракции покровных культур отделялись (надземная биомасса покровных культур, корни покровных культур и сорняки), взвешивались, а затем измельчались до 4 см с помощью ножа из нержавеющей стали. Часть образца была извлечена, промыта деионизированной водой для уменьшения загрязнения почвы, а затем высушена при температуре 60 °C в течение 48 часов до тех пор, пока не прекратилось дальнейшее уменьшение веса, как описано в [ 17 , 18 ].
2.5.3 Минеральный азот почвы (SMN)
Все блоки были почвенными образцами с размером ячеек 15 см до начала эксперимента 2 августа 2018 года и 10 августа 2019 года. Почву просеивали до 4 мм, затем 50 г смешивали со 100 мл 2 M KCl. Дополнительную часть образца почвы (10 г) высушивали в печи при 102 °C в течение 24 часов для оценки процента влажности и получения сухого веса использованной почвы. Смесь почвы и KCl помещали в орбитальный шейкер на 1 час при 200 об/мин, затем центрифугировали при 2976× g в течение 4 минут, и жидкую фракцию фильтровали через фильтровальную бумагу Whatman № 40. Для каждого набора экстракций (всего 4) проводили два холостых опыта для определения и корректировки на предмет любого загрязнения. Было проведено три повтора каждой обработки вместе с калибровочными стандартами, выступающими в качестве контроля качества. Содержание азота в почве на гектар оценивалось путем умножения концентрации азота на глубину почвы (15 см) и насыпную плотность 1,2 кг/л.
2.5.4 Отбор проб почвы во время выращивания покровных культур
Образцы почвы были отобраны 25 февраля 2019 г. и 11 марта 2020 г., когда покровные культуры ещё росли, и проанализированы на содержание SMN, как описано выше. Образцы почвы были отобраны керноотборником на глубину 15 см с двух блоков, что позволило провести 4 повторности каждой обработки, поскольку обработка азотом ещё не проводилась, и, следовательно, на тот момент планировка была разделена на участки.
2.6 Статистический анализ
2.6.1 Биомасса покровных культур
Для анализа параметров роста покровных культур методом дисперсионного анализа (ANOVA) использовалась программа Genstat версии 18. Для анализа поглощения азота покровными культурами в результате неравного количества наблюдений, например, биомассы корней и сорняков, а также поглощения питательных веществ, применялся метод ограниченного максимального правдоподобия (REML), поскольку не все виды были проанализированы из-за недостаточного количества образцов. Для выявления различий между видами применялся незащищённый постфактум-анализ Фишера.
Анализ REML повторных измерений использовался для анализа показаний цептометра LAI с степенной моделью порядка 1, примененной к случайной составляющей номера дня, чтобы учесть корреляционную структуру неравномерно разнесенных повторных измерений.
Результаты считаются значимыми, если вероятность случайного события составляет менее 5% ( p < 0,05), а тенденции рассматриваются менее 10% ( p = 0,10) [ 29 ].
2.6.2. Графики
Графики были созданы с помощью Graphpad Prism версии 7.00 для Windows, Graphpad Software, Ла-Хойя, Калифорния, США, www.graphpad.com (дата обращения: 20 октября 2019 г.).
3. Результаты
3.1. Индекс листовой поверхности (ИЛП)
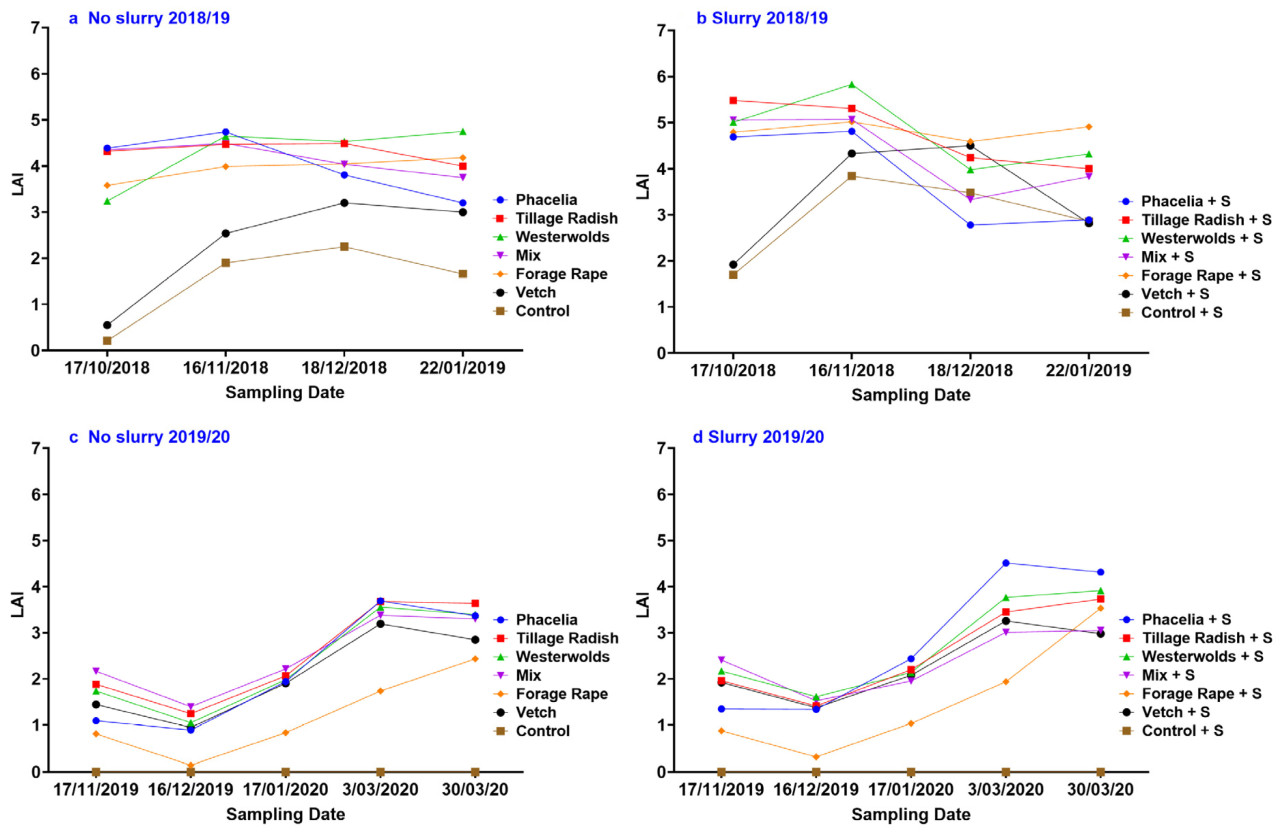
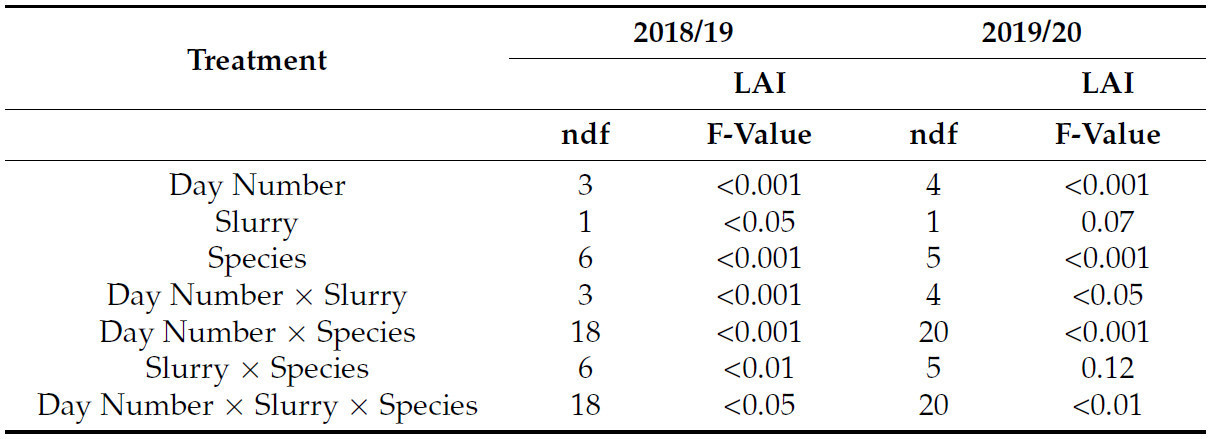
В 2018/19 году измерения цептометра показали, что в среднем жидкая взвесь значительно ( p < 0,05) увеличила LAI, но не в 2019/20 году. В оба года была обнаружена значительная взаимосвязь между числом дней и жидкой взвесью ( p < 0,001 в 2018/19 году, p < 0,05 в 2019/20 году) ( таблица 3 ). В 2018/19 году жидкая взвесь не только увеличила общий рост и LAI, но также и скорость роста и, следовательно, зрелость растений ( рисунок 1 a, b). Виды привели к значительным различиям в оба года ( p < 0,001) и продемонстрировали значительную взаимосвязь с числом дней ( p < 0,001). Следовательно, взаимодействие числа дней x жидкой взвесь x вида было значительным; p < 0,05 и p < 0,01 в 2018/19 и 2019/20 годах соответственно. Это взаимодействие можно наблюдать на рисунке 1 a–d, где при сравнении видов с навозной жижей и видов без навозной жижи LAI больше. Это особенно очевидно в 2018/19 году для таких видов, как фацелия, редис пахотный и вестервольд. Однако LAI значительно снизился после измерения, проведенного 16/11/18. В 2019/20 году отсроченный посев привел к значительно более низкому LAI для всех видов, чем в 2018/19 году. В контроле в 2019/20 году сорняки не росли, поэтому измерения не проводились. Фацелия с навозной жижей показала наибольший LAI с января и далее в 2019/20 году. Рапс кормовой, с навозной жижей и без нее, был видом с самым низким LAI в 2019/20 году, тогда как в 2018/19 году он дал самый высокий LAI.
3.2. Измерения биомассы покровных культур
В 2018/19 году сумма всей биомассы (покровной культуры, корней и сорняков) при измерении в феврале существенно зависела от навозной жижи ( p < 0,05), видов ( p < 0,05) и их взаимодействия ( p < 0,001) ( таблица 4 ). Однако в 2019/20 году сумма всей биомассы зависела только от видов при измерении в марте 2020 года ( p < 0,001) ( таблица 4 ).
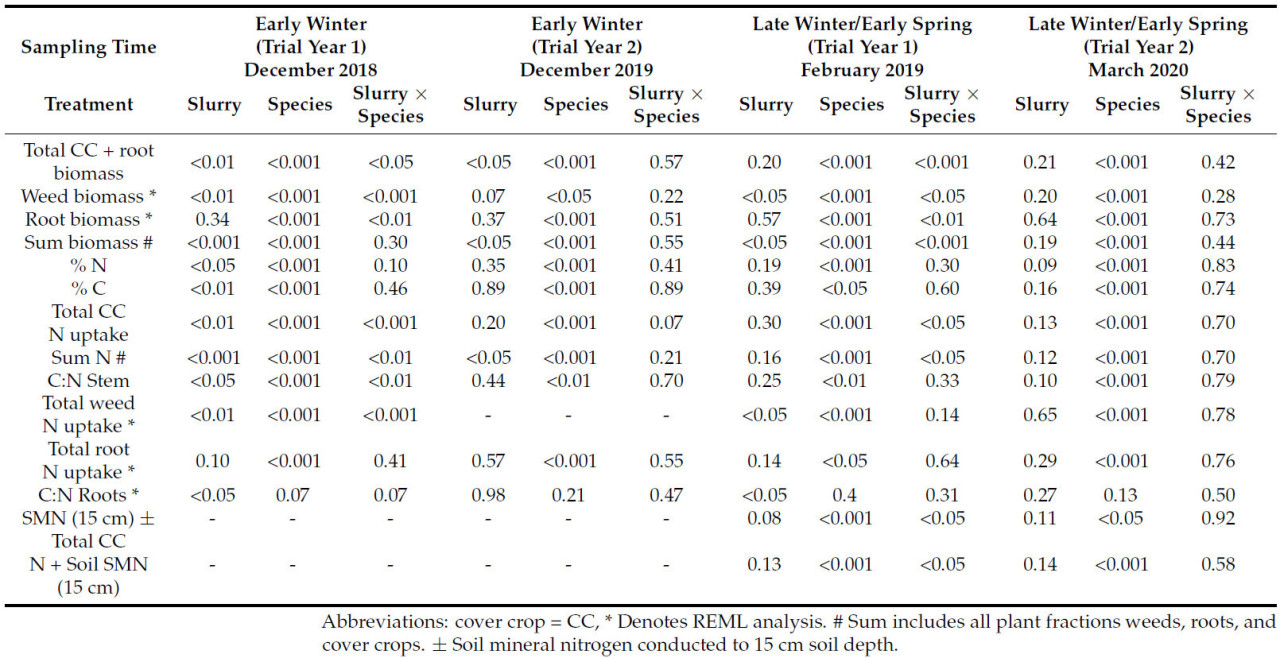
В 2019/20 году производство биомассы было значительно ниже, чем в 2018/19 году, в обе даты измерений. В 2018/19 году кормовой рапс с жидким навозом, по данным февральского измерения, имел наибольшую биомассу – 6689 кг/га, когда сорняки, корни и надземная покровная культура были объединены вместе. Однако вика без жидкого навоза произвела наименьшее количество биомассы покровной культуры ( рис. 2, а–г). В 2019/20 году вика также имела наименьшее производство биомассы среди всех видов покровных культур ( p < 0,05). Снижение потенциала роста между двумя годами лучше всего прослеживается на примере контрольного урожая 2019/20 года, где сорняки отсутствовали, тогда как в предыдущем году к февралю 2019 года их количество составляло почти 3000 кг/га. В 2019/20 году кормовой рапс дал наибольшую биомассу при измерении в марте (2673 кг/га). Все виды покровных культур, за исключением вики, имели схожую биомассу при измерении в марте ( p < 0,05).
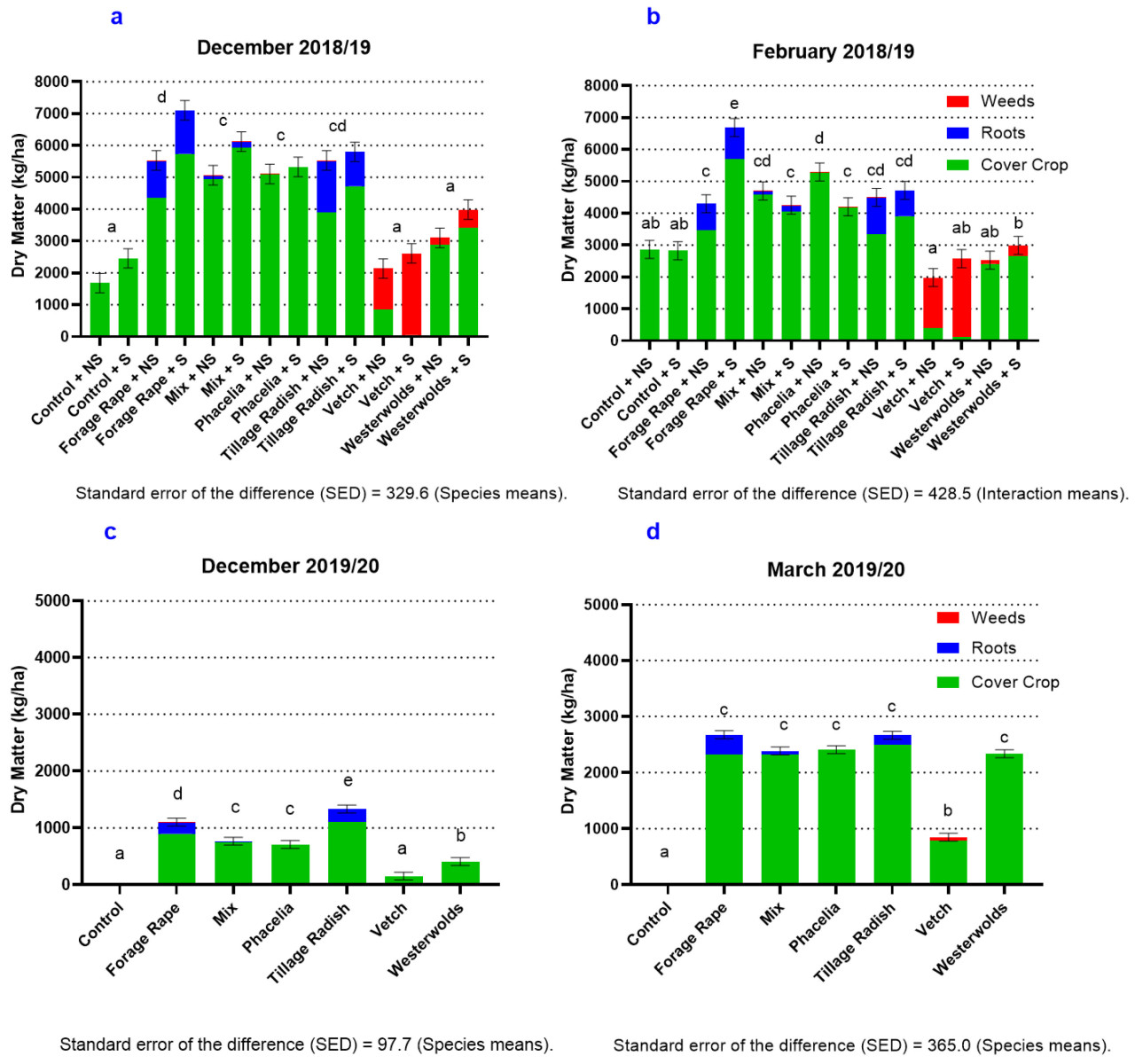
Контроль рассматривался как вид, чтобы можно было сравнить типичную практику и провести более значимые сравнения подавления сорняков между покровными культурами, которые будут оцениваться. Сорняки значительно увеличились при внесении навозной жижи ( p < 0,05) в 2018/19 году при измерении в декабре и феврале ( таблица 4 ), но на них не повлияли эти обработки в 2019/20 году ни в одну из дат отбора проб ( таблица 4 ). Виды, в обе даты отбора проб, значительно влияли на популяцию сорняков в оба года ( p < 0,001). В 2018/19 году вика произвела наибольшее количество сорняков ( p < 0,05), в среднем 2028 кг СВ/га, и привела к наибольшей биомассе сорняков в 2019/20 году ( p < 0,05) (59 кг/га) при измерении в феврале. В 2018/19 году кормовой рапс продемонстрировал наибольшее подавление сорняков, а при внесении жидкого навоза обеспечивал почти полное подавление в обе даты отбора проб.
Корни во все даты отбора проб в оба года значительно зависели от вида ( p < 0,001). В 2018/19 году при измерении в феврале редис пахотный дал самый большой стержневой корень и самую большую биомассу стержневых корней ( p < 0,05), составившую 1145 кг/га. Однако биомасса корней у этого вида была снижена из-за навозной жижи ( p < 0,05). В марте 2019/20 года кормовой рапс показал почти вдвое большую биомассу корней, чем редька пахотная (350 кг/га против 165 кг/га соответственно), но общая биомасса корней у обоих видов была значительно ниже в 2019/20 году по сравнению с предыдущим годом.
3.3. Соотношение C:N в покровных культурах в ответ на внесение жидкого навоза
На соотношение C:N существенное влияние ( p < 0,01) оказывали виды в оба года, при более поздних датах отбора проб в феврале и марте ( таблица 5 ). В оба года редька и вика имели самое низкое соотношение C:N по сравнению со всеми другими видами ( таблица 5 ). В 2018/19 году смесь всех видов и вестервольда имела значительно ( p < 0,05) более высокое соотношение C:N по сравнению со всеми другими видами, при этом в 2019/20 году у вестервольда снова было самое высокое соотношение C:N.
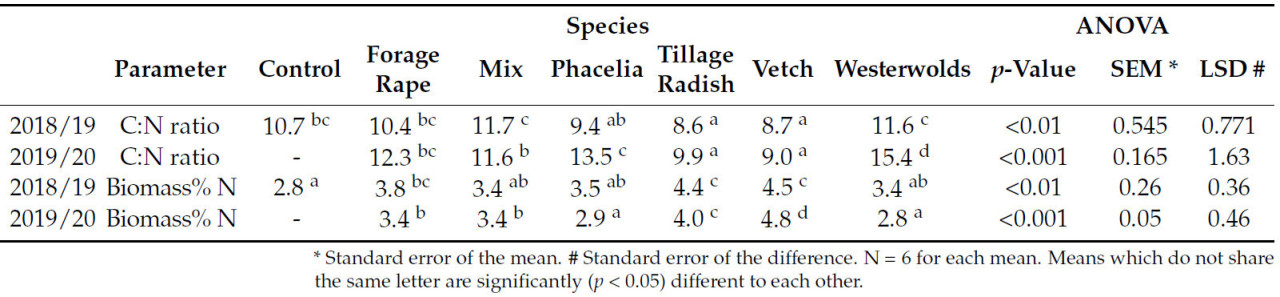
В декабрьской выборке опыта 2018/19 года навозная жижа значительно ( p < 0,05) снизила соотношение C:N покровной культуры и привела к значительным различиям между видами ( p < 0,001), что привело к значительному взаимодействию ( p < 0,05) ( таблица 4 ). Единственным видом в том году, который существенно ( p < 0,05) не снизил свое соотношение C:N в ответ на навозную жижу, была вика. Даже сорняки, восстановленные в контрольной группе, показали значительное снижение ( p < 0,05) соотношения C:N в ответ на навозную жижу. Однако в декабрьской выборке опыта 2019/20 года на соотношение C:N влияли только виды ( p < 0,01) ( таблица 4 ).
3.4. Поглощение азота покровными культурами
Рисунок 3 а показывает поглощение N покровной культурой для дат отбора проб в декабре и феврале. В феврале испытания 2018/19 г. общая сумма N покровной культуры (покровная культура, корни и сорняки) была подвержена влиянию навозной жижи ( p < 0,001), видов ( p < 0,001) и взаимодействия ( p < 0,001) ( таблица 4 ). Таблица 4 показывает, что в феврале 2018/19 г. виды привели к значительному ( p < 0,001) эффекту, как и взаимодействие с навозной жижей ( p < 0,05), но обработка навозной жижей не была значимой. Это перекрестное взаимодействие из-за эффектов навозной жижи, как увеличивающих, так и уменьшающих среднее усвоение N видами. Перекрестное взаимодействие задокументировано Грейс-Мартин (2020). Рисунок 3 а показывает, что кормовой рапс с жидким навозом имел наибольшее поглощение N при измерении в феврале, которое увеличивалось между декабрем и февралем. Рисунок 3 а также показывает, что фацелия без жидкого навоза увеличивала поглощение N между двумя временными точками, тогда как внесение жидкого навоза в этот вид вызывало снижение поглощения N. В 2019/20 г. общее поглощение N покровными культурами зависело только от вида ( p < 0,001). Рисунок 3 б демонстрирует гораздо более низкое поглощение N покровными культурами в 2019/20 г. по сравнению с 2018/19 г. В 2018/19 г. было обнаружено, что у редиса, выращиваемого на пашне (с жидким навозом), максимальное содержание N в биомассе составило 104 кг N/га, тогда как в 2018/19 г. у редиса, выращиваемого на пашне (с жидким навозом), поглощение N составило 233 кг/га (отбор проб в феврале). В 2018/19 году при внесении навозной жижи под кормовой рапс поглощение азота значительно ( p < 0,05) увеличилось – со 166 до 284 кг N/га. На рисунке 3 , а показано не только поглощение азота под воздействием навозной жижи, но и виды растений, которые увеличили или уменьшили поглощение азота биомассой в период между датами отбора проб. Ни один из видов не снизил поглощение азота биомассой в период с декабря по март в 2019/20 году.
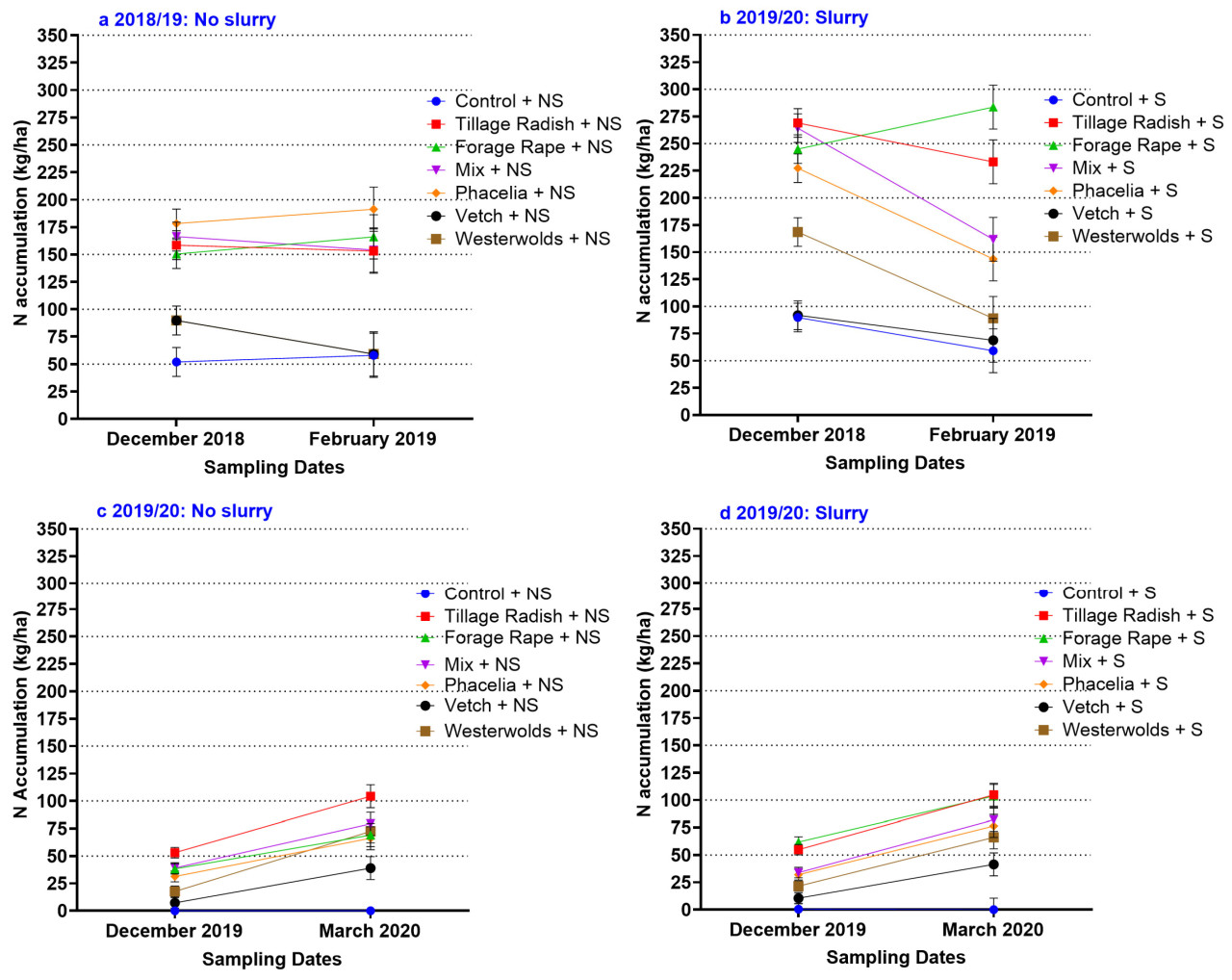
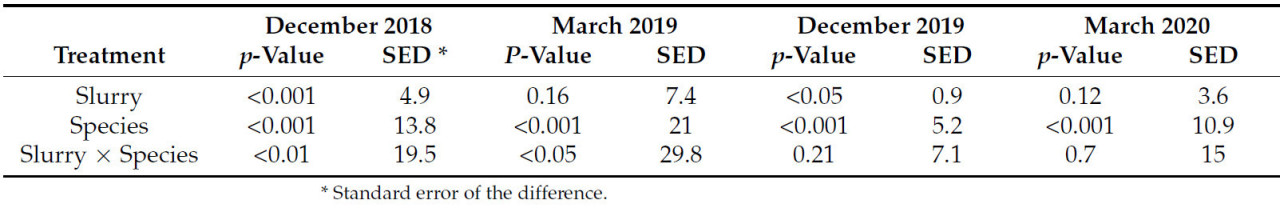
3.5 Общее поглощение азота покровными культурами (февраль 2019 г. и март 2020 г.)
Общее поглощение N состоит из N, содержащегося во фракциях надземной биомассы покровной культуры, корней покровной культуры, сорняков и SMN, отобранных на глубине 15 см, и показано на рисунке 4 a, b. В 2018/19 году все фракции N были выше, чем в 2019/20 году (N покровной культуры, корневой N, N сорняков и почвенный SMN). В 2018/19 году на общее поглощение N влияли виды, которые показали значительное взаимодействие с навозной жижей ( p < 0,05), но эффект обработки навозной жижей был незначительным ( таблица 6 ). Эффект навозной жижи вызвал как увеличение, так и уменьшение общего поглощения N и, таким образом, представлял собой перекрестное взаимодействие. В 2018/19 году, когда навозная жижа применялась для обработки редиса и кормового рапса, было получено в общей сложности 294 кг N/га и 299 кг N/га соответственно. Это значение было значительно выше, чем при отсутствии навозной жижи в отношении этих видов, а также по сравнению со всеми другими вариантами обработки, за исключением фацелии без навозной жижи. Навозная жижа снизила поглощение азота фацелией при отборе проб в феврале. В 2019/20 году контрольная культура привела к наименьшему общему поглощению азота из-за отсутствия биомассы у сорняков. В 2019/20 году редис и кормовой рапс, как и в предыдущем году, обеспечили наибольшее общее поглощение азота ( p < 0,05) ( рисунок 4, а), что привело к 114 кг азота/га и 94 кг азота/га, секвестрированного в почву.
В 2018/19 опытном году и вика, и вестервольд не привели к большему поглощению N, чем в контрольном варианте при измерении в феврале 2019 года ( Рисунок 4 а). Без навозной жижи фацелия в 2018/19 году поглощала 241 кг/га N, что на 170 кг N/га больше, чем в контрольном варианте без навозной жижи. Несмотря на более низкое поглощение N SMN и покровной культурой в 2019/20 опытном году, редька для обработки почвы обеспечила 118 кг N/га ( Рисунок 4 б). Это на 107 кг N/га больше, чем в контрольном варианте. В 2018/19 году в ответ на навозную жижу контрольная культура увеличила общее поглощение N всего на 37 кг/га. Это показывает, насколько эффективно некоторые виды покровных культур секвестрируют N по сравнению с оставлением земель под паром.
4. Обсуждение
4.1 Влияние навозной жижи на поглощение азота покровными культурами
В ответ на жидкую жижу кормовой рапс и редька пахотная накопили 123 кг N/га и на 114 кг N/га больше N в своей биомассе, сорняках и SMN в 2018/19 году. Аналогичным образом, Thilakarathna и др. [ 6 ] обнаружили, что только небобовые культуры увеличили свою биомассу в ответ на жидкую жижу. Однако в 2019/20 году покровные культуры не увеличили значительно свою биомассу или поглощение N в ответ на жидкую жижу. Это объясняется задержкой сева, которая привела к абиотическим условиям, которые ограничили рост, а не поступлением питательных веществ в почву ( рисунок 2 c, d). Контроль в 2019/20 году не накопил N в биомассе сорняков. Это, в сочетании с низкими количествами, извлеченными из SMN, показывает, что большие количества N были потеряны, скорее всего, за счет вымывания. Хотя в тот опытный год редька, выращиваемая под пашней, накопила 105 кг N/га в биомассе, которая, таким образом, сохранялась в почвенной системе, по сравнению с контрольным вариантом, представляющим собой пар, где азот не накапливался из-за отсутствия сорняков. Количество азота, накопленного в биомассе сорняков в контрольном варианте в 2018/19 году ( рисунок 4 а), было нетипичным. При обычной практике эти сорняки были бы уничтожены, чтобы предотвратить их семенное развитие. Если бы сорняки в контрольном варианте данного опыта были уничтожены для предотвращения роста, это могло бы изменить цикл азота в 2018/19 году.
Этот эксперимент продемонстрировал гораздо больший потенциал поглощения N покровными культурами по сравнению с экспериментом Бхогала и др. [ 31 ]. Более низкое поглощение N в том эксперименте частично объясняется тем, что одна из дат посева была намного более поздней, чем в этом эксперименте. Кроме того, в этом эксперименте фацелия, посеянная позже (10 сентября 2019 г.), сумела накопить 76 кг/га N (с навозной жижей), что снова намного выше, чем сообщают Бхогал и др. [ 31 ]. Эти авторы также обнаружили, что контрольный пар накапливал 25 кг/га сорняков. Следовательно, поглощение N покровной культурой ненамного выше, чем пар. В Дании из 90 полей, на которых в 2019 г. были отобраны образцы покровных культур, большинство накопили менее 40 кг N/га [ 32 ]. В Ирландии покровная культура горчицы накопила 31, 81 и 6 кг N/га за зиму в течение трех испытательных лет 2007–2009 соответственно [ 33 ].
В этом испытании в 2018/19 году без навозной жижи фацелия накопила 178 кг N/га в декабре 2018 года, что увеличилось до 191 кг N/га к февралю 2019 года. С навозной жижей фацелия накопила 227 кг N/га в декабре 2018 года, что снизилось до 143 кг N/га в феврале 2019 года. Без навозной жижи фацелия в испытательном году 2018/19 накопила наибольшее количество N по сравнению со всеми другими видами. Аналогичным образом, Ин 'т Зандт, Д и др. [ 34 ] обнаружили, что фацелия не реагировала на дополнительный N, тогда как кормовой рапс и пахотный редис реагировали. Кроме того, эти авторы пришли к выводу, что фацелия может не справляться с очень высокими концентрациями N. В этом исследовании фацелия отреагировала на внесение навозной жижи в декабре 2018 года, но к февралю 2019 года в биомассе, сорняках и почве под фацелией, внесённой навозной жижей, было обнаружено меньше азота. Это было связано с заморозками, которые привели к старению растений и, как следствие, к обратному переносу азота из биомассы в почву, что было подтверждено повышенным значением SMN, как было обнаружено в [ 35 ].
4.2. Выделение питательных веществ из покровных культур
LAI показывает не только, как покровные культуры росли и формировали листовой полог, но также дает важную информацию, такую как повреждение культуры как от заморозков, так и от естественного старения, что привело к снижению LAI. Это было связано с тем, что заморозки повредили листья, поэтому полог перехватывал меньше света, что приводило к снижению LAI. Влияние заморозков на чувствительные виды было очевидно в 2018/19 году по снижению как LAI ( Рисунок 1 ), так и биомассы покровной культуры между измерениями в декабре и феврале ( Рисунок 2 c, d). Это привело к снижению поглощения N биомассой ( Рисунок 3 a, b); в свою очередь, это увеличило восстановленное SMN ( Рисунок 4 a). Виды, которые не были затронуты, например, кормовой рапс, продолжали расти и демонстрировали эффекты, противоположные тем, которые были чувствительными. Заморозки были в 2019/20 году, но не повлияли ни на одну из комбинаций обработок ( Рисунок 3 c, d). Это очевидно, поскольку все виды обработки навозной жижей + увеличили их биомассу и поглощение азота в период с декабря по март 2019/20 года ( рисунок 3 и рисунок 4 б).
Сила заморозков/температура были схожими между годами ( Рисунок A3 ), но контрастные эффекты объясняются различиями в зрелости видов покровных культур и устойчивости к заморозкам. Это было обнаружено в похожем эксперименте, где более поздние посевы покровных культур, сопоставимые с датой посева во второй испытательный год, не были затронуты заморозками [ 36 ]. Влияние заморозков в 2018/19 увеличило количество SMN, восстановленного у восприимчивых видов, из-за старения, которое вызвало высвобождение питательных веществ. Это также было обнаружено Сторром и соавторами [ 35 ] при использовании смеси видов, чувствительных к заморозкам, по сравнению с устойчивыми к заморозкам. В 2018/19 фацелия и редька для обработки дали SMN 67 кг/га и 62 кг/га соответственно, когда вносилась жидкая навозная жижа. Для сравнения, без внесения жидкая навозная жижа кормовой рапс и контроль имели самое низкое SMN 11 кг N/га. Кроме того, это показывает, что покровные культуры лучше сохраняют азот и обеспечивают его поступление в товарные культуры.
Соотношение C:N в биомассе покровных культур является важным показателем для оценки и определения доступности питательных веществ из покровных культур [ 37 ]. Соотношения C:N у видов в этом эксперименте были очень низкими, независимо от обработки, т. е. навозной жижи, и по сравнению с другими исследованиями: Couëdel et al. (2018) [ 30 ] сообщили, что у таких видов, как рапс (кормовой рапс), редис и вика, соотношение C:N составляло 22,0, 24,4 и 10 соответственно. Büchi et al. [ 14 ] обнаружили, что у фацелии соотношение C:N составляло 37,5 и 33,8 с % N 1,05 и 1,34 соответственно на двух разных участках. Следовательно, участок с высоким содержанием SMN обеспечил более высокий процент содержания N в фацелии и более низкое соотношение C:N (124 кг N/га по сравнению с 74 кг N/га, полученными при предварительном отборе проб на глубине 90 см).
5. Выводы
Данный эксперимент демонстрирует, что покровные культуры накапливают большое количество азота в своей биомассе. Кормовой рапс с жидким навозом накапливал 299 кг/га азота в своей биомассе, сорняках и в составе азота сульфида азота. В то же время, при позднем посеве и менее благоприятных условиях выращивания редис полевой накапливал 120 кг/га азота в тех же фракциях. Более того, при раннем посеве фацелия и редис полевой могут одновременно повышать содержание азота в почве и сохранять больше азота в своей биомассе по сравнению с оставлением земли под паром.
Данное исследование демонстрирует не только важность раннего посева покровных культур, где это возможно, но и значительные различия в эффективности покровных культур по годам. В обоих годах покровные культуры в этом регионе играют важную роль в секвестировании питательных веществ по сравнению с оставлением земель под паром.
Это исследование показывает, что заморозки и сбор неустойчивых к заморозкам видов могут быть эффективным инструментом для прекращения выращивания покровных культур, в результате чего фацелия и редис начинают высвобождать N из биомассы в почву. Покровные культуры, устойчивые к заморозкам, могут иметь гораздо более медленные темпы минерализации из биомассы после прекращения их выращивания. Однако, если темпы минерализации достаточны, покровные культуры должны не только положительно влиять на последующую культуру, но и могут иметь потенциал для снижения ее потребности в неорганических азотных удобрениях. Там, где покровные культуры интегрируются с навозной жижей, кормовой рапс способен увеличивать биомассу и удерживать высокие уровни N в своих остатках. Это приводит к максимальному усвоению N 283 кг/га при сохранении низкого SMN. Однако синергия между навозной жижей и покровными культурами зависит от выбора видов и раннего посева, чтобы обеспечить достаточный рост.
Приложение А
Ссылки
1. Abdalla, M.; Hastings, A.; Cheng, K.; Yue, Q.; Chadwick, D.; Espenberg, M.; Truu, J.; Rees, B.; Smith, P. A critical review of the impacts of cover crops on nitrogen leaching, net greenhouse gas balance and crop productivity. Glob. Change Biol. 2019, 25, 2530–2543. [Google Scholar] [CrossRef]
2. Allingham, K.D.; Cartwright, R.; Donaghy, D.; Conway, J.S.; Goulding, K.W.T.; Jarvis, S.C. Nitrate leaching losses and their control in a mixed farm system in the Cotswold Hills, England. Soil Use Manag. 2002, 18, 421–427. [Google Scholar] [CrossRef]
3. Everett, L.; Wilson, M.; Pepin, R.; Coulter, J. Winter Rye Cover Crop with Liquid Manure Injection Reduces Spring Soil Nitrate but Not Maize Yield. Agronomy 2019, 9, 852. [Google Scholar] [CrossRef]
4. Liu, J.; Li, J.; Ma, Y.; Jia, Y.; Liang, Q. Apparent Accumulated Nitrogen Fertilizer Recovery in Long-Term Wheat–Maize Cropping Systems in China. Agronomy 2018, 8, 293. [Google Scholar] [CrossRef]
5. Huang, J.; Duan, Y.-h.; Xu, M.-g.; Zhai, L.-m.; Zhang, X.-b.; Wang, B.-r.; Zhang, Y.-z.; Gao, S.-d.; Sun, N. Nitrogen mobility, ammonia volatilization, and estimated leaching loss from long-term manure incorporation in red soil. J. Integr. Agric. 2017, 16, 2082–2092. [Google Scholar] [CrossRef]
6. Thilakarathna, M.; Serran, S.; Lauzon, J.; Janovicek, K.; Deen, B. Management of Manure Nitrogen Using Cover Crops. Agron. J. 2015, 107, 1595–1607. [Google Scholar] [CrossRef]
7. Aronsson, H.; Hansen, E.; Thomsen, I.; Liu, J.; Øgaard, A.; Kankanen, H.; Ulén, B. The ability of cover crops to reduce nitrogen and phosphorus losses from arable land in southern Scandinavia and Finland. J. Soil Water Conserv. 2016, 71, 41–55. [Google Scholar] [CrossRef]
8. Parkin, T.B.; Kaspar, T.; Singer, J.W. Cover crop effects on the fate of N following soil application of swine manure. Plant Soil 2006, 289, 141–152. [Google Scholar] [CrossRef]
9. Seman-Varner, R.; Varco, J.; O’Rourke, M. Nitrogen Benefits of Winter Cover Crop and Fall-Applied Poultry Litter to Corn. Agron. J. 2017, 109, 2881–2888. [Google Scholar] [CrossRef]
10. Wittwer, R.; Dorn, B.; Jossi, W.; Van der Heijden, M. Cover crops support ecological intensification of arable cropping systems. Sci. Rep. 2017, 7, 41911. [Google Scholar] [CrossRef] [PubMed]
11. Qin, Z.; Guan, K.; Zhou, W.; Peng, B.; Villamil, M.B.; Jin, Z.; Tang, J.; Grant, R.; Gentry, L.; Margenot, A.J.; et al. Assessing the impacts of cover crops on maize and soybean yield in the U.S. Midwestern agroecosystems. Field Crops Res. 2021, 273, 108264. [Google Scholar] [CrossRef]
12. Rosa, A.T.; Creech, C.F.; Elmore, R.W.; Rudnick, D.R.; Lindquist, J.L.; Fudolig, M.; Butts, L.; Werle, R. Implications of cover crop planting and termination timing on rainfed maize production in semi-arid cropping systems. Field Crops Res. 2021, 271, 108251. [Google Scholar] [CrossRef]
13. Couëdel, A.; Kirkegaard, J.; Alletto, L.; Justes, É. Chapter Two–Crucifer-legume cover crop mixtures for biocontrol: Toward a new multi-service paradigm. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 55–139. [Google Scholar]
14. Büchi, L.; Gebhard, C.; Liebisch, F.; Sinaj, S.; Ramseier, H.; Charles, R. Accumulation of biologically fixed nitrogen by legumes cultivated as cover crops in Switzerland. Plant Soil 2015, 393, 1–13. [Google Scholar] [CrossRef]
15. Carlsson, G.; Huss-Danell, K. Nitrogen fixation in perennial forage legumes in the field. Plant Soil 2003, 253, 353–372. [Google Scholar] [CrossRef]
16. Wendling, M.; Büchi, L.; Amosse, C.; Sinaj, S.; Walter, A.; Charles, R. Nutrient accumulation by cover crops with different root systems. Asp. Appl. Biol. 2015, 129, 91–95. [Google Scholar]
17. Couëdel, A.; Alletto, L.; Justes, E. Crucifer-legume cover crop mixtures provide effective sulphate catch crop and sulphur green manure services. Plant Soil 2018, 426, 61–76. [Google Scholar] [CrossRef]
18. Couëdel, A.; Alletto, L.; Kirkegaard, J.; Justes, É. Crucifer glucosinolate production in legume-crucifer cover crop mixtures. Eur. J. Agron. 2018, 96, 22–33. [Google Scholar] [CrossRef]
19. Roarty, S.; Hackett, R.; Schmidt, O. Earthworm populations in twelve cover crop and weed management combinations. Appl. Soil Ecol. 2017, 114, 142–151. [Google Scholar] [CrossRef]
20. Finney, D.; Kemanian, A.; Kaye, J. A Model–Data Fusion Approach for Predicting Cover Crop Nitrogen Supply to Corn. Agron. J. 2016, 108, 2527–2540. [Google Scholar]
21. Kaye, J.; Finney, D.; White, C.; Bradley, B.; Schipanski, M.; Alonso-Ayuso, M.; Hunter, M.; Burgess, M.; Mejia, C. Managing nitrogen through cover crop species selection in the U.S. mid-Atlantic. PLoS ONE 2019, 14, e0215448. [Google Scholar] [CrossRef]
22. Finney, D.; Buyer, J.S.; Kaye, J.P. Living cover crops have immediate impacts on soil microbial community structure and function. J. Soil Water Conserv. 2017, 72, 361–373. [Google Scholar] [CrossRef]
23. Hunter, M.C.; Schipanski, M.E.; Burgess, M.H.; LaChance, J.C.; Bradley, B.A.; Barbercheck, M.E.; Kaye, J.P.; Mortensen, D.A. Cover Crop Mixture Effects on Maize, Soybean, and Wheat Yield in Rotation. Agric. Environ. Lett. 2019, 4, 180051. [Google Scholar] [CrossRef]
24. Schappert, A.; Schumacher, M.; Gerhards, R. Weed Control Ability of Single Sown Cover Crops Compared to Species Mixtures. Agronomy 2019, 9, 294. [Google Scholar] [CrossRef]
25. Trinsoutrot Gattin, I.; Recous, S.; Bentz, B.; Nicolardot, B. Biochemical Quality of Crop Residues and Carbon and Nitrogen Mineralization Kinetics under Nonlimiting Nitrogen Conditions. Soil Sci. Soc. Am. J. SSSAJ 2000, 64, 918–926. [Google Scholar] [CrossRef]
26. Wendling, M.; Büchi, L.; Amosse, C.; Sinaj, S.; Walter, A.; Charles, R. Influence of root and leaf traits on the uptake of nutrients in cover crops. Plant Soil 2016, 409, 419–434. [Google Scholar] [CrossRef]
27. Lawson, A.; Cogger, C.; Bary, A.; Fortuna, A.-M. Influence of Seeding Ratio, Planting Date, and Termination Date on Rye-Hairy Vetch Cover Crop Mixture Performance under Organic Management. PLoS ONE 2015, 10, e0129597. [Google Scholar] [CrossRef]
28. Cottney, P.; Black, L.; White, E.; Williams, P.N. The Correct Cover Crop Species Integrated with Slurry Can Increase Biomass, Quality and Nitrogen Cycling to Positively Affect Yields in a Subsequent Spring Barley Rotation. Agronomy 2020, 10, 1760. [Google Scholar] [CrossRef]
29. VSN International. Genstat for Windows 18th; VSN International: Hemel Hempstead, UK, 2017. [Google Scholar]
30. Couëdel, A.; Alletto, L.; Tribouillois, H.; Justes, É. Cover crop crucifer-legume mixtures provide effective nitrate catch crop and nitrogen green manure ecosystem services. Agric. Ecosyst. Environ. 2018, 254, 50–59. [Google Scholar] [CrossRef]
31. Bhogal, A.; White, C.; Morris, N. Maximising the Benefits from Cover Crops Through Species Selection and Crop Management; Agriculture and Horticulture Development Board: Nottinghamshire, UK, 2020. [Google Scholar]
32. Kristensen, N.H. The story of catch crops in Denmark. In PlanteInnovation, SEGES, Denmark; SEGES, Ed.; Teagasc National Tillage Conference: County Kildare, Ireland, 2020. [Google Scholar]
33. Premrov, A.; Coxon, C.E.; Hackett, R.; Kirwan, L.; Richards, K.G. Effects of over-winter green cover on groundwater nitrate and dissolved organic carbon concentrations beneath tillage land. Sci. Total Environ. 2012, 438, 144–153. [Google Scholar] [CrossRef] [PubMed]
34. In ‘t Zandt, D.; Fritz, C.; Wichern, F. In the land of plenty: Catch crops trigger nitrogen uptake by soil microorganisms. Plant Soil 2017, 423, 1–14. [Google Scholar] [CrossRef]
35. Storr, T.; Simmons, R.; Hannam, J. Using frost sensitive cover crops for timely nitrogen mineralisation and soil moisture management. Soil Use Manag. 2020, 37, 427–435. [Google Scholar] [CrossRef]
36. Cottney, P.; Black, L.; Williams, P.; White, E. How Cover Crop Sowing Date Impacts upon Their Growth, Nutrient Assimilation and the Yield of the Subsequent Commercial Crop. Agronomy 2022, 12, 369. [Google Scholar] [CrossRef]
37. Justes, E.; Mary, B.; Nicolardot, B. Quantifying and modelling C and N mineralization kinetics of catch crop residues in soil: Parameterization of the residue decomposition module of STICS model for mature and non mature residues. Plant Soil 2009, 325, 171–185. [Google Scholar] [CrossRef]

Перевод статьи «Organic Manures Integrated with Cover Crops Affect Both Cover Crop and Commercial Crop Performance Including Soil Health: Effect on Cover Crop Growth, Nitrogen Uptake, and Soil Mineral N» авторов Cottney, P.; Black, L.; White, E.; Williams, P.N., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык



















































































































Комментарии (0)