Влияние биоугля из соломы пшеницы на почвенные микробные сообщества в условиях стресса, вызванного фенантреном
Потенциал биоугля влиять на изменения в почвенных микробных сообществах, вызванные стрессом от полициклических ароматических углеводородов (ПАУ) в сельскохозяйственных почвах, и тем самым способствовать биоремедиации загрязненной почвы, остается не до конца изученным. В данном исследовании в сельскохозяйственную почву, загрязненную фенантреном в концентрациях 2,5 и 25 мг/кг, вносили биоуголь из соломы пшеницы, полученный при 300 °C (W300) и 500 °C (W500), в различных количествах (1% и 2% от массы). Целью было изучить их влияние на структуру микробного сообщества и разложение фенантрена аутохтонными микроорганизмами.
Аннотация
Внесение биоугля как в слабо (PLS), так и в сильно (PHS) загрязненную почву привело к увеличению общей микробной/бактериальной биомассы, сохранению бактериального разнообразия и селективному обогащению определенных родов бактерий, которые подавлялись стрессом от фенантрена, за счет усиления сорбции и снижения биотоксичности. В PHS биоуголь, особенно W300, способствовал увеличению численности родов бактерий, разлагающих ПАУ, и гена деградации nidA за счет улучшения почвенных питательных веществ, что усиливало биодеградацию фенантрена. Однако в PLS биоуголь, в особенности W500, подавлял их численность из-за снижения биодоступности фенантрена для специфических деструкторов, тем самым препятствуя биодеградации фенантрена.
Эти результаты позволяют предположить, что внесение биоугля из соломы пшеницы, полученного при соответствующих температурах, может благоприятно влиять на почвенную микробную экологию и способствовать устранению ПАУ, предлагая устойчивую стратегию использования ресурсов соломы для защиты здоровья почв и качества сельскохозяйственной продукции.
1. Введение
Полициклические ароматические углеводороды (ПАУ), образующиеся в основном при неполном сгорании ископаемого топлива и биомассы, были признаны одними из основных органических загрязнителей почвы в Китае согласно национальному исследованию почв. Около 20% пахотных земель сильно загрязнены [1,2,3]. Будучи канцерогенными, тератогенными и мутагенными загрязнителями, ПАУ вызывают серьезную озабоченность из-за своей токсичности, устойчивости и повсеместного распространения, представляя угрозу для здоровья человека через пищевую цепь [4,5]. Поэтому крайне важно и срочно снизить экологические риски, которые несут ПАУ в сельскохозяйственных почвах, чтобы обеспечить безопасность продукции растениеводства.
Загрязнение почвы ПАУ, как было показано, снижает разнообразие и активность почвенных микробных сообществ [6], которые играют ключевую роль в функционировании агроэкосистем, включая плодородие почвы, продуктивность культур и устойчивость к стрессам [7]. Поддержание разнообразной структуры микробного сообщества благоприятствует почвенным экологическим процессам и стабильности, поскольку микробы играют важнейшую роль в энергетическом метаболизме, биогеохимических циклах питательных веществ и разложении органических загрязнителей в почве [8,9]. Таким образом, восстановление почвенной микробной экологии является imperative для улучшения и ремедиации загрязненных ПАУ сельскохозяйственных земель.
Производство биоугля путем пиролиза соломы и его внесение на поля для улучшения качества почвы, обеспечивая такие преимущества, как регулирование кислотности, связывание углерода, удержание влаги и сохранение питательных веществ, привлекает все больше внимания [10,11,12]. Использование технологий биоугля для переработки соломенных отходов открывает перспективы для их утилизации [13], что имеет большое значение для качества воздуха в Китае и во всем мире [14]. Биоуголь, богатый углеродом материал, считается высокоэффективным сорбентом для снижения экологических рисков, создаваемых ПАУ в сельскохозяйственных почвах [15]. Биоуголь может иммобилизовать ПАУ, снижая их биодоступность, перемещение и биоаккумуляцию в почве, тем самым уменьшая их токсичность для почвенных организмов, особенно для важных микробных сообществ [16]. Предыдущие исследования показали, что внесение 1% (по массе) биоугля из соломы кукурузы, полученного при 500 °C, способствовало выживанию специфических бактериальных таксонов (особенно Bacteroidetes, Latescibacteria, Saccharibacteria, Parcubacteria, Ignavibacterium, Basidiomycota и Zygomycota), которые значительно сократились под воздействием фенантрена в почве [17]. Богатая и устойчивая структура микробного сообщества необходима для здоровья почвы. Однако другие исследователи сообщали, что внесение 5% (по массе) биоугля из соломы кукурузы в почву, загрязненную ПАУ, не привело к значительным изменениям в пропорциях бактериальных родов, что указывает на отсутствие заметного влияния на состав микробного сообщества [18]. Остается неясным, может ли внесение биоугля из соломы в загрязненную ПАУ пахотную почву опосредовать изменения в почвенных микробных сообществах, вызванные стрессом от ПАУ, чтобы улучшить здоровье почвы, что потенциально зависит от исходного сырья для биоугля, температуры обработки и уровня внесения. Существует ограниченное понимание взаимного влияния применения биоугля и стресса от ПАУ на почвенные микробные сообщества, что требует дальнейшего изучения.
Поскольку солома, как травянистый материал, дает биоуголь, более богатый питательными веществами и имеющий более пористую структуру, чем древесные материалы [19,20], внесение биоугля из соломы в загрязненные и находящиеся в стрессе почвенные среды имеет потенциал для изменения физико-химических свойств почвы и обеспечения подходящего уровня pH, жизненного пространства и питательных веществ для роста микробов [21,22,23]. Это может увеличить микробную биомассу, разнообразие и активность, способствуя, в свою очередь, метаболизму и трансформации загрязнителей и снижая их опасность для почвенных микробов, что приводит к формированию более устойчивого микробного сообщества для безопасных аграрных практик [24,25]. Предыдущие исследования highlighted увеличение микробной деградации ПАУ в почве за счет биостимуляции биоуглем из соломы пшеницы и рапса, что объяснялось увеличением общей численности бактерий и обогащением специфических видов бактерий и функциональных генов, связанных с разложением ПАУ [26,27]. Однако Bao и др. наблюдали, что добавление как 1%, так и 4% биоугля значительно снижало биодеградацию фенантрена в почве, что могло быть связано с изменениями в составе консорциума микроорганизмов, разлагающих фенантрен, и ослаблением обилия активных деструкторов фенантрена (например, Sphaerobacter, неклассифицированные Diplorickettsiaceae, Pseudonocardia и Planctomyces) и генов PAH-RHDα, индуцированных биоуглем [28]. Эти противоречивые данные указывают на то, что то, как биоуголь влияет на реакцию почвенных микробных сообществ на токсический стресс от ПАУ и связанное с этим воздействие на биоремедиацию ПАУ, еще не до конца понятно, и лежащие в основе механизмы требуют дальнейшего изучения.
Цель данного исследования — оценить влияние биоугля из соломы пшеницы на почвенные микробные сообщества в условиях стресса от ПАУ и его влияние на разложение ПАУ аутохтонными микроорганизмами в почве. Сельскохозяйственные почвы с разными уровнями загрязнения фенантреном (2,5 и 25 мг/кг) обрабатывали биоуглем из соломы пшеницы, полученным при 300 и 500 °C, в различных дозах (1% и 2% по массе в пересчете на сухое вещество). Изучались изменения в почвенных микробных сообществах (а именно, общая микробная биомасса, активность внеклеточных ферментов, количество генов бактериальной 16S рРНК и PAH-деградирующего гена nidA, а также структура и разнообразие бактериального сообщества) под воздействием стресса от фенантрена и то, как эти изменения опосредуются биоуглем. Это исследование также изучило физиологические механизмы, лежащие в основе влияния реакции микробного сообщества на внесение биоугля в загрязненной ПАУ почве на биоремедиацию ПАУ.
2. Материалы и методы
2.1. Почва и биоуголь
Чистая почва с pH 6,58 и гранулометрическим составом 32,6% песка, 54,1% ила и 13,3% глины была отобрана с незагрязненного сельскохозяйственного поля в Нанкине, провинция Цзянсу, Китай, в сентябре 2023 года. Фоновые концентрации 16 приоритетных ПАУ и фенантрена в почве составили 1419,2 ± 14,5 и 169,3 ± 3,8 мкг/кг соответственно. Образец почвы (глубина 0–20 см) имел содержание общего органического углерода (TOC) 17,99 г/кг и растворенного органического углерода (DOC) 91,64 мг/кг. Содержание общего азота (TN), фосфора (TP) и калия в почве составило 1,15 г/кг, 0,77 г/кг и 14,95 г/кг соответственно. Содержание доступного азота (AN) и фосфора (AP) в tested почве составило 34,21 и 18,20 мг/кг соответственно. Собранную почву высушивали на воздухе, измельчали и просеивали через сито с размером ячеек 2 мм перед использованием.
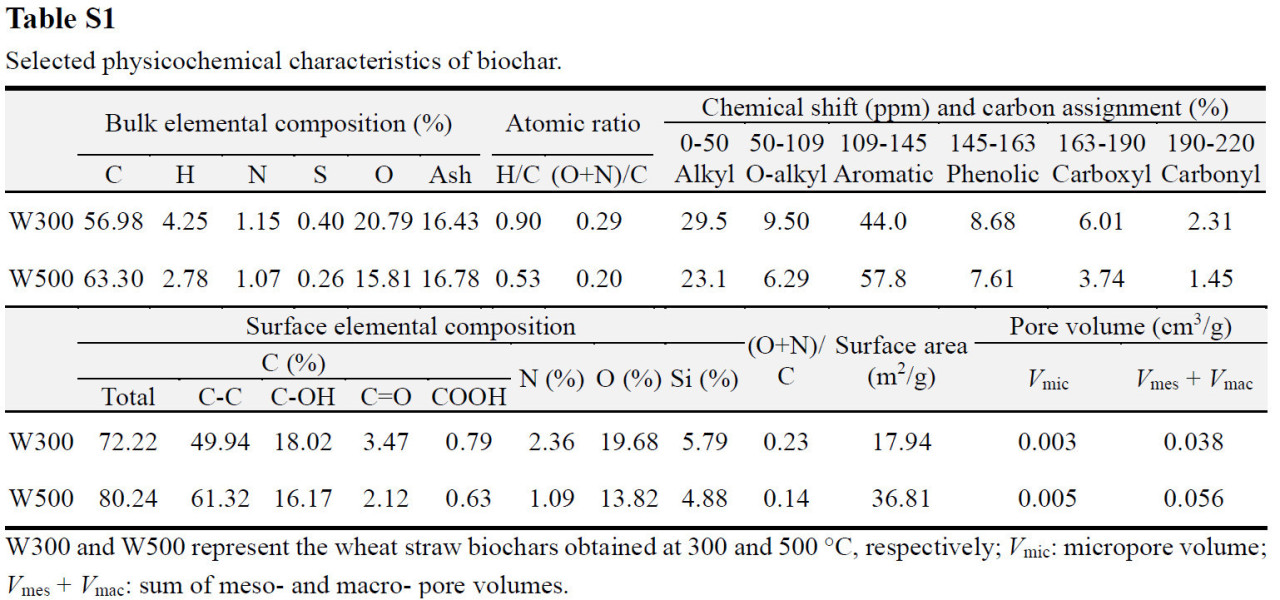
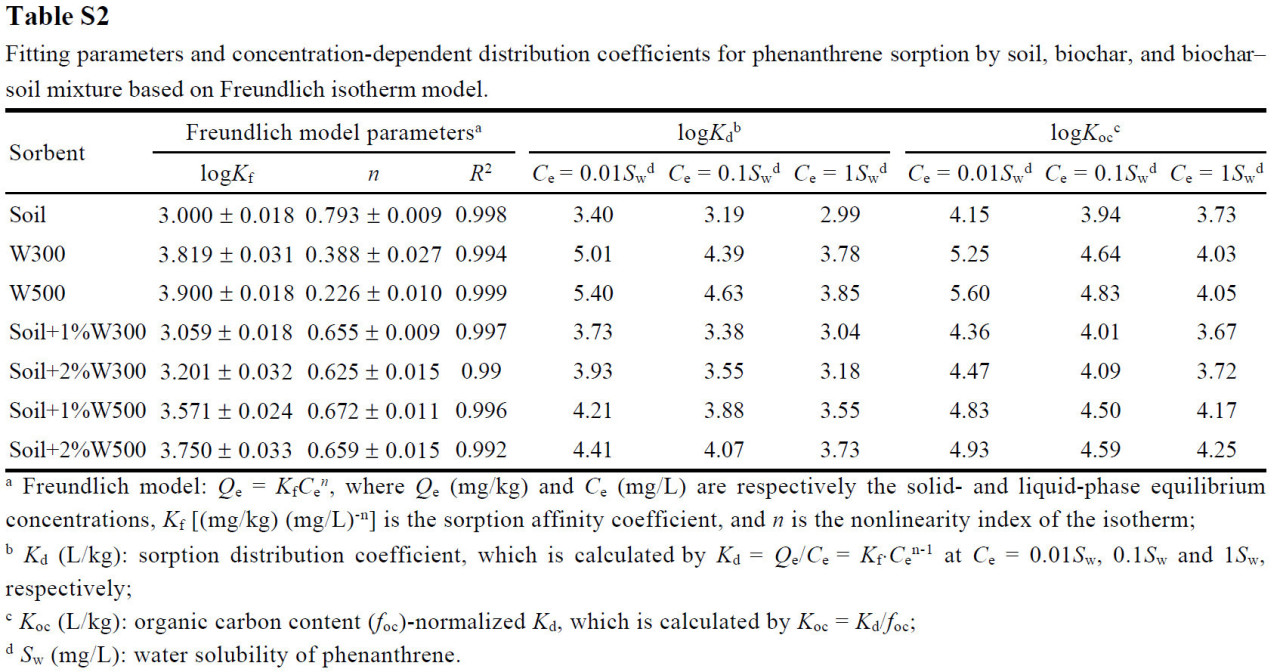
Биоуголь получали пиролизом соломы пшеницы, типичного сельскохозяйственного остатка, при 300 или 500 °C в бескислородных условиях в муфельной печи и маркировали как W300 или W500 соответственно. Биоугли, полученные при этих двух температурах, имели схожее содержание фенантрена: 169,3 ± 6,8 и 188,1 ± 8,9 мкг/кг соответственно. Эти концентрации были значительно ниже уровней загрязнения (2,5 и 25 мг/кг) фенантрена в исследуемых почвах, что делало их влияние незначительным. Основные физико-химические характеристики биоуглей подробно описаны в Таблице S1 в Дополнительных материалах, как сообщалось в нашем недавнем исследовании [29]. Изотермы сорбции фенантрена исследуемой почвой, биоуглями и их смесями с почвой исследовали с помощью техники равновесного насыщения, а параметры подгонки по модели Фрейндлиха приведены в Таблице S2.
2.2. Эксперименты по инкубации
Для получения почвы с разным уровнем загрязнения фенантреном почву разделили на две части. Каждую часть обрабатывали раствором фенантрена в ацетоне и тщательно перемешивали после испарения растворителя в вытяжном шкафу для получения низкой (2,5 мг/кг) и высокой (25 мг/кг) начальных концентраций, обозначенных как PLS и PHS соответственно. Затем в загрязненную почву вносили и тщательно перемешивали 1% или 2% W300 или W500 (по массе в пересчете на сухое вещество). Десять вариантов опыта обозначили как PL, PL1%W3, PL1%W5, PL2%W3, PL2%W5, PH, PH1%W3, PH1%W5, PH2%W3 и PH2%W5. Почву без обработки фенантреном или биоуглем использовали в качестве контрольной (CK). Для каждого варианта 120 г (в сухом весе) почвы переносили в стеклянный стакан на 250 мл и увлажняли стерильной деионизированной водой до доведения влажности почвы до 30%, что соответствовало полевой влагоемкости. Каждый вариант опыта выполняли в трех повторностях и инкубировали в климатической камере в темноте при 25 °C и относительной влажности 60% в течение 21 дня. В конце периода инкубации из каждой повторности различных вариантов отбирали 2 г почвы для определения концентрации фенантрена. Еще 3 г почвы отбирали на 21-й день для анализа активности почвенного фермента (полифенолоксидазы) и структуры бактериального сообщества. Чтобы оценить влияние самого биоугля на структуру микробного сообщества в незагрязненной почве, почву, обработанную биоуглем, но без внесения фенантрена (т.е. 1%W300, 1%W500, 2%W300 и 2%W500), готовили и культивировали по той же процедуре.
2.3. Экстракция и определение фенантрена
Для экстракции фенантрена из почвы использовали метод ускоренной растворительной экстракции (ASE) [29,30]. Вкратце, 1 г каждого лиофилизированного образца почвы гомогенизировали с 4 г диатомовой земли для подготовки к ASE и подвергали экстракции гексаном/ацетоном (4:1, об./об.) с использованием экстрактора Dionex ASE 350 (ThermoFisher Scientific, Уилмингтон, NC, США). Удельные параметры экстракции: давление газа-носителя 0,8 МПа, давление в экстракторе 1500 psi, температура экстракции 100 °C, время нагрева 5 мин, время статической экстракции 5 мин, объем растворителя, составляющий 60% от 34 мл экстракционной ячейки, интервал продувки азотом 60 с и 2 цикла экстракции. Затем полученные экстракты упаривали до объема примерно 1 мл с помощью роторного вакуум-испарителя (RE-52, Qingdao Mingbolm, Циндао, Китай) с последующей очисткой на колонке для твердофазной экстракции с силикагелем (CNWBOND, Shanghai Anpel Laboratory Technologies Inc., Шанхай, Китай). Элюирование с колонки проводили с использованием 12 мл смеси гексан/дихлорметан (1:1, об./об.). Этот элюат затем упаривали досуха под слабым потоком азота, восстанавливали в 1 мл метанола в качестве конечного объема и добавляли 200 нг фенантрен-d10, подготавливая его для последующего анализа методом высокоэффективной жидкостной хроматографии (ВЭЖХ).
Приблизительно 10 мкл образца использовали для исследования концентрации фенантрена в экстракте с помощью системы ВЭЖХ Dionex UltiMate 3000 (ThermoFisher Scientific, Уилмингтон, NC, США), оборудованной аналитической колонкой с обращенной фазой Agilent ZORBAX Eclipse Plus C18 (5 мкм, 4,6 мм × 250 мм) и УФ-детектором. Подвижная фаза представляла собой смесь ацетонитрила и деионизированной воды (80:20, об./об.), протекающую со скоростью 1 мл/мин. Детектирование фенантрена проводили при УФ-длине волны 254 нм, и количественное определение его концентрации основывалось на калибровке по внутреннему стандарту. Анализ показал средний процент извлечения фенантрена из почвы 96,0 ± 1,6% и предел обнаружения (LOD) 5,4 нг/г.
2.4. Углерод микробной биомассы почвы
После 21-дневного периода инкубации содержание углерода микробной биомассы в почве оценивали с помощью метода экстракции после обработки хлороформом [31,32]. Свежие образцы почвы массой 10 г каждый получали из различных повторностей вариантов, причем часть (5 г) подвергали обработке парами безэтанольного хлороформа в условиях отсутствия света в вакуум-эксикаторе в течение 24 ч. Впоследствии как обработанные, так и необработанные хлороформом почвы подвергали экстракции путем интенсивного встряхивания с 20 мл 0,5 M K2SO4 на ротационном шейкере в течение 0,5 ч. Затем экстракты фильтровали через шприцевые фильтры 0,45 мкм. Содержание органического углерода в этих фильтратах определяли с помощью анализатора TOC Multi N/C 3100 (Analytik Jena, Йена, Германия). Содержание углерода микробной биомассы вычисляли по следующему уравнению:
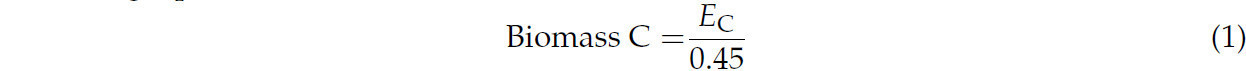
Здесь Ec (мг C/кг почвы) представляет собой разницу масс TOC, экстрагированного из обработанных и необработанных хлороформом почв.
2.5. Активность почвенных ферментов
Полифенолоксидаза (PPO), присутствующая в различных бактериях, грибах и растительных тканях, может катализировать превращение фенолов, полученных при окислительном метаболизме ПАУ, в хиноны [33,34]. Активность PPO во влажной почве после 21 дня инкубации измеряли с использованием набора для определения активности твердофазной PPO (Beijing Solarbio Science & Technology Co., Ltd., Пекин, Китай), чтобы отразить способность и потенциал микробов разлагать ПАУ [35,36]. К 0,1 г воздушно-сухой почвы добавляли 500 мкл реакционного субстрата и инкубировали при 30 °C в течение 1 ч. Затем добавляли 200 мкл буферного раствора и 1750 мкл эфира для экстракции на вортексе. После отстаивания смеси при комнатной температуре в течение 0,5 ч 1 мл надосадочной жидкости анализировали с помощью УФ-видиого спектрофотометра (UV-2700, Shimadzu, Киото, Япония) при длине волны 430 нм. Активность PPO рассчитывали по следующей формуле:
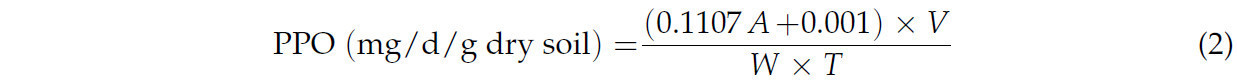
Здесь A представляет значение поглощения, T — время реакции (1 ч = 1/24 д), V — общий объем реакции (1,75 мл), а W — вес образца почвы (0,1 г).
2.6. Высокопроизводительное секвенирование
Геномную ДНК экстрагировали из 0,5 г образца влажной почвы с использованием набора FastDNA® Spin Kit for Soil (MP Biomedicals, Санта-Ана, Калифорния, США) в соответствии с инструкциями производителя. Экстракт ДНК анализировали с помощью 1% агарозного гель-электрофореза, а его чистоту измеряли с помощью спектрофотометра УФ-видимого диапазона NanoDrop 2000 (Thermo Scientific, Уилмингтон, NC, США). Для анализа структуры и разнообразия бактериального сообщества универсальную праймерную пару 515F (5′-GTGCCAGCMGCCGCGG-3′) и 907R (5′-CCGTCAATTCMTTTRAGTTT-3′) использовали для амплификации V4–V5 областей гена бактериальной 16S рРНК с помощью термоциклера для ПЦР ABI GeneAmp® 9700 (Applied Biosystems, Фостер-Сити, Калифорния, США). По сравнению с праймерами 338F/806R, нацеленными на области V3–V4, праймеры, нацеленные на области V4–V5, могут минимизировать завышение оценки прокариотического разнообразия, вызванного внутригеномной гетерогенностью в гене 16S рРНК [37]. Кроме того, они эффективно обнаруживают определенные виды бактерий, такие как важная аммиак-окисляющая группа Thaumarchaeota, которая может иметь более слабую амплификацию в областях V3–V4 [38,39]. Секвенирующие библиотеки создавали с использованием набора TruSeqTM DNA Sample Prep Kit (Illumina, Сан-Диего, Калифорния, США) в соответствии с протоколами производителя. Высокопроизводительное секвенирование проводили на платформе Illumina MiSeq PE300 (Сан-Диего, Калифорния, США), предоставленной Majorbio Bio-Pharm Technology Co., Ltd. (Шанхай, Китай). Детальные процедуры ПЦР-амплификации, очистки и обработки данных секвенирования в соответствии со стандартными протоколами обобщены в Дополнительных материалах. Все последовательности, нацеленные на ген 16S рРНК, были сгруппированы в операционные таксономические единицы (OTU) с порогом сходства 97% с использованием USEARCH-UPARSE (версия 11) [40], и любые химерные последовательности были идентифицированы и впоследствии исключены. Таксономическую классификацию репрезентативной последовательности каждого OTU проводили с помощью классификатора RDP (версия 2.13) по последней базе данных Silva 16S рРНК (версия 138) с порогом достоверности 70% [41,42].
Альфа-разнообразие, выраженное как индекс Шеннона, анализировали с помощью Mothur (версия 1.30.2) для оценки выравненности и богатства видов в почвенных бактериальных сообществах. Расстояние Брея–Куртиса, представляющее бета-разнообразие, рассчитывали с использованием QIIME 2 (версия 2024.2) для определения вариации структуры бактериального сообщества различных групп обработки по сравнению с контролем. Анализ главных координат (PCoA) с использованием взвешенного расстояния UniFrac был выполнен с помощью пакета R vegan (v2.6-4) для визуализации сходств и различий между почвенными бактериальными сообществами. Избыточный анализ (RDA) был выполнен с использованием пакета vegan v2.6-4 для интуитивного демонстрирования относительного влияния экологических факторов на почвенные бактериальные сообщества.
2.7. Количественная ПЦР
Количества гена бактериальной 16S рРНК и ключевого гена деградации ПАУ (nidA) измеряли с помощью количественной ПЦР в реальном времени (qPCR). Амплификацию гена 16S рРНК достигали с использованием прямого праймера Eub338F (5′-ACTCCTACGGGAGGCAGCAG-3′) и обратного праймера Eub518R (5′-ATTACCGCGGCTGCTGG-3′). Амплификацию гена nidA достигали с использованием прямого праймера nidAF (5′-TTCCCGAGTACGAGGGATAC-3′) и обратного праймера nidAR (5′-TCACGTTGATGAACGACAAA-3′). Серийные 10-кратные разведения рекомбинантной плазмиды (pMD18-T), содержащей амплифицированный фрагмент либо гена 16S рРНК (4,49 × 10^8 копий/мкл), либо гена nidA (4,21 × 10^8 копий/мкл), готовили для построения стандартной кривой для последующего количественного определения. Реакционная смесь для qPCR состояла из 5 мкл 2× ChamQ SYBR Color qPCR Master Mix (Vazyme Biotech, Нанкин, Китай), 0,4 мкл каждого праймера (5 мкМ), 0,2 мкл 50× ROX Reference Dye, 1 мкл матрицы ДНК и 3 мкл ddH2O, что давало конечный объем 10 мкл. qPCR проводили на системе ABI 7300 Real-Time PCR (Applied Biosystems, Фостер-Сити, Калифорния, США) со следующим термическим профилем: начальная выдержка при 95 °C в течение 10 мин, затем 40 циклов денатурации при 95 °C в течение 15 с, отжиг при 58 °C в течение 30 с и элонгация при 72 °C в течение 30 с. ( Прим. перев.: в оригинале, вероятно, опечатка: "30 min" должно быть "30 s" ).
2.8. Статистический анализ
Статистические анализы проводили с использованием SPSS Statistics 20 с порогом значимости p < 0,05. Значимые различия между группами статистически анализировали с помощью однофакторного дисперсионного анализа (ANOVA) с последующим тестом множественных сравнений Тьюки. Значимые корреляции определяли с помощью корреляционного анализа Спирмена с использованием пакета R psych (v2.1.3).
3. Результаты и обсуждение
3.1. Биодеградация фенантрена аутохтонными почвенными микроорганизмами
Проценты биодеградации фенантрена аутохтонными микроорганизмами в почве, обработанной и необработанной биоуглем из соломы пшеницы после 21 дня инкубации, представлены на Рисунке 1. Доля микробной деградации фенантрена в сильно загрязненной почве (PHS: 32,95–50,53%) была значительно ниже, чем в слабо загрязненной почве (PLS: 54,39–77,75%). Это несоответствие, предположительно, связано с токсичностью высоких концентраций фенантрена для микроорганизмов, что ингибирует их рост и активность, о чем свидетельствует снижение обилия и разнообразия бактериального сообщества (Рисунок 2b и Рисунок 3a). Не было значительных различий в биодеградации фенантрена между почвой, обработанной W500, и контролем (PH) (Рисунок 1). Внесение W300 значительно способствовало биодеградации фенантрена (p < 0,05). По сравнению с контролем (PL), обработка 1%W300 не оказала значительного влияния на биодеградацию фенантрена. Биодеградация фенантрена была явно подавлена после применения 2%W300, 1%W500 и 2%W500 (p < 0,05). Для определенного биоугля более высокие уровни внесения приводили к более низким остаткам фенантрена в PHS; однако в PLS повышенные уровни внесения приводили к более высоким остаткам фенантрена (Рисунок 1). Это изменение можно объяснить различной ролью биоугля из соломы пшеницы в почвах с разным уровнем загрязнения. В PLS более высокие уровни внесения определенного биоугля приводили к более сильной сорбции и иммобилизации фенантрена, чем более низкие уровни (Таблица S2), уменьшая биодоступную фракцию фенантрена для микробной деградации и, как следствие, снижая его уменьшение (Рисунок 1). Это последовательно suggests, что значительное подавление биодеградации фенантрена W500 в PLS было в первую очередь связано с его усиленной сорбцией (p < 0,05, Рисунок 1). И наоборот, поскольку PHS содержала более высокую долю несвязанного фенантрена, чем PLS, биоуголь, внесенный в PHS, оказывал относительно более слабые ограничения на биодоступность фенантрена. Исследуемый биоуголь в PHS играл более значительную роль в поставке необходимых питательных веществ (Таблица S3), таких как общий и доступный азот и фосфор, таким образом стимулируя рост микробов, а не усиливая сорбцию и ингибируя биодоступность [43]. В результате внесение определенного биоугля на более высоких уровнях может быть более эффективным для биодеградации фенантрена, чем на более низких уровнях (Рисунок 1). В соответствии с этим, только W300, который содержал больше питательных веществ и приводил к более высокому содержанию N и P в почве после внесения, чем W500 (Таблицы S1 и S3), значительно усиливал микробную деградацию фенантрена в PHS (p < 0,05, Рисунок 1).
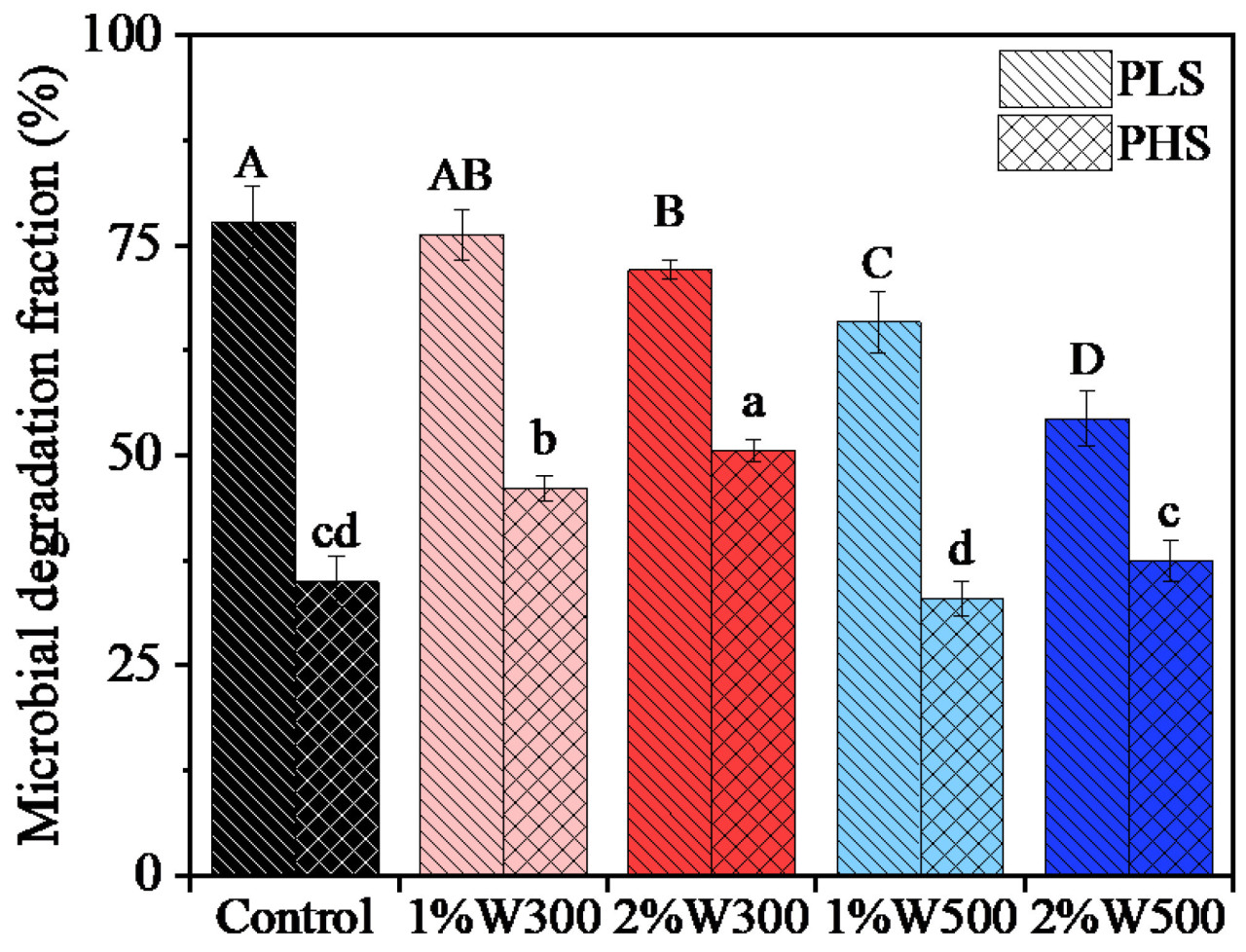
Рисунок 1. Процент биодеградации фенантрена аутохтонными микроорганизмами в почве при различных вариантах применения биоугля после 21 дня инкубации. PLS и PHS обозначают слабо и сильно загрязненную фенантреном почву соответственно. Control: без внесения биоугля. W300 и W500 обозначают биоугли из соломы пшеницы, полученные при 300 и 500 °C соответственно, а 1% и 2% — уровни внесения биоугля. Разные буквы обозначают значимые различия между вариантами при p < 0,05.
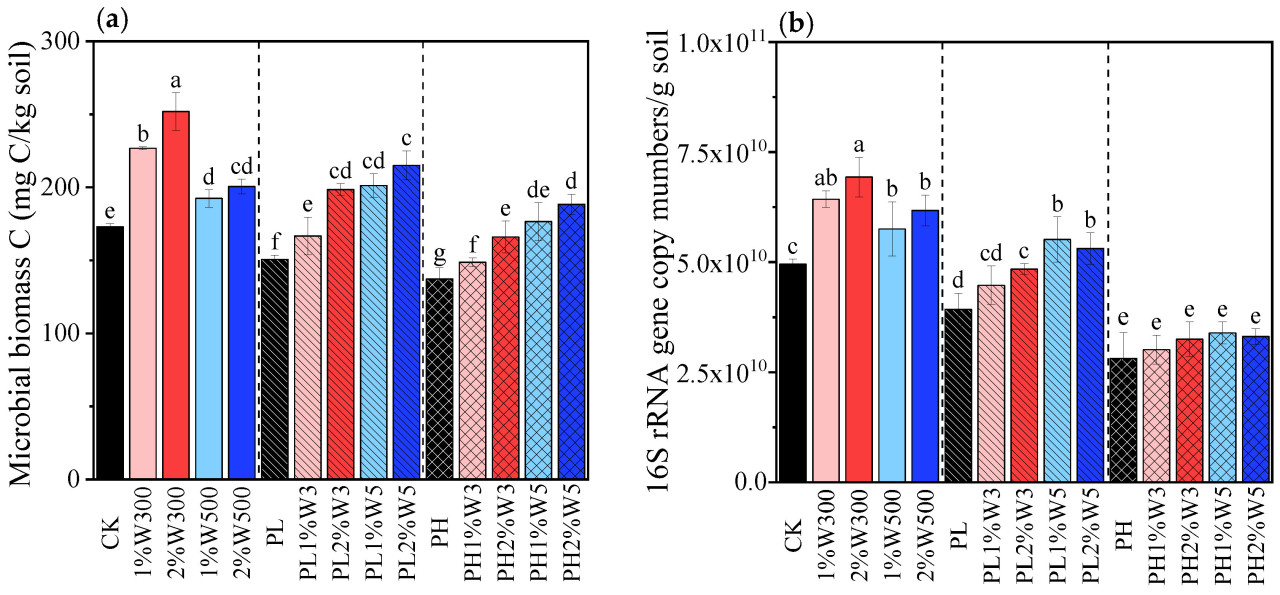
Рисунок 2. Содержание углерода микробной биомассы (a) и общее количество бактерий (b) в незагрязненной, слабо (PLS) и сильно (PHS) загрязненной почве при различных вариантах обработки биоуглем после 21 дня инкубации. CK: исходная почва без обработки; PL и PH: почвы, загрязненные низкой (2,5 мг/кг) и высокой (25 мг/кг) концентрациями фенантрена соответственно; 1%Wn00 и 2%Wn00: чистые почвы с внесением 1% и 2% биоугля из соломы пшеницы соответственно; PL/PH + 1%Wn и 2%Wn: слабо/сильно загрязненные почвы с внесением 1% и 2% биоугля из соломы пшеницы соответственно; «n»: температуры обработки 300 и 500 °C. Разные буквы обозначают значимые различия между вариантами при p < 0,05.
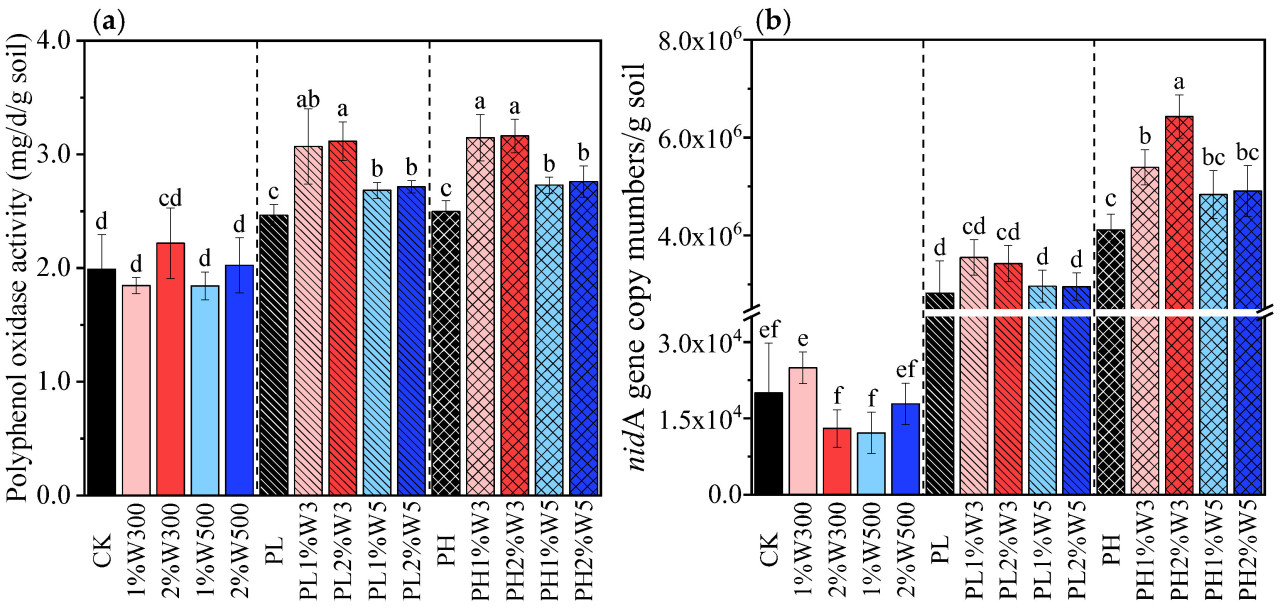
Рисунок 3. Активность полифенолоксидазы (a) и количество гена nidA (b) в незагрязненной, слабо (PLS) и сильно (PHS) загрязненной почве при различных вариантах обработки биоуглем после 21 дня инкубации. CK: исходная почва без обработки; PL и PH: почвы, загрязненные низкой (2,5 мг/кг) и высокой (25 мг/кг) концентрациями фенантрена соответственно; 1%Wn00 и 2%Wn00: чистые почвы с внесением 1% и 2% биоугля из соломы пшеницы соответственно; PL/PH + 1%Wn и 2%Wn: слабо/сильно загрязненные почвы с внесением 1% и 2% биоугля из соломы пшеницы соответственно; «n»: температуры обработки 300 и 500 °C. Разные буквы обозначают значимые различия между вариантами при p < 0,05.
3.2. Численность и активность почвенных микроорганизмов
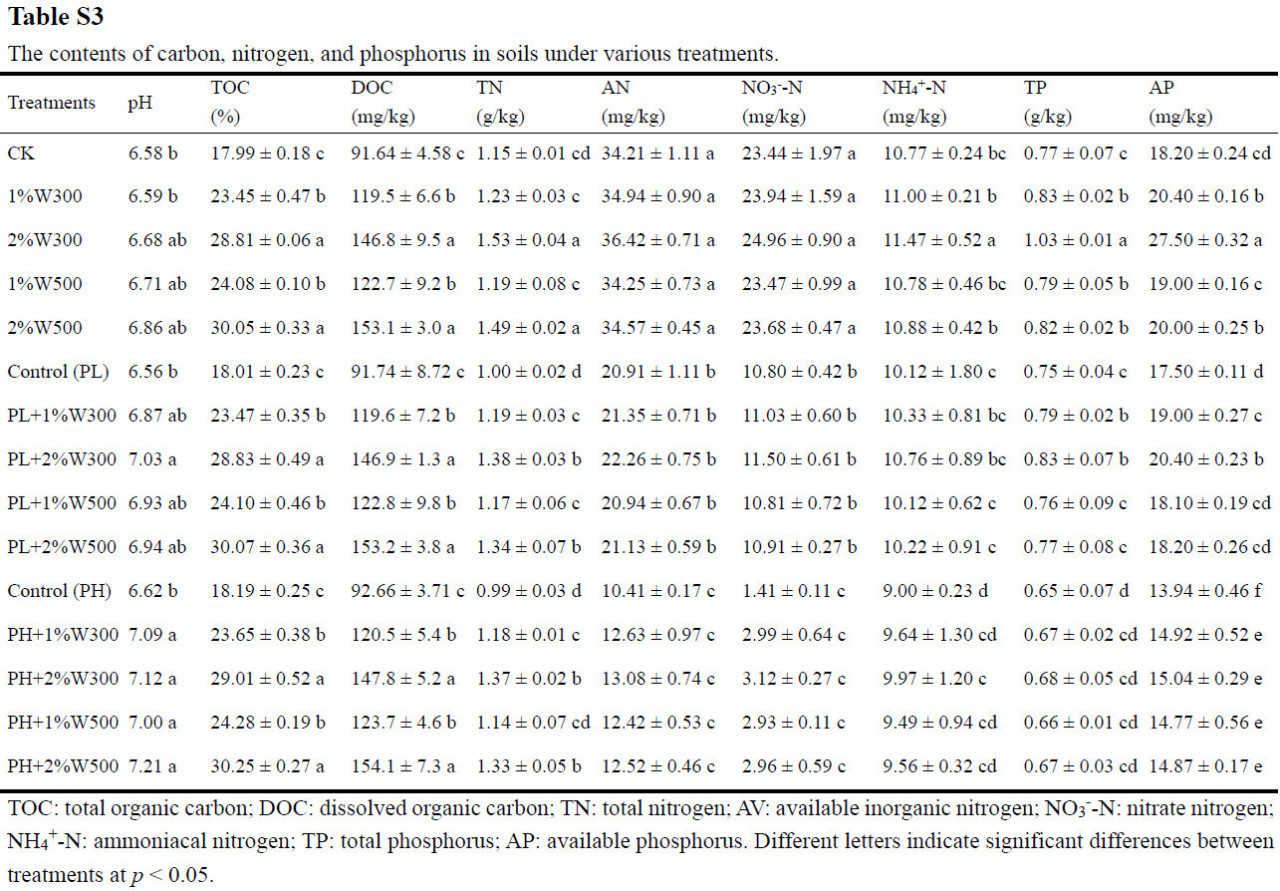
Внесение биоугля в незагрязненную почву вызвало значительное увеличение (13,51–55,09%) содержания углерода микробной биомассы (Рисунок 2a), поскольку он выступил в качестве трудноразлагаемого углеродного субстрата и источника необходимых питательных веществ (N и P) для роста всего микробного сообщества [44,45]. Наблюдалось заметное увеличение содержания C, N и P в почве, включая как общие, так и доступные фракции (например, TOC, DOC, TN, растворенный неорганический NH4+-N, TP и AP), в ответ на внесение биоугля (Таблица S3). Более того, улучшение питательного режима было более выраженным при внесении W300 по сравнению с W500, что привело к большему увеличению углерода микробной биомассы в почве, обработанной W300, по сравнению с почвой, обработанной W500 (Рисунок 2a). Однако другие исследователи обнаружили, что биоуголь не оказывал значительного влияния на численность микробов/бактерий или состав сообщества в пахотных почвах [46,47,48]. Эти противоречивые данные могут проистекать из различий в типе и дозе биоугля, физико-химических характеристиках почвы, продолжительности воздействия и экспериментальных условиях в этих исследованиях. Предыдущее исследование показало, что биоуголь, полученный газификацией древесины, был довольно устойчивым, и его внесение в высоких дозах (60 т/га) в слабощелочную почву в ходе годичного полевого эксперимента значительно увеличивало содержание TOC в почве, но не влияло на содержание TN и AP, и, следовательно, не способствовало увеличению численности микробов и активности ферментов [19]. Это доказывает превосходные питательные свойства биоугля из соломы, использованного в данном исследовании, вероятно, обусловленные поглощением и удержанием ключевых питательных веществ, таких как N и P, в течение жизненного цикла травянистых растений [49]. Этот эффект улучшения питательного режима имеет ключевое значение для стимулирования роста микробов в почве по сравнению со способностью связывания углерода у древесного биоугля с более высоким содержанием углерода [50].
Содержание углерода микробной биомассы в почве значительно снижалось с увеличением стресса от фенантрена (Рисунок 2a), что указывает на токсическое воздействие фенантрена на почвенные микроорганизмы. Примечательно, что токсический стресс, вызванный фенантреном для почвенных микробных сообществ, ослаблялся при внесении биоугля из соломы пшеницы: содержание углерода микробной биомассы в почве, обработанной биоуглем, увеличивалось на 13,31–53,35% и 10,62–47,56% по сравнению с контролем (PL и PH) соответственно. Помимо стимуляции питательными веществами, это наблюдение можно в первую очередь объяснить исключительными сорбционными свойствами биоугля из соломы, обусловленными высоким содержанием лигнина в соломе сельскохозяйственных культур, что способствует созданию пористой структуры, введению функциональных групп (например, гидроксильных и альдегидных) и увеличению гидрофобности [51,52,53], тем самым снижая биотоксичность фенантрена. Среди tested биоуглей W500 показал себя лучше, чем W300, в увеличении углерода биомассы всего микробного сообщества (Рисунок 2a), вероятно, благодаря его большей сорбционной силе и емкости (Таблица S2); это проистекает из его более высокой ароматичности, площади поверхности и пористости, а также объемной и поверхностной гидрофобности (Таблица S1), что позволяет ему более эффективно снижать биодоступность и токсичность фенантрена в почве. Результаты данного исследования демонстрируют, что иммобилизующий эффект W500 на фенантрен более значителен, чем у W300, что может объяснять, почему доля биодеградации фенантрена в вариантах с W500 была значительно ниже, чем в вариантах с W300, независимо от уровня загрязнения (Рисунок 1). Кроме того, углерод микробной биомассы в загрязненной почве показал положительную корреляцию с увеличением уровня внесения определенного соломенного биоугля (Рисунок 2a) из-за увеличения содержания трудноразлагаемых (ароматических) углеродных доменов [54], что позволяет усилить сорбцию ароматических соединений (т.е. фенантрена) в почве и уменьшить их неблагоприятное воздействие на популяции микробов [55,56]. Ослабление токсичности загрязнителей для почвенных микробных сообществ с помощью биоугля сообщалось в предыдущих исследованиях [57,58,59]. Было обнаружено, что биоуголь способствовал увеличению численности, разнообразия и функциональности микробов в почве, загрязненной углеводородами, включая бактериальные и грибковые сообщества, подверженные воздействию таких веществ, как нефть и ПАУ [17,60].
Количество копий гена бактериальной 16S рРНК в незагрязненной почве также значительно увеличилось после внесения биоугля (Рисунок 2b). Стресс от фенантрена вызвал существенное снижение количества гена 16S рРНК, и этот эффект был особенно заметен при высоком уровне загрязнения (p < 0,05), что согласуется с результатами, наблюдаемыми для углерода микробной биомассы (Рисунок 2a). В слабо загрязненной почве количество бактериального гена заметно увеличилось в ответ на внесение биоугля, и стимулирующий эффект был более выраженным у W500 (p < 0,05); однако в сильно загрязненной почве значительных изменений не наблюдалось (Рисунок 2b). Стимулирование углерода микробной биомассы и общей численности бактерий, вызванное внесением биоугля, было более значительным в PLS, чем в PHS. Это различие может быть связано с тем, что доля подвижного фенантрена была намного ниже в PLS, чем в PHS [29]. Внесение биоугля в PLS снижало биодоступность фенантрена и смягчало его токсическое воздействие в большей степени за счет сорбции и иммобилизации, что оказалось более выгодным для общего роста микробных/бактериальных сообществ, чем его влияние на PHS.
PPO служит критически важным показателем окислительного потенциала и биохимической трансформационной активности в почвенных микробных сообществах. Этот фермент можно использовать для оценки микробных метаболических процессов, участвующих в аэробном разложении трудноразлагаемых ароматических соединений, включая ПАУ. Как показано на Рисунке 3a, внесение биоугля не оказало значительного влияния на активность PPO в незагрязненной почве, тогда как в слабо и сильно загрязненных почвах активность PPO значительно стимулировалась внесением биоугля. Более того, активность PPO в почве была заметно выше в вариантах с W300, чем в вариантах с W500. Эта тенденция согласуется с тем, что доля биодеградации фенантрена была больше в почве, обработанной W300, чем в почве, обработанной W500, независимо от уровня загрязнения (Рисунок 1), что указывает на то, что активность этого фермента коррелирует с эффективностью микробной деградации стойких органических соединений (например, ПАУ).
3.3. Разнообразие и структура бактериального сообщества почвы
Стресс от фенантрена значительно снизил альфа-разнообразие почвенного бактериального сообщества, выраженное индексом Шеннона (Рисунок 4a), что согласуется с предыдущими исследованиями [61,62]. Когда биоуголь из соломы пшеницы вносили как в слабо, так и в сильно загрязненные почвы, индексы альфа-разнообразия демонстрировали более высокие значения, чем те, которые наблюдались только при загрязнении фенантреном (Рисунок 4a). Это указывает на то, что применение биоугля смягчило снижение разнообразия бактериального сообщества почвы, вызванное загрязнением фенантреном. Аналогичные результаты были также получены в другом исследовании, где добавление биоугля из соломы пшеницы в почву, загрязненную ПАУ, помогло сохранить разнообразие бактериального сообщества [43]. Примечательно, что биоуголь, внесенный отдельно в незагрязненную почву, не оказал значительного влияния на альфа-разнообразие бактерий (Рисунок 4a). Следовательно, благотворное влияние биоугля на поддержание разнообразия бактериального сообщества в загрязненной почве можно в первую очередь объяснить его способностью сорбировать токсичные соединения, тем самым ослабляя токсический стресс для почвенного бактериального сообщества, вызванный загрязнителями (т.е. фенантреном), а не обеспечением питательными веществами или местами обитания для почвенных бактерий или изменением физико-химических свойств почвы [63,64]. Кроме того, благодаря превосходной сорбционной силе и емкости по сравнению с W300, W500 более эффективно снижал биодоступность и токсичность фенантрена, что, как следствие, привело к более выраженному увеличению альфа-разнообразия бактерий в загрязненной почве по сравнению с W300 (Рисунок 4a). Это положительное влияние усиливалось с увеличением уровня внесения конкретного биоугля. Увеличение индекса Шеннона свидетельствует о более разнообразном бактериальном сообществе. Однако Li и др. обнаружили, что внесение 1% (по массе) биоугля из соломы кукурузы, полученного при 500 °C, в почву, загрязненную фенантреном, не вызвало значительных изменений индекса Шеннона, но снизило видовое богатство бактериального сообщества, описываемое индексом Чао [17]. Результаты этого исследования противоречат результатам нашего исследования относительно влияния биоугля на разнообразие почвенных бактерий, главным образом из-за различных типов исходного сырья для биоугля. В упомянутом исследовании использовалась солома кукурузы, которая имеет более низкое содержание лигнина, чем солома пшеницы, использованная в нашем исследовании [65], что приводит к получению биоугля с более высокой устойчивостью к разложению [66]. Это подтверждается увеличением соотношения C/N в почве в результате применения биоугля из соломы кукурузы [67], что, в свою очередь, подавляло рост специфических бактериальных популяций в почве.
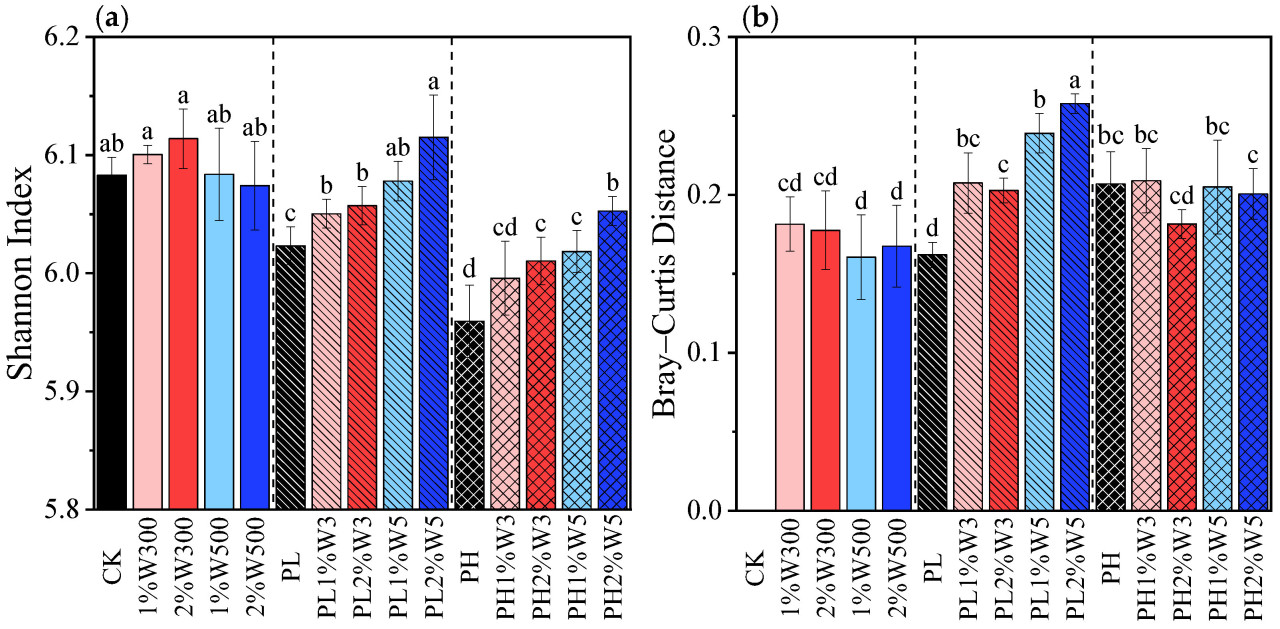
Рисунок 4. Разнообразие бактериального сообщества в незагрязненной, слабо (PLS) и сильно (PHS) загрязненной почве при различных вариантах обработки биоуглем после 21 дня инкубации. (a) Альфа-разнообразие, измеренное с помощью индекса Шеннона; (b) бета-разнообразие, измеренное с помощью расстояния Брея–Куртиса между каждой обработкой и CK. CK: исходная почва без обработки; PL и PH: почвы, загрязненные низкой (2,5 мг/кг) и высокой (25 мг/кг) концентрациями фенантрена соответственно; 1%Wn00 и 2%Wn00: чистые почвы с внесением 1% и 2% биоугля из соломы пшеницы соответственно; PL/PH + 1%Wn и 2%Wn: слабо/сильно загрязненные почвы с внесением 1% и 2% биоугля из соломы пшеницы соответственно; «n»: температуры обработки 300 и 500 °C. Разные буквы обозначают значимые различия между вариантами при p < 0,05.
Оба биоугля в tested дозах в слабо загрязненной почве заметно увеличили бета-разнообразие почвенного бактериального сообщества, о чем свидетельствует большее расстояние Брея–Куртиса по сравнению с контролем (PL) (p < 0,05, Рисунок 4b), что подразумевает сдвиг в структуре почвенного бактериального сообщества из-за применения биоугля. Кроме того, что касается различий между почвенными бактериальными сообществами после обработки биоуглем и контролем (PL), то различия, вызванные W500, были больше, чем вызванные W300. Однако не было значительных различий в структуре бактериального сообщества между различными вариантами обработки биоуглем в сильно загрязненной почве и контролем (PH) (Рисунок 4b), что согласуется с имеющейся литературой [43,68]. Эти находки указывают на то, что чистое влияние биоугля на структуру бактериального сообщества было значительным в PLS, но минимальным в PHS. Это различие также можно объяснить большим снижением биодоступности фенантрена и его токсичности для почвенных бактерий, вызванным внесением биоугля в PLS, где доля несвязанного фенантрена была ниже, чем в PHS [29].
На основе анализа RDA, почвенные бактериальные сообщества в отсутствие фенантрена после различных обработок биоуглем (т.е. 1%W300, 2%W300, 1%W500 и 2%W500) не показали отчетливого отделения от CK и не отличались друг от друга (Рисунок 5), что раскрывает, что структура бактериального сообщества в незагрязненной почве не была значительно затронута только внесением биоугля. Стресс от фенантрена сильно сместил структуру почвенного бактериального сообщества, что отразилось в результатах RDA, которые показали, что бактериальные сообщества с фенантреном образовывали отдельные кластеры от сообществ без фенантрена, независимо от присутствия биоугля. Это различие было особенно evident при высоких уровнях загрязнения (Рисунок 5). Анализ RDA далее показал, что резкие сдвиги в структуре бактериального сообщества, вызванные стрессом от фенантрена, опосредовались внесением биоугля в почву, о чем свидетельствуют тупые углы между этими двумя атрибутами (т.е. PHE и BC), показывающие отрицательную корреляцию между эффектами биоугля и стресса от фенантрена на структуру бактериального сообщества [69]. Кроме того, острые углы наблюдались между влиянием свойств почвы и биоугля (Рисунок 5), что позволяет предположить, что применение биоугля из пиролизованной соломы пшеницы с относительно низким уровнем устойчивого углерода модифицировало питательные свойства почвы, specifically TOC, TN и NO3−-N, тем самым модулируя структуру бактериального сообщества в загрязненной фенантреном почве. Аналогично, предыдущие исследования продемонстрировали, что применение биоугля из пиролиза соломы риса положительно влияло на структуру микробного сообщества как в незагрязненных, так и в загрязненных ПАУ почвах, обеспечивая доступные питательные вещества и подходящие места обитания для микробов и модифицируя физико-химические свойства почвы [70,71]. В отличие от полевого исследования, предполагающего положительное влияние биоугля из газификации соломы (2,773 и 16,16 т/га) на питательные свойства почвы и бактериальные популяции [66], Baldoni и др. сообщили об отсутствии значительных изменений в содержании TN и AP в почве после применения биоугля из газификации древесины в полевых испытаниях при существенно высокой дозе 60 т/га [19]. Это несоответствие в результатах исследований можно объяснить более высоким уровнем устойчивости древесного биоугля по сравнению с соломенным, который играет большую роль в увеличении содержания TOC в почве и запасов углерода, а не в прямой стимуляции питательных веществ. Результаты данного исследования позволяют предположить, что стресс от фенантрена, внесение соломенного биоугля и содержание NO3−-N в почве являются тремя основными факторами, влияющими на структуру бактериального сообщества в сельскохозяйственной почве.
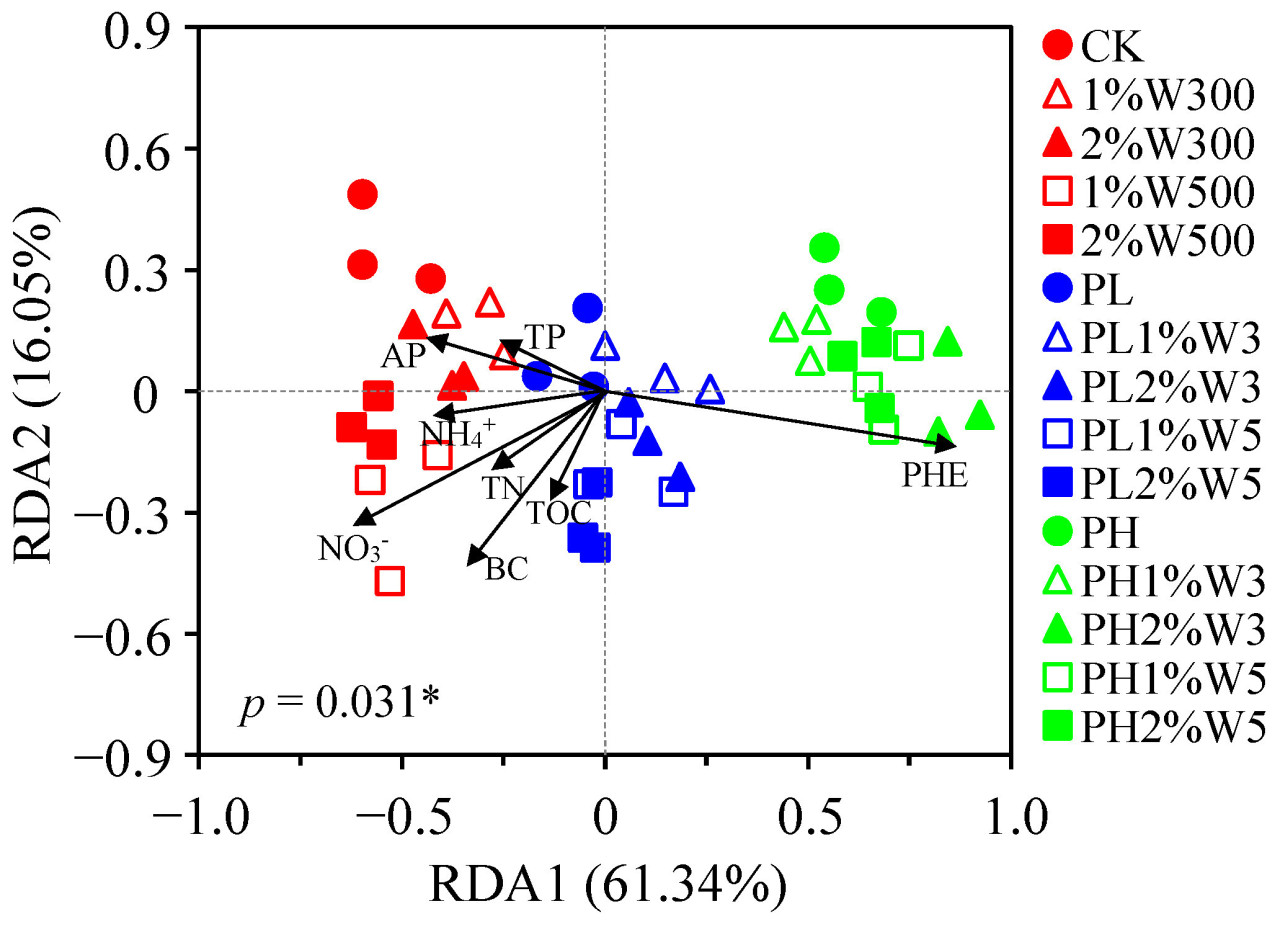
Рисунок 5. Избыточный анализ (RDA) бактериальных сообществ и переменных окружающей среды, т.е. концентрации фенантрена (PHE), внесения биоугля (BC), общего органического углерода (TOC), общего азота (TN), нитратного азота (NO3−-N), аммонийного азота (NH4+-N), общего (TP) и доступного фосфора (AP) в почве, обработанной биоуглем из соломы пшеницы после 21 дня инкубации. CK: исходная почва без обработки; PL и PH: почвы, загрязненные низкой (2,5 мг/кг) и высокой (25 мг/кг) концентрациями фенантрена соответственно; 1%Wn00 и 2%Wn00: чистые почвы с внесением 1% и 2% биоугля из соломы пшеницы соответственно; PL/PH + 1%Wn и 2%Wn: слабо/сильно загрязненные почвы с внесением 1% и 2% биоугля из соломы пшеницы соответственно; «n»: температуры обработки 300 и 500 °C. * обозначает значимые различия при p < 0,05.
В целом, высокий стресс от фенантрена приводил к явно более разным и значительно менее разнообразным бактериальным сообществам, что позволяет предположить, что загрязнение действительно является критическим фактором формирования структуры почвенного бактериального сообщества. Внесение биоугля благоприятствовало модуляции структуры бактериального сообщества в загрязненной фенантреном почве за счет снижения биотоксичности фенантрена и улучшения питательных свойств почвы, что может быть полезным для биодеградации и разрушения фенантрена в почве.
3.4. Обилие родов бактерий-деструкторов и функциональных генов в почве
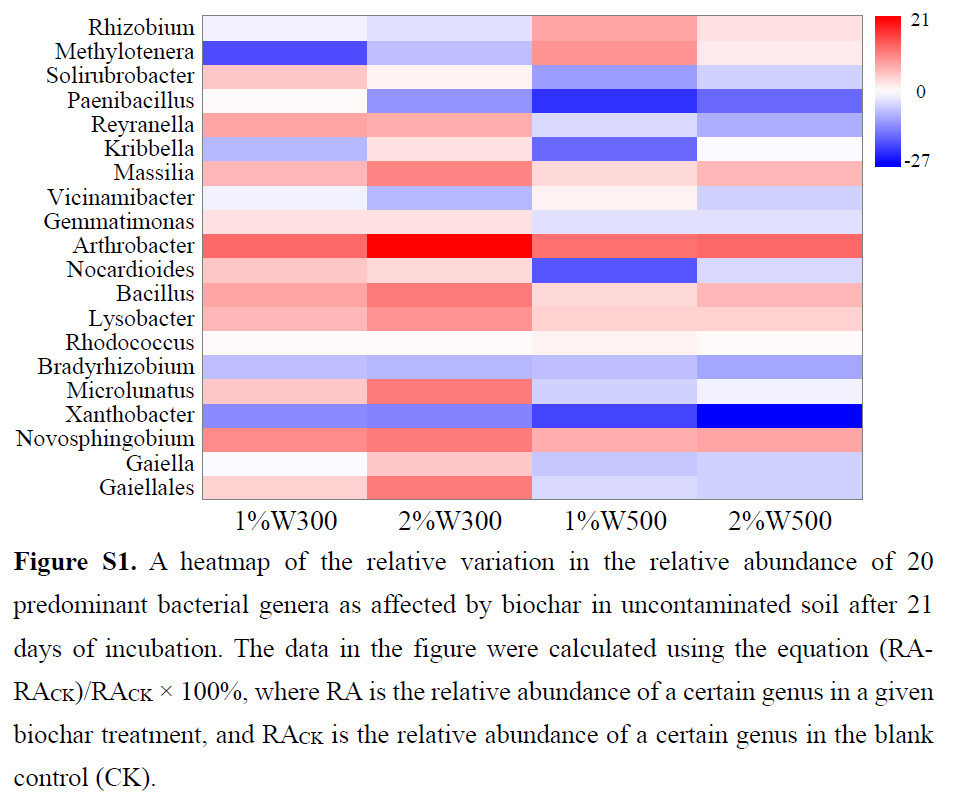
Всего в каждой обработанной почве было идентифицировано 456 бактериальных родов, 20 из которых показали относительную численность, превышающую 1%, что составляло приблизительно 49,62–60,28% от всей бактериальной популяции. Среди 20 наиболее распространенных бактериальных родов подмножество гетеротрофных бактерий (т.е. Arthrobacter) было в основном обогащено в незагрязненной почве после применения биоугля из соломы пшеницы (Рисунок S1), что, возможно, было обусловлено их лучшей утилизацией доступных углеродных ресурсов (например, устойчивого лигнина), полученных из биоугля [72]. Совместный метаболизм природного почвенного углерода и углерода, полученного из биоугля, наблюдался для обогащения определенных видов бактерий [73]. Девять из этих преобладающих бактериальных родов, таких как Lysobacter, Massilia, Bacillus, Reyranella и Gaiellales, были dramatically обогащены (15–65%) под воздействием стресса от фенантрена (Рисунок 6a, c). В то же время восемь бактериальных родов, таких как Paenibacillus, Arthrobacter и Kribbella, были значительно подавлены (15–25%) стрессом от загрязнения (Рисунок 6a, c). Несколько бактериальных родов (например, Rhizobium, Arthrobacter, Nocardioides, Paenibacillus и Gemmatimonas) продемонстрировали более выраженное снижение в контроле PH по сравнению с контролем PL (Рисунок 6a, c), что указывает на токсический стресс; их количество увеличивалось после внесения биоугля в загрязненную почву (выделено красным на Рисунке 6b, d). Это явление implies, что биоуголь способствовал выживанию этих вышеупомянутых бактериальных родов, которые были подвержены стрессу от фенантрена, в основном за счет усиления сорбции и ослабления биотоксичности. И наоборот, некоторые отдельные роды, включая Bacillus, Lysobacter, Massilia, Novosphingobium и Vicinamibacter, были более обогащены в контроле PH по сравнению с контролем PL (Рисунок 6a, c), что signifies степень устойчивости к стрессу. Эти устойчивые к токсичности роды, известные своей способностью разлагать ПАУ в почвенных системах [74,75,76], как было обнаружено, процветают в условиях повышенного стресса от фенантрена, поскольку это соединение служило источником углерода, поддерживающим их пролиферацию в бактериальных сообществах.
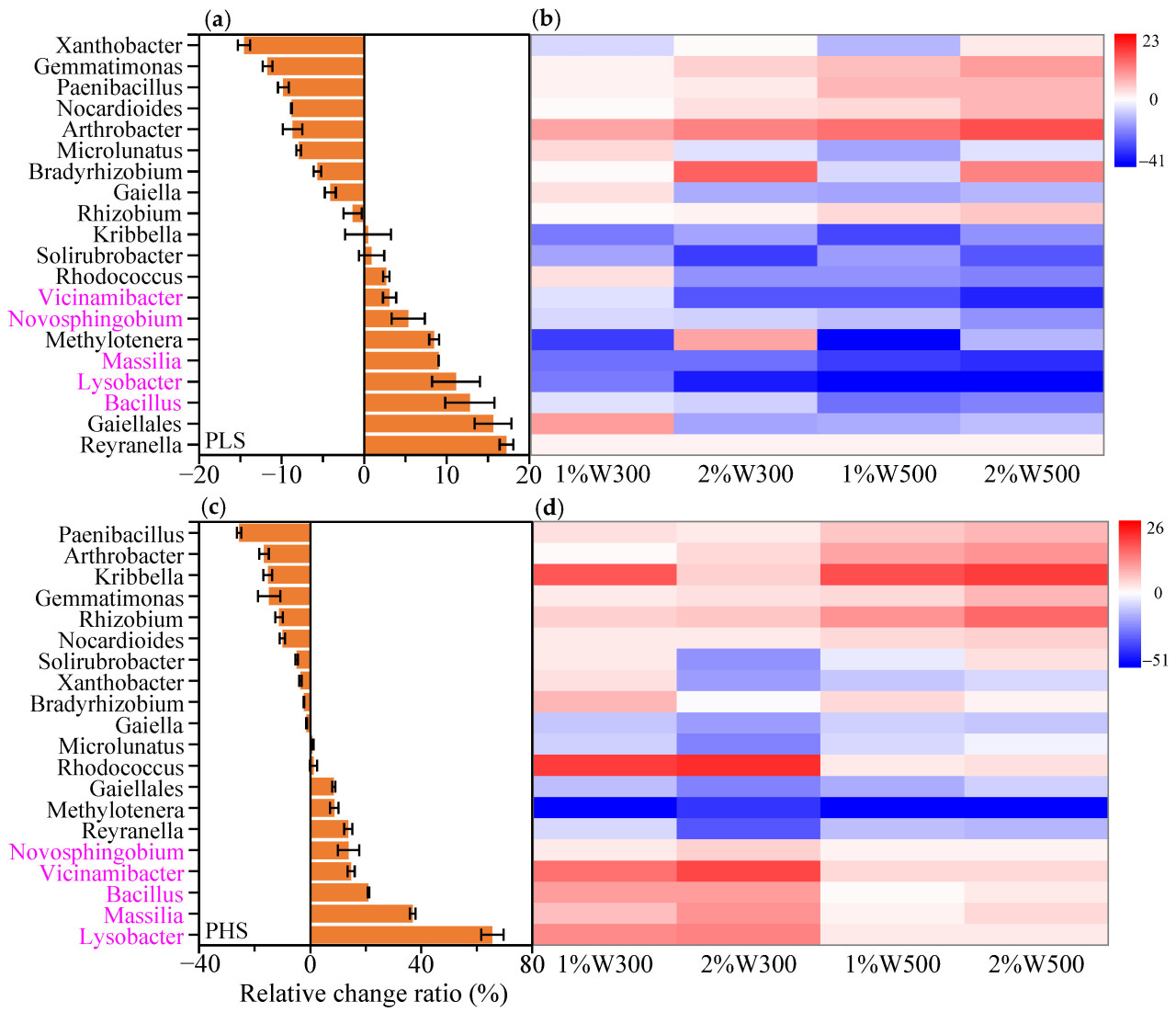
Рисунок 6. Процентное изменение относительной численности 20 основных бактериальных родов в почве после загрязнения низкой (a) и высокой (c) концентрациями фенантрена в течение 21 дня, и тепловая карта относительной скорости изменения относительной численности этих родов под влиянием биоугля в слабо (b) и сильно (d) загрязненных почвах после инкубации в течение 21 дня. Данные были рассчитаны по формуле (RA-RAcontrol)/RAcontrol × 100%, где RA и RAcontrol — относительная численность определенного рода в данном варианте обработки биоуглем и в контроле (CK или контроль (PL/PH)) соответственно. W300 и W500 обозначают биоугли из соломы пшеницы, полученные при 300 и 500 °C соответственно, а 1% и 2% — уровни внесения биоугля. Розово-красным цветом выделены бактериальные роды, предположительно являющиеся потенциальными деструкторами.
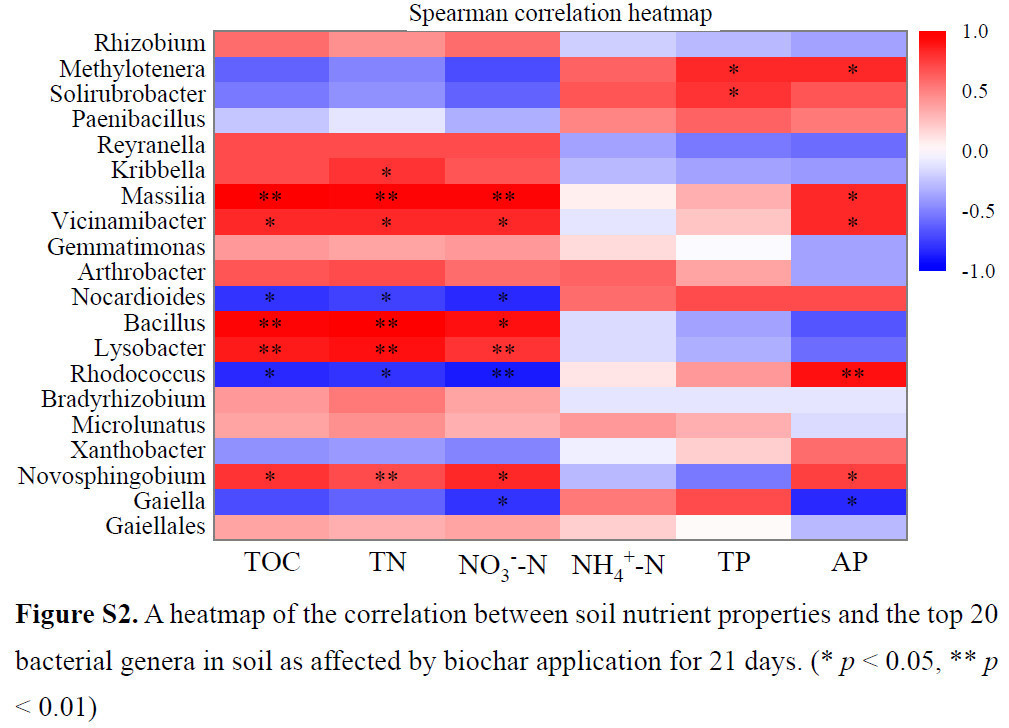
Реакция потенциальных деструкторов ПАУ на биоуголь различалась между слабо и сильно загрязненными фенантреном почвами. Специфические роды-деструкторы, которые показали большое обогащение due to внесению биоугля в PHS (красный на Рисунке 6d), были подавлены в PLS при внесении биоугля (синий на Рисунке 6b). Это различие можно объяснить distinct функциями, которые биоуголь выполняет в почвах, загрязненных различными концентрациями фенантрена. Хотя пять родов бактерий-деструкторов были сильно обогащены в контроле PL по сравнению с CK, биоуголь сдерживал их увеличение (Рисунок 6a, b). Учитывая, что в PLS уровни подвижного фенантрена были заметно ниже, чем в PHS, внесение биоугля в PLS накладывало более строгие ограничения на доступную фракцию фенантрена для деструктивных бактерий по сравнению с его внесением в PHS [29], что оказалось пагубным для пролиферации пяти бактериальных сообществ на уровне рода. Следовательно, их численность явно снижалась после внесения биоугля (синий на Рисунке 6b), что способствовало затруднению биодеградации фенантрена в PLS (Рисунок 1), особенно при применении W500. Аналогично, Omoni и др. наблюдали сокращение популяций микробов, разлагающих фенантрен, с увеличением добавок биоугля в почву, в первую очередь из-за снижения микробной доступности фенантрена [77]. Введение биоугля в PHS further увеличило относительную численность пяти родов-деструкторов (красный на Рисунке 6d). Более того, реакция этих пяти родов значительно увеличивалась с увеличением содержания питательных веществ в почве (т.е. TOC, TN, растворенного неорганического NO3− и AP) в результате внесения биоугля (Рисунок S2), что указывает на то, что биоуголь стимулировал expansion специфических популяций бактерий-деструкторов, косвенно улучшая питательные свойства почвы [78,79]. Последовательно, результаты RDA показали, что питательные свойства почвы (т.е. содержание TOC, TN и NO3−-N) были явно затронуты tested биоуглем из соломы пшеницы с относительно низким уровнем устойчивости [66], впоследствии влияя на структуру бактериального сообщества в исследуемой почве (Рисунок 5). Следовательно, в PHS, где уровни подвижного фенантрена, вероятно, были выше, чем в PLS, биоуголь играл более ключевую роль в обеспечении необходимыми питательными веществами (Таблица S3), создании благоприятной среды обитания для бактерий и исправлении физико-химических характеристик почвы, а не в уменьшении биодоступности субстрата для бактерий [29,80]. Эффективная биодеградация фенантрена в PHS при внесении биоугля, особенно усиленная биодеградация, наблюдавшаяся с W300 (Рисунок 1), может проистекать из его благотворного воздействия на эти роды-деструкторы. W300 был более благоприятен для роста бактерий, чем W500, в первую очередь due to его превосходному обеспечению питательными веществами (например, N и P) и более нейтральному pH (Таблица S3), что приводило к более выраженному улучшению относительной численности пяти бактериальных родов (Рисунок 6d). Эти более обогащенные роды бактерий-деструкторов в PHS при применении W300 в конечном итоге способствовали увеличению биодеградации фенантрена в вариантах с W300 по сравнению с вариантами с W500 (Рисунок 1). Внесение биоугля в PLS оказало негативное влияние на бактериальные роды, участвующие в деградации фенантрена (синий на Рисунке 6b). Тем не менее, биомасса ключевых бактерий, разлагающих ПАУ, значительно увеличилась после внесения биоугля в PHS (красный на Рисунке 6d), что объясняет уменьшение различий в биодеградации фенантрена между PLS и PHS после внесения биоугля (Рисунок 1).
Усиление стресса от фенантрена также значительно стимулировало количество PAH-деградирующего гена nidA (Рисунок 3b); он кодирует α-субъединицу диоксигеназы, гидроксилирующей кольцо ПАУ (PAH-RHDα), которая инициирует аэробный метаболизм ПАУ [81,82]. Аналогично, предыдущие исследования показали, что функциональные гены, ответственные за деградацию ПАУ, могут быть обогащены и сверхэкспрессированы при стрессе от загрязнения [83,84,85]. Количество гена nidA было further увеличено за счет применения биоугля из соломы пшеницы в загрязненной почве (Рисунок 3b). Биоуголь, внесенный в PLS, оказывал лишь незначительное или пренебрежимо малое положительное влияние на биомассу сообществ, несущих nidA. Возможное объяснение этому заключается в том, что сорбция фенантрена на биоугле снизила его способность стимулировать сообщества, несущие ген nidA. Этот неблагоприятный эффект был более глубоким при применении W500 по сравнению с применением W300, о чем свидетельствует значительно более низкое количество гена nidA в PLS, обработанной W500, по сравнению с обработанной W300 (Рисунок 3b). Это различие, вероятно, связано с более низкой биодоступностью фенантрена для потенциальных деструкторов, несущих nidA, в PLS, обработанной W500, по сравнению с обработанной W300, что является результатом того, что сродство и емкость сорбции W500 сильнее, чем у W300 (Таблица S2). Эти находки раскрывают, что количество генов деградации ПАУ в значительной степени зависит от микробной доступности ПАУ. Аналогично, Xia и др. обнаружили значительную положительную корреляцию между числом копий гена nahAc, который действует как универсальный биомаркер для бактерий, разлагающих нафталин, в донных отложениях и свободной растворенной концентрацией нафталина в поровой воде [86].
По сравнению с внесением биоугля в PLS, внесение биоугля в PHS оказалось более выгодным для положительного ответа катаболического гена-биомаркера nidA на стресс от фенантрена (Рисунок 3b). Это различие также может быть результатом distinct функций биоугля в почвах, загрязненных различными концентрациями фенантрена. В PHS tested биоуголь играл более crucial роль в питательной стимуляции и микробной пролиферации потенциальных деструкторов, несущих nidA, чем в усилении сорбции и ингибировании биодоступности, как observeds в PLS. Большее количество питательных веществ (например, N и P), предоставляемое W300 по сравнению с W500, приводило к явно более высокому количеству генов nidA в PHS, обработанной W300, по сравнению с обработками W500, все из которых были substantially больше, чем в контроле (PH) (Рисунок 3b и Таблица S3). По сравнению с контролем (PH), заметное увеличение количества деструкторов, несущих nidA, способствовало значительному облегчению биодеградации фенантрена в PHS при применении W300 (Рисунок 1). Эти находки согласуются с полученными в исследовании Ahmad и др., которое продемонстрировало, что применение 1% биоугля увеличивало количество гена nidA в загрязненном пиреном sediment, полученном из Жемчужной реки, тем самым способствуя усилению метаболизма пирена [87].
4. Выводы
В данном исследовании внесение биоугля из соломы пшеницы в загрязненную сельскохозяйственную почву положительно модулировало изменения в почвенных микробных сообществах, подвергшихся стрессу от фенантрена. В частности, биоуголь увеличивал общую биомассу микробного/бактериального сообщества, поддерживал бактериальное разнообразие и избирательно способствовал росту определенных бактериальных родов, которые были подавлены стрессом от фенантрена. Эти улучшения можно объяснить повышенной сорбцией фенантрена в почве, обработанной этим типом биоугля, что ослабляло его токсическое воздействие на почвенные микробы. Кроме того, некоторые бактериальные роды и функциональные гены, участвующие в деградации фенантрена, были обогащены стрессом от фенантрена и были further стимулированы внесением биоугля в сильно загрязненную почву. Это явление было в первую очередь связано с улучшением питательного режима почвы (например, TOC, TN, NO3−-N и AP), индуцированным биоуглем, особенно выраженным при использовании W300, который содержал относительно низкий уровень устойчивого углерода, обеспечивая больше питательных веществ и, таким образом, способствуя биодеградации и удалению фенантрена из почвы. Однако в слабо загрязненной почве применение биоугля, особенно более сорбционного W500, подавляло численность этих потенциальных родов-деструкторов и специфического PAH-деградирующего гена nidA за счет снижения биодоступности фенантрена для деструкторов, следовательно, препятствуя биодеградации фенантрена. Эти находки раскрывают, что использование биоугля, полученного из соломенных ресурсов при оптимальных температурах, для улучшения загрязненной ПАУ сельскохозяйственной почвы представляет собой жизнеспособную стратегию для гармонизации почвенных микробных экосистем и смягчения экологических рисков ПАУ в сельскохозяйственном производстве.
Дополнительные материалы
Следующие вспомогательные материалы можно загрузить по адресу: https://www.mdpi.com/article/10.3390/agriculture15010077/s1


![Обработка данных секвенирования [88,89]](https://storage.agriexpert.ru/storage/images/articles/5626/block/73115/model/xl/djxYn9BQiNA7.jpg)
Ссылки
1. Чжан, QY; Гао, M.; Сан, XH; Ван, Y.; Юань, CL; Сан, HW. Общенациональное распределение полициклических ароматических углеводородов в почве Китая и их связь с бактериальным сообществом. J. Environ. Sci. 2023 , 128 , 1–11. [ Google Scholar ] [ CrossRef ] [ PubMed ]
2. Хань, Дж.; Лян, Ю.С.; Чжао, Б.; Ван, И.; Син, Ф.Т.; Цинь, Л.Б. Географическое распределение полициклических ароматических углеводородов (ПАУ) в Китае и их источники, анализ оценки риска. Окружающая среда. Загрязнение. 2019 , 251 , 312–327. [ Google Scholar ] [ CrossRef ] [ PubMed ]
3. Сан, Дж. Т.; Пан, Л. Л.; Ли, Ж. Х.; Цзэн, К. Т.; Ван, Л. В.; Чжу, Л. З. Сравнение тепличного и открытого грунтового выращивания в Китае: характеристики почвы, загрязнение и микробное разнообразие. Environ. Pollut. 2018 , 243 , 1509–1516. [ Google Scholar ] [ CrossRef ] [ PubMed ]
4. Zheng, H.; Xing, XL; Hu, TP; Zhang, Y.; Zhang, JQ; Zhu, GH; Li, Y.; Qi, SH Сжигание биомассы в наибольшей степени способствовало риску развития рака у людей, подвергшихся воздействию ПАУ, содержащихся в почве в экономическом районе Чэнду на западе Китая. Ecotoxicol. Environ. Saf. 2018 , 159 , 63–70. [ Google Scholar ] [ CrossRef ]
5. Маллах, MA; Ли, CX; Маллах, MA; Норин, S.; Лю, Y.; Саид, M.; Си, H.; Ахмед, B.; Фэн, FF; Миржат, AA; и др. Полициклические ароматические углеводороды и их влияние на здоровье человека: обзор. Chemosphere 2022 , 296 , 133948. [ Google Scholar ] [ CrossRef ] [ PubMed ]
6. И, М.Л.; Чжан, Л.Л.; Ли, И.; Цянь, И. Структурные, метаболические и функциональные характеристики микробных сообществ почвы в ответ на стресс, вызванный бензо[а]пиреном. J. Hazard. Mater. 2022 , 431 , 128632. [ Google Scholar ] [ CrossRef ]
7. Хартманн, М.; Сикс, Дж. Структура почвы и функции микробиома в агроэкосистемах. Nat. Rev. Earth Environ. 2023 , 4 , 4–18. [ Google Scholar ] [ CrossRef ]
8. Грэм, Э.Б.; Кнельман, Дж.Э. Влияние формирования микробного сообщества почвы на восстановление экосистемы: закономерности, процессы и потенциал. Microb. Ecol. 2023 , 85 , 809–819. [ Google Scholar ] [ CrossRef ]
9. Филиппо, Л.; Чену, К.; Капплер, А.; Риллиг, М.К.; Фирер, Н. Взаимодействие между микробными сообществами и свойствами почвы. Nat. Rev. Microbiol. 2024 , 22 , 226–239. [ Google Scholar ] [ CrossRef ] [ PubMed ]
10. Шаабан, М.; Ван Цвитен, Л.; Башир, С.; Юнас, А.; Нуньес-Дельгадо, А.; Чхайро, М.А.; Кубар, К.А.; Али, У.; Рана, М.С.; Мехмуд, М.А.; и др. Краткий обзор применения биоугля на сельскохозяйственных почвах для улучшения их состояния и борьбы с загрязнением. J. Environ. Manag. 2018 , 228 , 429–440. [ Google Scholar ] [ CrossRef ] [ PubMed ]
11. Dangi, S.; Gao, SD; Duan, YH; Wang, D. Структура микробного сообщества почвы, влияющая на источники биоугля и удобрений. Appl. Soil Ecol. 2020 , 150 , 103452. [ Google Scholar ] [ CrossRef ]
12. Xia, H.; Riaz, M.; Babar, S.; Yan, L.; Li, YX; Wang, XL; Wang, JY; Jiang, CC Оценка воздействия биоугля на микробы в кислых почвах: снижение токсичности алюминия и кислотности. J. Environ. Manag. 2023 , 345 , 118796. [ Google Scholar ] [ CrossRef ] [ PubMed ]
13. Qian, R.; Guo, R.; Yang, QX; Naseer, MA; Sun, BP; Wang, LL; Zhang, J.; Ren, XL; Chen, XL; Jia, ZK. Может ли переработка соломы обеспечить устойчивое сельское хозяйство на уровне мелких фермерских хозяйств? Пример полузасушливого региона. J. Clean Prod. 2024 , 439 , 140859. [ Google Scholar ] [ CrossRef ]
14. Lehmann, J.; Cowie, A.; Masiello, CA; Kammann, C.; Woolf, D.; Amonette, JE; Cayuela, ML; Camps-Arbestain, M.; Whitman, T. Биоуголь в смягчении последствий изменения климата. Nat. Geosci. 2021 , 14 , 883–892. [ Google Scholar ] [ CrossRef ]
15. Муртаза, Г.; Ахмед, З.; Элдин, С.М.; Али, И.; Усман, М.; Икбал, Р.; Ризван, М.; Абдель-Хамид, Великобритания; Хайдер, А.А.; Тарик, А. Биоуголь как зеленый сорбент для очистки загрязненных почв и связанных с ним рисков токсичности: критический обзор. Separations 2023 , 10 , 197. [ Google Scholar ] [ CrossRef ]
16. Бандара, Т.; Фрэнкс, А.; Сюй, Дж. М.; Болан, Н.; Ван, Х. Л.; Тан, С. Х. Механизмы химической и биологической иммобилизации потенциально токсичных элементов в почвах, обогащенных биоуглем. Crit. Rev. Environ. Sci. Technol. 2020 , 50 , 903–978. [ Google Scholar ] [ CrossRef ]
17. Ли, Сяньсянь; Сун, И.; Яо, С.; Бянь, И.Р.; Гу, Ч.Г.; Ян, Х.Л.; Ван, Ф.; Цзян, Х. Могут ли биоуголь и щавелевая кислота снизить токсический стресс, вызванный полициклическими ароматическими углеводородами в микробных сообществах почвы? Sci. Total Environ. 2019 , 695 , 133879. [ Google Scholar ] [ CrossRef ] [ PubMed ]
18. Бао, Х.Й.; Ван, Дж.Ф.; Чжан, Х.; Ли, Дж.; Ли, Х.; У, Ф.Й. Влияние биоугля и органических субстратов на биодеградацию полициклических ароматических углеводородов и структуру микробного сообщества в почвах, загрязненных ПАУ. J. Hazard. Mater. 2020 , 385 , 121595. [ Google Scholar ] [ CrossRef ]
19. Baldoni, N.; Francioni, M.; Trozzo, L.; Toderi, M.; Fornasier, F.; D'Ottavio, P.; Corti, G.; Cocco, S. Влияние биоугля, полученного газификацией древесины, на физико-химические свойства почвы и активность ферментов, а также на урожайность в системе производства пшеницы на слабощелочной почве. Biomass Bioenerg. 2023 , 176 , 106914. [ Google Scholar ] [ CrossRef ]
20. Хансен, В.; Мюллер-Стовер, Д.; Манкхольм, Л. Дж.; Пелтре, К.; Хауггард-Нильсен, Х.; Йенсен, Л. С. Влияние биоугля, полученного при газификации соломы и древесины, на секвестрацию углерода, отдельные показатели плодородия почвы и функциональные группы в почве: инкубационное исследование. Geoderma 2016 , 269 , 99–107. [ Google Scholar ] [ CrossRef ]
21. Чжан, LY; Цзин, YM; Сян, YZ; Чжан, RD; Лу, HB. Реакция изменений структуры и активности микробного сообщества почвы на добавление биоугля: метаанализ. Sci. Total Environ. 2018 , 643 , 926–935. [ Google Scholar ] [ CrossRef ] [ PubMed ]
22. Zheng, H.; Wang, X.; Luo, XX; Wang, ZY; Xing, BS. Эффекты отрицательной минерализации углерода, вызванные биоуглем, в прибрежной водно-болотной почве: роль агрегации почвы и микробной модуляции. Sci. Total Environ. 2018 , 610 , 951–960. [ Google Scholar ] [ CrossRef ] [ PubMed ]
23. Палансурья, К.Н.; Вонг, Дж.Т.Ф.; Хашимото, И.; Хуан, Л.Б.; Ринклебе, Дж.; Чанг, С.Х.; Болан, Н.; Ван, Х.Л.; Ок, И.С. Реакция микробных сообществ на почвы, обработанные биоуглем: критический обзор. Biochar 2019 , 1 , 3–22. [ Google Scholar ] [ CrossRef ]
24. Сингх, Х.; Нортап, Б.К.; Райс, К.В.; Прасад, П.В. В. Применение биоугля влияет на физические и химические свойства почвы, микробное разнообразие и урожайность сельскохозяйственных культур: метаанализ. Biochar 2022 , 4 , 8. [ Google Scholar ] [ CrossRef ]
25. Guo, MX; Shang, XT; Ma, YL; Zhang, KK; Zhang, L.; Zhou, YM; Gong, ZQ; Miao, RH. Фиторемедиация сельскохозяйственных почв, загрязненных полициклическими ароматическими углеводородами, с помощью биоугля: динамические реакции функциональных генов и микробного сообщества. Environ. Pollut. 2024 , 345 , 123476. [ Google Scholar ] [ CrossRef ]
26. Конг, Л. Л.; Гао, YY; Чжоу, QX; Чжао, XY; Сан, ZW. Биоуголь ускоряет биодеградацию ПАУ в почве, загрязненной нефтью, с помощью стратегии биостимуляции. J. Hazard. Mater. 2018 , 343 , 276–284. [ Google Scholar ] [ CrossRef ] [ PubMed ]
27. Чжао, XY; Мяо, RH; Го, MX; Шан, XT; Чжоу, YM; Чжу, JW. Биоуголь усиливает деградацию полициклических ароматических углеводородов в почве, засеянной райграсом: бактериальное сообщество и механизмы экспрессии генов деградации. Sci. Total Environ. 2022 , 838 , 156076. [ Google Scholar ] [ CrossRef ] [ PubMed ]
28. Бао, JQ; Ли, JB; Цзян, LF; Мэй, WP; Сун, MK; Хуан, DY; Ло, CL; Чжан, G. Новое понимание механизма, лежащего в основе влияния биоугля на деградацию фенантрена в загрязненной почве, полученное с помощью ДНК-SIP. J. Hazard. Mater. 2022 , 438 , 129466. [ Google Scholar ] [ CrossRef ] [ PubMed ]
29. Чжан, М.; Ло, И.К.; Чжу, И.Т.; Чжан, Х.Й.; Ван, Х.Л.; Ли, В.; Ли, П.П.; Хан, Дж.Г. Взгляд на механизмы, лежащие в основе биодеградации фенантрена в почве, обогащенной биоуглем: от биодоступности до микробных сообществ почвы. Biochar 2023 , 5 , 14. [ Google Scholar ] [ CrossRef ]
30. Чжан, YN; Ян, XL; Бянь, YR; Гу, CG; Ван, DZ; Цзян, X. Метод ускоренной экстракции растворителем, твердофазной экстракции и высокоэффективной жидкостной хроматографии для определения полициклических ароматических углеводородов в образцах почвы и дождевых червей. Chin. J. Anal. Chem. 2016 , 44 , 1514–1520. [ Google Scholar ] [ CrossRef ]
31. Сетиа, Р.; Верма, С.Л.; Маршнер, П. Измерение углерода микробной биомассы методом прямой экстракции – сравнение с экстракцией методом фумигации хлороформом. Eur. J. Soil Biol. 2012 , 53 , 103–106. [ Google Scholar ] [ CrossRef ]
32. Орен, А.; Ротбарт, Н.; Борисовер, М.; Бар-Тал, А. Экстракция фумигации хлороформом для измерения микробной биомассы почвы: обоснованность использования образцов, приближающихся к водонасыщению. Geoderma 2018 , 319 , 204–207. [ Google Scholar ] [ CrossRef ]
33. Li, X.; Zheng, R.; Bu, QH; Cai, QH; Liu, YF; Lu, Q.; Cui, JZ. Сравнение содержания ПАУ, потенциального риска в растительности и голой почве вблизи нефтяной скважины Дацин и оценка влияния свойств почвы на ПАУ. Environ. Sci. Pollut. R. 2019 , 26 , 25071–25083. [ Google Scholar ] [ CrossRef ] [ PubMed ]
34. Lu, Q.; Jiang, ZW; Feng, WX; Yu, CJ; Jiang, FZ; Huang, JY; Cui, JZ Исследование деградации полициклических ароматических углеводородов, индуцированной бактериальным сообществом, и образования гумуса при совместном компостировании отходов коровьего навоза с загрязненной почвой. J. Environ. Manag. 2023 , 326 , 116852. [ Google Scholar ] [ CrossRef ] [ PubMed ]
35. Чжань, XH; У, WZ; Чжоу, LX; Лян, JR; Цзян, TH. Взаимодействие растворенного органического вещества и фенантрена на ферментативную активность почвы. J. Environ. Sci. 2010 , 22 , 607–614. [ Google Scholar ] [ CrossRef ] [ PubMed ]
36. Ling, WT; Lu, XD; Gao, YZ; Liu, J.; Sun, YD. Активность полифенолоксидазы в субклеточных фракциях овсяницы тростниковой, загрязненной полициклическими ароматическими углеводородами. J. Environ. Qual. 2012 , 41 , 807–813. [ Google Scholar ] [ CrossRef ] [ PubMed ]
37. Сан, Д.Л.; Цзян, Х.; У, К.Л.; Чжоу, Нью-Йорк. Внутригеномная гетерогенность генов 16S рРНК приводит к переоценке прокариотического разнообразия. Appl. Environ. Microb. 2013 , 79 , 5962–5969. [ Google Scholar ] [ CrossRef ]
38. Walters, W.; Hyde, ER; Berg-Lyons, D.; Ackermann, G.; Humphrey, G.; Parada, A.; Gilbert, JA; Jansson, JK; Caporaso, JG; Fuhrman, JA; и др. Улучшенные праймеры для бактериального гена 16S рРНК (V4 и V4-5) и гена маркера внутреннего транскрибируемого спейсера грибов для исследований микробных сообществ. mSystems 2015 , 1 , e00009. [ Google Scholar ] [ CrossRef ] [ PubMed ]
39. Чжоу, X.; Лю, XY; Лю, MY; Лю, WX; Сю, JZ; Ли, YW. Сравнительная оценка пар праймеров 16S рРНК для идентификации нитрифицирующих гильдий в почвах при длительном внесении органических удобрений и управлении водными ресурсами. Front. Microbiol. 2024 , 15 , 1424795. [ Google Scholar ] [ CrossRef ]
40. Эдгар, Р. К. UPARSE: Высокоточные последовательности OTU из прочтений микробных ампликонов. Nat. Methods 2013 , 10 , 996–998. [ Google Scholar ] [ CrossRef ]
41. Ван, К.; Гаррити, Г.М.; Тидже, Дж.М.; Коул, Дж.Р. Наивный байесовский классификатор для быстрого определения последовательностей рРНК в новой бактериальной таксономии. Appl. Environ. Microb. 2007 , 73 , 5261–5267. [ Google Scholar ] [ CrossRef ] [ PubMed ]
42. Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, FO. Проект базы данных генов рибосомальной РНК SILVA: Улучшенная обработка данных и веб-инструменты. Nucleic Acids Res. 2012 , 41 , D590–D596. [ Google Scholar ] [ CrossRef ] [ PubMed ]
43. Сонг, И.; Бянь, И.Р.; Ван, Ф.; Сюй, М.; Ни, Н.; Ян, XL; Гу, Ч.Г.; Цзян, Х. Динамическое воздействие биоугля на структуру бактериального сообщества в почве, загрязненной полициклическими ароматическими углеводородами. J. Agric. Food Chem. 2017 , 65 , 6789–6796. [ Google Scholar ] [ CrossRef ] [ PubMed ]
44. Чжан, YY; Ван, T.; Янь, C.; Ли, YZ; Мо, F.; Хан, J. Стратегии жизненного цикла микроорганизмов и дисперсный органический углерод опосредуют образование углерода некромассы микроорганизмов и стабилизацию в ответ на добавление биоугля. Sci. Total Environ. 2024 , 950 , 175041. [ Google Scholar ] [ CrossRef ]
45. Гао, Л.; Ван, Р.; Шен, Г.М.; Чжан, Дж.Х.; Мэн, Г.Х.; Чжан, Дж.Г. Влияние биоугля на питательные вещества и структуру микробного сообщества почв, используемых для выращивания табака. J. Soil Sci. Plant Nutr. 2017 , 17 , 884–896. [ Google Scholar ] [ CrossRef ]
46. Рутильяно, ФА; Романо, М.; Марзайоли, Р.; Багливо, И.; Баронти, С.; Миглиетта, Ф.; Кастальди, С. Влияние добавления биоугля на микробное сообщество почвы в посевах пшеницы. Евро. Ж. Почвенная биол. 2014 , 60 , 9–15. [ Академия Google ] [ CrossRef ]
47. Luo, SS; Wang, SJ; Tian, L.; Li, SQ; Li, XJ; Shen, YF; Tian, CJ. Длительное применение биоугля влияет на микробное сообщество почвы и его потенциальную роль в полузасушливых сельскохозяйственных угодьях. Appl. Soil Ecol. 2017 , 117 , 10–15. [ Google Scholar ] [ CrossRef ]
48. Ляо, Х.К.; Чжэн, К.Л.; Лонг, Дж.; Гусман, И. Влияние биоугля на бактериальные сообщества ризосферы томатов и использование ими углерода, полученного из растений, в известковой почве. Geoderma 2021 , 396 , 115082. [ Google Scholar ] [ CrossRef ]
49. El-Naggar, A.; El-Naggar, AH; Shaheen, SM; Sarkar, B.; Chang, SX; Tsang, DCW; Rinklebe, J.; Ok, YS. Влияние состава биоугля на высвобождение питательных веществ в почве, минерализацию углерода и потенциальный экологический риск: обзор. J. Environ. Manag. 2019 , 241 , 458–467. [ Google Scholar ] [ CrossRef ]
50. Митчелл, П.Дж.; Симпсон, А.Дж.; Сунг, Р.; Шурман, Дж.С.; Томас, С.К.; Симпсон, М.Дж. Внесение биоугля и фосфорного удобрения изменило микробное сообщество лесной почвы и молекулярный состав органического вещества местной почвы. Биогеохимия 2016 , 130 , 227–245. [ Google Scholar ] [ CrossRef ]
51. Резерфорд, Д.В.; Вершоу, Р.Л.; Ростад, К.Э.; Келли, К.Н. Влияние условий формирования на биоуголь: композиционные и структурные свойства целлюлозы, лигнина и соснового биоугля. Biomass Bioenerg. 2012 , 46 , 693–701. [ Google Scholar ] [ CrossRef ]
52. Supanchaiyamat, N.; Jetsrisuparb, K.; Knijnenburg, JTN; Tsang, DCW; Hunt, AJ. Лигниновые материалы для адсорбции: современные тенденции, перспективы и возможности. Bioresour. Technol. 2019 , 272 , 570–581. [ Google Scholar ] [ CrossRef ] [ PubMed ]
53. Han, L.; Nie, X.; Wei, J.; Gu, MY; Wu, WP; Chen, MF. Влияние состава биополимерного сырья на физико-химические характеристики растворенного черного углерода из биоугля на основе лигноцеллюлозы. Sci. Total Environ. 2021 , 751 , 141491. [ Google Scholar ] [ CrossRef ] [ PubMed ]
54. Куприянчик, Д.; Хейл, С.; Циммерман, А. Р.; Харви, О.; Резерфорд, Д.; Абивен, С.; Кникер, Х.; Шмидт, Х. П.; Румпель, К.; Корнелиссен, Г. Сорбция гидрофобных органических соединений на разнообразном наборе углеродистых материалов с акцентом на биоуголь. Chemosphere 2016 , 144 , 879–887. [ Google Scholar ] [ CrossRef ] [ PubMed ]
55. Зама, Э.Ф.; Рид, Б.Дж.; Арп, Х.П.Х.; Сан, Г.Х.; Юань, Х.Й.; Чжу, Ю.Г. Достижения в исследованиях по использованию биоугля в почве для рекультивации: обзор. J. Soil Sediment 2018 , 18 , 2433–2450. [ Google Scholar ] [ CrossRef ]
56. Siedt, M.; Schäffer, A.; Smith, KEC; Nabel, M.; Roß-Nickoll, M.; van Dongen, JT. Сравнение соломы, компоста и биоугля с точки зрения их пригодности в качестве сельскохозяйственных почвенных добавок для влияния на структуру почвы, вымывание питательных веществ, микробные сообщества и судьбу пестицидов. Sci. Total Environ. 2021 , 751 , 141607. [ Google Scholar ] [ CrossRef ]
57. Хуан, XW; Ян, XL; Линь, JH; Фрэнкс, AE; Чэн, J.; Чжу, YJ; Ши, JC; Сю, JM; Юань, M.; Фу, XJ; и др. Биоуголь снизил токсичность атразина для соевых бобов, что было выявлено в микробном сообществе почвы и процессе сборки. Sci. Total Environ. 2022 , 834 , 155261. [ Google Scholar ] [ CrossRef ] [ PubMed ]
58. Ван, YS; Деверё, R.; Джордж, SE; Чен, JJ; Гао, B.; Ноерпель, M.; Шекель, K. Взаимодействие эффектов добавления биоугля и токсичности свинца на микробное сообщество почвы. J. Hazard. Mater. 2022 , 425 , 127921. [ Google Scholar ] [ CrossRef ] [ PubMed ]
59. Ян, X.; Ю, M.; Лю, SY; Саркар, B.; Лю, ZS; Ян, XL. Микробные реакции на внесение биоугля в почву, загрязненную потенциально токсичными элементами (ПТЭ): критический обзор эффектов и потенциальных механизмов. Biochar 2023 , 5 , 57. [ Google Scholar ] [ CrossRef ]
60. Zhen, MN; Chen, HK; Liu, QL; Song, BR; Wang, YZ; Tang, JC. Сочетание рамнолипида и биоугля для содействия фиторемедиации почвы, загрязненной нефтяными углеводородами, с использованием Spartina anglica . J. Environ. Sci. 2019 , 85 , 107–118. [ Google Scholar ] [ CrossRef ]
61. Лю, Дж.; Сян, И. Б.; Чжан, З. М.; Лин, В. Т.; Гао, И. З. Инокуляция эндофитной бактерии, разрушающей фенантрен, снижает уровень фенантрена и изменяет структуру бактериального сообщества в пшенице. Appl. Microbiol. Biot. 2017 , 101 , 5199–5212. [ Google Scholar ] [ CrossRef ] [ PubMed ]
62. Ци, Y.; У, YX; Чжи, QY; Чжан, Z.; Чжао, YL; Фу, G. Влияние полициклических ароматических углеводородов на состав бактериальных сообществ почвы в приливно-отливных водно-болотных угодьях дельты реки Хуанхэ в Китае. Микроорганизмы 2024 , 12 , 141. [ Google Scholar ] [ CrossRef ]
63. Чжан, MY; Ван, J.; Бай, SH; Чжан, YL; Тэн, Y.; Сюй, ZH. Фиторемедиация совместно загрязненной почвы с использованием биоугля: удаление загрязняющих веществ и свойства бактериального сообщества. Geoderma 2019 , 348 , 115–123. [ Google Scholar ] [ CrossRef ]
64. Чжу, Й.Ф.; Ге, Х.П.; Ван, Л.П.; Ю, Й.Н.; Чэн, Й.Дж.; Ма, Дж.; Чэнь, Ф. Биоуголь восстанавливает сложность сети редких и распространенных микробных таксонов в рекультивированной почве горнодобывающих районов для совместного предотвращения кадмиевого стресса. Front. Microbiol. 2022 , 13 , 972300. [ Google Scholar ] [ CrossRef ]
65. Капараджу, П.; Фелби, К. Характеристика лигнина в процессе окислительной и гидротермической предварительной обработки пшеничной соломы и кукурузной соломы. Bioresour. Technol. 2010 , 101 , 3175–3181. [ Google Scholar ] [ CrossRef ] [ PubMed ]
66. Хансен, В.; Мюллер-Стовер, Д.; Импарато, В.; Крог, П. Х.; Йенсен, Л. С.; Долмер, А.; Хауггард-Нильсен, Х. Влияние применения соломы или газифицированного биоугля на качество почвы и урожайность сельскохозяйственных культур: исследование на примере фермерского хозяйства. J. Environ. Manag. 2017 , 186 , 88–95. [ Google Scholar ] [ CrossRef ] [ PubMed ]
67. Гуль, С.; Уэйлен, Дж. К.; Томас, Б. В.; Сачдева, В.; Дэн, Х. Физико-химические свойства и микробные реакции в почвах, обработанных биоуглем: механизмы и будущие направления. Agric. Ecosyst. Environ. 2015 , 206 , 46–59. [ Google Scholar ] [ CrossRef ]
68. Tian, J.; Wang, JY; Dippold, M.; Gao, Y.; Blagodatskaya, E.; Kuzyakov, Y. Биоуголь влияет на круговорот органического вещества почвы и микробные функции, но не изменяет структуру микробного сообщества в рисовой почве. Sci. Total Environ. 2016 , 556 , 89–97. [ Google Scholar ] [ CrossRef ] [ PubMed ]
69. Чжоу, ZY; Ян, L.; Ван, MX; Чжоу, ZF. Добавка биоугля способствовала росту кукурузы и изменила состав бактериального сообщества в почве, загрязненной фенантреном. J. Soil Sci. Plant Nutr. 2023 , 23 , 3010–3022. [ Google Scholar ] [ CrossRef ]
70. Xu, HJ; Wang, XH; Li, H.; Yao, HY; Su, JQ; Zhu, YG. Влияние биоугля на состав микробного сообщества почвы и круговорот азота в кислой почве, засеянной рапсом. Environ. Sci. Technol. 2014 , 48 , 9391–9399. [ Google Scholar ] [ CrossRef ] [ PubMed ]
71. Чжан, GX; Хэ, LX; Го, XF; Хан, ZW; Цзи, L.; Хэ, QS; Хан, LF; Сан, K. Механизм действия биоугля как стратегии биостимуляции для удаления полициклических ароматических углеводородов из сильно загрязненной почвы на коксовом заводе. Geoderma 2020 , 375 , 114497. [ Google Scholar ] [ CrossRef ]
72. Фаррелл, М.; Кун, ТК; Макдональд, Л.М.; Мэддерн, Т.М.; Мерфи, Д.В.; Холл, П.А.; Сингх, Б.П.; Бауманн, К.; Крулл, Э.С.; Балдок, Дж.А. Микробное использование углерода, полученного из биоугля. Sci. Total Environ. 2013 , 465 , 288–297. [ Google Scholar ] [ CrossRef ] [ PubMed ]
73. Ли, XN; Сун, Y.; Бянь, YR; Гу, CG; Ян, XL; Ван, F.; Цзян, X. Взгляд на механизмы, лежащие в основе эффективной ризодеградации ПАУ в почве, обогащенной биоуглем: от микробных сообществ к метаболомике почвы. Environ. Int. 2020 , 144 , 105995. [ Google Scholar ] [ CrossRef ]
74. Лу, Л.; Чжан, Дж.; Пэн, Ч. Изменение характера рассеивания полициклических ароматических углеводородов (ПАУ) в почве и состава микробного сообщества вследствие добавления рамнолипидов. Вода, воздух, почва, загрязнение. 2019 , 230 , 107. [ Google Scholar ] [ CrossRef ]
75. Liu, ZS; Wang, KH; Cai, M.; Yang, ML; Wang, XK; Ma, HL; Yuan, YH; Wu, LH; Li, DF; Liu, SJ Agromyces chromiiresistens sp. nov., Novosphingobium album sp. nov., Sphingobium arseniciresistens sp. nov., Sphingomonas pollutisoli sp. nov. и Salinibacterium metalliresistens sp. nov.: пять новых представителей Microbacteriaceae и Sphingomonadaceae из загрязненной почвы. Front. Microbiol. 2023 , 14 , 1289-110. [ Google Scholar ]
76. Хан, SM; Ван, YX; Ли, Y.; Ши, KY. Исследование бактериального разнообразия в жильной почве, засаженной Cajanus cajan , методом высокопроизводительного секвенирования. Bioengineered 2021 , 12 , 6981–6995. [ Google Scholar ] [ CrossRef ] [ PubMed ]
77. Омони, В.Т.; Байду, П.К.; Фагбохунгбе, Миссури; Семпл, К.Т. Влияние обогащенного и необработанного биоугля на катаболизм 14 C-фенантрена в почве. Environ. Technol. Innov. 2020 , 20 , 101146. [ Google Scholar ] [ CrossRef ]
78. Сюй, Р.; Оббард, Дж. П. Биодеградация полициклических ароматических углеводородов в загрязненных нефтью пляжных отложениях, обработанных питательными добавками. J. Environ. Qual. 2004 , 33 , 861–867. [ Google Scholar ] [ CrossRef ]
79. Ли, Сянь Н.; Сун, И.; Ван, Ф.; Бянь, И. Р.; Цзян, Сянь. Комбинированное воздействие биоугля из кукурузной соломы и щавелевой кислоты на рассеивание полициклических ароматических углеводородов и структуру микробного сообщества в почве: механистическое исследование. J. Hazard. Mater. 2019 , 364 , 325–331. [ Google Scholar ] [ CrossRef ]
80. Zheng, XM; Xu, WH; Dong, J.; Yang, T.; Shangguan, ZC; Qu, J.; Li, X.; Tan, XF. Эффекты биоугля и его применение в микробной очистке загрязненной почвы: обзор. J. Hazard. Mater. 2022 , 438 , 129557. [ Google Scholar ] [ CrossRef ]
81. Чэнь, СК; Пэн, ДЖ; Дуань, ГЛ. Обогащение функциональных микробов и генов при деградации пирена в двух разных почвах. J. Soil Sediment 2016 , 16 , 417–426. [ Google Scholar ] [ CrossRef ]
82. Ляо, QH; Лю, H.; Лу, C.; Лю, J.; Вайги, MG; Лин, WT. Корневые экссудаты усиливают деградацию ПАУ и обилие генов, вызывающих деградацию, в почвах. Sci. Total Environ. 2021 , 764 , 144436. [ Google Scholar ] [ CrossRef ] [ PubMed ]
83. Wu, C.; Li, F.; Yi, SW; Ge, F. Генно-инженерная микробная ремедиация почв, загрязненных тяжелыми металлами и полициклическими ароматическими углеводородами: достижения и оценка экологического риска. J. Environ. Manag. 2021 , 296 , 113-185. [ Google Scholar ] [ CrossRef ]
84. Geng, SY; Xu, GM; Cao, W.; You, Y.; Zhu, Y.; Ding, AZ; Fan, FQ; Dou, JF. Распространение полициклических ароматических соединений и потенциально токсичных элементов, а также соответствующее формирование междоменного микробного сообщества в почве заброшенной заправочной станции. Environ. Res. 2022 , 212 Pt A , 113618. [ Google Scholar ] [ CrossRef ]
85. Tarigholizadeh, S.; Sushkova, S.; Rajput, VD; Ranjan, A.; Arora, J.; Dudnikova, T.; Barbashev, A.; Mandjieva, S.; Minkina, T.; Wong, MH Перенос и деградация ПАУ в системе почва-растение: обзор. J. Agric. Food Chem. 2024 , 72 , 46–64. [ Google Scholar ] [ CrossRef ]
86. Xia, XH; Xia, N.; Lai, YJ; Dong, JW; Zhao, PJ; Zhu, BT; Li, ZH; Ye, W.; Yuan, Y.; Huang, JX. Реакция генов, разрушающих ПАУ, на биодоступность ПАУ в вышележащих водах, взвешенных отложениях и отложенных отложениях реки Янцзы. Chemosphere 2015 , 128 , 236–244. [ Google Scholar ] [ CrossRef ] [ PubMed ]
87. Ахмад, М.; Ван, П.Д.; Ли, Дж.Л.; Ван, Р.Ф.; Дуань, Л.; Ло, Х.К.; Ирфан, М.; Пэн, З.К.; Инь, Л.З.; Ли, В.Дж. Влияние биостимуляторов на деградацию пирена, состав и функции прокариотического сообщества. Окружающая среда. Загрязнение. 2021 , 289 , 117863. [ Google Scholar ] [ CrossRef ]
88. Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: сверхбыстрый многофункциональный препроцессор FASTQ. Bioinformatics 2018 , 34 , i884–i890. [ Google Scholar ] [ CrossRef ] [ PubMed ]
89. Магоч, Т.; Зальцберг, С.Л. FLASH: Быстрая корректировка длины коротких прочтений для улучшения сборки генома. Bioinformatics 2011 , 27 , 2957–2963. [ Google Scholar ] [ CrossRef ]

Wang Z, Li J, Kang Y, Ran J, Song J, Jiang M, Li W, Zhang M. Effects of Wheat Straw-Derived Biochar on Soil Microbial Communities Under Phenanthrene Stress. Agriculture. 2025; 15(1):77.
Перевод статьи «Effects of Wheat Straw-Derived Biochar on Soil Microbial Communities Under Phenanthrene Stress» авторов Wang Z, Li J, Kang Y, Ran J, Song J, Jiang M, Li W, Zhang M., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык

















































































































Комментарии (0)