Оценка физиологических признаков генотипов Fragaria vesca в условиях дефицита воды
Засуха является одной из ключевых проблем изменения климата. Основная глобальная проблема, связанная с растущим дефицитом воды, заключается в том, что подавляющее большинство сельскохозяйственных культур – это виды и сорта, выведенные в результате селекционной работы, не учитывающей столь быстрого снижения доступности воды в почве. Основной целью проведенного исследования было сравнение физиолого-биохимической реакции на водный дефицит растений вида Fragaria vesca – двух культурных сортов и одного, собранного в естественной среде.
Аннотация
Двухлетний вегетационный эксперимент проводился в полиэтиленовом туннеле. Уровень влажности субстрата контролировался с помощью показаний тензиометра. Измерения параметров газообмена, флуоресценции хлорофилла «а», содержания фотосинтетических пигментов в листьях, индекса относительной оводненности листьев, общего урожая плодов, массы отдельных плодов и содержания K, Ca, Mg, Na, Cu, Zn, Mn, Mo, а также соотношения одновалентных и двухвалентных катионов в листьях, корнях и кроне растений проводились три раза в год в течение экспериментов. На основании однофакторного и двухфакторного дисперсионного анализа были обнаружены статистически значимые различия между дикорастущими растениями и культурными сортами в контрольных условиях, особенно по значениям флуоресценции хлорофилла и содержанию фотосинтетических пигментов. Для большинства измеряемых параметров в большинстве временных точек оценки был выявлен значительный основной эффект уровня влажности почвы. Однако, значимый эффект взаимодействия между уровнем влажности почвы и генотипом наблюдался реже. Значимые изменения в ответ на дефицит воды варьировались в зависимости от параметра и генотипа, от 2,5% до 106,1%. Для содержания химических элементов изменения достигали 157,1%. Результаты свидетельствуют о том, что растения, полученные из природных сред, демонстрируют лучшую адаптацию к условиям дефицита воды, что делает их пригодными для использования в селекционных программах, направленных на создание сортов, устойчивых к дефициту почвенной влаги. Однако следует учитывать ограничения исследования, в частности, отсутствие молекулярного анализа адаптивных механизмов растений.
1. Введение
Явления засухи и опустынивания, усугубленные изменением климата, отрицательно влияют на глобальную продовольственную безопасность и здоровье человека и могут даже представлять угрозу для мира во всем мире [ 1 , 2 , 3 ]. Климатические модели прогнозируют, что интенсивность засухи может удвоиться на 30% мировой площади суши при строгих сценариях смягчения последствий, что приведет к существенным последствиям для дефицита водоснабжения и спроса [ 4 ]. Стресс, вызванный дефицитом воды, представляет собой серьезную проблему для сельского хозяйства, делая устойчивость растений к нехватке воды критически важной для экономики [ 5 ]. В климате Польши периодические метеорологические засухи [ 6 ] нарушают водный баланс пострадавших районов, что приводит к засухам почвы, которые ухудшают продуктивность сельскохозяйственных культур [ 7 ]. В умеренном климате засуха является одним из основных факторов, ограничивающих урожайность сельскохозяйственных культур [ 4 , 8 ]. Основная глобальная проблема, связанная с растущим дефицитом воды, заключается в том, что большинство возделываемых культур, включая растения рода Fragaria , состоят из видов и сортов, выведенных путем селективного разведения, часто без должного учета текущего значительного сокращения доступности почвенной влаги [ 9 ].
Экономически наиболее значимым видом рода Fragaria является культурная клубника ( Fragaria × ananassa Duch.), плоды которой являются одними из наиболее широко потребляемых в мире [ 10 , 11 ]. В Польше растения рода Fragaria также имеют значительное экономическое значение [ 8 , 12 ]. В 2020 году доля растений Fragaria в общем урожае плодов с плодовых кустарников и ягодных плантаций достигла 26,0% [ 13 ]. Лесная земляника ( Fragaria vesca L.) в основном выращивается в любительских масштабах. Хотя F. vesca не выращивается в коммерческих целях, ее ценные признаки как исследовательской модели были широко освещены, особенно из-за ее короткого периода генерации, четко определенных циклов сезонного цветения и вегетативного роста и признаков одного гена, контролирующих такие аспекты, как цветение и окраска плодов [ 14 ].
Растения рода Fragaria очень чувствительны к дефициту воды из-за большой площади листовой поверхности, высокого содержания воды в плодах и поверхностной корневой системы. Эта чувствительность была подчеркнута в многочисленных исследованиях, в которых сообщается об ухудшении физиологических параметров у растений, подверженных условиям дефицита воды [ 3 , 15 , 16 , 17 , 18 , 19 , 20 , 21 ]. Однако эта особенность нежелательна при коммерческом выращивании, поскольку даже незначительные отклонения от оптимальных уровней полива могут привести к снижению урожайности, ухудшению качества плодов, снижению рентабельности культур и неудовлетворению потребительского и рыночного спроса. Следовательно, ключевой задачей для селекционеров является создание сортов с повышенной устойчивостью к дефициту воды в почве.
Важным предварительным шагом в селекционных программах, направленных на повышение устойчивости растений к дефициту воды за счет повышения эффективности водопользования, является оценка степени адаптации различных генотипов (сортов) к ограниченной доступности воды. Например, разработка засухоустойчивых сортов пшеницы и хлопка показала многообещающие результаты в поддержании продуктивности сельского хозяйства, несмотря на экологический стресс [ 22 , 23 ]. Различия в физиологических, морфологических и морфогенетических признаках на уровне сорта были задокументированы в многочисленных исследованиях [ 3 , 11 , 24 , 25 , 26 , 27 ]. Дикие сорта растений обладают генетическими ресурсами, которые повышают их устойчивость к абиотическим стрессам, как показано в исследованиях таких видов, как картофель [ 28 ] и ячмень [ 29 ]. Однако отсутствуют исследования, оценивающие различия в физиологических признаках и реакциях на дефицит почвенной влаги между культурными и дикими видами Fragaria .
Таким образом, целью данного исследования было сравнение физиологических реакций растений Fragaria vesca – двух сортов ('Rugia' и 'Baron von Solemacher') и образца лесной земляники, полученного из естественной среды обитания, для выявления генотипов с большей устойчивостью. Это позволит определить, проявляют ли дикорастущие растения меньшую чувствительность к засухе, и, следовательно, оправдано ли их использование в селекционных программах, направленных на создание сортов, устойчивых к дефициту воды. Эта цель была достигнута путем проверки следующих гипотез: (1) существуют физиологические различия между культурными и дикорастущими сортами; (2) дефицит воды отрицательно влияет на физиологические показатели растений Fragaria vesca ; (3) дикорастущие растения проявляют меньшую чувствительность к дефициту воды в субстрате по сравнению с культурными сортами.
2. Материалы и методы
2.1. Материалы
Растения, использованные в этом исследовании, включали Fragaria vesca сортов 'Rugia' и 'Baron von Solemacher' (саженцы получены от TOP-PLANT, Щецин, Польша), а также рассаду лесной земляники, собранную в естественной среде обитания в лесной местности недалеко от деревни Цемник, недалеко от Иньско, Польша (53°23′ с. ш., 15°34′ в. д.; 105 м над уровнем моря). Культурные сорта, использованные в исследовании, популярны в Польше. Оба сорта высокоурожайные и плодоносят с июня по сентябрь. Однако 'Baron von Solemacher' демонстрирует более низкую морозостойкость и большую восприимчивость к болезням по сравнению с 'Rugia' [ 30 ]. Эти характеристики сделали их подходящими кандидатами для экспериментального анализа.
2.2 Методы
Двухлетний эксперимент в горшках был проведен в период 2018–2019 гг. с использованием полностью рандомизированного дизайна с 15 повторениями (одно растение на повторение). Исследование было проведено в вегетационном зале факультета управления окружающей средой и сельского хозяйства Западно-Поморского технического университета в Щецине, Польша (53°25′ с. ш., 14°32′ в. д.; 25 м над уровнем моря). Использовались пластиковые контейнеры вместимостью 10 дм 3 , заполненные субстратом, состоящим из смеси 1:1 (по объему) песчано-илистой глинистой почвы, отобранной из пахотного гумусового горизонта (0–30 см) на Сельскохозяйственной опытной станции Западно-Поморского технического университета в Щецине, расположенной в Липнике (Польша), и песка. Физико-химические свойства почвы, использованной в эксперименте, представлены в таблице 1 . Для адаптации к новым условиям растения были высажены за год до начала измерений (в июне) по 3 растения в горшок. Сеянцы имели по три полностью развитых листа.
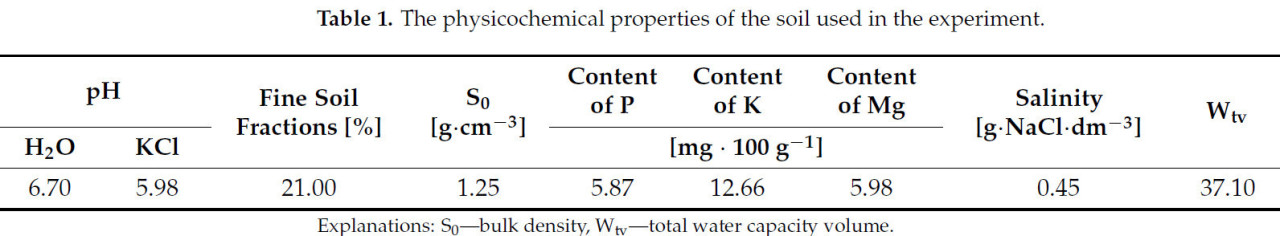
Таблица 1. Физико-химические свойства почвы, использованной в эксперименте.
Водный потенциал поддерживался на уровне от -10 кПа до -15 кПа в контрольных условиях (Ctrl) и от -30 кПа до -35 кПа в условиях дефицита воды (Def) в субстрате. Каждый горшок поливали с помощью индивидуальных капельниц, когда водный потенциал в субстрате повышался выше -15 кПа в контрольных условиях или выше -35 кПа в условиях дефицита воды. Полив проводили до значения -10 кПа в контрольном варианте и до -30 кПа в условиях дефицита воды. Принятый диапазон водного потенциала от -30 до -35 кПа рассматривался как стрессовые условия для изучаемых видов, основываясь на выводах разных авторов [ 3 ]. Необходимость полива определяли на основании показаний контактных почвенных тензиометров, расположенных на глубине 20 см в горшках каждого экспериментального варианта. Тензиометрические измерения успешно применялись в других исследованиях дефицита воды в растениях [ 31 , 32 , 33 , 34 , 35 ].
В период вегетации (с мая по октябрь) горшки находились в закрытой части вегетационного зала. В остальные месяцы их содержали в неотапливаемой теплице, поддерживая температуру выше 0 °C.
Подкормку проводили дважды в течение вегетационного периода: перед посадкой растений, при заполнении горшков и после окончания плодоношения. Во второй год исследований подкормку проводили ранней весной, перед началом вегетации и после окончания плодоношения. Использовалось многокомпонентное удобрение, специально разработанное для клубники, которое, по данным производителя (Agrecol Sp. z oo, Мешнары 2, Польша), содержит: 15,0% N (6,5% нитратного N, 8,5% аммонийного N), 6,0% P 2 O 5 (растворим в нейтральном цитрате аммония и воде, в том числе 5,2% водорастворимого P 2 O 5 ), 11,0% водорастворимого K 2 O, 0,04% общего B, 0,1% общей Cu, 0,2% общего Fe, 0,17% общего Mn, 0,01% общего Mo и 0,035% общего Zn. Норма внесения удобрения составляла 4 г на горшок за один раз.
В течение всего вегетационного периода систематически проводились морфологические наблюдения за растениями, удалялись формирующиеся столоны. Кроме того, растения регулярно осматривались на наличие вредителей. При превышении экономического порога вредоносности проводились химические обработки следующим образом:
- Decis Mega 50 EW против тли ( Aphidoea ) в дозе 0,25 л/га.
- Моспилан 20 СП, против трипсов ( Thysanoptera ), тлей ( Aphidoidea ) и тепличной белокрылки ( Trialeurodes vaporariorum ) в концентрации 4 г на 10 л воды, применяется из расчета 3 л на 100 м2 .
2.3 Измерения
В каждый год исследования три раза в течение вегетационного периода растений, во второй декаде июля, августа и сентября, проводились следующие измерения и анализы:
Содержание фотосинтетических пигментов оценивали по методу Арнона и др. [ 36 ], модифицированному Лихтенталером и Уэллберном [ 37 ] для хлорофилла «а», «b» и общего хлорофилла, а также по методу Хагера и Майера-Бертенрата [ 38 ] для каротиноидов;
индекс относительного содержания воды в листьях (RWC) по методу Ямасаки и Дилленбурга [ 39 ];
Параметры газообмена, т. е. интенсивность ассимиляции CO 2 нетто (P n ), интенсивность транспирации (E), устьичная проводимость H 2 O (g s ) и подустьичная концентрация CO 2 (ci ) , измеряли с помощью портативного газоанализатора TPS-2 с PLC-4 (PP Systems, Эймсбери, Массачусетс, США); коэффициент использования фотосинтетической воды (WUE) определяли на основе отношения P n /E;
Параметры флуоресценции хлорофилла «а» определяли на спектрофлуориметре Handy PEA (Hansatech Ltd., Кингс-Линн, Великобритания) по стандартной методике прибора (3 светодиода × 650 нм, максимальная интенсивность актиничного света 3000 мкмоль м⁻² с⁻¹ ) . Измерение проводили в каждом варианте на 15 случайно выбранных, полностью сформировавшихся листьях фасоли (повторность – измерение на одном листе) в предварительно затемненном в течение 20 мин месте с использованием фабричных зажимов (площадь облучения составляла 4 мм). Были измерены следующие параметры: начальная (нулевая) флуоресценция, индекс потери энергии возбуждения в антеннах мощности (F 0 ), максимальная флуоресценция, после восстановления акцепторов в ФС II и после адаптации к темной комнате (F M ), переменная флуоресценция, определяемая после темновой адаптации (параметр зависит от максимальной квантовой эффективности ФС II) (F V = F M − F 0 ), максимальная потенциальная эффективность фотохимической реакции в ФС II, определяемая после темновой адаптации, после восстановления акцепторов в ФС II (F V /F M ), время роста флуоресценции хлорофилла от начала измерения до достижения максимума (T FM ), и площадь над индукционной кривой флуоресценции хлорофилла «a» между точками F 0 и F M , пропорциональная размеру пула восстановленных акцепторов электронов пластохинона в ФС II (A M (Area)).
В каждом вегетационном сезоне общий урожай свежих плодов земляники определялся по весу, выраженному в граммах с горшка, а также по весу отдельного плода. В связи с тем, что лесная земляника плодоносит практически весь вегетационный период, сбор урожая проводился последовательно по мере достижения ягодами общей зрелости, охватывая период с июня по август каждого года эксперимента.
После каждого эксперимента определяли сырую и сухую массу корневой системы. Также измеряли содержание K, Ca, Mg, Na, Cu, Zn, Mn и Mo, а также соотношение одновалентных и двухвалентных элементов в листьях, корнях и кроне растений. Содержание кальция, магния, натрия и микроэлементов в тканях растений определяли методом атомно-абсорбционной спектрометрии (ААС) с использованием спектрофотометра серии iCE 3000 (ThermoScientific, Уолтем, Массачусетс, США) во влажном минерализованном материале, приготовленном на смеси азотной (HNO 3 ) и соляной кислот (HCl) в соотношении 1:1. Содержание фосфора определяли колориметрически с использованием спектрофотометра Marcel Mini. На основании полученных результатов дополнительно рассчитывали эквивалентное соотношение одновалентных и двухвалентных элементов, т. е. K : (Ca + Mg).
2.4 Метеорологические данные
Щецин расположен в субатлантической климатической зоне, где основными факторами, влияющими на климат, являются широта, рельеф местности, удалённость от моря и наличие крупных промышленных и производственных объектов. Эти факторы обуславливают относительно высокие среднегодовые температуры воздуха и низкую годовую амплитуду температур. Кроме того, для Щецина характерны мягкие зимы и прохладное лето.
Ключевые метеорологические элементы, имеющие отношение к проведенному исследованию, а также сравнение с многолетними средними значениями представлены в таблице 2. Данные, использованные в этом исследовании, были получены с ближайшей метеорологической станции IMGW Щецин-Домбье (ул. Пржестрженна, 10).
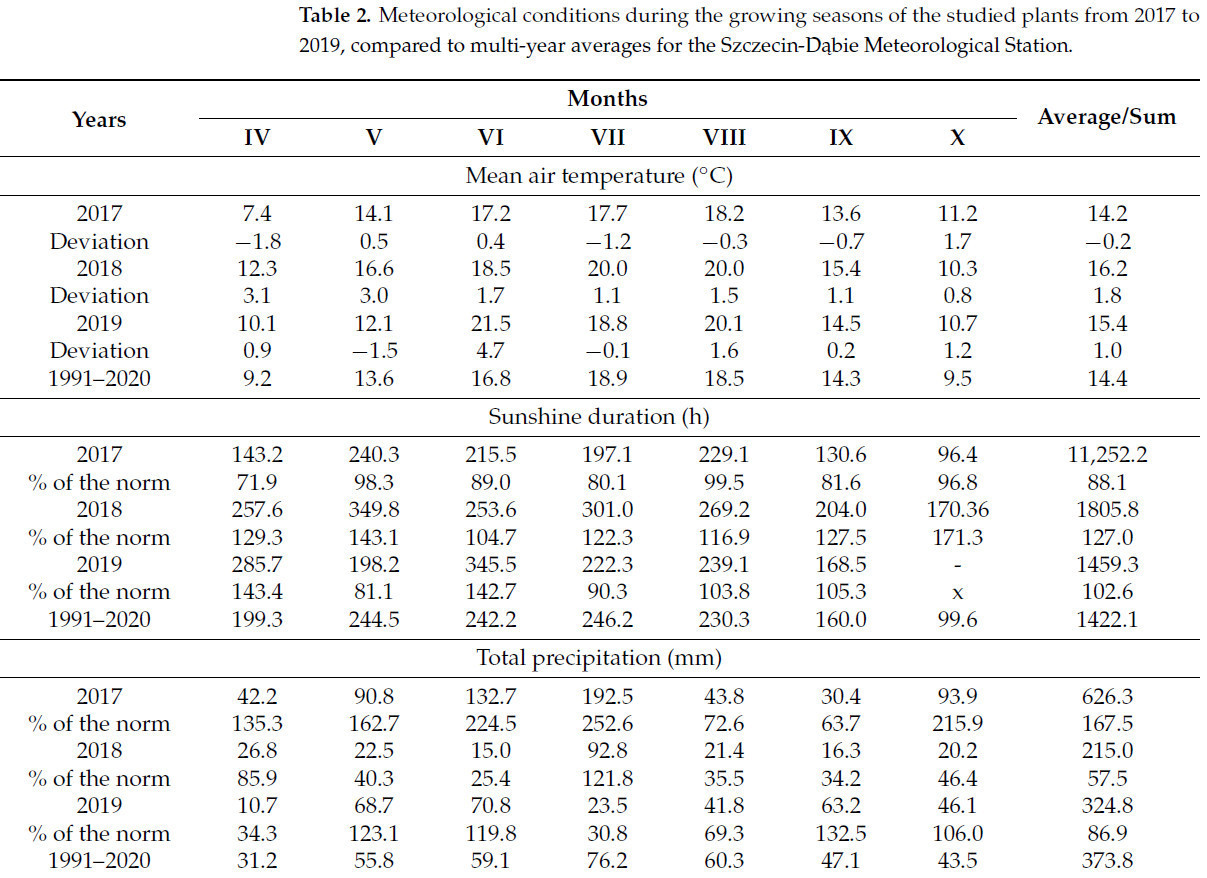
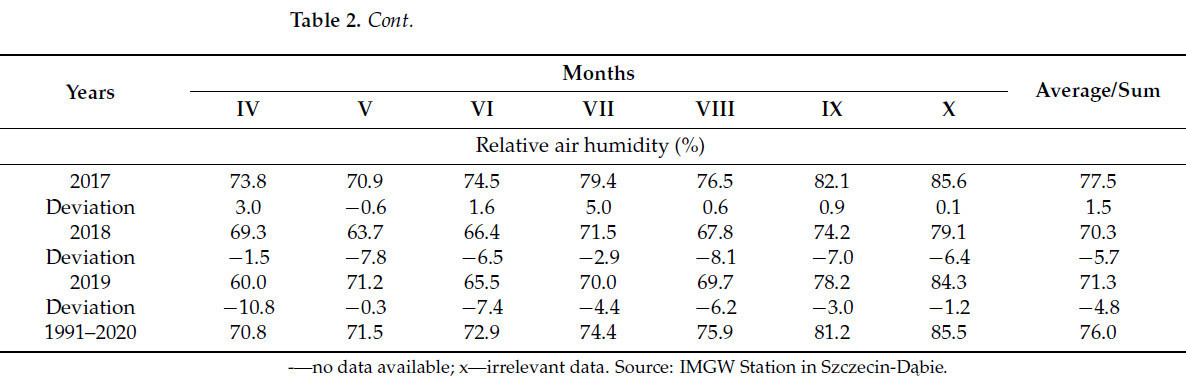
Таблица 2. Метеорологические условия в период вегетации исследуемых растений с 2017 по 2019 г. в сравнении со средними многолетними значениями для метеорологической станции Щецин-Домбе.
В первый год исследования (2017 г.) средняя температура воздуха в вегетационный период была на 0,2 °C ниже нормы. Самым холодным месяцем в исследуемый период был апрель (7,4 °C), самым тёплым – август (18,2 °C). Продолжительность солнечного сияния в вегетационный период в 2017 г. была на 21,9% ниже нормы. Ни в один месяц продолжительность солнечного сияния не превышала норму.
Во второй год исследований (2018 г.) средняя температура воздуха за вегетационный период превысила норму на 1,8 °C. Самая низкая средняя температура наблюдалась в октябре (10,3 °C), самая высокая – в июле и августе (20,0 °C). Продолжительность солнечного сияния за вегетационный период превысила норму на 27,0%, при этом все месяцы были выше нормы.
2.5 Статистический анализ
Результаты исследования были статистически проанализированы в программе STATISTICA версии 13.3 (StatSoft, Польша). Для сравнения генотипов, выращенных в контрольных условиях, использовался однофакторный дисперсионный анализ (ANOVA), а для оценки взаимодействий генотип × влажность субстрата – двухфакторный дисперсионный анализ (ANOVA) в полностью рандомизированном исследовании. Значимость различий между средними значениями определялась с помощью критерия множественных рангов Дункана при уровне значимости α = 0,05. Средним значениям, статистически не различавшимся, присваивались идентичные однобуквенные обозначения. Анализ проводился отдельно для каждого периода измерения.
В случае двухфакторного дисперсионного анализа средние значения, обозначенные одной и той же строчной буквой, не различались достоверно в контексте значимого эффекта взаимодействия. Аналогично, средние значения, обозначенные одной и той же заглавной буквой, не выявили значимых различий в отношении значимого основного эффекта (уровня влажности субстрата; SML). Основной эффект генотипа (G) не анализировался в двухфакторном анализе в связи с предположением, что реакции на водный стресс различаются у разных генотипов и могут изменяться в разных направлениях. Сравнение значений генотипов проводилось с помощью однофакторного дисперсионного анализа исключительно в контрольных условиях.
2.6. Радарные карты
На лепестковых диаграммах представлены значения выбранных анализируемых параметров. Для расчета значений параметров, представленных на диаграммах, использовались различия между значениями для растений, выращенных в контрольных условиях и в условиях дефицита влаги в почве.
Для параметров S , т. е. тех, для которых меньшее снижение в условиях дефицита воды по сравнению с контрольными условиями влажности указывало на лучшую адаптацию к стрессу (E, g s , P n , c i , WUE, F 0 , F M , F V , F V /F M , chla, chlb, chl total, морковь, RWC, урожайность свежей массы плодов, масса отдельного плода), использовалась следующая формула:
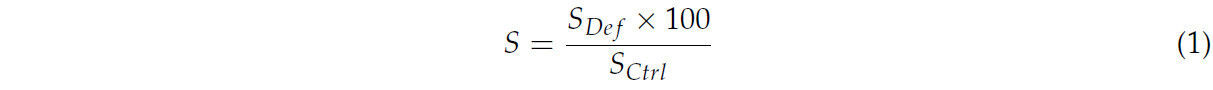

Для параметровD, т. е. тех, для которых меньший прирост в условиях дефицита воды по сравнению с контрольными условиями влажности указывал на лучшую адаптацию к стрессу (TFM, AM), использовалась следующая формула:
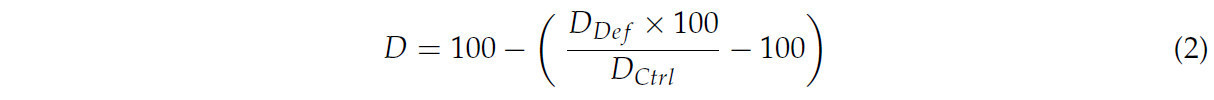
Диаграммы были построены на основе серии анализов, проверяющих значимость различий между средними значениями для отдельных типов объектов с использованием однофакторного дисперсионного анализа (ANOVA). Для средних значений, не различающихся достоверно, а также для значений, демонстрирующих тенденции, отклоняющиеся от общих закономерностей, то есть для параметров S, где значения в условиях дефицита были значительно выше, чем в контрольных условиях, и для параметров D, где значения в условиях дефицита были значительно ниже, чем в контрольных условиях, на радарных диаграммах было присвоено значение 100.
Сравнение изучаемых объектов проводилось путем вычисления площади многоугольника, образованного линией диаграммы.
3. Результаты
3.1 Оценка различий в физиологических признаках, химическом составе и урожайности между генотипами
3.1.1 Параметры газообмена
Результаты показали, что для каждого срока измерения самые высокие значения интенсивности транспирации (E) в условиях контрольной влажности субстрата были характерны для разных сортов ( Таблица 3 ). В первую дату измерения каждого года исследования сорт 'Rugia' демонстрировал значительно более низкое значение E по сравнению с сортом 'Baron von Solemacher' (в среднем на 13,6%) и растениями, полученными из естественной среды (в среднем на 13,0%). Для остальных дат, за исключением третьей даты измерения второго года исследования, сорт 'Rugia' чаще всего демонстрировал самое высокое значение E (в среднем на 35,3%). В последнюю дату измерения эксперимента этот сорт снова имел самое низкое значение E среди протестированных генотипов (на 8,4% ниже, чем 'Baron von Solemacher' и на 42,1% ниже, чем дикие растения).
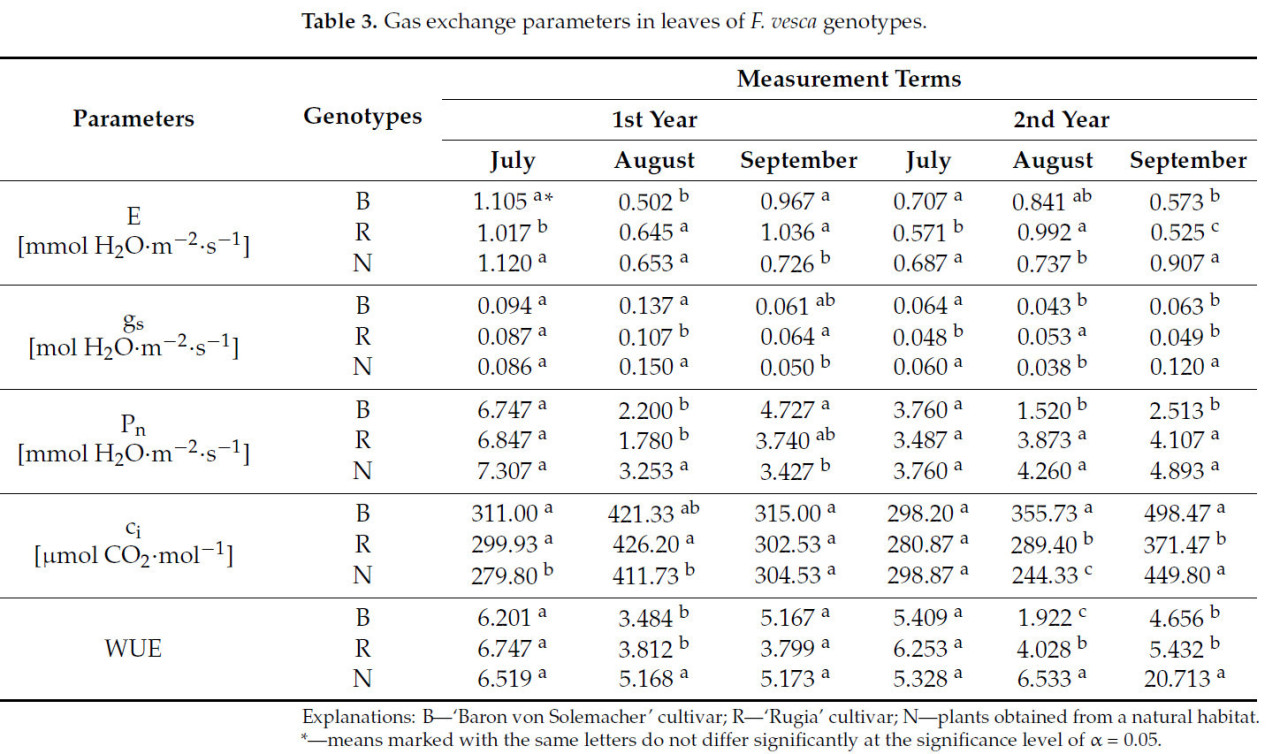
Таблица 3. Параметры газообмена в листьях генотипов F. vesca .
Аналогичные различия наблюдались для устьичной проводимости H2O ( gs ) . В первом семестре первого года исследования существенных различий между генотипами обнаружено не было. Сорт 'Rugia' продемонстрировал наименьшее значение gs во втором семестре первого года (в среднем на 25,3%), первом семестре второго года (в среднем на 22,5%) и, совместно с сортом 'Baron von Solemacher', в третьем семестре второго года (в среднем на 53,3%). Однако во втором семестре второго года наибольшее значение gs наблюдалось у сорта 'Rugia' среди анализируемых генотипов (в среднем на 23,6%).
Существенная дифференциация между анализируемыми генотипами в зависимости от даты измерения наблюдалась также по интенсивности чистой ассимиляции CO2 (Pn ) . В первую дату измерения каждого года различий в значениях Pn между генотипами не обнаружено . Растения, собранные в естественной среде, показали наивысшие значения Pn во вторую дату измерения первого года (в среднем на 38,8%), а также, наряду с сортом 'Rugia', во вторую и третью даты измерения второго года (в среднем на 53,1%).
Как и другие параметры газообмена, подустьичная концентрация CO2 ( c i ) также варьировала в зависимости от даты измерения. Во все даты сорт 'Baron von Solemacher' чаще всего демонстрировал наивысшее значение c i среди исследованных генотипов, особенно во втором триместре второго года исследования, где оно было на 18,6% выше, чем у 'Rugia', и на 31,3% выше, чем у дикорастущих растений.
Различий между исследуемыми генотипами по показателю эффективности фотосинтетического использования воды (ЭИВ) в первом и третьем триместрах первого года и в первом триместре второго года исследований не выявлено. В остальные триместра наибольшие значения ЭИВ наблюдались у растений, собранных в естественной среде обитания (в среднем на 53,2%).
3.1.2 Параметры флуоресценции хлорофилла «а»
В большинстве сроков измерений не наблюдалось различий в исходном (нулевом) значении флуоресценции (F0 ) между исследуемыми генотипами ( таблица 4 ). Исключение составил второй срок измерений второго года, когда наименьшее значение F0 наблюдалось у растений, собранных в естественной среде (в среднем на 14,0%).
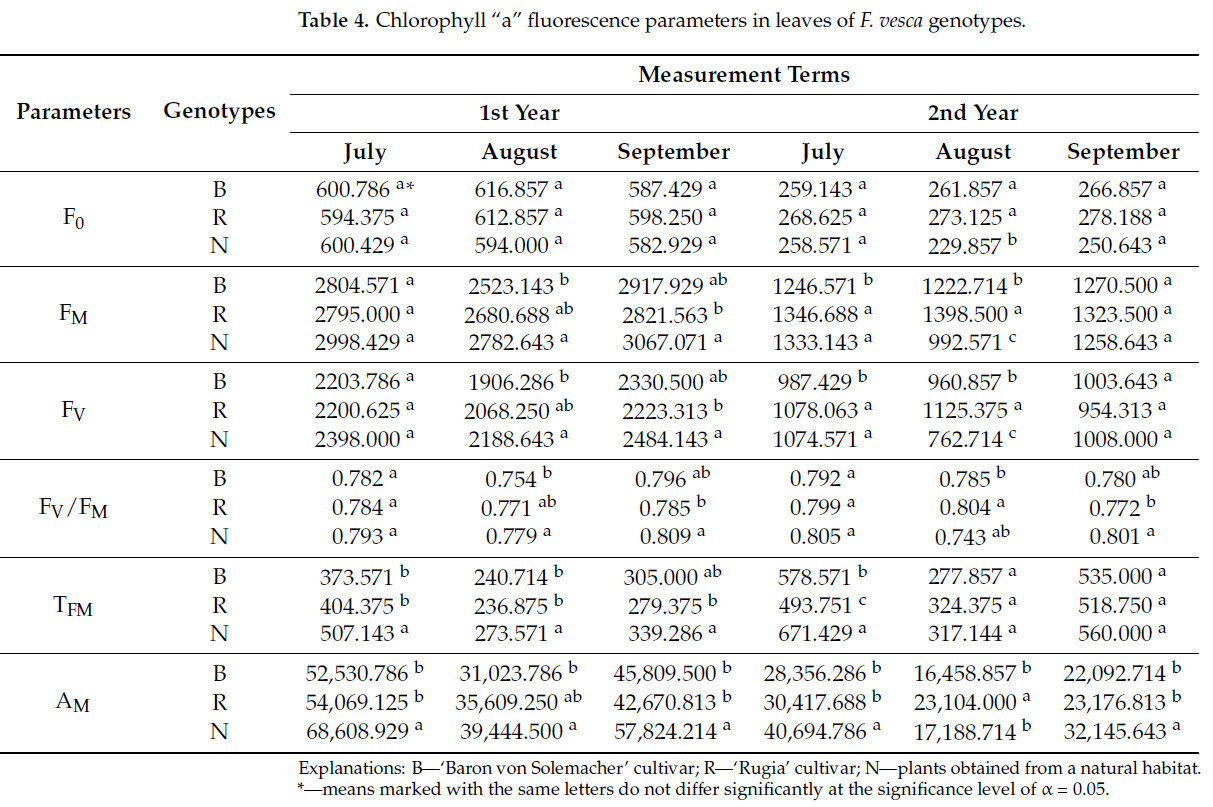
Таблица 4. Параметры флуоресценции хлорофилла «а» в листьях генотипов F. vesca .
По максимальной флуоресценции (F M ) различий между исследуемыми генотипами в первом семестре первого года и третьем семестре второго года исследования не наблюдалось. В остальных измерениях наиболее высокие значения F M чаще всего наблюдались у растений, собранных в естественной среде (например, во втором семестре первого года, где значение было на 9,3% выше, чем у сорта 'Baron von Solemacher', и в первом семестре второго года — на 6,5%); однако эти результаты не были однозначными.
Подобно F M , не наблюдалось различий в переменной флуоресценции (F V ) между исследуемыми генотипами в первом семестре первого года и третьем семестре второго года исследования. В остальные семестра сорт 'Baron von Solemacher' чаще всего характеризовался самым низким значением F V по сравнению с сортом 'Rugia' и растениями, собранными в естественной среде, особенно в первом семестре второго года исследования, где значение было на 8,4% ниже, чем у 'Rugia', и на 8,1% ниже, чем у дикорастущих растений.
Не было обнаружено различий между сравниваемыми генотипами по соотношению переменной к максимальной флуоресценции (F V /F M ) в течение первого триместра как первого, так и второго года исследования. Хотя результаты в другие триместры не были сопоставимы, можно заключить, что сорт 'Baron von Solemacher' чаще всего демонстрировал самые низкие значения F V /F M (например, во втором триместре первого года значение было на 3,2% ниже, чем у дикорастущих растений, а во втором триместре второго года значение было на 2,4% ниже, чем у 'Rugia'), в то время как растения, собранные в естественной среде, характеризовались самыми высокими значениями (например, в третьем триместре второго года значение было на 3,6% выше, чем у 'Rugia').
Во все сроки измерений, за исключением второго и третьего сроков второго года, где существенных различий не наблюдалось, наибольшее время нарастания флуоресценции (Т фм ) было зафиксировано у растений, собранных в естественной среде (в среднем на 17,5%). Однако следует отметить, что в третьем сроке первого года результаты не свидетельствовали об этом однозначно.
Для площади над кривой индукции флуоресценции (А М ) наиболее высокие значения чаще всего наблюдались у растений, собранных в естественной среде обитания (в среднем на 25,3%). Исключением стали второй триместр первого года, где результаты оказались неоднозначными, и второй триместр второго года, когда наибольшее значение А М было зафиксировано у сорта 'Rugia' (в среднем на 27,2%).
3.1.3 Содержание фотосинтетических пигментов
По большинству показателей в течение обоих лет исследования наибольшее содержание хлорофилла «а» в листьях наблюдалось у клубники, собранной в естественной среде, — в среднем на 29,8% ( таблица 5 ). Напротив, самые низкие значения, как правило, регистрировались для сорта «Rugia» (например, в третьем триместре первого года значение было на 27,0% ниже, чем у сорта «Baron von Solemacher» и на 41,4% ниже, чем у диких растений).
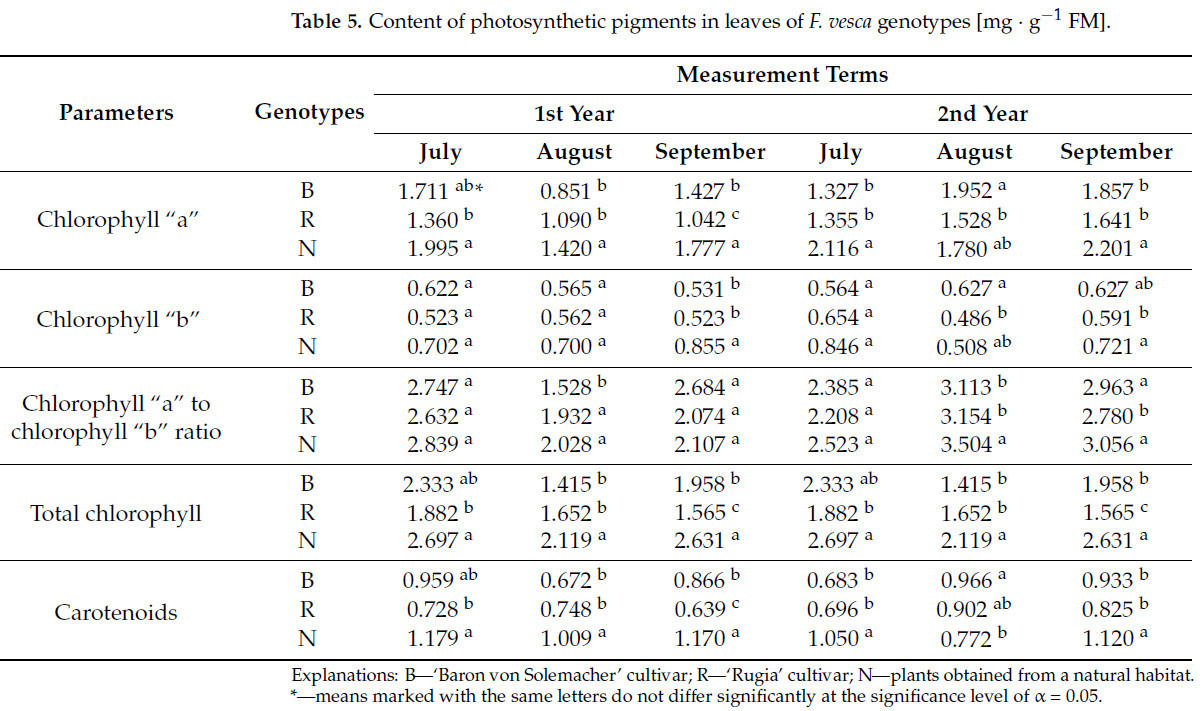
Таблица 5. Содержание фотосинтетических пигментов в листьях генотипов F. vesca [мг · г −1 ФМ].
Во все три периода измерений статистически значимых различий между изучаемыми генотипами по содержанию хлорофилла «b» в листьях не обнаружено. В третьем триместре первого года наибольшее содержание хлорофилла «b» наблюдалось в листьях растений земляники лесной, собранных в естественной среде (в среднем на 31,5%). Во втором триместре второго года сорт 'Baron von Solemacher' показал более высокое содержание хлорофилла «b» по сравнению с сортом 'Rugia' (на 22,5%). В третьем триместре того же года содержание хлорофилла «b» в листьях растений земляники лесной, собранных в естественной среде, было выше на 18,0%, чем у сорта 'Rugia'.
В первом и третьем семестрах первого года, а также в первом семестре второго года исследований различий между сравниваемыми генотипами по соотношению хлорофилла «а» к хлорофиллу «b» не наблюдалось. В остальные сроки исследования наибольшие значения этого показателя были выявлены у растений, собранных в естественной среде (например, во втором семестре второго года значение было на 11,2% выше, чем у сорта 'Baron von Solemacher', и на 10,0% выше, чем у сорта 'Rugia').
Содержание общего хлорофилла в листьях исследованных генотипов F. vesca демонстрировало тенденции, сходные с наблюдаемыми для ранее обсуждавшихся ассимиляционных пигментов. Во все сроки измерений, за исключением первого срока первого года и второго срока второго года, наибольшее содержание общего хлорофилла было обнаружено в листьях растений, собранных в естественной среде (в среднем на 30,3%). В первый срок первого года сорт 'Baron von Solemacher' также показал аналогичное высокое значение этого показателя.
При этом содержание каротиноидов в листьях изучаемых генотипов в большинстве сроков измерений было самым высоким у растений, собранных в естественной среде (в среднем на 30,3%). Исключением стал первый срок измерений первого года, когда сорт 'Baron von Solemacher' продемонстрировал столь же высокое содержание каротиноидов, и второй срок измерений второго года, когда этот сорт превзошел по содержанию каротиноидов землянику лесную, собранную в естественной среде, на 20,1%.
3.1.4 Относительное содержание воды в листьях (ОСЛ)
В первом семестре второго года исследования не наблюдалось значимых различий в относительном содержании воды (ОСВ) между изучаемыми генотипами. Кроме того, результаты, полученные в другие семестра, не позволили однозначно идентифицировать растения с самыми высокими или самыми низкими значениями ОСВ ( таблица 6 ). Только во втором семестре первого года наблюдалось самое высокое ОСВ в листьях растений земляники, собранных в естественной среде (на 15,8% больше, чем у сорта 'Baron von Solemacher', и на 15,2% больше, чем у сорта 'Rugia').
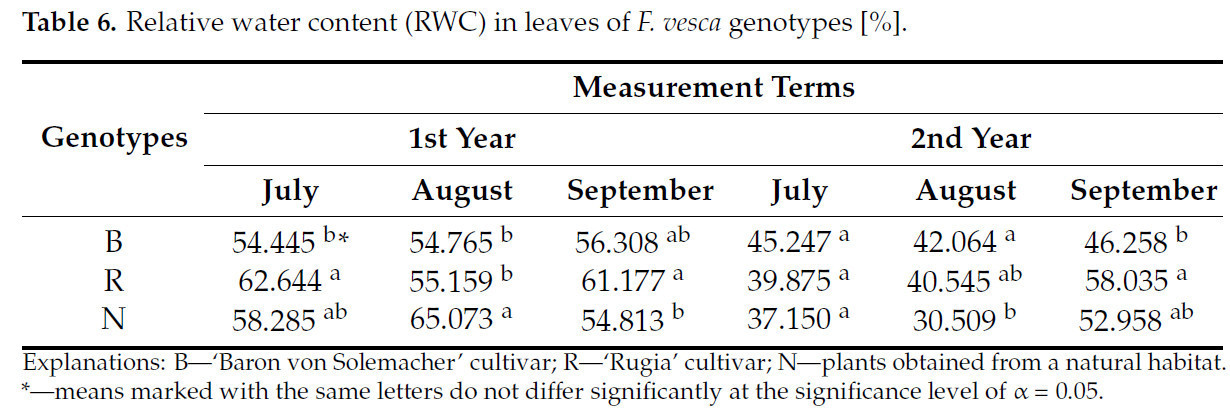
Таблица 6. Относительное содержание воды (RWC) в листьях генотипов F. vesca [%].
3.1.5. Урожайность и вес одного плода лесной земляники
В течение двухлетнего эксперимента цветение, отсутствовавшее у растений, собранных в естественной среде обитания, способствовало формированию плодов. На второй год сорт 'Baron von Solemacher' показал урожайность свежей массы плодов на 29,7% выше, чем сорт 'Rugia'. Однако в оба года исследований существенных различий между сортами 'Rugia' и 'Baron von Solemacher' по массе отдельных плодов не наблюдалось ( таблица 7 ).
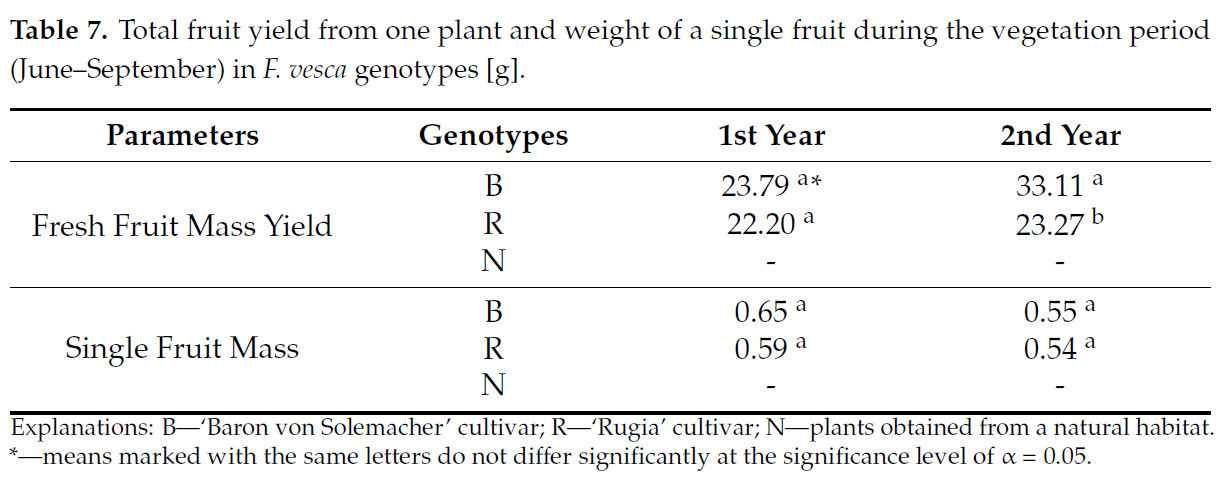
Таблица 7. Общая урожайность плодов с одного растения и масса одного плода за вегетационный период (июнь–сентябрь) у генотипов F. vesca [г].
3.1.6.Сырая и сухая масса корневой системы
По завершении двухлетнего эксперимента была измерена сырая и сухая масса корневой системы. Существенных различий между исследованными генотипами F. vesca по сырой и сухой массе корней не наблюдалось ( таблица 8 ).

Таблица 8. Сырая и сухая масса корневой системы генотипов F. vesca [г].
3.1.7 Химический состав
Анализ среднего содержания элементов в исследованных генотипах F. vesca не выявил существенных различий в концентрациях K, Ca, Mg, Na, Zn, Mn и Fe ( таблица 9 ). Однако сорт 'Rugia' показал содержание Mo на 25,7% ниже, чем у других генотипов, и на 31,7% выше, чем у растений, собранных в естественной среде.
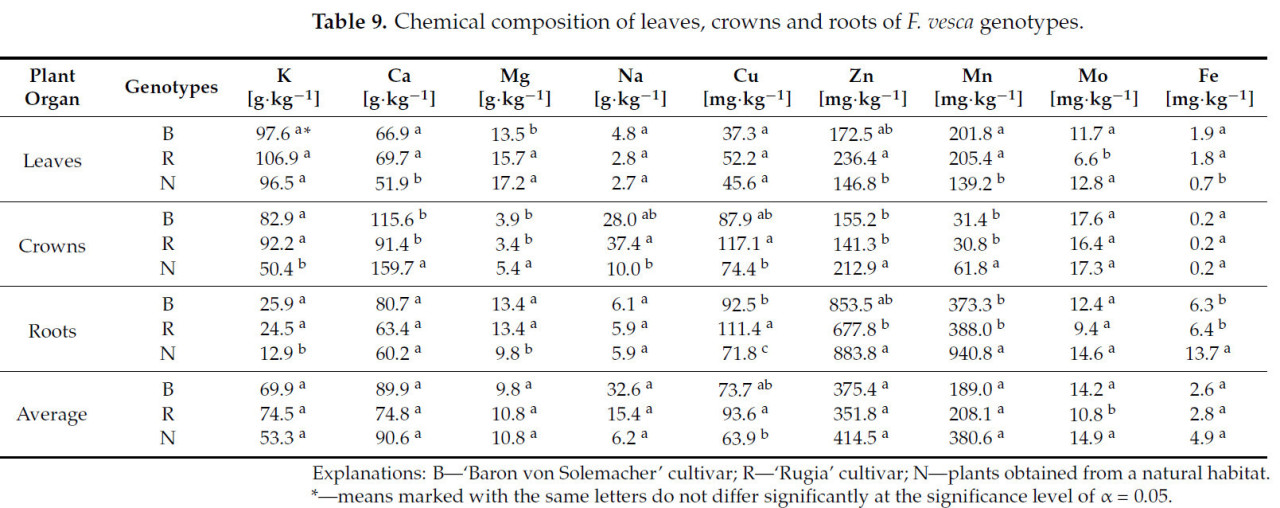
Таблица 9. Химический состав листьев, кроны и корней генотипов F. vesca .
Анализ содержания элементов в отдельных органах растений показал, что у лесной земляники, собранной в естественной среде, в листьях наблюдалось снижение концентрации Ca (в среднем на 24,0%), Mn (на 31,6%) и Fe (на 62,1%) по сравнению с сортами 'Rugia' и 'Baron von Solemacher', а также снижение концентрации Zn на 37,9% по сравнению с сортом 'Rugia'. Содержание Mg в листьях было наименьшим у сорта 'Baron von Solemacher' (в среднем на 17,8%), а содержание Mo – у сорта 'Rugia' (в среднем на 46,0%). Существенных различий между генотипами по содержанию K, Na и Cu в листьях не наблюдалось.
Изученные генотипы не выявили существенных различий в содержании Mo и Fe в кронах. Однако у растений, собранных в естественной среде, наблюдалось более высокое содержание Ca (в среднем на 35,2%), Mg (на 32,4%), Zn (на 30,4%) и Mn (на 49,7%) в кронах по сравнению с обоими сортами, а также более низкое содержание Na (на 274%) и Cu (на 57,4%) по сравнению с сортом 'Rugia'. Кроме того, у этих дикорастущих растений содержание K в кронах было ниже, чем у других генотипов (в среднем на 48,8%).
В корнях генотипы не различались по содержанию Ca, Na и Mo. Тем не менее, у растений из естественной среды обитания наблюдалось более низкое содержание K (в среднем на 48,8%) и Mg (на 26,9%), но более высокое содержание Mn (на 147,2%) и Fe (на 115,8%) в корнях по сравнению с культурными сортами. Содержание Zn в корнях у этих дикорастущих растений также было на 30,4% выше, чем у сорта 'Rugia', при этом содержание Cu было самым низким (в среднем на 29,0%), в отличие от сорта 'Rugia', у которого концентрация Cu в корнях была самой высокой.
3.2 Оценка различий в физиологических признаках, химическом составе и урожайности между генотипами в условиях различной влажности почвы
3.2.1 Параметры газообмена
Двухфакторный дисперсионный анализ (ANOVA) выявил значительное основное влияние фактора уровня влажности субстрата (SML) на различные физиологические параметры. В частности, интенсивность транспирации (E) показала значительное влияние в течение первых четырех сроков измерения, устьичная проводимость для H 2 O (g s ) в течение первых пяти сроков измерения, чистая интенсивность ассимиляции CO 2 (P n ) во все сроки измерения, подустьичная концентрация CO 2 (ci ) в течение первых четырех и последней даты измерения и эффективность использования воды фотосинтезом (WUE) в течение первых трех и пятой даты измерения ( Таблица S1 ). В целом, эти параметры показали снижение в условиях дефицита воды, за исключением g s и c i во вторую дату измерения, а также WUE в первую дату измерения, где наблюдалось увеличение значений (соответственно на 88,5%, 4,1% и 66,4%). Среднее снижение составило 36,7% для E, 30,0% для gs , 23,7% для Pn , 18,2% для ci и 57,6% для WUE ( таблица 10 ).
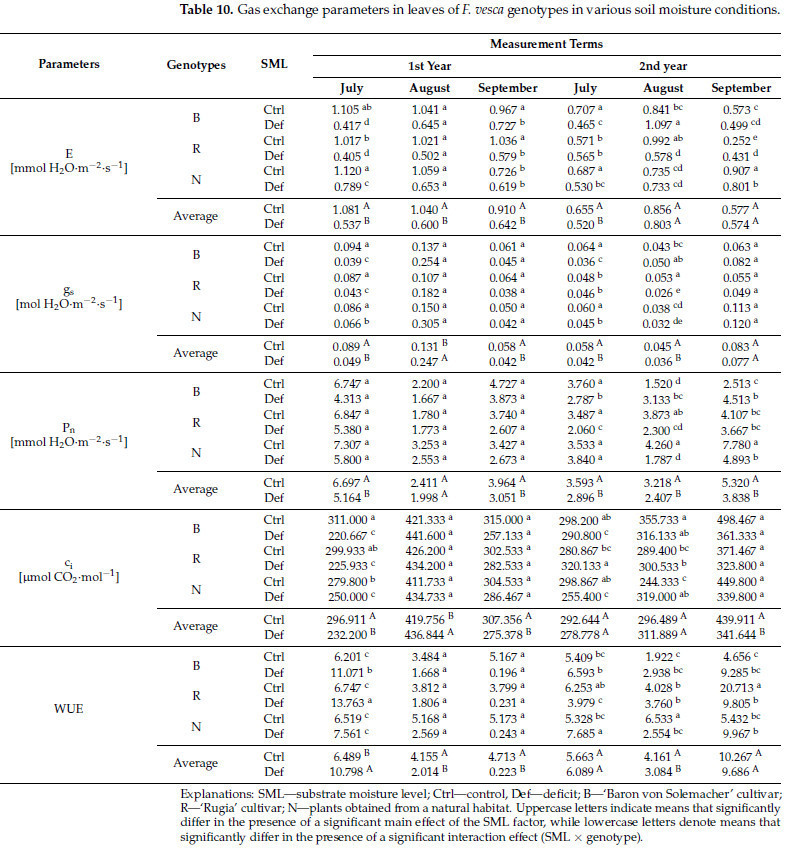
Таблица 10. Параметры газообмена в листьях генотипов F. vesca в условиях различной влажности почвы.
Значимый эффект взаимодействия генотипа × SML на интенсивность транспирации (E) наблюдался во всех терминах измерений, кроме второго. Для устьичной проводимости для H2O ( gs) и подустьичной концентрации CO2 ( ci ) значимые эффекты были обнаружены в первом, четвертом и пятом терминах; для чистой интенсивности ассимиляции CO2 (Pn ) — в последних трех терминах; и для эффективности использования фотосинтетической воды (WUE) — в первых и последних трех терминах измерений ( таблица S1 ). Постанализ выявил более выраженное снижение E в условиях дефицита воды у сортов 'Rugia' и 'Baron von Solemacher' (соответственно, в среднем на 48,7% и 40,4%, тогда как у диких растений оно составило 26,2%). Для g s сорта показали большее снижение в первом члене измерения (58,5% у 'Baron von Solemacher', 50,6% — у 'Rugia' и 23,3% — у дикорастущих растений). Однако в четвертом члене 'Rugia' продемонстрировала меньшее изменение по сравнению с растениями из естественной среды (статистически значимое изменение отсутствует по сравнению с 25,0% снижением), тогда как в пятом члене 'Baron von Solemacher' показал меньшее снижение по сравнению с другими растениями (50,9% и статистически значимое изменение отсутствует). В случае P n результаты были относительно непоследовательными. Например, у 'Baron von Solemacher' увеличение P n наблюдалось в последних двух членах измерения (на 106,1% и 79,6%). Однако в остальных членах значения P n в целом снижались, причем наибольшее снижение наблюдалось у растений из естественной среды (в среднем на 47,6%). Что касается c i , растения из естественной среды показали наименьшее снижение значения в первом сроке измерений (10,7% по сравнению со средним значением 26,9%). В четвертом сроке значения c i увеличились у сорта 'Rugia' на 14,0%, аналогично пятому сроку у растений из естественной среды (на 30,6%). Напротив, у сорта 'Baron von Solemacher' наблюдалось снижение значений c i на 2,5% в четвертом сроке. Суммируя изменения значений WUE по срокам измерений, самые неблагоприятные изменения наблюдались у сорта 'Rugia', который преимущественно отреагировал на дефицит воды снижением этого параметра (в среднем на 44,5%) ( таблица 10 ).
3.2.2 Параметры флуоресценции хлорофилла «а»
Значимый основной эффект фактора SML на F 0 наблюдался исключительно на четвертую дату измерения. Для F M и Fᵥ значимые эффекты были отмечены во вторых и последних трех терминах измерения, тогда как для T FM они наблюдались в течение первых двух и последних трех терминов. Аналогично, A M продемонстрировал значимые эффекты в последние три даты. Однако не было обнаружено значимых различий для F V / F M по отношению к фактору SML ни в одну из дат измерения ( таблица S2 ). Общее снижение значений F 0 (на 3,8%), F V (в среднем на 7,5%) и F M (в среднем на 12,1%) наблюдалось во все проанализированные даты, за исключением второй даты измерения, когда значения F V и F M были выше в условиях дефицита воды (соответственно, на 6,6% и 5,1%). Напротив, значения T FM и A M увеличились в условиях дефицита воды (соответственно, в среднем на 29,7% и 27,8%) ( таблица 11 ).
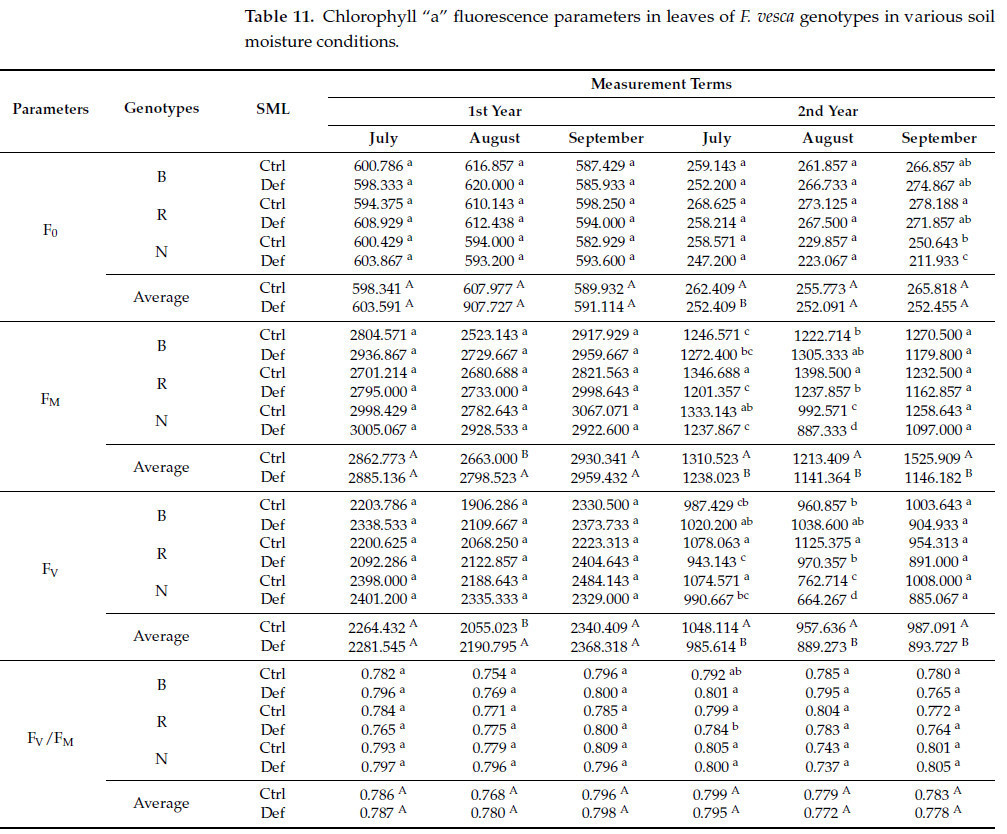
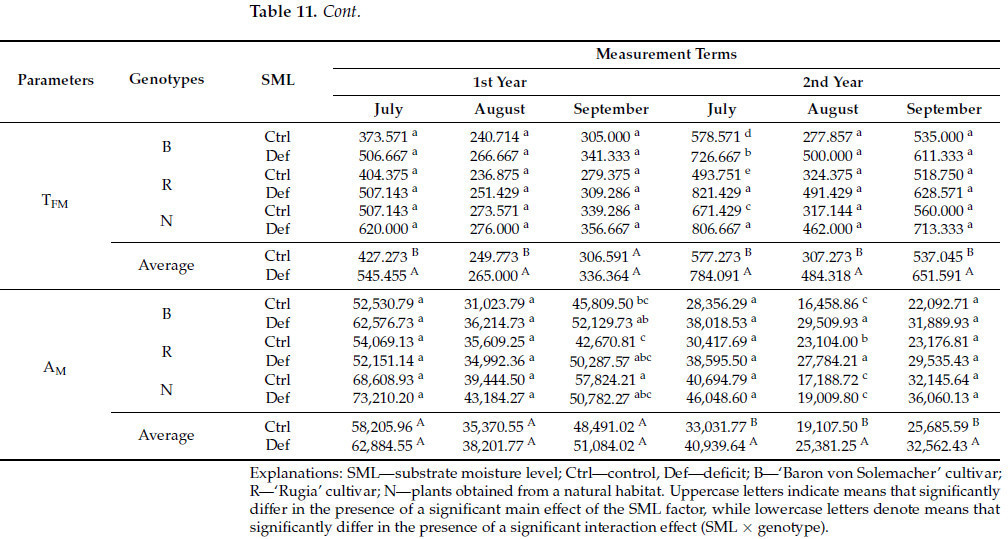
Таблица 11. Параметры флуоресценции хлорофилла «а» в листьях генотипов F. vesca в условиях различной влажности почвы.
Значимый эффект взаимодействия генотипа × SML на F 0 наблюдался исключительно в последний срок измерения. Для F M и F V значимые эффекты были отмечены в четвертом и пятом сроке измерения, тогда как для F V /F M и T FM они наблюдались в четвертом сроке. Значимые эффекты для A M были обнаружены в третьем, пятом и шестом сроках измерения ( таблица S2 ). Что касается F 0 , наиболее выраженные изменения в условиях дефицита воды произошли у растений, полученных из естественной среды (уменьшение на 15,4%). Для F M и F V наибольшее снижение в четвертую и пятую даты измерения наблюдалось у сорта 'Rugia' (соответственно, в среднем на 11,1% и 13,1%). Аналогичная тенденция была очевидна для F V /F M , где более существенное снижение было отмечено у 'Rugia' (на 1,9%). В случае сорта T FM самые неблагоприятные изменения в условиях дефицита воды зафиксированы у сорта 'Rugia' (увеличение на 66,4%), а в случае сорта A M – у сорта 'Baron von Solemacher' (увеличение на 79,3%) ( таблица 11 ).
3.2.3 Содержание фотосинтетических пигментов
Значительное основное влияние фактора SML на содержание хлорофилла «a» наблюдалось во втором, третьем, четвертом и пятом членах измерения. Для хлорофилла «b» значительные эффекты были обнаружены в первых трех и пятом членах. Соотношение хлорофиллов «a» и «b» показало значительные эффекты во втором и пятом членах, в то время как общее содержание хлорофилла значительно влияло во втором, третьем и пятом членах. Содержание каротиноидов показало значительные эффекты в первых пяти членах измерения ( Таблица S3 ). Однако результаты продемонстрировали заметную изменчивость. В первый, второй, четвертый и пятый сроки измерений в условиях дефицита воды наблюдалось увеличение содержания хлорофилла «а» (в среднем на 23,5%), хлорофилла «b» (в среднем на 28,2%), общего хлорофилла (в среднем на 27,5%) и каротиноидов (в среднем на 21,3%), сопровождавшееся уменьшением соотношения хлорофиллов «а» и «b» (в среднем на 7,8%). Напротив, в третью дату измерений было зафиксировано уменьшение содержания хлорофилла «а» (на 11,2%), хлорофилла «b» (на 19,3%), общего хлорофилла (на 13,7%) и каротиноидов (на 9,0%) ( таблица 12 ).
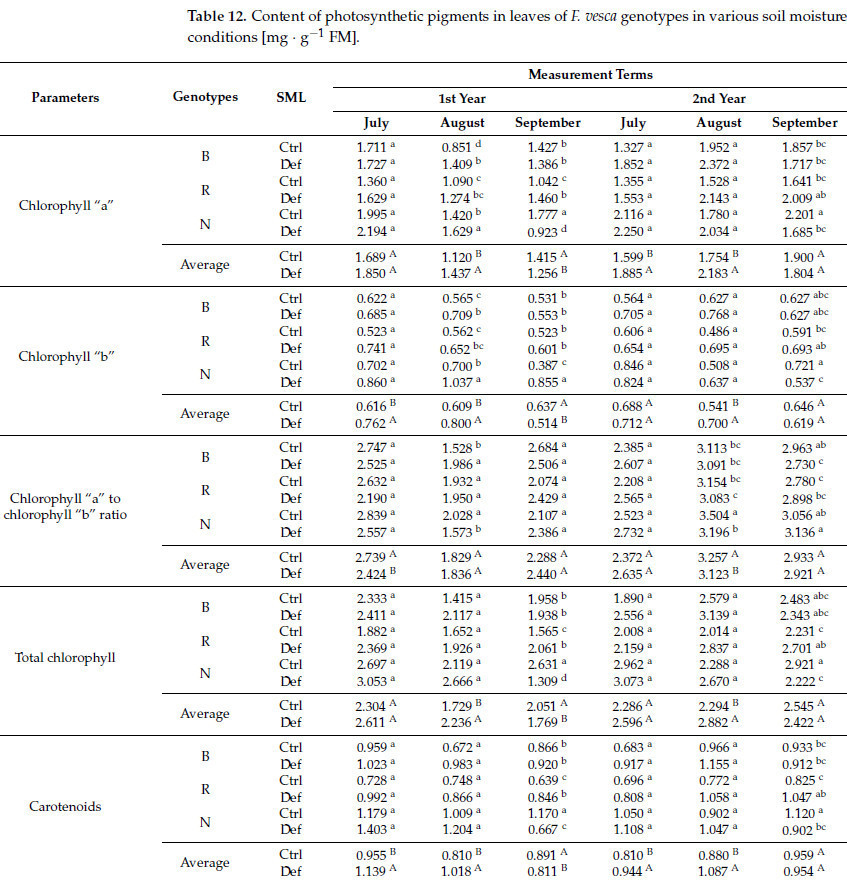
Таблица 12. Содержание фотосинтетических пигментов в листьях генотипов F. vesca в условиях различной влажности почвы [мг · г −1 ФМ].
Значимый эффект взаимодействия генотип × SML на содержание хлорофилла «а» и хлорофилла «b» наблюдался во втором, третьем и последнем терминах измерений. Для соотношения хлорофилла «а» к «b» значимые эффекты были отмечены во втором, пятом и шестом терминах, в то время как для общего содержания хлорофилла и каротиноидов значимые эффекты были зарегистрированы в третьем и шестом терминах ( таблица S3 ). Для хлорофилла «а» все генотипы отреагировали на дефицит воды во вторую дату измерения увеличением его содержания, при этом наиболее выраженное изменение наблюдалось у сорта 'Baron von Solemacher' (на 65,6%). Однако в третьем и шестом терминах растения из естественной среды показали снижение содержания хлорофилла «а» (в среднем на 35,8%). Аналогичная закономерность наблюдалась для хлорофилла «b», где увеличение содержания отмечалось в более ранние сроки (в среднем на 64,9%), но снижение на 25,5% произошло у дикорастущих растений в последний срок. Следовательно, наибольшее снижение общего содержания хлорофилла было зарегистрировано у дикорастущих растений (в среднем на 37,1%). Для соотношения хлорофилла «a» к «b» увеличение на 30,0% наблюдалось у культурных растений во второй срок измерения, в то время как у дикорастущих растений наблюдалось снижение на 22,4%. В пятый срок измерения наиболее существенное снижение этого соотношения было зарегистрировано у дикорастущих растений (на 8,8%), тогда как в шестой срок наибольшее снижение наблюдалось у сорта 'Baron von Solemacher' (на 7,9%). Что касается каротиноидов, то снижение содержания наблюдалось исключительно у дикорастущих растений в третий и шестой сроки (в среднем на 31,2%), тогда как сорт 'Rugia' продемонстрировал наиболее значительное увеличение содержания каротиноидов (в среднем на 29,7%) ( таблица 12 ).
3.2.4 Относительное содержание воды в листьях (ОСЛ)
Значительное влияние фактора SML на относительное содержание воды (ОВС) наблюдалось в первом, втором и пятом периодах измерения ( таблица S4 ). Во всех случаях значения ОВС снижались в условиях дефицита воды в среднем на 14,4% ( таблица 13 ).
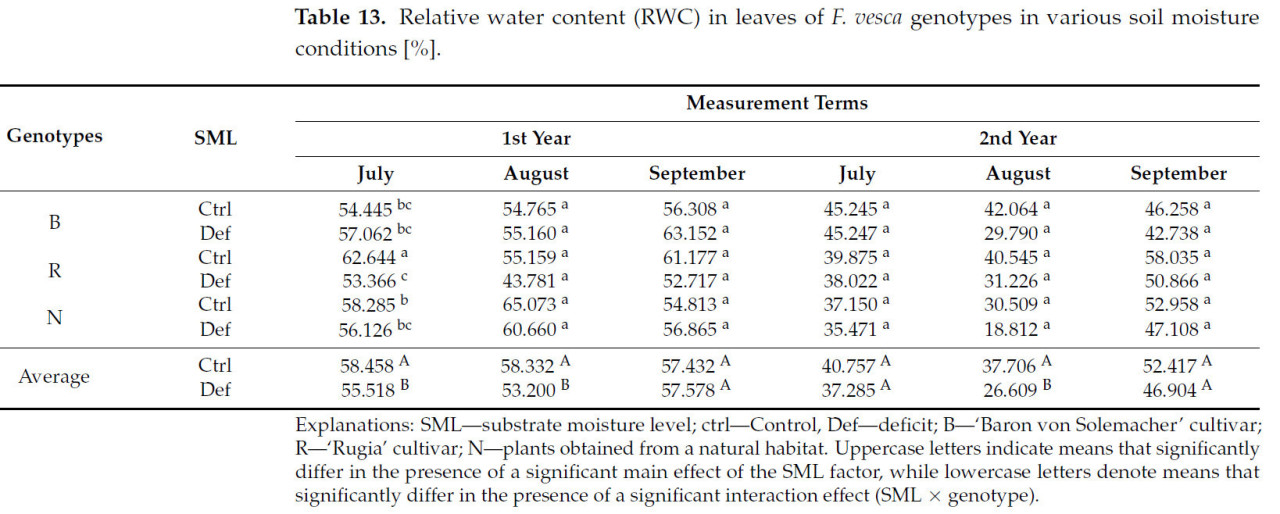
Таблица 13. Относительное содержание воды (ОСВ) в листьях генотипов F. vesca в условиях различной влажности почвы [%].
Значимый эффект взаимодействия генотипа × SML на RWC наблюдался только в первом периоде измерений ( таблица S4 ). В этот период наиболее существенное снижение RWC было зафиксировано у растений сорта «Rugia» (на 14,8%) ( таблица 13 ).
3.2.5. Урожайность и вес одного плода лесной земляники
У растений, полученных из естественной среды, цветение не наблюдалось, и, следовательно, они были исключены из анализа массы плодов, поскольку не дали плодов. Значительное влияние фактора SML было выявлено во все годы исследования как на урожайность свежей массы плодов, так и на массу отдельного плода ( таблица S5 ). В каждом случае растения, подвергавшиеся дефициту воды, давали плоды меньшей массы. Это представляло собой среднее снижение урожайности свежей массы плодов на 62,9% и среднее снижение массы отдельного плода на 35,6% ( таблица 14 ).
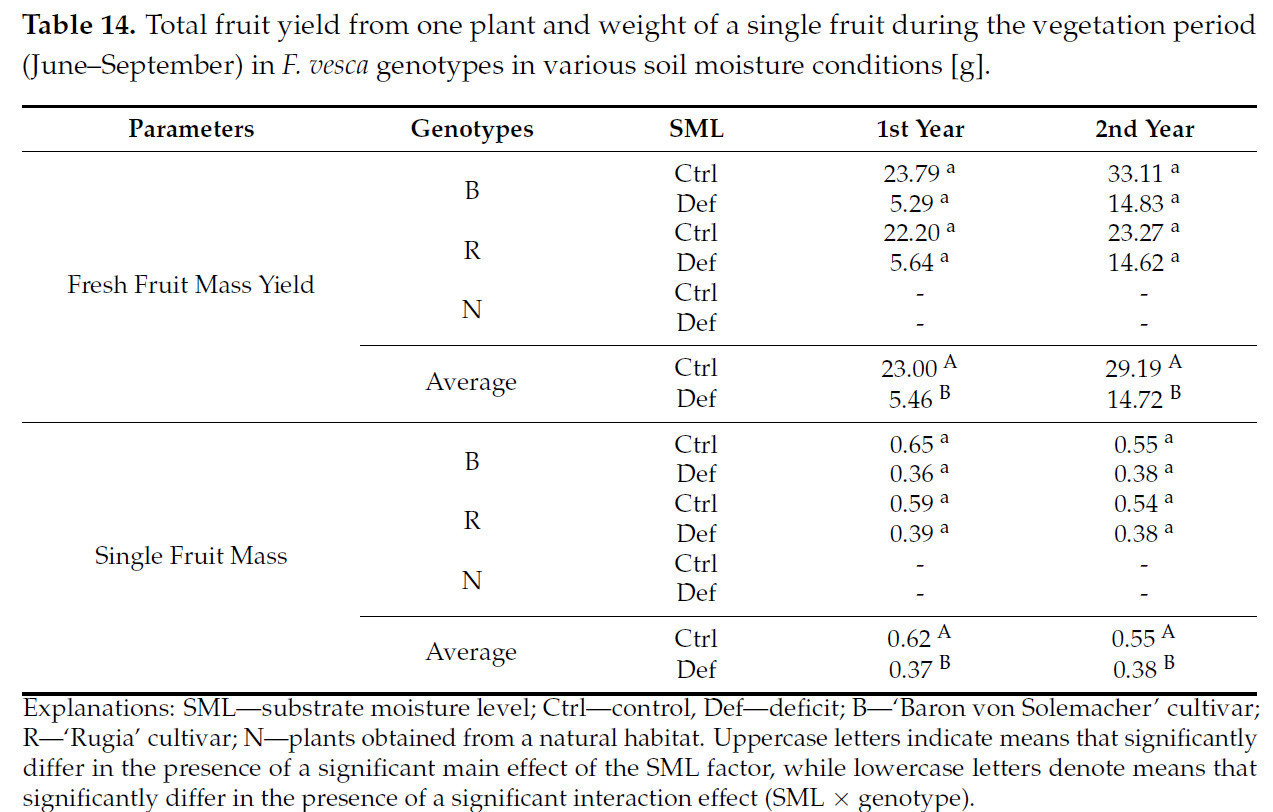
Таблица 14. Общая урожайность плодов с одного растения и масса одного плода за вегетационный период (июнь–сентябрь) у генотипов F. vesca в условиях различной влажности почвы [г].
Однако существенного влияния взаимодействия SML × генотип не наблюдалось ни в один из годов исследования ни на урожайность свежей массы плодов, ни на массу отдельного плода ( таблица S5 ).
3.2.6.Сырая и сухая масса корневой системы
Ни основной эффект SML, ни взаимодействие SML × генотип не были значимыми для сырой и сухой массы корневой системы F. vesca ( таблица S6 и таблица 15 ).
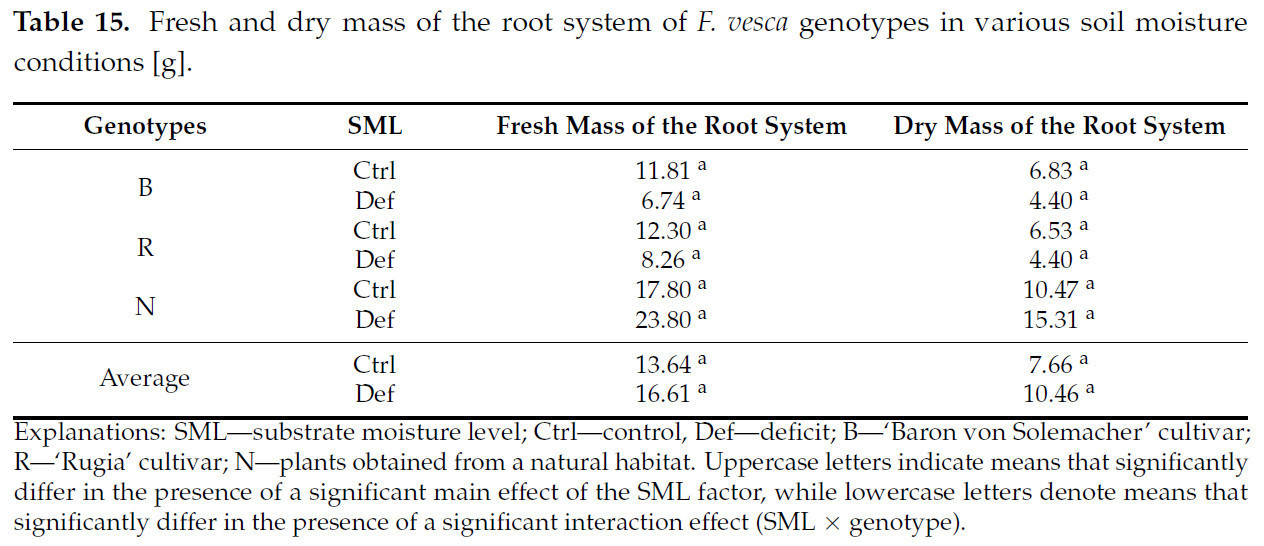
Таблица 15. Сырая и сухая масса корневой системы генотипов F. vesca в условиях различной влажности почвы [г].
3.2.7 Химический состав
Для содержания отдельных элементов в листьях значительный основной эффект фактора SML наблюдался для калия (K), магния (Mg), цинка (Zn) и марганца (Mn) ( таблица S7 ). В каждом случае содержание этих элементов увеличивалось в условиях засухи (соответственно на 8,5%, 10,9%, 32,8% и 20,6%) ( таблица 15 ). Значительный эффект взаимодействия SML × генотип был обнаружен для кальция (Ca), марганца (Mn) и железа (Fe) ( таблица S7 ). Наиболее выраженные изменения наблюдались у растений, полученных из естественной среды, которые показали более высокое содержание Zn (на 96,3%), Mn (на 62,2%) и Fe (на 157,1%) ( таблица 16 ).
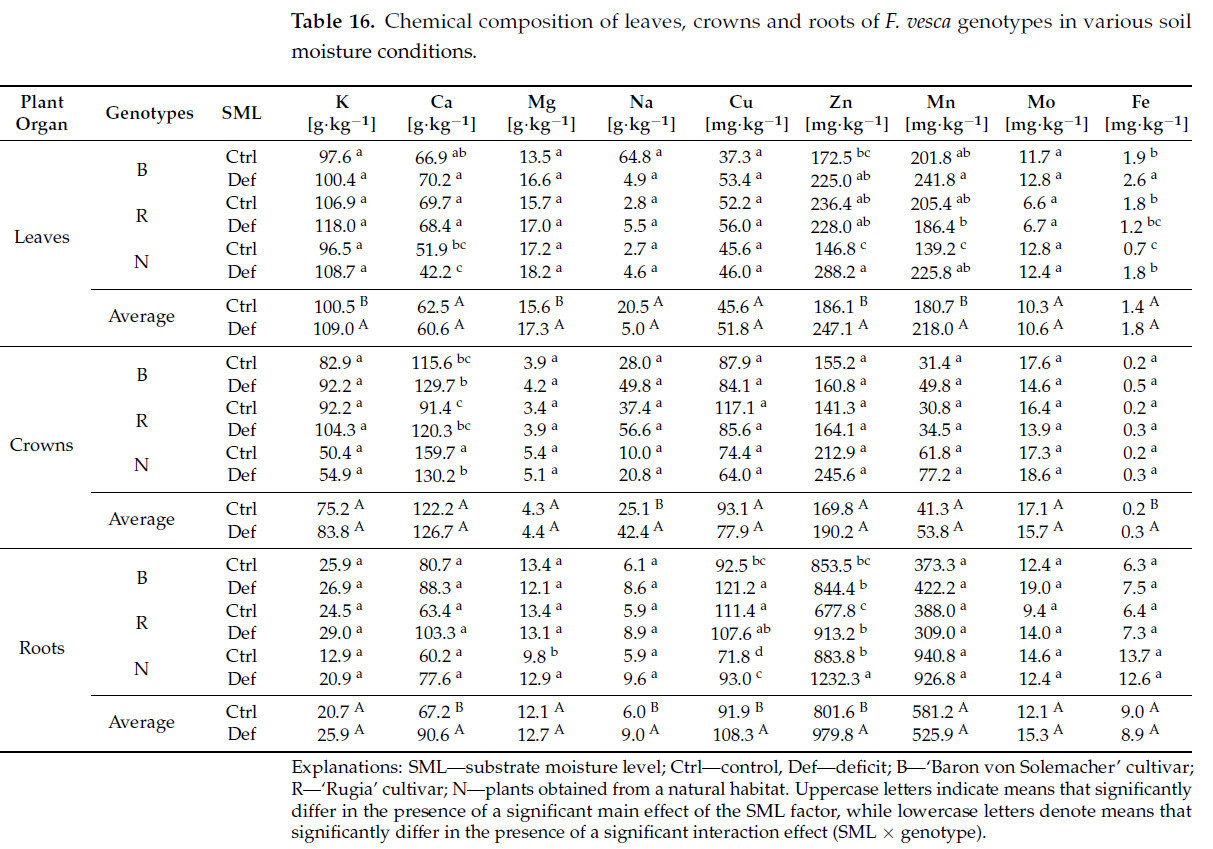
Таблица 16. Химический состав листьев, кроны и корней генотипов F. vesca в различных условиях влажности почвы.
Что касается содержания элементов в кронах, то значимый основной эффект фактора SML наблюдался для натрия (Na) и железа (Fe) ( таблица S7 ), причем содержание обоих элементов увеличивалось в условиях засухи (соответственно на 68,9% и 50,0%) ( таблица 16 ). Значимый эффект взаимодействия SML × генотип был выявлен только для кальция (Ca) ( таблица S7 ). Значимое снижение содержания Ca наблюдалось исключительно у растений, собранных в естественной среде (на 18,5%). Напротив, у культурных сортов содержание Ca увеличилось (в среднем на 21,9%); однако это увеличение не было статистически значимым.
Для содержания элементов в корнях значительный основной эффект фактора SML наблюдался для Ca, Na, Zn и меди (Cu) ( таблица S7 ), при этом содержание каждого элемента увеличивалось в условиях засухи (соответственно на 34,8%, 50,0%, 22,2% и 17,8%) ( таблица 15 ). Значительный эффект взаимодействия SML × генотип был выявлен для Mg, Cu и Zn ( таблица S7 ). Значительное увеличение содержания Mg в корнях в условиях дефицита воды наблюдалось исключительно у растений, полученных из естественной среды (на 31,6%). Эти растения, наряду с сортом 'Rugia', также показали наиболее выраженное увеличение содержания Zn (на 34,7% у 'Rugia' и на 39,4% у диких растений). Напротив, увеличение содержания Cu наблюдалось в сорте «Барон фон Солемахер» (на 31,0%) и в растениях, полученных из естественной среды (на 29,5%) ( таблица 16 ).
3.2.8. Радарные карты
На рисунке 1 представлен анализ изменений средних значений параметров, используемых для оценки физиологических характеристик растений F. vesca , сгруппированных по датам измерений и генотипам. Судя по площади графика, сорт 'Baron von Solemacher' продемонстрировал наименьшую адаптацию к условиям дефицита воды в субстрате и наиболее сильную негативную реакцию на стресс, вызванный засухой (на что указывает наименьшая площадь под кривой) в течение первого и второго триместров первого года исследований. Сорт 'Rugia' продемонстрировал наименьшую адаптацию в течение третьего триместра измерений обоих лет исследований, в то время как растения, собранные в естественной среде, показали наименьшую адаптацию в течение первого и второго триместров второго года.
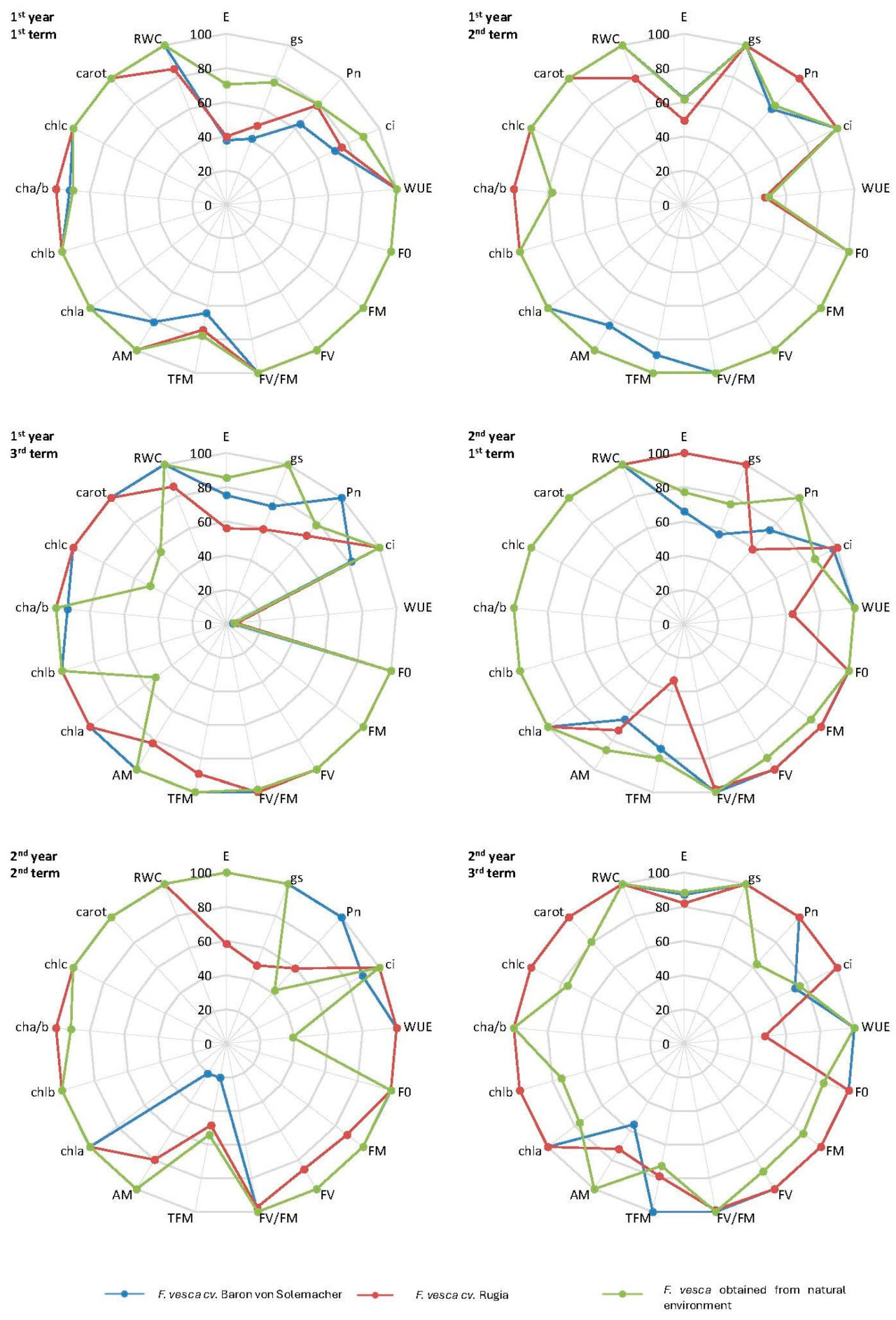
Рисунок 1. Изменения средних значений отдельных физиологических параметров генотипов F. vesca в условиях дефицита воды в субстрате по сравнению со средними значениями в условиях контрольной влажности. Пояснения: E — скорость транспирации H 2 O, g s — устьичная проводимость H 2 O, P n — чистая скорость ассимиляции CO 2 , c i — подустьичная концентрация CO 2 , WUE — коэффициент использования воды фотосинтезом, F 0 — начальная флуоресценция, F M — максимальная флуоресценция, F V — переменная флуоресценция, F V /F M — максимальная, потенциальная эффективность фотохимической реакции в ФС II, T FM — время роста флуоресценции хлорофилла, A M — площадь над индукционной кривой флуоресценции хлорофилла, chla — содержание хлорофилла «a», chlb — содержание хлорофилла «b», chla/b — отношение содержания хлорофилла «a» к «b», chlc — общее содержание хлорофилла, carot — содержание каротиноидов, RWC — относительный индекс содержания воды в листьях.
Напротив, сорт Baron von Solemacher продемонстрировал наибольшую адаптацию к засушливым условиям (что отражено наибольшей площадью под кривой) на третью дату первого года исследования и на вторую и третью дату второго года исследования. Растения, полученные из естественной среды, продемонстрировали наибольшую адаптацию на первую дату измерения обоих лет, в то время как сорт Rugia продемонстрировал наибольшую адаптацию на вторую дату первого года исследования.
4. Обсуждение
Физиологические признаки сортов культурных растений, в том числе рода Fragaria , демонстрируют существенную изменчивость, обусловленную как генетическими, так и экологическими факторами. Исследования показывают, что различия в параметрах газообмена, флуоресценции хлорофилла и урожайности выражены у разных видов и сортов Fragaria , что подчеркивает критическую роль генетического разнообразия в этих признаках [ 40 , 41 , 42 ]. Эти выводы согласуются с результатами нашего исследования. Хотя различия в физиологических признаках между сортами Fragaria хорошо документированы, важно учитывать, что факторы окружающей среды также могут оказывать глубокое влияние на эти характеристики, иногда затмевая генетические эффекты. Эта сложность подчеркивает необходимость продолжения исследований взаимодействий генотипа и окружающей среды, которые формируют физиологические реакции растений.
Физиологические характеристики растений F. vesca, растущих в естественной среде обитания, включая флуоресценцию хлорофилла и содержание ассимиляционных пигментов, превзошли таковые, наблюдаемые у культурных сортов в нашем исследовании, что, вероятно, отражает их адаптацию к естественным условиям. Эти растения процветают в разнообразных и часто сложных местах обитания, что повышает их способность к физиологической адаптации [ 43 , 44 ]. Механизмы адаптации к стрессу засухи у растений, как культурных, так и дикорастущих, имеют фундаментальные сходства. Однако масштаб и эффективность этих механизмов могут значительно различаться из-за различных экологических давлений, испытываемых дикими растениями [ 45 ]. Дикие растения показали менее выраженное снижение транспирации в ответ на тот же стресс засухи по сравнению с культурными сортами растений, что предполагает более низкую чувствительность к этому типу стресса.
Наблюдаемые различия в параметрах газообмена между дикими и культурными растениями F. vesca в условиях водного стресса можно отнести к присущему генетическим разнообразию и адаптивным признакам, присутствующим в диких разновидностях. Дикие F. vesca эволюционировали в естественных условиях, где водный стресс является общей проблемой, что привело к развитию физиологических и биохимических механизмов, которые повышают устойчивость к засухе. Напротив, культурные разновидности были селективно выведены по признакам, которые могут не отдавать приоритет устойчивости к засухе, что приводит к более выраженному снижению газообмена в аналогичных стрессовых условиях [ 46 , 47 ]. Большая генетическая изменчивость, присутствующая в природных популяциях из-за неконтролируемого скрещивания и мутаций, по сравнению с культурными разновидностями, приводит к более широкому диапазону адаптивных ответов на стрессовые факторы окружающей среды [ 48 ]. Эта генетическая изменчивость имеет решающее значение для экспрессии генов, чувствительных к засухе, которые способствуют улучшенному газообмену в условиях ограниченного количества воды. Например, исследования диких сородичей других культур, таких как арахис ( Arachis hypogaea L.), показали, что они обладают аллелями, которые обеспечивают устойчивость к абиотическим стрессам, включая засуху, что можно использовать для улучшения культурных сортов [ 49 ].
Культивируемые сорта F. vesca часто подвергаются воздействию агротехнических приемов, в которых урожайность ставится выше устойчивости к стрессу. Такая селекция может снижать экспрессию генов, связанных с устойчивостью к засухе, делая эти растения более восприимчивыми к водному стрессу. Следовательно, культурные сорта могут демонстрировать более выраженное снижение параметров газообмена при водном стрессе, поскольку у них отсутствуют мощные адаптивные черты, присущие их диким аналогам [ 50 ].
Похожие обработки могут вызывать противоположные изменения в других адаптивных механизмах. Наблюдаемое большее снижение содержания фотосинтетических пигментов в листьях растений F. vesca из естественной среды обитания в условиях дефицита почвенной влаги по сравнению с культурными сортами может быть результатом селективных методов селекции, благоприятствующих более высокой урожайности. Культурные сорта могут обладать улучшенными механизмами поддержания уровня пигментов в условиях стресса, такими как улучшенная эффективность использования воды и осмотическая регуляция [ 51 , 52 , 53 ]. Хотя растения могут изначально увеличивать выработку фотосинтетических пигментов при умеренном стрессе, сильный дефицит воды может подавлять эту реакцию, в конечном итоге приводя к чистому снижению содержания пигментов [ 51 , 52 ]. Дикие растения F. vesca , часто подвергающиеся колебаниям уровня влажности и другим абиотическим стрессам, могут демонстрировать более выраженную физиологическую реакцию на дополнительные стрессоры, такие как засуха [ 54 ]. Напротив, культурные сорта, обычно выращиваемые в более контролируемых условиях, могут лучше переносить водный стресс без значительного снижения содержания пигментов [ 54 ].
Анализ минерального состава показал, что дикие растения демонстрируют более существенные колебания содержания отдельных элементов в зависимости от условий влажности почвы, что свидетельствует о большей адаптивной гибкости. В контрольных условиях эти растения имели более высокое содержание кальция, в то время как при дефиците влаги уровни таких элементов, как калий и марганец увеличивались. Увеличение содержания калия связано с механизмами выживания растений во время стресса засухи [ 55 ]. Повышенные уровни калия могут также функционировать как механизм толерантности, ограничивая поглощение токсичных ионов корнями [ 56 , 57 ]. Марганец, критический компонент фотосинтетических белков и ферментов, играет центральную роль в реакции расщепления воды, которая поставляет электроны для последующих стадий фотосинтеза. Его дефицит может значительно ухудшить активность фотосистемы II (ФСII) [ 58 ]. Следовательно, наблюдаемое увеличение содержания марганца в тканях растений, подвергшихся воздействию засухи, вероятно, отражает активацию физиологических процессов, направленных на смягчение неблагоприятного воздействия дефицита воды на метаболизм растений.
Напротив, возделываемые сорта показали меньше или менее выраженные различия в содержании минералов в условиях различной влажности, затрагивая только выбранные элементы. Например, сорт 'Baron von Solemacher' показал повышенное содержание натрия и меди в условиях засухи, тогда как 'Rugia' показал большую стабильность в своем минеральном составе независимо от уровня влажности почвы. Натрий способствует осмотической адаптации в тканях растений в условиях низкого внешнего водного потенциала. У видов C4 натрий также играет роль как в световой, так и в темновой фазах фотосинтеза; например, он способствует транспорту пирувата в мезофилл хлоропласта и поддерживает структурную целостность гранул хлоропласта [ 59 ]. Медь, тем временем, имеет решающее значение на клеточном уровне, влияя на механизмы транскрипции и транспорта белков, окислительное фосфорилирование и накопление железа, каждый из которых существенно формирует реакцию растения на стресс засухи [ 60 ].
Другие исследования показывают, что дикие растения развивают более универсальные механизмы адаптации к изменяющимся и сложным условиям окружающей среды, что подтверждается их большим генетическим разнообразием. Например, исследования различных генотипов растений показывают, что дикие растения производят более высокие уровни антиоксидантных ферментов и осмопротекторов, которые повышают их выживаемость в условиях стресса [ 61 ]. Дикие растения также продемонстрировали более выраженные различия в содержании элементов в зависимости от уровня влажности субстрата. Их листья, выращенные в контрольных условиях, содержали больше кальция (Ca), тогда как в условиях засухи они показали повышенные концентрации калия (K), натрия, марганца и железа. Это предполагает способность динамически регулировать внутренний транспорт и распределение минералов в ответ на засуху [ 62 ], реакцию, характерную для диких растений, которые должны адаптироваться к естественно изменяющимся условиям окружающей среды, где доступность воды варьируется. Для сравнения, культурные сорта, которые были селективно выведены в первую очередь для высокой урожайности и однородности, могут не демонстрировать такой широкий спектр адаптивных реакций в условиях стресса [ 63 ].
Различия в содержании элементов между корнями диких растений и культурных сортов были еще более выражены. Корни диких растений показали более высокие концентрации калия, магния, натрия, меди и цинка, что указывает на повышенную способность эффективно поглощать и хранить минералы в условиях дефицита. В то время как повышенное накопление натрия и меди наблюдалось в корнях сорта 'Baron von Solemacher', а повышение натрия и цинка в корнях 'Rugia', эта реакция на дефицит воды была более обширной у диких растений, с более широким диапазоном накопления и перераспределения минералов [ 64 ]. Естественные популяции демонстрируют большее генетическое разнообразие, что обеспечивает более широкий спектр ответов на стрессовые факторы окружающей среды, такие как низкая влажность почвы. Эта генетическая изменчивость имеет решающее значение для эффективного поглощения и транслокации необходимых минералов, как показано в исследованиях, подчеркивающих роль генетической биообогащения у диких родственников культур [ 65 ]. Более того, дикие растения часто демонстрируют значительную фенотипическую пластичность, что позволяет им изменять характеристики корней и листьев в ответ на колебания уровня влажности, как показано в исследованиях Plantago lanceolata [ 66 ]. Виды, адаптированные к почвам с низким содержанием питательных веществ, также развивают специализированную морфологию корней и стратегии усвоения питательных веществ, которые улучшают усвоение минералов в условиях стресса [ 67 ].
Сорта, вероятно, были селекционированы с упором на урожайность, а не на устойчивость к изменчивости окружающей среды, что может ограничить их способность адаптироваться к условиям низкой влажности почвы. Такая ориентация на урожайность может привести к снижению генетического разнообразия и сужению спектра функциональных признаков, что потенциально снижает их способность накапливать необходимые минералы в сложных условиях.
В условиях дефицита воды (стресс засухи) многочисленные физиологические и молекулярные процессы либо активируются, либо ингибируются, чтобы регулировать рост и развитие растений, тем самым повышая стрессоустойчивость. Термины «дефицит воды» или «водный стресс» относятся к состоянию, при котором содержание воды в растении снижается до уровней, которые ухудшают его физиологические функции, такие как рост, устьичная проводимость и скорость фотосинтеза. Определение дефицита воды в абсолютных величинах является сложной задачей из-за сложного взаимодействия атмосферных, растительных и почвенных факторов. Кроме того, реакция растения на стресс может зависеть от условий, предшествующих самому стрессовому событию. Жак и соавторы [ 68 ] выделяют концепцию «стрессовой памяти», предполагающую, что растения обладают механизмами сохранения информации о стрессе, связанном с засухой, что потенциально приводит к повышенной устойчивости в последующие вегетационные периоды. Это явление также наблюдалось в нашем исследовании, где реакция растений на стресс засухи была менее выражена на второй год эксперимента.
Физиологические преимущества дикой Fragaria vesca обусловлены эволюционными адаптациями, обширным генетическим разнообразием и сложными метаболическими профилями, которые часто снижены у культурных форм. Хотя культурные сорта могут обладать характеристиками, благоприятными для коммерческого производства, такими как однородность и повышенная урожайность, им часто не хватает устойчивости и метаболического разнообразия, присущих диким популяциям. Этот компромисс подчёркивает важность диких видов в селекционных программах, поскольку они предоставляют генетический материал, способный повысить стрессоустойчивость и питательную ценность культурных сортов.
Хотя дикая F. vesca демонстрирует превосходную засухоустойчивость, важно признать, что культурные сорта могут обладать другими ценными признаками, такими как более высокая урожайность и улучшенное качество плодов. Это подчеркивает ценность интеграции диких генетических ресурсов в селекционные усилия для повышения устойчивости культурной клубники при сохранении желаемых агрономических характеристик. Данные свидетельствуют о том, что встречающиеся в природе растения демонстрируют значительную наследуемую генетическую изменчивость по признакам, связанным с устойчивостью к стрессу, что подразумевает, что дикая F. vesca может содержать неиспользованные генетические ресурсы, ценные для улучшения культурных сортов [ 69 ]. Баланс между засухоустойчивостью и другими агрономическими признаками остается ключевым направлением будущих исследований и разработок в области селекции Fragaria .
Однако наше исследование имеет ряд существенных ограничений. Во-первых, поскольку эксперименты проводились в контролируемых условиях в горшках, результаты могут быть не полностью применимы к полевым условиям, где на физиологические реакции влияют многочисленные дополнительные факторы. Несмотря на это, нашей целью было выделить и изучить один фактор, максимально минимизировав влияние других. Во-вторых, отсутствие молекулярного и протеомного анализа не позволило нам окончательно определить механизмы, лежащие в основе адаптации растений к стрессу. Такой анализ дал бы более детальное понимание и, вероятно, подтвердил бы наши выводы. В-третьих, отсутствие данных об эвапотранспирации означает, что мы не можем однозначно количественно оценить общие потери воды растениями и окружающей средой, включая как транспирацию (потерю воды через устьица), так и испарение с поверхности почвы. Тем не менее, тензиометрические измерения широко считаются надежным методом с высокой точностью, предлагающим эффективную альтернативу более трудоемкому гравиметрическому методу. Наконец, некоторые измерения могут показаться несколько непоследовательными из-за длительной продолжительности исследования и влияния неконтролируемых факторов, таких как ветер, вредители и влажность, которые не были учтены в наших анализах.
5. Выводы
- Данное исследование позволило провести сравнительный анализ физиологических реакций растений земляники лесной (Fragaria vesca) , включая два культурных сорта ('Rugia' и 'Baron von Solemacher') и популяцию лесной земляники, полученную из естественной среды обитания. Основные результаты следующие:
- Генотипы F. vesca демонстрируют различия в физиологических признаках. Дикорастущие растения демонстрируют иные физиологические особенности по сравнению с культурными сортами, включая более высокую флуоресценцию хлорофилла и большее содержание ассимиляционных пигментов.
- Дефицит воды в субстрате существенно влияет на физиологические характеристики растений F. vesca . Засуха приводит к снижению параметров газообмена, эффективности использования воды, параметров флуоресценции хлорофилла «а», относительной оводненности листьев и массы плодов. Напротив, засуха увеличивает содержание таких элементов, как K, Ca, Mg, Na, Cu, Zn и Mn, в различных органах растения.
- Генотипы F. vesca различаются по физиологическим реакциям на дефицит воды в субстрате. Растения из естественной среды обитания демонстрируют меньшую чувствительность к физиологическим реакциям на дефицит воды по сравнению с культурными сортами.
Дополнительные материалы
Следующая вспомогательная информация может быть загружена по адресу: https://www.mdpi.com/article/10.3390/agriculture15010070/s1 , Таблица S1: Результаты двухфакторного дисперсионного анализа параметров газообмена, Таблица S2: Результаты двухфакторного дисперсионного анализа параметров флуоресценции хлорофилла «а», Таблица S3: Результаты двухфакторного дисперсионного анализа содержания фотосинтетических пигментов, Таблица S4: Результаты двухфакторного дисперсионного анализа относительного содержания воды (RWC), Таблица S5: Результаты двухфакторного дисперсионного анализа общей урожайности плодов с одного растения и веса одного плода, Таблица S6: Результаты двухфакторного дисперсионного анализа сырой и сухой массы корневой системы, Таблица S7: Результаты двухфакторного дисперсионного анализа химического состава листьев, кроны и корней
Ссылки
1. Hamdan, M.; Shafar, J. Effect of water irrigation techniques on growth and yield of rock melon (Cucumis melo Linn cv. Glamour). Adv. Plant Sci. Technol. 2017, 24, 80–84. [Google Scholar]
2. Ghaderi, N.; Normohammadi, S.; Javadi, T. Morpho-physiological responses of strawberry (Fragaria × ananassa) to exogenous salicylic acid application under drought stress. J. Agric. Sci. Technol. 2015, 17, 167–178. [Google Scholar]
3. Klamkowski, K.; Treder, W.; Wójcik, K. Effects of long-term water stress on leaf gas exchange, growth and yield of three strawberry cultivars. J. Agric. Sci. Technol. 2015, 17, 167–178. [Google Scholar]
4. Naumann, G.; Alfieri, L.; Wyser, K.; Mentaschi, L.; Betts, R.A.; Carrao, H.; Spinoni, J.; Vogt, J.; Feyen, L. Global Changes in Drought Conditions Under Different Levels of Warming. Geophys. Res. Lett. 2018, 45, 3285–3296. [Google Scholar] [CrossRef]
5. Shao, H.-B.; Bu, L.-Y.; Jaleel, C.; Zhao, C.-X. Water-deficit stress-induced anatomical changes in higher plants. Comptes Rendus Biol. 2008, 331, 215–225. [Google Scholar] [CrossRef] [PubMed]
6. Doroszewski, A.; Jadczyszyn, J.; Kozyra, J.; Pudełko, R.; Stuczyński, T.; Mizak, K.; Łopatka, A.; Koza, P.; Górski, T.; Wróblewska, E. Podstawy systemu monitoringu suszy rolniczej (Fundamentals of the agricultural drought monitoring system). Water Environ. Rural Areas 2015, 2, 77–91. [Google Scholar]
7. Boczoń, A.; Wróbel, M. Wpływ suszy na pobór wody przez sosnę zwyczajną (Pinus sylvestris L.) o różnej pozycji w drzewostanie (The influence of drought on the water uptake by Scots pines (Pinus sylvestris L.) at different positions in the tree stand). For. Res. Work 2015, 76, 370–376. [Google Scholar]
8. Treder, W.; Wójcik, K.; Tryngiel-Grać, A.; Krzewińska, D.; Klamkowski, K. Rozwój nawodnień roślin sadowniczych w świetle badań ankietowych (Development of irrigation of orchard plants reflected by survey investigations). Infrastruct. Ecol. Rural Areas 2011, 5, 61–69. [Google Scholar]
9. Dietz, K.; Zörb, C.; Geilfus, C. Drought and crop yield. Plan Biol. 2021, 23, 881–893. [Google Scholar] [CrossRef]
10. Liu, F.; Savic, S.; Jensen, C.; Shahnazari, A.; Jacobsen, S.; Stikic, R.; Andersen, M. Water relations and yield of lysimeter-grown strawberries under limited irrigation. Sci. Hortic. 2007, 111, 128–132. [Google Scholar] [CrossRef]
11. Żebrowska, J.; Dyduch, M. Quantitative assessment of the morphogenetic efficiency in strawberry Fragaria × ananassa Duch. Folia Hortic. 2009, 21, 53–64. [Google Scholar] [CrossRef]
12. Statistics Poland, Agriculture Department. Crop Production in 2016; Statistical Publishing Establishment: Warsaw, Poland, 2017.
13. Statistics Poland, Agriculture Department. Production of Agricultural and Horticultural Crops in 2020; Statistical Publishing Establishment: Warsaw, Poland, 2021.
14. Alsheikh, M.; Suso, H.; Robson, M.; Battey, N.; Wetten, A. Appropriate choice of antibiotic and Agrobacterium strain improves transformation of antibiotic-sensitive Fragaria vesca and F. v. semperflorens. Plant Cell Rep. 2002, 20, 1173–1180. [Google Scholar] [CrossRef]
15. Caulet, R.-P.; Gradinariu, G.; Iurea, D.; Morariu, A. Influence of furostanol glycosides treatments on strawberry (Fragaria × ananassa Duch.) growth and photosynthetic characteristics under drought conditions. Sci. Hort. 2014, 169, 179–188. [Google Scholar] [CrossRef]
16. Klamkowski, K.; Treder, K. wpływ deficytu wody na wymianę gazową liści, wzrost i plonowanie dwóch odmian truskawki uprawianych pod osłonami (Effect of water deficit on gas exchange parameters, growth and yield of two strawberry cultivars grown under greenhouse conditions). Infrastruct. Ecol. Rural Areas 2011, 5, 105–113. [Google Scholar]
17. Sun, C.; Li, X.; Hu, Y.; Zhao, P.; Xu, T.; Sun, J.; Gao, X. Proline, sugars, and antioxidant enzymes respond to drought stress in the leaves of strawberry plants. Hortic. Sci. Technol. 2015, 33, 625–632. [Google Scholar] [CrossRef]
18. Yin, B.; Wang, Y.; Liu, P.; Hu, J.; Zhen, W. Effects of vesicular-arbuscular mycorrhiza on the protective system in strawberry leaves under drought stress. Front. Agric. China 2010, 4, 165–169. [Google Scholar] [CrossRef]
19. Ibrahim, M.H.; Nulit, R.; Sakimin, S.Z. Influence of drought stress on growth, biochemical changes and leaf gas exchange of strawberry (Fragaria × ananassa Duch.) in Indonesia. AIMS Agric. Food. 2022, 7, 37–60. [Google Scholar]
20. Jiang, L.; Song, R.; Wang, X.; Wang, J.; Wu, C. Transcriptomic and Metabolomic Analyses Provide New Insights into the Response of Strawberry (Fragaria × ananassa Duch.) to Drought Stress. Horticulturae 2024, 10, 734. [Google Scholar] [CrossRef]
21. Mikiciuk, G.; Mikiciuk, M.; Hawrot-Paw, M. Influence of superabsorbent polymers on the chemical composition of strawberry (Fragaria × ananassa Duch.) and biological activity in the soil. Folia Hortic. 2015, 27, 63–69. [Google Scholar] [CrossRef][Green Version]
22. Krishna, K.V.; Manasa, N.; Krishna, N.V. Drought Resistance in Wheat (Triticum aestivum L.). J. Adv. Biol. Biotechnol. 2024, 27, 409–415. [Google Scholar] [CrossRef]
23. Sun, F.; Chen, Q.; Chen, Q.; Jiang, M.; Qu, Y. Yield-based drought tolerance index evaluates the drought tolerance of cotton germplasm lines in the interaction of genotype-by-environment. PeerJ 2023, 11, e14367. [Google Scholar] [CrossRef] [PubMed]
24. Bartczak, M.; Lisiecka, J.; Knaflewski, M. Correlation between selected parameters of planting material and strawberry yield. Folia Hortic. 2010, 22, 9–12. [Google Scholar] [CrossRef]
25. Błaszczyk, J. Influence of harvest date and storage conditions on the content of chlorophyll pigments in pear peels. Folia Hortic. 2012, 24, 91–95. [Google Scholar] [CrossRef]
26. Ghaderi, N.; Siosemardeh, A. Response to drought stress of two strawberry cultivars (cv. Kurdistan and Selva). Hortic. Environ. Biotechnol. 2011, 52, 6–12. [Google Scholar] [CrossRef]
27. Zahedi, S.M.; Hosseini, M.S.; Fahadi Hoveizeh, N.; Kadkhodaei, S.; Vaculík, M. Physiological and biochemical responses of commercial strawberry cultivars under optimal and drought stress conditions. Plants 2023, 12, 496. [Google Scholar] [CrossRef]
28. Docimo, T.; Paesano, A.; D’Agostino, N.; D’Amelia, V.; Garramone, R.; Carputo, D.; Aversano, R. Exploring CDF gene family in wild potato under salinity stress unveils promising candidates for developing climate-resilient crops. Sci. Rep. 2024, 14, 24619. [Google Scholar] [CrossRef] [PubMed]
29. Najafipour, B.; Mirlohi, A.; Majidi, M.M.; Saeidi, G.; Abtahi, M. Wild barley genomic resources for drought adaptability and quality improvement. J. Cereal Sci. 2023, 114, 103802. [Google Scholar] [CrossRef]
30. Żurawicz, E. Truskawka i Poziomka (Strawberry and Wild Strawberry); Państwowe Wydawnictwo Rolnicze i Leśne: Warsaw, Poland, 2005. [Google Scholar]
31. Kaya, C.; Şenbayram, M.; Akram, N.A.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. Sulfur-enriched leonardite and humic acid soil amendments enhance tolerance to drought and phosphorus deficiency stress in maize (Zea mays L.). Sci. Rep. 2020, 10, 6432. [Google Scholar] [CrossRef]
32. Wang, X.; Fu, J.; Min, Z.; Zou, D.; Liu, H.; Wang, J.; Zheng, H.; Jia, L.; Yang, L.; Xin, W.; et al. Response of rice with overlapping growth stages to water stress by assimilates accumulation and transport and starch synthesis of superior and inferior grains. Int. J. Mol. Sci. 2022, 23, 11157. [Google Scholar] [CrossRef] [PubMed]
33. Frost, C.J.; Nyamdari, B.; Tsai, C.J.; Harding, S.A. The tonoplast-localized sucrose transporter in Populus (PtaSUT4) regulates whole-plant water relations, responses to water stress, and photosynthesis. PLoS ONE 2012, 7, e44467. [Google Scholar] [CrossRef]
34. Küçükyumuk, C.; Yıldız, H.; Meriç, M.K. The response of Braeburn apple to regulated deficit irrigation. J. Agric. Sci. 2020, 26, 154–163. [Google Scholar] [CrossRef]
35. Jones, H.G. Monitoring plant and soil water status: Established and novel methods revisited and their relevance to studies of drought tolerance. J. Exp. Bot. 2007, 58, 119–130. [Google Scholar] [CrossRef] [PubMed]
36. Arnon, D.I.; Allen, M.B.; Whatley, F.R. Photosynthesis by isolated chloroplasts. IV. General concept and comparison of three photochemical reactions. Biochim. Biophys. Acta 1956, 20, 449–461. [Google Scholar] [CrossRef]
37. Lichtenthaler, H.; Wellburn, A. Determinations of total carotenoids and chlorophyll a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1966, 11, 591–592. [Google Scholar] [CrossRef]
38. Hager, A.; Mayer-Berthenrath, T. Die Isolierung und quantative Bestimung der Carotenoide und Chlorophyll von Blatern, Algen und isolierten Chloroplasten mit Hilfe Dunnschichtchromatographischer Methoden. Planta 1966, 69, 198–217. [Google Scholar] [CrossRef] [PubMed]
39. Yamasaki, S.; Rebello Dillenburg, I. Measurements of leaf relative water content in Araucaria angustifolia. Rev. Bras. Fisiol. Veg. 1999, 11, 69–75. [Google Scholar]
40. Cameron, J.S.; Hartley, C.A. Gas exchange characteristics of Fragaria chiloensis genotypes. Hort. Sci. 1990, 25, 327–329. [Google Scholar] [CrossRef]
41. Gao, S.; Yan, Q.; Chen, L.; Song, Y.; Li, J.; Fu, C.; Dong, M. Effects of ploidy level and haplotype on variation of photosynthetic traits: Novel evidence from two Fragaria species. PLoS ONE 2017, 12, e0179899. [Google Scholar] [CrossRef]
42. Lapshin, V.I.; Yakovenko, V.V.; Shcheglov, S.N.; Podorojny, V.N. A methodical approach for evaluating the variability of productivity and fruit quality in the genetic collections of strawberry (Fragaria × ananassa Duch.). Vavilovskij Žurnal Genet. I Sel. 2019, 23, 675–682. [Google Scholar] [CrossRef]
43. Kiprono, D.; Thagun, C.; Kodama, Y. Light-dependent chloroplast relocation in wild strawberry (Fragaria vesca). Plant Signal. Behav. 2024, 19, 2342744. [Google Scholar] [CrossRef]
44. Na, Y.W.; Jeong, H.J.; Lee, S.Y.; Choi, H.G.; Kim, S.H.; Rho, I.R. Chlorophyll fluorescence as a diagnostic tool for abiotic stress tolerance in wild and cultivated strawberry species. Hortic. Environ. Biotechnol. 2014, 55, 280–286. [Google Scholar] [CrossRef]
45. Malambane, G.; Madumane, K.; Sewelo, L.T.; Batlang, U. Drought stress tolerance mechanisms and their potential common indicators to salinity, insights from the wild watermelon (Citrullus lanatus): A review. Front. Plant Sci. 2023, 13, 1074395. [Google Scholar] [CrossRef]
46. Skender, A.; Ajdinović, T.; Bećirspahić, D. The comparison of phenotypic characteristics of current varieties and wild species of Fragaria. Genetika 2015, 47, 45–52. [Google Scholar] [CrossRef]
47. Ulrich, D.; Olbricht, K. Diversity of metabolite patterns and sensory characters in wild and cultivated strawberries 1. J. Berry Res. 2014, 4, 11–17. [Google Scholar] [CrossRef]
48. Wei, Y.; Kong, Y.; Li, H.; Yao, A.; Han, J.; Zhang, W.; Li, X.; Han, D. Genome-Wide Characterization and Expression Profiling of the AP2/ERF Gene Family in Fragaria vesca L. Int. J. Mol. Sci. 2024, 25, 7614. [Google Scholar] [CrossRef] [PubMed]
49. Brasileiro, A.; Morgante, C.; Araujo, A.; Leal-Bertioli, S.; Silva, A.; Martins, A.; Vinson, C.; Santos, C.; Bonfim, O.; Togawa, R.; et al. Transcriptome profiling of wild Arachis from water-limited environments uncovers drought tolerance candidate genes. Plant Mol. Biol. Rep. 2015, 33, 1876–1892. [Google Scholar] [CrossRef]
50. Schulze, J.; Erhardt, A.; Stoll, P. Reduced clonal reproduction indicates low potential for establishment of hybrids between wild and cultivated strawberries (Fragaria vesca × F. × ananassa). Ecol. Res. 2013, 28, 43–52. [Google Scholar] [CrossRef]
51. Rugienius, R.; Bendokas, V.; Siksnianas, T.; Stanys, V.; Sasnauskas, A.; Kazanaviciute, V. Characteristics of Fragaria vesca yield parameters and anthocyanin accumulation under water deficit stress. Plants 2021, 10, 557. [Google Scholar] [CrossRef]
52. Rokosa, M.; Mikiciuk, M. Assessment of physiological and morphological traits of plants of the genus Fragaria under conditions of water deficit-a study review. Acta Sci. Pol. Hortorum Cultus 2020, 19, 21–39. [Google Scholar] [CrossRef]
53. Vinskienė, J.; Bendokas, V.; Stanys, V.; Sasnauskas, A.; Rugienius, R. The effect of osmotic stress, lighting spectrum and temperature on growth and gene expression related to anthocyanin biosynthetic pathway in wild strawberry (L.). Folia Hortic. 2023, 35, 419–431. [Google Scholar] [CrossRef]
54. Nezhadahmadi, A.; Faruq, G.; Rashid, K. The impact of drought stress on morphological and physiological parameters of three strawberry varieties in different growing conditions. Pak. J. Agric. Sci. 2015, 52, 79–92. [Google Scholar]
55. Al-Yasi, H.; Attia, H.; Alamer, K.; Hassan, F.; Ali, E.; Elshazly, S.; Siddique, K.; Hessini, K. Impact of drought on growth, photosynthesis, osmotic adjustment, and cell wall elasticity in Damask rose. Plant Physiol. Biochem. 2020, 150, 133–139. [Google Scholar] [CrossRef] [PubMed]
56. Perin, E.; Messias, R.; Galli, V.; Borowski, J.; Souza, E.; Avila, L.; Bamberg, A.; Rombaldi, C. Mineral content and antioxidant compounds in strawberry fruit submitted to drough stress. Food Sci. Technol. Int. 2019, 39, 245–254. [Google Scholar] [CrossRef]
57. Keutgen, A.J.; Pawelzik, E. Quality and nutritional value of strawberry fruit under long term salt stress. Food Chem. 2008, 107, 1413–1420. [Google Scholar] [CrossRef]
58. Buchanan, B.; Gruissem, W.; Jones, R. Photosynthesis. In Biochemistry and Molecular Biology of Plants; Wiley Blackwell: Hoboken, NJ, USA; American Society of Plants Biologists: Rockville, MD, USA, 2000; pp. 568–629. ISBN 978-0-470-71421-8. [Google Scholar]
59. Martinez, J.P.; Ledent, J.F.; Bajji, M.; Kinet, J.M.; Lutts, S. Effect of water stress on growth, Na+ and K+ accumulation and water use efficiency in relation to osmotic adjustment in two populations of Atriplex halimus L. Plant Growth Regul. 2003, 41, 63–73. [Google Scholar] [CrossRef]
60. Yurela, I. Copper in plants. Braz. J. Plant Physiol. 2005, 17, 145–156. [Google Scholar] [CrossRef]
61. Gupta, B.; Shrestha, J. Abiotic stress adaptation and tolerance mechanisms in crop plants. Front. Plant Sci. 2023, 14, 1278895. [Google Scholar] [CrossRef]
62. Munné-Bosch, S. Vitamin E function in stress sensing and signaling in plants. Dev. Cell 2019, 48, 290–292. [Google Scholar] [CrossRef] [PubMed]
63. Lata, C.; Prasad, M. Role of DREBs in regulation of abiotic stress responses in plants. J. Exp. Bot. 2011, 62, 4731–4748. [Google Scholar] [CrossRef]
64. Craine, J.M.; Nippert, J.B.; Elmore, A.J.; Skibbe, A.M.; Hutchinson, S.L.; Brunsell, N.A. Timing of climate variability and grassland productivity. Proc. Natl. Acad. Sci. USA 2012, 09, 3401–3405. [Google Scholar] [CrossRef]
65. Maqbool, A.; Bakhsh, A.; Aksoy, E. Effects of natural variations on biofortification. In Wild Germplasm for Genetic Improvement in Crop Plants; Academic Press: Cambridge, MA, USA, 2021; pp. 115–138. [Google Scholar]
66. Dawson, W.; Bòdis, J.; Bucharova, A.; Catford, J.; Duncan, R.; Fraser, L.; Groenteman, R.; Kelly, R.; Moore, J.; Pärtel, M.; et al. Root traits vary as much as leaf traits and have consistent phenotypic plasticity among 14 populations of a globally widespread herb. Funct. Ecol. 2024, 4, 926–941. [Google Scholar] [CrossRef]
67. Bowsher, A.W.; Miller, B.J.; Donovan, L.A. Evolutionary divergences in root system morphology, allocation, and nitrogen uptake in species from high-versus low-fertility soils. Funct. Plant Biol. 2015, 43, 129–140. [Google Scholar] [CrossRef] [PubMed]
68. Jacques, C.; Salon, C.; Barnard, R.L.; Vernoud, V.; Prudent, M. Drought stress memory at the plant cycle level: A review. Plants 2021, 10, 1873. [Google Scholar] [CrossRef] [PubMed]
69. Omondi, E.O.; Lin, C.Y.; Huang, S.M.; Liao, C.A.; Lin, Y.P.; Oliva, R.; van Zonneveld, M. Landscape genomics reveals genetic signals of environmental adaptation of African wild eggplants. Ecol. Evol. 2024, 14, e11662. [Google Scholar] [CrossRef]

Rokosa M, Mikiciuk M, Ptak P. Assessment of Physiological Traits of Fragaria vesca Genotypes Under Water Deficit Conditions. Agriculture. 2025; 15(1):70. https://doi.org/10.3390/agriculture15010070
Перевод статьи «Assessment of Physiological Traits of Fragaria vesca Genotypes Under Water Deficit Conditions» авторов Rokosa M, Mikiciuk M, Ptak P., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык

















































































































Комментарии (0)