Молекулярный анализ генов устойчивости к пирикуляриозу Pita2 и Pi2/Pi9 у некоторых образцов риса (Oryza sativa L.) и отобранных гибридов
Выявление основных генов устойчивости к пирикуляриозу у генотипов Oryza sativa L. является ключом к повышению и поддержанию резистентности. Создание сортов риса с длительной устойчивостью к пирикуляриозу стало важной целью в селекционных программах. В данном исследовании в течение 2020 и 2021 годов была проведена оценка десяти египетских и интродуцированных сортов.
Аннотация
Сначала был проведен полевой эксперимент в соответствии с рандомизированной полной блочной схемой с тремя повторениями, и было установлено, что египетский сорт Sakha 101 имел самую высокую урожайность зерна (53,27 г). Испанские сорта Hispagrán и Puebla оказались самыми раннеспелыми (110 и 108 дней соответственно), а также имели наибольшую массу 1000 зерен, что делает их приоритетными донорами для этих признаков; однако у них были наименьшие средние значения по количеству метелок. Во-вторых, эти сорта были подвергнуты воздействию восемнадцати изолятов пяти штаммов Pyricularia oryzae (IH, IC, ID, IE и II). Египетские сорта Giza 177, Giza 179, Sakha 106, Giza 182, GZ1368-5-5-4 и GZ6296 показали 100% устойчивость, в то время как устойчивость Hispagrán составила 16,6%, за которым следовал Sakha 101 с 27,8%.
Для углубленного изучения десяти сортов мы использовали STS-, SCAR- и CAPS-маркеры для выявления и анализа аллелей основных генов устойчивости к пирикуляриозу с широким спектром действия Pi2, Pi9 и Pita2. В контексте рассмотрения пирамидирования генов как эффективной стратегии для достижения широкой и длительной устойчивости к пирикуляриозу, молекулярный профиль также был проведен для восемнадцати индивидуальных растений F2, полученных от гибридизации сортов Giza 177 (устойчивый) × Puebla (восприимчивый). Также было получено восемнадцать индивидуальных растений F2 от скрещивания сортов Giza 177 × Hispagrán (сильно восприимчивый).
Проведение молекулярного сканирования с помощью доминантного STS-маркера YL153/YL154 на десяти сортах позволило обнаружить наличие гена Pita2, который проявлялся уникальной полосой у сорта Puebla. При сканировании 18 растений второго поколения, полученных от скрещивания Giza 177 × Puebla, у 11 индивидуальных растений из 18 была обнаружена полоса, унаследованная от Puebla. Растения F2, полученные от скрещивания Giza 177 × Puebla и амплифицированные с CAPS-маркером RG64-431/RG64-432, имели большее количество аллелей Pi2, в то время как растения F2 от скрещивания Giza 177 × Hispagrán, амплифицированные со SCAR-маркером, сцепленным с Pi9, превзошли по этому показателю своих родителей.
Наши результаты показали, что молекулярные маркеры сыграли важную роль в определении направления эволюции признаков устойчивости к пирикуляриозу.
1. Введение
Пирикуляриоз риса развил высокую скорость эволюции из-за значительной нестабильности генома, вызывая колоссальные потери урожая риса в размере 70–80%, который является основным источником питания в развивающихся странах [1]. Гриб-возбудитель Magnaporthe oryzae может инфицировать все части растения риса, включая листья, влагалища листьев, шеей метелки, метелки, цветоножки, семена и корни [2,3]. Классическая селекция на создание сортов с длительной устойчивостью остается основным прямым, экономически эффективным и экологически безопасным методом поддержания устойчивости к пирикуляриозу. Более того, селекционеры разработали множество устойчивых к пирикуляриозу сортов, адаптированных к различным рисоводческим регионам мира. Однако длительность устойчивости по-прежнему остается проблемой для селекционеров риса [4].
Нарушения устойчивости, такие как потеря устойчивости сортом Reiho в первый год производства в Египте, приводят к серьезным эпидемиям [5]. Существует насущная необходимость в устойчивости к болезням с широким спектром действия, определяемой как устойчивость к большинству географически различных рас или изолятов одного и того же патогена [6,7,8].
Растения продуцируют гены устойчивости (R-гены), которые могут напрямую или косвенно распознавать секретируемые эффекторы патогена, чтобы инициировать второй уровень защиты, называемый иммунитетом, запускаемым эффекторами (ETI), culminрующий в гиперчувствительной гибели клеток [9,10,11]. Было идентифицировано и картировано более 100 основных R-генов [12,13]. Хромосомы 6 и 12 у риса отличаются тем, что содержат ключевые гены устойчивости к пирикуляриозу, которые являются фундаментальными в архитектуре устойчивости [14,15]. R-гены с широким спектром действия Pita и Pita2 расположены на хромосоме 12, а также как минимум восемь других генов [16,17]. Pita2 кодирует R-белки, отличные от Pita, и обеспечивает более широкий спектр устойчивости [18,19,20]. Также Pita2 показывает устойчивость ко всем вирулентным по Pita изолятам и некоторым вирулентным по Pita изолятам [21,22]. Кроме того, на хромосоме 6 установлено, что локус Pi2/Pi9 расположен близко к центромере 6 [23], и этот локус включает гены Pi2, Pigm, Piz-t и Pi9 [24]. Ген Pi9, принадлежащий к семейству NBS-LRR (nucleotide-binding site leucine-rich repeat), считается ключевым моментом в выяснении молекулярной основы устойчивости к болезням с широким спектром действия [6].
Предыдущие исследования иллюстрируют важность кластерного расположения R-генов в связи с эволюцией новых специфичностей устойчивости, выработанных под давлением популяций патогенов на протяжении эволюционной истории [23,25]. На сегодняшний день идентифицировано тринадцать аллелей гена Pi9, которые различаются по спектрам устойчивости к пирикуляриозу. Эти вариации являются основной причиной долговечности гена Pi9 против Magnaporthe oryzae [26]. На основе полимеразной цепной реакции (ПЦР) были разработаны маркеры для скрининга различных R-генов устойчивости к пирикуляриозу. ДНК-маркеры представляют собой важный инструмент для повышения точности идентификации генов устойчивости к пирикуляриозу в селекционных программах [27,28,29].
Несколько египетских коммерческих сортов риса с широкоспектральной устойчивостью использовались в качестве источников устойчивости; однако их R-гены не были охарактеризованы. Цель данного исследования заключалась в следующем: (1) оценить важные признаки у десяти генотипов, чтобы продемонстрировать их фенотипическую архитектуру и изучить генетический прогресс, дисперсию и наследуемость; (2) провести скрининг реакции устойчивости к листовому пирикуляриозу в питомниках пирикуляриоза (естественное заражение) и в теплице (18 изолятов), чтобы понять, как на пирикуляриоз риса влияют условия окружающей среды; (3) исследовать R-гены (Pi2, Pi9 и Pita2) у генотипов риса и растений F2.
2. Материалы и методы
2.1. Полевой эксперимент
Исследование проводилось на опытных полях Центра исследований и обучения рисоводству в Сахе, Кафр-эш-Шейх, в течение 3 сезонов в 2020, 2021 и 2022 годах.
Генетические материалы, использованные в данном исследовании, включали десять египетских коммерческих сортов и перспективных линий, а также два интродуцированных сорта (Таблица 1). В летние сезоны 2020 и 2021 годов эксперимент проводился в соответствии с рандомизированной полной блочной схемой (RCBD) в трех повторностях. Растения выращивались в трех рядах с расстоянием 20 см между рядами и 15 см между растениями. Эксперимент проводился в соответствии с рекомендуемой для региона сельскохозяйственной практикой. Удобрения вносились в рекомендуемых дозах и сроки: через 15 дней после пересадки из расчета 40 кг N/fed и 15 кг P₂O₅/fed. Прополка проводилась вручную.
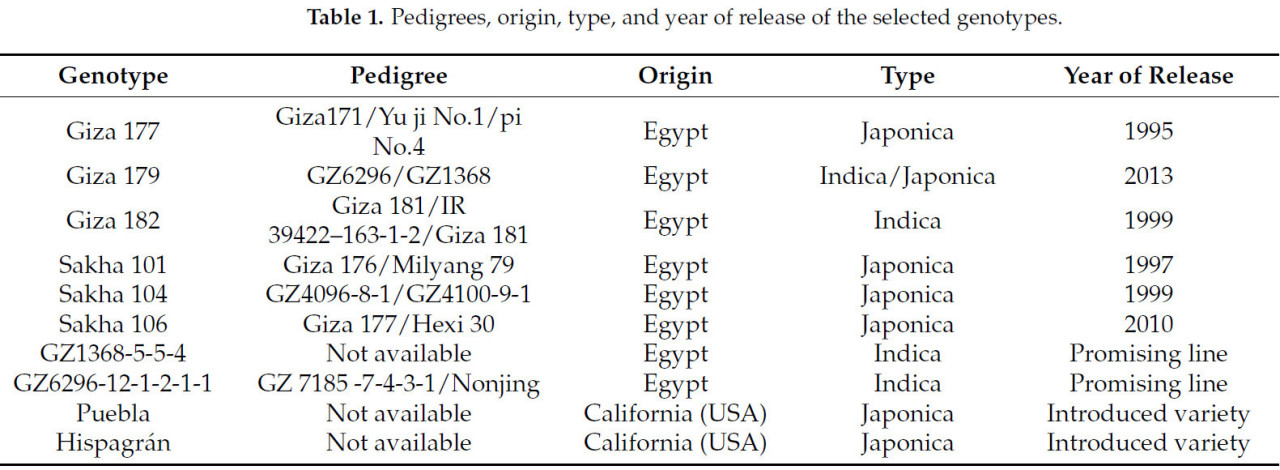
Таблица 1. Родословные, происхождение, тип и год выпуска выбранных генотипов.
2.2. Изучаемые признаки
Изучаемые признаки (количественные параметры) регистрировались для десяти сортов риса, выращенных при затоплении. Изучаемыми признаками были: число дней до созревания, количество метелок на растение, масса 1000 зерен (г) и урожай зерна с растения (г) согласно методике IRRI [30]. Фенотипический и генотипический коэффициенты изменчивости (PCV и GCV) для всех изучаемых признаков были рассчитаны по формуле, приведенной в работе Бертона [31]. Генетический прогресс в процентах от среднего (GAM) и наследуемость в широком смысле для всех признаков как отношение генетической дисперсии к общей фенотипической дисперсии были рассчитаны, как сообщалось Джонсоном и др. [32].
2.3. Анализ данных генетических параметров
Данные были проанализированы с помощью дисперсионного анализа для проверки значимости различий между изучаемыми генотипами с использованием пакета компьютерных программ "MSTAT" версии 5.4.
2.4. Заражение пирикуляриозом
В 2022 году сорта оценивались в питомниках пирикуляриоза в Сахе и Геммизе с 1 июня. Сорта высевались на грядки и удобрялись 60 кг N/феддан и навозом из расчета 8 м³/феддан. Участки грядок имели ширину 1 метр и длину 10,5 м. Восприимчивые контрольные сорта высевались после каждых 10 линий изучаемых сортов в качестве распространителя, а также высевались по всей границе питомника пирикуляриоза в качестве пограничной линии. Восприимчивые контроли, за которыми следовал один ряд устойчивых контролей, высевались попеременно, окружая изучаемые сорта. Затем они оставались подверженными естественному заражению пирикуляриозом на стадии проростков. Примерно через 40 дней после посева поражения пирикуляриозом оценивались по шкале от 0 до 9.
Заражение в теплице проводилось в том же сезоне (2022) с использованием 18 изолятов Pyricularia oryzae, полученных из сортов Сахa 101 и Сахa 104 из шести рисоводческих губернаторств. Пораженные пирикуляриозом листья собирались со старых рисовых полей. Сначала образцы листьев промывались гипохлоритом натрия, а затем дистиллированной водой перед инкубацией в чашках Петри при 32 °C для усиления споруляции. После инкубации отдельные споры отбирались под микроскопом с помощью иглы-петли и помещались на банановую среду. Рост гриба начинался в течение 7 дней после помещения спор на среду [33]. Исследования патогенности и идентификации рас проводились для полученных грибных изолятов в условиях теплицы с искусственным заражением. Международные дифференциальные сорта (I.D.V.) использовались для идентификации физиологических рас пирикуляриоза. Генотипы высевались в пластиковые лотки (30 × 20 × 15 см). Каждый лоток состоял из 10 рядов; из них 2 ряда засевались восприимчивыми контрольными сортами риса. Лотки содержались в теплице при 25–30 °C и удобрялись мочевиной (46,5% N; 5 г/лоток). Проростки на стадии 3–4 листьев (около 3–4 недель после посева) инокулировали суспензией спор (100 мл) концентрации 5 × 10⁴ спор/мл с использованием электрического распылителя. Инокулированные проростки выдерживались в увлажненной камере с влажностью воздуха не менее 90% и температурой 25–28 °C в течение 24 ч, а затем перемещались в те же условия в теплице. Каждое растение оценивалось на степень поражения пирикуляриозом риса по шкале от 0 до 9 [34].
2.5. Семена F2
Скрещивание проводилось для создания двух разных гибридов: первого скрещивания Giza 177 и Puebla, и второго скрещивания Giza 177 и Hispagrán. В период цветения применялся метод массовой кастрации согласно Джодону [35] и модифицированный Бутани [36], с использованием горячей воды (42–44 °C в течение 10 мин). Растения отбирались из F1 и продвигались до F2. Восемнадцать индивидуальных растений F2 от каждого скрещивания были использованы для экстракции ДНК, чтобы показать наследование генов устойчивости к пирикуляриозу.
2.6. Экстракция ДНК
В 2022 году геномная ДНК из десяти генотипов и восемнадцати индивидуальных растений F2 Giza 177 (устойчивый) × Puebla (восприимчивый) и восемнадцати индивидуальных растений F2 Giza 177 (устойчивый) × Hispagrán (сильно восприимчивый) была экстрагирована и проколичествована из трехнедельных незрелых молодых листьев согласно модифицированному методу CTAB, опубликованному Мюрреем и Томпсоном [37]. Экстрагированная ДНК хранилась при -20 °C. Спектрофотометрическая техника и электрофорез в агарозном геле использовались для подтверждения и количественного определения экстрагированной геномной ДНК. В этом исследовании использовались три маркера (STS, CAPS и SCAR маркеры), как показано в Таблице 2. Последовательности праймеров были загружены непосредственно с сайта Gramene (www.gramene.org). ПЦР-реакции проводились в объеме 20 мкл, содержащем 2,0 мкл ДНК (15 нг/мкл), 6,0 мкл dd.H₂O, 10,0 мкл 10 × ПЦР master max буфера, 1,0 мкл прямого праймера (30 нг/мкл) и 1,0 мкл обратного праймера (30 нг/мкл). Смесь реакционной смеси подвергалась следующим условиям: начальная денатурация при 94 °C в течение 5 мин, а затем 35 циклов амплификации со следующими параметрами: денатурация матрицы при 94 °C в течение 1 мин, отжиг праймеров при 50–62 °C в течение 24 с и элонгация при 72 °C в течение 2 мин. После завершения 35-го цикла проводилась финальная элонгация при 72 °C в течение 10 мин с последующим хранением при 4 °C. Использовались ПЦР-термоциклеры марок Biometra и Applied Biosystems. ДНК-фрагменты разделялись в 3% агарозном геле [38] и окрашивались бромистым этидием, а размер полос измерялся и анализировался с помощью программного обеспечения myImage analysis версии 1.1.753.

Таблица 2. Названия маркеров, хромосомная локализация (CL), молекулярный размер (MS), температура отжига (AT) и последовательности ДНК-праймеров, использованных в ПЦР.
3. Результаты и обсуждение
3.1. Средние значения показателей
Была проведена фенотипическая оценка (Таблица 3) десяти генотипов риса. По средним значениям дней до созревания выделялись сорта с наиболее длительным периодом от семени до семени, что, возможно, помогает уйти от болезни и может быть умной стратегией для решения проблемы нехватки воды. Таким образом, эти генотипы важны в селекционных программах. Сорта были разделены на три группы: первая группа скороспелости включает Puebla и Hispagrán, которые показали наименьшие средние значения 108 и 110 дней соответственно. Вторая группа включает дни до созревания в диапазоне от 123 до 135 дней, тогда как третья группа проводит самое длительное время до созревания — 136 и 143 дня у Sakha 104 и Sakha 101 соответственно. Драз и др. [42] сообщили, что Sakha 101 показал наибольшее среднее значение по дням до созревания (140 дней).
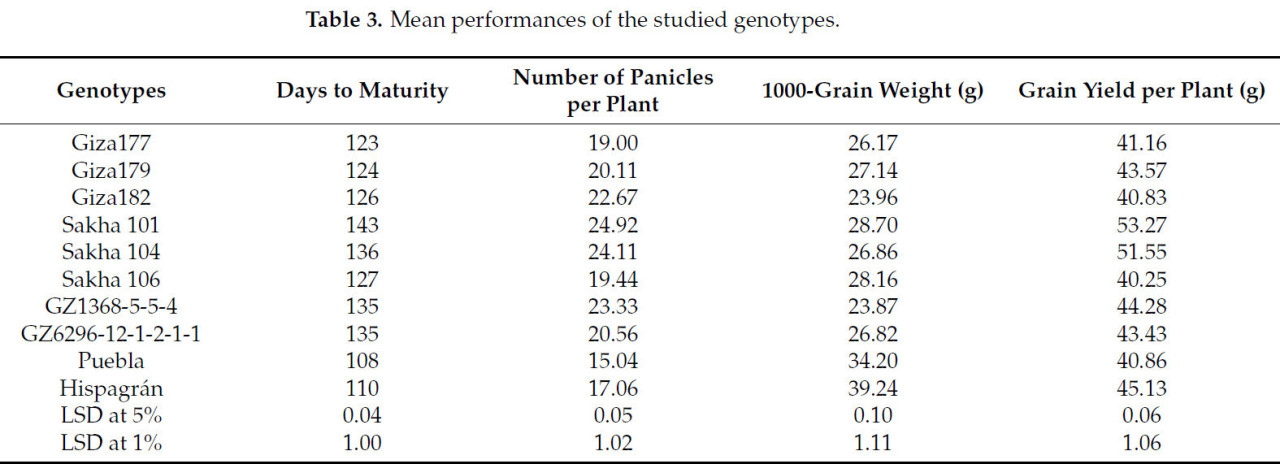
Таблица 3. Средние показатели изучаемых генотипов.
Что касается количества метелок на растение, результаты в Таблице 3 ясно показывают, что Sakha 101 зарегистрировал наивысшее среднее значение (24,92), второе наивысшее среднее значение было у Sakha 104 (24,11), за которым следуют GZ1368-5-5-4 (23,33) и затем Giza 182 с 22,67. С другой стороны, испанские сорта Puebla и Hispagrán показали наименьшее количество метелок на растение — 15,04 и 17,06 соответственно. Giza 177, Sakha 106, Giza 179 и GZ6296-12-1-2-1-1 дали средние средние значения (19,00, 19,44, 20,11 и 20,56 соответственно).
Масса 1000 зерен испанских сортов заняла самые желательные средние значения: 34,20 г и 39,24 г для Hispagrán и Puebla соответственно, тогда как египетские сорта показали средние и низкие показатели в диапазоне от 23,87 г до 28,70 г для GZ1368-5-5-4 и Sakha 101 соответственно. Урожай зерна — это конечный и самый важный параметр, и более высокий потенциал урожайности — конечная цель селекционера в развивающихся странах. Самый высокий урожай зерна с растения был у Sakha101 и Sakha 104 — 53,27 г и 51,55 г соответственно. Абд Аллах и др. [43] также сообщали, что испанские и дельта-средиземноморские генотипы в целом имели низкую урожайность зерна, более короткую продолжительность вегетационного периода и более высокую массу 1000 зерен.
3.2. Дисперсионный анализ для сортов и параметров
Фенотипическая дисперсия складывалась из генотипической дисперсии и дисперсии среды. Она оказалась высокой для дней до созревания (132,22), низкой для количества метелок на растение (13,44) и средней для массы 1000 зерен (27,49). Генетическая дисперсия, которая относится к трем основным компонентам — аддитивной, доминантной и эпистатической, — показала широкую изменчивость среди генотипов. Дни до созревания получили самую высокую генетическую изменчивость — 131,60 (Таблица 4), тогда как низкая генетическая дисперсия была обнаружена для количества метелок на растение (12,17). Существовала прямая связь между фенотипическим коэффициентом вариации (PCV%) и генотипическим коэффициентом вариации (GCV%). Наивысшие значения PCV% и GCV% были обнаружены для массы 1000 зерен, тогда как самые низкие PCV% и GCV% были обнаружены для количества дней до созревания.
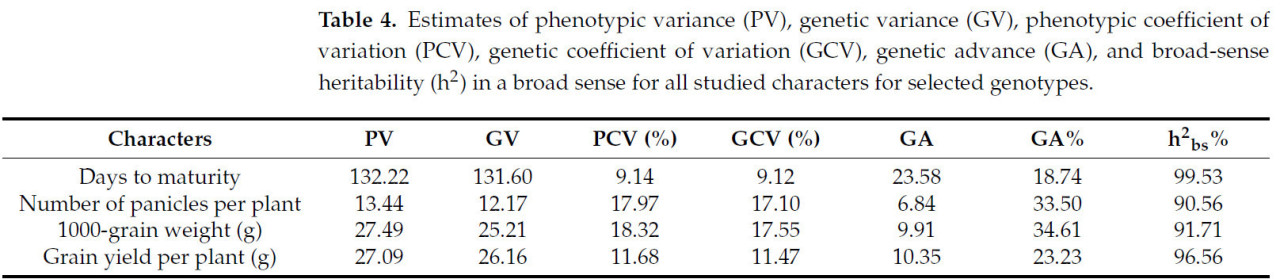
Таблица 4. Оценки фенотипической дисперсии (PV), генетической дисперсии (GV), фенотипического коэффициента вариации (PCV), генетического коэффициента вариации (GCV), генетического прогресса (GA) и наследуемости в широком смысле (h²) для всех изучаемых признаков у выбранных генотипов.
Что касается генетического прогресса (GA), надежного значимого параметра, который выражает генетический выигрыш и сочетается с наследуемостью, самые высокие значения были у дней до созревания (23,58). С другой стороны, самое низкое значение было зарегистрировано для количества метелок на растение (6,84). Между тем, процент генетического прогресса (GA%) представлял высокие значения для массы 1000 зерен (34,61%), за которым следовало количество метелок на растение (33,50%), а самый низкий процент был обнаружен для количества дней до созревания (18,74%).
Оценка наследуемости помогает селекционерам распределять ресурсы, crucial для эффективного отбора для достижения оптимизированного генетического выигрыша при ограниченных ресурсах [44]. Наследуемость в широком смысле была высокой для выбранных признаков; значения варьировались от 90,56 до 99,53%. Самый высокий процент был у дней до созревания, а самый низкий процент — у количества метелок на растение.
3.3. Скрининг пирикуляриоза при искусственном и естественном заражении
Десять генотипов были оценены в полевых условиях при естественном заражении (питомник пирикуляриоза). Источник заражения в питомнике пирикуляриоза исходил от восприимчивых сортов, высаженных в том же распространителе. Поражения листьев оценивались согласно Стандартной системе оценки IRRI (1996) с использованием шкалы 0–9 [40]. Как показано в Таблице 5, египетские генотипы Giza 177, Giza 179, Giza 182, GZ1368-5-5-4 и GZ6296-12-1-2-1-1 были оценены как устойчивые [45], Sakha 101, Sakha 104 и Puebla — как восприимчивые сорта, а Hispagrán — как сильно восприимчивый [46].
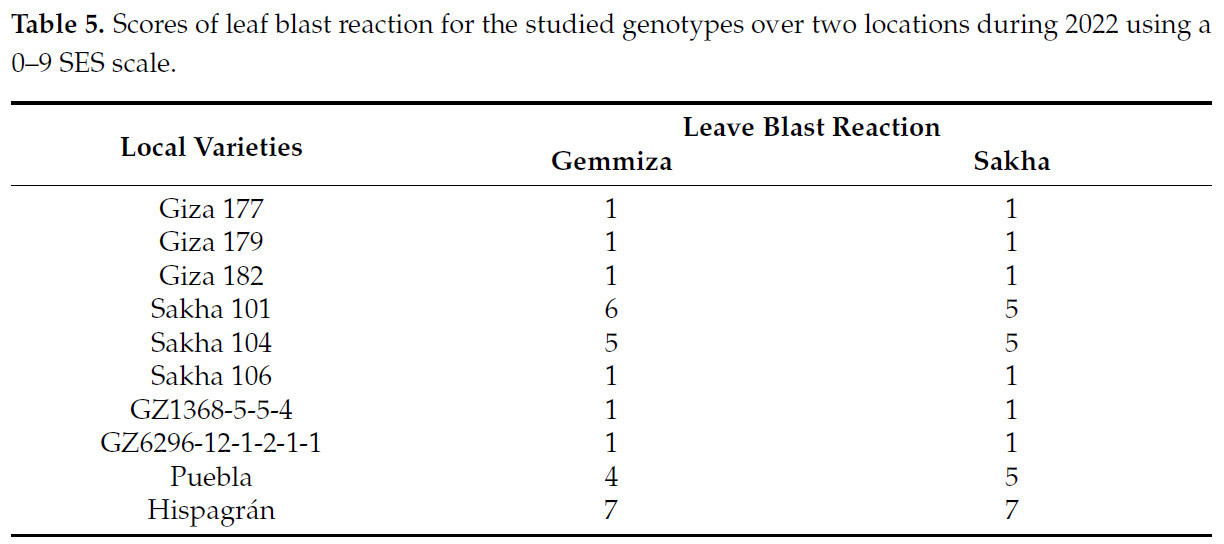
Таблица 5. Оценки реакции листьев на пирикуляриоз у изучаемых генотипов в двух локациях в течение 2022 года с использованием шкалы SES 0–9.
Восемнадцать изолятов пирикуляриоза были собраны с восприимчивых сортов Sakha 101 и Sakha 104 из шести рисоводческих губернаторств в течение сезона 2022 года. 18 изолятов были categorizedрованы в пять рас: IC, II, IH, ID и IE, как показано в Таблице 6. Процент вирулентности рас был самым высоким у ID, IC, а затем IH, в то время как IE показал низкий процент вирулентности (Рисунок 1).
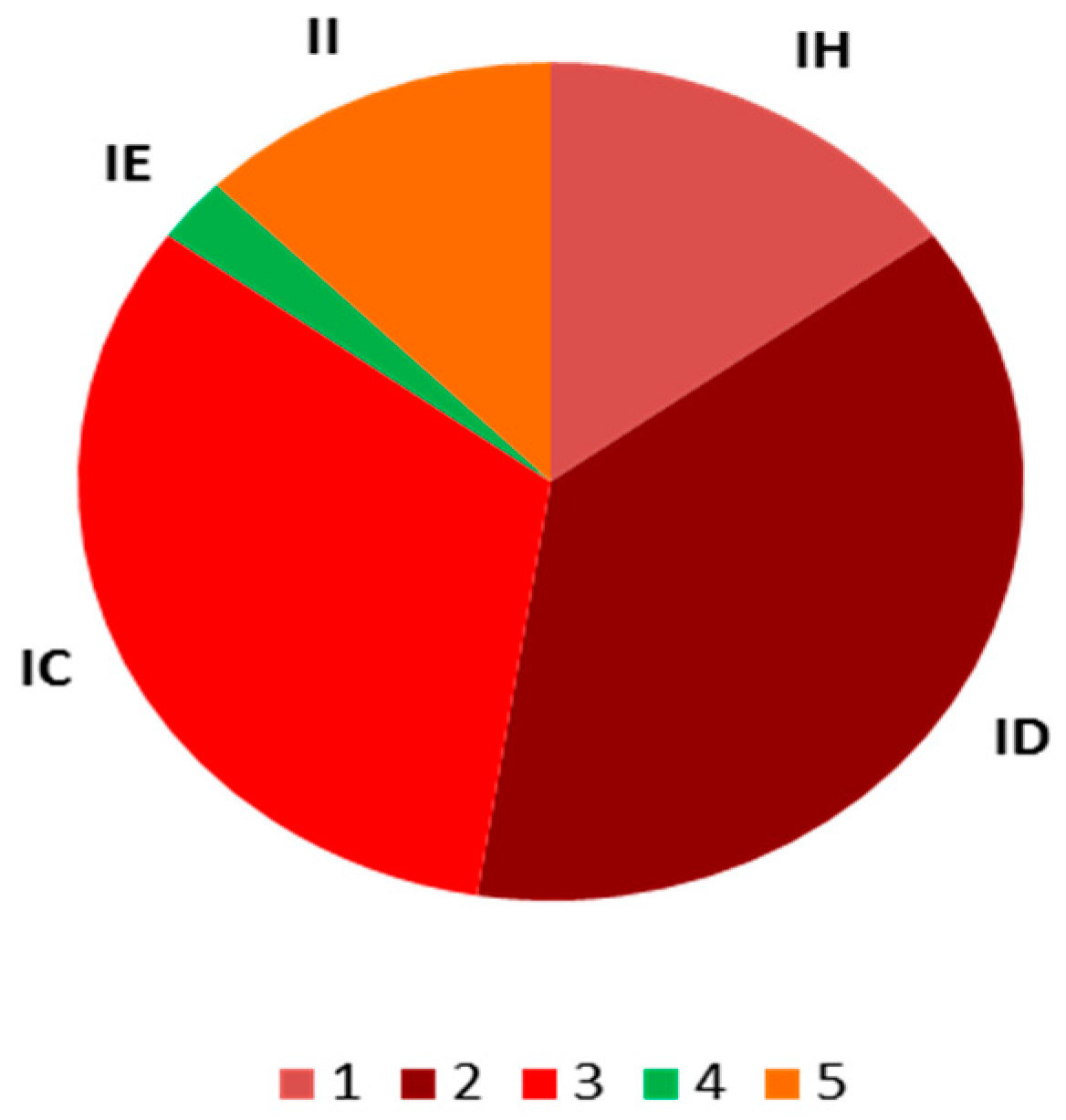
Рисунок 1. Процент вирулентности рас: (1) IH, (2) ID, (3) IC, (4) IE, (5) II.
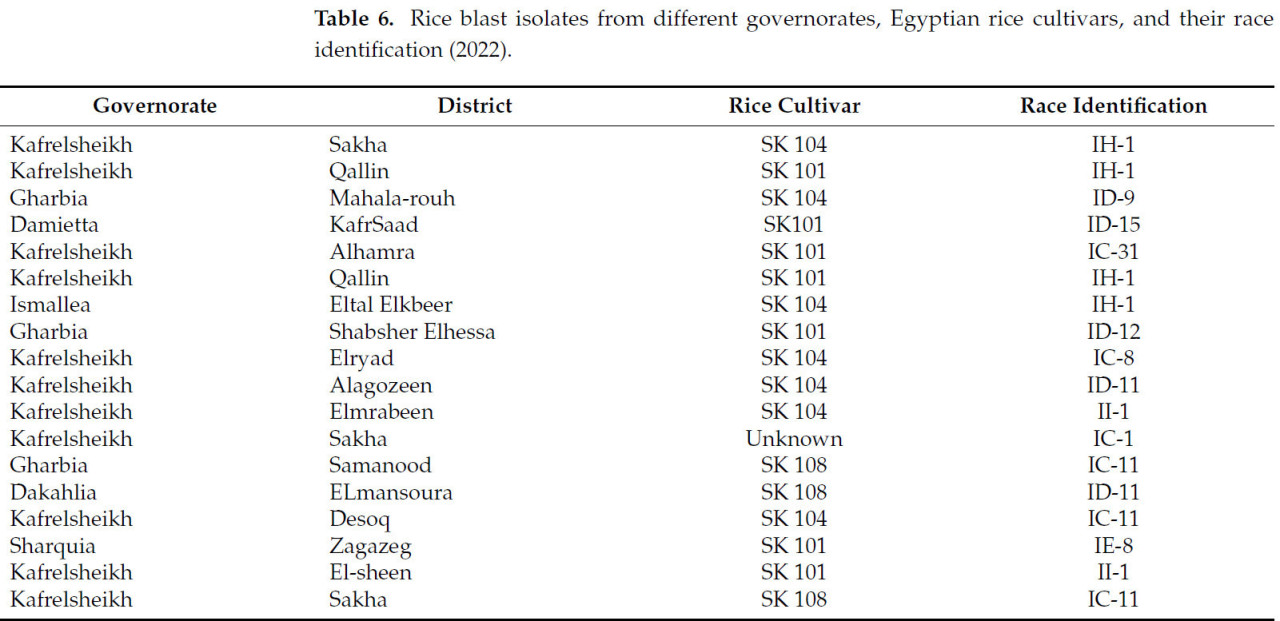
Таблица 6. Изоляты пирикуляриоза риса из разных губернаторств, египетские сорта риса и их идентификация рас (2022).
Изоляты были идентифицированы в соответствии с их реакцией на международные дифференциальные сорта на уровне рас (Таблица 7).
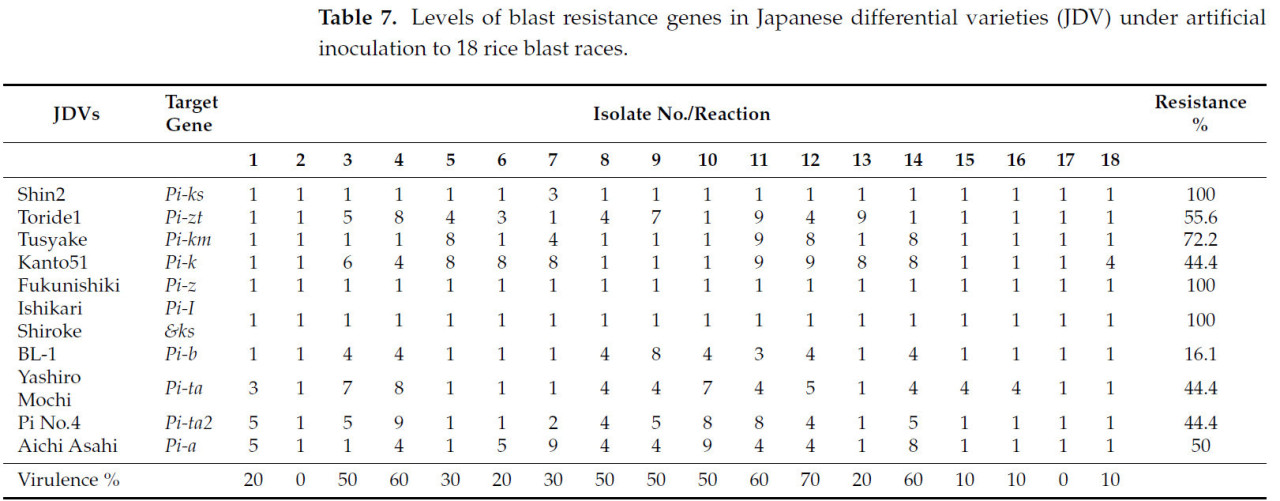
Таблица 7. Уровни генов устойчивости к пирикуляриозу у японских дифференциальных сортов (JDV) при искусственной инокуляции 18 расами пирикуляриоза.
Идентификация и мониторинг рас определяют сдвиг рас; собранные данные указали, что групповая раса IC была наиболее распространенной в 2018 и 2019 годах [47], но расы IB, IF и IG отсутствовали, в то время как Сели и др. [48] классифицировали изоляты на семь расовых групп, т.е. расы IA, IC, ID, IB, IG, IH и II.
Результаты (Таблица 8, Рисунок 2) показывают, что Hispagrán является сильно восприимчивым с процентом устойчивости 16,6%.
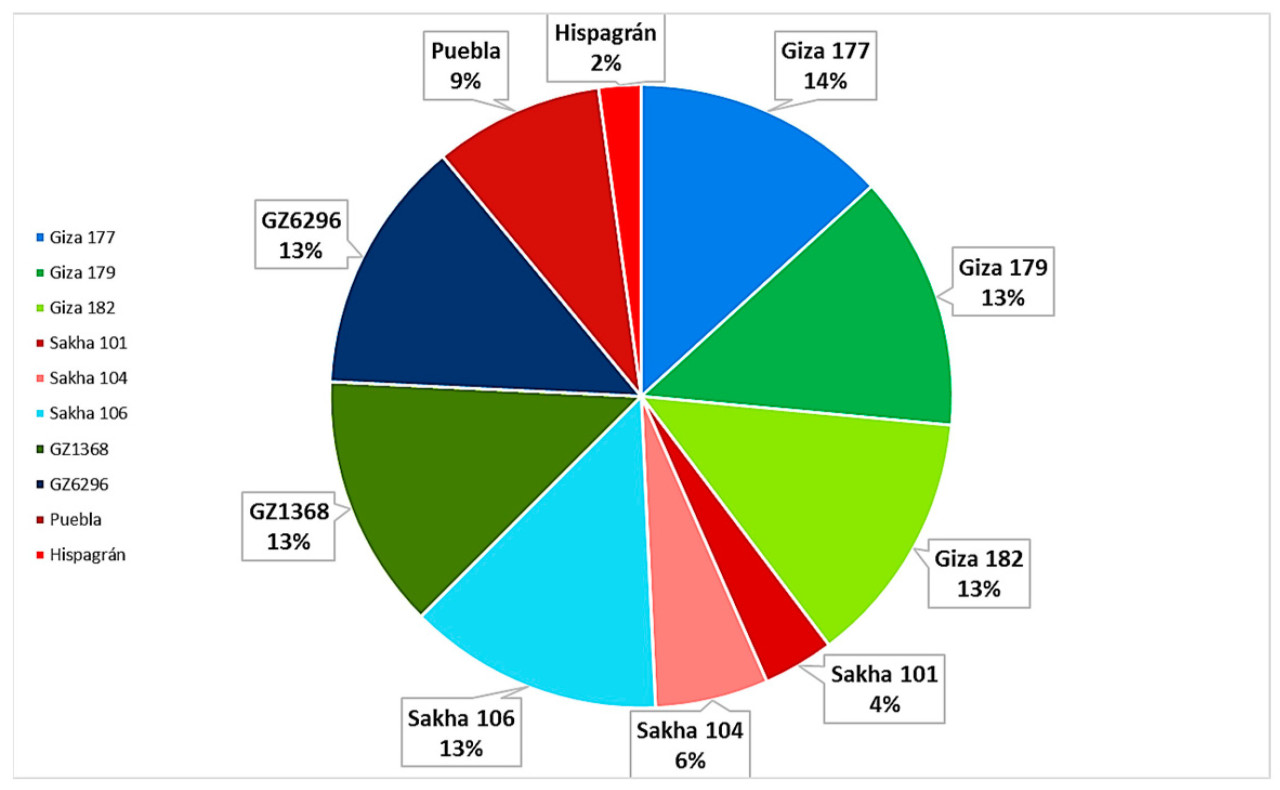
Рисунок 2. Процент устойчивости к пирикуляриозу десяти генотипов риса. (1) Giza 177, (2) Giza 179, (3) Giza 182, (4) Sakha 101, (5) Sakha 104, (6) Sakha 106, (7) GZ1368-5-5-4, (8) GZ6296-12-1-2-1-1, (9) Puebla, (10) Hispagrán.
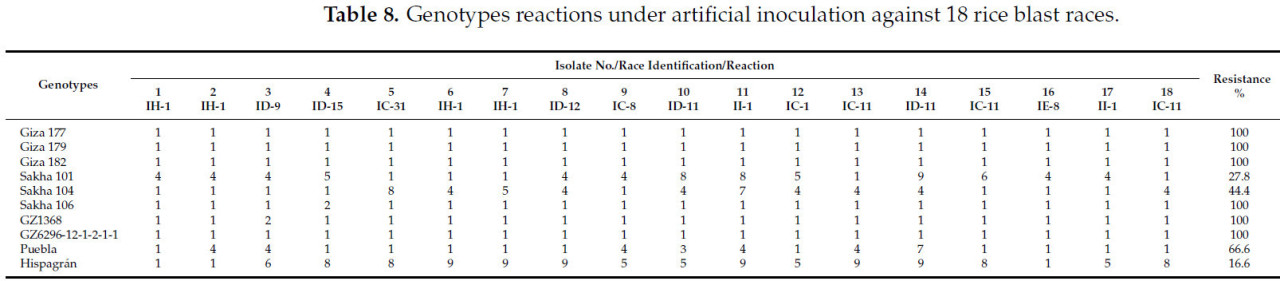
Таблица 8. Реакции генотипов при искусственной инокуляции против 18 рас пирикуляриоза риса.
Sakha 101 также был сильно восприимчивым с процентом устойчивости 27,7%. В то время как Sakha 104 был восприимчивым, его устойчивость к восемнадцати изолятам составила 44,4%, а Puebla была восприимчивой с устойчивостью 66,6%. Другие египетские сорта показали 100% устойчивость.
На Рисунке 3 показаны поражения листового пирикуляриоза, образовавшиеся в результате заражения изолятом № 11 (раса II-1) на сильно восприимчивых сортах Hispagrán и Sakha 101, восприимчивом сорте Puebla и устойчивых сортах Giza 182 и Giza 177.

Рисунок 3. Поражения листового пирикуляриоза на некоторых сортах.
3.4. Идентификация генов устойчивости к пирикуляриозу на основе маркеров
Выбранные генотипы риса и индивидуальные растения популяций F2, полученные от скрещиваний Giza 177 × Hispagrán и Giza 177 × Puebla, были подвергнуты скринингу генов устойчивости к пирикуляриозу с использованием ДНК-маркеров. Три праймера (SCAR, CAPS и STS маркеры) были использованы для идентификации генов устойчивости к пирикуляриозу, как показано на Рисунках 4-11. Распределение генов устойчивости и их ожидаемых полос представлены в Таблицах 9 и 10.

Рисунок 4. Скрининг десяти изучаемых генотипов риса с использованием доминантного STS-маркера YL155/YL87 для Pita2; (1) Giza 177, (2) Giza 179, (3) Giza 182, (4) Sakha 101, (5) Sakha 104, (6) Sakha 106, (7) GZ1368-5-5-4, (8) GZ6296-12-1-2-1-1, (9) Puebla, (10) Hispagrán, (M) маркер молекулярной массы 1000 п.н.
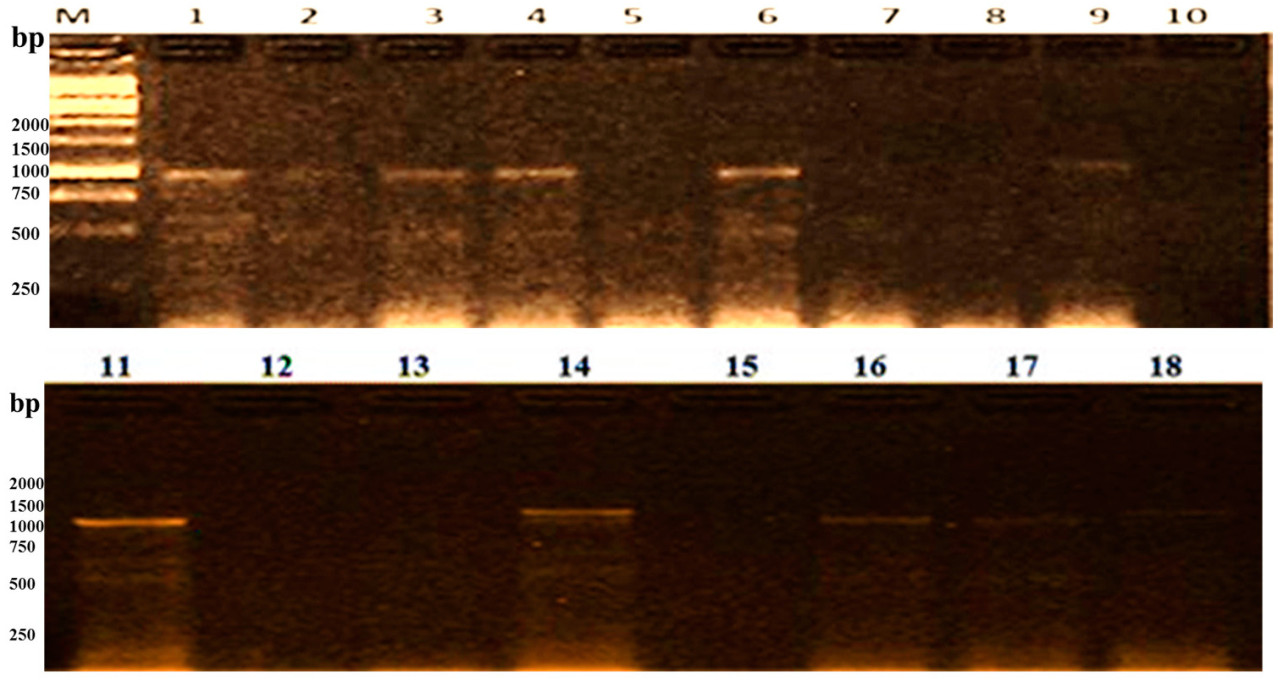
Рисунок 5. Скрининг восемнадцати индивидуальных растений F2 от скрещивания Puebla × Giza 177 с использованием доминантного STS-маркера YL155/YL87.

Рисунок 6. Скрининг десяти изучаемых генотипов риса с использованием SCAR-маркера RG64-431/RG64-432, тесно сцепленного с геном Pi2; (1) Giza 177, (2) Giza 179, (3) Giza 182, (4) Sakha 101, (5) Sakha 104, (6) Sakha 106, (7) GZ1368-5-5-4, (8) GZ6296-12-1-2-1-1, (9) Puebla, (10) Hispagrán, (M) маркер молекулярной массы 50 п.н.
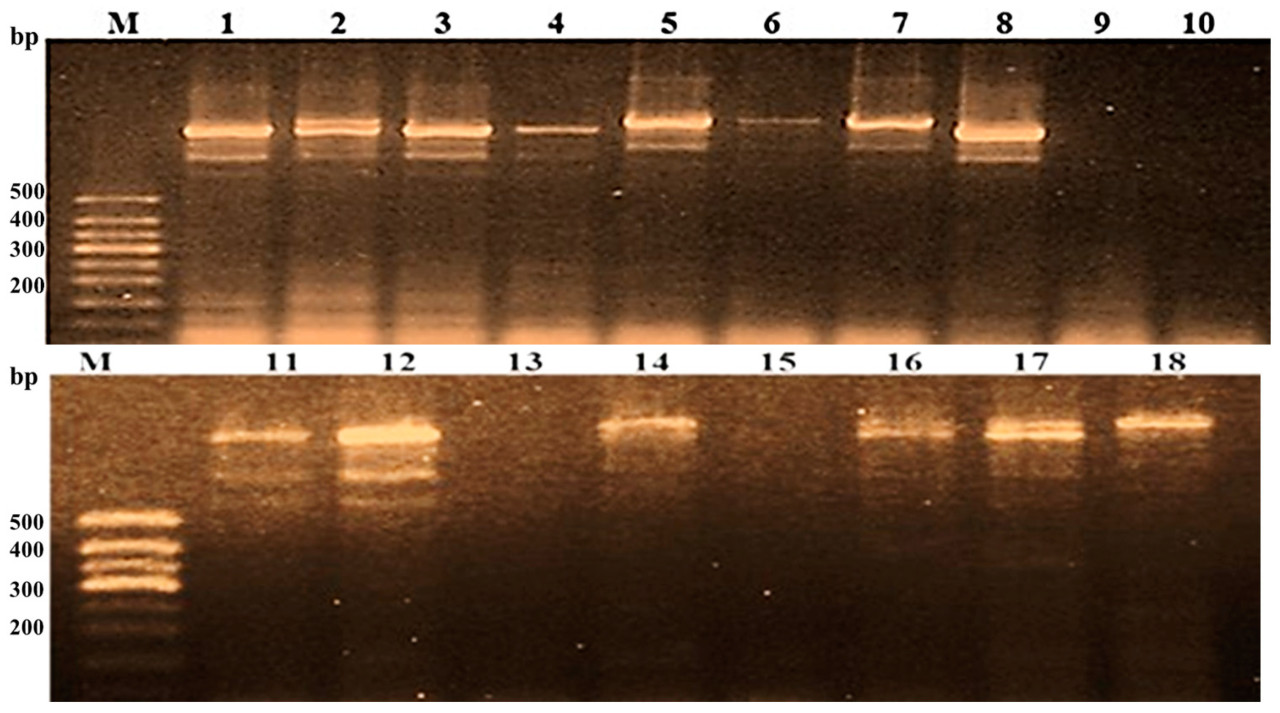
Рисунок 7. Скрининг восемнадцати индивидуальных растений F2 от скрещивания Puebla × Giza 177 с использованием CAPS-маркера RG64-431/RG64-432.
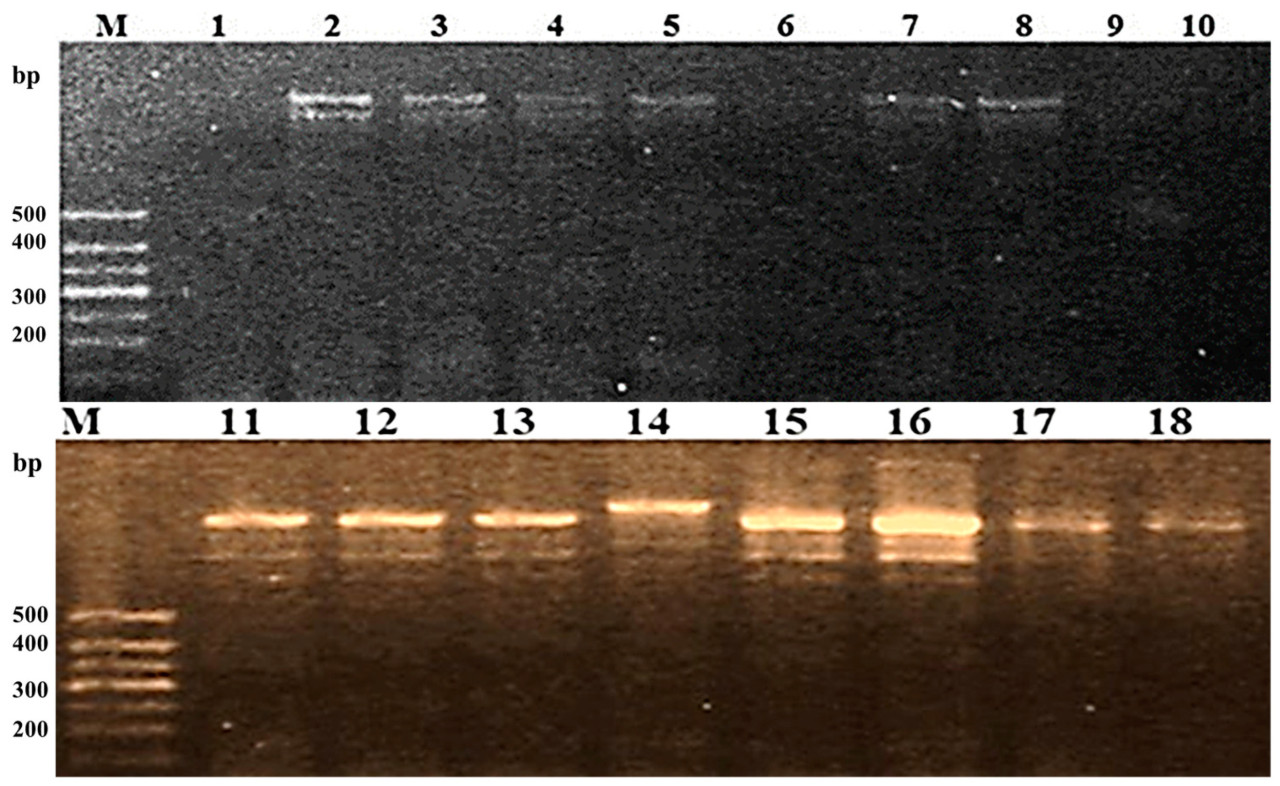
Рисунок 8. Скрининг восемнадцати индивидуальных растений F2 от скрещивания Hispagrán × Giza 177 с использованием CAPS-маркера RG64-431/RG64-432.
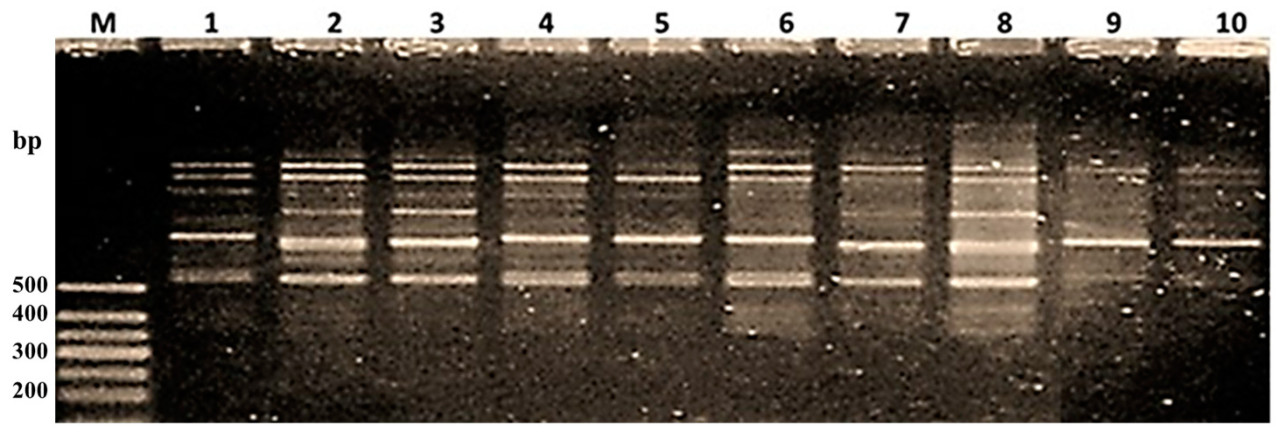
Рисунок 9. Скрининг десяти изучаемых генотипов риса с использованием SCAR-маркера PBA12; (1) Giza 177, (2) Giza 179, (3) Giza 182, (4) Sakha 101, (5) Sakha 104, (6) Sakha106, (7) GZ1368-5-5-4, (8) GZ6296-12-1-2-1-1, (9) Puebla, (10) Hispagrán, (M) маркер молекулярной массы 50 п.н.

Рисунок 10. Скрининг восемнадцати индивидуальных растений F2 от скрещивания Puebla × Giza 177 с использованием SCAR-маркера PBA12.
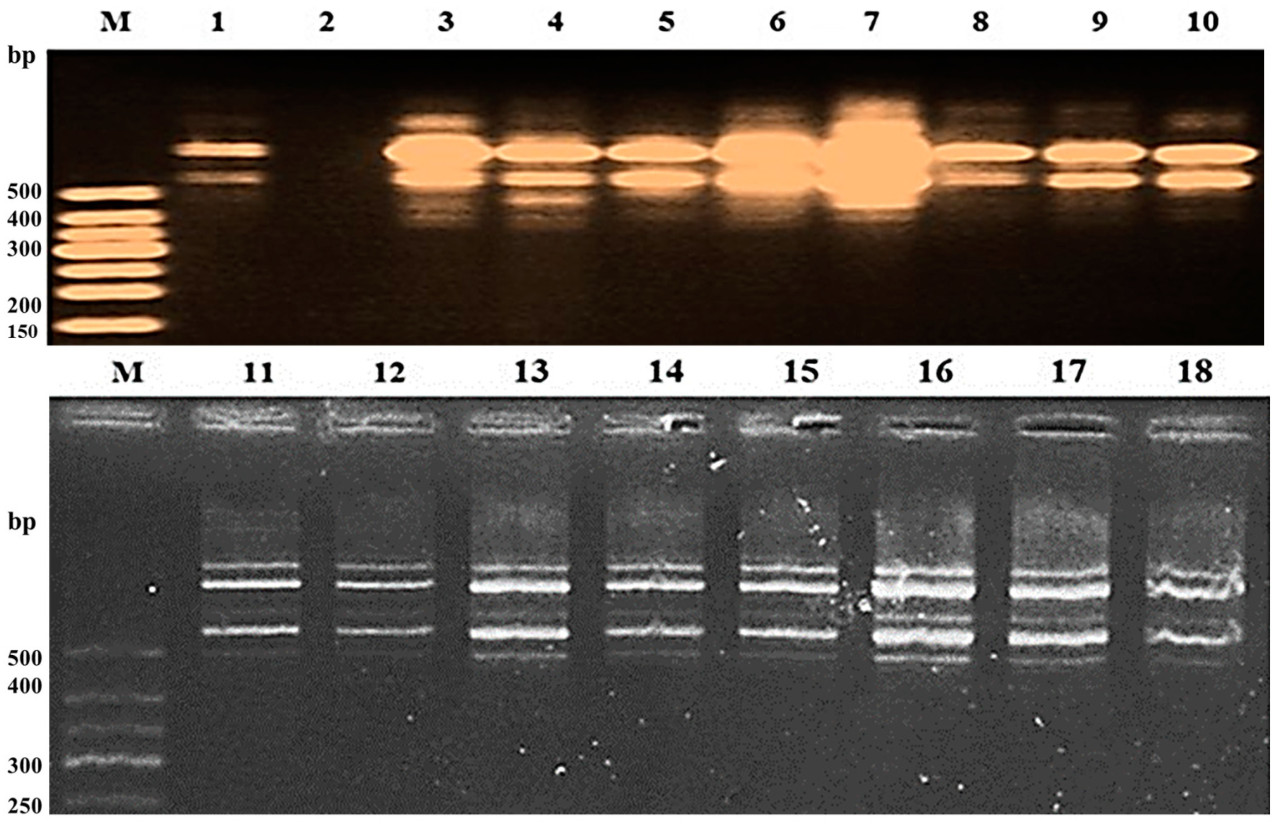
Рисунок 11. Скрининг восемнадцати индивидуальных растений F2 от скрещивания Hispagrán × Giza 177 с использованием SCAR-маркера PBA12.
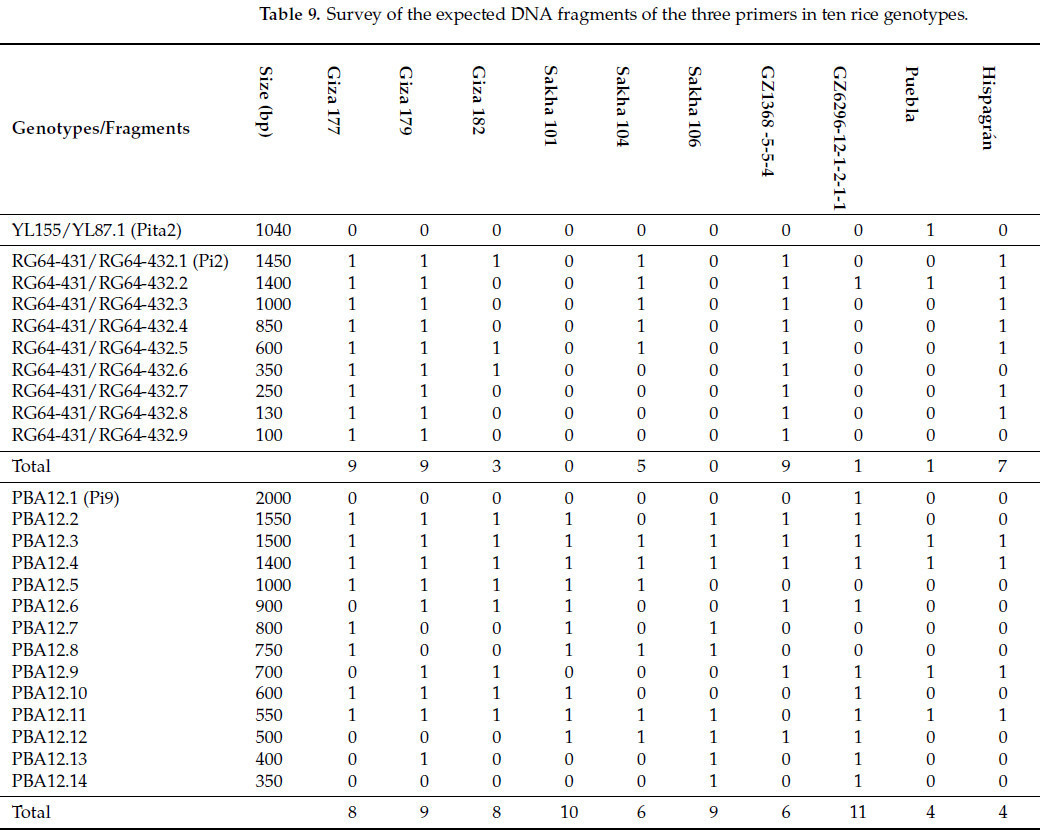
Таблица 9. Обзор ожидаемых ДНК-фрагментов трех праймеров у десяти генотипов риса.

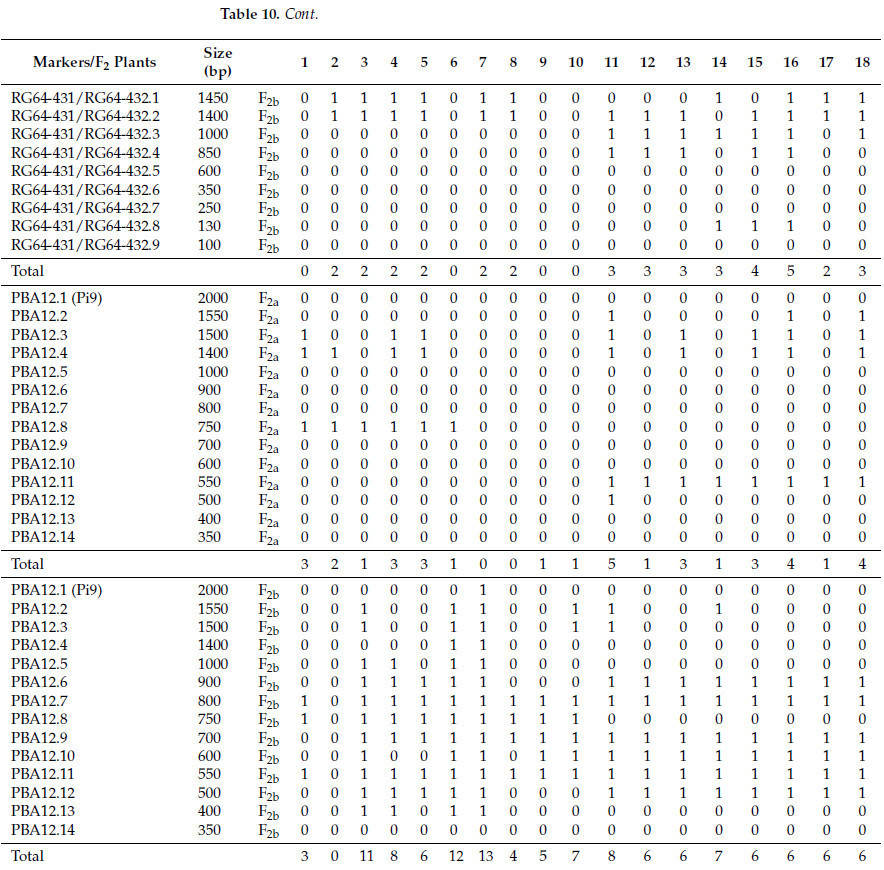
Таблица 10. ДНК-фрагменты трех праймеров в популяции F2 Giza 177 × Puebla (F2a) и Giza 177 × Hispagrán (F2b).
Доминантные молекулярные маркеры YL155 и YL187 разработаны для амплификации ожидаемых полос с молекулярной массой 1040 п.н. для гена устойчивости Pita2 [49]. Данные Рисунка 4 подтверждают наличие/отсутствие гена Pita2. Ожидаемая полоса (1040 п.н.) была полиморфной и обнаружена только у одного генотипа — у восприимчивого интродуцированного сорта Puebla (Таблица 9). Несмотря на это, ген Pita2 отсутствует у Giza 177 и не был унаследован от его родителей.
Синхронный скрининг восемнадцати растений F2 от скрещивания Puebla × Giza 177 был важной возможностью для подтверждения наличия Pita2. Доминантный аллель был унаследован одиннадцатью растениями, как показано на Рисунке 5 и в Таблице 10. Связь между Pita и Pita2 была установлена Менгом и др. [10] на Филиппинах и в Бурунди, которые пришли к выводу, что опосредованная Pita2 полевая устойчивость была относительно распространена. Более того, сообщения предполагают, что Pita2 обеспечивает более широкий спектр устойчивости, чем Pita [21,22]. В этом исследовании мы обнаружили, что при использовании японских сортов процент устойчивости этих генов был равен генам Pita и Pita2 (44%), что делает ген восприимчивым в условиях, характерных для 2022 года в Египте (Таблица 7).
CAPS-маркер (cleaved amplified polymorphic sequence) RG64-431/RG64-432 тесно сцеплен с Pi2, который расположен на хромосоме 6 [50]. Он амплифицировал девять полиморфных фрагментов в диапазоне от 1450 п.н. до 100 п.н. и ожидаемую полосу с молекулярным размером 1040 п.н. у десяти изучаемых сортов, как указано в Таблице 9. Желательный конструктивный пул гена Pi2 был обнаружен у Giza 177, Giza 179 и GZ1368-5-5-4, которые exhibited все девять полос, в то время как аллели Pi2 отсутствовали у Sakha 101 и Sakha 106. Маркер выявляет только одну полосу с тем же молекулярным размером 1400 п.н. у восприимчивого сорта Puebla и устойчивого сорта GZ6296-12-1-2-1-1. Среди генотипов Sakha 104 дала пять полос, а Hispagrán — семь полос.
Восемнадцать индивидуальных растений F2, полученных от Giza 177 × Puebla, были проскринированы на Pi2 (Таблица 10, Рисунок 7). Ожидаемая полоса с молекулярным размером 1400 п.н. была амплифицирована у двенадцати растений. Фрагмент с молекулярным размером 250 п.н. отсутствовал у всех растений. На Рисунке 7 видно, что восемь аллелей были накоплены в одном растении, а еще два растения были лишены полос Pi2. Также четыре растения несли семь полос, и два растения показали наличие только одной полосы. Было отмечено, что ни одно растение не содержало всех фрагментов, аналогичных Giza 177. С точки зрения популяционной генетики, наиболее элементарным шагом в эволюции является изменение частоты генов [51].
Анализ R-генов в популяции F2 углубил наше понимание архитектуры генов и их накопления. Восемнадцать индивидуальных растений F2 Giza 177 × Hispagrán были амплифицированы с CAPS-маркером RG64-431/RG64-432, как показано на Рисунке 8; максимальное количество амплифицированных фрагментов составило пять. Четыре растения были лишены аллеля Pi2, и только одно растение дало пять фрагментов, в то время как тринадцать растений показали ожидаемую полосу (1400 п.н.).
Растения F2 от Giza 177 × Puebla дают большее накопление и вариацию аллелей гена Pi2, чем растения F2 от Giza 177 × Hispagrán. Это делает популяцию F2, полученную от Giza 177 × Puebla, содержащей лучший шанс иметь превосходный аллель для гена Pi2.
SCAR-маркер (sequence characterized amplified region) является кодоминантным маркером, который обнаружил двенадцать полиморфных полос с молекулярными размерами от 2000 п.н. до 350 п.н. и две мономорфные полосы с молекулярными размерами 1400 п.н. и 1500 п.н. Сорта, которые показали ожидаемую полосу с молекулярным размером 400 п.н. (Таблица 9, Рисунок 9), были Giza 179, Sakha 106 и GZ6296-12-1-2-1-1. Более того, GZ6296-12-1-2-1-1 — это сорт индика, который амплифицировал наибольшее количество фрагментов (11), поэтому этот сорт считается генетическим пулом для Pi9. Он также рассматривается как важный источник этого гена в селекционных программах путем гибридизации для получения широкоспектральных устойчивых к пирикуляриозу сортов [52,53]. Эти результаты согласуются с У и др. [54], которые сообщили, что Pi9 присутствовал только в образцах типа индика.
Для Giza 177 × Puebla восемнадцать растений F2 были оценены с SCAR-маркером; паттерны полос показаны на Рисунке 10 и в Таблице 10. Накопление пяти полос в одном растении и два растения, лишенные полос, позволяют нам заключить, что все фрагменты не унаследованы от двух родителей. Большинство растений содержали одну полосу.
Растения второго поколения, полученные от Giza 177 × Hispagrán, варьировались от одного индивидуального растения, содержащего 13 фрагментов, до растения, лишенного фрагментов; эти результаты проиллюстрированы на Рисунке 11. Ожидаемый фрагмент с молекулярным размером 400 п.н. был обнаружен у четырех растений. Разница и разнообразие во фрагментах являются свидетельством расширения базового генетического пула этого гена и увеличения аллельного разнообразия внутри растений F2, полученных от скрещивания устойчивого отца (Giza 177) и сильно восприимчивого отца (Hispagrán), в то время как генетическая база была меньше для растений, полученных между устойчивым отцом и только восприимчивым отцом (Puebla). Комбинация аллелей имеет первостепенное значение для использования генетических ресурсов в улучшении сельскохозяйственных культур, а также использования различных геномов для эффективного использования с целью увеличения аллельного разнообразия гена Pi9 в плане улучшения признака устойчивости к пирикуляриозу.
Традиционная селекция путем интродукции сортов и гибридизации является эффективным способом пирамидирования генов не только для противодействия нестабильности физиологических рас пирикуляриоза, но и для увеличения способности сортов адаптироваться к изменению климата и засухе, поскольку эти условия увеличивают severity пирикуляриоза. Поскольку сорт Puebla является восприимчивым в египетских условиях, но содержит ген Pita2, который способствовал эволюции признака устойчивости к пирикуляриозу.
Основная цель ДНК-маркеров — выявить генетическую и молекулярную архитектуру, лежащую в основе фенотипов, и идентифицировать генотипы и фенотипы, лежащие в основе адаптации к различным условиям окружающей среды. Генотипическое разнообразие обычно является результатом добавления или удаления целых повторяющихся единиц или мотивов. Следовательно, разные особи проявляют вариации как различия в количестве повторов. Другими словами, полиморфизмы, наблюдаемые в маркерах, являются результатом различий в количестве повторов мотива, вызванных проскальзыванием цепи полимеразы при репликации ДНК или ошибками рекомбинации. Выявление аллелей у растений F2 является ключом к производству эволюционных сортов, поскольку признаки устойчивости к пирикуляриозу риса являются не только качественными или количественными, но и теми и другими вместе. Эти признаки называются пороговыми признаками, которые являются полигенными и выражаются в категориях и дают два типа устойчивости: частичную (количественную) и полную (качественную).
4. Выводы
Наши результаты позволяют предположить, что агрегирование всех аллелей широкоспектральных генов устойчивости к пирикуляриозу является сложной задачей. Обнаружение фрагментов, связанных с геном, с помощью молекулярных маркеров для выяснения его наличия и отсутствия у растений, является важным инструментом, помогающим в отборе растений второго поколения для экономии времени и усилий, в дополнение к выбору родительских форм в программах гибридизации. Использование кодоминантных молекулярных маркеров важно для выяснения фрагментов, связанных с наличием гена, а затем для выяснения наличия рекомбинаций для формирования нового аллеля, который может быть эффективным в устойчивости у потомства. Также, непроведение молекулярного сканирования родительских форм перед гибридизацией на изучаемый признак и подтверждение передачи признака второму поколению после фенотипического отбора считается слепым отбором и зависит от везения.
Ссылки
1. Simkhada, K.; Thapa, R. Rice Blast, A Major Threat to the Rice Production and its Various Management Techniques. Turk. J. Agric. Food Sci. Technol. 2022, 10, 147–157. [Google Scholar] [CrossRef]
2. Zhang, H.; Zheng, X.; Zhang, Z. The Magnaporthe grisea species complex and plant pathogenesis. Mol. Plant Pathol. 2016, 17, 796–804. [Google Scholar] [CrossRef] [PubMed]
3. Wilson, R.A.; Talbot, N.J. Under pressure: Investigating the biology of plant infection by Magnaporthe oryzae. Nat. Rev. Microbiol. 2009, 7, 185–195. [Google Scholar] [CrossRef]
4. Miah, G.; Rafii, M.Y.; Ismail, M.R.; Puteh, A.B.; Rahim, H.A.; Asfaliza, R.; Latif, M.A. Blast resistance in rice: A review of conventional breeding to molecular approaches. Mol. Biol. Rep. 2013, 40, 2369–2388. [Google Scholar] [CrossRef]
5. Reddy, A.P.k. Recent epidemics of rice blast in India and Egypt. Plant Dis. 1987, 71, 850. [Google Scholar] [CrossRef]
6. Qu, S.; Liu, G.; Zhou, B.; Bellizzi, M.; Zeng, L.; Dai, L.; Han, B.; Wang, G.L. The broad-spectrum blast resistance gene Pi9 encodes a nucleotide-binding site leucine-rich repeat protein and is a member of a multigene family in rice. Genetics 2006, 172, 1901–1914. [Google Scholar] [CrossRef]
7. Kou, Y.; Wang, S. Broad-spectrum and durability: Understanding of quantitative disease resistance. Curr. Opin. Plant Biol. 2010, 13, 181–185. [Google Scholar] [CrossRef]
8. Li, W.; Wang, K.; Chern, M.; Liu, Y.; Zhu, Z.; Liu, J.; Zhu, X.; Yin, J.; Ran, L.; Xiong, J.; et al. Sclerenchyma cell thickening through enhanced lignification induced by OsMYB30 prevents fungal penetration of rice leaves. New Phytol. 2020, 226, 1850–1863. [Google Scholar] [CrossRef] [PubMed]
9. Kawano, Y.; Kawano-Kaneko, T.; Shimamoto, K. Rho family GTPase-dependent immunity in plants and animals. Front. Plant Sci. 2014, 5, 522. [Google Scholar] [CrossRef] [PubMed]
10. Meng, X.; Xiao, G.; Jeanie, M.; Yanoria, T.; Siazon, P.M.; Padilla, J.; Opulencia, R.; Bigirimana, J.; Habarugira, G.; Wu, J.; et al. The broad-spectrum rice blast resistance (R) gene Pita2 encodes a novel R protein unique from Pita. Rice 2020, 13, 1–15. [Google Scholar] [CrossRef] [PubMed]
11. Lopez-Moya, F.; Suarez-Fernandez, M.; Lopez-Llorca, L.V. Molecular mechanisms of chitosan interactions with fungi and plants. Int. J. Mol. Sci. 2019, 20, 332. [Google Scholar] [CrossRef] [PubMed]
12. Yamasaki, Y.; Kiyosawa, S. Studies on inheritance of resistance of rice varieties to blast. 1. Inheritance of resistance of Japanese varieties to several strains of the fungus. Bull. Natl. Inst. Agric. Sci. 1966, 14, 39–69. [Google Scholar]
13. Sharma, T.R.; Rai, A.K.; Gupta, S.K.; Vijayan, J.; Devanna, B.N.; Ray, S. Rice blast management through host-plant resistance: Retrospect and prospects. Agric. Res. 2012, 1, 37–52. [Google Scholar] [CrossRef]
14. Liu, X.Q.; Wei, J.L.; Zhang, J.C.; Wang, C.T.; Liu, X.Q.; Zhang, X.M.; Wang, L.; Pan, Q.H. Genetic Variation of Rice Blast Resistance Genes in Oryza sativa and Its Wild Relatives. Int. J. Plant Sci. 2011, 172, 970–979. [Google Scholar] [CrossRef]
15. Ashkani, S.; Rafii, M.Y.; Shabanimofrad, M.; Ghasemzadeh, A.; Ravanfar, S.A.; Latif, M.A. Molecular progress on the mapping and cloning of functional genes for blast disease in rice (Oryza sativa L.): Current status and future considerations. Crit. Rev. Biotechnol. 2016, 36, 353–367. [Google Scholar] [CrossRef]
16. Lei, C.; Hao, K.; Yang, Y.; Ma, J.; Wang, S.; Wang, J.; Cheng, Z.; Zhao, S.; Zhang, X.; Guo, X.; et al. Identification and fine mapping of two blast resistance genes in rice cultivar 93–11. Crop J. 2013, 1, 2–14. [Google Scholar] [CrossRef]
17. Deng, Y.; Zhai, K.; Xie, Z.; Yang, D.; Zhu, X.; Liu, J.; Wang, X.; Qin, P.; Yang, Y.; Zhang, G.; et al. Epigenetic regulation of antagonistic receptors confers rice blast resistance with yield balance. Science 2017, 355, 962–965. [Google Scholar] [CrossRef] [PubMed]
18. Zhao, H.; Wang, X.; Jia, Y.; Minkenberg, B.; Wheatley, M.; Fan, J.; Jia, M.H.; Famoso, A.; Edwards, J.D.; Wamishe, Y.; et al. The rice blast resistance gene Ptr encodes an atypical protein required for broad-spectrum disease resistance. Nat. Commun. 2018, 9, 2039. [Google Scholar] [CrossRef] [PubMed]
19. Xie, Z.; Yan, B.; Shou, J.; Tang, J.; Wang, X.; Zhai, K.; Liu, J.; Li, Q.; Luo, M.; Deng, Y.; et al. A nucleotide-binding site-leucine-rich repeat receptor pair confers broad-spectrum disease resistance through physical association in rice. Philos. Trans. R. Soc. B 2019, 374, 20180308. [Google Scholar] [CrossRef] [PubMed]
20. Meng, Q.; Gupta, R.; Min, C.W.; Kim, J.; Kramer, K.; Wang, Y.; Park, S.R.; Finkemeier, I.; Kim, S.T. A proteomic insight into the MSP1 and flg22 induced signaling in Oryza sativa leaves. J. Proteom. 2019, 196, 120–130. [Google Scholar] [CrossRef]
21. Kiyosawa, S. Gene analysis of blast resistance in exotic varieties of rice (1971). JPN Agric. Res. Quart. 1971, 6, 1–8. [Google Scholar]
22. Rybka, K.; Miyamoto, M.; Ando, I.; Saito, A.; Kawasaki, S. High resolution mapping of the Indica-derived rice blast resistance genes II. Pi-ta2 and Pi-ta and a consideration of their origin. Mol. Plant-Microbe Interact. 1997, 10, 517–524. [Google Scholar] [CrossRef]
23. Dai, Y.; Jia, Y.; Correll, J.; Wang, X.; Wang, Y. Diversification and evolution of the avirulence gene AVR-Pita1 in field isolates of Magnaporthe oryzae. Fungal Genet. Biol. 2010, 47, 973–980. [Google Scholar] [CrossRef] [PubMed]
24. Xiao, N.; Wu, Y.; Pan, C.; Yu, L.; Chen, Y.; Liu, G.; Li, Y.; Zhang, X.; Wang, Z.; Dai, Z.; et al. Improving of rice blast resistances in Japonica by pyramiding major R genes. Front. Plant Sci. 2017, 7, 1918. [Google Scholar] [CrossRef]
25. Gladieux, P.; Oosterhout, C.V.; Fairhead, S.; Jouet, A.; Ortiz, D.; Ravel, S.; Shrestha, R.K.; Frouin, J.; He, X.; Zhu, Y.; et al. Extensive immune receptor repertoire diversity in disease-resistant rice landraces. Curr. Biol. 2024, 34, 3983–3995. [Google Scholar] [CrossRef]
26. Zhou, S.X.; Zhu, Y.; Wang, L.F.; Zheng, Y.P.; Chen, J.F.; Li, T.T.; Yang, X.M.; He, W.; Li, X.P.; Ma, X.C.; et al. Osa-miR1873 fine-tunes rice immunity against Magnaporthe oryzae and yield traits. J. Integr. Plant Biol. 2020, 62, 1213–1226. [Google Scholar] [CrossRef] [PubMed]
27. Hayashi, K.; Yoshida, H.; Ashikawa, I. Development of PCR-based allele-specific and InDel marker sets for nine rice blast resistance genes. Theor. Appl. Genet. 2006, 113, 251–260. [Google Scholar] [CrossRef] [PubMed]
28. Wang, Z.; Jia, Y.; Rutger, J.N.; Xia, Y. Rapid survey for presence of a blast resistance gene Pi-ta in rice cultivars using the dominant DNA markers derived from portions of the Pi-ta. Plant Breed. 2007, 126, 36–42. [Google Scholar] [CrossRef]
29. Susan, A.; Yadav, M.K.; Kar, S.; Aravindan, S.; Ngangkham, U.; Raghu, S.; Prabhukarthikeyan, S.R.; Keerthana, U.; Mukherjee, S.C.; Salam, J.L.; et al. Molecular identification of blast resistance genes in rice landraces from Northeastern India. Plant Pathol. 2019, 68, 537–554. [Google Scholar] [CrossRef]
30. IRRI. International Rice Research Descriptors for Rice; IRRI: Los Banos Laguna, Philippines, 1996; 52p. [Google Scholar]
31. Burton, G.W. Quantitative inheritance in grasses. In Proceedings of the 6th International Grassland Congress, State College, PA, USA, 17–23 August 1952; Volume 1, pp. 277–283. [Google Scholar]
32. Johnson, H.W.; Robinson, H.F.; Constock, R.E. Estimates of genetic and environmental variability in soybean. Agron. J. 1955, 47, 314–318. [Google Scholar] [CrossRef]
33. Kulmitra, A.K.; Sahu, N.; Sahu, M.K.; Kumar, R.; Kushram, T.; Kumar, V.B. Growth of Rice Blast Fungus Pyricularia oryzae (Cav.) on Different Solid and Liquid Media. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 1154–1160. [Google Scholar] [CrossRef][Green Version]
34. IRRI. Standard Evaluation System for Rice, 3rd ed.; International Rice Testing Programme: Manila, Philippines, 2013. [Google Scholar]
35. Jodon, N.E. Experiments on artificial hybridization of rice. J. Am. Soc. Agron. 1938, 30, 294–305. [Google Scholar] [CrossRef]
36. Butany, W.T. Mass emasculation in rice. Intern. Rice Com. Newsletter 1961, 9, 9–13. [Google Scholar]
37. Murray, M.G.; Thompson, W. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef] [PubMed]
38. Bassam, B.J.; Caetano-Anolles, G.; Gresshoff, P.M. Fast and sensitive silver staining of DNA in polyacrylamide gels. Anal. Biochem. 1991, 196, 80–83. [Google Scholar] [CrossRef]
39. Suh, J.P.; Roh, J.H.; Chop, Y.C.; Han, S.S.; Kim, Y.G.; Jena, K.K. The Pi40 gene for durable resistance to rice blast and molecular analysis of Pi-40 advanced backcross breeding lines. Phytopathology 2009, 99, 243–250. [Google Scholar] [CrossRef] [PubMed]
40. Hittalmani, S.; Foolad, M.R.; Mew, T.; Rodriguez, R.L.; Hung, N. Development of a PCR-based marker to identify rice blast resistance gene, pi-2(t), in a segregating population. Theor. Appl. Genet. 1995, 91, 9–14. [Google Scholar] [CrossRef]
41. Liu, G.; Lu, G.; Zeng, L.; Wang, G.L. Two broad-spectrum blast resistance genes, Pi9(t) and Pi2(t), are physicallylinked on rice chromosome 6. Mol. Genet. Genom. 2003, 267, 472–480. [Google Scholar] [CrossRef] [PubMed]
42. Draz, A.E.; Badawi, A.T.; Aidy, I.R.; El-Hissewy, A.A.; Maximos, M.A.; Bastawisi, A.O.; El-Kady, A.A.; El-Mowafi, H.F.; El-Abd, A.B.; Abd Allah, A.A.; et al. Sakha 101 a high yielding, Short grain rice variety. Egypt. J. Agric. Res. 2004, 82, 29–42. [Google Scholar]
43. Abd Allah, A.A.; Badawy, S.A.; Badawi, A.T. Strategies and Methodologies of Improving Rice Varieties in Egypt. Egypt. J. Agron. 2012, 34, 201–211. [Google Scholar] [CrossRef][Green Version]
44. Smalley, M.D.; Daub, J.L.; Hallauer, A.R. Estimation of heritability in maize by parent offspring regression. Maydica 2004, 49, 221–229. [Google Scholar]
45. Badawi, T. Rice research accomplishments in Egypt. In Activités de Recherche sur le riz en Climat Méditerranéen: Actes du Séminaire D’Arles; Chataigner, J., Ed.; CIHEAM-IAMM: Montpellier, France; 194p, (Cahiers Options Méditerranéennes; n. 24(2)). Consultation Technique du Réseau Coopératif Interrégional FAO/REU/RNE de Recherche sur le Riz en Climat Méditerranéen, 1996/09/04-07, Arles (France); Available online: http://om.ciheam.org/option.php?IDOM=736 (accessed on 23 June 2014).
46. Castejón-Muñoz, M.; Lara-Álvarez, I.; Aguilar, M. Resistance of rice cultivars to Pyricularia oryzae in Southern Spain. Span. J. Agric. Res. 2007, 5, 59–66. [Google Scholar] [CrossRef][Green Version]
47. Awadallah, M.M.; Taha, A.S.; Tahoon, A.M. Genetic Variability, Insect and Disease Resistance of some Promising Rice Genotypes. J. Plant Pathol. 2021, 12, 413–421. [Google Scholar] [CrossRef]
48. Sehly, M.R.; Osman, Z.H.; Salem, E.A. Rice diseases. In Rice in Egypt; Rice Research and Training Center, Field Crop Research Institute: Giza, Egypt, 2002; p. 301. [Google Scholar]
49. Yan, L.; Yan, B.Y.; Peng, Y.L.; Ji, Z.J.; Zeng, Y.X.; Wu, H.L.; Yang, C.D. Molecular Screening of Blast Resistance Genes in Rice Germplasms Resistant to Magnaporthe oryzae. Rice Sci. 2017, 24, 41–47. [Google Scholar] [CrossRef]
50. Yu, Z.H.; Mackill, D.J.; Bonman, J.M.; Tanksley, S.D. Tagging genes for blast resistance in rice via linkage to RFLP markers. Theor. Appl. Genet. 1991, 81, 471–476. [Google Scholar] [CrossRef]
51. Kimura, M.; Crow, J.F. The number of alleles that can be maintained in a finite population (1964). Genetics 1964, 49, 725–738. [Google Scholar] [CrossRef]
52. Wu, Y.; Yua, L.; Xiao, N.; Dia, Z.; Li, Y.; Pan, C.; Zhang, X.; Liu, G.; Li, A. Characterization and evaluation of rice blast resistance of Chinese indica hybrid rice parental lines. Crop J. 2017, 5, 509–517. [Google Scholar] [CrossRef]
53. Nizolli, V.O.; Pegoraro, C.; Oliveira, A.C. Rice blast: Strategies and challenges for improving genetic resistance. Crop Breed. Appl. Biotechnol. 2021, 21, 1–15. [Google Scholar] [CrossRef]
54. Wu, Y.; Xiao, N.; Yu, L.; Pan, C.; Li, Y.; Zhang, X.; Liu, G.; Dai, Z.; Pan, X.; Li, A. Combination Patterns of Major R Genes Determine the Level of Resistance to the M. oryzae in Rice (Oryza sativa L.). PLoS ONE 2015, 10, e0126130. [Google Scholar] [CrossRef]

Essa WM, Bassuony NN, Tahoon AE-a, Mosalam AM, Zsembeli J. Molecular Profiling for Blast Resistance Genes Pita2 and Pi2/Pi9 in Some Rice (Oryza sativa L.) Accessions and Selected Crosses. Agriculture. 2025; 15(1):61. https://doi.org/10.3390/agriculture15010061
Перевод статьи «Molecular Profiling for Blast Resistance Genes Pita2 and Pi2/Pi9 in Some Rice (Oryza sativa L.) Accessions and Selected Crosses» авторов Essa WM, Bassuony NN, Tahoon AE-a, Mosalam AM, Zsembeli J., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык

















































































































Комментарии (0)