Грибковое биоразнообразие коры, листьев, стеблей и плодов яблони под укрытиями от дождя с сокращенным графиком применения фунгицидов
Применение укрытий от дождя является перспективной агрономической практикой для защиты урожая от осадков, которая снижает потребность в фунгицидах для борьбы с некоторыми патогенами, использующими влажность листовой поверхности. Однако сочетание отсутствия дождя и сокращенного графика обработки фунгицидами может повлиять на грибковые популяции, потенциально благоприятствуя агентам биоконтроля и/или другим патогенам.
Аннотация
В данном исследовании оценивали влияние данной практики на эпифитные и эндофитные грибковые сообщества, ассоциированные с корой, листьями, цветками и плодами двух сортов яблони (Фуджи и Голден Делишес), в течение двух сезонов. Яблони выращивали в двух условиях в саду, приближенном к коммерческому: (1) под укрытиями от дождя с сокращенным графиком применения фунгицидов и (2) без укрытий по стандартной схеме интегрированной защиты растений (IPM).
Использование укрытий от дождя в сочетании со сниженным количеством обработок фунгицидами влияет на общую структуру грибкового сообщества и численность конкретных таксонов. Наибольшее воздействие было отмечено на эпифитов листьев; также различия в грибковых сообществах наблюдались между двумя сортами яблони. Использование укрытий позволило сократить применение фунгицидов в саду, но привело к увеличению численности потенциальных патогенов, таких как мучнистая роса и парша яблони, по сравнению с интегрированной защитой растений в условиях открытого грунта. Понимание реакции микробиома растения на новые подходы, способствующие сокращению использования фунгицидов, может помочь в разработке стратегий, предотвращающих накопление потенциально новых патогенов.
1. Введение
Яблоня ( Malus × domestica Borkh.) является одной из наиболее широко культивируемых плодовых культур, насчитывающей около 7500 признанных сортов по всему миру [ 1 ]. Несмотря на коммерческую значимость, большинство сортов яблони восприимчивы к различным заболеваниям, в основном вызываемым грибками и бактериями, таким как парша яблони, мучнистая роса и бактериальный ожог [ 2 ]. Среди них парша яблони, вызываемая Venturia inaequalis (Cke.) Wint., является одним из наиболее важных заболеваний, влияющих на качество плодов и приводящих к значительным экономическим потерям [ 3 ]. В частности, когда погодные условия благоприятны для заболевания, инокулят патогена высок, а сорт восприимчив; для борьбы с паршой яблони необходимы многочисленные обработки фунгицидами [ 4 ]. Однако ужесточение нормативных ограничений на использование пестицидов и растущая обеспокоенность общественности по поводу остатков химических препаратов стимулировали поиск альтернативных методов, позволяющих снизить зависимость от химических фунгицидов, обеспечивая при этом высокое качество плодов и отсутствие остатков пестицидов [ 2 ]. Использование сеток, которые действуют как физический барьер для защиты деревьев от суровых условий окружающей среды, увеличивается, поскольку они улучшают здоровье растений, повышают качество плодов и снижают потребность в средствах защиты растений [ 5 ]. Помимо защиты растений от абиотических стрессов, таких как град, дождь, заморозки, колебания температуры и солнечная радиация [ 6 ], сетки также помогают против биотических агентов, таких как птицы и насекомые. Сочетание дождевых укрытий на сетках использовалось для защиты яблонь от дождевой воды, снижая заболеваемость теми грибковыми патогенами, которым нужна влажность листьев для заражения, такими как парша яблони и послеуборочные заболевания, особенно в таких регионах, как Южный Тироль и Швейцария [ 7 ]. Аналогичным образом, исследования по использованию дождевых укрытий показали их эффективность в снижении тяжести заболеваний и повышении рентабельности для нескольких культур, таких как вишня [ 8 ], клубника [ 9 ], виноградная лоза [ 10 ] и киви [ 11 ]. На основании этих данных, укрытия от дождя можно рассматривать как перспективный инструмент для интеграции в комплексные программы борьбы с вредителями (IPM) для сокращения использования фунгицидов. Однако, несмотря на их преимущества в виде сокращения количества осадков, выпадающих на растения, и продолжительности увлажнения листьев, укрытия от дождя также могут создавать микроклимат с более высокими температурами и повышенной влажностью в ночное время. Такие изменения микроклиматических условий могут влиять на физиологию растений и взаимодействие между ними и связанными с ними микроорганизмами [ 2 , 12] .]. Кроме того, сокращение фунгицидов теоретически может благоприятствовать вторичным патогенам, которые обычно контролируются полными схемами обработки фунгицидами [ 13 ]. Сообщества микроорганизмов, ассоциированных с растениями, играют жизненно важную роль в содействии росту и здоровью растений [ 14 ], и сохранение этих сообществ было предложено в качестве возможного подхода к борьбе с болезнями [ 15 ]. Несмотря на признанную важность этих сообществ, лишь немногие исследования оценили, могут ли укрытия от дождя влиять на сообщества микроорганизмов, ассоциированных с растениями, и если да, то как. В частности, Чжан и др. [ 16 ] обнаружили, что укрытия от дождя усиливают метаболизм углеводов, аминокислот и органических соединений в грушевых деревьях и увеличивают микробное разнообразие и богатство в ризосфере. Аналогичным образом, когда укрытия от дождя использовались вместе с удобрением почвы, они оказывали значительное влияние на бактериальную структуру, способствуя разнообразию и количеству ризосферных бактерий Panax notoginseng [ 16 ]. Более того, Суй и др. [ 11 ] обнаружили, что укрытия от дождя увеличили разнообразие эпифитных бактериальных и грибковых сообществ на киви по сравнению с системами выращивания в открытом грунте. Однако Чен и др. [ 12 ] обнаружили, что укрытия от дождя не всегда могут принести пользу плодородию почвы, микробному метаболизму источника углерода, круговороту органического вещества или микробному разнообразию ризосферы грушевых деревьев, предполагая, что влияние укрытий от дождя на микробные сообщества, связанные с растениями, может варьироваться в зависимости от растения-хозяина. На виноградной лозе сообщалось о дифференцированном влиянии агентов биологического контроля и химических обработок на бактериальные и грибковые сообщества листьев, защищенных от дождя, по сравнению с незащищенными [ 17 ].
Грибковые сообщества, связанные с яблонями, различаются в зависимости от тканей растения (например, лист, цветок, плод и кора), которые представляют собой различные микроместа обитания со специфическими особенностями, в которых обитают различные сообщества [ 18 ]. Такие факторы, как возраст тканей, местоположение сада и борьба с болезнями, могут влиять на состав грибковых и бактериальных сообществ, присутствующих на коре яблони [ 19 ], которые могут выступать в качестве резервуаров для фитопатогенов, которые могут распространяться на другие ткани-хозяева через ветер или брызги дождя в течение вегетационного сезона [ 20 ]. Кроме того, Маклафлин и др. [ 21 ] наблюдали, что условия окружающей среды являются ключевыми факторами, определяющими структуру и разнообразие грибковых сообществ на плодах яблони. На сегодняшний день ни одно исследование специально не оценивало, как интеграция укрытий от дождя в программы IPM может повлиять на биоразнообразие грибковых сообществ, связанных с яблонями. Целью данного исследования было изучение влияния использования укрытий от дождя с минимальным применением фунгицидов на состав и динамику грибковых сообществ, связанных с тканями коры, листьев, цветков и плодов двух сортов яблони (Fuji и Golden Delicious) в саду, похожем на промышленное. Сравнивая эпифитные и эндофитные грибковые сообщества, укрытые от дождя с уменьшенной дозой фунгицидов (IN), и не укрытые и оставленные в условиях открытого грунта со стандартной IPM (OUT) в течение нескольких сезонов, мы стремились оценить, как использование укрытий и связанных с ними уменьшенных доз фунгицидов влияет на разнообразие грибов, потенциально патогенных грибов и общее состояние яблонь.
2. Материалы и методы
2.1 Экспериментальный проект и сбор образцов
Исследование проводилось в саду промышленного назначения, расположенном на севере Италии (46.273893 с.ш., 11.027136 в.д., высота над уровнем моря 435 м). Сад был заложен в 2017 году двухлетними клонированными яблонями двух сортов (Golden и Fuji), привитыми на подвой M9. В 2019 году половина сада была укрыта дождезащитным укрытием (Microtex, патент № 0001422628) для защиты растений от осадков и сокращения периода обработки фунгицидами против парши яблонь, в то время как другая половина осталась открытой, как обычное открытое поле. Стандартная программа IPM была применена к саду ( Таблица 1 и Таблица S1 ), и обработки применялись только при соблюдении условий для заражения вредителями и патогенами яблони, как указано в местных рекомендациях по IPM ( https://www.provincia.tn.it/Documenti-e-dati/Documenti-di-funzionamento/Disciplinare-di-produzione-integrata-edizione-2023 ; дата обращения 10 марта 2023 г.). Из-за защиты от осадков и отсутствия увлажнения листьев из-за укрытия от дождя, закрытая часть (IN) сада не получала обработки фунгицидами против парши яблони (вызываемой Venturia inaequalis ), тогда как обработки против настоящей мучнистой росы (вызываемой Podosphaera leucotricha ) обычно применялись в закрытой части (IN) так же, как и в открытой части (OUT) ( Таблица 1 и Таблица S1 ). Почасовая температура, относительная влажность воздуха, количество осадков и влажность листьев регистрировались метеорологической станцией, расположенной поблизости. Были рассчитаны минимальная, максимальная и средняя недельная температура и относительная влажность, накопленные осадки и влажность листьев ( таблица S1 ). Заболеваемость паршой и мучнистой росой яблони на побегах и плодах регистрировалась в течение обоих лет для растений сорта Голден Делишес и выражалась в процентах зараженных листьев на каждом побеге с использованием методологии, предоставленной руководящими принципами Европейской организации по защите растений (ЕОКЗР, https://pp1.eppo.int/standards/ ; дата обращения: 1 марта 2019 г.; PP1/152(4), PP1/181(4), PP1/005(3), PP1/069(3)).
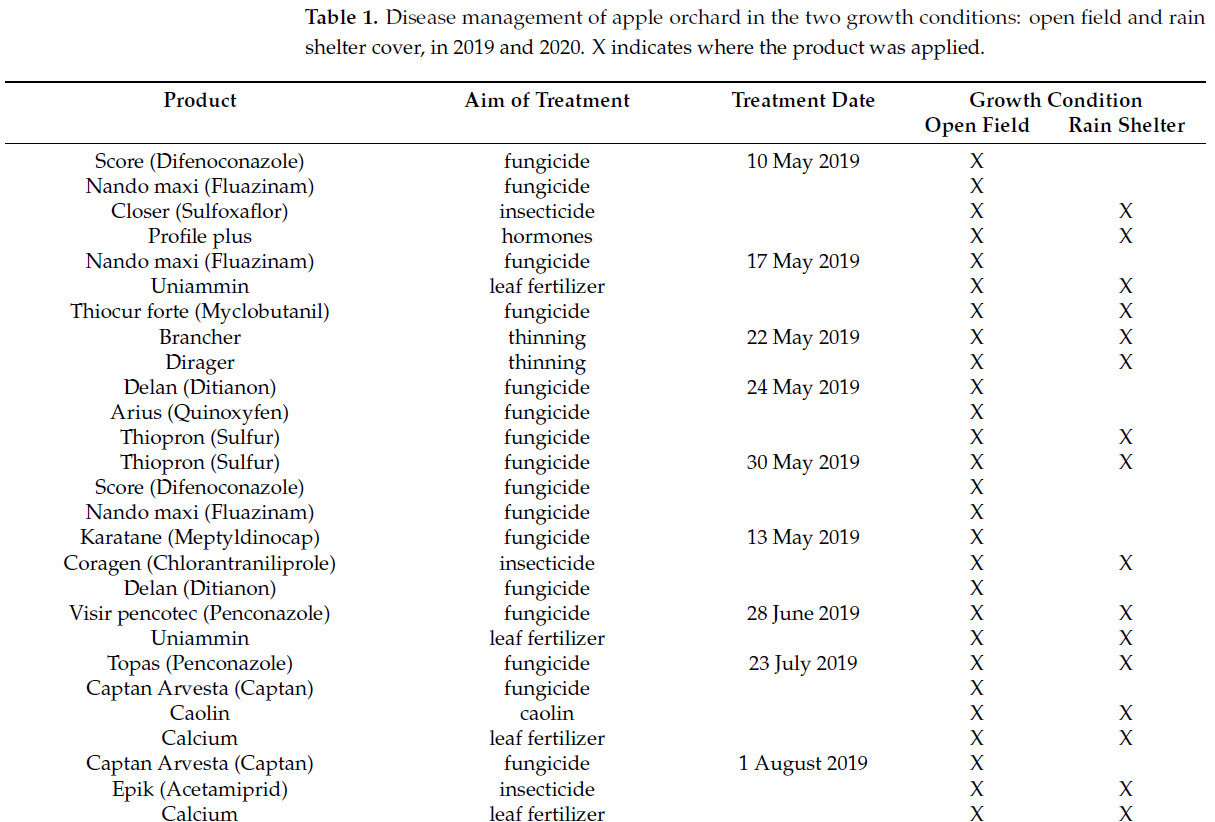
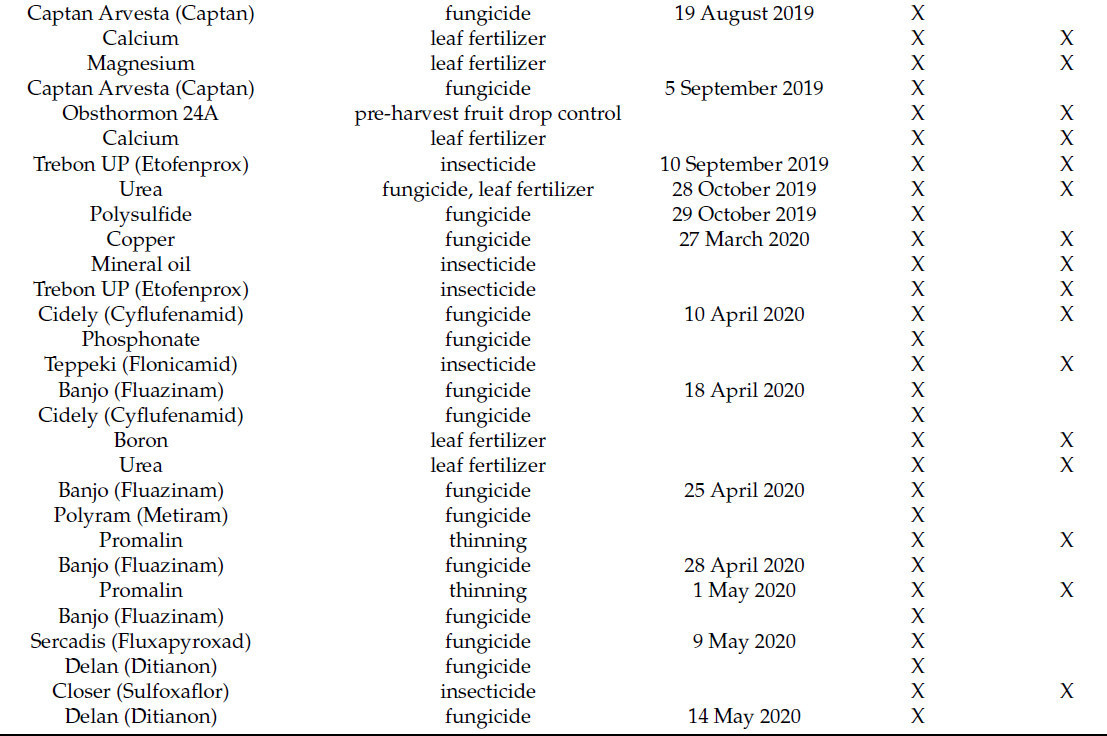
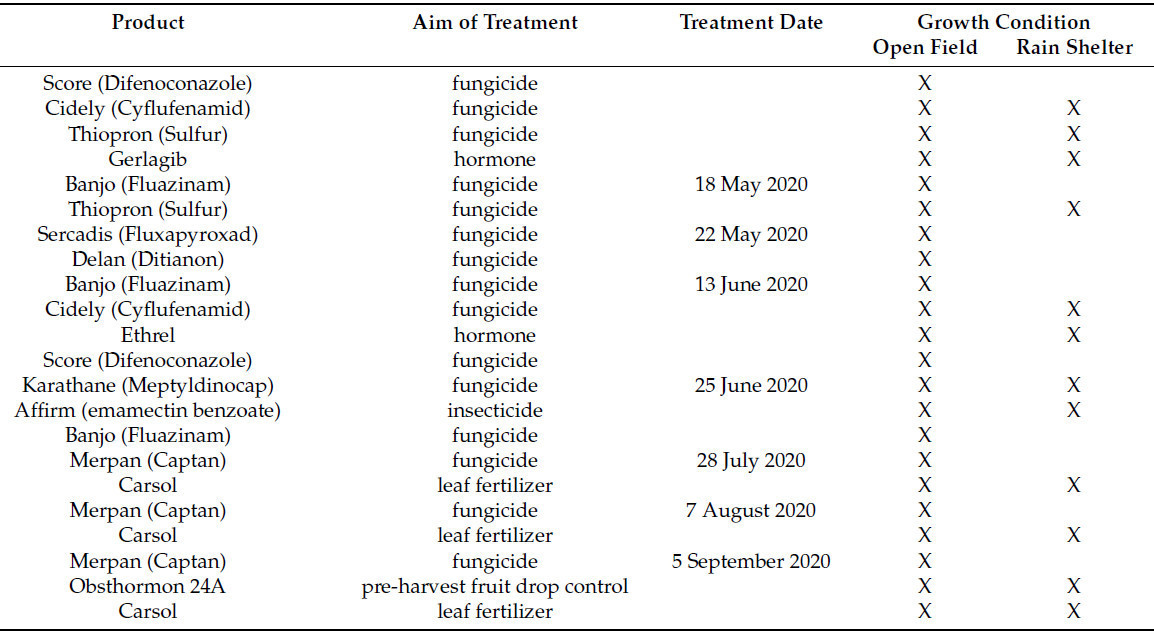
Таблица 1. Борьба с болезнями яблоневых садов в двух условиях выращивания: в открытом грунте и под укрытием от дождя в 2019 и 2020 годах. X указывает место применения препарата.
Образцы растений собирали в трех повторностях (повторности обозначены цифрами от 1 до 3) в течение двух последовательных сезонов (2019 и 2020 гг.) в трех временных точках за сезон (2 мая, T0, полное цветение; 4 июля, T1, зеленые плоды; и 17 сентября, T2, спелые плоды, в 2019 г. и 23 апреля, T3, полное цветение; 27 июля, T4, зеленые плоды; и 17 сентября, T5, спелые плоды, в 2020 г.). Каждая повторность состояла из пула из пяти случайно выбранных растений (пул растений), собранных в соответствии с планом выборки с разделенным участком в условиях OUT или IN, как описано ранее [20].
Образцы коры представляли собой пул из 30 завитков коры (0,5 г), собранных с пяти растений с помощью стерилизованного огнем скальпеля. Из того же пула растений случайным образом отбирали 50 здоровых листьев во все временные точки, 50 цветков – в моменты времени T0 и T3, а 50 плодов – в моменты времени T1, T2, T4 и T5, в зависимости от фенологической стадии, как описано ранее [ 18 ]. Образцы помещали в стерильные пластиковые пакеты и доставляли в лабораторию при температуре 10 °C в течение 1 часа.
2.2. Извлечение ДНК, амплификация и секвенирование
Метод, описанный в [ 18 ], был использован для анализа секвенирования ампликонов в сообществах грибов, связанных с растениями. Вкратце, для анализа сообществ грибов-эпифитов образцы листьев, цветов и плодов промывали в стерильном пакете, содержащем 500 мл NaCl 0,85%, дополненного 100 мкл/л Tween 80, и гомогенизировали при орбитальном встряхивании в течение 15 минут при 120 об/мин [ 18 ]. Каждую суспензию фильтровали через стерильную марлю, собирали в пробирки объемом 50 мл, центрифугировали при 10 000 × g в течение 20 минут при 4 °C. Полученные осадки смывов листьев, цветов и плодов хранили при температуре -20 °C до выделения ДНК для анализа сообществ грибов-эпифитов. Кору, промытые листья, промытые цветы и кожуру промытых плодов, полученные в стерильных условиях с использованием стерилизованного скальпеля, замораживали в жидком азоте для анализа преимущественно грибковых эндофитных сообществ. Каждый образец (0,5 г) измельчали в стерильных банках из нержавеющей стали, содержащих 2,5 мл холодного (4 °C) стерильного изотонического раствора (0,85% NaCl), с помощью миксера-мельницы-дезинтегратора (MM 400, Retsch, Германия) при 25 Гц в течение 45 с [ 18 ]. Аликвоты (500 мкл) полученных измельченных образцов коры, листьев, цветов и плодов хранили при температуре -20 °C до выделения ДНК. Геномную ДНК выделяли с использованием набора FastDNA spin для почвы (MP Biomedicals, Санта-Ана, Калифорния, США) в соответствии с инструкциями производителя. Концентрацию образцов ДНК определяли с помощью набора Qubit dsDNA Quantification Assay Kit (Invitrogn, Thermo Fisher Scientific, Уолтем, Массачусетс, США). Количество каждого образца ДНК доводили до 5,0 нг/мкл, а внутренний транскрибируемый спейсер 2 гриба (ITS2) амплифицировали с помощью метода вложенной ПЦР, который используется для ограничения амплификации ДНК хозяина в исследованиях секвенирования ампликонов эндофитов растений [ 22 ]. Вкратце, первая амплификация ITS гриба была проведена с использованием прямого праймера ITS1 (5′-CTTGGTCATTTAGAGGAAGTAA-3′) и обратного TW13 (5′-GGTCCGTGTTTCAAGACG-3′), который амплифицирует ITS гриба и часть большой субъединицы рибосомы [ 23 ]. Вторая ПЦР-амплификация была адаптирована из [ 24 ] с использованием продукта первой амплификации (3 мкл) с эквимолярными смесями прямых праймеров ITS3Mix (5′-CATCGATGAAGAACGCAG-3′, 5′-CAACGATGAAGAACGCAG-3′, 5′-CACCGATGAAGAACGCAG-3′, 5′-CATCGATGAAGAACGTAG-3′ и 5′-CATCGATGAAGAACGTGG-3′) [ 24 ] и обратных праймеров ITS4Mix (5′-TCCTCCGCTTATTGATATGC-3′ и 5′-TCCTSSSCTTATTGATATGC-3′) для увеличения покрытия царства грибов [ 25 ]. Все праймеры включали адаптеры Illumina (5′-TCGTCGGCAGCGTCAGATGTGTATAAGAGACAG-3′ и 5′-GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAG-3′ в прямом и обратном праймерах соответственно). Амплификации ITS грибов были получены с помощью системы FastStart High-Fidelity PCR (Roche), как описано ранее [ 18] с 30 циклами амплификации в первой и второй ITS-амплификации (95 °C в течение 30 с, 60 °C в течение 30 с и 72 °C в течение 30 с). Все реакции проводили в двух повторах и объединяли после амплификации.
Индексирование ДНК, количественная оценка и подготовка библиотеки для секвенирования Illumina MiSeq (PE300) были выполнены, как описано ранее [ 20 ], и были получены последовательности 360 образцов в течение двух сезонов (2019 и 2020 гг.) с тремя временными точками на сезон (T0, T1 и T2 сезона 2019 г., T3, T4 и T5 сезона 2020 г.), двумя условиями роста (образцы IN и OUT), четырьмя измельченными (измельчение коры, измельчение листьев, измельчение цветков и измельчение плодов) и тремя промытыми (мытье листьев, мытье цветков и мытье плодов) тканями, а также три репликата.
2.3 Обработка данных секвенирования ампликонов
Raw reads обрабатывали с помощью программного обеспечения MICCA (v.1.7.2) [ 26 ]. Парные конечные риды объединяли с помощью VSEARCH ( https://github.com/torognes/vsearch ; дата обращения 1 декабря 2020 г.) с минимальной длиной перекрытия 100 и максимальным количеством допустимых несовпадений 32. Праймеры обрезали с помощью Cutadapt v.1.18 [ 27 ], и объединенные риды короче 150 п.н. или с ожидаемой частотой ошибок более 0,5% удаляли. Отфильтрованные последовательности кластеризовали в Amplicon Sequence Variants (ASV) с помощью алгоритма UNOISE ( https://doi.org/10.1101/081257 ; дата обращения 1 декабря 2020 г.), доступного в MICCA. Таксономическое присвоение проводилось с использованием классификатора RDP версии 2.13 [ 28 ] на основе базы данных грибов UNITE + INSD версии 8.3 [ 29 , 30 ].
2.4 Статистический анализ
Статистический анализ данных секвенирования был выполнен с помощью пакета phyloseq R версии 1.44.0 [ 31 ] в соответствии с протоколом [ 32 ]. Считывания последовательностей были разрежены до 90% наименьших образцов, были рассчитаны индексы альфа-разнообразия, и был выполнен многомерный анализ с помощью анализа главных координат (PCoA) на основе матрицы расстояний сходства Брея-Кертиса. Тест PERMANOVA был применен с помощью пакета vegan R версии 2.6-5, а дифференциально распространенные ASV были рассчитаны с помощью пакета DESeq2 R версии 1.28.1 [ 33 ]. Графики были созданы с помощью пакета ggplot2 R версии 3.4.4 [ 34 ] и microViz версии 0.11.0 [ 35].
3. Результаты
3.1. Погодные условия и заболеваемость яблони паршой и мучнистой росой
Средняя температура в 2019 году колебалась от 1 °C до 21 °C, а в 2020 году – от 0,7 °C до 20 °C, а относительная влажность воздуха – от 50% до 90% в 2019 году и от 58,5% до 85% в 2020 году. В варианте «ВНЕ ПОЧВЫ» сумма осадков составила 1412,5 мм и 1151 мм в 2019 и 2020 годах соответственно ( таблица S1 ). В варианте «ВНУТРЬ» благодаря укрытию от дождя удалось предотвратить попадание осадков на растения, а увлажнение листьев было незначительным, поэтому применялся сокращенный режим обработки фунгицидами ( таблица 1 и таблица S1 ).
В 2019 году заболеваемость паршой и мучнистой росой яблони в условиях открытого и закрытого грунта была одинаковой (критерий Манна–Уитни, p > 0,05). Для парши яблони заболеваемость на листьях (процент пораженных листьев на каждом побеге) составила 5,15% ± 11,92% и 1,21% ± 4,53% в условиях открытого и закрытого грунта соответственно. Для мучнистой росы заболеваемость составила 2,91% ± 7,29% и 1,22% ± 3,32% в условиях открытого и закрытого грунта соответственно. Однако в 2020 году заболеваемость паршой и мучнистой росой яблони внутри укрытия от дождя была выше, чем снаружи (критерий Манна–Уитни, p ≤ 0,05). Поражённость яблонь паршой составила 4,20% ± 8,52% и 0,10% ± 1,00% в условиях IN и OUT соответственно, тогда как поражённость мучнистой росой составила 9,84% ± 23,36% и 0,48% ± 3,09% в условиях IN и OUT соответственно. Симптомов на плодах не наблюдалось в оба сезона.
3.2. Состав грибного сообщества тканей растений яблони
Секвенирование ампликона ITS ДНК, выделенной из тканей коры, цветов, листьев и плодов яблонь, дало 54 378 141 необработанных прочтений (151 050 ± 33 677 на образец), из которых 45 300 325 остались после фильтрации и шумоподавления ( таблица S2 ). Всего было обнаружено 18 624 грибковых ASV ( таблица S3 ), относящихся к семи отделам ( рисунок 1 ). В грибковых сообществах доминировали Ascomycota (49,98% ± 0,04%), за ними следовали Basidiomycota (17,92% ± 0,02%), а другие отделы имели среднюю относительную распространенность ниже 0,10%, такие как Chytridiomycota, Entomophtoromycota, Mortierellomycota, Mucoromycota и Rozellomycota.
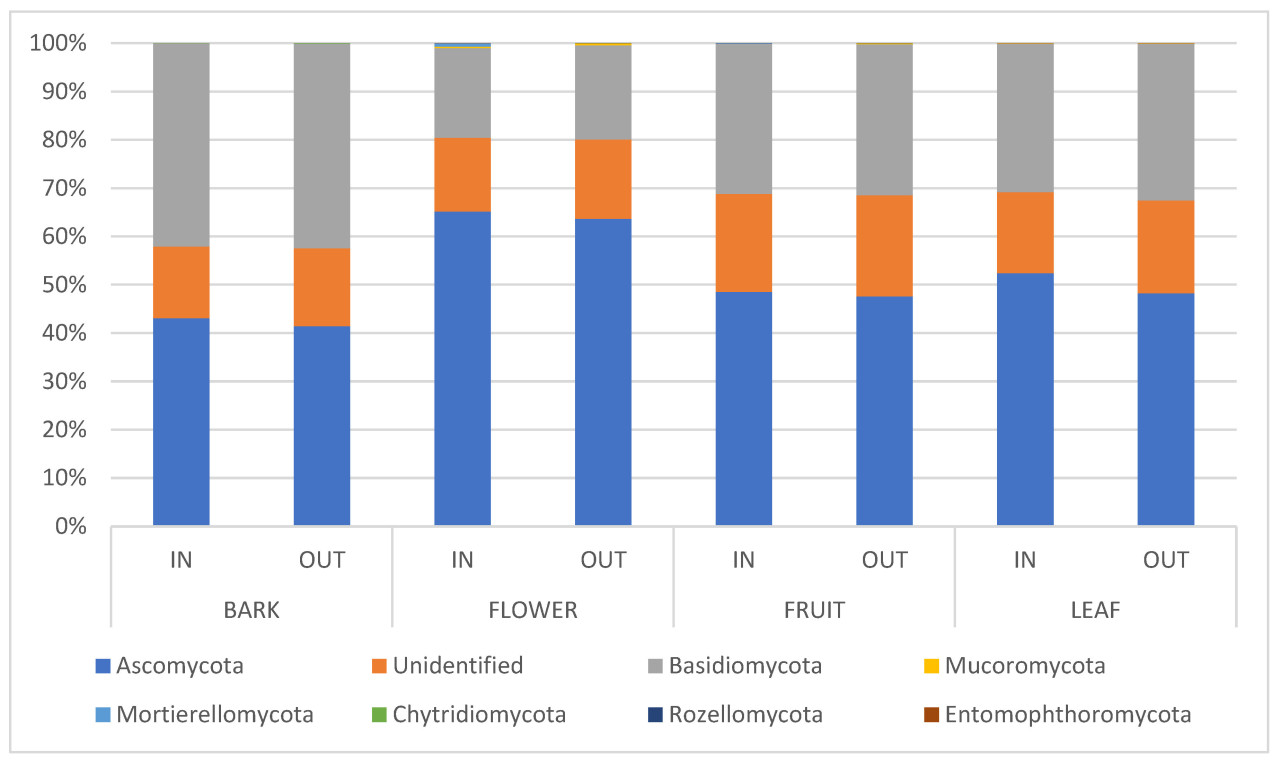
Рисунок 1. Относительное обилие грибковых сообществ на уровне филума в каждой ткани растения яблони сортов Голден и Фуджи, отобранных внутри (ВНУТРИ) и снаружи (СНАРУЖИ) укрытия от дождя. Участки ВНУТРИ и СНАРУЖИ были обработаны в соответствии с комплексными рекомендациями по борьбе с вредителями, при этом участок ВНУТРИ не обрабатывался фунгицидами против парши яблони.
Отдел аскомицетов (Ascomycota) представлен преимущественно дотидеомицетами (41,88% относительной численности), леотиомицетами (3,45%) и сордариомицетами (2,59%). Кроме того, основными классами базидиомицетов являются микроботриомицеты (17,83%), тремелломицеты (4,82%), цистобазидиомицеты (4,78%) и агарикомицеты (3,03%). Наиболее многочисленными семействами аскомицетов были Aureobasidiaceae (14,62%), Cladosporiaceae (12,11%), Didimellaceae (5,51%), Pleosporaceae (4,38%) и Erysiphaceae (2,96%), тогда как наиболее многочисленными семействами базидиомицетов были Sporidiobolaceae (17,18%), Cystobasidiaceae (1,90%), Buckleyzimaceae (1,58%) и Bulleribasidiaceae (1,42%). При рассмотрении всех образцов растений во всех условиях наиболее распространенными родами были Aureobasidium (14,58%), Cladosporium (12,11%), Alternaria (3,98%), Podosphaera (2,89%), Sporidiobolus (2,43%), Cystobasidium (1,90%), Buckleyzyma (1,58%), Rhodotorula (1,46%), Vishniacozyma (1,37%), Symmetrospora (1,21%) и Filobasidium (1,16%).
Состав наиболее распространенных семейств грибов эпифитов и эндофитов варьировался в зависимости от времени отбора проб, сорта и условий роста ( Рисунок 2 , Таблица S4 ). Среди листовых эпифитов относительное обилие Erysiphaceae (на T3 и T5) и Aureobasidiaceae (на T4) было выше в условиях IN по сравнению с условиями OUT, в то время как Cystobasidiaceae и Buckleyzymaceae было ниже в условиях IN по сравнению с условиями OUT в T3 и T4 (дифференциальный анализ обилия, основанный на отрицательном биномиальном распределении, скорректированный по частоте ложных открытий, p < 0,01; Рисунок 2 , Таблица S4 ). Что касается листовых эндофитов, семейство Pleosporaceae было более многочисленным в условиях IN (дифференциальный анализ обилия, основанный на отрицательном биномиальном распределении, скорректированный по частоте ложных открытий, p < 0,01; Рисунок 2 , Таблица S4 ). При рассмотрении эндофитов плодов основные различия были на этапе T2, когда обилие Steccherinaceae и Physalacriaceae было выше в условиях IN по сравнению с условиями OUT, тогда как Entylomataceae и Botryosphaeriaceae были более многочисленны в условиях OUT (дифференциальный анализ обилия, основанный на отрицательном биномиальном распределении, скорректированном по частоте ложных обнаружений, p < 0,01; Рисунок 2 , Таблица S4 ).
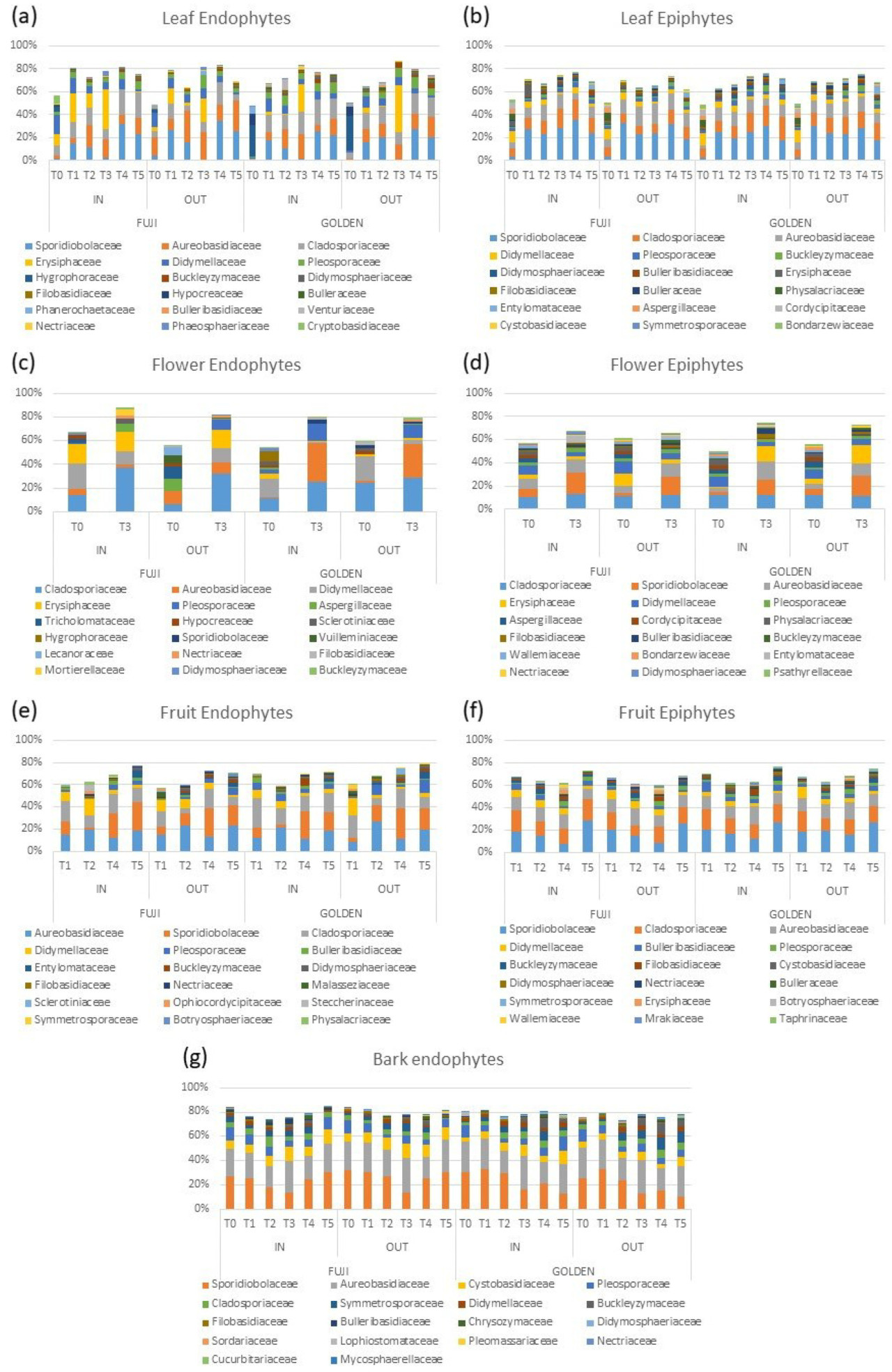
Рисунок 2. Состав наиболее распространенных семейств грибов эндофитов и эпифитов различных тканей яблони. Таксономический состав указан для каждого сорта растения (Голден или Фуджи), времени отбора проб (T0, T1, T2, T3, T4 и T5) и условий роста, например, внутри (IN) или снаружи (OUT) укрытия от дождя.
Наличие укрытия от дождя и снижение дозы фунгицида не повлияли на альфа-разнообразие (богатство и равномерность) сообществ эндофитов и эпифитов грибов в условиях «вход» в течение двух лет и в разных тканях, не показав существенной разницы по сравнению с условиями «вход» ( рисунок 3 ). Однако была отмечена существенная разница в бета-разнообразии эпифитов и эндофитов между условиями «вход» и «выход» (PERMANOVA, p ≤ 0,05; рисунок 4 и таблица S5 ).
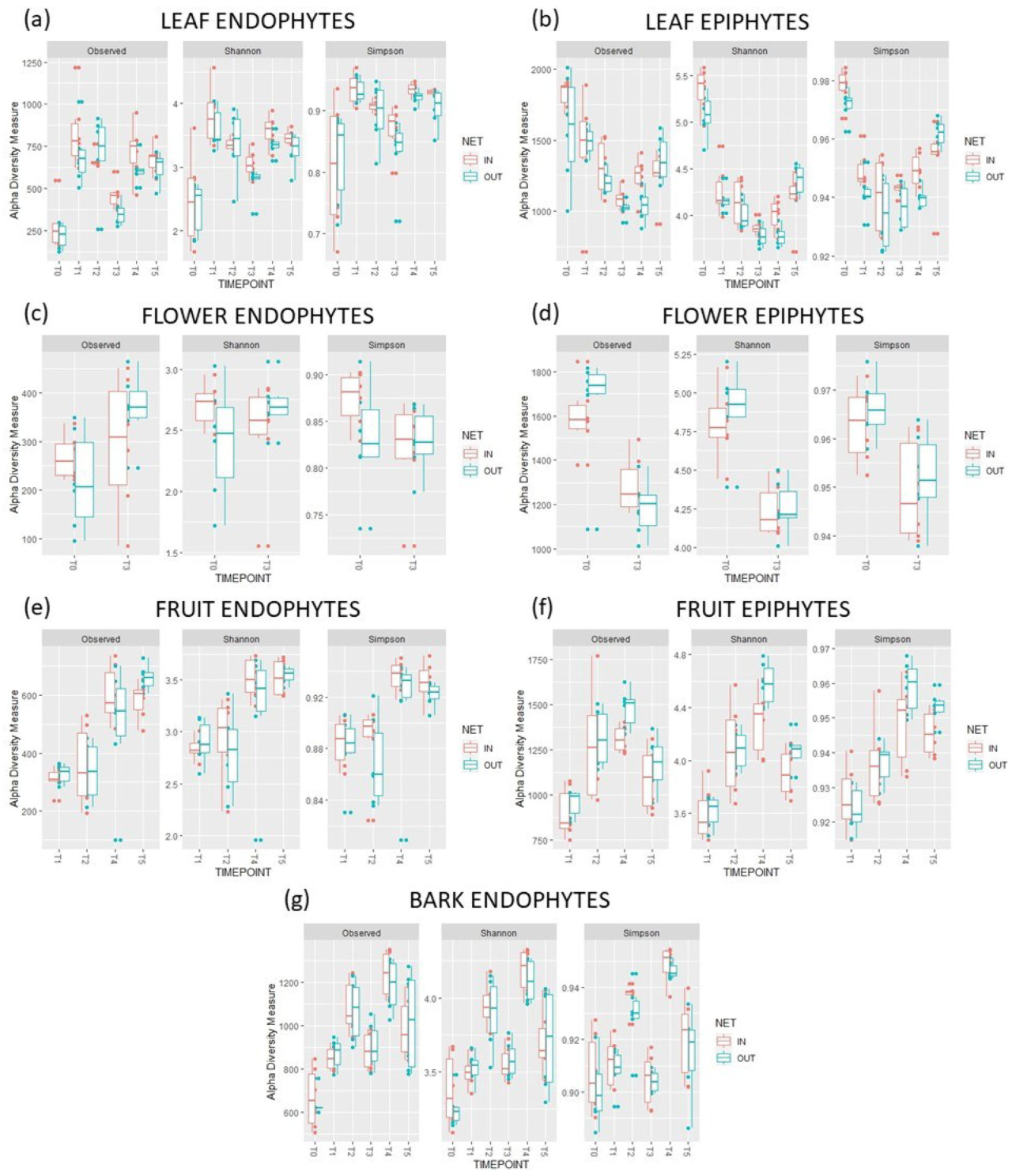
Рисунок 3. Индексы видового богатства (наблюдаемые) и разнообразия (по Шеннону и Симпсону) эндофитов и эпифитов различных тканей яблони. Состав таксонов представлен для каждого времени отбора проб (T0, T1, T2, T3, T4 и T5) и условий роста в условиях «в помещении» (укрытие от дождя + сокращенное применение фунгицидов) и «в открытом грунте» (открытый грунт + стандартная программа IPM).
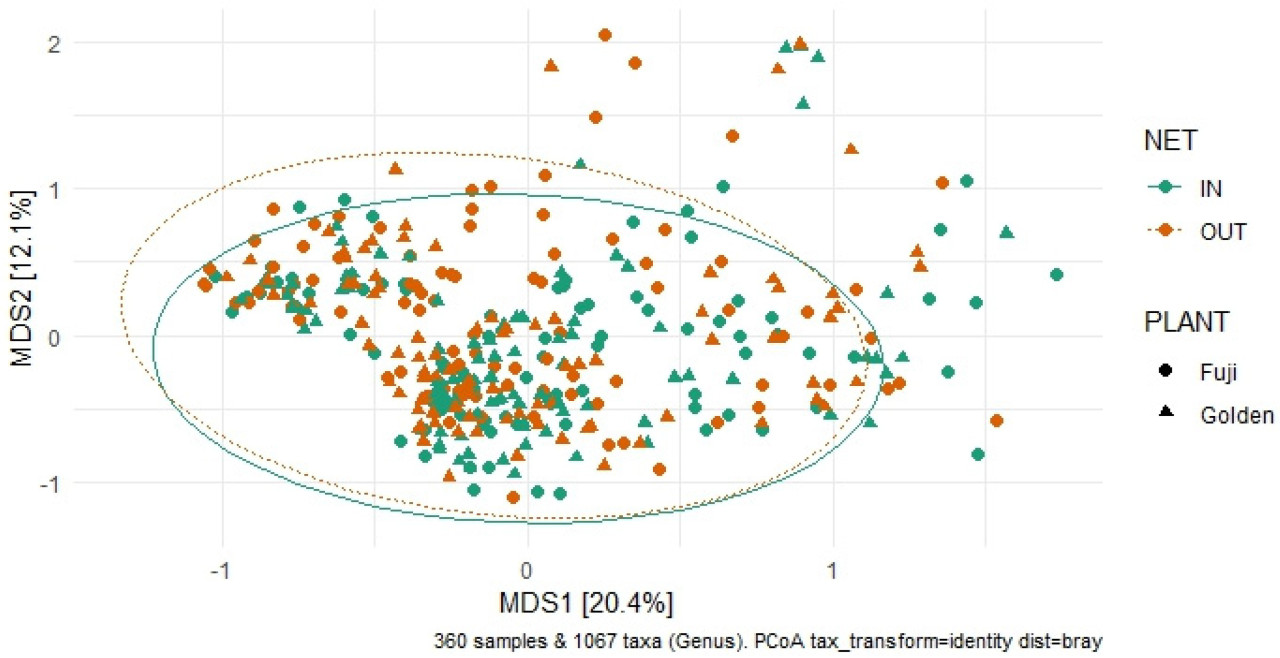
Рисунок 4. Анализ главных координат (PCoA) на основе матрицы расстояний несходства Брея-Кертиса данных секвенирования Illumina грибковых сообществ в тканях яблони (сорт Fuji, обозначенный круглым символом, и сорт Golden, обозначенный треугольным символом) в условиях IN (укрытие от дождя + сокращенное применение фунгицидов; зеленый цвет) и в условиях OUT (открытое поле + стандартная программа IPM; оранжевый цвет).
Анализ дифференциального обилия показал, что 15 видов ASV (аннотированных на уровне рода и с обилием > 0,05% в одном из двух условий) были более многочисленны внутри укрытия от дождя, чем снаружи ( таблица 2 ), и они включали Alternaria , Claviceps, Cladosporium , Lophiostoma , Podosphaera , Pseudopithomyces , Venturia и Vishniacozyma . Листовые эпифиты были наиболее пострадавшим сообществом в условиях IN и OUT, особенно во второй год тестирования, когда их состав значительно различался между условиями IN и OUT во все три времени отбора проб (T3, T4 и T5; PERMANOVA p < 0,05; таблица S5 ). Анализ дифференциального обилия был применен для определения того, какие таксоны грибов сообщества эпифитов листьев различаются в двух условиях роста, и выявил роды Prosthemium и Aureobasidium как более многочисленные в условиях ВЫХОДА (0,17% ВЫХОДА против 0,00% ВХОДА и 0,05% ВЫХОДА против 0,02% ВХОДА соответственно; согласно отрицательному биномиальному распределению, скорректированное значение p < 0,05), в то время как Claviceps и Venturia были более многочисленны в условиях ВХОДА (0,20% ВХОДА против 0,17% ВЫХОДА и 0,07% ВХОДА против 0,00% ВХОДА; согласно отрицательному биномиальному распределению, скорректированное значение p < 0,05).
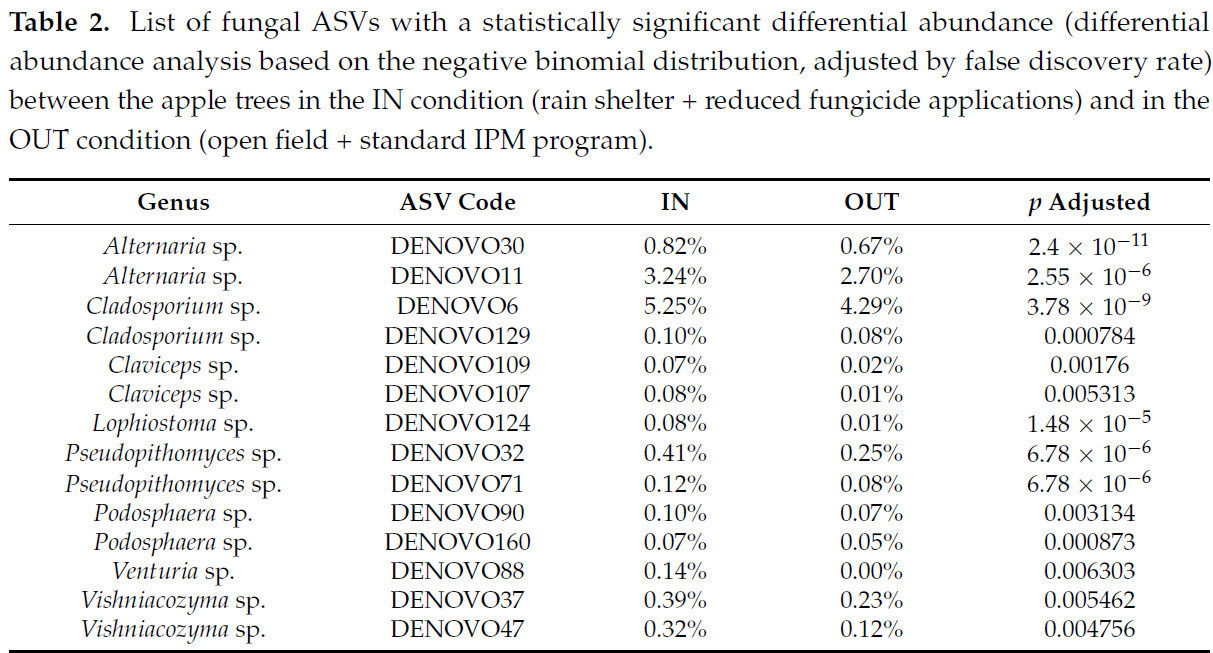
Таблица 2. Список грибковых ASV со статистически значимой дифференциальной численностью (анализ дифференциальной численности на основе отрицательного биномиального распределения, скорректированного с учетом частоты ложных обнаружений) между яблонями в условиях IN (укрытие от дождя + сокращенное применение фунгицидов) и в условиях OUT (открытый грунт + стандартная программа IPM).
Более того, состав грибкового сообщества растений сорта Фуджи отличался от такового у растений сорта Голден Делишес (PERMANOVA, p = 0,001 , рисунок 4 и таблица S5 ), а дифференциальный анализ численности подтвердил различия между двумя сортами яблок. В частности, Botrytis , Heterobasidion , Hygrophorus , Pseudopithomyces , Starmerella , Symmetrospora , Tilleptiopsis , Venturia и Vishniacozyma были более многочисленны у сорта Голден Делишес по сравнению с Фуджи, тогда как Diplodia , Fusarium и Podosphaera были более многочисленны у Фуджи по сравнению с Голден Делишес ( таблица 3 ).
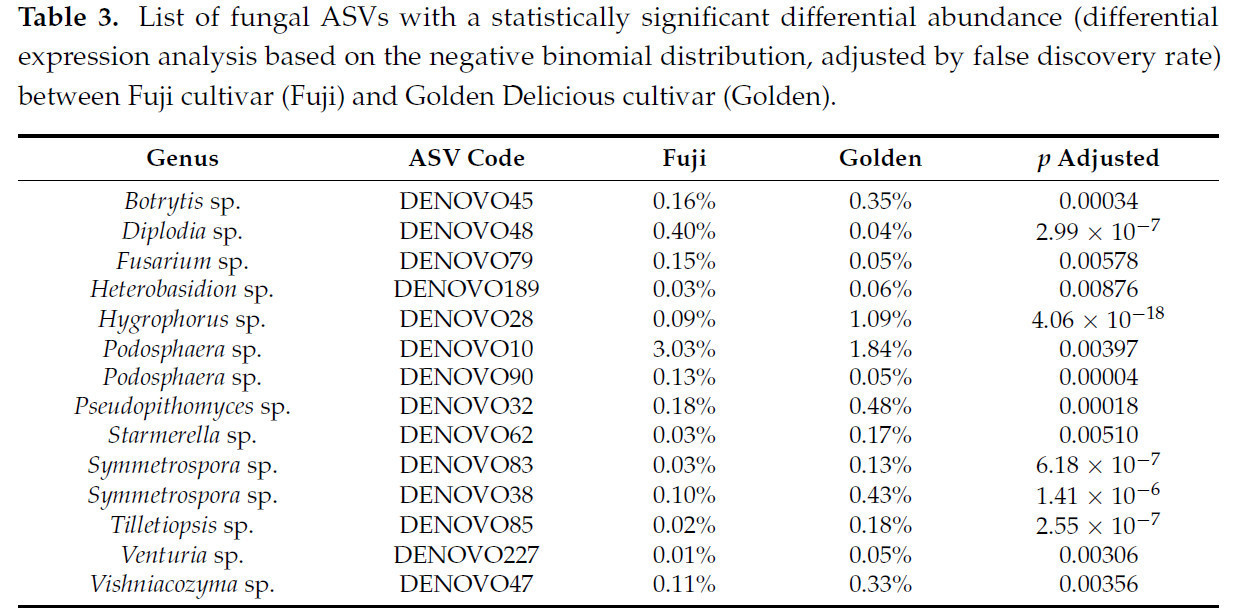
Таблица 3. Список грибковых ASV со статистически значимой дифференциальной распространенностью (анализ дифференциальной экспрессии на основе отрицательного биномиального распределения, скорректированный с учетом частоты ложных обнаружений) между сортом Фуджи (Fuji) и сортом Голден Делишес (Golden).
4. Обсуждение
Использование укрытий от дождя стало многообещающей агрономической практикой для защиты сельскохозяйственных культур от осадков, тем самым снижая потребность в фунгицидах, особенно против болезней, которым способствуют дожди и длительные периоды увлажнения листьев. Эта практика может быть интегрирована в программы IPM для поддержки устойчивого сельскохозяйственного производства [ 36 ]. Ключевым аспектом защиты растений является понимание того, как агрономические практики в IPM влияют на микробиом растений, поскольку микроорганизмы играют важную роль в здоровье растений. Поскольку предыдущие исследования показали, что агрономические практики могут существенно влиять на микробиом, связанный с яблоней [ 19 , 37 , 38 ], эта работа специально изучает, как укрытия от дождя в сочетании с уменьшенным использованием фунгицидов влияют на разнообразие грибов. В частности, целью нашего исследования была оценка совместного воздействия укрытия от дождя и уменьшенного использования фунгицидов в рамках программы IPM, специально направленной на борьбу с патогенами, такими как парша яблони, которые развиваются под воздействием дождя и увлажнения листьев, с сокращенным графиком внесения фунгицидов на двух сортах яблони в саду коммерческого типа. Таким образом, наша экспериментальная установка учитывает отдельные эффекты, поскольку уменьшение расхода фунгицида является следствием укрытия от дождя.
Состав грибкового сообщества совпадает с предыдущими выводами о микробиоме яблони из того же региона [ 18 ] и включает ASV, таксономически аннотированные как потенциальные антагонисты (например, Aureobasidium sp.) и потенциальные фитопатогены (например, Alternaria sp., Cladosporium sp. и Podosphaera sp.). Среди наиболее распространенных таксонов были идентифицированы семь родов дрожжей ( Sporidiobolus , Cystobasidium , Buckleyzyma , Rhodotorula , Vishniacozyma , Symmetrospora и Filobasidium ), многие из которых принадлежат к Basidiomycetes. Это согласуется с предыдущими выводами о распространенности базидиомицетовых дрожжей на филлоплане [ 39 , 40 ] и на плодах яблони [ 41 ]. Дрожжи филлопланы проявляют обширную биодеградативную активность; Они могут усваивать многие растительные компоненты, извлекать пользу из растительных выделений, конкурировать за питательные вещества и защищать растение от фитопатогенных грибов [ 42 ]. Среди дрожжевых родов Vishniacozyma был значительно более многочисленным в условиях IN по сравнению с условиями OUT, и этот род также был обнаружен более многочисленным в сорте Golden Delicious по сравнению с Fuji, что свидетельствует о его способности адаптироваться к определенным условиям окружающей среды.
Результаты показывают, что укрытие от дождя в сочетании с уменьшенным применением фунгицидов влияет как на общую структуру грибкового сообщества, так и на обилие конкретных таксонов, и эффект может быть получен из сочетания уменьшенного внесения фунгицида и измененных условий микроклимата под укрытием от дождя (например, небольшое повышение температуры и/или влажности, изменение интенсивности солнечного света, увлажнения листьев, интенсивности ветра и распространения инокулята). В частности, растения яблони в состоянии IN показали повышенное обилие ASV, таксономически аннотированных как потенциальные грибковые патогены. Например, ASV рода Podosphaera , который включает возбудителя настоящей мучнистой росы, были более многочисленны в состоянии IN по сравнению с условием OUT, что согласуется с оценкой заболевания настоящей мучнистой росой в 2020 году. Обилие Podosphaera было выше в состоянии IN по сравнению с условием OUT, хотя фунгициды против этого патогена применялись также в состоянии IN. Аналогичным образом, ранее было обнаружено, что частота возникновения настоящей мучнистой росы выше под укрытиями от дождя, достигая уровня более 70% на растениях сорта Голден Делишес [ 2 ]. Аналогичным образом, исследование виноградных лоз сообщило об увеличении тяжести поражения настоящей мучнистой росой у растений, выращиваемых под укрытиями от дождя, по сравнению с контрольными условиями без применения фунгицидов [ 10 ]. Это было связано с влажным микроклиматом, в котором листья не смачивались под укрытиями от дождя, что способствовало прорастанию спор и заражению [ 2 , 43 ]. Кроме того, этот фитопатоген лучше процветает в затененных условиях, которые присутствуют под укрытиями от дождя из-за сниженного облучения по сравнению с условиями открытого грунта [ 10 ]. Кроме того, ASV, аннотированные как Venturia sp. были более многочисленными в состоянии IN по сравнению с состоянием OUT, что согласуется с более высокой частотой парши яблони в состоянии IN по сравнению с состоянием OUT в 2020 году, что указывает на то, что условия для заражения паршой яблони, хотя и минимальные, могут также возникать в состоянии IN или что отсутствие специфических фунгицидов позволяет развиваться более крупным популяциям V. inaequalis . Таким образом, избегать применения фунгицидов против парши яблони под укрытиями от дождя может быть слишком рискованно, поскольку некоторые листья, близкие к сетке, все еще могут намокать во время сильных боковых дождей и провоцировать инфекции, хотя и в ограниченной степени [ 2 ]. Это говорит о том, что укрытия от дождя могут эффективно контролировать Venturiasp. при коммерческом выращивании без применения фунгицидов, хотя результирующая эффективность может быть несколько ниже по сравнению со стандартной программой IPM. Кроме того, следует отметить, что эффективность укрытий от дождя с уменьшенным внесением фунгицидов зависит от восприимчивости сорта яблони и заболеваемости в саду [ 2 ]. Другие роды грибов, такие как Cladosporium и Alternaria, были более многочисленны в условиях IN. Грибы, принадлежащие к этим родам, связаны с сердцевинной гнилью, внутренней сухой гнилью плодов яблони после сбора урожая или хранения [ 44 ].
Несмотря на увеличение относительной численности ASV, таксономически аннотированных как потенциальные фитопатогены в условиях IN, укрытие от дождя с уменьшенным количеством фунгицидов также оказало положительное влияние, способствуя обилию предполагаемых антагонистов, таких как Vishniacozyma , которые могут включать возможные агенты биологического контроля [ 45 ]. С другой стороны, Aureobasidium, устойчивый черный дрожжеподобный гриб, устойчивый к высыханию и УФ-излучению [ 46 ], был более многочисленным в эпифитном сообществе в условиях OUT по сравнению с условиями IN, предполагая, что условия окружающей среды за пределами укрытий от дождя могли ограничивать рост других грибов, но не Aureobasidium . Аналогичным образом, род Prosthemium, обычный член микробиома ветвей деревьев лиственных пород [ 47 ], был более многочисленным в условиях OUT. Prosthemium считается эндофитным или эпифитным таксоном на листьях Betulaceae [ 48 ]. Этот род редко встречается в микробиоме яблони, и он был обнаружен как обильный таксон грибковых эндофитов в корнях больных пересаженных яблонь [ 49 ], хотя он не был связан с причиной заболевания.
В нашем исследовании грибковое сообщество также различалось между двумя сортами яблони. Род Tilletiopsis, который включает потенциальных возбудителей белой дымки послеуборочного расстройства [ 50 ], был более многочисленным в Golden Delicious по сравнению с Fuji, вместе с Botrytis sp. и Venturia sp. С другой стороны, Diplodia , род, включающий виды, которые, как сообщается, вызывают болезни яблони (например, пятнистость листьев «лягушачий глаз», черная гниль, отмирание и язвы) [ 51 ], был более многочисленным в Fuji, вместе с патогенными родами Podosphera и Fusarium . Эти результаты отражают влияние генотипа растения на ассоциированные эндофитные микробные сообщества в различных условиях окружающей среды и сезонах, как наблюдалось в других исследованиях комбинаций подвоя и привоя яблони [ 52 , 53 ]. Многие отчеты демонстрируют, что генотипы растений яблони различаются по ассоциированной с ними микробиоте на уровне сорта [ 54 ]. Более того, некоторые сорта яблони показали большее разнообразие микробиома, чем другие [ 55 ]. При развитии некоторых патогенов, таких как V. inaequalis, эффекты восприимчивости сорта наблюдались в экспериментальных садах без защиты фунгицидами [ 56 , 57 ], что позволило отнести сорт Golden Delicious к широкой группе, считающейся умеренно восприимчивой к парше яблони. Кроме того, коммерческие сорта яблони имеют разные уровни устойчивости к болезни мучнистой росы яблони [ 58 ]. По данным Биггса и соавторов [ 59 ], нет никаких доказательств того, что устойчивость к парше коррелирует с устойчивостью к мучнистой росе; в то время как некоторые устойчивые к парше сорта обладают дополнительной устойчивостью к мучнистой росе, многие другие сорта ею не обладают.
Способность сеток изменять условия окружающей среды, оказывая значительное влияние на освещенность, скорость ветра и влажность листьев, а также тенденция к снижению влажности и повышению температуры воздуха [ 60 ], может оказывать существенное влияние на грибковые сообщества, подвергающиеся их воздействию. Фактически, эпифитные сообщества, подвергавшиеся более сильному воздействию, пострадали сильнее, тогда как эндофитные сообщества, обладающие более защищенной средой обитания в растительных тканях, были в меньшей степени подвержены влиянию условий окружающей среды [ 61 ].
5. Выводы
В заключение следует отметить, что использование укрытий от дождя в сочетании с сокращением применения фунгицидов в программах комплексной защиты растений (IPM) может влиять на структуру и состав грибковых сообществ яблонь, в первую очередь затрагивая эпифитные грибы на листьях. Хотя использование укрытий от дождя способствует снижению расхода фунгицидов в саду, оно может увеличить численность АСВ, таксономически аннотированных как потенциальные патогены, по сравнению с IPM в условиях открытого грунта, скорее всего, из-за меньшего расхода фунгицидов. Следовательно, включение дополнительных методов биологического контроля в дополнение к укрытиям от дождя может снизить риск накопления популяций патогенов и/или улучшить управление болезнями без увеличения использования химических фунгицидов. Хотя необходимы дальнейшие исследования для количественной оценки независимых эффектов укрытий от дождя и снижения применения специфических фунгицидов в различных условиях, наше исследование показывает, что микробиом растений существенно реагирует на изменения в методах агротехники. Этот результат подчеркивает важность включения таких исследований для повышения устойчивости производства яблок и разработки оптимизированных стратегий борьбы с патогенами.
Дополнительные материалы
Следующая вспомогательная информация может быть загружена по адресу: https://www.mdpi.com/article/10.3390/agriculture15010017/s1 , Таблица S1: Управление болезнями и климатические данные сезонов 2019 и 2020 годов; Таблица S2: Необработанные данные, полученные с помощью секвенирования ампликона ITS ДНК, выделенной из коры, цветов, листьев и тканей плодов яблонь; Таблица S3: Грибковые ASV, полученные с помощью секвенирования ампликона ITS ДНК, выделенной из коры, цветов, листьев и тканей плодов яблонь; Таблица S4: Дифференциальный анализ численности на уровне семейств на основе отрицательного биномиального распределения, скорректированного по частоте ложных обнаружений между внутри укрытия и снаружи укрытия для каждого грибкового сообщества в каждую временную точку; Таблица S5: Результаты теста PERMANOVA (функция adonis2 в веганском пакете R) для грибковых сообществ между растениями внутри и снаружи укрытия.
Ссылки
1. Kemp, C. Apple revival: How science is bringing historic varieties back to life. Nature 2023, 622, 446–449. [Google Scholar] [CrossRef] [PubMed]
2. Zavagli, F.; Wenneker, M.; Vilardell, P.; Spinelli, F.; Naef, A.; Micheli, F.; Kelderer, M.; Holthusen, H.H.F.; Donati, I.; Cabrefiga, J.; et al. Benefits and limitations of apple production under rainproof covers. Acta Hortic. 2023, 1378, 253–260. [Google Scholar] [CrossRef]
3. Bowen, J.K.; Mesarich, C.H.; Bus, V.G.M.; Beresford, R.M.; Plummer, K.M.; Templeton, M.D. Venturia inaequalis: The causal agent of apple scab. Mol. Plant Pathol. 2011, 12, 105–122. [Google Scholar] [CrossRef] [PubMed]
4. Holb, I.J. Timing of first and final sprays against apple scab combined with leaf removal and pruning in organic apple production. Crop Prot. 2008, 27, 814–822. [Google Scholar] [CrossRef]
5. Manja, K.; Aoun, M. The use of nets for tree fruit crops and their impact on the production: A review. Sci. Hortic. 2019, 246, 110–122. [Google Scholar] [CrossRef]
6. Bastías, R.M.; Boini, A. Apple Production under Protective Netting Systems. In Apple Cultivation—Recent Advances; IntechOpen: London, UK, 2023. [Google Scholar]
7. Boutry, C.; Kelderer, M.; Holtz, T.; Baumgartner, F.; Friedli, M. Testing the effect of a rainproof protection net on the apple production regarding disease and pest damages. In Proceedings of the 20th International Conference on Organic Fruit-Growing, Online, 21–23 February 2022. [Google Scholar]
8. Zhang, H.; Tu, K.; Qiu, Z.; Wen, Z.; Li, Q.; Wen, X. Effects of different rain shelter coverings on volatile organic compounds in mature fruit and postharvest quality of sweet cherry. CYTA J. Food 2021, 19, 465–475. [Google Scholar] [CrossRef]
9. Claire, D.; Watters, N.; Gendron, L.; Boily, C.; Pépin, S.; Caron, J. High productivity of soilless strawberry cultivation under rain shelters. Sci. Hortic. 2018, 232, 127–138. [Google Scholar] [CrossRef]
10. Du, F.; Deng, W.; Yang, M.; Wang, H.; Mao, R.; Shao, J.; Fan, J.; Chen, Y.; Fu, Y.; Li, C.; et al. Protecting grapevines from rainfall in rainy conditions reduces disease severity and enhances profitability. Crop Prot. 2015, 67, 261–268. [Google Scholar] [CrossRef]
11. Sui, Y.; Zhao, Q.; Wang, Z.; Liu, J.; Jiang, M.; Yue, J.; Lan, J.; Liu, J.; Liao, Q.; Wang, Q.; et al. A Comparative Analysis of the Microbiome of Kiwifruit at Harvest Under Open-Field and Rain-Shelter Cultivation Systems. Front. Microbiol. 2021, 12, 757719. [Google Scholar] [CrossRef]
12. Chen, X.M.; Zhang, Q.; Zeng, S.M.; Chen, Y.; Guo, Y.Y.; Huang, X.Z. Rhizosphere soil affects pear fruit quality under rain-shelter cultivation. Can. J. Plant Sci. 2020, 100, 683–691. [Google Scholar] [CrossRef]
13. Ellis, M.A.; Ferree, D.C.; Funt, R.C.; Madden, L.V. Effects of an Apple Scab-Resistant Cultivar on Use Patterns of Inorganic and Organic Fungicides and Economics of Disease Control. Plant Dis. 1998, 82, 428–433. [Google Scholar] [CrossRef] [PubMed]
14. Compant, S.; Samad, A.; Faist, H.; Sessitsch, A. A review on the plant microbiome: Ecology, functions, and emerging trends in microbial application. J. Adv. Res. 2019, 19, 29–37. [Google Scholar] [CrossRef]
15. Adam, E.; Groenenboom, A.E.; Kurm, V.; Rajewska, M.; Schmidt, R.; Tyc, O.; Weidner, S.; Berg, G.; de Boer, W.; Falcão Salles, J. Controlling the Microbiome: Microhabitat Adjustments for Successful Biocontrol Strategies in Soil and Human Gut. Front. Microbiol. 2016, 7, 1079. [Google Scholar] [CrossRef] [PubMed]
16. Zhang, Q.; Pang, X.; Chen, X.; Ye, J.; Lin, S.; Jia, X. Rain-shelter cultivation influence rhizosphere bacterial community structure in pear and its relationship with fruit quality of pear and soil chemical properties. Sci. Hortic. 2020, 269, 109419. [Google Scholar] [CrossRef]
17. Perazzolli, M.; Antonielli, L.; Storari, M.; Puopolo, G.; Pancher, M.; Giovannini, O.; Pindo, M.; Pertot, I. Resilience of the Natural Phyllosphere Microbiota of the Grapevine to Chemical and Biological Pesticides. Appl. Environ. Microbiol. 2014, 80, 3585–3596. [Google Scholar] [CrossRef] [PubMed]
18. Longa, C.M.O.; Antonielli, L.; Bozza, E.; Sicher, C.; Pertot, I.; Perazzolli, M. Plant organ and sampling time point determine the taxonomic structure of microbial communities associated to apple plants in the orchard environment. Microbiol. Res. 2022, 258, 126991. [Google Scholar] [CrossRef] [PubMed]
19. Arrigoni, E.; Albanese, D.; Longa, C.M.O.; Angeli, D.; Donati, C.; Ioriatti, C.; Pertot, I.; Perazzolli, M. Tissue age, orchard location and disease management influence the composition of fungal and bacterial communities present on the bark of apple trees. Environ. Microbiol. 2020, 22, 2080–2093. [Google Scholar] [CrossRef]
20. Arrigoni, E.; Antonielli, L.; Pindo, M.; Pertot, I.; Perazzolli, M. Tissue age and plant genotype affect the microbiota of apple and pear bark. Microbiol. Res. 2018, 211, 57–68. [Google Scholar] [CrossRef] [PubMed]
21. McLaughlin, M.S.; Yurgel, S.N.; Abbasi, P.A.; Prithiviraj, B.; Ali, S. Impacts of abiotic factors on the fungal communities of ‘Honeycrisp’ apples in Canada. Microb. Biotechnol. 2023, 16, 1639–1656. [Google Scholar] [CrossRef] [PubMed]
22. Perazzolli, M.; Vicelli, B.; Antonielli, L.; Longa, C.M.O.; Bozza, E.; Bertini, L.; Caruso, C.; Pertot, I. Simulated global warming affects endophytic bacterial and fungal communities of Antarctic pearlwort leaves and some bacterial isolates support plant growth at low temperatures. Sci. Rep. 2022, 12, 18839. [Google Scholar] [CrossRef]
23. Klaubauf, S.; Inselsbacher, E.; Zechmeister-Boltenstern, S.; Wanek, W.; Gottsberger, R.; Strauss, J.; Gorfer, M. Molecular diversity of fungal communities in agricultural soils from Lower Austria. Fungal Divers. 2010, 44, 65–75. [Google Scholar] [CrossRef]
24. Tedersoo, L.; Bahram, M.; Põlme, S.; Kõljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Global diversity and geography of soil fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef] [PubMed]
25. Keiblinger, K.M.; Schneider, M.; Gorfer, M.; Paumann, M.; Deltedesco, E.; Berger, H.; Jöchlinger, L.; Mentler, A.; Zechmeister-Boltenstern, S.; Soja, G.; et al. Assessment of Cu applications in two contrasting soils—Effects on soil microbial activity and the fungal community structure. Ecotoxicology 2018, 27, 217–233. [Google Scholar] [CrossRef]
26. Albanese, D.; Fontana, P.; De Filippo, C.; Cavalieri, D.; Donati, C. MICCA: A complete and accurate software for taxonomic profiling of metagenomic data. Sci. Rep. 2015, 5, 9743. [Google Scholar] [CrossRef] [PubMed]
27. Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet.J. 2011, 17, 10. [Google Scholar] [CrossRef]
28. Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
29. Kessy, A.; Allan, K.; Timo, P.; Raivo, P.; Filipp, I.; Henrik, N.R.; Kõljalg, H. Urmas: Full UNITE+INSD dataset for eukaryotes. UNITE Community. 2021. Available online: https://mothur.org/wiki/unite_its_database/ (accessed on 23 December 2024).
30. Porter, T.M. Terrimporter/UNITE_ITSClassifier: UNITE v2.0; Zenodo.2021; Zenodo: Geneva, Switzerland, 2021. [Google Scholar] [CrossRef]
31. McMurdie, P.J.; Holmes, S. phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
32. Nicola, L.; Landínez-Torres, A.Y.; Zambuto, F.; Capelli, E.; Tosi, S. The Mycobiota of High Altitude Pear Orchards Soil in Colombia. Biology 2021, 10, 1002. [Google Scholar] [CrossRef] [PubMed]
33. Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
34. Wickham, H. ggplot2; Use R! Springer International Publishing: Cham, Germany, 2016; ISBN 978-3-319-24275-0. [Google Scholar]
35. Barnett, D.; Arts, I.; Penders, J. microViz: An R package for microbiome data visualization and statistics. J. Open Source Softw. 2021, 6, 3201. [Google Scholar] [CrossRef]
36. Brun, L.; Combe, F.; Gros, C.; Walser, P.; Saudreau, M. Protecting Apricot Orchards with Rain Shelters Reduces Twig Blight Damage Caused by Monilinia spp. and Makes It Possible to Reduce Fungicide Use. Agronomy 2023, 13, 1338. [Google Scholar] [CrossRef]
37. Schaeffer, R.N.; Pfeiffer, V.W.; Basu, S.; Brousil, M.; Strohm, C.; DuPont, S.T.; Vannette, R.L.; Crowder, D.W. Orchard Management and Landscape Context Mediate the Pear Floral Microbiome. Appl. Environ. Microbiol. 2021, 87, 1–13. [Google Scholar] [CrossRef]
38. Coller, E.; Cestaro, A.; Zanzotti, R.; Bertoldi, D.; Pindo, M.; Larger, S.; Albanese, D.; Mescalchin, E.; Donati, C. Microbiome of vineyard soils is shaped by geography and management. Microbiome 2019, 7, 1–15. [Google Scholar] [CrossRef]
39. Inácio, J.; Pereira, P.; De Carvalho, M.; Fonseca, Á.; Amaral-Collaço, M.T.; Spencer-Martins, I. Estimation and diversity of phylloplane mycobiota on selected plants in a Mediterranean-type ecosystem in Portugal. Microb. Ecol. 2002, 44, 344–353. [Google Scholar] [CrossRef] [PubMed]
40. Wang, K.; Sipilä, T.P.; Overmyer, K. The isolation and characterization of resident yeasts from the phylloplane of Arabidopsis thaliana. Sci. Rep. 2016, 6, 1–13. [Google Scholar] [CrossRef]
41. Kristjuhan, A.; Kristjuhan, K.; Tamm, T. Richness of yeast community associated with apple fruits in Estonia. Heliyon 2024, 10, e27885. [Google Scholar] [CrossRef] [PubMed]
42. Middelhoven, W.J. Identity and biodegradative abilities of yeasts isolated from plants growing in an arid climate. Antonie van Leeuwenhoek, Int. J. Gen. Mol. Microbiol. 1997, 72, 81–89. [Google Scholar] [CrossRef]
43. Huang, R.; Jiang, J.; Su, Y.; Yu, H.; Shen, L.; Liu, Y.; Qin, Y.; Song, Y. Microbial community succession and volatile compounds changes during spontaneous fermentation of Cabernet Sauvignon (Vitis vinifera L.) under rain-shelter cultivation. Food Chem. X 2023, 17, 100618. [Google Scholar] [CrossRef]
44. Watpade, S.; Lal, M.K.; Tiwari, R.K.; Kumar, R.; Naga, K.C.; Kumar, R.; Pramanick, K.K.; Kumari, H.; Devi, E.; Pal, D. Fusarium core rot disease affects physicochemical and pathophysiological attributes of apple (Malus × domestica Borkh.) fruit. Postharvest Biol. Technol. 2024, 208, 112648. [Google Scholar] [CrossRef]
45. Gorordo, M.F.; Lucca, M.E.; Sangorrín, M.P. Biocontrol Efficacy of the Vishniacozyma Victoriae in Semi-Commercial Assays for the Control of Postharvest Fungal Diseases of Organic Pears. Curr. Microbiol. 2022, 79, 259. [Google Scholar] [CrossRef] [PubMed]
46. Rensink, S.; van Nieuwenhuijzen, E.J.; Sailer, M.F.; Struck, C.; Wösten, H.A.B. Use of Aureobasidium in a sustainable economy. Appl. Microbiol. Biotechnol. 2024, 108, 202. [Google Scholar] [CrossRef] [PubMed]
47. Kowalski, T.; Kehr, R.D. Endophytic fungal colonization of branch bases in several forest tree species. Sydowia 1992, 44, 137–168. [Google Scholar]
48. Kamiyama, M.; Hirayama, K.; Tanaka, K.; Mel’nik, V.A. Transfer of Asterosporium orientale to the genus Prosthemium (Pleosporales, Ascomycota): A common coelomycetous fungus with stellate conidia occurring on twigs of Betula spp. Mycoscience 2009, 50, 438–441. [Google Scholar] [CrossRef]
49. Popp, C.; Wamhoff, D.; Winkelmann, T.; Maiss, E.; Grunewaldt-Stöcker, G. Molecular identification of Nectriaceae in infections of apple replant disease affected roots collected by Harris Uni-Core punching or laser microdissection. J. Plant Dis. Prot. 2020, 127, 571–582. [Google Scholar] [CrossRef]
50. Boekhout, T.; Gildemacher, P.; Theelen, B.; Müller, W.H.; Heijne, B.; Lutz, M. Extensive colonization of apples by smut anamorphs causes a new postharvest disorder. FEMS Yeast Res. 2006, 6, 63–76. [Google Scholar] [CrossRef] [PubMed]
51. Bhat, A.H.; Shah, M.D.; Padder, B.A.; Shah, Z.A.; Dar, E.A.; Fayaz, U.; Nain, M.S.; Ali, M.A.; Al-Hemaid, F.M.; Stępień, P.; et al. Morphological, pathogenic and genetic diversity in Diplodia seriata associated with black rot canker of apple in India. Sci. Rep. 2023, 13, 15682. [Google Scholar] [CrossRef] [PubMed]
52. Liu, J.; Abdelfattah, A.; Wasserman, B.; Wisniewski, M.; Droby, S.; Fazio, G.; Mazzola, M.; Wu, X. Contrasting effects of genotype and root size on the fungal and bacterial communities associated with apple rootstocks. Hortic. Res. 2022, 9, uhab013. [Google Scholar] [CrossRef]
53. Liu, J.; Abdelfattah, A.; Norelli, J.; Burchard, E.; Schena, L.; Droby, S.; Wisniewski, M. Apple endophytic microbiota of different rootstock/scion combinations suggests a genotype-specific influence. Microbiome 2018, 6, 18. [Google Scholar] [CrossRef]
54. Papp-Rupar, M.; Karlstrom, A.; Passey, T.; Deakin, G.; Xu, X. The Influence of Host Genotypes on the Endophytes in the Leaf Scar Tissues of Apple Trees and Correlation of the Endophytes with Apple Canker (Neonectria ditissima) Development. Phytobiomes J. 2022, 6, 127–138. [Google Scholar] [CrossRef]
55. Britt, E.; Bösch, Y.; Perren, S.; Naef, A.; Frey, J.E.; Bühlmann, A. The apple fruit microbiome: Influence of orchard management, cultivar, storage time and storage atmosphere. Acta Hortic. 2021, 1325, 317–324. [Google Scholar] [CrossRef]
56. Brun, L.; Didelot, F.; Parisi, L. Effects of apple cultivar susceptibility to Venturia inaequalis on scab epidemics in apple orchards. Crop Prot. 2008, 27, 1009–1019. [Google Scholar] [CrossRef]
57. Dewdney, M.; Charest, J.; Paulitz, T.; Carisse, O. Multivariate analysis of apple cultivar susceptibility to Venturia inaequalis under greenhouse conditions. Can. J. Plant Pathol. 2003, 25, 387–400. [Google Scholar] [CrossRef]
58. Strickland, D.A.; Hodge, K.T.; Cox, K.D. An Examination of Apple Powdery Mildew and the Biology of Podosphaera leucotricha from Past to Present. Plant Health Prog. 2021, 22, 421–432. [Google Scholar] [CrossRef]
59. Biggs, A.R.; Sundin, G.W.; Rosenberger, D.A.; Yoder, K.S.; Sutton, T.B. Relative Susceptibility of Selected Apple Cultivars to Apple Scab Caused by Venturia inaequalis. Plant Health Prog. 2010, 11, 20. [Google Scholar] [CrossRef]
60. Kjaer, K.H.; Petersen, K.K.; Bertelsen, M. Protective rain shields alter leaf microclimate and photosynthesis in organic apple production. Acta Hortic. 2016, 1134, 317–325. [Google Scholar] [CrossRef]
61. Gomes, T.; Pereira, J.A.; Benhadi, J.; Lino-Neto, T.; Baptista, P. Endophytic and Epiphytic Phyllosphere Fungal Communities Are Shaped by Different Environmental Factors in a Mediterranean Ecosystem. Microb. Ecol. 2018, 76, 668–679. [Google Scholar] [CrossRef] [PubMed]

Longa CMO, Nicola L, Pindo M, Bozza E, Sicher C, Bondesan D, Pertot I, Perazzolli M. Fungal Biodiversity of Apple Bark, Leaves, Stems, and Fruit Under Rain Shelters with Reduced Fungicide Schedule. Agriculture. 2025; 15(1):17. https://doi.org/10.3390/agriculture15010017
Перевод статьи «Fungal Biodiversity of Apple Bark, Leaves, Stems, and Fruit Under Rain Shelters with Reduced Fungicide Schedule» авторов Longa CMO, Nicola L, Pindo M, Bozza E, Sicher C, Bondesan D, Pertot I, Perazzolli M., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык

















































































































Комментарии (0)