Совершенствование системы CRISPR-Cas и её эффективное применение в растениях
Создание системы CRISPR–Cas открыло беспрецедентные возможности для исследования генома растений и улучшения генетики сельскохозяйственных культур.
Аннотация
В последние годы эта система постоянно совершенствовалась для удовлетворения потребностей человека посредством расширения и модификации белков Cas, диверсификации областей воздействия и оптимизации векторов CRISPR. В данном обзоре мы систематически описываем белки Cas II класса, используемые в растениях, деактивированный Cas9 (dCas9) и его роль в регуляции транскрипции, системы точного редактирования, варианты белка Cas9, а также методы и примеры систем CRISPR–Cas, нацеленные на различные области с различной широтой охвата. Кроме того, мы описываем планы оптимизации, основанные на конструкциях CRISPR, которые позволяют преодолеть плейотропию генов или ускорить получение растений без трансгенов, а также применение систем CRISPR в селекции растений. Наконец, мы обсуждаем теорию и развитие «CRISPR плюс», а также комплексное применение существующих систем у большего числа видов.
1. Введение
Растущие климатические изменения и разнообразие потребностей человека требуют эффективной системы для достижения быстрого улучшения растений [ 1 ]. С 2013 года система CRISPR – CRISPR-ассоциированный белок (Cas), основанная на адаптивной фаговой иммунной системе прокариот, была впервые применена к растениям [ 2 , 3 ]. Исследователи отдают ей предпочтение за такие преимущества, как простота использования, высокая эффективность и стабильность, и ее применение привело к генетическому улучшению многих видов [ 4 , 5 ].
Для содействия применению системы CRISPR–Cas усилиями многих исследовательских групп были разработаны постоянные улучшения и эволюции системы [ 4 ]. Это включало расширение и модификацию белков Cas, разнообразие областей редактирования и добавление кофакторов, среди прочего. Различные белки Cas могут быть нацелены на разные шаблоны и обеспечивать меньший объем [ 6 , 7 ]. На основе модификаций белка Cas9 сгенерированные редакторы оснований, включая редакторы оснований цитозина (CBE) и редакторы оснований аденина (ABE), могут достигать точного редактирования оснований. А генерация первичных редакторов (PE) достигает точного редактирования в более широком диапазоне [ 8 , 9 ]. Специфические варианты Streptococcus pyogenes Cas9 (SpCas9), такие как SpCas9-NG и SpRY, позволяют редактировать гены без ограничений, связанных с мотивом, прилегающим к протоспейсеру (PAM), что может способствовать разработке более эффективных одиночных направляющих РНК (sgRNA) [ 10 , 11 , 12 , 13 ]. Дезактивированный Cas9 (dCas9) сохраняет свою способность таргетироваться на определенные регионы, что позволяет регулировать транскрипцию генов [ 14 , 15 , 16 , 17 ]. Кроме того, характеристики системы CRISPR–Cas позволяют экспрессировать несколько sgRNA одновременно, что позволяет целенаправленно редактировать несколько генов, что удобно для изучения функциональной избыточности между генами. Более того, область редактирования не ограничивается кодирующими последовательностями (CDS) генов; Цис-регуляторные элементы (CRE) в промоторе и расположенных выше открытых рамках считывания (uORF) также могут быть целевыми [ 18 , 19 ]. Для лучшего и более быстрого понимания функций генов был также проведен скрининг CRISPR на нескольких видах, что привело к появлению множества мутантов с различными фенотипами [ 20 , 21 ]. В конструкциях CRISPR–Cas использование тканеспецифичных промоторов способствует повышению эффективности редактирования и изучению плейотропных генов [ 22 , 23 ]. Более того, добавление кофакторов, включая обычные репортерные гены для идентификации трансгенных растений, морфогенетические гены для стимуляции регенерации и гены мужской стерильности (МС) или эмбриональные летальные гены для снижения или прямого блокирования образования трансгенных растений у потомства, может находить различные применения [ 24 , 25 , 26 , 27]. Эти стратегии, которые постоянно совершенствуются, служат руководством для эффективного использования системы CRISPR–Cas в растениях. После создания подходящей системы CRISPR–Cas, получение регенерированных растений также находится в центре внимания исследователей, а методы доставки реагентов для редактирования и стратегии регенерации также совершенствуются [ 28 , 29 ]. Эти результаты указывают на то, что системы CRISPR–Cas демонстрируют большой потенциал, особенно у видов, функциональные исследования которых только начинаются, и для которых необходимо подробное руководство по применению.
Этот обзор фокусируется на интегрированных приложениях системы CRISPR–Cas, которые относительно легко реализовать в большинстве лабораторий. Расширение и модификация белков Cas, различные целевые области с различной широтой, оптимизация набора инструментов CRISPR, модернизация методов доставки и применение системы CRISPR–Cas в практике селекции растений систематически обобщены в этом обзоре ( Рисунок 1 ), чтобы предоставить читателям навигационную карту всеобъемлющей системы CRISPR–Cas. В этом обзоре также обсуждаются новые возможности применения систем CRISPR в будущем, с меньшим акцентом на конкретных функциях генов. Наша цель - предоставить справочную информацию для исследователей, занимающихся функцией генов или генетическим улучшением сельскохозяйственных культур, и способствовать лучшему применению системы CRISPR для большего количества видов растений.

Рисунок 1. Обзор расширений, модификаций, обновлений и оптимизаций системы CRISPR, а также методов ее доставки и применения в селекционной практике.
2. Разнообразие и модификация Cas-белков
Археи и бактерии достигают адаптивного иммунитета посредством серии систем CRISPR–Cas, каждая из которых состоит из комбинации белков Cas и CRISPR РНК (crRNA) [ 30 ]. Различные системы CRISPR–Cas делятся на две категории: системы класса I (типы I, III и IV) используют несколько белков Cas и crRNA для формирования комплексов, тогда как системы класса II (типы II, V и VI) объединяют крупные однокомпонентные белки Cas с crRNA для опосредования целевого расщепления, например, хорошо охарактеризованного Cas9 [ 7 , 31 ].
Поскольку белок dCas9 по-прежнему сохраняет свою способность к таргетированию, регуляция транскрипции может быть достигнута путем связывания dCas9 с активаторами или супрессорами транскрипции [ 32 , 33 ]. Большинство изменений, вызванных двухцепочечными разрывами (DSB), являются нокаутирующими мутациями, а полученные правки часто случайны, что затрудняет удовлетворение требований улучшения селекции. Для решения этой проблемы были предложены редакторы оснований, которые объединяют никазу Cas9 (Cas9n) с цитидин- или аденозиндезаминазой для получения целевого редактирования оснований [ 34 , 35 ], а создание PE решает ограниченное количество переходов оснований и обеспечивает точные вставки и делеции [ 8 ]. SpCas9 является наиболее часто используемым белком Cas, но из-за его характеристики распознавания NGG PAM (где N - любой нуклеотид), подходящие или эффективные sgRNA для требуемой области не могут быть разработаны для множества генов. Таким образом, было разработано несколько вариантов Cas9, нацеленных на более широкий спектр PAM [ 12 ].
2.1. Cas-белки класса II
Cas9 относится к белкам Cas класса II типа II, типа V и типа VI, а также используется в растениях, предоставляя новые возможности для систем CRISPR–Cas из-за различий в структурах, размерах, способностях к нацеливанию и специфичности ( Рисунок 2 ). Cas9 содержит домен нуклеазы HNH, встроенный в домен RuvC, который вместе расщепляет двухцепочечную ДНК (dsDNA) [ 6 ]. Белки Cas12, которые относятся к типу V, содержат один домен нуклеазы RuvC, который нацеливается и коллатерально расщепляет одноцепочечную РНК (ssRNA) и ssDNA, а также может выполнять надрез и расщепление dsDNA [ 7 ]. Белок Cas13 типа VI нацелен на ssRNA, что обеспечивает уникальные преимущества в регуляции уровней РНК и защите от вирусов [ 36 ].

Рисунок 2. Различные системы Cas класса II, применяемые в растениях. ( A ) Система Cas9 содержит домен нуклеазы HNH, встроенный в домен RuvC, и требует комбинации crRNA и трансактивирующей crRNA (tracrRNA), что в конечном итоге приводит к образованию тупых концов, расположенных проксимально выше по течению от PAM NGG. ( B ) Система Cas12a требует только crRNA и производит ступенчатые концы и крупные делеции, расположенные дистально ниже по течению от PAM TTTV (V = A, C или G). ( C ) Система Cas12b требует как crRNA, так и tracrRNA и производит ступенчатые концы и крупные делеции, расположенные дистально ниже по течению от PAM VTTV. ( D ) CasΦ примерно в два раза меньше Cas9 и распознает PAM TBN (B = G, T или C). ( E ) Система Cas13 содержит два структурно различных домена связывания нуклеотидов (HEPN) высших эукариот и прокариот, которые обеспечивают активность РНКазы, распознают и расщепляют одноцепочечную РНК, но не могут расщеплять двуцепочечную ДНК.
2.1.1.Cas12a
CRISPR–Cas12a (Cpf1) является еще одной хорошо изученной системой в дополнение к Cas9 [ 7 ]. Эта система использует T-богатый PAM (TTTV) для идентификации целевой ДНК, что облегчает нацеливание генных промоутеров и других AT-богатых сайтов [ 37 ]. В отличие от Cas9, которому требуется объединение crRNA и tracrRNA в sgRNA, Cas12a требует только crRNA и обладает РНКазной активностью для выполнения собственного процессинга crRNA. Таким образом, система CRISPR–Cas12a является хорошей платформой для мультимодального редактирования [ 38 , 39 , 40 , 41 ]. Сайт расщепления Cas12a расположен дистально ниже последовательности PAM, а не проксимально выше, как сайт расщепления Cas9; Cas12a производит ступенчатые концы и большие делеции, тогда как Cas9 производит DSB с тупыми концами, что приводит к небольшим вставкам и делециям [ 42 ]. Активность Cas12a по исследованию ДНК, расщеплению и высвобождению продукта была дополнительно охарактеризована с помощью анализа флуоресценции отдельных молекул и биохимических анализов [ 43 ]. Анализ активности редактирования при различных температурах показывает, что система Cas12a чувствительна к температуре, причем высокие температуры повышают ее эффективность редактирования [ 44 ]. Среди некоторых улучшений, эффективность редактирования Cas12a также была улучшена. Модифицированный массив тРНК–crRNA позволил редактировать несколько генов и успешно редактировать целевые сайты, которые были пропущены массивами crRNA [ 45 ]. Было исследовано в общей сложности 17 новых ортологов Cas12a, и было доказано, что Ev1Cas12a и Hs1Cas12a имеют высокую эффективность редактирования, при этом Hs1Cas12a показывает более низкую температурную чувствительность [ 46 ]. Более того, более эффективный вариант LbCas12a был получен посредством насыщающего мутагенеза, а LbCas12a-RRV в сочетании с RV и D156R продемонстрировал большую эффективность редактирования в рисе и тополе [ 47 ].
2.1.2.Cas12b
CRISPR–Cas12b (C2c1) содержит RuvC и Nuc домены. Эта система также имеет тенденцию нацеливаться на T-богатые PAM и производить ступенчатые концы. Но, как и Cas9, Cas12b требует как crRNA, так и tracrRNA [ 48 , 49 , 50 ]. Более того, Cas12b считается привлекательным вариантом из-за его небольшого размера и высокой специфичности, но оптимальная температура для расщепления ДНК Cas12b, как правило, превышает 40 °C, что затрудняет его использование в растительных клетках [ 51 ]. Эффективность редактирования AaCas12b при различных температурных условиях и продолжительностях была оценена на хлопке ( Gossypium hirsutum ) из-за его устойчивости к высоким температурам, и результаты показали, что эффективность редактирования была наибольшей при температуре 45 °C и продолжительности 4 дня. AaCas12b индуцировал делеции 1–16 пар оснований (п.н.), причем большинство делеций имели длину 9–14 п.н.; эти делеции больше, чем те, которые индуцирует Cas9 (1–5 п.н.) [ 52 ]. Интересно, что было обнаружено, что эффективность редактирования этой системы зависела от структуры хроматина [ 53 ]. У риса сравнивали эффективность редактирования четырех белков Cas12b; среди них наиболее эффективным был AaCas12b, который преимущественно распознавал PAM ATTV и GTTG [ 54 ]. Сравнение пяти направляющих РНК-скаффолдов показало, что Aac и Aa1.2 значительно увеличивали эффективность редактирования без нецелевой активности, а целевые мутации в основном передавались зародышевой линии [ 55 ]. У Arabidopsis Cas12b чаще всего индуцировал делеции 5–13 п.н., что согласуется с результатами по хлопку и рису. Более того, Cas12b может успешно индуцировать множественное редактирование генома, приводя к делеции примерно 1 кб между двумя целевыми участками [ 56 ]. Система редактирования генома с использованием мультиплексных прямых повторов (DR)-спейсеров (iMAGE) на основе Cas12i3, которая использует другой белок Cas12, также обеспечивает более высокую частоту хромосомных структурных вариаций по сравнению с Cas9 [ 57 ].
2.1.3.CasΦ
CasΦ (Cas12j) — это гиперкомпактный белок Cas, обнаруженный в гигантских фагах [ 58 ]. CasΦ примерно в два раза меньше SpCas9 и Cas12a (~70 кДа), что означает, что его можно упаковывать в векторы на основе вирусов [ 7 ]. Белок CasΦ распознает TBN PAM, особенно NTTV; использует один домен RuvC для расщепления ДНК, а также может использоваться для процессинга crRNA. CasΦ был доставлен в виде рибонуклеопротеинов (РНП) для нацеливания гена PDS3 в протопластах Arabidopsis , и в целевом сайте были вызваны делеции 8–10 п.н. [ 58 ]. В клетках риса и томата в целевом сайте, дистальнее PAM, часто встречается делеция 7–13 п.н., и протоспейсеры длиной 18 п.н. или длиннее более эффективны. Более того, регенерация протопластов может быть более эффективной при редактировании, опосредованном CasΦ, у растений [ 59 ]. CasΦ нечувствителен к температуре, но чувствителен к хроматиновому окружению, и эффективность редактирования увеличивалась, когда целевой участок не был метилирован. По сравнению с CasΦ дикого типа (WT), варианты vCasΦ и nCasΦ демонстрируют более высокую эффективность и специфичность [ 60 ].
2.1.4.Cas13
В системе CRISPR–Cas типа VI белок Cas13, который содержит два структурно различных нуклеотидсвязывающих домена (HEPN) высших эукариот и прокариот, которые обеспечивают активность РНКазы, распознает и расщепляет одноцепочечную РНК, но не может расщепить двуцепочечную ДНК; Cas13 также может воздействовать на пре-crRNA и обрабатывать ее для создания функциональной crRNA [ 61 , 62 ]. Эта система делится на шесть подтипов: VI-A (Cas13a, C2c2), VI-B (Cas13b, C2c4), VI-C (Cas13c, C2c7), VI-D (Cas13d), VI-X (Cas13X) и VI-Y (Cas13Y) [ 36 ]. Cas13X и Cas13Y (от 775 до 803 аминокислот [аа]) представляют собой два компактных семейства, полученные из метагеномных наборов данных [ 63 ]. Abudayyeh et al. [ 64 ] использовали LwaCas13a для нацеливания на EPSPS , HCT и PDS в протопластах риса, и большинство направляющих достигли более 50% нокдауна. И применение этой системы в сельскохозяйственных культурах является особенно многообещающим для вмешательства в вирусы растений [ 65 ]. CRISPR–Cas13a использовался для вмешательства в вирус мозаики репы (TuMV) в Nicotiana benthamiana и Arabidopsis , а уровни снижения зависели от целеуказанных РНК [ 66 , 67 ]. Кроме того, CRISPR–Cas13 эффективно мешал вирусу картофеля Y (PVY), вирусу южной черной полосатой карликовости риса (SRBSDV), вирусу мозаики риса полос (RSMV) и вирусу хлоротической карликовости батата (SPCSV)-RNase3 [ 68 , 69 , 70 ]. Вариант CasRX (Cas13d из Ruminococcus flavefaciens ) является наименьшим из вариантов Cas13a-d, но показал наибольшую интерференционную активность в N. benthamiana [ 71 , 72 ]. Одновременное нацеливание на несколько вирусов РНК картофеля было достигнуто с помощью эндогенной системы процессинга тРНК (полицистронной тРНК-гРНК [PTG]) [ 73 ]. Cas13d также использовался для нацеливания на различные мишени РНК, включая микроРНК (miRNA), длинные некодирующие РНК (lncRNA) и кольцевые РНК (circRNA) [ 74 ]. Более того, что удивительно, crRNA, предназначенная для управления Cas13, может вызывать значительное снижение уровней РНК без белка Cas13. Cas13-независимое управляемое индуцированное подавление генов (GIGS) наблюдалось у табака, томатов и Arabidopsis и может быть распространено на crRNA, разработанные Cas9, с длиной более 28 пар оснований [ 75 ]. Кроме того, CRISPR–Cas13 использовался для обнаружения и количественной оценки генов [ 76 ].
2.2. dCas9 и регуляция транскрипции
Сверхэкспрессия является наиболее часто используемым методом исследования функции гена. Однако из-за большой рабочей нагрузки клонирования, ограниченной емкости вектора и необходимости использования нескольких промоторов достижение мультигенной сверхэкспрессии традиционными методами является сложным и неэффективным [ 77 ]. Целевые мутации RuvC D10A и HNH H841A генерируют белок dCas9. В сочетании с направляющими РНК и транскрипционными эффекторами система может достигать активации или ингибирования гена ( Рисунок 3 ). Комбинация dCas9 и транскрипционных активаторов, таких как домен активации EDLL (dCas9-EDLL), домен активации TAL (dCas9-TAD) и VP64, слияние четырех тандемных повторов VP16 (dCas9-VP64), составляет систему активации CRISPR (CRISPRa) [ 32 , 33 ]. Однако активация целевого гена этими системами была менее эффективна, поэтому было создано второе поколение систем CRISPRa. Среди них dCas9-TV полагается на шесть копий мотива TAD, подобного активатору транскрипции (TALE), соединенного с VP128 [ 77 ], dCas9-SunTag объединяет тандемный массив пептидов GCN4, которые могут рекрутировать активаторы транскрипции VP64 [ 78 ], а CRISPR-Act2.0 основан на рекрутировании VP64 и модификации направляющей РНК-матрицы [ 79 ]. Различия в способности этих систем CRISPRa активировать целевые гены привели к созданию CRISPR-Act3.0, которая объединяет dCas9-VP64, gRNA2.0, 10xGCN4 SunTag и 2xTAD активатора [ 16 ]. А полученная платформа CRISPR-Combo, использующая каркасы gRNA1.0 и gRNA2.0 с протоспейсерами разной длины, достигла одновременного редактирования и активации [ 15 , 80 ]. Недавние исследования были сосредоточены на применении доменов активации транскрипции (AD) растений. Цинсельмейер и др. [ 14 ] отметили, что система, в которой VP64 заменяется AD растительного происхождения, такими как программируемые активаторы транскрипции (PTA), имеет значительные возможности для улучшения. Используя протопласты Arabidopsis и Seteria в качестве моделей двудольных и однодольных растений соответственно, были протестированы несколько AD, и было обнаружено, что DREB2 , AvrXa10 , DOF1 , AtHSFA6b и DREB1 имели эффективность активации, равную или превышающую таковую у обычно используемого VP64. Однако необходима дальнейшая валидация. Более того, транскрипционные AD растений были идентифицированы в масштабе генома, и объединение этих AD с постоянно совершенствующимися системами CRISPR–Cas может способствовать дальнейшему развитию транскрипционной регуляции [ 81 ].
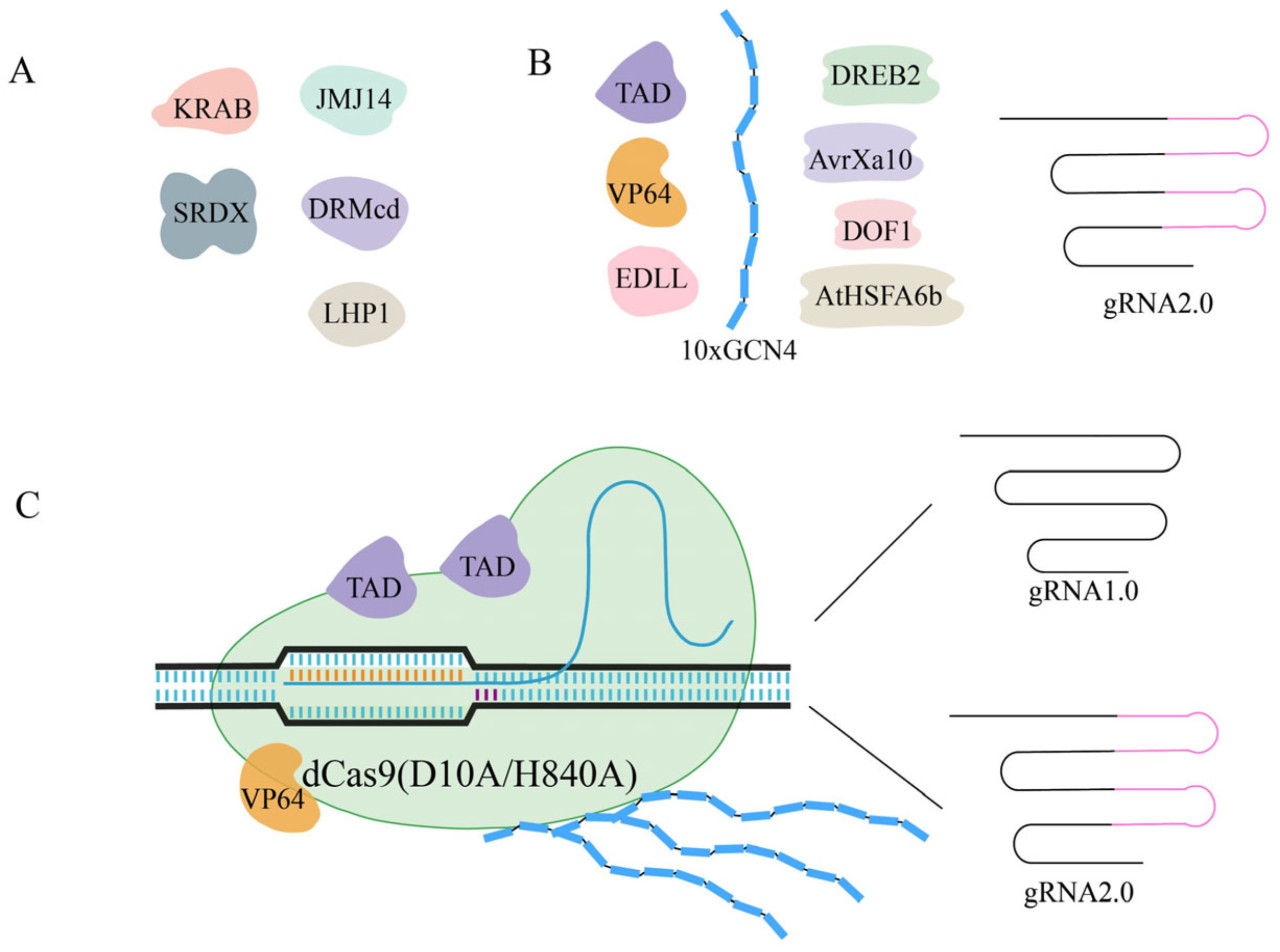
Рисунок 3. Сочетание белка dCas9 с транскрипционными эффекторами и эпигенетическими модификациями обеспечивает транскрипционную регуляцию целевых генов. ( A ) Эти транскрипционные эффекторы и эпигенетические модификации, слитые с dCas9, могут обеспечить ингибирование транскрипции. ( B ) Транскрипционные эффекторы, AD растительного происхождения и некоторые стратегии, которые могут усилить активацию целевых генов. ( C ) Платформа CRISPR–Combo с каркасами gRNA1.0 и gRNA2.0 позволяет одновременно осуществлять редактирование и активацию.
В настоящее время система CRISPRa практикуется во многих видах растений. Одним из потенциальных применений CRISPRa является настройка метаболома растений путем активации выбранных ферментов в определенных метаболических путях, например, селективное обогащение нарингенина, эриодиктиола, кемпферола и кверцетина было достигнуто в листьях N. benthamiana с помощью dCasEV2.1 [ 82 ]. В клетках винограда обе системы dCas9-VP64 и dCas9-TV эффективно активировали ген УДФ-глюкозофлавоноидгликозилтрансферазы ( UFGT ), а уровень экспрессии CBF4 был увеличен в 3,7–42,3 раза с помощью системы dCas9-TV [ 83 ]. Когда CRISPR-Act3.0 использовался для нацеливания на промоторы семи генов у груш, четыре из семи генов были активированы как минимум десять раз [ 84 ]. Система CRISPRa также была применена в Populus , и экспрессия целевых генов увеличилась в 1,2–7,0 раз [ 85 ]. Недавно партеногенез и активация транскрипции, опосредованная эпигенетической фабрикой, также были достигнуты в кукурузе и томате с помощью инструмента CRISPRa посредством специальных экспериментальных разработок [ 86 , 87 ]. Три новых зародышевых плазмы хлопка также были получены с помощью системы dCas9-TV [ 88 ]. Более того, был разработан химически индуцируемый инструмент CRISPRa, называемый ER Tag, путем объединения индуцируемого β-эстрадиолом XVE (LexA-VP16-ER) с dCas9-SunTag, и эта система способна временно контролировать экспрессию генов [ 89 ].
Когда dCas9 коэкспрессируется с направляющей РНК, образуется комплекс распознавания ДНК, который может специфически препятствовать удлинению транскрипции, связыванию РНК-полимеразы или связыванию фактора транскрипции (TF); эта переориентированная система CRISPR–Cas была обозначена как CRISPR-интерференция (CRISPRi). CRISPRi может избежать неопределенности РНК-интерференции (RNAi) в масштабе всего генома и одновременно ингибировать несколько генов [ 90 ]. Когда dCas9 был слит с доменом ассоциированного бокса Крюппеля (KRAB) Kox1, сигнал GFP снизился в пять раз [ 91 ]. Сочетание dCas9 с ингибиторным доменом SRDX ТФ ERF (dCas9:SRDX или dCas9-3X) достигло ингибирования транскрипции [ 32 , 33 ], а деактивированный Cas12a, слитый с SRDX, также продемонстрировал мощные эффекты ингибирования транскрипции в растениях [ 42 ]. С другой стороны, эпигенетическая стратегия является распространенной реализацией CRISPRi; эта стратегия использует систему SunTag с каталитическим доменом метилтрансферазы DRM N. tabacum (DRMcd) для эффективного нацеливания метилирования ДНК на промоторы FWA и SUPERMAN и достижения подавления генов [ 78 ]. Различные факторы, включая SUVH2, JMJ14, LHP1, HD2C, ELF7 и CPL2, которые представляют различные эпигенетические пути, были добавлены в систему SunTag. И подавление целевого гена наблюдалось при применении JMJ14 и LHP1, в то время как остальные факторы приводили к слабому подавлению [ 17 ]. Однако по сравнению с CRISPRa применение CRISPRi в растениях относительно ограничено, и эффективность ингибирования сильно варьируется. Таким образом, если взять CRISPRa в качестве ориентира, для повышения эффективности ингибирования CRISPRi необходимы более эффективные стратегии, такие как скрининг высокоэффективных супрессоров транскрипции и модификация sgRNA [ 19 ].
2.3. Точное редактирование
Классическая система CRISPR–Cas использует нуклеазы, такие как Cas9, в которой домен HNH расщепляет комплементарную цепь (целевую цепь), а домен RuvC расщепляет некомплементарную цепь (нецелевую цепь) [ 92 ], что приводит к образованию двуцепочечных разрывов (DSB), которые служат субстратами клеточных механизмов репарации ДНК, включая гомологичную направленную репарацию (HDR) и негомологичное соединение концов (NHEJ) [ 93 ]. NHEJ часто приводит к неточным мутациям, тогда как HDR может производить точные модификации, но ограничен присущей ему неэффективностью в высших организмах [ 19 , 34 , 94 ]. Кроме того, редактирование путем создания DSB обычно приводит к повреждению клеток [ 95 ]. Эффективные инструменты модификации генома требуют более точного редактирования и должны быть контролируемыми. Альтернативная стратегия заключается в использовании редакторов оснований, использующих Cas9n, содержащий точечную мутацию Asp10Ala (D10A), которая инактивирует домен нуклеазы RuvC SpCas9 и генерирует одноцепочечный разрыв («ник») в целевой цепи [ 9 ]. nCas9 (D10A) связывается с различными дезаминазами для достижения точного преобразования оснований. Система PE, представляющая собой эпохальный шаг вперед в точном редактировании, использует продуманную комбинацию nCas9 (H840A) с другими инструментами, которые инактивируют домен HNH [ 8 ].
2.3.1 Редактирование цитозиновых оснований
Широко используемая система CBE BE3 состоит из nCas9 (D10A), цитидиндезаминазы и ингибиторов урацил-ДНК-гликозилазы (UDG) (UGI) [ 34 , 94 ]. Цитидиндезаминаза дезаминирует цитидин в уридин на нецелевой цепи, а UGI предотвращает дезаминирование цитидина UDG в апиримидиновый сайт (AP). Когда nCas9 (D10A) вызывает разрыв в целевой цепи, активируется путь репарации ДНК, преимущественно заменяя несоответствие U:G желаемым U:A, что приводит к переходу основания C:G в T:A после репликации ДНК [ 1 ].
У растений исходная система CBE, основанная на крысиной цитидиндезаминазе APOBEC1, генерировала целевые мутации в позициях с третьего по девятое в протоспейсере и имела предпочтение к мотивам TC [ 96 ]. Впоследствии использовались различные цитидиндезаминазы, а использование цитидиндезаминазы 1 Petromyzon marinus (PmCDA1) повысило эффективность редактирования [ 97 ]. Более того, окно редактирования оснований было расширено до 17 п.н. с помощью человеческого APOBEC3A (hAPOBEC3A), и целевые мутации C-to-T были получены очень эффективно в пшенице, рисе и картофеле [ 98 ]. Два варианта CBE, основанные на дезаминазе A3Bctd (усеченной человеческой цитидиндезаминазе APOBEC3B), устранили независящее от sgRNA нецелевое редактирование в рисе и показали улучшенную специфичность [ 99 ]. Более того, Рен и др. [ 100 ] сравнили 21 различную систему CBE и обнаружили, что PmCDA1-CBE_V04 и A3A/Y130F-CBE_V04 обладают высокой эффективностью редактирования. Для оптимизации систем CBE также использовались различные инструменты. AlphaFold2 — это метод кластеризации белков на основе структуры [ 101 ]; поскольку структура белка определяет его функцию, AlphaFold2 обеспечивает простой способ обнаружения и разработки новых дезаминаз, что расширяет область применения систем CBE для большего количества видов [ 102 ].
В CBE активность UDG ингибируется UGI, тогда как сверхэкспрессия UDG запускает репарацию эксцизии оснований (BER); UDG распознает несоответствие U: G, вырезает урацил и создает основной сайт, который надрезается AP-лиазой. Этот надрез в сочетании с надрезом, образованным Cas9 вблизи DSB, должен приводить к точной делеции [ 1 , 103 ]. На основе этого обоснования были разработаны системы делеции, индуцированные слиянием APOBEC–Cas9 (AFID), включая цитидиндезаминазу, Cas9, UDG и AP-лиазу, для создания точных делеций [ 103 ]. В дополнение к делеции, расширения одноцепочечной ДНК, производимые AFID, также могут быть использованы для создания точных вставок или замен [ 9 ].
2.3.2 Редактирование аденинового основания
Сочетание аденозиндезаминазы с nCas9 (D10A) генерирует ABE. Аденозиндезаминаза дезаминирует аденозин в инозин, который распознаётся ДНК-полимеразой как гуанозин во время репарации и репликации ДНК, реализуя преобразование A:T в G:C. Природная адениндезаминаза встречается редко, и после нескольких раундов направленной эволюции были получены высокоактивные ABE с широкой совместимостью последовательностей (ABE6 и ABE7) [ 34 ]. Впоследствии ABE были быстро применены к рису, пшенице и рапсу [ 104 , 105 ]. По сравнению с CBE, ABE показали большую специфичность таргетирования [ 106 , 107 ]. Добавление трех последовательностей ядерной локализации SV40 к C-концу nCas9, усиленной sgRNA путем модификации каркаса sgRNA и промотора гена RPS5A использовалось для повышения эффективности редактирования [ 104 , 105 ]. По сравнению с широко используемым ABE-P1, ABE-P1S (упрощенная версия 1 для растений) более эффективен в рисе, возможно, из-за большей экспрессии белка [ 108 ]. Благодаря прерывистой и непрерывной эволюции дезаминазы с помощью фага (PANCE и PACE) был получен ABE8e, содержащий восемь дополнительных мутаций. По сравнению с ABE7.10 активность (k app ) ABE8e была увеличена в 590 раз, а способность катализировать дезаминирование ДНК была увеличена до 1100 раз [ 109 , 110 ]. Рисовый ABE8e (rABE8e), сочетающий мономерный TadA8e, bis-bpNLS и оптимизацию кодонов, продемонстрировал чрезвычайно высокую эффективность редактирования в специфическом окне редактирования [ 111 ]. Более того, TadA9 был создан путём включения мутаций V82S/Q154R в TadA8e, и он достиг сопоставимого или улучшенного редактирования по сравнению с TadA8e [ 112 ].
2.3.3. Редактирование на двух базах
CBE и ABE могут индуцировать только один тип модификации [ 113 ], поэтому были созданы редакторы двойных оснований. Редактор целевого эндогенного мутагенеза с насыщенными основаниями (STEME) объединяет nCas9 (D10A) с цитидиндезаминазой, аденозиндезаминазой и UGI. В протопластах риса STEME-1 достигал одновременных замен C-на-T и A-на-G с эффективностью до 15,10% [ 114 ]. Одновременное широкое редактирование, индуцированное одной системой (SWISS), мультиплексный ортогональный редактор оснований (MoBE) и система высокоэффективных редакторов двойных оснований растений (PhieDBEs) также являются мощными инструментами для создания мультиплексного редактирования оснований [ 35 , 115 , 116 ]. Однако эти системы основаны на одновременном применении цитидин- и аденозиндезаминаз, что приводит к увеличению размера вектора. TadDE, редактор с двумя основаниями, полученный из TadA-8e, может генерировать редактирование с двумя основаниями с более компактной структурой [ 117 ].
2.3.4 Редактирование трансверсии
CBE и ABE были широко развиты в растениях, но типы преобразования оснований ограничены. Однако в приложениях ABE и CBE исследователи наблюдали трансверсии C-to-G [ 118 , 119 ]. Для повышения эффективности этой трансверсии был создан редактор оснований C-to-G (CGBE), состоящий из nCas9, цитидиндезаминазы rAPOBEC1 или ее варианта R33A и урацил-ДНК N-гликозилазы, полученной из Escherichia coli (eUNG) или rXRCC1, полученной от крыс. eUNG преобразует урацил в сайт AP и инициирует BER [ 119 , 120 ]. CGBE добились редактирования C-to-G с разной эффективностью в рисе, томате и тополе. Основными побочными продуктами были правки C-to-T, а также вставки и удаления (индели) [ 121 , 122 ].
Для достижения эффективной трансверсии аденина ABE8e был объединен с вариантами мышиной алкиладенин-ДНК-гликозилазы (mAAG), что позволило эффективно осуществлять трансверсии оснований A-в-C и A-в-T [ 123 ]. У риса слияние белка N-метилпурин-ДНК-гликозилазы, высвобождающего гипоксантин (MPGv3, также называемого AAG), с rBE49b привело к образованию новой системы, названной pAKBE (K = G или T). По сравнению с клетками млекопитающих, pAKBE демонстрировал частые переходы A-в-G и низкую эффективность трансверсии A-в-T у риса [124, 125].
2.3.5. Монтаж премьер-версии
Революционная технология была разработана в 2019 году, и ее появление решило проблему ограниченного числа переходов оснований. Эта система может производить все виды замен оснований и делать целевые индели фрагментов ДНК без необходимости использования DSB или шаблонов донорской ДНК [ 1 , 8 ]. Система PE объединяет nCas9 (H840A), обратную транскриптазу (RT) и направляющую РНК для редактирования праймера (pegRNA), которая состоит из шаблона RT, содержащего генетическую информацию для желаемой мутации, и сайта связывания праймера (PBS). PBS спаривается с одноцепочечными разрывами ДНК, созданными nCas9 (H840A), запуская обратную транскрипцию и интегрируя генетическую информацию в геном [ 1 ]. Для улучшения PE1 были приняты различные стратегии; PE2 использует сконструированную RT, а PE3 добавляет дополнительную gRNA для создания еще одного разрыва в целевой цепи, что стимулирует репарацию ДНК [ 8 ].
С момента своего создания система PE была быстро внедрена в такие
растения, как рис, пшеница, кукуруза и томат. Благодаря оптимизации кодонов,
промотора и условий редактирования, обогащению с помощью отбора клеток с
отредактированными основаниями, разработке улучшенной sgRNA (esgRNA) и T m -направленной длины PBS, трансформации генов
слияния с P2A и использованию двойной pegRNA, эффективность PE в растениях улучшилась [
126 , 127 , 128 , 129 , 130 , 131 ]. Более того, оценка нецелевой активности
показала высокую специфичность этой системы [ 132 ]. ePPEplus достиг большего повышения
эффективности за счет введения замены V223A в RT сконструированного
растительного первичного редактора (ePPE), изменения сигналов ядерной локализации (NLS) и увеличения активности Cas9 [ 133 , 134 ]. Более
компактный и эффективный PE6
был получен с помощью эволюции с помощью фагов и белковой инженерии [ 135 ].
Более того, практика мультиплексного редактирования праймов в пшенице и рисе
улучшила гибкость и применимость PE [ 136 ]. По сравнению с sgRNA, используемыми с Cas9 или редакторами оснований, pegRNA сложнее проектировать, и они
играют важную роль в определении эффективности редактирования. Также были
разработаны вспомогательные инструменты проектирования и анализа, такие как PlantPegDesigner
( http://www.plantgenomeediting.net/ ) доступный 24 декабря 2024 г. [ 131 ], PE-Designer
( http://www.rgenome.net/pe-designer/
) доступный 24 декабря 2024 г., и PE-Analyzer
( http://www.rgenome.net/pe-analyzer/
) доступный 24 декабря 2024 г. [ 137 ].
Эффективное редактирование необходимо для точной работы с геномом, и было показано, что система репарации несоответствий ДНК (MMR) способствует образованию нежелательных побочных продуктов инделей. При коэкспрессии доминантно-негативного белка MMR (MLH1dn) с системой PE2/PE3 средняя эффективность редактирования PE4/PE5 увеличивалась с 2,8 до 7,7 раз [ 138 ]. Однако эффект у растений был слабым [ 139 ], в то время как прямое подавление OsMLH1 в системе ePE5c увеличивало эффективность с 1,30 до 2,11 раз. В то же время была введена система условного удаления для преодоления частичной стерильности, вызванной подавлением OsMLH1 [ 140 ].
Редактирование праймов также обеспечивает руководство для точных делеций крупных фрагментов; метод редактирования, основанный на паре pegRNA, PRIME-Del, может эффективно индуцировать делеции до 10 кб и одновременно вставлять короткие фрагменты в геном [ 141 ]. Другая система делеции и репарации на основе PE–Cas9 (PEDAR) была создана путем слияния нуклеазы Cas9 с обратной транскриптазой (RT) и объединения этого комплекса с двумя pegRNA, что позволяет эффективно создавать крупные точные геномные изменения [ 142 ]. С другой стороны, PrimeRoot (рекомбинация подходящих целей, опосредованная редактированием праймов) также может производить крупные вставки ДНК [ 143 ].
2.4. Варианты белка Cas9
В практических приложениях технологии CRISPR–Cas необходимость наличия PAM, таких как NGG или TTN, для распознавания мишени ограничивает геномные сайты, которые могут быть целевыми [ 144 ]. В результате было разработано несколько вариантов белка Cas9, нацеленных на более широкий спектр сайтов, что позволяет проводить целевое редактирование практически без PAM. Два варианта SpCas9, xCas9 и Cas9-NG, распознают более пермиссивные NG PAM. xCas9 демонстрирует почти такую же эффективность редактирования, как Cas9, на большинстве типичных NGG PAM, но показывает ограниченную активность на нетипичных NGH PAM (H = A, C или T). В случае несовпадения sgRNA xCas9 обладает лучшей специфичностью нацеливания, чем Cas9 [ 10 , 11 , 145 ]. Два варианта Cas9-NG, Cas9-NGv1 и Cas9-NG, были разработаны для нацеливания на NGN PAM [ 146 ], с кристаллическими структурами, указывающими на то, что потеря третьего специфического к основанию взаимодействия азотистого основания компенсируется новыми неспецифическими к основанию взаимодействиями, что позволяет распознавать NG PAM [ 144 ]. Эффективность редактирования Cas9-NG намного выше, чем у Cas9-NGv1 на участках NG PAM, но эффективность редактирования на NGG PAM относительно низкая. Cas9-NG также обладает более высокой специфичностью, чем Cas9, на NGG PAM [ 10 , 11 ].
Для обеспечения более широкого таргетирования PAM был разработан вариант SpG, способный таргетировать расширенный набор NGN PAM; дальнейшая оптимизация привела к разработке почти свободного от PAM варианта SpRY (NRN и NYN PAM; R = A или G и Y = C или T) [ 12 ]. В растениях SpG поддерживает NGD PAM (D = A, G или T); хотя он менее эффективен, чем SpCas9-NG, SpRY достигает эффективного редактирования в широком диапазоне геномных локусов, что указывает на предпочтение NGD и NAN PAM [ 13 , 147 , 148 ]. Несколько других вариантов SpCas9, таких как SpCas9-NRRH, SpCas9-NRCH и SpCas9-NRTH, распознают NRNH PAM, тем самым еще больше расширяя диапазон редактирования системы SpCas9 в растениях [ 149 ]. К счастью, в то же время, когда разрабатывались различные варианты Cas9, также были разработаны соответствующие системы регуляции транскрипции и редакторы оснований, и их эффективность была продемонстрирована на различных растениях [ 16 , 149 ].
3. Системы CRISPR–Cas, нацеленные на разные области с разной широтой охвата
3.1. Нацеливание на CDS
Редактирование CDS генов является наиболее распространенным применением CRISPR. Поскольку функции разных членов одного и того же семейства генов часто избыточны, необходимо производить мутанты, которые содержат несколько генов, мутировавших одновременно [ 150 ]. Системы CRISPR–Cas предоставляют множество эффективных способов экспрессии Cas9 с несколькими gRNA. На основе лигирования Golden Gate или сборки Gibson были получены тандемные кассеты экспрессии sgRNA, каждая из которых содержала промотор и терминатор полимеразы (Pol) III. Благодаря этому подходу до восьми кассет экспрессии sgRNA были вставлены в один бинарный вектор CRISPR–Cas9, и семь из восьми целевых генов были мутированы в трех растениях T0 [ 151 ]. Система эндогенной тРНК-процессинга осуществляет мультиплексное редактирование посредством эндогенных тРНК-процессирующих РНК-ферментов (РНКаза P и РНКаза Z), которые распознают компоненты тРНК и вырезают их из PTG в отдельные гРНК [ 152 ]. Рибонуклеаза 4 типа йерсинии (Csy) (Csy4) системы CRISPR также использовалась для одновременной экспрессии нескольких гРНК, а комбинация систем экспрессии Csy4 и тРНК была почти в два раза эффективнее одного промотора Pol III для экспрессии гРНК. А способность к мутагенезу была улучшена за счет добавления экзонуклеазы Trex2 [ 153 ]. Благодаря активности процессинга crRNA Cas12a, интронные crRNA также могут быть расщеплены для редактирования нескольких генов [ 154 ]. Более того, sgRNA, управляемые различными промоторами Pol III, были собраны несколькими рестриктазами [ 155 ], и широко используются изокаудомеры, такие как Spe I/Nhe I, BamH I/Bgl II, Xba I/Nhe I и Xho I/Sal I. Промежуточные векторы были переварены и, наконец, введены в вектор редактирования [ 156 ]. Теоретически, эта система может быть использована для редактирования неограниченного количества генов одновременно [ 156 ]. За счет уменьшения длины промотора sgRNA было собрано 12 кассет экспрессии sgRNA [ 157 ]. Кроме того, были проведены структурно-направленный сайт-мутагенез и случайный скрининг; было отобрано более 35 вариантов каркаса sgRNA и идентифицировано множество функциональных вариантов. Используя эти варианты и различные тРНК, с помощью ПЦР слияния можно синтезировать единицы спейсер-скеффолд-тРНК-спейсер, содержащие до 9 мишеней; В сочетании с лигированием Golden Gate можно эффективно клонировать до 24 целевых участков за один шаг [ 158 ].
Выбор sgRNA является важным фактором, определяющим эффективность и специфичность систем CRISPR–Cas. На сегодняшний день создано множество веб-сайтов, посвященных дизайну sgRNA генома растений ( таблица 1 ), включая CRISPR-P 2.0 ( http://cbi.hzau.edu.cn/CRISPR2/ ), доступ к которому был осуществлен 24 декабря 2024 г. [ 159 ], локальный инструмент для дизайна sgRNA CRISPR-Local ( http://crispr.hzau.edu.cn/CRISPR-Local/ ), доступ к которому был осуществлен 24 декабря 2024 г. [ 160 ], и интегрированный инструментарий CRISPR-GE ( http://skl.scau.edu.cn/ ), доступ к которому был осуществлен 24 декабря 2024 г. [ 161 ]. Другие комплексные инструменты, включая Cas designer ( http://rgenome.net/cas-designer/ ), доступ к которому осуществлен 24 декабря 2024 года, и CRISPOR ( http://crispor.org ), доступ к которому осуществлен 24 декабря 2024 года, также охватывают значительный объем информации о геноме растений, а всестороннее рассмотрение результатов с нескольких веб-сайтов поможет в выборе лучших sgRNA [ 162 , 163 , 164 ].


Таблица 1. Наборы инструментов для проектирования sgRNA и анализа результатов редактирования.
3.2. CRISPR-скрининг
Система CRISPR–Cas9, специфичная только к целевым участкам в 20 нуклеотидов (нт), обеспечивает высокопроизводительное редактирование, что способствует функциональным геномным исследованиям и может обеспечить ценные генетические ресурсы для улучшения сельскохозяйственных культур [ 169 ]. Высокое качество, высокий охват и равномерное распределение являются гарантией эффективного применения скрининга CRISPR [ 21 ]. Были разработаны два популярных метода скрининга CRISPR: скрининг с использованием массивов и скрининг с использованием пулов; благодаря характеристикам применения скрининга с использованием массивов, в растениях чаще используются скрининговые пулы sgRNA [ 170 ]. На сегодняшний день скрининги CRISPR были проведены на многих растениях ( таблица 2 ). Практика большого скрининга CRISPR первоначально проводилась на рисе, который содержит 88 541 sgRNA для нацеливания на 34 234 гена, и было получено 91 004 трансгенных растения с эффективностью редактирования около 83,9%. В этих трансгенных линиях наблюдались фенотипические изменения фертильности, угла побега и цвета листьев [ 171 ]. В том же году была также зарегистрирована библиотека, содержащая 25 604 sgRNA, нацеленных на 12 802 гена, которые экспрессируются в стебле риса в высокой степени, что позволило получить 14 000 растений T0 [ 172 ]. В томате было разработано 4379 sgRNA для 990 ТФ и получено 487 положительных растений, только 92 из которых содержали одну sgRNA, с эффективностью редактирования 23%. Доля растений, содержащих одну sgRNA, была увеличена до 42% путем трансформации небольшой библиотеки, нацеленной на 30 ТФ [ 173 ]. В рапсе также была создана библиотека CRISPR, охватывающая 10 480 генов посредством 18 414 sgRNA, и было получено 1·104 растений T0 с эффективностью редактирования 55,80% [ 174 ].
Фактически, текущее построение библиотеки CRISPR по всему геному по-прежнему сосредоточено в основном на рисе. У других видов были созданы библиотеки CRISPR с небольшими объемами или специфическими функциями, а мутантные библиотеки подходящего размера демонстрируют преимущества в изучении специальных функций. Первоначально, у томата, было получено 15 мутантных линий путем нацеливания 54 иммуноассоциированных лейциновых повторов подсемейства XII генов с 165 sgRNA; фенотипический анализ показал, что Fls2.1 играет важную роль в реакции листьев на flg22 [ 175 ]. Используя семьдесят векторов CRISPR–Cas9 для нацеливания генов-кандидатов с потенциальными функциями в образовании клубеньков или семенах у сои, было получено в общей сложности 407 линий T0, содержащих все sgRNA, со средней частотой мутаций 59,2%, из которых 35,6% линий несли множественные мутации [ 176 ]. В кукурузе 1368 sgRNA, нацеленных на 1244 гена, идентифицированных с помощью генетического картирования и сравнительного геномного анализа, были объединены и произвели 4356 устойчивых к глифосату растений T0, содержащих 743 целевых гена, и было обнаружено много неожиданных фенотипических изменений [ 177 ]. В рисе также были созданы библиотеки, специфичные для CRISPR, связанные с MS, рецептор-подобной киназой (RLK) и генами, предпочитающими семена [ 178 , 179 , 180 ]. Интересно, что библиотеки CRISPR, основанные на различных признаках, также были созданы в древесном растении (хлопке), включая гены, которые дифференциально экспрессируются во время развития растения, эндогенные гены, связанные с устойчивостью к насекомым, и семейство генов кальций-зависимой протеинкиназы (CDPK) [ 181 , 182 , 183 ]. Обнаружение многоцелевой вставки в этих исследованиях также указало на важность создания специфических библиотек CRISPR, что имеет преимущество при анализе одного и того же признака.
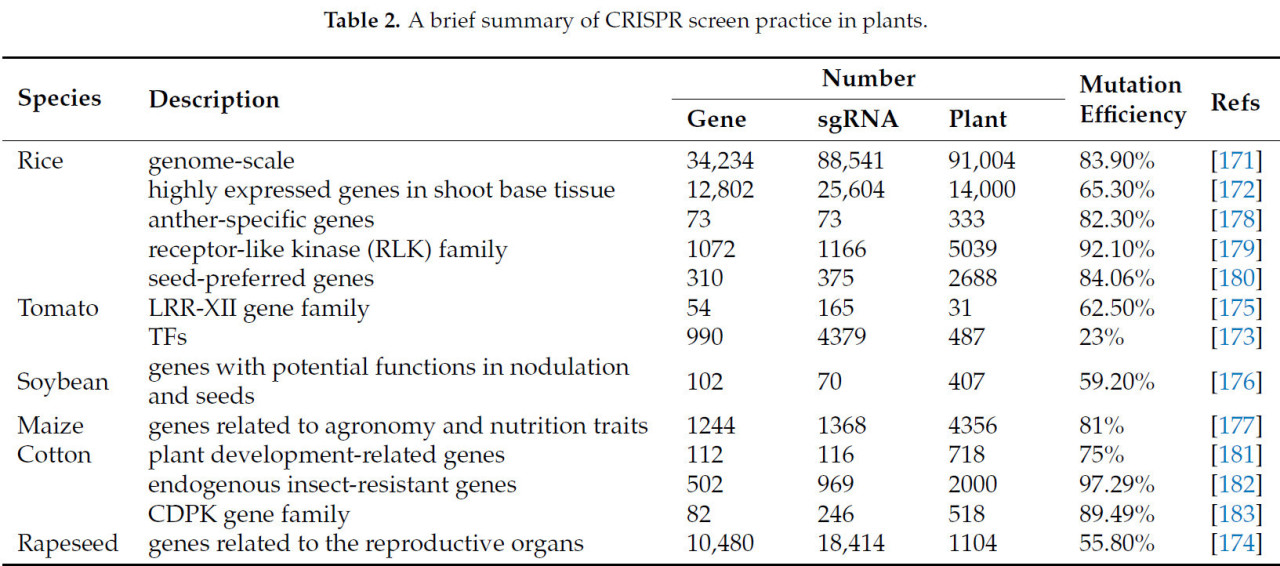
Таблица 2. Краткое описание практики скрининга CRISPR у растений.
Идентификация sgRNA растений T0 в основном основана на целевом секвенировании амплификации. Чен и др. [ 179 ] разработали маркеры длины фрагмента для различения конвейера gRNAs (FLASH); выровненная библиотека CRISPR, основанная на конвейере FLASH, использовалась для нацеливания 1072 членов семейства RLK в рисе, и быстрая идентификация gRNAs была достигнута без секвенирования. В последнее время было предложено много новых систем для обеспечения руководства по лучшему использованию библиотек CRISPR. Для преодоления функциональной избыточности был разработан набор инструментов под названием multi-knock. Всего было разработано 59 129 оптимальных sgRNA, каждая из которых нацелена на 2–10 генов в пределах семейства, всего 16 152 гена, и разделено на 10 функциональных подбиблиотек для обеспечения гибких и целевых генетических скринингов; путем преобразования одной из этих подбиблиотек были обнаружены новые TRP со скрытыми функциями из-за генетической избыточности [ 150 ]. Объединив селекцию с редактированием генов кукурузы, была предложена система BREEDIT, основанная на мультиплексных схемах редактирования генома и гибридизации с использованием CRISPR, для установления связи между традиционной селекцией и генной инженерией [ 184 ]. Все эти системы могут быть распространены на другие виды.
CRISPR-скрины также использовались для CDS отдельных генов путем разработки широкого спектра sgRNA, которые, как было показано, обеспечивают насыщающий мутагенез и способствуют эволюции белка. Батт и др. [ 185 ] разработали 119 sgRNA на основе участков, прилегающих к PAM в CDS SF3B1, и получили мутанты с различной степенью устойчивости к ингибиторам сплайсинга. Из-за случайности мутаций, вносимых CRISPR–Cas9, впоследствии были применены точные системы редактирования. В сочетании с библиотеками gRNA была осуществлена направленная эволюция генов устойчивости к гербицидам, включая ацетил-КоА-карбоксилазу (ACC) [ 186 ], ацетолактатсинтазу 1 (ALS1) [ 187 ] и 5-енолпирувилшикимат-3-фосфатсинтазу (EPSPS) [ 100 ]. Для прямой эволюции OsACC также использовались редакторы двойных оснований и система насыщающего мутагенеза, опосредованного библиотекой первичного редактирования (PLSM) [ 114 , 188 ]. Совсем недавно Ван и др. [ 189 ] разработали эффективную систему, специфичную для зародышевой линии, на основе улучшенного редактора оснований и способности Arabidopsis производить много семян; с помощью этой системы были получены устойчивые к гербицидам варианты EPSPS, ALS и HPPD.
Секвенирование по Сэнгеру является распространенным методом обнаружения мутаций, и инструменты биоинформатики, такие как Tracking of Indels by Decomposition (TIDE; http://tide.nki.nl ), дата обращения 24 декабря 2024 г. и Inference of CRISPR Edits (ICE; https://ice.synthego.com ), дата обращения 24 декабря 2024 г., могут быть использованы для декодирования данных секвенирования по Сэнгеру, содержащих различные типы мутаций [ 4 ]. Однако чувствительность секвенирования по Сэнгеру составляет всего около 15 %; поэтому правки с низкой частотой, вероятно, будут проигнорированы. Кроме того, это громоздко и дорого при применении объединенных стратегий, в то время как высокопроизводительное секвенирование следующего поколения (NGS) на основе штрихкодов показало уникальные преимущества в скрининге CRISPR. Также были разработаны аналитические инструменты, такие как Hi-TOM2 [ 165 ], CRISPResso2 [ 166 ], CRISPR-GRANT [ 167 ] и CrisprStitch [ 168 ] ( таблица 1 ). Эти инструменты позволяют удобно анализировать большие наборы данных, генерируемые NGS.
3.3. Нацеливание CRE на промотор и uORF
Системы CRISPR–Cas способны генерировать множество видов генетических вариантов. Для плейотропных генов, активных в нескольких органах или участвующих в нескольких регуляторных путях, редактирование CRE может дать значительно лучшие результаты, чем редактирование CDS. В сочетании с CRISPR-скринингом можно получить множество цис-регуляторных аллелей с несвязанными регуляторными признаками, что может способствовать одомашниванию сельскохозяйственных культур и улучшению количественных признаков [ 18 ].
Многие практики редактирования CRE были реализованы на растениях. Нацеливая эффекторный связывающий элемент (EBEP thA4 ) CsLOB1 , можно достичь повышенной устойчивости к раку цитрусовых [ 190 ]. CLAVATA 3 (CLV3) и WUSCHEL (WUS) поддерживают высококонсервативную петлю отрицательной обратной связи для регуляции пролиферации клеток. В томате мутация CRE, опосредованная CRISPR–Cas9, использовалась для характеристики взаимосвязей между CLV3 и количественными признаками и выяснения аддитивных и синергических отношений между консервативными последовательностями [ 191 , 192 ]. Более того, Хендельман и соавторы [ 193 ] сконструировали много аллелей промотора WUSCHEL HOMEOBOX9 ( WOX9 ) в томате, выявив скрытую плеотропную роль WOX9, и распространили ее на другие виды. Пептидный сигнал CLAVATA3/EMBRYO SURROUNDING REGION-RELATED (CLE) в пути обратной связи CLV3-WUS также регулирует размер меристемы. Использование CRISPR–Cas9 для создания слабых аллелей CLE улучшило признаки, связанные с урожайностью [ 194 ]. Кроме того, благодаря целенаправленному редактированию промотора Wx и трёх основных генов, связанных со сроками колошения, удалось точно настроить уровни амилозы и сроки колошения [ 195 , 196 ].
У риса мутация Ideal Plant Architecture 1 ( IPA1 ) увеличила количество зерен на метелку, но уменьшила количество побегов. Путем трансформации 39 векторов, содержащих 2–4 sgRNA в промоторе, нетранслируемых областях (UTR) и нижестоящих областях, была выполнена делеция тайлинга. Делеция 54 п.н. в промоторе одновременно увеличила количество и размер метелок [ 197 ]. Этот метод представляет собой новый тип скрининга CRISPR по сравнению со стандартным, вовлекая две sgRNA, одновременно нацеливающиеся на последовательность, что приводит к большой делеции. Фенотипическая регуляция, достигаемая путем целевого редактирования в определенных сайтах, также дает руководство для идентификации сайтов связывания ТФ [ 197 ]. Между тем, модель глубокого обучения BPNet, которая использует последовательности ДНК для прогнозирования профиля связывания иммунопреципитации хроматина с разрешением по основаниям (ChIP)-нексуса плюрипотентных ТФ, была валидирована с помощью точечной мутации, опосредованной CRISPR [ 198 ]. Эти результаты показывают, что редактирование на основе промотора играет важную роль в разобщении плейотропии генов и характеристике количественных связей между CRE и количественными признаками. Однако некоторые свойства промоторов, включая обогащение оснований AT, низкую сложность последовательности и сильную повторяемость элементов, накладывают определенные требования на систему CRISPR–Cas [ 199 ]. Белок Cas12a и инструменты точного редактирования являются высокоэффективными альтернативными инструментами [ 18 , 200 ]. В рисе система редактирования промотора CRISPR–Cas12a (CAPE) ввела континуумы количественной вариации признаков (QTV) для нескольких признаков посредством редактирования промоторов генов [ 201 ]. Таким образом, системы редактирования, нацеленные на промоторные области, имеют большой потенциал.
Функции генов богаты и разнообразны, и для создания желаемых признаков, в дополнение к нокауту, также важную роль играет стимуляция экспрессии генов. Будучи эффективными CRE в мРНК, uORF играют важную роль в контроле инициации трансляции нижестоящих первичных ORF (pORF) [ 202 ], и они проявляются в качестве общего механизма, который ингибирует уровни белка [ 203 , 204 ]. Было показано, что редактирование последовательностей uORF позволяет достичь сверхэкспрессии [ 19 ]. В салате редактирование uORF LsGGP2 , который участвует в биосинтезе витамина C, улучшило антиоксидантную способность и содержание аскорбата [ 203 ]. Редактирование высококонсервативной uORF FvebZIPs1.1 позволило увеличить содержание сахара в клубнике [ 205 ]. Кроме того, при редактировании uORF гена SlGGP1 наблюдалось обогащение аскорбатом, что было связано с МС у томатов [ 206 ]. Многократный мутагенез uORF гена PSBS1 у риса выявил фенотип сверхэкспрессии и двух-трехкратное увеличение содержания белка OsPsbS1 [ 207 ]. Эти результаты свидетельствуют о том, что редактирование uORF представляет собой универсальный и эффективный способ управления трансляцией мРНК, который может быть применен для изучения механизмов биологических процессов или повышения качества и урожайности сельскохозяйственных культур.
4. Оптимизация векторов CRISPR–Cas
4.1. Применение тканеспецифичных промоторов
Промоторы вируса мозаики цветной капусты (CaMV) 35S и убиквитина часто используются для управления экспрессией Cas9, но применение этой системы в Arabidopsis относительно неэффективно по сравнению с рисом, и многие мутации не наследуются в следующем поколении [ 151 ]. Для достижения эффективной доставки целевых мутаций потомству белок Cas9 управлялся тканеспецифичными промоторами, включая промоторы INCURVATA2 и YAO , оба преимущественно экспрессируются в активно делящихся клетках [ 208 , 209 ]. Гены SPOROCYTELESS ( SPL ), DD45 и томата LAT52 экспрессируются в спорогенных клетках и микроспороцитах, яйцеклетках и клетках пыльцы соответственно. Промоторы этих генов использовались для управления экспрессией Cas9 в клетках зародышевой линии, что привело к увеличению скорости наследуемых генных мутаций [ 210 ]. Мики и др. [ 211 ] сообщили о методе последовательной трансформации, с помощью которого они сравнили четыре промотора, специфичных для зародышевой линии ( DD45 , Lat52 , YAO и CDC45 ). Было обнаружено, что промотор DD45 повысил частоту замен последовательностей. Промотор яйцеклеток ( EC1 ) также использовался для эффективного нацеливания генов (GT) [ 212 ]. Гомозиготные двойные мутанты try cpc (TRIPTYCHON и CAPRICE) были обнаружены в поколении T1 с помощью pWUS и показали улучшенную эффективность редактирования по сравнению с промотором EC1.2 [ 213 ].
Из-за плейотропии генов традиционные системы редактирования не могут эффективно анализировать функцию генов в конкретных тканях. Поэтому Декаестекер и др. [ 214 ] предложили систему тканеспецифического нокаута на основе CRISPR (CRISPR-TSKO), которая может генерировать соматические мутации в определенных клетках, тканях и органах. В томатах редактирование генома для специфичного для плода гена с использованием промотора фосфоенолпируваткарбоксилазы 2 ( PPC2 ) оказалось эффективным инструментом, помогающим обойти плейотропные эффекты генов [ 215 ]. Кроме того, пыльца хлопка была трансформирована векторами экспрессии Cas9, управляемыми промоторами GhPLIM2b и GhMYB24 с помощью вакуумной инфильтрации Agrobacterium , и целевое редактирование генов в пыльце хлопка было успешно достигнуто [ 216 ]. С другой стороны, растения сидячие и полагаются на множество сигналов и защитных механизмов для реагирования на внешнюю среду. Многие гены участвуют в этих механизмах, и большинство из них имеют важные биологические функции для роста и развития растений [ 22 ]. Нокаут этих генов, вероятно, окажет значительное влияние на морфологию и урожайность, в то время как тканеспецифический нокаут будет отличным способом выяснить основные механизмы защитных реакций [ 22 , 23 ]. Более того, для многих описанных негативных регуляторных генов можно использовать тканеспецифическое редактирование генома (TSGE) для оптимизации их применения. Применение CRISPR-TSGE требует промоутеров, которые хорошо функционируют в различных тканях, и было описано много тканеспецифических промоутеров и нескольких систем TSGE, которые обеспечивают руководство для этих применений [23]. Другим важным применением TSGE является повышение устойчивости к патогенам. Вирусы систематически заражают и реплицируются в определенных тканях растений, используя вирус-индуцируемые или тканеспецифические промоторы, где вирусы реплицируются для экспрессии Cas9 для локально нокаутированных партнеров по взаимодействию растений, может ограничивать систематическую передачу и репликацию вирусов [ 22 ]. В сочетании с BE, PE и другими инструментами TSGE может быть использован более эффективно.
4.2 Ускорение получения растений без трансгенов
Использование системы CRISPR–Cas9 привело к революционным улучшениям в сельскохозяйственных культурах [ 1 ]. Однако из-за существования трансгенов постоянная редактирующая активность белка Cas9 затрудняет различение унаследованных и вновь генерируемых мутаций и увеличивает нецелевые эффекты. Это ограничение создает проблему для его дальнейшего применения, а оценка наследуемости и фенотипической стабильности растений, отредактированных с помощью CRISPR, требует удаления конструкций [ 217 ]. Кроме того, удаление этих конструкций может быть предпосылкой для коммерческих применений отредактированных сельскохозяйственных культур. ПЦР-амплификация является распространенным способом выделения растений без трансгенов из потомства, но этот процесс занимает много времени и трудоемок. Широко используемые репортерные гены, такие как mCherry, GFP и его производное RFP, можно добавлять в конструкции Cas9 [ 218 , 219 ], что позволяет легко отличать растения, содержащие трансгены, от растений без них, но для этих флуоресцентных маркеров требуются дополнительные инструменты. Репортерный ген β-глюкуронидазы (GUS) также широко используется в растениях, но для него требуются дорогостоящие субстраты [ 220 ].
Появление RUBY, который преобразует тирозин в яркий красный беталаин, делает присутствие трансгенов четко видимым невооруженным глазом [ 221 , 222 ]. В кукурузе была разработана система ViMeBox, которая усиливает экспрессию DsRED2, и через тканеспецифичные промоторы делает семена, содержащие Cas9, видимыми невооруженным глазом при естественном освещении [ 223 ]. Аналогичным образом, в кукурузе была установлена система репортера флуоресценции семян (SFR), которая экспрессирует DsRED в эндосперме, зародыше или обеих тканях [ 224 ]. Более того, одна система совместной экспрессии, Cas9-PF, экспрессировала ПРОДУКЦИЮ АНТОЦИАНОВЫХ ПИГМЕНТОВ 1 ( PAP1 ) и Цветущий локус T ( FT ) через промоторы вируса желтого скручивания листьев цеструма ( CmYLCV ) и убиквитина 10 арабидопсиса ( AtUbi10 ) соответственно. Эта система может ускорить создание целевых мутантов и растений без трансгенов [ 225 ]. Другие морфогенетические гены, такие как GLABRA1 ( GL1 ) и GL2 , играют роль в формировании трихом, а мутации в этих генах приводят к потере волосков на листьях и стеблях; была создана система, основанная на видимом отборе на основе мутаций GL2 (GBVS) [ 226 ]. Эти маркерные гены могут управляться одним промотором или быть связаны в тандеме с белком Cas9 через пептид P2A, чтобы указывать на уровень экспрессии белка Cas9 [ 219 ].
Добавление репортерных генов обеспечивает удобный способ различать растения с трансгенами и без них, а прямое устранение трансгенного потомства с помощью генов суицида или генов MS оказало бы значительное влияние на использование технологии CRISPR для улучшения сельскохозяйственных культур. Вводя кассеты экспрессии REG2-BARNASE и 35S-CMS2 в обычный вектор CRISPR–Cas9, Хе и др. [ 217 ] сконструировали систему Transgene Killer CRISPR (TKC); когда растение достигало репродуктивной стадии, токсичные белки убивали мужские гаметофиты и эмбрионы, содержащие конструкции. Однако конкретные применения редактирования генома, такие как редактирование с индукцией гаплоидов, требуют сохранения трансгенов [ 227 , 228 ]. Для кукурузы была предложена система самоэлиминации пыльцы CRISPR–Cas (PSEC), а для управления геном альфа-амилазы AA1 с целью предотвращения передачи трансгена использовался специфичный для пыльцы промотор ( полигалактуроназа 47 , ZmPG47 ) [ 26 ]. Система CRISPR–Cas9 с использованием флуоресцентного маркера и убийц пыльцы (FMPKC) использовала DsRed2 для различения трансгенного потомства и убийц пыльцы ( RAmy1A или orfH79 ) с целью снижения доли трансгенных семян [ 27 ], устанавливая баланс между отделением семян без трансгенов и получением трансгенных семян.
4.3. Применение морфогенетических генов
Для большинства растений культивирование тканей, опосредованное Agrobacterium , по-прежнему является основным методом регенерации, а высокая эффективность регенерации является важной гарантией для достижения целевого редактирования. Активация гена AP2/ERF TF WOUND INDUCED DEDIFFERENTIATION 1 ( WIND1 ) значительно способствовала регенерации почек у Brassica napus и Arabidopsis , но подавляла рост листьев [ 229 ]. Было показано, что Baby boom ( BBM ) и WUS повышают эффективность регенерации у многих видов, включая Zea mays , Sorghum bicolor , Saccharum officinarum , Oryza sativa , Malus domestica и т. д. [ 230 , 231 , 232 ], но эти гены также могут вызывать некоторые аномальные фенотипы, такие как дисплазия, искажение и снижение фертильности. Гены фактора регуляции роста ( GRF ) относятся к небольшому семейству специфичных для растений ТФ, которые образуют транскрипционный комплекс с взаимодействующим с GRF фактором (GIF), который играет роль в регуляции роста и развития растений [ 233 , 234 ]. Сверхэкспрессия GRF5 улучшила эффективность трансформации двудольных и однодольных видов, таких как рапс, соя, подсолнечник и кукуруза [ 235 ]. Сверхэкспрессия пшеничного белка слияния GRF4-GIF1 значительно улучшила эффективность и скорость регенерации у пшеницы, тритикале, риса и даже двудольных цитрусовых [ 236 ]. Трансгенные растения были фертильными без очевидных дефектов развития, и эффективная регенерация могла быть вызвана без экзогенных цитокининов [ 236 ]. Сравнение нескольких регуляторов развития (DR) в томате, включая PLETHORA 5 ( PLT5 ), WIND1 , ENHANCED SHOOT REGENERATION 1 ( ESR1 ), WUS и слияние WUS и BBM ( WUS - P2A - BBM ), показало, что растения, сверхэкспрессирующие PLT5, имели самую высокую эффективность регенерации. PLT5 также способствовал эффективности регенерации у B. rapa и Capsicum annum , но также наблюдались некоторые морфологические изменения [ 237 ]. Недавно были обнаружены некоторые гены, которые могут повышать эффективность регенерации, включая WOX5 , DOF5.6, DOF3.4 и LAX1 были обнаружены в пшенице, некоторые из которых также эффективны в кукурузе, сое и ячмене [ 25 , 238 , 239 ]. Несколько генов, включая LAX2 , LAX1 и LOX3 , также были обнаружены в хлопке с помощью технологии секвенирования отдельных клеток в сочетании с генетическим анализом [ 240 ].
Открытие этих морфогенетических генов дает рекомендации по высокоэффективной регенерации растений. Для генов, которые могут вызывать неблагоприятный рост, можно использовать слабые промоторы или тандемные репортерные гены для снижения негативных фенотипов. Условная система Cre/loxP SSR также может использоваться со специфическими обработками для устранения неблагоприятных факторов после формирования почек [ 241 ]. Помимо обычных фитогормонов, аденозинмонофосфат (АМФ) также обладает способностью повышать эффективность регенерации [ 242 ]. С другой стороны, несколько исследовательских групп применили тройную векторную систему, в которой морфогенетические гены добавляются для дальнейшего повышения эффективности регенерации сорго и кукурузы [ 24 , 241 , 243 ].
5. Система CRISPR–Cas и методы селекции растений
5.1. Обход культуры тканей или отсутствие трансгенов
Хотя было обнаружено, что некоторые DR способствуют регенерации растений, процесс культивирования тканей по-прежнему сложен и в определенной степени зависит от генотипа. Поэтому было предложено несколько новых стратегий трансформации. Agrobacterium , несущий DR и кассеты sgRNA, был введен в растения со сверхэкспрессией Cas9, из которых была удалена меристема, и растения с отредактированным геном были получены непосредственно из сгенерированных почек в N. benthamiana , томате, картофеле и винограде [ 244 ]. Поскольку вирусы растений обладают способностью перемещаться между клетками [ 29 ], некоторые вирусы РНК с положительной цепью, такие как вирус погремушки табака (TRV) и вирус мозаики полос ячменя (BSMV), были использованы для доставки sgRNA в клетки растений со сверхэкспрессией Cas9 [ 245 , 246 , 247 , 248 , 249 ]. В качестве альтернативы, кассеты Cas9 и sgRNA могут быть одновременно вставлены в вирусы РНК с отрицательной цепью, включая вирус желтой полосатой мозаики ячменя (BYSMV) или рабдовирус желтой сети Sonchus (SYNV) [ 250 , 251 ], все из которых успешно достигают целевого редактирования. С другой стороны, последовательности РНК, такие как тРНК и FT, обладают способностью перемещаться на большие расстояния внутри растения [ 248 ]; основываясь на этой характеристике, реагенты CRISPR могут быть перенесены с подвоя на привой путем прививки. Исследователи создали подвой, содержащий последовательности, подобные Cas9-тРНК (TLS) и слияния gRNA-TLS, а в привоях, привитых на подвой, были обнаружены целевые делеции у потомков [ 252 ]. Однако изготовление подвоя является этапом, ограничивающим скорость, и реализация этого подхода на однодольных растениях является сложной задачей. Кроме того, наночастицы также рассматриваются как новые векторы доставки для редактирования генома, но они ограничены размером нуклеаз [ 28 ].
Существует три типа реагентов для доставки CRISPR–Cas9: экспрессионные плазмиды (ДНК), транскрипты in vitro (IVT) и предварительно собранные комплексы РНП, которые включают белки Cas9 и направляющую РНК [ 19 ]. Среди них РНП быстро деградируют после попадания в клетку, обеспечивая мгновенную экспрессию нуклеаз Cas и снижая нецелевые эффекты [ 253 ]. Трансформация РНП с помощью полиэтиленгликоля (ПЭГ) использовалась во многих важных видах растений, включая рис, пшеницу, кукурузу, виноград и яблоко [ 254 , 255 , 256 ]. Трансфицированные протопласты использовались для получения регенерированных растений с различной эффективностью редактирования у разных видов [ 257 ]. Учитывая потенциальную токсичность ПЭГ, также использовались несколько других методов трансформации, включая липофекцию, бомбардировку частицами и электропорацию [ 258 , 259 , 260 ].
5.2. Гаплоидная индукция
Гаплоидная селекция может стабилизировать генетический фон гибридов за два поколения, в то время как традиционный метод требует от шести до восьми поколений, и ключевым фактором для его успешной реализации является эффективная гаплоидная индукция (HI) [ 9 ]. Мутации со сдвигом рамки считывания в гене MATRILINEAL [ MTL , также известный как NOT LIKE DAD ( NLD ) или PHOSPHOLIPASE A ( PLA1 )], кодирующем специфичную для пыльцы фосфолипазу, могут вызвать элиминацию патрилинейных хромосом в зиготах [ 261 , 262 , 263 ], что приводит к образованию гаплоидных зародышей кукурузы и далее распространяется на рис и пшеницу [ 264 , 265 ]. Более того, было также показано , что мембранный белок домена неизвестной функции 679 ( DMP ), фосфолипаза D3 ( PLD3 ) и POD65 запускают материнскую HI [ 266 , 267 , 268 , 269 ]. У Arabidopsis инженерия вариантов центромер-специфического гистона 3 ( CENH3 ) может запустить эффективную отцовскую HI [ 270 ], что также применялось к пшенице [ 271 ]. Новые системы, которые сочетают методы HI с CRISPR–Cas9, такие как HI-редактирование, редактирование генома с помощью индукторов гаплоидов (IMGE) и редактирование генов с последующим образованием удвоенных гаплоидов (GEDH) [ 227 , 228 , 269 ], могут преодолеть барьеры, связанные с зависимостью от генотипа, и достичь быстрой гомозиготности.
5.3. Рассеянный склероз и манипуляция самонесовместимостью
Благодаря гибридной силе гибридизация является важным способом повышения урожайности и качества в селекции сельскохозяйственных культур. Для производства коммерческих гибридных семян необходимо избегать самоопыления, а создание линий MS является практичным и эффективным методом. Хотя было зарегистрировано много линий MS, перенос признаков MS в элитные сорта традиционными методами требует много времени и усилий [ 1 ]. Система CRISPR–Cas предлагает быстрый метод создания MS. Редактирование генов мужской стерильности 1 ( MS1 , кодирующего белок-переносчик липидов, закрепленный на гликозилфосфатидилинозитоле) и MS45 (кодирующего фермент, подобный стриктозидинсинтазе) у пшеницы, SlSTR1 у томата и других генов, связанных с MS [ 272 , 273 , 274 ], может быстро производить зародышевую плазму MS; и эти стратегии могут быть распространены на другие виды [ 275 ]. Кроме того, путем воздействия на ген термочувствительной генной мужской стерильности (TGMS) TMS5 или ген фотопериодически-чувствительной генной мужской стерильности (PGMS) CARBON-STARVED ANTHERS ( CSA ) у риса были созданы более гибкие линии MS [ 275 ].
Самонесовместимость также широко распространена у цветковых растений, включая диплоидный картофель, рапс и капусту. Эта характеристика способствует перекрестному опылению для поддержания высокого уровня гетерозиготности внутри популяции и снижает наличие вредных гомозиготных рецессивных генов [ 9 ]. Однако у этих видов трудно получить стабильные гомозиготные инбредные линии, что затрудняет фундаментальные исследования и селекцию сельскохозяйственных культур. S-РНКаза контролирует самонесовместимость у пасленовых, и самосовместимый картофель был создан путем редактирования S-РНКазы [ 276 ]. В то время как ген фарнезилпирофосфатсинтазы ( FPS2 ) функционирует при S-РНКазо-независимой односторонней несовместимости, редактирование FPS2 может восстановить самонесовместимость [ 277 ]. У рапса и капусты самонесовместимость подавляется редактированием генов протеинкиназы М-локуса ( MLPK ) и рецепторной киназы S-рецептора ( SRK ) соответственно [ 278 , 279 ]. Более того, ожидается, что выяснение механизма самонесовместимости растений позволит преодолеть барьер межвидового размножения [ 1 ].
5.4. Новое одомашнивание
Селективное разведение для получения высокой урожайности и высокого качества привело к потере генетического разнообразия во многих одомашненных культурах, оставив их уязвимыми для биологических и абиотических стрессов, в то время как дикие виды сохраняют разнообразный генофонд и могут иметь врожденную устойчивость к неблагоприятным условиям [ 280 ]. При адекватном анализе генома можно добиться быстрого одомашнивания. Одомашнивание de novo, основанное на редактировании генома, достигло хорошего баланса между производительностью и адаптацией к окружающей среде. Орфанные культуры, которые выращиваются и потребляются в ограниченных регионах и имеют нежелательные характеристики, похожие на их диких сородичей, также являются важными материалами для одомашнивания de novo [ 281 , 282 , 283 , 284 ]. Эта стратегия была использована для достижения быстрого улучшения нескольких признаков у вишни сизой ( Physalis pruinosa ), дикого томата и аллополиплоидного риса путем одновременного редактирования нескольких генов [ 285 , 286 , 287 , 288 ]. Дикие виды или редкие культуры других растений, такие как киноа ( Chenopodium quinoa ), дикий картофель ( Solanum spp.) и люцерна ( Medicago sativa ), также имеют выраженные сильные и слабые стороны, а целевое воздействие на гены, регулирующие нежелательные признаки, может способствовать улучшению видов растений [ 1 ]. Системы трансформации диких видов и точная идентификация регуляторных генов для целевых признаков стали основой для эффективного использования одомашнивания de novo.
6. Выводы и перспективы на будущее
Постоянное распознавание белков Cas, повышение эффективности нацеливания, расширение широты нацеливания и оптимизация вспомогательных мер позволяют видеть, что постоянное совершенствование системы CRISPR–Cas обеспечивает надежную поддержку ее эффективного применения в растениях, а сочетание различных классификаций, представленных в данном обзоре, может еще больше способствовать ее применению. В будущем развитие систем CRISPR можно разделить на два аспекта: дальнейшее совершенствование и интеграция существующих систем в большее количество видов.
Для эффективного применения системы CRISPR в растениях, помимо всестороннего изучения системы CRISPR, также необходимы грамотное проектирование и модификация в соответствии с экспериментальными задачами. Использование CRISPR в сочетании с регуляторными элементами, мультиомикой или инструментами анализа белков может привести к появлению новых приложений. Кроме того, появляется всё больше инструментов для идентификации и модификации компонентов систем CRISPR. С другой стороны, хотя системы CRISPR занимают видное место во многих трансгенных технологиях и было получено множество превосходных линий, для массового производства этих превосходных линий всё ещё необходимы существенные исследования. Соответствующая политика надзора и общественное признание являются важными факторами для выхода этих систем на рынок. Полное определение стабильности редактируемых линий и создание систем CRISPR с более точными и меньшими нецелевыми эффектами – это цели, к которым исследователи должны прилагать постоянные усилия.
Широкомасштабное применение CRISPR остаётся важным подходом для быстрого понимания функций генов, особенно для видов с отстающими фундаментальными исследованиями. Однако платформы, позволяющей получить все или большинство белков Cas или их вариантов, не существует. Поэтому тем, кто впервые хочет исследовать эту область, относительно сложно получить необходимые реагенты. В будущем может потребоваться платформа для сбора опубликованной информации, которая будет способствовать эффективному применению системы CRISPR-Cas в растениях. С другой стороны, ожидается, что сочетание скрининга CRISPR и CRISPRa может улучшить понимание функций генов. Скрининг CRISPR, ориентированный на определённые семейства генов или определённые признаки, обладает уникальными преимуществами. В связи с важностью полногеномного скрининга CRISPR для вида растений, для получения как можно большего количества регенерированных растений и представления их общественности в виде базы данных могут потребоваться скоординированные усилия многих лабораторий. В будущем эта база данных может стать основой для нового исследования.
Ссылки
1. Zhu, H.; Li, C.; Gao, C. Applications of CRISPR–Cas in agriculture and plant biotechnology. Nat. Rev. Mol. Cell Biol. 2020, 21, 661–677. [Google Scholar] [CrossRef] [PubMed]
2. Li, W.; Teng, F.; Li, T.; Zhou, Q. Simultaneous generation and germline transmission of multiple gene mutations in rat using CRISPR-Cas systems. Nat. Biotechnol. 2013, 31, 684–686. [Google Scholar] [CrossRef] [PubMed]
3. Shan, Q.; Wang, Y.; Li, J.; Zhang, Y.; Chen, K.; Liang, Z.; Zhang, K.; Liu, J.; Xi, J.J.; Qiu, J.-L.; et al. Targeted genome modification of crop plants using a CRISPR-Cas system. Nat. Biotechnol. 2013, 31, 686–688. [Google Scholar] [CrossRef]
4. Cardi, T.; Murovec, J.; Bakhsh, A.; Boniecka, J.; Bruegmann, T.; Bull, S.E.; Eeckhaut, T.; Fladung, M.; Galovic, V.; Linkiewicz, A.; et al. CRISPR/Cas-mediated plant genome editing: Outstanding challenges a decade after implementation. Trends Plant Sci. 2023, 28, 1144–1165. [Google Scholar] [CrossRef]
5. Zhang, Y.; Malzahn, A.A.; Sretenovic, S.; Qi, Y. The emerging and uncultivated potential of CRISPR technology in plant science. Nat. Plants 2019, 5, 778–794. [Google Scholar] [CrossRef]
6. Yan, W.X.; Hunnewell, P.; Alfonse, L.E.; Carte, J.M.; Keston-Smith, E.; Sothiselvam, S.; Garrity, A.J.; Chong, S.; Makarova, K.S.; Koonin, E.V.; et al. Functionally diverse type V CRISPR-Cas systems. Science 2019, 363, 88–91. [Google Scholar] [CrossRef] [PubMed]
7. Wada, N.; Osakabe, K.; Osakabe, Y. Expanding the plant genome editing toolbox with recently developed CRISPR–Cas systems. Plant Physiol. 2022, 188, 1825–1837. [Google Scholar] [CrossRef]
8. Anzalone, A.V.; Randolph, P.B.; Davis, J.R.; Sousa, A.A.; Koblan, L.W.; Levy, J.M.; Chen, P.J.; Wilson, C.; Newby, G.A.; Raguram, A.; et al. Search-and-replace genome editing without double-strand breaks or donor DNA. Nature 2019, 576, 149–157. [Google Scholar] [CrossRef]
9. Li, B.; Sun, C.; Li, J.; Gao, C. Targeted genome-modification tools and their advanced applications in crop breeding. Nat. Rev. Genet. 2024, 25, 603–622. [Google Scholar] [CrossRef] [PubMed]
10. Zhong, Z.; Sretenovic, S.; Ren, Q.; Yang, L.; Bao, Y.; Qi, C.; Yuan, M.; He, Y.; Liu, S.; Liu, X.; et al. Improving Plant Genome Editing with High-Fidelity xCas9 and Non-canonical PAM-Targeting Cas9-NG. Mol. Plant 2019, 12, 1027–1036. [Google Scholar] [CrossRef]
11. Ren, B.; Liu, L.; Li, S.; Kuang, Y.; Wang, J.; Zhang, D.; Zhou, X.; Lin, H.; Zhou, H. Cas9-NG Greatly Expands the Targeting Scope of the Genome-Editing Toolkit by Recognizing NG and Other Atypical PAMs in Rice. Mol. Plant 2019, 12, 1015–1026. [Google Scholar] [CrossRef]
12. Walton, R.T.; Christie, K.A.; Whittaker, M.N.; Kleinstiver, B.P. Unconstrained genome targeting with near-PAMless engineered CRISPR-Cas9 variants. Science 2020, 368, 290–296. [Google Scholar] [CrossRef]
13. Xu, Z.; Kuang, Y.; Ren, B.; Yan, D.; Yan, F.; Spetz, C.; Sun, W.; Wang, G.; Zhou, X.; Zhou, H. SpRY greatly expands the genome editing scope in rice with highly flexible PAM recognition. Genome Biol. 2021, 22, 6. [Google Scholar] [CrossRef]
14. Zinselmeier, M.H.; Casas-Mollano, J.A.; Cors, J.; Sychla, A.; Heinsch, S.C.; Voytas, D.F.; Smanski, M.J. Optimized dCas9 programmable transcriptional activators for plants. Plant Biotechnol. J. 2024, 22, 3202–3204. [Google Scholar] [CrossRef] [PubMed]
15. Pan, C.; Qi, Y. CRISPR-Combo–mediated orthogonal genome editing and transcriptional activation for plant breeding. Nat. Protoc. 2023, 18, 1760–1794. [Google Scholar] [CrossRef] [PubMed]
16. Pan, C.; Wu, X.; Markel, K.; Malzahn, A.A.; Kundagrami, N.; Sretenovic, S.; Zhang, Y.; Cheng, Y.; Shih, P.M.; Qi, Y. CRISPR–Act3.0 for highly efficient multiplexed gene activation in plants. Nat. Plants 2021, 7, 942–953. [Google Scholar] [CrossRef] [PubMed]
17. Wang, M.; Zhong, Z.; Gallego-Bartolomé, J.; Li, Z.; Feng, S.; Kuo, H.Y.; Kan, R.L.; Lam, H.; Richey, J.C.; Tang, L.; et al. A gene silencing screen uncovers diverse tools for targeted gene repression in Arabidopsis. Nat. Plants 2023, 9, 460–472. [Google Scholar] [CrossRef] [PubMed]
18. Schmitz, R.J.; Grotewold, E.; Stam, M. Cis-regulatory sequences in plants: Their importance, discovery, and future challenges. Plant Cell 2022, 34, 718–741. [Google Scholar] [CrossRef] [PubMed]
19. Zhang, R.-X.; Li, B.-B.; Yang, Z.-G.; Huang, J.-Q.; Sun, W.-H.; Bhanbhro, N.; Liu, W.-T.; Chen, K.-M. Dissecting Plant Gene Functions Using CRISPR Toolsets for Crop Improvement. J. Agric. Food Chem. 2022, 70, 7343–7359. [Google Scholar] [CrossRef]
20. Liu, T.; Zhang, X.; Li, K.; Yao, Q.; Zhong, D.; Deng, Q.; Lu, Y. Large-scale genome editing in plants: Approaches, applications, and future perspectives. Curr. Opin. Biotechnol. 2023, 79, 102875. [Google Scholar] [CrossRef] [PubMed]
21. Pan, C.; Li, G.; Bandyopadhyay, A.; Qi, Y. Guide RNA library-based CRISPR screens in plants: Opportunities and challenges. Curr. Opin. Biotechnol. 2023, 79, 102883. [Google Scholar] [CrossRef] [PubMed]
22. Ali, Z.; Mahfouz, M.M.; Mansoor, S. CRISPR-TSKO: A Tool for Tissue-Specific Genome Editing in Plants. Trends Plant Sci. 2020, 25, 123–126. [Google Scholar] [CrossRef]
23. Singha, D.L.; Das, D.; Sarki, Y.N.; Chowdhury, N.; Sharma, M.; Maharana, J.; Chikkaputtaiah, C. Harnessing tissue-specific genome editing in plants through CRISPR/Cas system: Current state and future prospects. Planta 2021, 255, 28. [Google Scholar] [CrossRef]
24. Li, J.; Pan, W.; Zhang, S.; Ma, G.; Li, A.; Zhang, H.; Liu, L. A rapid and highly efficient sorghum transformation strategy using GRF4-GIF1/ternary vector system. Plant J. 2023, 117, 1604–1613. [Google Scholar] [CrossRef]
25. Wang, K.; Shi, L.; Liang, X.; Zhao, P.; Wang, W.; Liu, J.; Chang, Y.; Hiei, Y.; Yanagihara, C.; Du, L.; et al. The gene TaWOX5 overcomes genotype dependency in wheat genetic transformation. Nat. Plants 2022, 8, 110–117. [Google Scholar] [CrossRef] [PubMed]
26. Wang, H.; Qi, X.; Zhu, J.; Liu, C.; Fan, H.; Zhang, X.; Li, X.; Yang, Q.; Xie, C. Pollen self-elimination CRISPR–Cas genome editing prevents transgenic pollen dispersal in maize. Plant Commun. 2023, 4, 100637. [Google Scholar] [CrossRef] [PubMed]
27. Yu, D.; Zhou, T.; Xu, N.; Sun, X.; Song, S.; Liu, H.; Sun, Z.; Lv, Q.; Chen, J.; Tan, Y.; et al. Novel CRISPR/Cas9 system assisted by fluorescence marker and pollen killer for high-efficiency isolation of transgene-free edited plants in rice. Plant Biotechnol. J. 2024, 22, 1649–1651. [Google Scholar] [CrossRef]
28. Demirer, G.S.; Silva, T.N.; Jackson, C.T.; Thomas, J.B.; Ehrhardt, D.W.; Rhee, S.Y.; Mortimer, J.C.; Landry, M.P. Nanotechnology to advance CRISPR–Cas genetic engineering of plants. Nat. Nanotechnol. 2021, 16, 243–250. [Google Scholar] [CrossRef]
29. Shen, Y.; Ye, T.; Li, Z.; Kimutai, T.H.; Song, H.; Dong, X.; Wan, J. Exploiting viral vectors to deliver genome editing reagents in plants. aBIOTECH 2024, 5, 247–261. [Google Scholar] [CrossRef]
30. Pacesa, M.; Pelea, O.; Jinek, M. Past, present, and future of CRISPR genome editing technologies. Cell 2024, 187, 1076–1100. [Google Scholar] [CrossRef]
31. Makarova, K.S.; Wolf, Y.I.; Iranzo, J.; Shmakov, S.A.; Alkhnbashi, O.S.; Brouns, S.J.J.; Charpentier, E.; Cheng, D.; Haft, D.H.; Horvath, P.; et al. Evolutionary classification of CRISPR–Cas systems: A burst of class 2 and derived variants. Nat. Rev. Microbiol. 2019, 18, 67–83. [Google Scholar] [CrossRef]
32. Piatek, A.; Ali, Z.; Baazim, H.; Li, L.; Abulfaraj, A.; Al-Shareef, S.; Aouida, M.; Mahfouz, M.M. RNA-guided transcriptional regulation in planta via synthetic dCas9-based transcription factors. Plant Biotechnol. J. 2014, 13, 578–589. [Google Scholar] [CrossRef] [PubMed]
33. Lowder, L.G.; Zhang, D.; Baltes, N.J.; Paul, J.W.; Tang, X.; Zheng, X.; Voytas, D.F.; Hsieh, T.-F.; Zhang, Y.; Qi, Y. A CRISPR/Cas9 Toolbox for Multiplexed Plant Genome Editing and Transcriptional Regulation. Plant Physiol. 2015, 169, 971–985. [Google Scholar] [CrossRef] [PubMed]
34. Gaudelli, N.M.; Komor, A.C.; Rees, H.A.; Packer, M.S.; Badran, A.H.; Bryson, D.I.; Liu, D.R. Programmable base editing of A•T to G•C in genomic DNA without DNA cleavage. Nature 2017, 551, 464–471. [Google Scholar] [CrossRef]
35. Li, C.; Zong, Y.; Jin, S.; Zhu, H.; Lin, D.; Li, S.; Qiu, J.-L.; Wang, Y.; Gao, C. SWISS: Multiplexed orthogonal genome editing in plants with a Cas9 nickase and engineered CRISPR RNA scaffolds. Genome Biol. 2020, 21, 141. [Google Scholar] [CrossRef]
36. Kavuri, N.R.; Ramasamy, M.; Qi, Y.; Mandadi, K. Applications of CRISPR/Cas13-Based RNA Editing in Plants. Cells 2022, 11, 2665. [Google Scholar] [CrossRef] [PubMed]
37. Zetsche, B.; Gootenberg, J.S.; Abudayyeh, O.O.; Slaymaker, I.M.; Makarova, K.S.; Essletzbichler, P.; Volz, S.E.; Joung, J.; van der Oost, J.; Regev, A.; et al. Cpf1 Is a Single RNA-Guided Endonuclease of a Class 2 CRISPR-Cas System. Cell 2015, 163, 759–771. [Google Scholar] [CrossRef]
38. Wang, M.; Mao, Y.; Lu, Y.; Tao, X.; Zhu, J.-K. Multiplex Gene Editing in Rice Using the CRISPR-Cpf1 System. Mol. Plant 2017, 10, 1011–1013. [Google Scholar] [CrossRef]
39. Fonfara, I.; Richter, H.; Bratovič, M.; Le Rhun, A.; Charpentier, E. The CRISPR-associated DNA-cleaving enzyme Cpf1 also processes precursor CRISPR RNA. Nature 2016, 532, 517–521. [Google Scholar] [CrossRef] [PubMed]
40. Zetsche, B.; Heidenreich, M.; Mohanraju, P.; Fedorova, I.; Kneppers, J.; DeGennaro, E.M.; Winblad, N.; Choudhury, S.R.; Abudayyeh, O.O.; Gootenberg, J.S.; et al. Multiplex gene editing by CRISPR–Cpf1 using a single crRNA array. Nat. Biotechnol. 2016, 35, 31–34. [Google Scholar] [CrossRef]
41. Yamano, T.; Nishimasu, H.; Zetsche, B.; Hirano, H.; Slaymaker, I.M.; Li, Y.; Fedorova, I.; Nakane, T.; Makarova, K.S.; Koonin, E.V.; et al. Crystal Structure of Cpf1 in Complex with Guide RNA and Target DNA. Cell 2016, 165, 949–962. [Google Scholar] [CrossRef] [PubMed]
42. Tang, X.; Lowder, L.G.; Zhang, T.; Malzahn, A.A.; Zheng, X.; Voytas, D.F.; Zhong, Z.; Chen, Y.; Ren, Q.; Li, Q.; et al. A CRISPR–Cpf1 system for efficient genome editing and transcriptional repression in plants. Nat. Plants 2017, 3, 17018. [Google Scholar] [CrossRef] [PubMed]
43. Singh, D.; Mallon, J.; Poddar, A.; Wang, Y.; Tippana, R.; Yang, O.; Bailey, S.; Ha, T. Real-time observation of DNA target interrogation and product release by the RNA-guided endonuclease CRISPR Cpf1 (Cas12a). Proc. Natl. Acad. Sci. USA 2018, 115, 5444–5449. [Google Scholar] [CrossRef]
44. Malzahn, A.A.; Tang, X.; Lee, K.; Ren, Q.; Sretenovic, S.; Zhang, Y.; Chen, H.; Kang, M.; Bao, Y.; Zheng, X.; et al. Application of CRISPR-Cas12a temperature sensitivity for improved genome editing in rice, maize, and Arabidopsis. BMC Biol. 2019, 17, 9. [Google Scholar] [CrossRef] [PubMed]
45. Hu, X.; Meng, X.; Li, J.; Wang, K.; Yu, H. Improving the efficiency of the CRISPR-Cas12a system with tRNA-crRNA arrays. Crop J. 2020, 8, 403–407. [Google Scholar] [CrossRef]
46. Li, G.; Zhang, Y.; Dailey, M.; Qi, Y. Hs1Cas12a and Ev1Cas12a confer efficient genome editing in plants. Front. Genome Ed. 2023, 5, 1251903. [Google Scholar] [CrossRef]
47. Zhang, L.; Li, G.; Zhang, Y.; Cheng, Y.; Roberts, N.; Glenn, S.E.; DeZwaan-McCabe, D.; Rube, H.T.; Manthey, J.; Coleman, G.; et al. Boosting genome editing efficiency in human cells and plants with novel LbCas12a variants. Genome Biol. 2023, 24, 102. [Google Scholar] [CrossRef] [PubMed]
48. Shmakov, S.; Abudayyeh, O.O.; Makarova, K.S.; Wolf, Y.I.; Gootenberg, J.S.; Semenova, E.; Minakhin, L.; Joung, J.; Konermann, S.; Severinov, K.; et al. Discovery and Functional Characterization of Diverse Class 2 CRISPR-Cas Systems. Mol. Cell 2015, 60, 385–397. [Google Scholar] [CrossRef]
49. Yang, H.; Gao, P.; Rajashankar, K.R.; Patel, D.J. PAM-Dependent Target DNA Recognition and Cleavage by C2c1 CRISPR-Cas Endonuclease. Cell 2016, 167, 1814–1828. [Google Scholar] [CrossRef]
50. Wu, D.; Guan, X.; Zhu, Y.; Ren, K.; Huang, Z. Structural basis of stringent PAM recognition by CRISPR-C2c1 in complex with sgRNA. Cell Res. 2017, 27, 705–708. [Google Scholar] [CrossRef] [PubMed]
51. Teng, F.; Cui, T.; Feng, G.; Guo, L.; Xu, K.; Gao, Q.; Li, T.; Li, J.; Zhou, Q.; Li, W. Repurposing CRISPR-Cas12b for mammalian genome engineering. Cell Discov. 2018, 4, 63. [Google Scholar] [CrossRef]
52. Wang, P.; Zhang, J.; Sun, L.; Ma, Y.; Xu, J.; Liang, S.; Deng, J.; Tan, J.; Zhang, Q.; Tu, L.; et al. High efficient multisites genome editing in allotetraploid cotton (Gossypium hirsutum) using CRISPR/Cas9 system. Plant Biotechnol. J. 2017, 16, 137–150. [Google Scholar] [CrossRef] [PubMed]
53. Wang, Q.; Alariqi, M.; Wang, F.; Li, B.; Ding, X.; Rui, H.; Li, Y.; Xu, Z.; Qin, L.; Sun, L.; et al. The application of a heat-inducible CRISPR/Cas12b (C2c1) genome editing system in tetraploid cotton (G. hirsutum) plants. Plant Biotechnol. J. 2020, 18, 2436–2443. [Google Scholar] [CrossRef]
54. Ming, M.; Ren, Q.; Pan, C.; He, Y.; Zhang, Y.; Liu, S.; Zhong, Z.; Wang, J.; Malzahn, A.A.; Wu, J.; et al. CRISPR–Cas12b enables efficient plant genome engineering. Nat. Plants 2020, 6, 202–208. [Google Scholar] [CrossRef]
55. Gurel, F.; Wu, Y.; Pan, C.; Cheng, Y.; Li, G.; Zhang, T.; Qi, Y. On- and Off-Target Analyses of CRISPR-Cas12b Genome Editing Systems in Rice. CRISPR J. 2023, 6, 62–74. [Google Scholar] [CrossRef] [PubMed]
56. Wu, F.; Qiao, X.; Zhao, Y.; Zhang, Z.; Gao, Y.; Shi, L.; Du, H.; Wang, L.; Zhang, Y.J.; Zhang, Y.; et al. Targeted mutagenesis in Arabidopsis thaliana using CRISPR-Cas12b/C2c1. J. Integr. Plant Biol. 2020, 62, 1653–1658. [Google Scholar] [CrossRef] [PubMed]
57. Lv, P.; Su, F.; Chen, F.; Yan, C.; Xia, D.; Sun, H.; Li, S.; Duan, Z.; Ma, C.; Zhang, H.; et al. Genome editing in rice using CRISPR/Cas12i3. Plant Biotechnol. J. 2023, 22, 379–385. [Google Scholar] [CrossRef]
58. Pausch, P.; Al-Shayeb, B.; Bisom-Rapp, E.; Tsuchida, C.A.; Li, Z.; Cress, B.F.; Knott, G.J.; Jacobsen, S.E.; Banfield, J.F.; Doudna, J.A. CRISPR-CasΦ from huge phages is a hypercompact genome editor. Science 2020, 369, 333–337. [Google Scholar] [CrossRef] [PubMed]
59. Liu, S.; Sretenovic, S.; Fan, T.; Cheng, Y.; Li, G.; Qi, A.; Tang, X.; Xu, Y.; Guo, W.; Zhong, Z.; et al. Hypercompact CRISPR–Cas12j2 (CasΦ) enables genome editing, gene activation, and epigenome editing in plants. Plant Commun. 2022, 3, 100453. [Google Scholar] [CrossRef]
60. Li, Z.; Zhong, Z.; Wu, Z.; Pausch, P.; Al-Shayeb, B.; Amerasekera, J.; Doudna, J.A.; Jacobsen, S.E. Genome editing in plants using the compact editor CasΦ. Proc. Natl. Acad. Sci. USA 2023, 120, e2216822120. [Google Scholar] [CrossRef]
61. Ali, Z.; Mahas, A.; Mahfouz, M. CRISPR/Cas13 as a Tool for RNA Interference. Trends Plant Sci. 2018, 23, 374–378. [Google Scholar] [CrossRef] [PubMed]
62. Shmakov, S.; Smargon, A.; Scott, D.; Cox, D.; Pyzocha, N.; Yan, W.; Abudayyeh, O.O.; Gootenberg, J.S.; Makarova, K.S.; Wolf, Y.I.; et al. Diversity and evolution of class 2 CRISPR–Cas systems. Nat. Rev. Microbiol. 2017, 15, 169–182. [Google Scholar] [CrossRef] [PubMed]
63. Xu, C.; Zhou, Y.; Xiao, Q.; He, B.; Geng, G.; Wang, Z.; Cao, B.; Dong, X.; Bai, W.; Wang, Y.; et al. Programmable RNA editing with compact CRISPR–Cas13 systems from uncultivated microbes. Nat. Methods 2021, 18, 499–506. [Google Scholar] [CrossRef]
64. Abudayyeh, O.O.; Gootenberg, J.S.; Essletzbichler, P.; Han, S.; Joung, J.; Belanto, J.J.; Verdine, V.; Cox, D.B.T.; Kellner, M.J.; Regev, A.; et al. RNA targeting with CRISPR–Cas13. Nature 2017, 550, 280–284. [Google Scholar] [CrossRef] [PubMed]
65. Wolter, F.; Puchta, H. The CRISPR/Cas revolution reaches the RNA world: Cas13, a new Swiss Army knife for plant biologists. Plant J. 2018, 94, 767–775. [Google Scholar] [CrossRef]
66. Aman, R.; Mahas, A.; Butt, H.; Ali, Z.; Aljedaani, F.; Mahfouz, M. Engineering RNA Virus Interference via the CRISPR/Cas13 Machinery in Arabidopsis. Viruses 2018, 10, 732. [Google Scholar] [CrossRef] [PubMed]
67. Aman, R.; Ali, Z.; Butt, H.; Mahas, A.; Aljedaani, F.; Khan, M.Z.; Ding, S.; Mahfouz, M. RNA virus interference via CRISPR/Cas13a system in plants. Genome Biol. 2018, 19, 1. [Google Scholar] [CrossRef]
68. Zhan, X.; Zhang, F.; Zhong, Z.; Chen, R.; Wang, Y.; Chang, L.; Bock, R.; Nie, B.; Zhang, J. Generation of virus-resistant potato plants by RNA genome targeting. Plant Biotechnol. J. 2019, 17, 1814–1822. [Google Scholar] [CrossRef]
69. Zhang, T.; Zhao, Y.; Ye, J.; Cao, X.; Xu, C.; Chen, B.; An, H.; Jiao, Y.; Zhang, F.; Yang, X.; et al. Establishing CRISPR/Cas13a immune system conferring RNA virus resistance in both dicot and monocot plants. Plant Biotechnol. J. 2019, 17, 1185–1187. [Google Scholar] [CrossRef]
70. Yu, Y.; Pan, Z.; Wang, X.; Bian, X.; Wang, W.; Liang, Q.; Kou, M.; Ji, H.; Li, Y.; Ma, D.; et al. Targeting of SPCSV-RNase3 via CRISPR-Cas13 confers resistance against sweet potato virus disease. Mol. Plant Pathol. 2021, 23, 104–117. [Google Scholar] [CrossRef] [PubMed]
71. Konermann, S.; Lotfy, P.; Brideau, N.J.; Oki, J.; Shokhirev, M.N.; Hsu, P.D. Transcriptome Engineering with RNA-Targeting Type VI-D CRISPR Effectors. Cell 2018, 173, 665–676. [Google Scholar] [CrossRef]
72. Mahas, A.; Aman, R.; Mahfouz, M. CRISPR-Cas13d mediates robust RNA virus interference in plants. Genome Biol. 2019, 20, 263. [Google Scholar] [CrossRef]
73. Zhan, X.; Liu, W.; Nie, B.; Zhang, F.; Zhang, J. Cas13d-mediated multiplex RNA targeting confers a broad-spectrum resistance against RNA viruses in potato. Commun. Biol. 2023, 6, 855. [Google Scholar] [CrossRef]
74. Sarkar, J.; Jyoti, T.P.; Sahana, S.; Bhattacharya, A.; Chandel, S.; Singh, R. CRISPR–Cas13d in plant biology: An insight. Plant Biotechnol. Rep. 2024, 18, 301–311. [Google Scholar] [CrossRef]
75. Sharma, V.K.; Marla, S.; Zheng, W.; Mishra, D.; Huang, J.; Zhang, W.; Morris, G.P.; Cook, D.E. CRISPR guides induce gene silencing in plants in the absence of Cas. Genome Biol. 2022, 23, 6. [Google Scholar] [CrossRef] [PubMed]
76. Abudayyeh, O.O.; Gootenberg, J.S.; Kellner, M.J.; Zhang, F. Nucleic Acid Detection of Plant Genes Using CRISPR-Cas13. CRISPR J. 2019, 2, 165–171. [Google Scholar] [CrossRef]
77. Li, Z.; Zhang, D.; Xiong, X.; Yan, B.; Xie, W.; Sheen, J.; Li, J.-F. A potent Cas9-derived gene activator for plant and mammalian cells. Nat. Plants 2017, 3, 930–936. [Google Scholar] [CrossRef] [PubMed]
78. Papikian, A.; Liu, W.; Gallego-Bartolomé, J.; Jacobsen, S.E. Site-specific manipulation of Arabidopsis loci using CRISPR-Cas9 SunTag systems. Nat. Commun. 2019, 10, 729. [Google Scholar] [CrossRef]
79. Lowder, L.G.; Zhou, J.; Zhang, Y.; Malzahn, A.; Zhong, Z.; Hsieh, T.-F.; Voytas, D.F.; Zhang, Y.; Qi, Y. Robust Transcriptional Activation in Plants Using Multiplexed CRISPR-Act2.0 and mTALE-Act Systems. Mol. Plant 2018, 11, 245–256. [Google Scholar] [CrossRef]
80. Pan, C.; Li, G.; Malzahn, A.A.; Cheng, Y.; Leyson, B.; Sretenovic, S.; Gurel, F.; Coleman, G.D.; Qi, Y. Boosting plant genome editing with a versatile CRISPR-Combo system. Nat. Plants 2022, 8, 513–525. [Google Scholar] [CrossRef]
81. Morffy, N.; Van den Broeck, L.; Miller, C.; Emenecker, R.J.; Bryant, J.A.; Lee, T.M.; Sageman-Furnas, K.; Wilkinson, E.G.; Pathak, S.; Kotha, S.R.; et al. Identification of plant transcriptional activation domains. Nature 2024, 632, 166–173. [Google Scholar] [CrossRef]
82. Selma, S.; Sanmartín, N.; Espinosa-Ruiz, A.; Gianoglio, S.; Lopez-Gresa, M.P.; Vázquez-Vilar, M.; Flors, V.; Granell, A.; Orzaez, D. Custom-made design of metabolite composition in N. benthamiana leaves using CRISPR activators. Plant Biotechnol. J. 2022, 20, 1578–1590. [Google Scholar] [CrossRef] [PubMed]
83. Ren, C.; Li, H.; Liu, Y.; Li, S.; Liang, Z. Highly efficient activation of endogenous gene in grape using CRISPR/dCas9-based transcriptional activators. Hortic. Res. 2022, 9, uhab037. [Google Scholar] [CrossRef]
84. Ming, M.; Long, H.; Ye, Z.; Pan, C.; Chen, J.; Tian, R.; Sun, C.; Xue, Y.; Zhang, Y.; Li, J.; et al. Highly efficient CRISPR systems for loss-of-function and gain-of-function research in pear calli. Hortic. Res. 2022, 9, uhac148. [Google Scholar] [CrossRef] [PubMed]
85. Yao, T.; Yuan, G.; Lu, H.; Liu, Y.; Zhang, J.; Tuskan, G.A.; Muchero, W.; Chen, J.-G.; Yang, X. CRISPR/Cas9-based gene activation and base editing in Populus. Hortic. Res. 2023, 10, uhad085. [Google Scholar] [CrossRef] [PubMed]
86. García-Murillo, L.; Valencia-Lozano, E.; Priego-Ranero, N.A.; Cabrera-Ponce, J.L.; Duarte-Aké, F.P.; Vizuet-de-Rueda, J.C.; Rivera-Toro, D.M.; Herrera-Ubaldo, H.; de Folter, S.; Alvarez-Venegas, R. CRISPRa-mediated transcriptional activation of the SlPR-1 gene in edited tomato plants. Plant Sci. 2023, 329, 111617. [Google Scholar] [CrossRef] [PubMed]
87. Qi, X.; Gao, H.; Lv, R.; Mao, W.; Zhu, J.; Liu, C.; Mao, L.; Li, X.; Xie, C. CRISPR/dCas-mediated gene activation toolkit development and its application for parthenogenesis induction in maize. Plant Commun. 2023, 4, 100449. [Google Scholar] [CrossRef]
88. Yu, L.; Li, Z.; Ding, X.; Alariqi, M.; Zhang, C.; Zhu, X.; Fan, S.; Zhu, L.; Zhang, X.; Jin, S. Developing an efficient CRISPR–dCas9–TV-derived transcriptional activation system to create three novel cotton germplasm materials. Plant Commun. 2023, 4, 100600. [Google Scholar] [CrossRef] [PubMed]
89. Zhang, C.; Tang, Y.; Tang, S.; Chen, L.; Li, T.; Yuan, H.; Xu, Y.; Zhou, Y.; Zhang, S.; Wang, J.; et al. An inducible CRISPR activation tool for accelerating plant regeneration. Plant Commun. 2024, 5, 100823. [Google Scholar] [CrossRef]
90. Qi, L.S.; Larson, M.H.; Gilbert, L.A.; Doudna, J.A.; Weissman, J.S.; Arkin, A.P.; Lim, W.A. Repurposing CRISPR as an RNA-Guided Platform for Sequence-Specific Control of Gene Expression. Cell 2013, 152, 1173–1183. [Google Scholar] [CrossRef]
91. Gilbert, L.A.; Larson, M.H.; Morsut, L.; Liu, Z.; Brar, G.A.; Torres, S.E.; Stern-Ginossar, N.; Brandman, O.; Whitehead, E.H.; Doudna, J.A.; et al. CRISPR-Mediated Modular RNA-Guided Regulation of Transcription in Eukaryotes. Cell 2013, 154, 442–451. [Google Scholar] [CrossRef] [PubMed]
92. Babu, K.; Kathiresan, V.; Kumari, P.; Newsom, S.; Parameshwaran, H.P.; Chen, X.; Liu, J.; Qin, P.Z.; Rajan, R. Coordinated Actions of Cas9 HNH and RuvC Nuclease Domains Are Regulated by the Bridge Helix and the Target DNA Sequence. Biochemistry 2021, 60, 3783–3800. [Google Scholar] [CrossRef] [PubMed]
93. Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable Dual-RNA–Guided DNA Endonuclease in Adaptive Bacterial Immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
94. Komor, A.C.; Kim, Y.B.; Packer, M.S.; Zuris, J.A.; Liu, D.R. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature 2016, 533, 420–424. [Google Scholar] [CrossRef] [PubMed]
95. Kosicki, M.; Tomberg, K.; Bradley, A. Repair of double-strand breaks induced by CRISPR–Cas9 leads to large deletions and complex rearrangements. Nat. Biotechnol. 2018, 36, 765–771. [Google Scholar] [CrossRef]
96. Zong, Y.; Wang, Y.; Li, C.; Zhang, R.; Chen, K.; Ran, Y.; Qiu, J.-L.; Wang, D.; Gao, C. Precise base editing in rice, wheat and maize with a Cas9-cytidine deaminase fusion. Nat. Biotechnol. 2017, 35, 438–440. [Google Scholar] [CrossRef]
97. Shimatani, Z.; Kashojiya, S.; Takayama, M.; Terada, R.; Arazoe, T.; Ishii, H.; Teramura, H.; Yamamoto, T.; Komatsu, H.; Miura, K.; et al. Targeted base editing in rice and tomato using a CRISPR-Cas9 cytidine deaminase fusion. Nat. Biotechnol. 2017, 35, 441–443. [Google Scholar] [CrossRef] [PubMed]
98. Zong, Y.; Song, Q.; Li, C.; Jin, S.; Zhang, D.; Wang, Y.; Qiu, J.-L.; Gao, C. Efficient C-to-T base editing in plants using a fusion of nCas9 and human APOBEC3A. Nat. Biotechnol. 2018, 36, 950–953. [Google Scholar] [CrossRef] [PubMed]
99. Jin, S.; Fei, H.; Zhu, Z.; Luo, Y.; Liu, J.; Gao, S.; Zhang, F.; Chen, Y.-H.; Wang, Y.; Gao, C. Rationally Designed APOBEC3B Cytosine Base Editors with Improved Specificity. Mol. Cell 2020, 79, 728–740. [Google Scholar] [CrossRef] [PubMed]
100. Ren, Q.; Sretenovic, S.; Liu, G.; Zhong, Z.; Wang, J.; Huang, L.; Tang, X.; Guo, Y.; Liu, L.; Wu, Y.; et al. Improved plant cytosine base editors with high editing activity, purity, and specificity. Plant Biotechnol. J. 2021, 19, 2052–2068. [Google Scholar] [CrossRef] [PubMed]
101. Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
102. Huang, J.; Lin, Q.; Fei, H.; He, Z.; Xu, H.; Li, Y.; Qu, K.; Han, P.; Gao, Q.; Li, B.; et al. Discovery of deaminase functions by structure-based protein clustering. Cell 2023, 186, 3182–3195. [Google Scholar] [CrossRef] [PubMed]
103. Wang, S.; Zong, Y.; Lin, Q.; Zhang, H.; Chai, Z.; Zhang, D.; Chen, K.; Qiu, J.-L.; Gao, C. Precise, predictable multi-nucleotide deletions in rice and wheat using APOBEC–Cas9. Nat. Biotechnol. 2020, 38, 1460–1465. [Google Scholar] [CrossRef] [PubMed]
104. Li, C.; Zong, Y.; Wang, Y.; Jin, S.; Zhang, D.; Song, Q.; Zhang, R.; Gao, C. Expanded base editing in rice and wheat using a Cas9-adenosine deaminase fusion. Genome Biol. 2018, 19, 59. [Google Scholar] [CrossRef]
105. Kang, B.-C.; Yun, J.-Y.; Kim, S.-T.; Shin, Y.; Ryu, J.; Choi, M.; Woo, J.W.; Kim, J.-S. Precision genome engineering through adenine base editing in plants. Nat. Plants 2018, 4, 427–431. [Google Scholar] [CrossRef]
106. Jin, S.; Zong, Y.; Gao, Q.; Zhu, Z.; Wang, Y.; Qin, P.; Liang, C.; Wang, D.; Qiu, J.-L.; Zhang, F.; et al. Cytosine, but not adenine, base editors induce genome-wide off-target mutations in rice. Science 2019, 364, 292–295. [Google Scholar] [CrossRef] [PubMed]
107. Zhou, C.; Sun, Y.; Yan, R.; Liu, Y.; Zuo, E.; Gu, C.; Han, L.; Wei, Y.; Hu, X.; Zeng, R.; et al. Off-target RNA mutation induced by DNA base editing and its elimination by mutagenesis. Nature 2019, 571, 275–278. [Google Scholar] [CrossRef] [PubMed]
108. Hua, K.; Tao, X.; Liang, W.; Zhang, Z.; Gou, R.; Zhu, J.K. Simplified adenine base editors improve adenine base editing efficiency in rice. Plant Biotechnol. J. 2019, 18, 770–778. [Google Scholar] [CrossRef]
109. Lapinaite, A.; Knott, G.J.; Palumbo, C.M.; Lin-Shiao, E.; Richter, M.F.; Zhao, K.T.; Beal, P.A.; Liu, D.R.; Doudna, J.A. DNA capture by a CRISPR-Cas9–guided adenine base editor. Science 2020, 369, 566–571. [Google Scholar] [CrossRef] [PubMed]
110. Richter, M.F.; Zhao, K.T.; Eton, E.; Lapinaite, A.; Newby, G.A.; Thuronyi, B.W.; Wilson, C.; Koblan, L.W.; Zeng, J.; Bauer, D.E.; et al. Phage-assisted evolution of an adenine base editor with improved Cas domain compatibility and activity. Nat. Biotechnol. 2020, 38, 883–891. [Google Scholar] [CrossRef] [PubMed]
111. Wei, C.; Wang, C.; Jia, M.; Guo, H.X.; Luo, P.Y.; Wang, M.G.; Zhu, J.K.; Zhang, H. Efficient generation of homozygous substitutions in rice in one generation utilizing an rABE8e base editor. J. Integr. Plant Biol. 2021, 63, 1595–1599. [Google Scholar] [CrossRef] [PubMed]
112. Yan, D.; Ren, B.; Liu, L.; Yan, F.; Li, S.; Wang, G.; Sun, W.; Zhou, X.; Zhou, H. High-efficiency and multiplex adenine base editing in plants using new TadA variants. Mol. Plant 2021, 14, 722–731. [Google Scholar] [CrossRef] [PubMed]
113. Grünewald, J.; Zhou, R.; Lareau, C.A.; Garcia, S.P.; Iyer, S.; Miller, B.R.; Langner, L.M.; Hsu, J.Y.; Aryee, M.J.; Joung, J.K. A dual-deaminase CRISPR base editor enables concurrent adenine and cytosine editing. Nat. Biotechnol. 2020, 38, 861–864. [Google Scholar] [CrossRef]
114. Li, C.; Zhang, R.; Meng, X.; Chen, S.; Zong, Y.; Lu, C.; Qiu, J.-L.; Chen, Y.-H.; Li, J.; Gao, C. Targeted, random mutagenesis of plant genes with dual cytosine and adenine base editors. Nat. Biotechnol. 2020, 38, 875–882. [Google Scholar] [CrossRef]
115. Zheng, Z.; Liu, T.; Chai, N.; Zeng, D.; Zhang, R.; Wu, Y.; Hang, J.; Liu, Y.; Deng, Q.; Tan, J.; et al. PhieDBEs: A DBD-containing, PAM-flexible, high-efficiency dual base editor toolbox with wide targeting scope for use in plants. Plant Biotechnol. J. 2024, 22, 3164–3174. [Google Scholar] [CrossRef]
116. Zhang, A.; Shan, T.; Sun, Y.; Chen, Z.; Hu, J.; Hu, Z.; Ming, Z.; Zhu, Z.; Li, X.; He, J.; et al. Directed evolution rice genes with randomly multiplexed sgRNAs assembly of base editors. Plant Biotechnol. J. 2023, 21, 2597–2610. [Google Scholar] [CrossRef]
117. Fan, T.; Cheng, Y.; Wu, Y.; Liu, S.; Tang, X.; He, Y.; Liao, S.; Zheng, X.; Zhang, T.; Qi, Y.; et al. High performance TadA-8e derived cytosine and dual base editors with undetectable off-target effects in plants. Nat. Commun. 2024, 15, 5103. [Google Scholar] [CrossRef]
118. Chen, L.; Park, J.E.; Paa, P.; Rajakumar, P.D.; Prekop, H.-T.; Chew, Y.T.; Manivannan, S.N.; Chew, W.L. Programmable C:G to G:C genome editing with CRISPR-Cas9-directed base excision repair proteins. Nat. Commun. 2021, 12, 1384. [Google Scholar] [CrossRef] [PubMed]
119. Kurt, I.C.; Zhou, R.; Iyer, S.; Garcia, S.P.; Miller, B.R.; Langner, L.M.; Grünewald, J.; Joung, J.K. CRISPR C-to-G base editors for inducing targeted DNA transversions in human cells. Nat. Biotechnol. 2020, 39, 41–46. [Google Scholar] [CrossRef] [PubMed]
120. Zhao, D.; Li, J.; Li, S.; Xin, X.; Hu, M.; Price, M.A.; Rosser, S.J.; Bi, C.; Zhang, X. Glycosylase base editors enable C-to-A and C-to-G base changes. Nat. Biotechnol. 2020, 39, 35–40. [Google Scholar] [CrossRef]
121. Tian, Y.; Shen, R.; Li, Z.; Yao, Q.; Zhang, X.; Zhong, D.; Tan, X.; Song, M.; Han, H.; Zhu, J.K.; et al. Efficient C-to-G editing in rice using an optimized base editor. Plant Biotechnol. J. 2022, 20, 1238–1240. [Google Scholar] [CrossRef]
122. Sretenovic, S.; Liu, S.; Li, G.; Cheng, Y.; Fan, T.; Xu, Y.; Zhou, J.; Zheng, X.; Coleman, G.; Zhang, Y.; et al. Exploring C-To-G Base Editing in Rice, Tomato, and Poplar. Front. Genome Ed. 2021, 3, 756766. [Google Scholar] [CrossRef] [PubMed]
123. Chen, L.; Hong, M.; Luan, C.; Gao, H.; Ru, G.; Guo, X.; Zhang, D.; Zhang, S.; Li, C.; Wu, J.; et al. Adenine transversion editors enable precise, efficient A•T-to-C•G base editing in mammalian cells and embryos. Nat. Biotechnol. 2023, 42, 638–650. [Google Scholar] [CrossRef]
124. Wu, X.; Ren, B.; Liu, L.; Qiu, S.; Li, X.g.; Li, P.; Yan, F.; Lin, H.; Zhou, X.; Zhang, D.; et al. Adenine base editor incorporating the N-methylpurine DNA glycosylase MPGv3 enables efficient A-to-K base editing in rice. Plant Commun. 2023, 4, 100668. [Google Scholar] [CrossRef] [PubMed]
125. Tong, H.; Wang, X.; Liu, Y.; Liu, N.; Li, Y.; Luo, J.; Ma, Q.; Wu, D.; Li, J.; Xu, C.; et al. Programmable A-to-Y base editing by fusing an adenine base editor with an N-methylpurine DNA glycosylase. Nat. Biotechnol. 2023, 41, 1080–1084. [Google Scholar] [CrossRef]
126. Lin, Q.; Zong, Y.; Xue, C.; Wang, S.; Jin, S.; Zhu, Z.; Wang, Y.; Anzalone, A.V.; Raguram, A.; Doman, J.L.; et al. Prime genome editing in rice and wheat. Nat. Biotechnol. 2020, 38, 582–585. [Google Scholar] [CrossRef] [PubMed]
127. Xu, W.; Zhang, C.; Yang, Y.; Zhao, S.; Kang, G.; He, X.; Song, J.; Yang, J. Versatile Nucleotides Substitution in Plant Using an Improved Prime Editing System. Mol. Plant 2020, 13, 675–678. [Google Scholar] [CrossRef]
128. Jiang, Y.Y.; Chai, Y.P.; Lu, M.H.; Han, X.L.; Lin, Q.; Zhang, Y.; Zhang, Q.; Zhou, Y.; Wang, X.-C.; Gao, C.; et al. Prime editing efficiently generates W542L and S621I double mutations in two ALS genes in maize. Genome Biol. 2020, 21, 257. [Google Scholar] [CrossRef]
129. Lu, Y.; Tian, Y.; Shen, R.; Yao, Q.; Zhong, D.; Zhang, X.; Zhu, J.K. Precise genome modification in tomato using an improved prime editing system. Plant Biotechnol. J. 2020, 19, 415–417. [Google Scholar] [CrossRef] [PubMed]
130. Xu, R.; Li, J.; Liu, X.; Shan, T.; Qin, R.; Wei, P. Development of Plant Prime-Editing Systems for Precise Genome Editing. Plant Commun. 2020, 1, 100043. [Google Scholar] [CrossRef] [PubMed]
131. Lin, Q.; Jin, S.; Zong, Y.; Yu, H.; Zhu, Z.; Liu, G.; Kou, L.; Wang, Y.; Qiu, J.-L.; Li, J.; et al. High-efficiency prime editing with optimized, paired pegRNAs in plants. Nat. Biotechnol. 2021, 39, 923–927. [Google Scholar] [CrossRef]
132. Jin, S.; Lin, Q.; Luo, Y.; Zhu, Z.; Liu, G.; Li, Y.; Chen, K.; Qiu, J.-L.; Gao, C. Genome-wide specificity of prime editors in plants. Nat. Biotechnol. 2021, 39, 1292–1299. [Google Scholar] [CrossRef] [PubMed]
133. Zong, Y.; Liu, Y.; Xue, C.; Li, B.; Li, X.; Wang, Y.; Li, J.; Liu, G.; Huang, X.; Cao, X.; et al. An engineered prime editor with enhanced editing efficiency in plants. Nat. Biotechnol. 2022, 40, 1394–1402. [Google Scholar] [CrossRef] [PubMed]
134. Ni, P.; Zhao, Y.; Zhou, X.; Liu, Z.; Huang, Z.; Ni, Z.; Sun, Q.; Zong, Y. Efficient and versatile multiplex prime editing in hexaploid wheat. Genome Biol. 2023, 24, 156. [Google Scholar] [CrossRef] [PubMed]
135. Doman, J.L.; Pandey, S.; Neugebauer, M.E.; An, M.; Davis, J.R.; Randolph, P.B.; McElroy, A.; Gao, X.D.; Raguram, A.; Richter, M.F.; et al. Phage-assisted evolution and protein engineering yield compact, efficient prime editors. Cell 2023, 186, 3983–4002. [Google Scholar] [CrossRef]
136. Gupta, A.; Liu, B.; Raza, S.; Chen, Q.-J.; Yang, B. Modularly assembled multiplex prime editors for simultaneous editing of agronomically important genes in rice. Plant Commun. 2024, 5, 100741. [Google Scholar] [CrossRef] [PubMed]
137. Hwang, G.-H.; Jeong, Y.K.; Habib, O.; Hong, S.-A.; Lim, K.; Kim, J.-S.; Bae, S. PE-Designer and PE-Analyzer: Web-based design and analysis tools for CRISPR prime editing. Nucleic Acids Res. 2021, 49, W499–W504. [Google Scholar] [CrossRef]
138. Chen, P.J.; Hussmann, J.A.; Yan, J.; Knipping, F.; Ravisankar, P.; Chen, P.-F.; Chen, C.; Nelson, J.W.; Newby, G.A.; Sahin, M.; et al. Enhanced prime editing systems by manipulating cellular determinants of editing outcomes. Cell 2021, 184, 5635–5652. [Google Scholar] [CrossRef]
139. Jiang, Y.; Chai, Y.; Qiao, D.; Wang, J.; Xin, C.; Sun, W.; Cao, Z.; Zhang, Y.; Zhou, Y.; Wang, X.-C.; et al. Optimized prime editing efficiently generates glyphosate-resistant rice plants carrying homozygous TAP-IVS mutation in EPSPS. Mol. Plant 2022, 15, 1646–1649. [Google Scholar] [CrossRef]
140. Liu, X.; Gu, D.; Zhang, Y.; Jiang, Y.; Xiao, Z.; Xu, R.; Qin, R.; Li, J.; Wei, P. Conditional knockdown of OsMLH1 to improve plant prime editing systems without disturbing fertility in rice. Genome Biol. 2024, 25, 131. [Google Scholar] [CrossRef] [PubMed]
141. Choi, J.; Chen, W.; Suiter, C.C.; Lee, C.; Chardon, F.M.; Yang, W.; Leith, A.; Daza, R.M.; Martin, B.; Shendure, J. Precise genomic deletions using paired prime editing. Nat. Biotechnol. 2021, 40, 218–226. [Google Scholar] [CrossRef] [PubMed]
142. Jiang, T.; Zhang, X.-O.; Weng, Z.; Xue, W. Deletion and replacement of long genomic sequences using prime editing. Nat. Biotechnol. 2021, 40, 227–234. [Google Scholar] [CrossRef] [PubMed]
143. Sun, C.; Lei, Y.; Li, B.; Gao, Q.; Li, Y.; Cao, W.; Yang, C.; Li, H.; Wang, Z.; Li, Y.; et al. Precise integration of large DNA sequences in plant genomes using PrimeRoot editors. Nat. Biotechnol. 2023, 42, 316–327. [Google Scholar] [CrossRef]
144. Nishimasu, H.; Shi, X.; Ishiguro, S.; Gao, L.; Hirano, S.; Okazaki, S.; Noda, T.; Abudayyeh, O.O.; Gootenberg, J.S.; Mori, H.; et al. Engineered CRISPR-Cas9 nuclease with expanded targeting space. Science 2018, 361, 1259–1262. [Google Scholar] [CrossRef] [PubMed]
145. Hua, K.; Tao, X.; Han, P.; Wang, R.; Zhu, J.-K. Genome Engineering in Rice Using Cas9 Variants that Recognize NG PAM Sequences. Mol. Plant 2019, 12, 1003–1014. [Google Scholar] [CrossRef] [PubMed]
146. Endo, M.; Mikami, M.; Endo, A.; Kaya, H.; Itoh, T.; Nishimasu, H.; Nureki, O.; Toki, S. Genome editing in plants by engineered CRISPR–Cas9 recognizing NG PAM. Nat. Plants 2018, 5, 14–17. [Google Scholar] [CrossRef]
147. Ren, Q.; Sretenovic, S.; Liu, S.; Tang, X.; Huang, L.; He, Y.; Liu, L.; Guo, Y.; Zhong, Z.; Liu, G.; et al. PAM-less plant genome editing using a CRISPR–SpRY toolbox. Nat. Plants 2021, 7, 25–33. [Google Scholar] [CrossRef]
148. Ren, J.; Meng, X.; Hu, F.; Liu, Q.; Cao, Y.; Li, H.; Yan, C.; Li, J.; Wang, K.; Yu, H.; et al. Expanding the scope of genome editing with SpG and SpRY variants in rice. Sci. China Life Sci. 2021, 64, 1784–1787. [Google Scholar] [CrossRef]
149. Li, J.; Xu, R.; Qin, R.; Liu, X.; Kong, F.; Wei, P. Genome editing mediated by SpCas9 variants with broad non-canonical PAM compatibility in plants. Mol. Plant 2021, 14, 352–360. [Google Scholar] [CrossRef]
150. Hu, Y.; Patra, P.; Pisanty, O.; Shafir, A.; Belew, Z.M.; Binenbaum, J.; Ben Yaakov, S.; Shi, B.; Charrier, L.; Hyams, G.; et al. Multi-Knock—A multi-targeted genome-scale CRISPR toolbox to overcome functional redundancy in plants. Nat. Plants 2023, 9, 572–587. [Google Scholar] [CrossRef]
151. Ma, X.; Zhang, Q.; Zhu, Q.; Liu, W.; Chen, Y.; Qiu, R.; Wang, B.; Yang, Z.; Li, H.; Lin, Y.; et al. A Robust CRISPR/Cas9 System for Convenient, High-Efficiency Multiplex Genome Editing in Monocot and Dicot Plants. Mol. Plant 2015, 8, 1274–1284. [Google Scholar] [CrossRef] [PubMed]
152. Xie, K.; Minkenberg, B.; Yang, Y. Boosting CRISPR/Cas9 multiplex editing capability with the endogenous tRNA-processing system. Proc. Natl. Acad. Sci. USA 2015, 112, 3570–3575. [Google Scholar] [CrossRef]
153. Čermák, T.; Curtin, S.J.; Gil-Humanes, J.; Čegan, R.; Kono, T.J.Y.; Konečná, E.; Belanto, J.J.; Starker, C.G.; Mathre, J.W.; Greenstein, R.L.; et al. A Multipurpose Toolkit to Enable Advanced Genome Engineering in Plants. Plant Cell 2017, 29, 1196–1217. [Google Scholar] [CrossRef]
154. Ding, D.; Chen, K.; Chen, Y.; Li, H.; Xie, K. Engineering Introns to Express RNA Guides for Cas9- and Cpf1-Mediated Multiplex Genome Editing. Mol. Plant 2018, 11, 542–552. [Google Scholar] [CrossRef]
155. Zhang, Z.; Mao, Y.; Ha, S.; Liu, W.; Botella, J.R.; Zhu, J.-K. A multiplex CRISPR/Cas9 platform for fast and efficient editing of multiple genes in Arabidopsis. Plant Cell Rep. 2015, 35, 1519–1533. [Google Scholar] [CrossRef]
156. Wang, C.; Shen, L.; Fu, Y.; Yan, C.; Wang, K. A Simple CRISPR/Cas9 System for Multiplex Genome Editing in Rice. J. Genet. Genom. 2015, 42, 703–706. [Google Scholar] [CrossRef]
157. Hao, Y.; Zong, W.; Zeng, D.; Han, J.; Chen, S.; Tang, J.; Zhao, Z.; Li, X.; Ma, K.; Xie, X.; et al. Shortened snRNA promoters for efficient CRISPR/Cas-based multiplex genome editing in monocot plants. Sci. China Life Sci. 2020, 63, 933–935. [Google Scholar] [CrossRef] [PubMed]
158. Wang, Y.; Li, X.; Liu, M.; Zhou, Y.; Li, F. Guide RNA scaffold variants enabled easy cloning of large gRNA cluster for multiplexed gene editing. Plant Biotechnol. J. 2023, 22, 460–471. [Google Scholar] [CrossRef]
159. Liu, H.; Ding, Y.; Zhou, Y.; Jin, W.; Xie, K.; Chen, L.-L. CRISPR-P 2.0: An Improved CRISPR-Cas9 Tool for Genome Editing in Plants. Mol. Plant 2017, 10, 530–532. [Google Scholar] [CrossRef]
160. Sun, J.; Liu, H.; Liu, J.; Cheng, S.; Peng, Y.; Zhang, Q.; Yan, J.; Liu, H.-J.; Chen, L.-L.; Birol, I. CRISPR-Local: A local single-guide RNA (sgRNA) design tool for non-reference plant genomes. Bioinformatics 2019, 35, 2501–2503. [Google Scholar] [CrossRef]
161. Xie, X.; Ma, X.; Zhu, Q.; Zeng, D.; Li, G.; Liu, Y.-G. CRISPR-GE: A Convenient Software Toolkit for CRISPR-Based Genome Editing. Mol. Plant 2017, 10, 1246–1249. [Google Scholar] [CrossRef] [PubMed]
162. Park, J.; Bae, S.; Kim, J.-S. Cas-Designer: A web-based tool for choice of CRISPR-Cas9 target sites. Bioinformatics 2015, 31, 4014–4016. [Google Scholar] [CrossRef]
163. Concordet, J.-P.; Haeussler, M. CRISPOR: Intuitive guide selection for CRISPR/Cas9 genome editing experiments and screens. Nucleic Acids Res. 2018, 46, W242–W245. [Google Scholar] [CrossRef]
164. Hassan, M.M.; Zhang, Y.; Yuan, G.; De, K.; Chen, J.-G.; Muchero, W.; Tuskan, G.A.; Qi, Y.; Yang, X. Construct design for CRISPR/Cas-based genome editing in plants. Trends Plant Sci. 2021, 26, 1133–1152. [Google Scholar] [CrossRef]
165. Sun, T.; Liu, Q.; Chen, X.; Hu, F.; Wang, K. Hi-TOM 2.0: An improved platform for high-throughput mutation detection. Sci. China Life Sci. 2024, 67, 1532–1534. [Google Scholar] [CrossRef] [PubMed]
166. Clement, K.; Rees, H.; Canver, M.C.; Farouni, R.; Gehrke, J.M.; Hsu, J.Y.; Cole, M.A.; Liu, D.R.; Joung, J.K.; Bauer, D.E.; et al. CRISPResso2 provides accurate and rapid genome editing sequence analysis. Nat. Biotechnol. 2019, 37, 215–216. [Google Scholar] [CrossRef] [PubMed]
167. Fu, H.; Shan, C.; Kang, F.; Yu, L.; Li, Z.; Yin, Y. CRISPR-GRANT: A cross-platform graphical analysis tool for high-throughput CRISPR-based genome editing evaluation. BMC Bioinform. 2023, 24, 219. [Google Scholar] [CrossRef] [PubMed]
168. Han, Y.; Liu, G.; Wu, Y.; Bao, Y.; Zhang, Y.; Zhang, T. CrisprStitch: Fast evaluation of the efficiency of CRISPR editing systems. Plant Commun. 2024, 5, 100783. [Google Scholar] [CrossRef] [PubMed]
169. Huang, Y.; Shang, M.; Liu, T.; Wang, K. High-throughput methods for genome editing: The more the better. Plant Physiol. 2022, 188, 1731–1745. [Google Scholar] [CrossRef]
170. Gaillochet, C.; Develtere, W.; Jacobs, T.B. CRISPR screens in plants: Approaches, guidelines, and future prospects. Plant Cell 2021, 33, 794–813. [Google Scholar] [CrossRef]
171. Lu, Y.; Ye, X.; Guo, R.; Huang, J.; Wang, W.; Tang, J.; Tan, L.; Zhu, J.-K.; Chu, C.; Qian, Y. Genome-wide Targeted Mutagenesis in Rice Using the CRISPR/Cas9 System. Mol. Plant 2017, 10, 1242–1245. [Google Scholar] [CrossRef]
172. Meng, X.; Yu, H.; Zhang, Y.; Zhuang, F.; Song, X.; Gao, S.; Gao, C.; Li, J. Construction of a Genome-Wide Mutant Library in Rice Using CRISPR/Cas9. Mol. Plant 2017, 10, 1238–1241. [Google Scholar] [CrossRef] [PubMed]
173. Bi, M.; Wang, Z.; Cheng, K.; Cui, Y.; He, Y.; Ma, J.; Qi, M. Construction of transcription factor mutagenesis population in tomato using a pooled CRISPR/Cas9 plasmid library. Plant Physiol. Biochem. 2023, 205, 108094. [Google Scholar] [CrossRef] [PubMed]
174. He, J.; Zhang, K.; Yan, S.; Tang, M.; Zhou, W.; Yin, Y.; Chen, K.; Zhang, C.; Li, M. Genome-scale targeted mutagenesis in Brassica napus using a pooled CRISPR library. Genome Res. 2023, 33, 798–809. [Google Scholar] [CrossRef]
175. Jacobs, T.B.; Zhang, N.; Patel, D.; Martin, G.B. Generation of a Collection of Mutant Tomato Lines Using Pooled CRISPR Libraries. Plant Physiol. 2017, 174, 2023–2037. [Google Scholar] [CrossRef]
176. Bai, M.; Yuan, J.; Kuang, H.; Gong, P.; Li, S.; Zhang, Z.; Liu, B.; Sun, J.; Yang, M.; Yang, L.; et al. Generation of a multiplex mutagenesis population via pooled CRISPR-Cas9 in soya bean. Plant Biotechnol. J. 2019, 18, 721–731. [Google Scholar] [CrossRef] [PubMed]
177. Liu, H.-J.; Jian, L.; Xu, J.; Zhang, Q.; Zhang, M.; Jin, M.; Peng, Y.; Yan, J.; Han, B.; Liu, J.; et al. High-Throughput CRISPR/Cas9 Mutagenesis Streamlines Trait Gene Identification in Maize. Plant Cell 2020, 32, 1397–1413. [Google Scholar] [CrossRef] [PubMed]
178. Ma, K.; Han, J.; Hao, Y.; Yang, Z.; Chen, J.; Liu, Y.-G.; Zhu, Q.; Chen, L. An effective strategy to establish a male sterility mutant mini-library by CRISPR/Cas9-mediated knockout of anther-specific genes in rice. J. Genet. Genom. 2019, 46, 273–275. [Google Scholar] [CrossRef] [PubMed]
179. Chen, K.; Ke, R.; Du, M.; Yi, Y.; Chen, Y.; Wang, X.; Yao, L.; Liu, H.; Hou, X.; Xiong, L.; et al. A FLASH pipeline for arrayed CRISPR library construction and the gene function discovery of rice receptor-like kinases. Mol. Plant 2022, 15, 243–257. [Google Scholar] [CrossRef]
180. Zhao, D.; Chen, S.; Han, Y.; Liu, G.; Liu, J.; Yang, Q.; Zhang, T.; Shen, J.; Fan, X.; Zhang, C.; et al. A CRISPR/Cas9-mediated mutant library of seed-preferred genes in rice. Plant Biotechnol. J. 2024, 22, 3012–3014. [Google Scholar] [CrossRef] [PubMed]
181. Ramadan, M.; Alariqi, M.; Ma, Y.; Li, Y.; Liu, Z.; Zhang, R.; Jin, S.; Min, L.; Zhang, X. Efficient CRISPR/Cas9 mediated Pooled-sgRNAs assembly accelerates targeting multiple genes related to male sterility in cotton. Plant Methods 2021, 17, 16. [Google Scholar] [CrossRef]
182. Sun, L.; Alariqi, M.; Wang, Y.; Wang, Q.; Xu, Z.; Zafar, M.N.; Yang, G.; Jia, R.; Hussain, A.; Chen, Y.; et al. Construction of Host Plant Insect-Resistance Mutant Library by High-Throughput CRISPR/Cas9 System and Identification of A Broad-Spectrum Insect Resistance Gene. Adv. Sci. 2023, 11, 2306157. [Google Scholar] [CrossRef] [PubMed]
183. Wang, F.; Liang, S.; Wang, G.; Hu, T.; Fu, C.; Wang, Q.; Xu, Z.; Fan, Y.; Che, L.; Min, L.; et al. CRISPR/Cas9-mediated generation of a mutant library of cotton CDPK gene family for identifying insect-resistant genes. Plant Commun. 2024, 12, 101047. [Google Scholar] [CrossRef]
184. Lorenzo, C.D.; Debray, K.; Herwegh, D.; Develtere, W.; Impens, L.; Schaumont, D.; Vandeputte, W.; Aesaert, S.; Coussens, G.; De Boe, Y.; et al. BREEDIT: A multiplex genome editing strategy to improve complex quantitative traits in maize. Plant Cell 2023, 35, 218–238. [Google Scholar] [CrossRef] [PubMed]
185. Butt, H.; Eid, A.; Momin, A.A.; Bazin, J.; Crespi, M.; Arold, S.T.; Mahfouz, M.M. CRISPR directed evolution of the spliceosome for resistance to splicing inhibitors. Genome Biol. 2019, 20, 73. [Google Scholar] [CrossRef]
186. Liu, X.; Qin, R.; Li, J.; Liao, S.; Shan, T.; Xu, R.; Wu, D.; Wei, P. A CRISPR-Cas9-mediated domain-specific base-editing screen enables functional assessment of ACCase variants in rice. Plant Biotechnol. J. 2020, 18, 1845–1847. [Google Scholar] [CrossRef] [PubMed]
187. Kuang, Y.; Li, S.; Ren, B.; Yan, F.; Spetz, C.; Li, X.; Zhou, X.; Zhou, H. Base-Editing-Mediated Artificial Evolution of OsALS1 In Planta to Develop Novel Herbicide-Tolerant Rice Germplasms. Mol. Plant 2020, 13, 565–572. [Google Scholar] [CrossRef]
188. Xu, R.; Liu, X.; Li, J.; Qin, R.; Wei, P. Identification of herbicide resistance OsACC1 mutations via in planta prime-editing-library screening in rice. Nat. Plants 2021, 7, 888–892. [Google Scholar] [CrossRef]
189. Wang, X.; Pan, W.; Sun, C.; Yang, H.; Cheng, Z.; Yan, F.; Ma, G.; Shang, Y.; Zhang, R.; Gao, C.; et al. Creating large-scale genetic diversity in Arabidopsis via base editing-mediated deep artificial evolution. Genome Biol. 2024, 25, 215. [Google Scholar] [CrossRef]
190. Peng, A.; Chen, S.; Lei, T.; Xu, L.; He, Y.; Wu, L.; Yao, L.; Zou, X. Engineering canker-resistant plants through CRISPR/Cas9-targeted editing of the susceptibility gene CsLOB1 promoter in citrus. Plant Biotechnol. J. 2017, 15, 1509–1519. [Google Scholar] [CrossRef] [PubMed]
191. Rodríguez-Leal, D.; Lemmon, Z.H.; Man, J.; Bartlett, M.E.; Lippman, Z.B. Engineering Quantitative Trait Variation for Crop Improvement by Genome Editing. Cell 2017, 171, 470–480. [Google Scholar] [CrossRef]
192. Wang, X.; Aguirre, L.; Rodríguez-Leal, D.; Hendelman, A.; Benoit, M.; Lippman, Z.B. Dissecting cis-regulatory control of quantitative trait variation in a plant stem cell circuit. Nat. Plants 2021, 7, 419–427. [Google Scholar] [CrossRef] [PubMed]
193. Hendelman, A.; Zebell, S.; Rodriguez-Leal, D.; Dukler, N.; Robitaille, G.; Wu, X.; Kostyun, J.; Tal, L.; Wang, P.; Bartlett, M.E.; et al. Conserved pleiotropy of an ancient plant homeobox gene uncovered by cis-regulatory dissection. Cell 2021, 184, 1724–1739. [Google Scholar] [CrossRef] [PubMed]
194. Liu, L.; Gallagher, J.; Arevalo, E.D.; Chen, R.; Skopelitis, T.; Wu, Q.; Bartlett, M.; Jackson, D. Enhancing grain-yield-related traits by CRISPR–Cas9 promoter editing of maize CLE genes. Nat. Plants 2021, 7, 287–294. [Google Scholar] [CrossRef]
195. Huang, L.; Li, Q.; Zhang, C.; Chu, R.; Gu, Z.; Tan, H.; Zhao, D.; Fan, X.; Liu, Q. Creating novel Wx alleles with fine-tuned amylose levels and improved grain quality in rice by promoter editing using CRISPR/Cas9 system. Plant Biotechnol. J. 2020, 18, 2164–2166. [Google Scholar] [CrossRef]
196. Zhou, S.; Cai, L.; Wu, H.; Wang, B.; Gu, B.; Cui, S.; Huang, X.; Xu, Z.; Hao, B.; Hou, H.; et al. Fine-tuning rice heading date through multiplex editing of the regulatory regions of key genes by CRISPR-Cas9. Plant Biotechnol. J. 2023, 22, 751–758. [Google Scholar] [CrossRef]
197. Song, X.; Meng, X.; Guo, H.; Cheng, Q.; Jing, Y.; Chen, M.; Liu, G.; Wang, B.; Wang, Y.; Li, J.; et al. Targeting a gene regulatory element enhances rice grain yield by decoupling panicle number and size. Nat. Biotechnol. 2022, 40, 1403–1411. [Google Scholar] [CrossRef] [PubMed]
198. Avsec, Ž.; Weilert, M.; Shrikumar, A.; Krueger, S.; Alexandari, A.; Dalal, K.; Fropf, R.; McAnany, C.; Gagneur, J.; Kundaje, A.; et al. Base-resolution models of transcription-factor binding reveal soft motif syntax. Nat. Genet. 2021, 53, 354–366. [Google Scholar] [CrossRef]
199. Tang, X.; Zhang, Y. Beyond knockouts: Fine-tuning regulation of gene expression in plants with CRISPR-Cas-based promoter editing. New Phytol. 2023, 239, 868–874. [Google Scholar] [CrossRef]
200. Cui, Y.; Cao, Q.; Li, Y.; He, M.; Liu, X.; Cubas, P. Advances in cis-element- and natural variation-mediated transcriptional regulation and applications in gene editing of major crops. J. Exp. Bot. 2023, 74, 5441–5457. [Google Scholar] [CrossRef]
201. Zhou, J.; Liu, G.; Zhao, Y.; Zhang, R.; Tang, X.; Li, L.; Jia, X.; Guo, Y.; Wu, Y.; Han, Y.; et al. An efficient CRISPR–Cas12a promoter editing system for crop improvement. Nat. Plants 2023, 9, 588–604. [Google Scholar] [CrossRef]
202. Zhang, H.; Wang, Y.; Lu, J. Function and Evolution of Upstream ORFs in Eukaryotes. Trends Biochem. Sci. 2019, 44, 782–794. [Google Scholar] [CrossRef]
203. Zhang, H.; Si, X.; Ji, X.; Fan, R.; Liu, J.; Chen, K.; Wang, D.; Gao, C. Genome editing of upstream open reading frames enables translational control in plants. Nat. Biotechnol. 2018, 36, 894–898. [Google Scholar] [CrossRef]
204. Si, X.; Zhang, H.; Wang, Y.; Chen, K.; Gao, C. Manipulating gene translation in plants by CRISPR–Cas9-mediated genome editing of upstream open reading frames. Nat. Protoc. 2020, 15, 338–363. [Google Scholar] [CrossRef] [PubMed]
205. Xing, S.; Chen, K.; Zhu, H.; Zhang, R.; Zhang, H.; Li, B.; Gao, C. Fine-tuning sugar content in strawberry. Genome Biol. 2020, 21, 230. [Google Scholar] [CrossRef] [PubMed]
206. Deslous, P.; Bournonville, C.; Decros, G.; Okabe, Y.; Mauxion, J.-P.; Jorly, J.; Gadin, S.; Brès, C.; Mori, K.; Ferrand, C.; et al. Overproduction of ascorbic acid impairs pollen fertility in tomato. J. Exp. Bot. 2021, 72, 3091–3107. [Google Scholar] [CrossRef] [PubMed]
207. Patel-Tupper, D.; Kelikian, A.; Leipertz, A.; Maryn, N.; Tjahjadi, M.; Karavolias, N.G.; Cho, M.-J.; Niyogi, K.K. Multiplexed CRISPR—Cas9 mutagenesis of rice PSBS1 noncoding sequences for transgene—Free overexpression. Sci. Adv. 2024, 10, eadm7452. [Google Scholar] [CrossRef] [PubMed]
208. Hyun, Y.; Kim, J.; Cho, S.W.; Choi, Y.; Kim, J.-S.; Coupland, G. Site-directed mutagenesis in Arabidopsis thaliana using dividing tissue-targeted RGEN of the CRISPR/Cas system to generate heritable null alleles. Planta 2014, 241, 271–284. [Google Scholar] [CrossRef] [PubMed]
209. Yan, L.; Wei, S.; Wu, Y.; Hu, R.; Li, H.; Yang, W.; Xie, Q. High-Efficiency Genome Editing in Arabidopsis Using YAO Promoter-Driven CRISPR/Cas9 System. Mol. Plant 2015, 8, 1820–1823. [Google Scholar] [CrossRef] [PubMed]
210. Mao, Y.; Zhang, Z.; Feng, Z.; Wei, P.; Zhang, H.; Botella, J.R.; Zhu, J.K. Development of germ-line-specific CRISPR-Cas9 systems to improve the production of heritable gene modifications in Arabidopsis. Plant Biotechnol. J. 2015, 14, 519–532. [Google Scholar] [CrossRef] [PubMed]
211. Miki, D.; Zhang, W.; Zeng, W.; Feng, Z.; Zhu, J.-K. CRISPR/Cas9-mediated gene targeting in Arabidopsis using sequential transformation. Nat. Commun. 2018, 9, 1967. [Google Scholar] [CrossRef]
212. Wolter, F.; Klemm, J.; Puchta, H. Efficient in planta gene targeting in Arabidopsis using egg cell-specific expression of the Cas9 nuclease of Staphylococcus aureus. Plant J. 2018, 94, 735–746. [Google Scholar] [CrossRef] [PubMed]
213. Kong, W.; Wang, M.; Huang, L.; Wu, F.; Tao, J.; Mo, B.; Yu, Y. A high-efficient and naked-eye visible CRISPR/Cas9 system in Arabidopsis. Planta 2023, 257, 30. [Google Scholar] [CrossRef] [PubMed]
214. Decaestecker, W.; Buono, R.A.; Pfeiffer, M.L.; Vangheluwe, N.; Jourquin, J.; Karimi, M.; Van Isterdael, G.; Beeckman, T.; Nowack, M.K.; Jacobs, T.B. CRISPR-TSKO: A Technique for Efficient Mutagenesis in Specific Cell Types, Tissues, or Organs in Arabidopsis. Plant Cell 2019, 31, 2868–2887. [Google Scholar] [CrossRef]
215. Feder, A.; Jensen, S.; Wang, A.; Courtney, L.; Middleton, L.; Van Eck, J.; Liu, Y.; Giovannoni, J.J. Tomato fruit as a model for tissue-specific gene silencing in crop plants. Hortic. Res. 2020, 7, 142. [Google Scholar] [CrossRef]
216. Lei, J.; Dai, P.; Li, J.; Yang, M.; Li, X.; Zhang, W.; Zhou, G.; Guo, W.; Liu, X. Tissue-Specific CRISPR/Cas9 System of Cotton Pollen with GhPLIMP2b and GhMYB24 Promoters. J. Plant Biol. 2020, 64, 13–21. [Google Scholar] [CrossRef]
217. He, Y.; Zhu, M.; Wang, L.; Wu, J.; Wang, Q.; Wang, R.; Zhao, Y. Programmed Self-Elimination of the CRISPR/Cas9 Construct Greatly Accelerates the Isolation of Edited and Transgene-Free Rice Plants. Mol. Plant 2018, 11, 1210–1213. [Google Scholar] [CrossRef]
218. Gao, X.; Chen, J.; Dai, X.; Zhang, D.; Zhao, Y. An Effective Strategy for Reliably Isolating Heritable and Cas9-Free Arabidopsis Mutants Generated by CRISPR/Cas9-Mediated Genome Editing. Plant Physiol. 2016, 171, 1794–1800. [Google Scholar] [CrossRef]
219. Wang, J.; Chen, H. A novel CRISPR/Cas9 system for efficiently generating Cas9-free multiplex mutants in Arabidopsis. aBIOTECH 2019, 1, 6–14. [Google Scholar] [CrossRef]
220. Deng, Y.; Duan, A.; Li, T.; Wang, H.; Xiong, A. A betaxanthin-based visible and fluorescent reporter for monitoring plant transformation. Crop J. 2023, 11, 666–671. [Google Scholar] [CrossRef]
221. He, Y.; Zhang, T.; Sun, H.; Zhan, H.; Zhao, Y. A reporter for noninvasively monitoring gene expression and plant transformation. Hortic. Res. 2020, 7, 152. [Google Scholar] [CrossRef] [PubMed]
222. Lee, K.; Kang, M.; Ji, Q.; Grosic, S.; Wang, K. New T-DNA binary vectors with NptII selection and RUBY reporter for efficient maize transformation and targeted mutagenesis. Plant Physiol. 2023, 192, 2598–2603. [Google Scholar] [CrossRef]
223. Xu, J.; Yin, Y.; Jian, L.; Han, B.; Chen, Z.; Yan, J.; Liu, X. Seeing is believing: A visualization toolbox to enhance selection efficiency in maize genome editing. Plant Biotechnol. J. 2021, 19, 872–874. [Google Scholar] [CrossRef] [PubMed]
224. Yan, Y.; Zhu, J.; Qi, X.; Cheng, B.; Liu, C.; Xie, C. Establishment of an efficient seed fluorescence reporter-assisted CRISPR/Cas9 gene editing in maize. J. Integr. Plant Biol. 2021, 63, 1671–1680. [Google Scholar] [CrossRef] [PubMed]
225. Liu, Y.; Zeng, J.; Yuan, C.; Guo, Y.; Yu, H.; Li, Y.; Huang, C. Cas9-PF, an early flowering and visual selection marker system, enhances the frequency of editing event occurrence and expedites the isolation of genome-edited and transgene-free plants. Plant Biotechnol. J. 2019, 17, 1191–1193. [Google Scholar] [CrossRef]
226. Kong, X.; Pan, W.; Sun, N.; Zhang, T.; Liu, L.; Zhang, H. GLABRA2-based selection efficiently enriches Cas9-generated nonchimeric mutants in the T1 generation. Plant Physiol. 2021, 187, 758–768. [Google Scholar] [CrossRef]
227. Kelliher, T.; Starr, D.; Su, X.; Tang, G.; Chen, Z.; Carter, J.; Wittich, P.E.; Dong, S.; Green, J.; Burch, E.; et al. One-step genome editing of elite crop germplasm during haploid induction. Nat. Biotechnol. 2019, 37, 287–292. [Google Scholar] [CrossRef] [PubMed]
228. Wang, B.; Zhu, L.; Zhao, B.; Zhao, Y.; Xie, Y.; Zheng, Z.; Li, Y.; Sun, J.; Wang, H. Development of a Haploid-Inducer Mediated Genome Editing System for Accelerating Maize Breeding. Mol. Plant 2019, 12, 597–602. [Google Scholar] [CrossRef] [PubMed]
229. Iwase, A.; Mita, K.; Nonaka, S.; Ikeuchi, M.; Koizuka, C.; Ohnuma, M.; Ezura, H.; Imamura, J.; Sugimoto, K. WIND1-based acquisition of regeneration competency in Arabidopsis and rapeseed. J. Plant Res. 2015, 128, 389–397. [Google Scholar] [CrossRef]
230. Lowe, K.; Wu, E.; Wang, N.; Hoerster, G.; Hastings, C.; Cho, M.-J.; Scelonge, C.; Lenderts, B.; Chamberlin, M.; Cushatt, J.; et al. Morphogenic Regulators Baby boom and Wuschel Improve Monocot Transformation. Plant Cell 2016, 28, 1998–2015. [Google Scholar] [CrossRef]
231. Chen, J.; Tomes, S.; Gleave, A.P.; Hall, W.; Luo, Z.; Xu, J.; Yao, J.-L. Significant improvement of apple (Malus domestica Borkh.) transgenic plant production by pre-transformation with a Baby boom transcription factor. Hortic. Res. 2022, 9, uhab014. [Google Scholar] [CrossRef]
232. Zhang, T.-Q.; Lian, H.; Zhou, C.-M.; Xu, L.; Jiao, Y.; Wang, J.-W. A Two-Step Model for de Novo Activation of WUSCHEL during Plant Shoot Regeneration. Plant Cell 2017, 29, 1073–1087. [Google Scholar] [CrossRef]
233. Omidbakhshfard, M.A.; Proost, S.; Fujikura, U.; Mueller-Roeber, B. Growth-Regulating Factors (GRFs): A Small Transcription Factor Family with Important Functions in Plant Biology. Mol. Plant 2015, 8, 998–1010. [Google Scholar] [CrossRef] [PubMed]
234. Liebsch, D.; Palatnik, J.F. MicroRNA miR396, GRF transcription factors and GIF co-regulators: A conserved plant growth regulatory module with potential for breeding and biotechnology. Curr. Opin. Plant Biol. 2020, 53, 31–42. [Google Scholar] [CrossRef] [PubMed]
235. Kong, J.; Martin-Ortigosa, S.; Finer, J.; Orchard, N.; Gunadi, A.; Batts, L.A.; Thakare, D.; Rush, B.; Schmitz, O.; Stuiver, M.; et al. Overexpression of the Transcription Factor GROWTH-REGULATING FACTOR5 Improves Transformation of Dicot and Monocot Species. Front. Plant Sci. 2020, 11, 572319. [Google Scholar] [CrossRef]
236. Debernardi, J.M.; Tricoli, D.M.; Ercoli, M.F.; Hayta, S.; Ronald, P.; Palatnik, J.F.; Dubcovsky, J. A GRF–GIF chimeric protein improves the regeneration efficiency of transgenic plants. Nat. Biotechnol. 2020, 38, 1274–1279. [Google Scholar] [CrossRef] [PubMed]
237. Lian, Z.; Nguyen, C.D.; Liu, L.; Wang, G.; Chen, J.; Wang, S.; Yi, G.; Wilson, S.; Ozias-Akins, P.; Gong, H.; et al. Application of developmental regulators to improve in planta or in vitro transformation in plants. Plant Biotechnol. J. 2022, 20, 1622–1635. [Google Scholar] [CrossRef]
238. Yu, Y.; Yu, H.; Peng, J.; Yao, W.J.; Wang, Y.P.; Zhang, F.L.; Wang, S.R.; Zhao, Y.; Zhao, X.Y.; Zhang, X.S.; et al. Enhancing wheat regeneration and genetic transformation through overexpression of TaLAX1. Plant Commun. 2024, 5, 100738. [Google Scholar] [CrossRef] [PubMed]
239. Liu, X.; Bie, X.M.; Lin, X.; Li, M.; Wang, H.; Zhang, X.; Yang, Y.; Zhang, C.; Zhang, X.S.; Xiao, J. Uncovering the transcriptional regulatory network involved in boosting wheat regeneration and transformation. Nat. Plants 2023, 9, 908–925. [Google Scholar] [CrossRef]
240. Zhu, X.; Xu, Z.; Wang, G.; Cong, Y.; Yu, L.; Jia, R.; Qin, Y.; Zhang, G.; Li, B.; Yuan, D.; et al. Single-cell resolution analysis reveals the preparation for reprogramming the fate of stem cell niche in cotton lateral meristem. Genome Biol. 2023, 24, 194. [Google Scholar]
241. Zhang, Q.; Zhang, Y.; Lu, M.-H.; Chai, Y.-P.; Jiang, Y.-Y.; Zhou, Y.; Wang, X.-C.; Chen, Q.-J. A Novel Ternary Vector System United with Morphogenic Genes Enhances CRISPR/Cas Delivery in Maize. Plant Physiol. 2019, 181, 1441–1448. [Google Scholar] [CrossRef] [PubMed]
242. Lee, H.G.; Jang, S.Y.; Jie, E.Y.; Choi, S.H.; Park, O.-S.; Bae, S.H.; Kim, H.-S.; Kim, S.W.; Hwang, G.-S.; Seo, P.J. Adenosine monophosphate enhances callus regeneration competence for de novo plant organogenesis. Mol. Plant 2023, 16, 1867–1870. [Google Scholar] [CrossRef] [PubMed]
243. Che, P.; Anand, A.; Wu, E.; Sander, J.D.; Simon, M.K.; Zhu, W.; Sigmund, A.L.; Zastrow-Hayes, G.; Miller, M.; Liu, D.; et al. Developing a flexible, high-efficiency Agrobacterium-mediated sorghum transformation system with broad application. Plant Biotechnol. J. 2018, 16, 1388–1395. [Google Scholar] [CrossRef]
244. Maher, M.F.; Nasti, R.A.; Vollbrecht, M.; Starker, C.G.; Clark, M.D.; Voytas, D.F. Plant gene editing through de novo induction of meristems. Nat. Biotechnol. 2019, 38, 84–89. [Google Scholar] [CrossRef]
245. Ali, Z.; Abul-faraj, A.; Li, L.; Ghosh, N.; Piatek, M.; Mahjoub, A.; Aouida, M.; Piatek, A.; Baltes, N.J.; Voytas, D.F.; et al. Efficient Virus-Mediated Genome Editing in Plants Using the CRISPR/Cas9 System. Mol. Plant 2015, 8, 1288–1291. [Google Scholar] [CrossRef] [PubMed]
246. Nagalakshmi, U.; Meier, N.; Liu, J.-Y.; Voytas, D.F.; Dinesh-Kumar, S.P. High-efficiency multiplex biallelic heritable editing in Arabidopsis using an RNA virus. Plant Physiol. 2022, 189, 1241–1245. [Google Scholar] [CrossRef]
247. Liu, D.; Xuan, S.; Prichard, L.E.; Donahue, L.I.; Pan, C.; Nagalakshmi, U.; Ellison, E.E.; Starker, C.G.; Dinesh-Kumar, S.P.; Qi, Y.; et al. Heritable base-editing in Arabidopsis using RNA viral vectors. Plant Physiol. 2022, 189, 1920–1924. [Google Scholar] [CrossRef] [PubMed]
248. Ellison, E.E.; Nagalakshmi, U.; Gamo, M.E.; Huang, P.-J.; Dinesh-Kumar, S.; Voytas, D.F. Multiplexed heritable gene editing using RNA viruses and mobile single guide RNAs. Nat. Plants 2020, 6, 620–624. [Google Scholar] [CrossRef] [PubMed]
249. Li, T.; Hu, J.; Sun, Y.; Li, B.; Zhang, D.; Li, W.; Liu, J.; Li, D.; Gao, C.; Zhang, Y.; et al. Highly efficient heritable genome editing in wheat using an RNA virus and bypassing tissue culture. Mol. Plant 2021, 14, 1787–1798. [Google Scholar] [CrossRef]
250. Gao, Q.; Xu, W.Y.; Yan, T.; Fang, X.D.; Cao, Q.; Zhang, Z.J.; Ding, Z.H.; Wang, Y.; Wang, X.B. Rescue of a plant cytorhabdovirus as versatile expression platforms for planthopper and cereal genomic studies. New Phytol. 2019, 223, 2120–2133. [Google Scholar] [CrossRef] [PubMed]
251. Ma, X.; Zhang, X.; Liu, H.; Li, Z. Highly efficient DNA-free plant genome editing using virally delivered CRISPR–Cas9. Nat. Plants 2020, 6, 773–779. [Google Scholar] [CrossRef]
252. Yang, L.; Machin, F.; Wang, S.; Saplaoura, E.; Kragler, F. Heritable transgene-free genome editing in plants by grafting of wild-type shoots to transgenic donor rootstocks. Nat. Biotechnol. 2023, 41, 958–967. [Google Scholar] [CrossRef]
253. Liang, Z.; Chen, K.; Li, T.; Zhang, Y.; Wang, Y.; Zhao, Q.; Liu, J.; Zhang, H.; Liu, C.; Ran, Y.; et al. Efficient DNA-free genome editing of bread wheat using CRISPR/Cas9 ribonucleoprotein complexes. Nat. Commun. 2017, 8, 14261. [Google Scholar] [CrossRef] [PubMed]
254. Malnoy, M.; Viola, R.; Jung, M.-H.; Koo, O.-J.; Kim, S.; Kim, J.-S.; Velasco, R.; Nagamangala Kanchiswamy, C. DNA-Free Genetically Edited Grapevine and Apple Protoplast Using CRISPR/Cas9 Ribonucleoproteins. Front. Plant Sci. 2016, 7, 1904. [Google Scholar] [CrossRef] [PubMed]
255. Liang, Z.; Chen, K.; Yan, Y.; Zhang, Y.; Gao, C. Genotyping genome-edited mutations in plants using CRISPR ribonucleoprotein complexes. Plant Biotechnol. J. 2018, 16, 2053–2062. [Google Scholar] [CrossRef] [PubMed]
256. Sant’Ana, R.R.A.; Caprestano, C.A.; Nodari, R.O.; Agapito-Tenfen, S.Z. PEG-Delivered CRISPR-Cas9 Ribonucleoproteins System for Gene-Editing Screening of Maize Protoplasts. Genes 2020, 11, 1029. [Google Scholar] [CrossRef] [PubMed]
257. Zhang, Y.; Iaffaldano, B.; Qi, Y. CRISPR ribonucleoprotein-mediated genetic engineering in plants. Plant Commun. 2021, 2, 100168. [Google Scholar] [CrossRef]
258. Banakar, R.; Schubert, M.; Collingwood, M.; Vakulskas, C.; Eggenberger, A.L.; Wang, K. Comparison of CRISPR-Cas9/Cas12a Ribonucleoprotein Complexes for Genome Editing Efficiency in the Rice Phytoene Desaturase (OsPDS) Gene. Rice 2020, 13, 4. [Google Scholar] [CrossRef]
259. Liu, W.; Rudis, M.R.; Cheplick, M.H.; Millwood, R.J.; Yang, J.-P.; Ondzighi-Assoume, C.A.; Montgomery, G.A.; Burris, K.P.; Mazarei, M.; Chesnut, J.D.; et al. Lipofection-mediated genome editing using DNA-free delivery of the Cas9/gRNA ribonucleoprotein into plant cells. Plant Cell Rep. 2019, 39, 245–257. [Google Scholar] [CrossRef]
260. Lee, M.H.; Lee, J.; Choi, S.A.; Kim, Y.-S.; Koo, O.; Choi, S.H.; Ahn, W.S.; Jie, E.Y.; Kim, S.W. Efficient genome editing using CRISPR–Cas9 RNP delivery into cabbage protoplasts via electro-transfection. Plant Biotechnol. Rep. 2020, 14, 695–702. [Google Scholar] [CrossRef]
261. Liu, C.; Li, X.; Meng, D.; Zhong, Y.; Chen, C.; Dong, X.; Xu, X.; Chen, B.; Li, W.; Li, L.; et al. A 4-bp Insertion at ZmPLA1 Encoding a Putative Phospholipase A Generates Haploid Induction in Maize. Mol. Plant 2017, 10, 520–522. [Google Scholar] [CrossRef] [PubMed]
262. Kelliher, T.; Starr, D.; Richbourg, L.; Chintamanani, S.; Delzer, B.; Nuccio, M.L.; Green, J.; Chen, Z.; McCuiston, J.; Wang, W.; et al. MATRILINEAL, a sperm-specific phospholipase, triggers maize haploid induction. Nature 2017, 542, 105–109. [Google Scholar] [CrossRef] [PubMed]
263. Gilles, L.M.; Khaled, A.; Laffaire, J.B.; Chaignon, S.; Gendrot, G.; Laplaige, J.; Bergès, H.; Beydon, G.; Bayle, V.; Barret, P.; et al. Loss of pollen-specific phospholipase NOT LIKE DAD triggers gynogenesis in maize. EMBO J. 2017, 36, 707–717. [Google Scholar] [CrossRef]
264. Yao, L.; Zhang, Y.; Liu, C.; Liu, Y.; Wang, Y.; Liang, D.; Liu, J.; Sahoo, G.; Kelliher, T. OsMATL mutation induces haploid seed formation in indica rice. Nat. Plants 2018, 4, 530–533. [Google Scholar] [CrossRef] [PubMed]
265. Liu, C.; Zhong, Y.; Qi, X.; Chen, M.; Liu, Z.; Chen, C.; Tian, X.; Li, J.; Jiao, Y.; Wang, D.; et al. Extension of the in vivo haploid induction system from diploid maize to hexaploid wheat. Plant Biotechnol. J. 2019, 18, 316–318. [Google Scholar] [CrossRef] [PubMed]
266. Zhong, Y.; Liu, C.; Qi, X.; Jiao, Y.; Wang, D.; Wang, Y.; Liu, Z.; Chen, C.; Chen, B.; Tian, X.; et al. Mutation of ZmDMP enhances haploid induction in maize. Nat. Plants 2019, 5, 575–580. [Google Scholar] [CrossRef]
267. Zhong, Y.; Chen, B.; Li, M.; Wang, D.; Jiao, Y.; Qi, X.; Wang, M.; Liu, Z.; Chen, C.; Wang, Y.; et al. A DMP-triggered in vivo maternal haploid induction system in the dicotyledonous Arabidopsis. Nat. Plants 2020, 6, 466–472. [Google Scholar] [CrossRef]
268. Li, Y.; Lin, Z.; Yue, Y.; Zhao, H.; Fei, X.; E, L.; Liu, C.; Chen, S.; Lai, J.; Song, W. Loss-of-function alleles of ZmPLD3 cause haploid induction in maize. Nat. Plants 2021, 7, 1579–1588. [Google Scholar] [CrossRef] [PubMed]
269. Impens, L.; Lorenzo, C.D.; Vandeputte, W.; Wytynck, P.; Debray, K.; Haeghebaert, J.; Herwegh, D.; Jacobs, T.B.; Ruttink, T.; Nelissen, H.; et al. Combining multiplex gene editing and doubled haploid technology in maize. New Phytol. 2023, 239, 1521–1532. [Google Scholar] [CrossRef] [PubMed]
270. Kuppu, S.; Ron, M.; Marimuthu, M.P.A.; Li, G.; Huddleson, A.; Siddeek, M.H.; Terry, J.; Buchner, R.; Shabek, N.; Comai, L.; et al. A variety of changes, including CRISPR/Cas9-mediated deletions, in CENH3 lead to haploid induction on outcrossing. Plant Biotechnol. J. 2020, 18, 2068–2080. [Google Scholar] [CrossRef]
271. Lv, J.; Yu, K.; Wei, J.; Gui, H.; Liu, C.; Liang, D.; Wang, Y.; Zhou, H.; Carlin, R.; Rich, R.; et al. Generation of paternal haploids in wheat by genome editing of the centromeric histone CENH3. Nat. Biotechnol. 2020, 38, 1397–1401. [Google Scholar] [CrossRef]
272. Singh, M.; Kumar, M.; Albertsen, M.C.; Young, J.K.; Cigan, A.M. Concurrent modifications in the three homeologs of Ms45 gene with CRISPR-Cas9 lead to rapid generation of male sterile bread wheat (Triticum aestivum L.). Plant Mol. Biol. 2018, 97, 371–383. [Google Scholar] [CrossRef] [PubMed]
273. Okada, A.; Arndell, T.; Borisjuk, N.; Sharma, N.; Watson-Haigh, N.S.; Tucker, E.J.; Baumann, U.; Langridge, P.; Whitford, R. CRISPR/Cas9-mediated knockout of Ms1 enables the rapid generation of male-sterile hexaploid wheat lines for use in hybrid seed production. Plant Biotechnol. J. 2019, 17, 1905–1913. [Google Scholar] [CrossRef]
274. Du, M.; Zhou, K.; Liu, Y.; Deng, L.; Zhang, X.; Lin, L.; Zhou, M.; Zhao, W.; Wen, C.; Xing, J.; et al. A biotechnology-based male-sterility system for hybrid seed production in tomato. Plant J. 2020, 102, 1090–1100. [Google Scholar] [CrossRef] [PubMed]
275. Chen, G.; Zhou, Y.; Kishchenko, O.; Stepanenko, A.; Jatayev, S.; Zhang, D.; Borisjuk, N. Gene editing to facilitate hybrid crop production. Biotechnol. Adv. 2021, 46, 107676. [Google Scholar] [CrossRef] [PubMed]
276. Ye, M.; Peng, Z.; Tang, D.; Yang, Z.; Li, D.; Xu, Y.; Zhang, C.; Huang, S. Generation of self-compatible diploid potato by knockout of S-RNase. Nat. Plants 2018, 4, 651–654. [Google Scholar] [CrossRef]
277. Qin, X.; Li, W.; Liu, Y.; Tan, M.; Ganal, M.; Chetelat, R.T. A farnesyl pyrophosphate synthase gene expressed in pollen functions in S-RNase-independent unilateral incompatibility. Plant J. 2018, 93, 417–430. [Google Scholar] [CrossRef]
278. Chen, F.; Yang, Y.; Li, B.; Liu, Z.; Khan, F.; Zhang, T.; Zhou, G.; Tu, J.; Shen, J.; Yi, B.; et al. Functional Analysis of M-Locus Protein Kinase Revealed a Novel Regulatory Mechanism of Self-Incompatibility in Brassica napus L. Int. J. Mol. Sci. 2019, 20, 3303. [Google Scholar] [CrossRef] [PubMed]
279. Ma, C.; Zhu, C.; Zheng, M.; Liu, M.; Zhang, D.; Liu, B.; Li, Q.; Si, J.; Ren, X.; Song, H. CRISPR/Cas9-mediated multiple gene editing in Brassica oleracea var. capitata using the endogenous tRNA-processing system. Hortic. Res. 2019, 6, 20. [Google Scholar] [CrossRef] [PubMed]
280. Razzaq, A.; Wani, S.H.; Saleem, F.; Yu, M.; Zhou, M.; Shabala, S.; Foyer, C. Rewilding crops for climate resilience: Economic analysis and de novo domestication strategies. J. Exp. Bot. 2021, 72, 6123–6139. [Google Scholar] [CrossRef] [PubMed]
281. Ye, C.-Y.; Fan, L. Orphan Crops and their Wild Relatives in the Genomic Era. Mol. Plant 2021, 14, 27–39. [Google Scholar] [CrossRef] [PubMed]
282. Huang, X.; Huang, S.; Han, B.; Li, J. The integrated genomics of crop domestication and breeding. Cell 2022, 185, 2828–2839. [Google Scholar] [CrossRef]
283. Zhang, J.; Yu, H.; Li, J. De novo domestication: Retrace the history of agriculture to design future crops. Curr. Opin. Biotechnol. 2023, 81, 102946. [Google Scholar] [CrossRef]
284. Curtin, S.; Qi, Y.; Peres, L.E.P.; Fernie, A.R.; Zsögön, A. Pathways to de novo domestication of crop wild relatives. Plant Physiol. 2022, 188, 1746–1756. [Google Scholar] [CrossRef]
285. Li, T.; Yang, X.; Yu, Y.; Si, X.; Zhai, X.; Zhang, H.; Dong, W.; Gao, C.; Xu, C. Domestication of wild tomato is accelerated by genome editing. Nat. Biotechnol. 2018, 36, 1160–1163. [Google Scholar] [CrossRef]
286. Zsögön, A.; Čermák, T.; Naves, E.R.; Notini, M.M.; Edel, K.H.; Weinl, S.; Freschi, L.; Voytas, D.F.; Kudla, J.; Peres, L.E.P. De novo domestication of wild tomato using genome editing. Nat. Biotechnol. 2018, 36, 1211–1216. [Google Scholar] [CrossRef] [PubMed]
287. Yu, H.; Lin, T.; Meng, X.; Du, H.; Zhang, J.; Liu, G.; Chen, M.; Jing, Y.; Kou, L.; Li, X.; et al. A route to de novo domestication of wild allotetraploid rice. Cell 2021, 184, 1156–1170. [Google Scholar] [CrossRef] [PubMed]
288. Lemmon, Z.H.; Reem, N.T.; Dalrymple, J.; Soyk, S.; Swartwood, K.E.; Rodriguez-Leal, D.; Van Eck, J.; Lippman, Z.B. Rapid improvement of domestication traits in an orphan crop by genome editing. Nat. Plants 2018, 4, 766–770. [Google Scholar] [CrossRef] [PubMed]

Tan W, Wang Z, Liu L. The Continuous Improvement of the Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR)–CRISPR-Associated Protein System Has Led to Its Highly Efficient Application in Plants. Agriculture. 2025; 15(1):29. https://doi.org/10.3390/agriculture15010029
Перевод статьи «The Continuous Improvement of the Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR)–CRISPR-Associated Protein System Has Led to Its Highly Efficient Application in Plants» авторов Tan W, Wang Z, Liu L., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык

















































































































Комментарии (0)