Влияние засухи, жары, избыточного освещения и засоления на производство кофе: стратегии смягчения стресса с помощью селекции и питания растений
Абиотические стрессы серьезно нарушают физиологию растений на молекулярном, биохимическом и морфологическом уровнях, часто вызывая необратимые повреждения. Для обеспечения устойчивого производства кофе крайне важно понять, как такие неблагоприятные факторы окружающей среды, как засуха, жара, избыточное освещение и засоление, влияют на рост растений, и разработать стратегии для смягчения их последствий.
Аннотация
Несмотря на ограниченное количество исследований по данной теме, обобщение существующих знаний может дать ценную информацию о том, как кофейные растения реагируют на такие стрессы. В частности, критически важно понять, способны ли кофейные растения переносить ущерб, вызванный этими стрессами, и какие механизмы они для этого используют. Цель данного обзора — (i) обобщить ключевые выводы о влиянии засухи, жары, избыточного освещения и засоления на кофейные растения и их механизмах адаптации; и (ii) изучить селекцию и питание растений в качестве потенциальных стратегий для смягчения этих абиотических стрессов и повышения продуктивности кофе.
1. Введение
Кофе является одним из важнейших товаров в международной сельскохозяйственной торговле и представляет собой значительный источник дохода для многих стран, включая Бразилию — крупнейшего в мире производителя и экспортера кофе [1]. По этой причине растет обеспокоенность regarding производством кофе, поскольку эта культура считается рисковой, особенно с учетом предстоящих изменений климата, которые, по прогнозам, будут продолжаться на протяжении XXI века [2]. Прогнозируемые погодные изменения включают нарушения режима осадков, более частые засушливые периоды, высокие температуры и смещение географических регионов, пригодных для выращивания кофе [3].
Приблизительно 99% мирового производства кофе приходится на два вида: Арабику (Coffea arabica L., 1753) и Робусту (Coffea canephora Pierre ex A. Froehner, 1897), оба принадлежат к порядку Gentianales, семейству Rubiaceae [4]. Арабика лучше растет при температурах от 18 до 22 °C, а Робуста — от 22 до 28 °C. Повреждение качества зерен и урожая у обоих видов может происходить за пределами этих температурных диапазонов [5], что свидетельствует о значительной чувствительности к изменениям климатических условий. Кроме того, повторяющиеся периоды засухи, сопровождающиеся избыточным освещением и/или эпизодами заморозков, вызывали нестабильность ежегодного предложения кофе [6,7], влияя на развитие растений, цветение, завязывание плодов и продуктивность как Арабики, так и Робусты [4,8].
Засушливые условия, избыточное освещение, высокие температуры, недостаточная доступность питательных веществ в почве и засоление являются наиболее важными абиотическими стрессами в сельском хозяйстве [9,10,11,12,13]. Абиотические стрессы влияют на физиологическое состояние растений, оказывая заметное воздействие на молекулярном, биохимическом и морфологическом уровнях и могут вызывать необратимые повреждения, представляя собой основные причины потерь урожайности [11]. Эта тема весьма актуальна для производства кофе, но остается менее изученной по сравнению с исследованиями на других культурах (Рисунок 1A). Усилия по пониманию того, как кофейные растения реагируют на различные абиотические стрессы, и разработке стратегий смягчения их последствий значительно активизировались с 2006 года (Рисунок 1B).
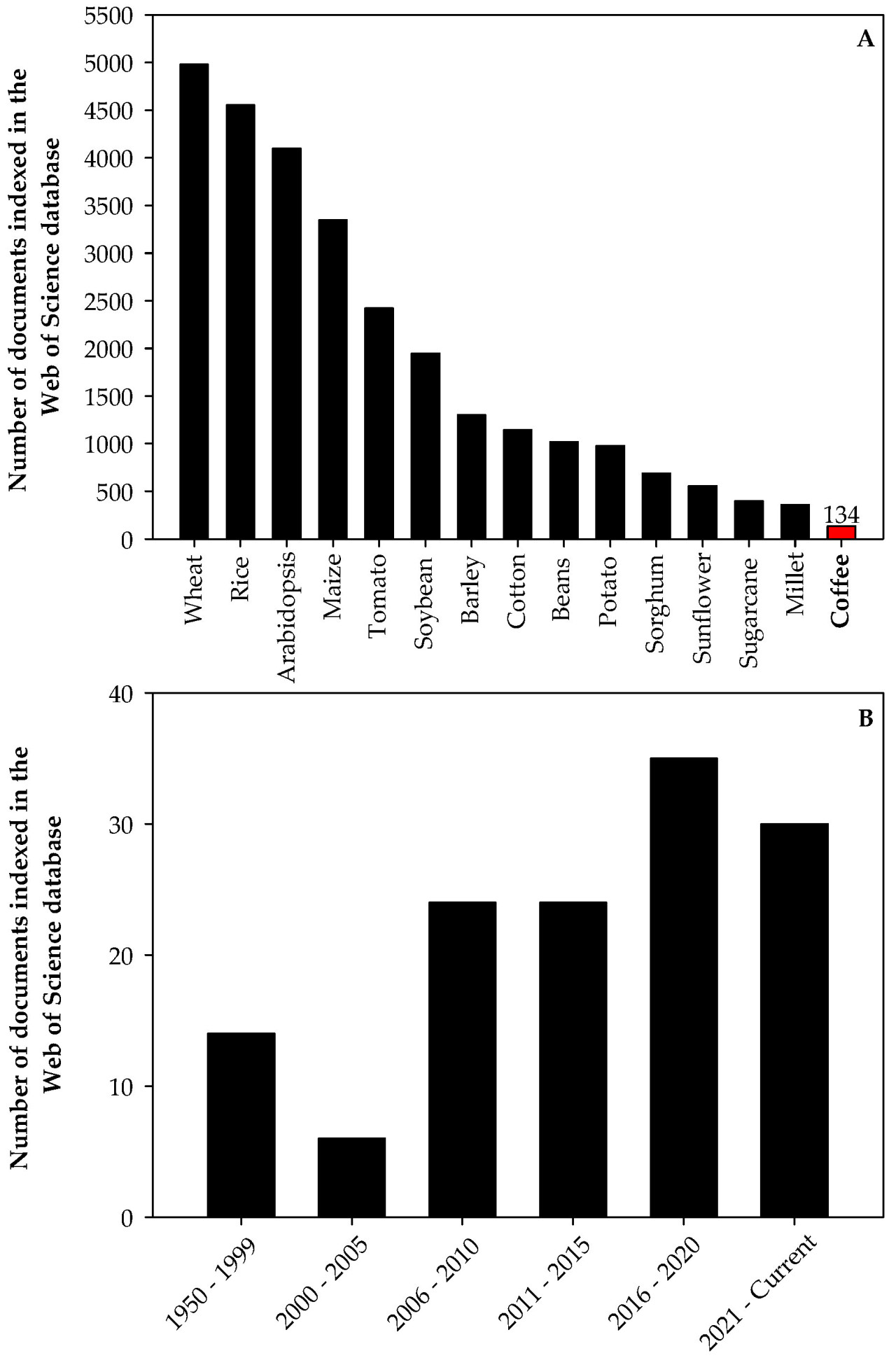
Рисунок 1. Количество документов, индексированных в базе данных Web of Science, о стрессе у наиболее важных экономических культур (за исключением рода Arabidopsis, который используется в качестве референтного модельного растения) в мире (A) и количество документов о стрессе у кофе (B).
Основываясь на приведенных данных, наши цели в этом кратком обзоре заключались в том, чтобы (i) обобщить основные результаты, касающиеся воздействия засухи, жары, избыточного освещения и засоления на производство кофе и основных механизмов, используемых растениями для противодействия неблагоприятным факторам окружающей среды; и (ii) рассмотреть селекцию и питание растений в качестве стратегий для смягчения абиотического стресса в кофейных насаждениях.
2. Факторы абиотического стресса и их связь с ростом кофейного растения
В этом разделе мы описываем в общем виде основные эффекты засухи, жары, избыточного освещения и засоления на рост растений и представляем основные механизмы, используемые кофейными растениями для борьбы с неблагоприятными факторами окружающей среды (Рисунок 2).

Рисунок 2. Сводка симптомов стресса, индуцированных засухой, жарой и избыточным освещением, а также засолением у кофейных растений, а также механизмов толерантности и стратегий для улучшения устойчивости растений к этим стрессам.
2.1. Стресс от засухи у кофе
Большинство исследований по абиотическому стрессу у кофе сосредоточено на стрессе от засухи и было опубликовано за последние 15 лет (Рисунок 3). Засуха считается наиболее стрессовым фактором окружающей среды, влияющим на рост и урожайность растений, и ожидается, что ее негативные последствия станут еще более заметными из-за изменения климата [2,14]. В этом сценарии поддержание роста и урожайности сельскохозяйственных культур в одних из самых стрессовых условий, таких как водный дефицит, представляет собой одну из основных проблем современного сельского хозяйства [15].
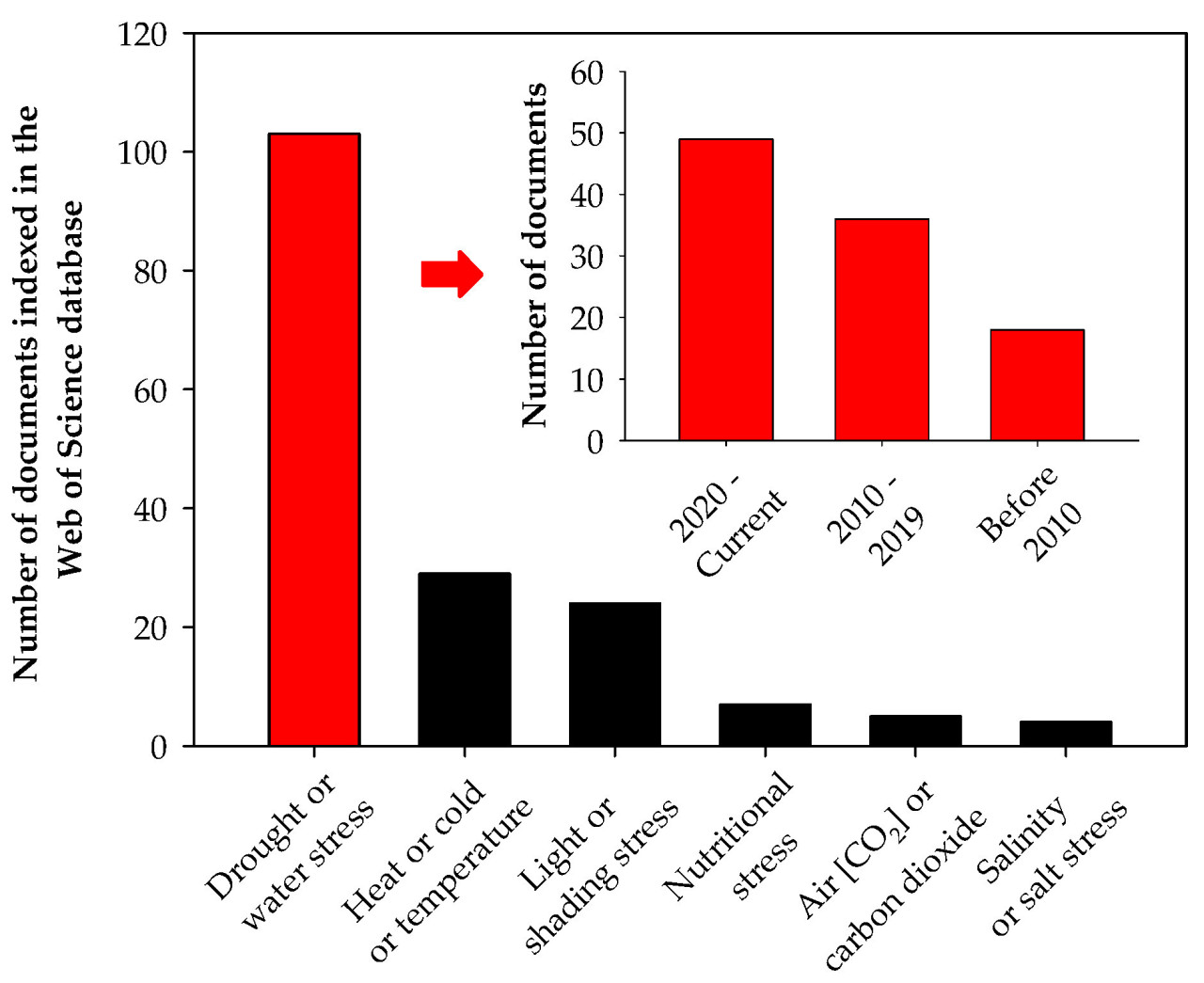
Рисунок 3. Количество документов, индексированных в базе данных Web of Science, касающихся засухи или водного стресса; жары, холода или температурного стресса; светового стресса или затенения; пищевого стресса; стресса от концентрации CO₂ в воздухе; и стресса от засоления или засоленности у кофе.
Водный дефицит у растений характеризуется дисбалансом между поглощением воды корнями и потерей воды при транспирации листьями; т.е. дефицит воды возникает, когда транспирация с поверхности листа превышает поглощение воды корнями [16]. Многие факторы могут вызывать водный дефицит у растений, включая факторы окружающей среды, такие как недостаточное количество осадков, высокая испаряемость, понижение уровня грунтовых вод, уплотнение почвы и удержание воды почвенными частицами [17], а также связанные с растением, например, состояние корневой системы. Период аномально сухой погоды, приводящий к дефициту почвенной влаги и, как следствие, к водному дефициту растения, определяется как засуха. Основные симптомы синдрома засухи у растений варьируются в зависимости от вида, интенсивности стресса, стадии развития, условий роста и взаимодействия с другими факторами окружающей среды. Основные морфологические симптомы обычно включают потерю тургора листьев, торможение роста клеток, поникание, увядание, этиоляцию, пожелтение и преждевременный листопад [18,19,20]. У кофейных растений основные симптомы стресса от засухи представлены в Таблице 1 и на Рисунке 4.

Рисунок 4. Симптомы стресса от засухи у кофейных растений включают потерю тургора листьев (a,c,d), увядание (a,c,d), поникание (b,e), пожелтение (d,e), некроз (b–e) и преждевременный листопад (b,e).
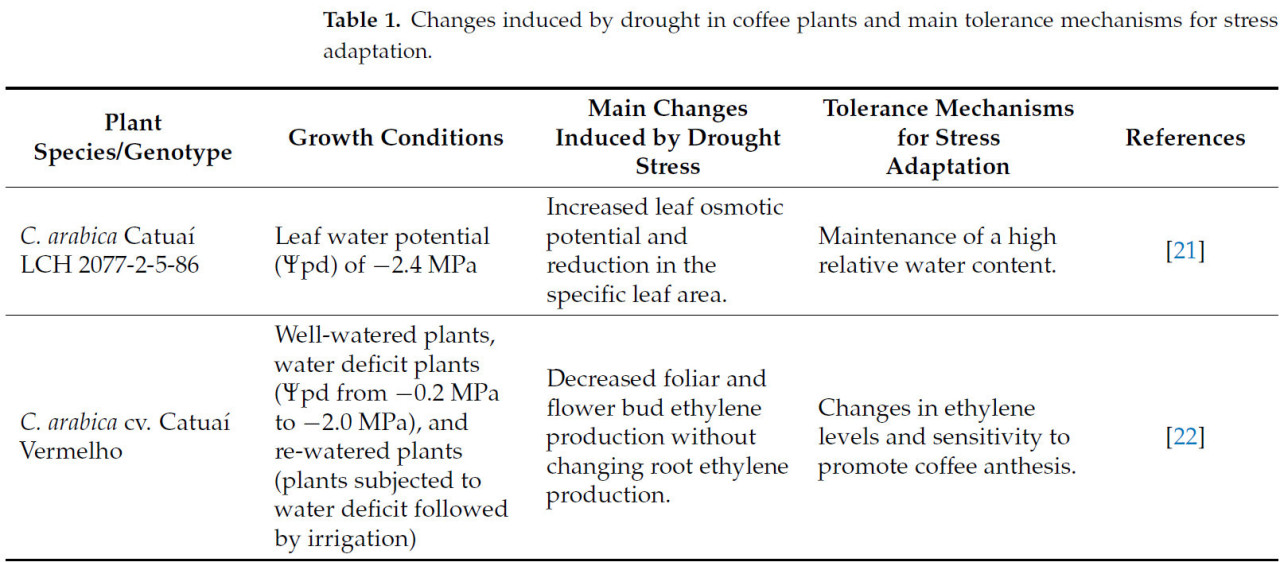
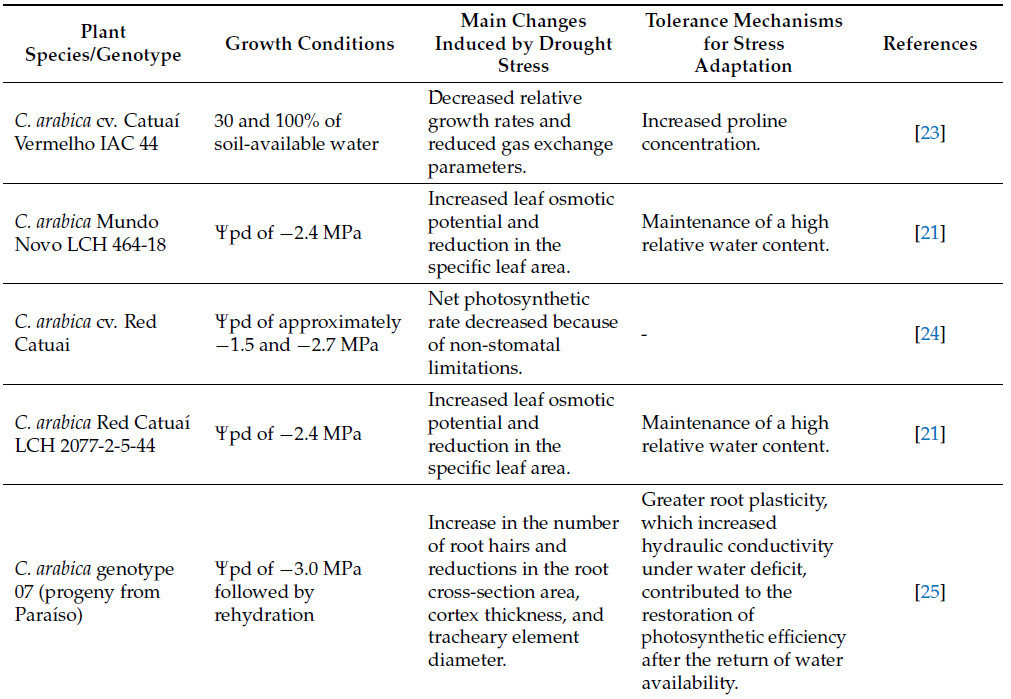
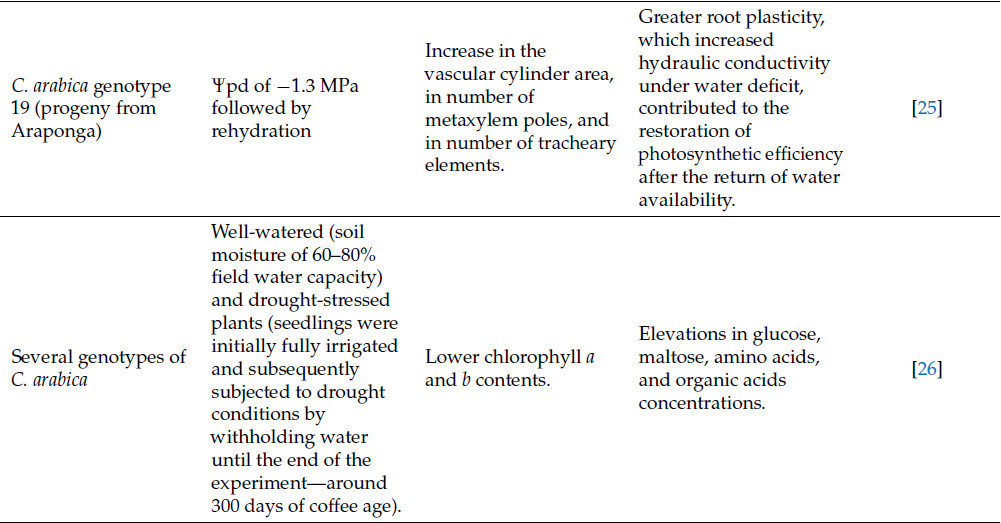
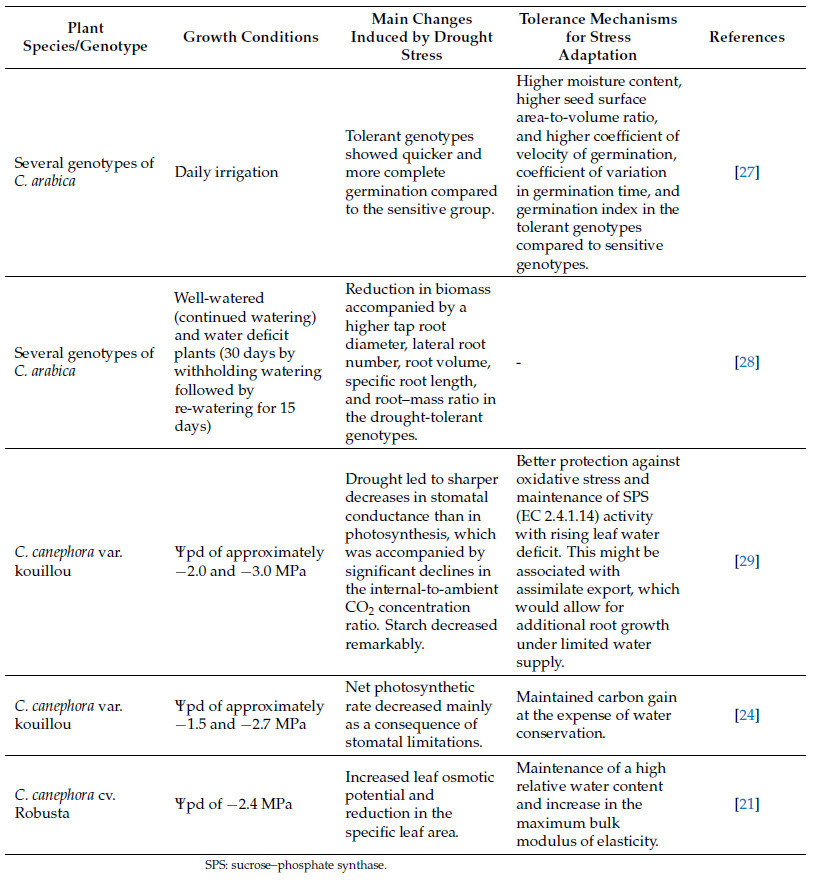
Таблица 1. Изменения, индуцированные засухой у кофейных растений, и основные механизмы толерантности для адаптации к стрессу.
В очень упрощенной цепи реакций стресс от водного дефицита вызывает закрытие устьиц в ответ на снижение водного потенциала ксилемы, предотвращая избыточную потерю водяного пара из листьев в атмосферу. Однако снижение транспирации повышает температуру листьев и уменьшает приток CO₂ путем диффузии в мезофилл листьев. При непрерывном поглощении фотосинтетически активной радиации (ФАР) этими листьями с низкой транспирацией биохимическая энергия (НАДФН и АТФ) из фотохимической фазы фотосинтеза будет меньше использоваться биохимической фазой из-за меньшей доступности CO₂ для рибулозо-1,5-бисфосфаткарбоксилазы/оксигеназы (RuBisCO), вызывая перенасыщение электронной транспортной цепи в тилакоидах хлоропластов. Из-за биофизической природы поглощения ФАР, комплекса выделения кислорода и возбуждения реакционного центра в фотосистеме II (PSII) свободные электроны от фотолиза воды реагируют со свободным кислородом в просвете хлоропласта и вызывают накопление активных форм кислорода (АФК), которое должно уравновешиваться антиоксидантной системой (AOS). Если AOS не может нейтрализовать АФК, окислительный стресс может разрушить клетки, нанося тканям необратимый ущерб [30,31].
Кофейные растения, подверженные стрессу от засухи, используют следующие стратегии для смягчения возможного ущерба: (i) уменьшение площади листа для снижения транспирации; (ii) поддержание движения сахаров от источников к стокам; (iii) изменение морфологии листьев и развития корней; (iv) сигнализация от побега к корню о водном статусе; (v) максимизация фотосинтеза по сравнению с транспирацией; и (vi) замена поврежденных АФК белков и фосфолипидов [25]. Было показано, что у толерантных клонов C. canephora транскрипционная память может способствовать устойчивости к засухе во время циклов воздействия засухи [32]. Более конкретно, подход RNAseq выявил дифференциально экспрессируемые гены у толерантных (826) и чувствительных (135) клонов и их обогащенные категории. Такой анализ показал, что чувствительные клоны могут запускать реакцию на окислительный стресс, возможно, приводящую к запрограммированной гибели клеток, при воздействии множественных эпизодов засухи [32]. Однако акклиматизация толерантных растений, по-видимому, включает вторичный антиоксидантный метаболизм и ответ на абсцизовую кислоту (ABA). Дополнительно было идентифицировано 49 генов памяти в толерантных клонах, в основном связанных с путем ABA, сворачиванием белков и биотическим стрессом [32].
Большинство исследований ответов растений на засуху рассматривают ее как единичное, изолированное событие, в отличие от природных условий, где повторяющиеся эпизоды засухи часто совпадают с другими факторами окружающей среды. Чувствительные и толерантные к засухе клоны C. canephora, подвергнутые множественным циклам засухи, развили дифференцированную акклиматизацию, которая потенцировала их защитные механизмы [14], с более значительной акклиматизацией у толерантных клонов по сравнению с чувствительными. Растения, подвергнутые множественным засушливым событиям, показали более высокие скорости фотосинтеза и высокую активность RuBisCO и других ферментов, связанных с углеродным и антиоксидантным метаболизмом, по сравнению с теми, которые подвергались засухе впервые. Дополнительно, акклиматизация к множественным засушливым событиям включала экспрессию тренируемых генов, связанных с устойчивостью к засухе, а также была связана с глубоким репрограммированием метаболитов с согласованными изменениями в центральных метаболических процессах, таких как дыхание и фотодыхание, что позволяло им находиться в «состоянии готовности», чтобы успешно справляться с дальнейшими засушливыми событиями [14]. Как дыхание, так и фотодыхание могут зависеть от концентрации CO₂ в воздухе, поэтому крайне важно понять, как повышенная концентрация CO₂ в воздухе, вызванная изменением климата, может повлиять на производство кофе.
Повышенная концентрация CO₂ в воздухе, вероятно, смягчает стресс от засухи. Повышенная [CO₂] часто снижает устьичную проводимость и плотность устьиц листа [33,34]. Дополнительно, арабика, выращиваемая в течение 4 лет при высокой [CO₂], показала уменьшение площади листовой поверхности кроны, связанное с улучшением чистого фотосинтеза [35]. Этот факт связывают с повышенной активностью RuBisCO и снижением оксигеназной активности, что уменьшает фотодыхание и продукцию АФК [34,36]. У кофе, подверженного стрессу от засухи, повышенная [CO₂] улучшила скорость ассимиляции углерода при неизменной устьичной проводимости, увеличила эффективность использования воды, скорость дыхания и накопление биомассы, а также снизила скорость фотодыхания и окислительный стресс, независимо от полива. Повышенная [CO₂] также способствовала ключевым аллометрическим изменениям, связанным с устойчивостью к засухе у кофе, таким как большее распределение биомассы в сторону корней с увеличением длины корней [10].
2.2. Стресс от жары и избыточного освещения у кофе
Как сидячие организмы, растения в значительной степени страдают от теплового стресса, который ограничивает их рост, метаболизм и урожайность во всем мире [30]. Интерес к пониманию ответных реакций растений на тепловой стресс растет, поскольку существует обеспокоенность тем, как экосистемы будут затронуты изменением климата. Прогнозируется, что глобальные среднегодовые температуры повысятся на 0,3–4,8 °C к 2100 году [2], и многие районы, вероятно, прогреются выше среднемирового показателя с увеличением количества волн жары и последствий, усугубляемых одновременными засушливыми явлениями.
Первичные реакции растений на тепловой стресс включают изменения в их морфологии, такие как увеличение корневой системы и уменьшение количества и проводимости устьиц. Дополнительно, листья скручиваются, складываются и уменьшают свою площадь, чтобы избежать потери воды при эвапотранспирации [37,38]. Согласно Лиме и др. [9], клеточная стенка листа кофейных растений, подвергшихся тепловому стрессу, отличается от таковой у растений без стресса по составу моносахаридов и профилям молекулярных масс полисахаридов. Тепловой стресс увеличил содержание гемицеллюлозы в листе, тогда как содержание пектина уменьшилось почти на 50% по сравнению с растениями в контрольных условиях. Тем не менее, конечный выход нерастворимого остатка не пострадал в конце тепловой обработки. Также наблюдались изменения в палисадных клетках и ультраструктурные повреждения хлоропластов. Эти данные показывают, что химический профиль полимеров клеточной стенки кофе и структурная анатомия клеток изменяются под воздействием теплового стресса [9].
Высокие температуры в природе часто сопровождаются избыточным освещением [39], что может ограничивать рост кофе, поскольку кофейные растения традиционно известны тем, что их нужно выращивать в тени. Однако кофе можно выращивать и при полном солнечном освещении без потери урожайности. Мартинс и др. [40] показали, что некоторые кофейные растения могут настраивать свой метаболический аппарат к избыточному свету за счет заметного увеличения антиоксидантной способности, связанного с усиленным потреблением восстановительных эквивалентов. Фотодыхание и альтернативные пути являются основными, ответственными за потребление восстановителей при избыточном освещении. В таких условиях было обнаружено, что первичный и вторичный метаболизм перепрограммируются для снижения регуляции промежуточных продуктов цикла трикарбоновых кислот (TCA) и повышения регуляции аминокислот, сахаров и сахарных спиртов, полиаминов и флавоноидов [40]. Эти данные свидетельствуют, что метаболические изменения в первую очередь связаны с избеганием окислительного стресса, а не представляют собой адаптацию, позволяющую растениям использовать дополнительный свет для улучшения своей фотосинтетической производительности.
Хотя некоторые кофейные растения могут настраивать свой метаболический аппарат к условиям избыточного освещения [40], другие растения или генотипы кофе чувствительны к жаре и избыточному свету. Некоторые из основных визуальных симптомов, вызванных избыточным светом и жарой, включают ожоги, солнечные ожоги и солнечную чесотку (Рисунок 5). Солнечный ожог происходит, когда кофейное растение поглощает большое количество энергии от солнца и не может ее рассеять, что приводит к окислительному повреждению. Этот тип повреждения обычно наблюдается после обрезки и посадки молодых деревьев, так как интенсивный солнечный свет (и жара) вызывает разрушение хлорофилла в листе [41]. Первоначальное повреждение проявляется в виде бледных, обесцвеченных или выцветших участков, которые становятся коричневыми и некротическими, пока finalmente не опадают. Если только повреждение не является чрезвычайно сильным, растения обычно восстанавливаются после повреждений от солнечных ожогов, но может произойти задержка вегетативного и/или репродуктивного роста, влияющая на урожайность. К сожалению, существует лишь несколько исследований, оценивающих реакцию кофейных растений на стрессы от жары и избыточного света (Таблица 2), что затрудняет применение эффективных стратегий для смягчения этих стрессовых условий.

Рисунок 5. Начальные симптомы солнечного ожога на листьях и плодах кофе проявляются как бледные, обесцвеченные или выцветшие участки, которые становятся коричневыми (a,c) и некротическими (b,d,e).
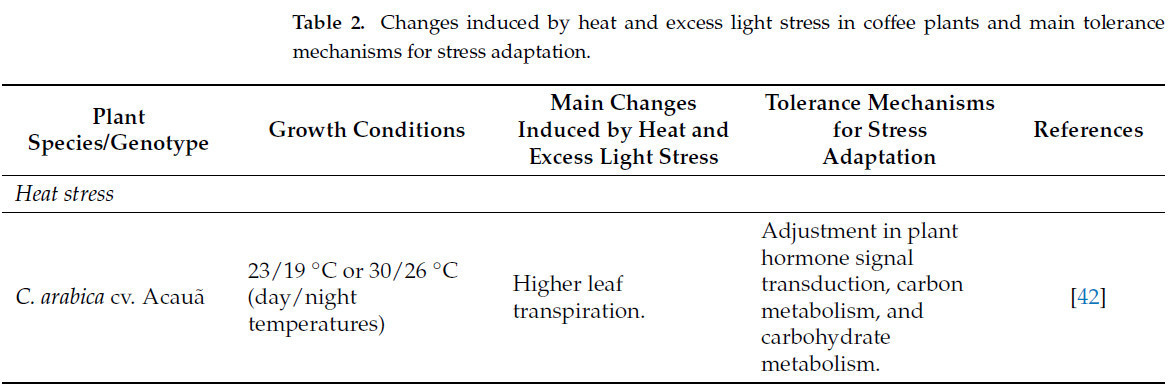
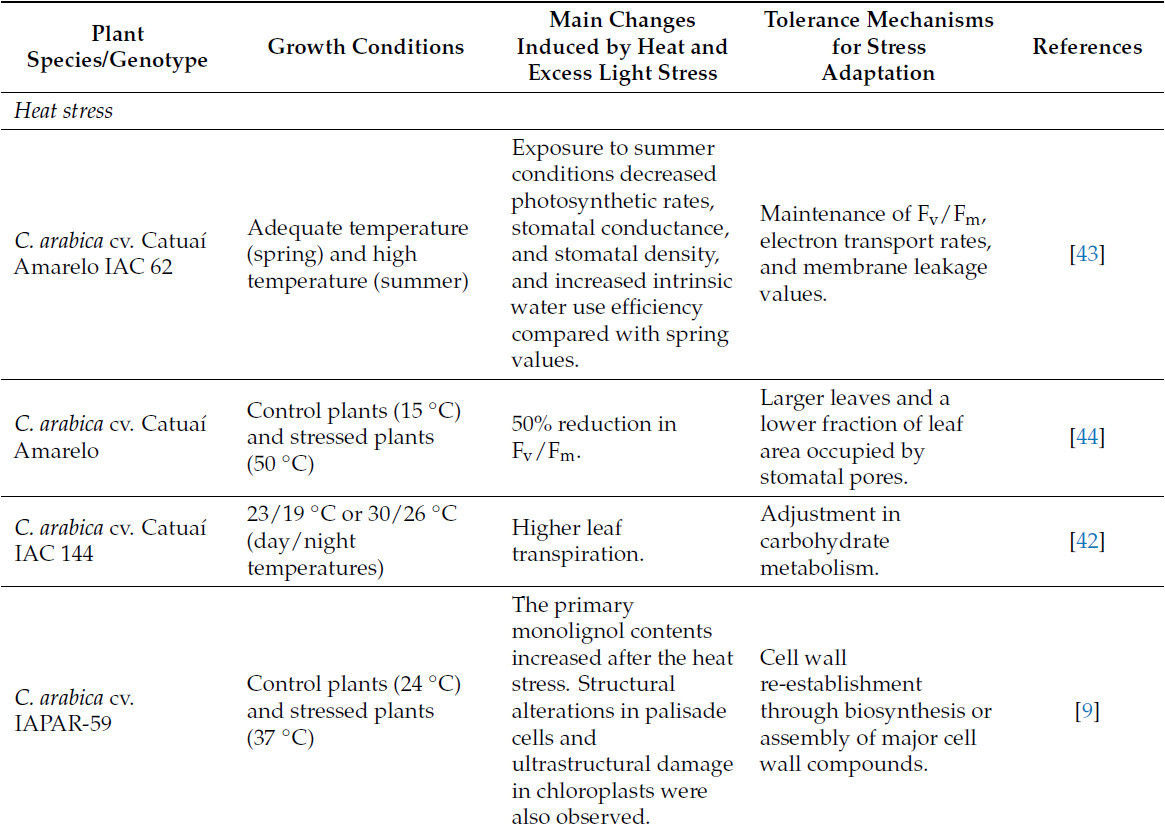
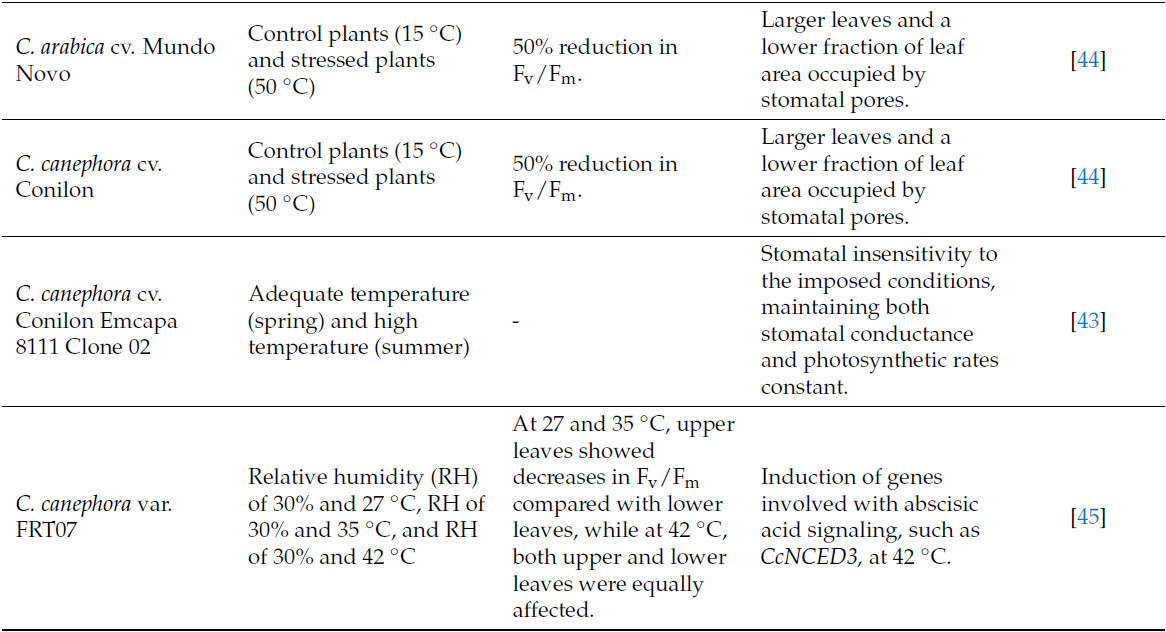
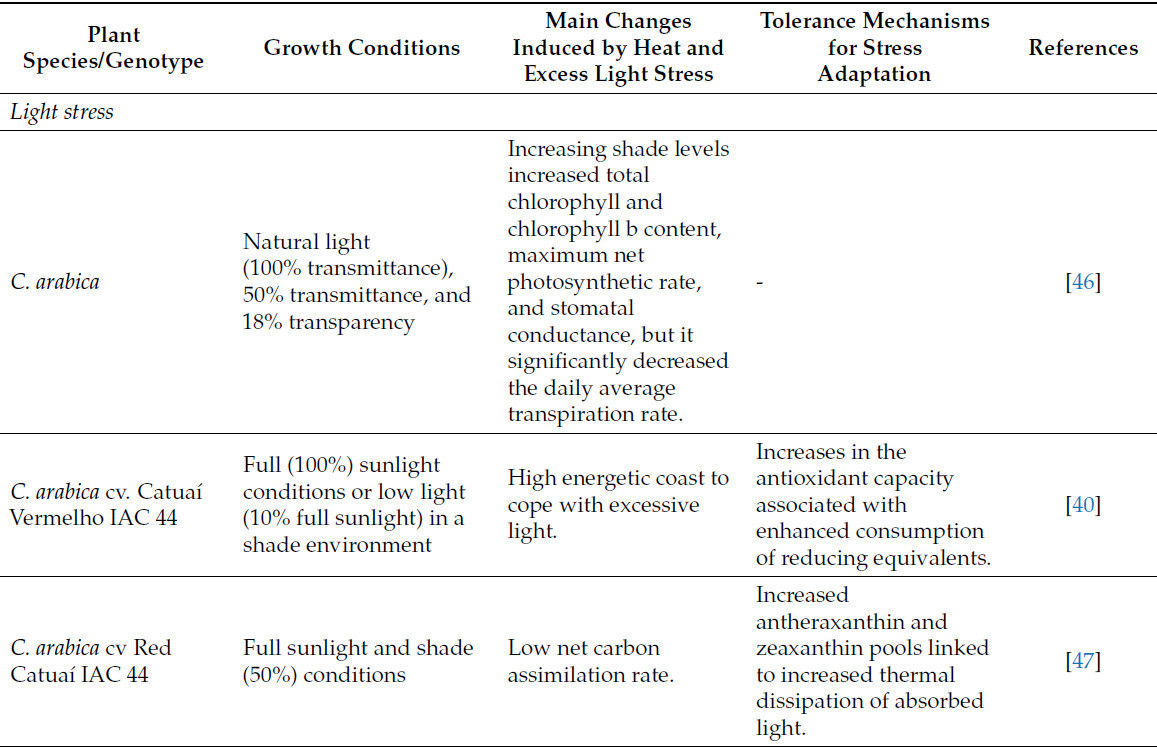

Таблица 2. Изменения, индуцированные стрессом от жары и избыточного света у кофейных растений, и основные механизмы толерантности для адаптации к стрессу.
2.3. Стресс от засоления у кофе
Засоление является одним из основных абиотических стрессов в сельском хозяйстве, вызывающим снижение роста, физиологические аномалии и снижение урожайности культур во всем мире [49]. Солевой стресс остается значительным фактором, ограничивающим рост, в основном в засушливых и полузасушливых зонах [50]. Засоленность почвы — это мера концентрации всех растворимых солей в почвенном растворе, которая обычно выражается как электропроводность (ЕС) [51]. Почва считается засоленной, когда ее pH ниже 8,5, ЕС выше 4,0 дСм м⁻¹ (что равно 40 мМ NaCl), а процент обменного натрия (ESP) превышает 15% [52].
Засоление почвы может происходить естественным путем или в результате деятельности человека. Естественные геологические, гидрологические и почвенные процессы составляют некоторые естественные причины, но изменение климата и управление водными ресурсами могут ускорить засоление. Как в засушливых, так и в полузасушливых землях эвапотранспирация играет жизненно важную роль в педогенезе засоленных и щелочных почв. Засоление почвы, вызванное антропогенной деятельностью, в основном связано с неправильными методами орошения. Часто для орошения используется вода плохого качества, поэтому со временем соли накапливаются в почве, если управление системой орошения не обеспечивает вымывание солей из почвенного профиля. Другими антропогенными причинами являются обезлесение, накопление солей, переносимых по воздуху или водой, в почвах, загрязнение химическими веществами и перевыпас [53]. По оценкам, более 100 стран сталкиваются с засоленными условиями в почве или грунтовых водах [54].
Корни обычно являются первым органом, который ощущает солевой сигнал после воздействия на растения засоленных условий [38]. Модификации архитектуры корней в ответ на засоление опосредованы подавлением деления клеток, инициации, удлинения или перенаправления роста от соли. Самая ранняя визуальная реакция растения на засоление — снижение скорости расширения листа, за которым следует прекращение расширения по мере усиления стресса. Также воздействие засоления приводит к закрытию устьиц, что снижает интенсивность фотосинтеза из-за уменьшения устьичной проводимости, которая ограничивает доступ CO₂ для цикла Кальвина-Бенсона [54,55].
У C. arabica засоление вызвало структурные повреждения в клетках мезофилла и изменения в организации пектинов, гемицеллюлоз и составе лигнина в клеточной стенке [56]. Помимо наблюдаемых изменений в составе, размер фракций гемицеллюлозы из листьев кофе увеличился при засолении, что свидетельствует об увеличении сшивки между полисахаридами. Установление более прочных поперечных связей между полисахаридами и лигнификация клеточных стенок могут способствовать упрочнению клетки, приводя к ограничению диффузии, таким образом действуя как барьер для проникновения соли. Увеличение отрицательно заряженных полисахаридов клеточной стенки, как указывалось, играет роль в борьбе с солью, облегчая транспорт ионов при высоких концентрациях соли. Однако также возможно, что отрицательно заряженные полисахариды задерживают проникновение ионов натрия (Na⁺) [56].
Количество исследований, оценивающих влияние засоления на кофейные растения, очень мало (Рисунок 3), но изменение климата может ускорить засоление, изменяя доступность воды [49]. Поэтому необходимо проводить новые исследования, чтобы лучше понять, как кофейные растения справляются с засолением, и найти ранние решения этой проблемы.
3. Стратегии для повышения устойчивости кофейных растений к абиотическому стрессу
Частота упомянутых выше стрессовых факторов будет прогрессивно увеличиваться в будущем из-за роста числа событий, связанных с изменением климата. Таким образом, крайне важно разработать или лучше понять стратегии для смягчения неблагоприятных эффектов этих проблем в сельском хозяйстве и повышения устойчивости и урожайности культур. В последние годы было протестировано несколько методов, таких как разработка генотипов, более толерантных к изменению климата, с помощью применения традиционных и современных методов селекции, а также использование регуляторов роста растений, осмопротекторов, управления питанием и водой, сроков посадки, прайминга семян, микробной обработки семян и арбускулярной микоризы. Применение биоугля, каолина, хитозана, суперабсорбента, экстракта дрожжей и экстракта морских водорослей также оценивалось как инструменты для смягчения стресса у растений [57].
Свежая кофейная шелуха для биоугля (BC) была протестирована как многообещающая альтернатива, поскольку ее можно вносить в почву для улучшения физических, химических и микробиологических характеристик, одновременно повышая урожайность культур [58,59]. Свойства почвы, такие как удержание воды [60], pH [61,62], гидравлическая проводимость и доступность питательных веществ, увеличиваются с применением BC [63]. Оценивая четыре различные дозы биоугля (0, 4, 8 и 16 т га⁻¹) в комбинации с прогрессивным снижением орошения (25, 50, 75 и 90% воды, потерянной через эвапотранспирацию), эти авторы показали, что применение 8 т га⁻¹ BC ослабляло эффекты умеренного водного дефицита у кофейных растений. Снижение орошения негативно влияло на максимальную квантовую выходность фотосистемы II (Fv/Fm), газообмен листьев, биомассу и водный статус. С другой стороны, применение 8 т га⁻¹ BC увеличивало фотосинтез у хорошо орошаемых растений (~6 мкмоль м⁻² с⁻¹) и менее орошаемых растений (~3,5 мкмоль м⁻² с⁻¹) по сравнению с растениями, выращенными без применения BC (хорошо орошаемые растения: ~3,9 мкмоль м⁻² с⁻¹; и менее орошаемые растения: ~1,8 мкмоль м⁻² с⁻¹). Авторы заключили, что использование BC можно рекомендовать для производства кофе не только для связывания углерода и возвращения его в почву, но и для ослабления эффектов умеренного водного дефицита [64]. Другие продукты вносились в почву или использовались в виде листовых подкормок для минимизации стресса, вызванного засухой. Было показано, что мелатонин (фитомелатонин) играет интересную роль в толерантности к абиотическому стрессу [65,66]. Мелатонин усиливал активность антиоксидантных ферментов и снижал генерацию свободных радикалов, перекиси водорода (H₂O₂) и малонового диальдегида у яблони, винограда, кукурузы, подсолнечника, томата и пшеницы [67]. Обработка мелатонином повышает толерантность растений к водному дефициту [68,69] за счет поддержания тургорного давления в листьях на удовлетворительном уровне [70]. Что касается реакции кофе, Кампос и др. [71] обнаружили, что 300 мкМ мелатонина, внесенного в почву через 4 недели после пересадки, способствовали повышению устойчивости к засухе за счет увеличения роста корней и снижения водного потенциала листьев. Поддержание водного потенциала листьев мелатонином могло благоприятствовать газообмену и, следовательно, биохимическим реакциям, поскольку среди растений, подвергшихся водному дефициту, те, которые обрабатывались мелатонином, показали более высокую устьичную проводимость и более высокие скорости фотосинтеза и транспирации, позволяя большему снабжению ассимилятами растущих тканей. Эти результаты подчеркивают, что мелатонин может повышать устойчивость кофейных растений к засухе. Другие стратегии, такие как предварительное воздействие на сеянцы кофе дефицитного орошения [72] или прививка чувствительных генотипов на толерантные подвои [73], также были протестированы и показали перспективные результаты в повышении устойчивости растений к абиотическому стрессу, особенно к водному дефициту. Тем не менее, в этом разделе мы выделяем селекцию растений и питание растений как потенциальные стратегии, помогающие кофейным растениям расти в стрессовых условиях.
3.1. Селекция растений на устойчивость к абиотическому стрессу
Генетическая селекция кофе — это ключевая практика, которая направлена на разработку генотипов кофе с желательными агрономическими признаками, такими как большая урожайность, устойчивость к болезням и вредителям, лучшее качество зерен и адаптация к различным климатическим условиям. Генотипы кофе значительно различаются по своим реакциям на условия окружающей среды [27,28]. Это изменение связано с генетическими различиями, влияющими на то, как каждый генотип взаимодействует с осадками, температурой и плодородием почвы. Существуют свидетельства значительной межиндивидуальной изменчивости в реакции на высокие температуры [74], что могло бы быть полезно учитывать вместе с данными относительно устойчивости к водному стрессу, поскольку средняя глобальная температура непрерывно повышается [2]. Однако улучшение кофе с помощью традиционной селекции происходит медленно из-за многолетней природы растения. С другой стороны, за последние 30 лет был достигнут значительный прогресс в биотехнологии кофе, создав множество возможностей в этой области исследований [75]. В настоящее время программы селекции кофе в основном направлены на идентификацию генов устойчивости к стрессам окружающей среды и транскрипционных факторов (TFs), чтобы позволить разработку новых генотипов. Пиросеквенирование РНК, экстрагированной из апикальных меристем побегов толерантных и восприимчивых к засухе генотипов C. arabica, подвергнутых стрессу от засухи, показало, что экспрессия гена CaSTK1 (протеинкиназа), который кодирует предполагаемый ответ на окислительный стресс (серин/треонин протеинкиназа), была сильно индуцирована засухой у толерантного генотипа [76]. Аналогичные профили наблюдались для CaSAMT1 (SAM-зависимая метилтрансфераза), гена, кодирующего предполагаемую S-аденозил-L-метионин-зависимую метилтрансферазу, CaSLP1 (развитие растения), CaMAS1 (биосинтез ABA) и генов nsLTP (белок переноса липидов) [76]. Эти авторы предположили, что nsLTP может быть связан с более толстой кутикулой, наблюдаемой на абаксиальной поверхности листа толерантных генотипов по сравнению с восприимчивыми. Ген GolS является одним из основных генов, вовлеченных в биосинтез олигосахаридов семейства раффинозы (RFOs), которые включают раффинозу, стахиозу и вербаскозу. У кофе были идентифицированы и охарактеризованы три изоформы GolS (CaGolS1, 2 и 3) в условиях абиотического стресса. Более того, было указано, что транскрипты CaM6PR, CaPMI и CaMTD, вовлеченные в биосинтез маннита, модулируются различными способами в ответ на абиотический стресс, такой как засуха, жара и засоление [77]. Гены, регулирующие RFO в кофе, необходимы для поддержания клеточной осмопротекции при засухе и засолении, что делает их перспективными генами для программ генетической селекции [77].
Транскрипционные факторы, такие как белки, связывающие dehydration responsive element (DREB), очень важны для растений, сталкивающихся с стрессами окружающей среды, поскольку они активируют экспрессию многих генов, вовлеченных в защиту клеток, детоксикацию и репарацию, среди прочего [78]. Регуляция генов, подобных DREB кофе, изучалась у кофейных растений, подвергнутых воздействию холода, жары, низкой относительной влажности, экзогенного внесения ABA и стресса от сильного света [79]. У C. arabica ген CaERF017 был повышен в ответ на холод, а гены CaERF053 и CaERF014 были повышены в ответ на низкую влажность и высокую температуру соответственно. Гены CcDREB1B, CcRAP2.4, CcERF027, CcDREB1D и CcTINY были повышены при стрессе от засухи у C. canephora [79]. Изучение этих генов у кофе может значительно способствовать разработке устойчивых к стрессу генотипов кофе, обеспечивая устойчивость производства кофе перед лицом изменения климата.
Хотя генетическая трансформация у кофе осуществима и широко исследовалась, разработка и коммерциализация трансгенных генотипов кофе, созданных для повышения устойчивости к засухе, жаре и засолению, все еще ограничены. Устойчивость к абиотическому стрессу — это сложный признак, часто контролируемый множеством генов. Это делает сложным идентификацию и манипуляцию специфическими генетическими факторами, способствующими улучшенной толерантности [80]. Также кофейное растение имеет сложный геном, и достижение стабильной и эффективной трансформации может быть технически сложным. Это включает проблемы, связанные с вставкой гена, экспрессией и наследованием в последующих поколениях. Наконец, большая часть исследований по генетической трансформации кофе была сосредоточена на признаках, таких как устойчивость к болезням (например, кофейная ржавчина) и качество кофе (например, содержание кофеина). Как результат, было относительно меньше внимания разработке генотипов, специфически для устойчивости к абиотическому стрессу [81]. Однако продолжаются исследования для решения этих проблем.
Достижения в геномных и биотехнологических инструментах, таких как CRISPR/Cas9, могут предложить новые возможности для разработки генотипов кофе с усиленной устойчивостью к абиотическому стрессу в ближайшем будущем. CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats) — это естественный механизм защиты, обнаруженный у бактерий. Он состоит из повторяющихся последовательностей ДНК, перемежающихся «спейсерными» последовательностями, полученными из вирусов, которые ранее атаковали бактерии. Эти спейсеры служат генетической памятью о прошлых инфекциях. Белок Cas9 — это фермент, который действует как молекулярные ножницы, способные разрезать ДНК в специфических местах. В такой технике создается синтетическая молекула РНК, которая соответствует специфической целевой последовательности ДНК, затем gRNA комбинируется с ферментом Cas9 для формирования комплекса, который может найти и разрезать целевую ДНК. Естественные механизмы репарации клетки активируются, и во время этого процесса репарации возможно внести специфические изменения, такие как вставка, удаление или замена последовательностей ДНК [82]. Однако не сообщалось о геномном редактировании для агрономически значимых признаков у кофе. Два исследования, проведенных с растениями C. canephora, использовали ген PDS (фитоен десатураза) в качестве целевого гена. Эффективность редактирования составила 30,4% [83] и 76,9% [84], с 7,6% и 54% гомозиготных мутаций соответственно, что обнадеживает для программ селекции, но все еще представляет проблему, учитывая аллополиплоидную природу кофе.
3.2. Питание растений как инструмент для повышения устойчивости кофе к абиотическому стрессу
Разработка решений для борьбы с изменениями температуры, доступности воды и интенсивности света, вызванными изменением климата, является значительной задачей для сельскохозяйственных исследователей. Абиотические стрессы влияют на поглощение, транслокацию и использование питательных веществ, что приводит к нарушению ионного гомеостаза в растительных клетках [12]. Например, снижение доступности воды может ограничить доступность питательных веществ в почве, уменьшить поглощение питательных веществ корнями и, finalmente, снизить их концентрацию в тканях растений [85]. Стресс от засухи оказывает неблагоприятное влияние на концентрации азота (N) и фосфора (P) [86], но не имеет определенного воздействия на концентрации других питательных веществ в тканях растений [87]. У кофейных растений негативный эффект засухи на концентрацию N связан со снижением Vmax (максимальная скорость поглощения), Km (константа Михаэлиса-Ментен) и Cmin (внешняя концентрация, при которой чистое поглощение ионов равно нулю), связанных с поглощением нитрата (NO₃⁻) [88]. Дисбаланс питательных веществ значительно влияет на модель роста, антиоксидантные защитные механизмы и устойчивость к биотическим и абиотическим стрессам [12]. Поэтому правильное снабжение питательными веществами необходимо для смягчения стресса у растений и увеличения производства продовольствия [12,89].
Сообщается, что многие питательные вещества растений смягчают стресс у растений, участвуя в событиях, которые варьируются от восприятия стресса до приобретения толерантности [89]. Например, N участвует в (i) сигнальных процессах через оксид азота, (ii) процессах осморегуляции и активации компонентов антиоксидантной системы и (iii) увеличении пластичности корней для поглощения воды и питательных веществ и в построении углеводов [12]. В свою очередь, P связан с процессами, такими как (i) поддержание тургора клеток и стабильности мембран, (ii) распределение углеводов и корректировка архитектуры корней, и (iii) действие как антиоксидантной, так и фотосинтетической систем [12,89]. Питательные вещества и полезные элементы играют ключевые роли в защите растений от биотических и абиотических стрессов (см. [12]), но некоторые элементы изучались больше, чем другие. Таблица 3 обобщает основные результаты применения питательных веществ и полезных элементов для смягчения стресса растений в производстве кофе. Количество исследований, оценивающих вклад питательных веществ, полезных элементов (Таблица 3) и элементов, не считающихся полезными для растений, таких как йод (I), в смягчение абиотического стресса в производстве кофе, невелико. Однако большинство исследований показали перспективные результаты в отношении повышения устойчивости растений и улучшения качества зерен. Для примера, Андраде и др. [90] сообщили, что применение I (2,5 мг дм³ KIO₃) ослабляло стресс от водного дефицита у C. arabica cv. Catuaí 99 за счет увеличения фотосинтетической эффективности, относительного содержания воды и индекса толерантности к водному дефициту, содержания фотосинтетических пигментов и совместимых осмолитов. Авторы также наблюдали, что применение I стимулировало антиоксидантную ферментативную систему, позволяя более высокую стабильность клеточных мембран. Эти результаты увлекательны, поскольку повреждение клеточных мембран в условиях водного дефицита является важным фактором, приводящим к нарушению ионного гомеостаза у растений [12], что снижает способность растения справляться со стрессом. Питание растений могло бы служить мощным инструментом для смягчения стресса [89]. Дальнейшие исследования необходимы для оценки роли минеральных элементов (включая те, которые традиционно не считаются полезными или необходимыми для растений) в снижении различных типов стресса.
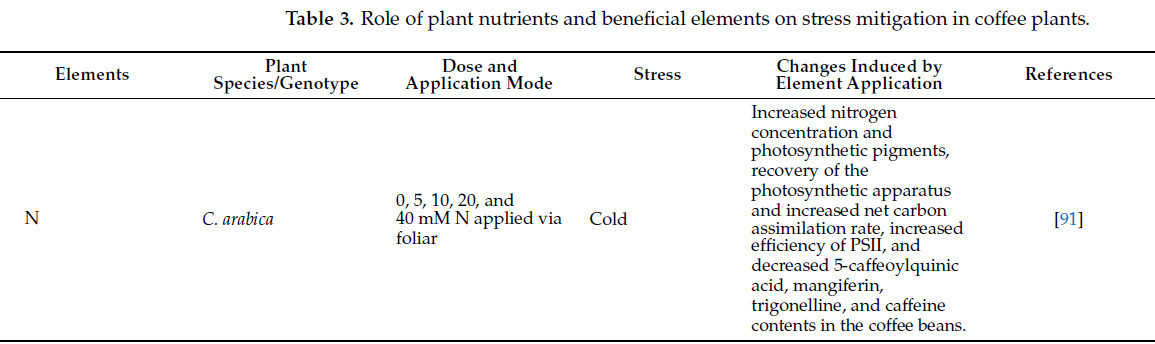
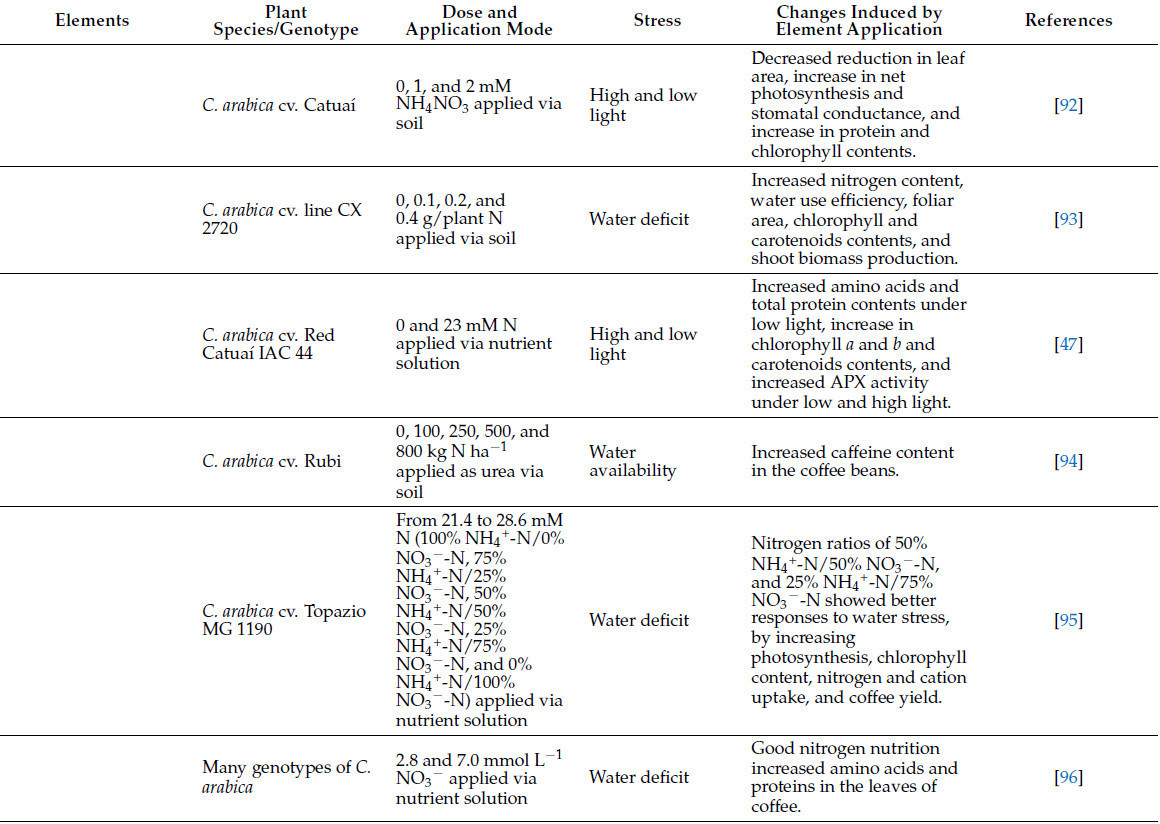
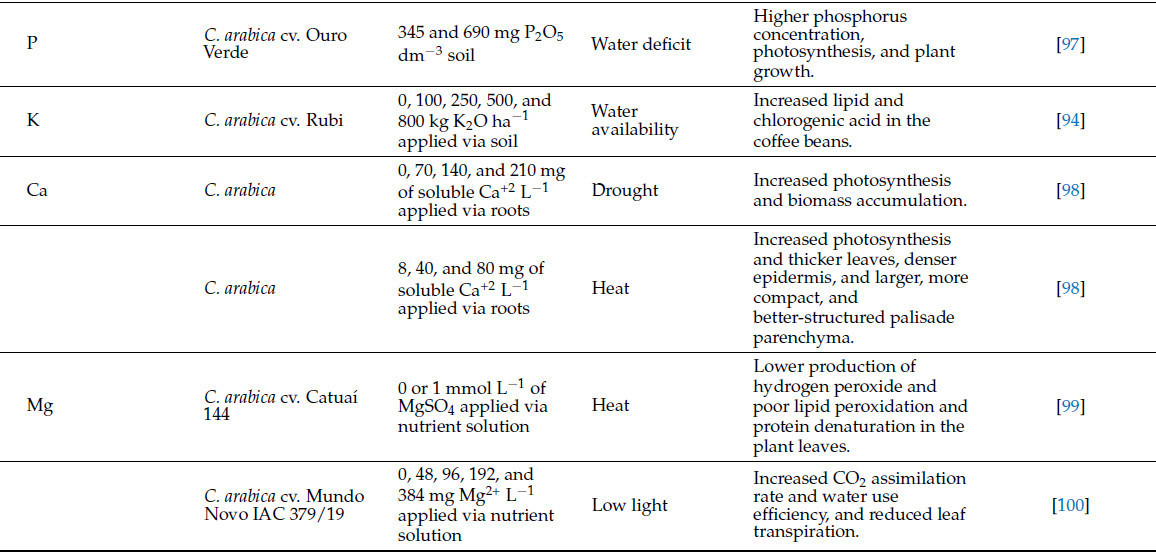
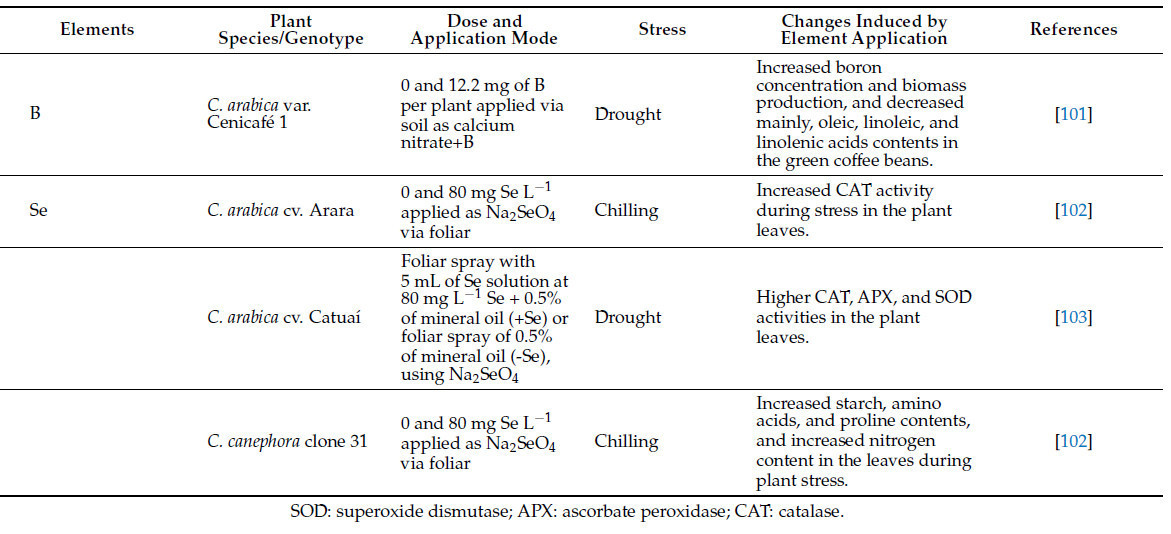
Таблица 3. Роль питательных веществ и полезных элементов растений в смягчении стресса у кофейных растений.
4. Заключительные замечания и перспективы на будущее
Количество исследований, оценивающих ущерб, вызванный засухой, жарой, избыточным освещением и засолением, в производстве кофе очень мало по сравнению с другими культурами, что ограничивает понимание ответных реакций растений на эти стрессы окружающей среды и, следовательно, способность использовать стратегии более эффективно для смягчения стресса у растений. Многие учреждения, такие как Всемирная метеорологическая организация (WMO), часто указывают, что климат меняется год от года, что имеет глубокие последствия для производства кофе. Поэтому срочно необходимо провести гораздо больше исследований, чтобы понять реакции кофе на засуху, жар, избыточное освещение и засоление, чтобы поддержать применение таких стратегий, как селекция растений и/или питание растений для смягчения стресса.
Некоторые исследования указали, что память растений крайне важна для кофейных растений, чтобы справляться с засухой, и на основе этих исследований было предложено предварительное воздействие на сеянцы кофе дефицитным орошением с перспективными результатами в отношении смягчения стресса у растений. Такие результаты связаны с более быстрой адаптацией метаболизма растения, вызванной повторяющимися эпизодами одного и того же стресса. Эту адаптацию растения можно получить из программ селекции растений и лучшего питания растений. Из-за многолетней природы кофейных растений селекция растений часто медленная. Однако программы селекции растений могли бы быть ускорены с помощью новых инструментов, таких как CRISPR, и лучшего понимания реакций кофе на стрессы окружающей среды. Более того, программы селекции растений, связанные с кофе, должны быть сосредоточены на получении растений, более толерантных к стрессам окружающей среды.
Питание растений — это другая альтернатива, которую можно использовать для поддержки адаптации метаболизма растений, но необходимо управлять минеральными питательными веществами более целостно, следуя Целям устойчивого развития. Таким образом, альтернативой использованию минеральных питательных веществ является применение биостимуляторов [89]. Однако лишь несколько исследований оценили эффект биостимуляторов на защиту кофе от засухи, жары, избыточного освещения и/или засоления. Дополнительно, питание растений как инструмент для смягчения стресса у растений должно следовать основному концепту управления питательными веществами 4R (внесение правильного источника питательных веществ растений в правильной дозе, в правильное время и в правильном месте).
References
1. USDA—United States Department of Agriculture. Brazil: Coffee Annual. 2023. Available online: https://fas.usda.gov/data/brazil-coffee-annual-8 (accessed on 16 July 2024).
2. WMO—World Meteorological Organization. State of the Global Climate 2023. WMO-No. 1347. 2024. Available online: https://library.wmo.int/idurl/4/68835 (accessed on 21 October 2024).
3. Koutouleas, A.; Sarzynski, T.; Bordeaux, M.; Bosselmann, A.S.; Campa, C.; Etienne, H.; Turreira-García, N.; Rigal, C.; Vaast, P.; Ramalho, J.C.; et al. Shaded-Coffee: A Nature-Based Strategy for Coffee Production under Climate Change? A Review. Front. Sustain. Food Syst. 2022, 6, 877476. [Google Scholar] [CrossRef]
4. DaMatta, F.M.; Avila, R.T.; Cardoso, A.A.; Martins, S.C.V.; Ramalho, J.C. Physiological and agronomic performance of the coffee crop in the context of climate change and global warming: A Review. J. Agric. Food Chem. 2018, 66, 5264–5274. [Google Scholar] [CrossRef] [PubMed]
5. Magrach, A.; Ghazoul, J. Climate and pest-driven geographic shifts in global coffee production: Implications for forest cover, biodiversity and carbon storage. PLoS ONE 2015, 10, e0133071. [Google Scholar] [CrossRef] [PubMed]
6. Rigal, C.; Xu, J.; Vaast, P. Young shade trees improve soil quality in intensively managed coffee systems recently converted to agroforestry in Yunnan Province, China. Plant Soil 2020, 453, 119–137. [Google Scholar] [CrossRef]
7. Braga, G.B.; Imbuzeiro, H.M.A.; Pires, G.F.; Oliveira, L.R.D.; Barbosa, R.A.; Vilela, K.D.F. Frost risk and rural insurance in Brazil. Rev. Bras. Meteorol. 2021, 36, 703–711. [Google Scholar] [CrossRef]
8. Jayakumar, M.; Rajavel, M.; Surendran, U.; Gopinath, G.; Ramamoorthy, K. Impact of climate variability on coffee yield in India—With a micro-level case study using long-term coffee yield data of humid tropical Kerala. Clim. Change 2017, 145, 335–349. [Google Scholar] [CrossRef]
9. Lima, R.B.; dos Santos, T.B.; Vieira, L.G.E.; Ferrarese, M.L.L.; Ferrarese-Filho, O.; Donatti, L.; Boeger, M.R.T.; Petkowicz, C.L.O. Heat stress causes alterations in the cell-wall polymers and anatomy of coffee leaves (Coffea arabica L.). Carbohydr. Polym. 2013, 93, 135–143. [Google Scholar] [CrossRef]
10. Avila, R.T.; Almeida, W.L.; Costa, L.C.; Machado, K.L.G.; Barbosa, M.L.; Souza, R.P.B.; Martino, P.B.; Juárez, M.A.T.; Marçal, D.M.S.; Martins, S.C.V.; et al. Elevated air [CO2] improves photosynthetic performance and alters biomass accumulation and partitioning in drought-stressed coffee plants. Environ. Exp. Bot. 2020, 177, 104137. [Google Scholar] [CrossRef]
11. dos Santos, T.B.; da Silva Ferreira, M.F.; Marques, I.; Oliveira, S.C.; Zaidan, I.R.; Oliveira, M.G.; Rodrigues, W.P.; Ribas, A.F.; Guyot, R.; Ramalho, J.C.; et al. Current challenges and genomic advances towards the development resilient coffee genotypes to abiotic stresses. In Genomic Designing for Abiotic Stress Resistant Technical Crops; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2022; pp. 41–69. [Google Scholar]
12. Kumari, V.V.; Banerjee, P.; Verma, V.C.; Sukumaran, S.; Chandran, M.A.S.; Gopinath, K.A.; Venkatesh, G.; Yadav, S.K.; Singh, V.K.; Awasthi, N.K. Plant nutrition: An Effective Way to alleviate abiotic stress in agricultural crops. Int. J. Mol. Sci. 2022, 23, 8519. [Google Scholar] [CrossRef]
13. Sharma, M.; Kumar, P.; Verma, V.; Sharma, R.; Bhargava, B.; Irfan, M. Understanding plant stress memory response for abiotic stress resilience: Molecular insights and prospects. Plant Physiol. Biochem. 2022, 179, 10–24. [Google Scholar] [CrossRef]
14. Menezes-Silva, P.E.; Sanglard, L.M.; Ávila, R.T.; Morais, L.E.; Martins, S.C.; Nobres, P.; Patreze, C.M.; Ferreira, M.A.; Araújo, W.L.; Fernie, A.R. Photosynthetic and metabolic acclimation to repeated drought events play key roles in drought tolerance in coffee. J. Exp. Bot. 2017, 68, 4309–4322. [Google Scholar] [CrossRef] [PubMed]
15. Kabbadj, A.; Makoudi, B.; Mouradi, M.; Pauly, N.; Frendo, P.; Ghoulam, C. Physiological and biochemical responses involved in water deficit tolerance of nitrogen-fixing Vicia faba. PLoS ONE 2017, 12, e0190284. [Google Scholar] [CrossRef] [PubMed]
16. Salehi-Lisar, S.Y.; Bakhshayeshan-Agdam, H. Drought Stress in Plants: Causes, Consequences, and Tolerance; Hossain, M., Wani, S., Bhattacharjee, S., Burritt, D., Tran, L.S., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; Volume 1, pp. 1–16. [Google Scholar]
17. Gimenez, C.; Gallardo, M.; Thompson, R.B. Plant water relations. In Encyclopedia of Soils in the Environment; Hillel, D., Ed.; Elsevier: Oxford, UK, 2005; pp. 231–238. [Google Scholar]
18. Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef]
19. Jaleel, C.A.; Manivannan, P.; Wahid, A.; Farooq, M.; Somasundaram, R.; Panneerselvam, R. Drought stress in plants: A review on morphological characteristics and pigments composition. Int. J. Agric. Biol. 2009, 11, 100–105. [Google Scholar]
20. Bhargava, S.; Sawant, K. Drought stress adaptation: Metabolic adjustment and regulation of gene expression. Plant Breed. 2013, 132, 21–32. [Google Scholar] [CrossRef]
21. DaMatta, F.M.; Maestri, M.; Barros, R.S.; Regazzi, A.J. Water relations of coffee leaves (Coffea arabica and C. canephora) in response to drought. J. Hortic. Sci.-India 1993, 68, 741–746. [Google Scholar] [CrossRef]
22. Lima, A.A.; Santos, I.S.; Torres, M.E.L.; Cardon, C.H.; Caldeira, C.F.; Lima, R.R.; Davies, W.J.; Dodd, I.C.; Chalfun-Junior, A. Drought and re-watering modify ethylene production and sensitivity, and are associated with coffee anthesis. Environ. Exp. Bot. 2021, 181, 104289. [Google Scholar] [CrossRef]
23. Cavatte, P.C.; Oliveira, A.A.G.; Morais, L.E.; Martins, S.C.V.; Sanglard, L.M.V.P.; DaMatta, F.M. Could shading reduce the negative impacts of drought on coffee? A morphophysiological analysis. Physiol. Plant. 2012, 144, 111–122. [Google Scholar] [CrossRef]
24. DaMatta, F.M.; Maestri, M.; Barros, R.S. Phothosynthetic performance of two coffee species under drought. Photosynthetica 1997, 34, 257–264. [Google Scholar] [CrossRef]
25. de Andrade, L.I.F.; Linhares, P.C.A.; da Fonseca, T.M.; Silva, A.A.; Santos, J.P.; Pereira, M.P.; Silva, V.A.; Marchiori, P.E.R. Photosynthetic efficiency and root plasticity promote drought tolerance in coffee genotypes. Acta Physiol. Plant. 2022, 44, 109. [Google Scholar] [CrossRef]
26. Chekol, H.; Warkineh, B.; Shimber, T.; Mierek-Adamska, A.; Dąbrowska, G.B.; Degu, A. Drought Stress Responses in Arabica Coffee Genotypes: Physiological and Metabolic Insights. Plants 2024, 13, 828. [Google Scholar] [CrossRef] [PubMed]
27. Chekol, H.; Bezuayehu, Y.; Warkineh, B.; Shimber, T.; Mierek-Adamska, A.; Dąbrowska, G.B.; Degu, A. Unraveling Drought Tolerance and Sensitivity in Coffee Genotypes: Insights fromSeed Traits, Germination, and Growth-Physiological Responses. Agriculture 2023, 13, 1754. [Google Scholar] [CrossRef]
28. Aman, M.; Worku, M.; Shimbir, T.; Astatkie, T. Root traits and biomass production of drought-resistant and drought-sensitive arabica coffee varieties growing under contrasting watering regimes. Agrosyst. Geosci. Environ. 2024, 7, e20488. [Google Scholar] [CrossRef]
29. Praxedes, S.C.; DaMatta, F.M.; Loureiro, M.E.; Ferrão, M.A.G.; Cordeiro, A.T. Effects of long-term soil drought on photosynthesis and carbohydrate metabolism in mature robusta coffee (Coffea canephora Pierre var. kouillou) leaves. Environ. Exp. Bot. 2006, 56, 263–273. [Google Scholar] [CrossRef]
30. Hassan, M.U.; Chattha, M.U.; Khan, I.; Chattha, M.B.; Barbanti, L.; Aamer, M.; Iqbal, M.M.; Nawaz, M.; Mahmood, A.; Ali, A.; et al. Heat stress in cultivated plants: Nature, impact, mechanisms, and mitigation strategies—A review. Plant Biosyst. 2021, 155, 211–234. [Google Scholar] [CrossRef]
31. Carvalho, M.H.C. Drought stress and reactive oxygen species. Plant Signal. Behav. 2008, 3, 156–165. [Google Scholar] [CrossRef]
32. de Guedes, F.A.F.; Nobres, P.; Ferreira, D.C.R.; Menezes-Silva, P.E.; Ribeiro-Alves, M.; Correa, R.L.; DaMatta, F.M.; Alves-Ferreira, M. Transcriptional memory contributes to drought tolerance in coffee (Coffea canephora) plants. Environ. Exp. Bot. 2018, 147, 220–233. [Google Scholar] [CrossRef]
33. Woodward, F.I. Stomatal numbers are sensitive to increases in CO2 from preindustrial levels. Nature 1987, 327, 617–618. [Google Scholar] [CrossRef]
34. Ainsworth, E.A.; Rogers, A. The response of photosynthesis and stomatal conductance to rising [CO2]: Mechanisms and environmental interactions: Photosynthesis and stomatal conductance responses to rising [CO2]. Plant Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef]
35. Rakocevic, M.; Ribeiro, R.V.; Marchiori, P.E.R.; Filizola, H.F.; Batista, E.R. Structural and functional changes in coffee trees after 4 years under free air CO2 enrichment. Ann. Bot. 2018, 121, 1065–1078. [Google Scholar] [CrossRef]
36. Leakey, A.D.B.; Ainsworth, E.A.; Bernacchi, C.J.; Rogers, A.; Long, S.P.; Ort, D.R. Elevated CO2 effects on plant carbon, nitrogen, and water relations: Six important lessons from FACE. J. Exp. Bot. 2009, 60, 2859–2876. [Google Scholar] [CrossRef] [PubMed]
37. Sicher, R.C.; Timlin, D.; Bailey, B. Responses of growth and primary metabolism of water-stressed barley roots to rehydration. J. Plant Physiol. 2012, 169, 686–695. [Google Scholar] [CrossRef]
38. Li, N.; Euring, D.; Cha, J.Y.; Lin, Z.; Lu, M.; Huang, L.J.; Kim, W.Y. Plant hormone-mediated regulation of heat tolerance in response to global climate change. Front. Plant Sci. 2021, 11, 2318. [Google Scholar] [CrossRef]
39. Gerganova, M.; Popova, A.V.; Stanoeva, D.; Velitchkova, M. Tomato plants acclimate better to elevated temperature and high light than to treatment with each factor separately. Plant Physiol. Biochem. 2016, 104, 234–241. [Google Scholar] [CrossRef]
40. Martins, S.C.V.; Araújo, W.L.; Tohge, T.; Fernie, A.R.; DaMatta, F.M. In high-light-acclimated coffee plants the metabolic machinery is adjusted to avoid oxidative stress rather than to benefit from extra light enhancement in photosynthetic yield. PLoS ONE 2014, 9, e94862. [Google Scholar] [CrossRef]
41. Roda, N.d.M.; Branchi, B.A.; Longo, R.M.; Pontin, J.; Abreu, D.P.d.; Santos, P.R.d.; Campostrini, E. The Advantages of Using Kaolin-Based Particle Films to Improve Coffee Production in the Minas Gerais Cerrado Biome. Sustainability 2022, 14, 4485. [Google Scholar] [CrossRef]
42. de Oliveira, R.R.; Ribeiro, T.H.C.; Cardon, C.H.; Fedenia, L.; Maia, V.A.; Barbosa, B.C.F.; Caldeira, C.F.; Klein, P.E.; Chalfun-Junior, A. Elevated Temperatures Impose Transcriptional Constraints and Elicit Intraspecific Differences Between Coffee Genotypes. Front. Plant Sci. 2020, 11, 1113. [Google Scholar] [CrossRef]
43. Rodrigues, W.P.; Silva, J.R.; Ferreira, L.S.; Machado Filho, J.A.; Figueiredo, F.A.M.M.A.; Ferraz, T.M.; Bernado, W.P.; Bezerra, L.B.S.; Abreu, D.P.; Cespom, L.; et al. Stomatal and photochemical limitations of photosynthesis in coffee (Coffea spp.) plants subjected to elevated temperatures. Crop Pasture Sci. 2018, 69, 317–325. [Google Scholar] [CrossRef]
44. Vilas-Boas, T.; Duarte, A.A.; Della Torre, F.; Lovato, M.B.; Lemos-Filho, J.P. Does acclimation in distinct light conditions determine differences in the photosynthetic heat tolerance of coffee plants? Plant Biol. 2023, 25, 1101–1108. [Google Scholar] [CrossRef]
45. Thioune, E.-H.; McCarthy, J.; Gallagher, T.; Osborne, B. A humidity shock leads to rapid, temperature dependent changes in coffee leaf physiology and gene expression. Tree Physiol. 2017, 37, 367–379. [Google Scholar]
46. Liu, X.G.; Wan, M.D.; Wu, H.; Yang, Q.L. Photosynthetic response and use of water and light of Arabica coffee leaf under different irrigation and light levels. Oxid. Commun. 2016, 39, 873–883. [Google Scholar]
47. Pompelli, M.F.; Martins, S.C.V.; Antunes, W.C.; Chaves, A.R.M.; DaMatta, F.M. Photosynthesis and photoprotection in coffee leaves is affected by nitrogen and light availabilities in winter conditions. J. Plant Physiol. 2010, 167, 1052–1060. [Google Scholar] [CrossRef] [PubMed]
48. Rodríguez-López, N.F.; Cavatte, P.C.; Silva, P.E.M.; Martins, S.C.V.; Morais, L.E.; Medina, E.F.; DaMatta, F.M. Physiological and biochemical abilities of robusta coffee leaves for acclimation to cope with temporal changes in light availability. Physiol. Plant. 2013, 149, 4555. [Google Scholar] [CrossRef]
49. FAO—Food and Agriculture Organization of the United Nations. Soil Letters Salt-Affected Soils Are a Global Issue. Intergovernmental Technical Panel on Soils. 2021. Available online: https://openknowledge.fao.org/server/api/core/bitstreams/8b5c687f-5a9b-4034-a4e2-0457066c1ae3/content (accessed on 16 July 2024).
50. Majeed, A.; Muhammad, Z. Salinity: A major agricultural problem—Causes, impacts on crop productivity and management strategies. In Plant Abiotic Stress Tolerance; Hasanuzzaman, M., Hakeem, K., Nahar, K., Alharby, H., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 83–99. [Google Scholar]
51. Zaman, M.; Shahid, S.A.; Heng, L. Guideline for Salinity Assessment, Mitigation and Adaptation Using Nuclear and Related Techniques; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
52. Chhabra, R. Classification of salt-affected soils. Arid Land Res. Manag. 2004, 19, 61–79. [Google Scholar] [CrossRef]
53. Yadav, S.; Irfan, M.; Ahmad, A.; Hayat, S. Causes of salinity and plant manifestations to salt stress: A review. J. Environ. Biol. 2011, 32, 667–685. [Google Scholar]
54. Hnilickova, H.; Hnilicka, F.; Martinkova, J.; Kraus, K. Effects of Salt Stress on Water Status; Photosynthesis and Chlorophyll Fluorescence of Rocket. Plant Soil Environ. 2017, 63, 362–367. [Google Scholar] [CrossRef]
55. Mbarki, S.; Sytar, O.; Cerda, A.; Zivcak, M.; Rastogi, A.; He, X.; Zoghlami, A.; Abdelly, C.; Brestic, M. Strategies to mitigate the salt stress effects on photosynthetic apparatus and productivity of crop plants. In Salinity Responses and Tolerance in Plants; Kumar, V., Wani, S., Suprasanna, P., Tran, L.S., Eds.; Springer: Berlin/Heidelberg, Germany, 2018; Volume 1, pp. 85–136. [Google Scholar]
56. Lima, R.B.; dos Santos, T.B.; Vieira, L.G.E.; Ferrarese, M.D.L.L.; Ferrarese-Filho, O.; Donatti, L.; Petkowicz, C.L.O. Salt stress alters the cell wall polysaccharides and anatomy of coffee (Coffea arabica L.) leaf cells. Carbohydr. Polym. 2014, 112, 686–694. [Google Scholar] [CrossRef]
57. Oyebamiji, Y.O.; Adigun, B.A.; Shamsudin, N.A.A.; Ikmal, A.M.; Salisu, M.A.; Malike, F.A.; Lateef, A.A. Recent Advancements in Mitigating Abiotic Stresses in Crops. Horticulturae 2024, 10, 156. [Google Scholar] [CrossRef]
58. Lehmann, J.; Amonette, J.E.; Roberts, K. Role of biochar in mitigation of climate change. In Handbook of Climate Change and Agroecosystems; Hillel, D., Rosenzweig, C., Eds.; Imperial College Press: London, UK, 2010; Volume 1, pp. 343–363. [Google Scholar]
59. Jeffery, S.; Verheijen, F.G.; Kammann, C.; Abalos, D. Biochar effects on methane emissions from soils: A meta-analysis. Soil Biol. Biochem. 2016, 101, 251–258. [Google Scholar] [CrossRef]
60. Amoakwah, E.; Frimpong, K.A.; Okae-Anti, D.; Arthur, E. Soil water retention, air flow and pore structure characteristics after corn cob biochar application to a tropical sandy loam. Geoderma 2017, 307, 189–197. [Google Scholar] [CrossRef]
61. Sorrenti, G.; Masiello, C.A.; Dugan, B.; Toselli, M. Biochar physico-chemical properties as affected by environmental exposure. Sci. Total Environ. 2016, 563–564, 237–246. [Google Scholar] [CrossRef] [PubMed]
62. Zulfiqar, F.; Wei, X.; Shaukat, N.; Chen, J.; Raza, A.; Younis, A.; Nafees, M.; Abideen, Z.; Zaid, A.; Latif, N.; et al. Effects of biochar and biochar-compost mix on growth, performance and physiological responses of potted Alpinia zerumbet. Sustainability 2021, 13, 11226. [Google Scholar] [CrossRef]
63. Ahmad, M.; Lee, S.S.; Lee, S.E.; Al-Wabel, M.I.; Tsang, D.C.W.; Ok, Y.S. Biochar-induced changes in soil properties affected immobilization/mobilization of metals/metalloids in contaminated soils. J. Soils Sediments 2017, 17, 717–730. [Google Scholar] [CrossRef]
64. Reyes-Herrera, D.F.; Sánchez-Reinoso, A.D.; Lombardini, L.; Restrepo-Díaz, H. Physiological Responses of Coffee (Coffea arabica L.) Plants to Biochar Application under Water Deficit Conditions. Not. Bot. Horti Agrobot. Cluj-Napoca 2023, 51, 12873. [Google Scholar] [CrossRef]
65. Wan, J.; Zhang, P.; Wang, R.; Sun, L.; Ju, Q.; Xu, J. Comparative physiological responses and transcriptome analysis reveal the roles of melatonin and serotonin in regulating growth and metabolism in Arabidopsis. BMC Plant Biol. 2018, 18, 362. [Google Scholar] [CrossRef]
66. Zhao, C.; Nawaz, G.; Cao, Q.; Xu, T. Melatonin is a potential target for improving horticultural crop resistance to abiotic stress. Sci. Hortic. 2022, 291, 110560. [Google Scholar] [CrossRef]
67. Ayyaz, A.; Shahzadi, A.K.; Fatima, S.; Yasin, G.; Zafar, Z.U.; Athar, H.U.R.; Farooq, M.A. Uncovering the role of melatonin in plant stress tolerance. Theor. Exp. Plant Physiol. 2022, 34, 335–346. [Google Scholar] [CrossRef]
68. Weeda, S.; Zhang, N.; Zhao, X.; Ndip, G.; Guo, Y.; Buck, G.A.; Fu, C.; Ren, S. Arabidopsis transcriptome analysis reveals key roles of melatonin in plant defense systems. PLoS ONE 2014, 9, e93462. [Google Scholar] [CrossRef]
69. Wei, Y.; Zeng, H.; Hu, W.; Chen, L.; He, C.; Shi, H. Comparative transcriptional profiling of melatonin synthesis and catabolic genes indicates the possible role of melatonin in developmental and stress responses in rice. Front. Plant Sci. 2016, 7, 676. [Google Scholar] [CrossRef]
70. Ye, J.; Wang, S.; Deng, X.; Yin, L.; Xiong, B.; Wang, X. Melatonin increased maize (Zea mays L.) seedling drought tolerance by alleviating drought-induced photosynthetic inhibition and oxidative damage. Acta Physiol. Plant. 2016, 38, 48. [Google Scholar]
71. Campos, C.N.; Ávila, R.G.; Souza, K.R.D.; Azevedo, L.M.; Alves, J.D. Melatonin reduces oxidative stress and promotes drought tolerance in young Coffea arabica L. plants. Agric. Water Manag. 2019, 211, 37–47. [Google Scholar] [CrossRef]
72. Sseremba, G.; Tongoona, P.B.; Musoli, P.; Eleblu, J.S.Y.; Melomey, L.D.; Bitalo, D.N.; Atwijukire, E.; Mulindwa, J.; Aryatwijuka, N.; Muhumuza, E.; et al. Viability of Deficit Irrigation Pre-Exposure in Adapting Robusta Coffee to Drought Stress. Agronomy 2023, 13, 674. [Google Scholar] [CrossRef]
73. Silva, V.A.; Antunes, W.C.; Guimarães, B.L.S.; Paiva, R.M.C.; Silva, V.F.; Ferrão, M.A.G.; DaMatta, F.M.; Loureiro, M.E. Physiological response of Conilon coffee clone sensitive to drought grafted onto tolerant rootstock. Pesqui. Agropecu. Bras. 2010, 45, 457–464. [Google Scholar]
74. Teixeira, A.L.; Souza, F.D.F.; Pereira, A.A.; Oliveira, A.C.B.D.; Rocha, R.B. Selection of arabica coffee progenies tolerant to heat stress. Cienc. Rural 2015, 45, 1228–1234. [Google Scholar] [CrossRef]
75. Naik, B.J.; Kim, S.C.; Seenaiah, R.; Basha, P.A.; Song, E.Y. Coffee cultivation techniques, impact of climate change on coffee production, role of nanoparticles and molecular markers in coffee crop improvement, and challenges. J. Plant Biotechnol. 2021, 48, 207–222. [Google Scholar] [CrossRef]
76. Mofatto, L.S.; Carneiro, F.D.A.; Vieira, N.G.; Duarte, K.E.; Vidal, R.O.; Alekcevetch, J.C.; Cotta, M.G.; Verdeil, J.-L.; Fabienne, L.-M.; Lartaud, M.; et al. Identification of candidate genes for drought tolerance in coffee by high-throughput sequencing in the shoot apex of different Coffea arabica cultivars. BMC Plant Biol. 2016, 16, 94. [Google Scholar] [CrossRef]
77. De Carvalho, K.; Petkowicz, C.L.O.; Nagashima, G.T.; Bespalhok Filho, J.C.; Vieira, L.G.E.; Pereira, L.F.P.; Domingues, D.S. Homeologous genes involved in mannitol synthesis reveal unequal contributions in response to abiotic stress in Coffea arabica. Mol. Genet. Genom. 2014, 289, 951–963. [Google Scholar] [CrossRef]
78. Atkinson, N.J.; Urwin, P.E. The interaction of plant biotic and abiotic stresses: From genes to the field. J. Exp. Bot. 2012, 63, 3523–3544. [Google Scholar] [CrossRef]
79. Torres, L.F.; Reichel, T.; Déchamp, E.; de Aquino, S.O.; Duarte, K.E.; Alves, G.S.C.; Silva, A.T.; Cotta, M.G.; Costa, T.S.; Diniz, L.E.C.; et al. Expression of DREB-like genes in Coffea canephora and C. arabica subjected to various types of abiotic stress. Trop. Plant Biol. 2019, 12, 98–116. [Google Scholar] [CrossRef]
80. Duque, A.S.; Almeida, A.M.; Bernardes da Silva, A.; Marques da Silva, J.; Farinha, A.P.; Santos, D.; Fevereiro, P.; Araújo, S.S. Abiotic stress responses in plants: Unraveling the complexity of genes and networks to survive. In Abiotic Stress: Plant Responses and Applications in Agriculture; Vahdati, K., Leslie, C., Eds.; INTECH Open: Rijeka, Croatia; London, UK, 2013; pp. 49–102. [Google Scholar]
81. Mishra, M.K.; Slater, A. Recent Advances in the Genetic Transformation of Coffee. Biotechnol. Res. Int. 2012, 2012, 580857. [Google Scholar] [CrossRef]
82. Vats, S.; Kumawat, S.; Kumar, V.; Patil, G.B.; Joshi, T.; Sonah, H.; Sharma, T.R.; Deshmukh, R. Genome editing in plants: Exploration of technological advancements and challenges. Cell 2019, 8, 1386. [Google Scholar] [CrossRef] [PubMed]
83. Breitler, J.-C.; Dechamp, E.; Campa, C.; Rodrigues, L.A.Z.; Guyot, R.; Marraccini, P.; Etienne, H. CRISPR/Cas9-mediated efficient targeted mutagenesis has the potential to accelerate the domestication of Coffea canephora. Plant Cell Tissue Organ Cult. 2018, 134, 383–394. [Google Scholar] [CrossRef]
84. Casarin, T.; Freitas, N.C.; Pinto, R.T.; Breitler, J.-C.; Rodrigues, L.A.Z.; Marraccini, P.; Etienne, H.; Diniz, L.E.C.; Andrade, A.C.; Paiva, L.V. Multiplex CRISPR/Cas9-mediated knockout of the phytoene desaturase gene in Coffea canephora. Sci. Rep. 2022, 12, 17270. [Google Scholar] [CrossRef]
85. Waraich, E.A.; Ahmad, R.; Ashraf, M.Y.; Saifullah Ahmad, M. Improving agricultural water use efficiency by nutrient management in crop plants. Acta Agric. Scand. B Soil. Plant Sci. 2011, 61, 291–304. [Google Scholar] [CrossRef]
86. He, M.; Dijkstra, F.A. Drought effect on plant nitrogen and phosphorus: A meta-analysis. New Phytol. 2014, 204, 924–931. [Google Scholar] [CrossRef]
87. Silva, E.C.; Nogueira, R.J.M.C.; Silva, M.A.; Albuquerque, M.B. Drought Stress and Plant Nutrition. Plant Stress 2011, 5, 32–41. [Google Scholar]
88. Martinez, H.E.P.; Souza, B.P.; Caixeta, E.T.; Carvalho, F.P.; Clemente, J.M. Water deficit changes nitrate uptake and expression of some nitrogen related genes in coffee-plants (Coffea arabica L.). Sci. Hortic. 2020, 267, 109254. [Google Scholar] [CrossRef]
89. Rabêlo, F.H.S. Role of Plant Nutrition in Mitigating Biotic and Abiotic Stresses in Plants. Inform. Agron. 2024, 2, 5–22. (In Portuguese) [Google Scholar]
90. Andrade, O.V.S.; Lima, J.S.; Neves, T.T.; Benevenute, P.A.N.; Santos, L.C.; Nascimento, V.L.; Guilherme, L.R.G.; Marchiori, P.E.R. The role of potassium iodate in mitigating the damages of water deficit in coffee plants. J. Soil Sci. Plant Nutr. 2024, 24, 5772–5788. [Google Scholar] [CrossRef]
91. Acidri, R.; Sawai, Y.; Sugimoto, Y.; Sasagawa, D.; Masunaga, T.; Yamamoto, S.; Nishihara, E. Foliar nitrogen supply enhances the recovery of photosynthetic performance of cold-stressed coffee (Coffea arabica L.) seedlings. Photosynthetica 2020, 58, 951–960. [Google Scholar] [CrossRef]
92. Nunes, M.A.; Ramalho, J.D.C.; Dias, M.A. Effect of Nitrogen Supply on the Photosynthetic Performance of Leaves from Coffee Plants Exposed to Bright Light. J. Exp. Bot. 1993, 44, 893–899. [Google Scholar] [CrossRef]
93. Salamanca-Jimenez, A.; Doane, T.A.; Horwath, W.R. Performance of Coffee Seedlings as Affected by Soil Moisture and Nitrogen Application. Adv. Agron. 2016, 136, 221–244. [Google Scholar]
94. Vinecky, F.; Davrieux, F.; Mera, A.C.; Alves, G.S.C.; Lavagnini, G.; Leroy, T.; Bonnot, F.; Rocha, O.C.; Bartholo, G.F.; Guerra, A.F.; et al. Controlled irrigation and nitrogen, phosphorous and potassium fertilization affect the biochemical composition and quality of Arabica coffee beans. J. Agric. Sci. 2017, 155, 902–918. [Google Scholar] [CrossRef]
95. Ramirez-Builes, V.H.; Kusters, J.; Thiele, E.; Lopez-Ruiz, J.C. Physiological and Agronomical Response of Coffee to Different Nitrogen Forms with and without Water Stress. Plants 2024, 13, 1387. [Google Scholar] [CrossRef]
96. Rocha, B.C.P.; Martinez, H.E.P.; Ribeiro, C.; Brito, D.S. Nitrogen Metabolism in Coffee Plants Subjected to Water Deficit and Nitrate Doses. Braz. Arch. Biol. Technol. 2023, 66, e23210060. [Google Scholar]
97. Rakocevic, M.; Marchiori, P.E.R.; Zambrosi, F.C.B.; Machado, E.C.; Maia, A.H.N.; Ribeiro, R.V. High phosphorus supply enhances leaf gas exchange and growth of young Arabica coffee plants under water deficit. Exp. Agric. 2022, 58, e30. [Google Scholar] [CrossRef]
98. Ramírez-Builes, V.H.; Küsters, J.; de Souza, T.R.; Simmes, C. Calcium Nutrition in Coffee and Its Influence on Growth, Stress Tolerance, Cations Uptake, and Productivity. Front. Agron. 2020, 2, 590892. [Google Scholar] [CrossRef]
99. Silva, D.M.; Souza, K.R.D.; Boas, L.V.V.; Alves, Y.S.; Alvez, J.D. The effect of magnesium nutrition on the antioxidant response of coffee seedlings under heat stress. Sci. Hortic. 2017, 224, 115–125. [Google Scholar] [CrossRef]
100. Dias, K.G.L.; Guimarães, P.T.G.; Furtini Neto, A.E.; Silveira, H.R.O.; Lacerda, J.J.J. Effect of Magnesium on Gas Exchange and Photosynthetic Efficiency of Coffee Plants Grown under Different Light Levels. Agriculture 2017, 7, 85. [Google Scholar] [CrossRef]
101. Ramirez-Builes, V.H.; Küsters, J.; Thiele, E.; Leal-Varon, L.A. Boron Nutrition in Coffee Improves Drought Stress Resistance and, Together with Calcium, Improves Long-Term Productivity and Seed Composition. Agronomy 2024, 14, 474. [Google Scholar] [CrossRef]
102. de Sousa, G.F.; Silva, M.A.; de Morais, E.G.; Van Opbergen, G.A.Z.; Van Opbergen, G.G.A.Z.; de Oliveira, R.R.; Amaral, D.; Brown, P.; Chalfun-Junior, A.; Guilherme, L.R.G. Selenium enhances chilling stress tolerance in coffee species by modulating nutrient, carbohydrates, and amino acids content. Front. Plant Sci. 2022, 13, 1000430. [Google Scholar] [CrossRef] [PubMed]
103. Sousa, G.F.d.; Silva, M.A.; Carvalho, M.R.d.; Morais, E.G.d.; Benevenute, P.A.N.; Van Opbergen, G.A.Z.; Van Opbergen, G.G.A.Z.; Guilherme, L.R.G. Foliar Selenium Application to Reduce the Induced-Drought Stress Effects in Coffee Seedlings: Induced Priming or Alleviation Effect? Plants 2023, 12, 3026. [Google Scholar] [CrossRef] [PubMed]

Borgo L, Rabêlo FHS, Marchiori PER, Guilherme LRG, Guerra-Guimarães L, Resende MLVd. Impact of Drought, Heat, Excess Light, and Salinity on Coffee Production: Strategies for Mitigating Stress Through Plant Breeding and Nutrition. Agriculture. 2025; 15(1):9. https://doi.org/10.3390/agriculture15010009
Перевод статьи «Impact of Drought, Heat, Excess Light, and Salinity on Coffee Production: Strategies for Mitigating Stress Through Plant Breeding and Nutrition» авторов Borgo L, Rabêlo FHS, Marchiori PER, Guilherme LRG, Guerra-Guimarães L, Resende MLVd., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык

















































































































Комментарии (0)