Как лимонная кислота влияет на снижение заболеваемости мягкой гнилью батата и на улучшение его качества после сбора урожая
Лимонная кислота (ЛА) является распространенной, без запаха и нетоксичной пищевой добавкой. Мягкая гниль, вызываемая патогеном Rhizopus stolonifer, является основным заболеванием батата (Ipomoea batatas (L.) Lam), проявляющимся после сбора урожая. Основная цель данного исследования — определить механизм ингибирующего действия ЛА на Rhizopus stolonifer, возбудителя мягкой гнили батата. Для установления практической применимости ЛА были использованы как in vitro, так и in vivo методики.
Аннотация
Целью экспериментов in vitro в данном исследовании было выяснение воздействия 0,5% (в/в) раствора ЛА на ингибирование роста Rhizopus stolonifer, включая морфологию мицелия и расширение колонии. Эксперименты in vivo проводились на сортах батата «Синьсян» с применением 0,5% (в/в) раствора ЛА в качестве предварительной обработки.
В частности, ткани, обработанные 0,5% ЛА, сохраняли лучшее качество внешнего вида и текстурные характеристики; активность пероксидазы, β-1,3-глюканазы, хитиназы и фенилаланинаммиак-лиазы была повышена. И наоборот, та же обработка привела к снижению активности полифенолоксидазы, каталазы, аскорбатпероксидазы, целлюлазы и полигалактозидазы. Более того, было обнаружено, что обработка ЛА поддерживает повышенный уровень общих фенолов и флавоноидов в тканях батата.
В итоге, исследование демонстрирует, что 0,5% ЛА усиливает устойчивость батата к мягкой гнили путем активации связанных с защитой ферментов, подавления активности разрушающих клеточную стенку ферментов и стимулирования накопления антимикробных соединений. Эти результаты свидетельствуют в пользу использования ЛА в качестве послеуборочной обработки для снижения заболеваемости батата мягкой гнилью.
1. Введение
Батат является важной продовольственной и товарной культурой, корни которого богаты витаминами, крахмалом, растворимыми сахарами, белками и полифенолами [1,2]. Однако неправильное обращение может вызвать механические повреждения батата во время сбора урожая, транспортировки и хранения, тем самым увеличивая риск грибковых инфекций. Мягкая гниль, вызываемая патогеном Rhizopus stolonifer, представляет собой основное послеуборочное заболевание, поражающее батат (Ipomoea batatas (L.) Lam), что приводит к значительным потерям урожая и качества [3]. При заражении мягкой гнилью патоген активирует ферменты, разрушающие клеточную стенку, что приводит к деградации клеточной стенки и последующему размягчению и гниению клубней. Дополнительно, заражение Rhizopus stolonifer может привести к потере некоторых питательных веществ из батата. Хотя химические средства борьбы эффективны в предотвращении и сдерживании болезней, они несут экологические риски и вызывают обеспокоенность по поводу остатков химических веществ [4]. Следовательно, существует насущная потребность в выявлении натуральных, нетоксичных и не оставляющих остатков методов обработки для предотвращения и контроля мягкой гнили во время хранения и сохранения питательного качества батата.
Лимонная кислота (CAC) является природной органической кислотой растений, известной своими антимикробными и антиоксидантными свойствами [5]. В качестве безопасной пищевой добавки было показано, что CAC задерживает созревание, потемнение и развитие болезней у различных фруктов и овощей, тем самым продлевая их срок годности. Например, было показано, что замачивание плодов груши в растворе CAC (2%, в/в) в течение 15 минут эффективно снижает повреждение пенициллом у груш 'Yali' и сохраняет их качество [6]. Dai и др. [7] обнаружили, что погружение в раствор CAC может эффективно ингибировать потемнение каллуса, причем раствор CAC 2 × 10−3 г L−1 показал превосходный ингибирующий эффект. Скорость потемнения каллуса многолетнего райграса составила 15% после двух недель субкультивирования. Tapia-Rodriguez и др. [8] обнаружили, что комбинация CAC и CaCl2 может продлить срок годности, сохранить антиоксидантную способность и улучшить сенсорные характеристики свежего нарезанного киви.
Предварительные экспериментальные результаты показали, что 0,5% CAC, 1% CAC и 2% CAC могут снизить потери, вызванные заражением Rhizopus stolonifer, но батат, обработанный 0,5% CAC, сохранял лучшее качество. Исходя из концепции экономической и экологической целесообразности в производственной практике, 0,5% CAC была выбрана в качестве оптимальной концентрации для теста на устойчивость батата к мягкой гнили, индуцированной CAC. Изучение методов обработки CAC на заболеваемость мягкой гнилью и качество хранения батата имеет существенное практическое значение. В данном исследовании изучалось влияние обработки CAC (0,5%, в/в) на заболеваемость мягкой гнилью и качество хранения батата. В этом исследовании мы впервые в литературе сообщаем о различной восприимчивости разных участков клубня батата к мягкой гнили. Мы также исследуем механизмы, лежащие в основе ингибирующего действия CAC (0,5% в/в) на Rhizopus stolonifer и его влияния на повышение устойчивости батата к болезням. Ожидается, что результаты будут способствовать разработке эффективных стратегий для предотвращения и контроля мягкой гнили во время хранения батата.
2. Материалы и методы
2.1. Экспериментальные материалы и обработки
Батат (сорт 'Синьсян') был собран вручную через 120 дней после посадки в сельскохозяйственной полевой зоне Университета Чжэцзян A & F в июне 2021 года. Rhizopus stolonifer был предоставлен Сюйчжоуским институтом сельскохозяйственных наук, провинция Цзянсу, Китай. Лимонную кислоту (CAC) приобрели у Shanghai Yuanye Bio-Technology Co., Ltd. Для эксперимента были тщательно отобраны клубни батата одинакового размера, свободные от механических повреждений, болезней, вредителей и прорастания. Сначала клубни мыли деионизированной водой, затем дезинфицировали в 1% растворе гипохлорита натрия в течение 10 минут, после чего тщательно промывали деионизированной водой для удаления остаточного хлора. Во-вторых, образцы высушивали стерильными бумажными полотенцами и хранили в стерильных условиях. Клубни батата случайным образом разделили на две группы, каждая состояла из трех повторностей. Экспериментальную группу погружали в 0,5% раствор CAC на 5 минут, затем высушивали на воздухе, в то время как контрольную группу обрабатывали стерильной деионизированной водой. Впоследствии на обоих концах и в середине каждого клубня батата наносили проколы диаметром 5 мм с помощью стерильного пробойника, и каждую рану инокулировали суспензией спор Rhizopus stolonifer с концентрацией 1 × 10⁶ КОЕ/мл.
Свежие образцы корней собирали из здоровой ткани, расположенной в пределах 2 см от областей, прилегающих к пораженной мякоти [9]. Эти образцы мелко нарезали с помощью стерильного дисера, быстро замораживали в жидком азоте и хранили при -80 °C для последующего анализа.
Порошок корней готовили следующим образом: Корни батата нарезали на тонкие ломтики толщиной 5 мм с помощью слайсера (HM 355S, Thermo Fisher Instrument Co., Ltd., Шанхай, Китай), замораживали в жидком азоте и лиофилизировали при -80 °C для удаления влаги в сублимационной сушилке (GAMMA1-16LSC, Marin Christ Co., Ltd., Остероде, Германия). Затем обезвоженные секции корней измельчали в мелкий порошок с помощью молотковой мельницы-циклона (JXFM110, Daji Electric Instrument Co., Ltd., Ханчжоу, Китай). Полученный порошок просеивали через сито с размером ячеек 100 меш для обеспечения однородности и хранили в темной, сухой, низкотемпературной среде для сохранения его качества. Для каждой группы обработки три случайно выбранных, одинаковых по размеру корня батата мыли, сушили и обрабатывали, как описано выше.
2.2. Противогрибковая активность CAC in vitro против Rhizopus stolonifer
2.2.1. Приготовление суспензий спор и питательных сред
Приготовление питательной среды следует методологиям, описанным Arrebola и др. [10] и Li и др. [11]. Приготовление включает следующие спецификации:
Контрольная группа (CK): Картофельно-декстрозный агар (PDA) готовили с использованием стерильной деионизированной воды.
Группа обработки (0,5% в/в CAC): Питательную среду PDA готовили с использованием раствора CAC (0,5% об/об).
Rhizopus stolonifer культивировали на чашках с PDA в течение 7 дней при температуре 28 °C и относительной влажности 85%. Споры собирали, осторожно промывая мицелий стерильной деионизированной водой и впоследствии фильтруя полученную суспензию через стерильную марлю. Концентрацию суспензии спор доводили до 1 × 10⁶ КОЕ/мл, используя гемоцитометр для точного определения.
2.2.2. Измерительные показатели
Оценка диаметра колонии Rhizopus stolonifer: Семь дней после инокуляции культуры Rhizopus stolonifer центрально инокулировали на чашки с PDA для каждой группы обработки, а затем измеряли согласно Chai и др. [12]. Чашки инкубировали в перевернутом положении при температуре 28 °C и относительной влажности 85%. Диаметры колоний измеряли через 12, 24, 48, 72, 96 и 120 часов после инокуляции, используя метод крест-накрест для точности.
Морфология роста мицелия Rhizopus stolonifer: Аликвоту 20 мкл суспензии спор равномерно распределяли на среду PDA, дополненную 0,5% в/в CAC. После того как среда полностью впитала суспензию, чашки инкубировали при 28 °C и 85% относительной влажности в искусственном климатическом инкубаторе (SAFE, Saifu Experimental Instrument Co., Ltd., Нинбо, Китай). После пяти дней инкубации небольшие образцы гиф Rhizopus stolonifer собирали для микроскопического исследования для оценки морфологии мицелия. Впоследствии изображения захватывали и сохраняли для последующего анализа.
2.3. Противогрибковая активность в отношении Rhizopus stolonifer In Vivo
2.3.1. Выявление устойчивости к мягкой гнили в различных участках клубня батата
Устойчивость клубней батата к мягкой гнили оценивали с использованием модифицированного экспериментального протокола, описанного Yang и др. [13]. Клубни батата инокулировали суспензией спор Rhizopus stolonifer при стандартизированной концентрации и наносили в определенные места. Степень мягкой гнили количественно определяли путем оценки фенотипического выражения болезни как в продольных, так и в поперечных сечениях инокулированных областей. Этот метод использовали для выявления различий в устойчивости к мягкой гнили в различных участках клубней батата и для оценки эффективности обработки 0,5% CAC в/в в повышении устойчивости к мягкой гнили.
2.3.2. Влияние 0,5% в/в CAC на текстурные характеристики
Текстурные атрибуты ломтиков батата оценивали с использованием анализатора профиля текстуры (TPA) (модель TMS-PRO, Food Technology Corporation, Дрейнсвилл, Вирджиния, США) в соответствии с методологией, описанной Aiessandrini и др. [14]. Настройки TPA были установлены следующим образом: скорость перед тестом: 30 мм/мин, скорость теста: 60 мм/мин, скорость после теста: 90 мм/мин, степень сжатия: 60%, диапазон обнаружения силы: 0–500 Н, диаметр цилиндрического зонда: 5 мм, время выдержки: 5 с, и сила срабатывания: 0,2 Н.
2.3.3. Влияние 0,5% в/в CAC на активность ферментов, связанных с защитой
Процедура экстракции ферментов: Образцы клубней батата, каждый весом 1,0 г, гомогенизировали в 1 мл предварительно охлажденного фосфатного буферного раствора (PBS, 50 мМ, pH 7,8) с использованием охлажденных ступки и пестика. Полученный гомогенат затем переносили в пробирку на 15 мл и дополняли дополнительными 4 мл того же PBS. Эту смесь центрифугировали при 12 000× g в течение 10 минут при 4 °C, после чего надосадочную жидкость осторожно собирали для оценки активности пероксидазы (POD) и аскорбатпероксидазы (APX). Для определения активности каталазы (CAT) свежие образцы клубней батата (1,0 г) гомогенизировали в 1 мл предварительно охлажденного боратно-боратного буфера (50 мМ, pH 8,8). Гомогенат затем дополняли 4 мл того же буфера и центрифугировали в указанных выше условиях. Надосадочную жидкость впоследствии собирали для анализа активности CAT. Для определения активности полифенолоксидазы (PPO) свежие образцы клубней батата (1,0 г) гомогенизировали в 1 мл предварительно охлажденного ацетатно-ацетатного буфера (0,05 М, pH = 5,5). Гомогенат затем дополняли 4 мл того же буфера и центрифугировали в указанных выше условиях. Надосадочную жидкость впоследствии собирали для анализа активности PPO. Грубые экстракты ферментов для активности хитиназы (CHI) и β-1,3-глюканазы (GLU) готовили путем гомогенизации образцов клубней батата (1,0 г) в 1 мл предварительно охлажденного ацетатно-ацетатного буферного раствора (100 мМ, pH 5,0), содержащего 1 мМ ЭДТА, поливинилполипирролидон (PVPP, 2% в/в) и 5 мМ β-меркаптоэтанол. Этот гомогенат центрифугировали при 12 000× g в течение 20 минут при 4 °C, и надосадочную жидкость сохраняли в качестве грубого экстракта фермента для последующего анализа.
Анализ активности POD: Метод Wu и др. был адаптирован для определения активности POD [15]. Реакционная смесь состояла из 0,1 мл экстракта фермента, 0,1 мл 1,5% (об/об) гваякола, 2,7 мл 50 мМ фосфатного буфера (pH 7,8) и 0,1 мл 0,3 мМ перекиси водорода (H₂O₂). Поглощение при 470 нм контролировали с использованием водного бланка. Активность POD выражали в ед/кг, где ед = 0,01 OD470 мин⁻¹.
Анализ активности APX: Метод Apiamu и Asagba [16] был адаптирован для определения активности APX. Реакционная смесь включала 0,1 мл экстракта фермента, 0,1 мл 0,3 ммоль/л H₂O₂, 0,1 мл 7,5 ммоль/л аскорбиновой кислоты и 2,7 мл 50 ммоль/л фосфатного буфера (pH 7,8). Поглощение при 290 нм контролировали с использованием водного бланка. Активность APX выражали в ед/кг, где ед = 0,01 OD290 мин⁻¹.
Анализ активности CAT: Активность CAT определяли с использованием реакционной смеси, состоящей из 0,1 мл экстракта фермента, 0,1 мл 0,3 ммоль/л H2O2 и 2,8 мл 150 ммоль/л фосфатного буфера (pH 7,8). Водный бланк использовали в качестве контроля. Активность CAT выражали в ед/кг, где ед = 0,01 OD240 мин⁻¹.
Анализ активности PPO: Активность PPO определяли с использованием реакционной смеси, состоящей из 0,1 мл экстракта фермента, 0,5 мл 50 ммоль/л раствора пирокатехина и 2 мл 50 ммоль/л ацетатного буфера (pH 5,5) [17]. Водный бланк использовали в качестве контроля. Активность PPO выражали в ед/кг, где ед = 0,01 OD420 мин⁻¹.
Анализ активности CHI: Активность CHI определяли с использованием коммерческого набора для анализа (Beijing Solarbio Technology Co., Ltd., Пекин, Китай), с измерением поглощения при 585 нм. Одна единица (ед) активности CHI была определена как количество фермента, необходимое для высвобождения 1 мкг N-ацетилглюкозамина из хитина в час на грамм свежего образца. Активность CHI выражают в ед/кг.
Анализ активности GLU: Анализ активности GLU был адаптирован из Zong и др. [18]. Реакционная смесь содержала 0,1 мл экстракта фермента и 0,1 мл 10 г/л раствора ламинарина. Смесь инкубировали при 37 °C в течение 40 минут, затем добавляли 3 мл 3,5-динитросалициловой кислоты и инкубировали при 100 °C в течение 5 минут. Поглощение измеряли при 540 нм. Одна единица (ед) активности GLU представляла количество фермента, высвобождающего 1 мг глюкозы из ламинарина в час на грамм сырого веса. Активность GLU выражают в ед/кг.
2.3.4. Влияние 0,5% CAC на активность ферментов, разрушающих клеточную стенку
Процедура экстракции ферментов: Клубни батата (1,0 г) асептически растирали до пульпы с помощью предварительно охлажденных ступки и пестика в присутствии 1 мл 95% этанола. После центрифугирования при 8000× g в течение 10 минут при 4 °C осадок осторожно собирали. Повторяли процедуры экстракции и цикла центрифугирования. Конечный осадок затем ресуспендировали в 5 мл предварительно охлажденного экстракционного буфера, содержащего 1,8 М NaCl, и инкубировали на льду в течение 16 минут для обеспечения полной солюбилизации ферментов. После центрифугирования надосадочную жидкость осторожно собирали и хранили при 4 °C для последующего использования.
Анализ активности полигалактуроназы (PG): Процедура определения активности PG была модифицирована на основе работы Zdunek и др. [19]. Реакционная смесь состояла из 1 мл 50 мМ ацетатного буфера (pH 5,5), 0,5 мл 10 г/л раствора полигалактуроновой кислоты и 0,5 мл экстракта фермента. Эту смесь инкубировали при 37 °C в течение 1 часа, после чего добавляли 3 мл 3,5-динитросалициловой кислоты (DNS). Смесь затем нагревали до кипения в течение 5 минут и давали остыть перед разбавлением до конечного объема 25 мл дистиллированной водой. Поглощение регистрировали при 540 нм, с контрольной реакцией без фермента для сравнения. Активность PG количественно определяли как количество фермента, необходимое для высвобождения 1 мг восстанавливающих сахаров на грамм свежего образца в час. Активность PG выражали в ед/кг.
Анализ активности целлюлазы (Cx): Метод оценки активности Cx был адаптирован из Abu-Goukh [20]. Реакционную смесь, состоящую из 1,5 мл 10 г/л раствора карбоксиметилцеллюлозы (CMC) и 0,5 мл экстракта фермента, инкубировали при 37 °C в течение 1 часа. После этого добавляли 3 мл 3,5-динитросалициловой кислоты (DNS), и смесь доводили до кипения в течение 5 минут перед разбавлением дистиллированной водой до конечного объема 25 мл. Поглощение измеряли при 540 нм. Активность Cx рассчитывали как количество фермента, необходимое для высвобождения 1 мг восстанавливающих сахаров на грамм свежего образца в час. Активность Cx выражали в ед/кг.
2.3.5. Влияние 0,5% в/в CAC на метаболизм фенилпропана
Анализ активности фенилаланинаммиак-лиазы (PAL): Метод анализа активности PAL был адаптирован из Lister и др. [21]. Реакционная смесь содержала 0,5 мл экстракта фермента, 0,5 мл 20 ммоль/л L-фенилаланина и 3 мл 50 ммоль/л боратно-боратного буфера (pH 8,8). Смесь инкубировали при 37 °C в течение 1 часа. Поглощение при 290 нм измеряли с использованием водного бланка. Активность PAL выражали в ед/кг, где ед = 0,01 OD290 ч⁻¹.
Содержание общих фенолов: Метод Zhou и др. [22] был адаптирован для измерения содержания общих фенольных соединений. Свежую ткань клубня батата (1,0 г) гомогенизировали с 1 мл 1% соляно-кислотно-метанольного раствора в предварительно охлажденной ступке. Этот гомогенат затем переносили в пробирку на 15 мл и разбавляли до конечного объема 5 мл тем же растворителем. После центрифугирования при 12 000× g в течение 15 минут при 4 °C надосадочную жидкость собирали. Содержание общих фенольных соединений оценивали путем измерения поглощения при 280 нм с использованием 1% соляно-кислотно-метанольного раствора в качестве бланка, с результатами, выраженными в OD280/кг.
Содержание флавоноидов: Метод определения содержания флавоноидов аналогичен методу определения содержания общих фенолов. Поглощение измеряли при 325 нм относительно 1% соляно-кислотно-метанольного бланка. Содержание флавоноидов выражали в OD325/кг.
2.3.6. Влияние 0,5% в/в CAC на показатели питательного качества
Анализ содержания белка и растворимых сахаров: Для оценки содержания белка и растворимых сахаров в образцах порошка корней батата использовали ближнюю инфракрасную спектроскопию (NIRS) с помощью прибора FOSS Analytical A/S DS2500. Образцы анализировали в трех повторностях, и результаты сообщали в г/кг порошка корней.
2.4. Комплексная оценка эффектов CAC на качество батата
Анализ главных компонент (PCA): PCA проводили для упрощения размерности набора данных и оценки значимости различных атрибутов качества. Ключевые показатели качества батата включали твердость, связность, содержание белка, уровень растворимых сахаров, общие фенолы и флавоноиды. Методику PCA модифицировали на основе подхода, подробно описанного Sun и др. [23].
2.5. Статистический анализ
Организация и анализ данных: Данные систематизировали с использованием Microsoft Excel 2016 (Microsoft Corporation, Редмонд, Вашингтон, США). Статистический анализ использовал однофакторный дисперсионный анализ, и метод Дункана использовали для сравнения средних значений внутри обработок на уровне значимости p < 0,05. Анализ главных компонент и комплексные оценки выполняли в программном обеспечении SPSS 20.0 (International Business Machines Corporation, Армонк, Нью-Йорк, США). Корреляционный анализ и визуализация данных были выполнены с использованием программного обеспечения Origin, версия 2021 (OriginLab Corporation, Нортгемптон, Массачусетс, США). Каждая экспериментальная обработка была повторена три раза биологически и три раза технически.
3. Результаты и обсуждение
3.1. Влияние 0,5% в/в CAC на активность Rhizopus stolonifer
Диаметр колонии Rhizopus stolonifer прогрессивно увеличивался с длительным хранением. Однако группа, обработанная 0,5% в/в CAC, показала заметно уменьшенный диаметр колонии по сравнению с контрольной группой (CK) (Рисунок 1A). Подавляющее воздействие 0,5% в/в CAC на расширение колонии сохранялось, достигнув снижения размера на 32,54% относительно контроля после 120 часов (Рисунок 1B(f,l)). После 5-дневного периода инкубации морфологию мицелия Rhizopus stolonifer исследовали под микроскопом при 400-кратном увеличении. Мицелий контрольной группы был гладкой текстуры, с неповрежденным клеточным содержимым и активным ростом (Рисунок 2A). Напротив, клетки, обработанные 0,5% в/в CAC, показали потемнение, вакуолизацию и значительное изменение внутренней структуры (Рисунок 2B).
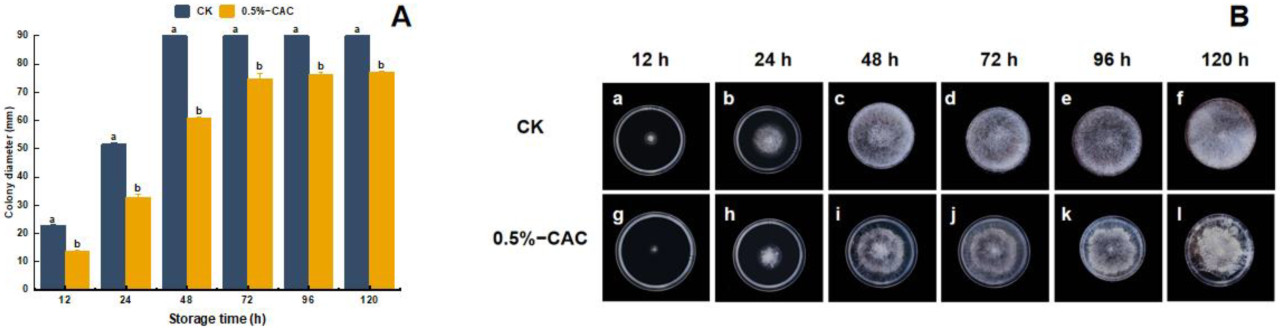
Рисунок 1. Диаметр колонии (A) и рост (B) Rhizopus stolonifer на среде PDA через 12 (a,g), 24 (b,h), 48 (c,i), 72 (d,j), 96 (e,k) и 120 часов (f,l). Статистически значимые различия между средними значениями (p < 0,05) обозначены буквами над столбцами гистограммы.
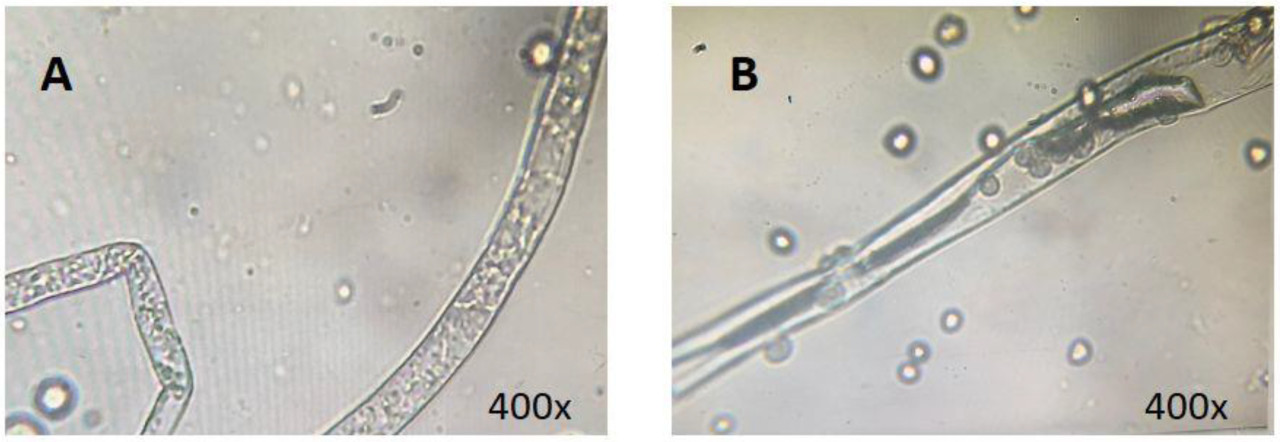
Рисунок 2. Оценка морфологии мицелия Rhizopus stolonifer после 5 дней инкубации. Сравнение между контрольной группой (CK) (A) и группой обработки 0,5% в/в CAC (B).
Rhizopus stolonifer размножается через образование спор и рост мицелия. Нарушение структуры Rhizopus stolonifer является эффективной стратегией для снижения его жизнеспособности и нарушения физиологических процессов. Wang и др. [24] продемонстрировали, что 1-октен-3-ол может ингибировать Rhizopus stolonifer, увеличивая проницаемость мембраны, изменяя морфологию мицелия и нарушая структуру органелл. Аналогично, Nickerson и Leastman [25] подробно описали ингибирующее действие цианобактерий на митохондриальный биосинтез Rhizopus stolonifer, что приводило к нарушению споруляции и образования зародышевой трубки. В данном исследовании обработка 0,5% CAC может снижать грибковую активность, разрушая структуру мицелия и ингибируя рост мицелия. Таким образом, мы заключаем, что обработка 0,5% CAC может эффективно предотвращать возникновение мягкой гнили у батата.
3.2. Физиологический ответ батата на мягкую гниль после обработки 0,5% в/в CAC
3.2.1. Влияние предварительной обработки 0,5% в/в CAC на процесс заражения Rhizopus stolonifer
Фенотип является прямым отражением генетических признаков и условий окружающей среды организмов. Оценка эффективности борьбы с болезнями, сохранности и хранения может быть эффективно проведена через фенотипическое наблюдение [26]. Исследовалось прогрессирование болезни в различных тканях батата после заражения Rhizopus stolonifer. При обследовании продольных срезов наблюдалось минимальное распространение пятен болезни, причем первичным способом заражения было боковое, а не вертикальное (Рисунок 3A). Поперечное сечение инокулированных участков клубней батата выявило отчетливые закономерности развития болезни. В частности, на 3-й день после инокуляции коричневое обесцвечивание появилось на концах клубней в контроле, в то время как центральная область оставалась незатронутой. К 7-му дню признаки гнили стали очевидны на обоих концах, а средняя часть показала признаки потемнения. После обработки 0,5% в/в CAC расширение поражений было сокращено через все участки клубня, в отличие от контрольных клубней, у которых наблюдалось прогрессирующее развитие поражений, изменение цвета тканей и последующий внутренний распад (Рисунок 3B). В этом исследовании результаты подчеркивают эффективность 0,5% в/в CAC в снижении инфекций Rhizopus stolonifer и сокращении заболеваемости мягкой гнилью. Выдвигается гипотеза, что изменение размера пятен болезни может быть связано с отсроченной активацией некоторых защитных механизмов у батата сразу после обработки 0,5% CAC.
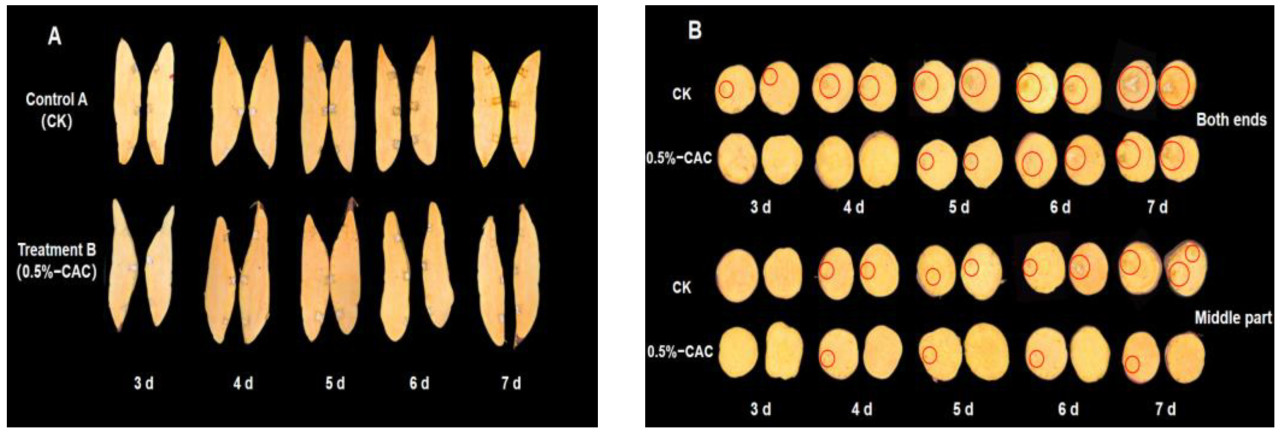
Рисунок 3. Фенотипы и прогрессирование поражений в различных участках клубня батата после инокуляции Rhizopus stolonifer. Вертикальные фенотипы (A) и поперечное сечение расширения поражений (B).
3.2.2. Влияние предварительной обработки 0,5% в/в CAC на текстурные характеристики при стрессе мягкой гнили
Твердость и связность являются ключевыми показателями текстуры, характеризующими сопротивление материала деформации и его внутреннюю прочность сцепления. Zhao и др. [27] обнаружили, что большая твердость у батата была связана с более сильной устойчивостью к мягкой гнили. В этом исследовании наблюдались сходные результаты (Рисунок 4). Контрольная группа испытала быстрое снижение текстурных характеристик после 2 дней, тогда как группа, обработанная 0,5% в/в CAC, проявляла более медленную скорость деградации текстуры, с значительным размягчением только после 3 дней. К 7-му дню батат, подвергнутый обработке 0,5% в/в CAC, сохранял на 3,57% и 4,88% большую связность в средней части и на обоих концах соответственно, чем в контроле (Рисунок 4A). Соответственно, твердость в обработанных CAC клубнях была повышена на 12,58% и 10,42% в средней и обеих конечных секциях по сравнению с контролем (Рисунок 4B). Батат, обработанный 0,5% в/в CAC, проявлял лучшие текстурные характеристики, что позволяет предположить, что эта обработка может улучшить устойчивость к мягкой гнили, поддерживая текстурные характеристики.
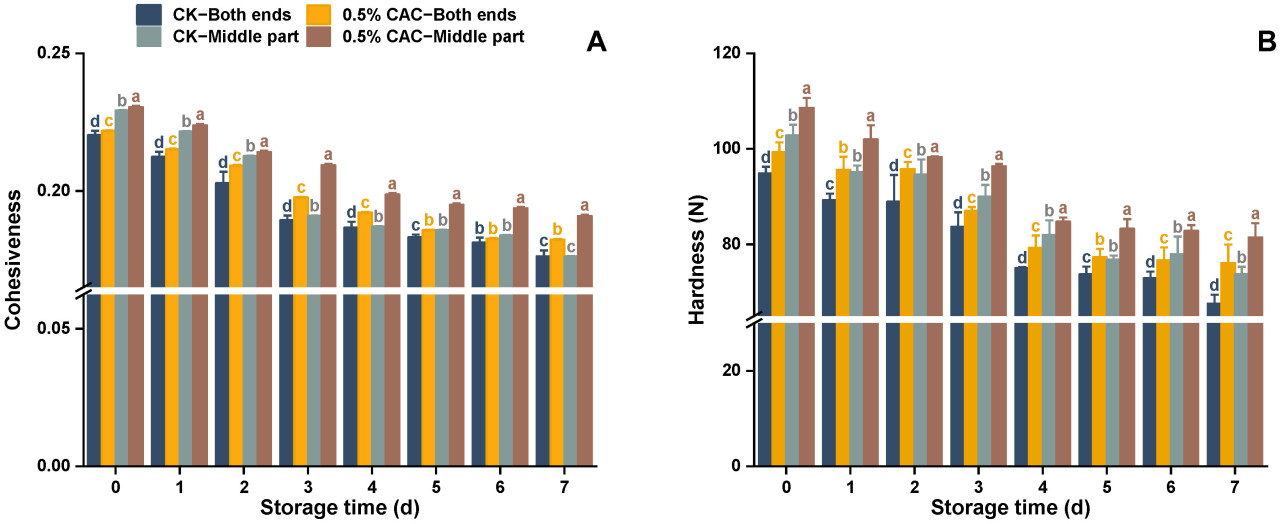
Рисунок 4. Изменения связности (A) и твердости (B) в различных участках клубня батата в течение 7-дневного заражения Rhizopus stolonifer. Статистически значимые различия (p < 0,05) между средними значениями обозначены буквами над столбцами гистограммы.
3.2.3. Влияние предварительной обработки 0,5% в/в CAC на активность ферментов, связанных с защитой, при стрессе мягкой гнили
POD является важной оксидоредуктазой, которая играет ключевую роль в ответе на атаку патогенов, окислительный стресс и другие проблемы. Она способствует превращению углеводов ткани в лигнин. Этот процесс усиливает лигнификацию, тем самым способствуя защите растений [28]. В этом исследовании применение 0,5% в/в CAC значительно повысило активность POD по сравнению с контролем (Рисунок 5A). Активность POD в клубнях батата демонстрировала двухфазный тренд, достигая пика на пятый день после обработки. В частности, на пятый день активность POD в середине контроля превышала таковую на обоих концах на 11,08%, в то время как в группе, обработанной 0,5% в/в CAC, средняя часть показала на 2,14% более высокую активность, чем оба конца. На пятый день активность POD на обоих концах и в средней части обработанных CAC клубней была заметно выше, будучи в 2,74 и 2,52 раза больше, соответственно, чем у контроля. Liu и др. [9] и Wang и др. [29] сообщали о сходных результатах и показали, что усиленная активность POD может улучшить устойчивость к болезням. Выдвигается предположение, что обработка 0,5% в/в CAC активирует защитные механизмы, запускает лигнификацию и укрепляет клеточную стенку, тем самым усиливая устойчивость батата и препятствуя заражению Rhizopus stolonifer.
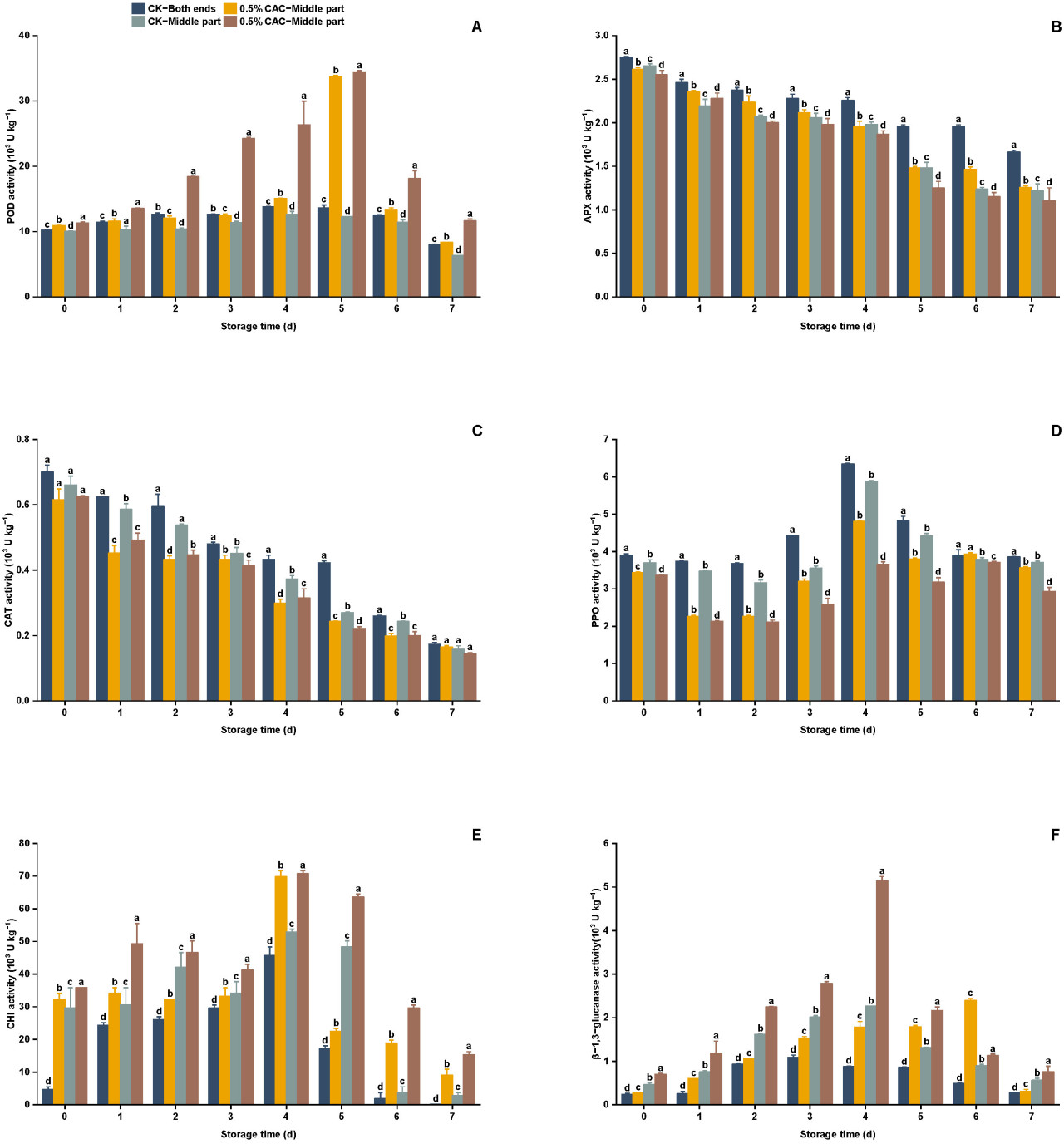
Рисунок 5. Активность ферментов, связанных с защитой, оценивалась в различных группах обработки батата в течение 7 дней хранения. Анализировались следующие ферменты: POD (A), APX (B), CAT (C), PPO (D), CHI (E) и GLU (F). Статистически значимые различия (p < 0,05) между средними значениями обозначены буквами над столбцами гистограммы.
H₂O₂ является ключевым компонентом механизма устойчивости к болезням у фруктов и овощей. Снижение активностей APX и CAT способствует накоплению H₂O₂, что помогает в процессе лигнификации поврежденных тканей, тем самым создавая барьер, который может смягчать распространение патогенов [30,31]. Роль APX и CAT в индуцированной устойчивости может варьироваться среди разных видов растений или даже среди разных тканей внутри одного и того же растения [32]. Zhu и др. [33] обнаружили, что метилжасмонат усиливает экспрессию и активность CAT и APX, что убирает избыток H₂O₂ и снижает окислительное повреждение белков, приводя к индуцированной устойчивости против серой гнили у томатов. И наоборот, Wang и др. [34] пришли к другому выводу в рисе. В этом исследовании активности APX и CAT в целом снижались во всех группах обработки. Обработка 0,5% в/в CAC заметно снижала активности APX и CAT по сравнению с контролем. В частности, на 7-й день хранения активность APX была снижена на 24,44% и 9,29% на концах и в середине обработанного CAC батата соответственно, по сравнению с контролем (Рисунок 5B). Соответственно, активность CAT в этих секциях была снижена на 5,04% и 8,80% соответственно (Рисунок 5C). Выдвигается гипотеза, что усиление устойчивости батата к болезням опосредовано ингибированием активностей APX и CAT, таким образом способствуя накоплению H₂O₂ и потенциально внося вклад в защитный ответ растения.
PPO является ключевым ферментом в процессе потемнения фруктов и овощей; повышенная активность PPO коррелирует с усиленным потемнением, размягчением и ускоренным старением фруктов и овощей [35,36]. Батат, обработанный 0,5% в/в CAC, проявлял превосходные фенотипические качества по сравнению с контролем при заражении, задерживая начало потемнения (Рисунок 3). Во время хранения активность PPO в клубнях батата следовала двухфазному тренду, достигая пика на четвертый день периода наблюдения. Примечательно, что применение 0,5% в/в CAC значительно подавляло активность PPO во всех секциях по сравнению с контролем (Рисунок 5D). В частности, на четвертый день активность PPO снизилась на 24,26% в концевых секциях и на 37,77% в средних секциях обработанных CAC клубней. Lin и др. [37] сообщали о сходных результатах и показали, что усиление активности PPO с помощью обработки ε-поли-L-лизином могло подавлять послеуборочные болезни маракуйи, вызванные Lasiodiplodia theobroma. Делается вывод, что обработка 0,5% в/в CAC может усиливать устойчивость батата к болезням, сдерживая окисление фенолов через PPO.
CHI и GLU, как белки, связанные с патогенезом (PR), являются неотъемлемой частью защитного механизма растения против грибковых патогенов. Эти ферменты разрушают клеточные стенки вторгающихся грибов, тем самым обеспечивая прямую линию защиты [38,39]. CHI обычно встречается во фруктах и овощах и усиливает устойчивость к болезням, расщепляя хитин, компонент грибковых клеточных стенок. Между тем, активность GLU тесно связана с послеуборочной устойчивостью фруктов и овощей к болезням, поскольку на нее влияют различные абиотические и биотические факторы [40]. Zhang и др. [41] сообщили, что олеаноловая и урсоловая кислоты могут усиливать устойчивость плодов к серой гнили, увеличивая активности CHI и GLU, таким образом замедляя гниение плодов. Эта корреляция между усиленной устойчивостью к болезням и повышенными активностями CHI и GLU была дополнительно поддержана Ye и др. [42] и Zhou и др. [43] в их соответствующих исследованиях. Сходные результаты были найдены в этом эксперименте. Активность CHI демонстрировала двухфазный тренд, достигая своего зенита на четвертый день по сравнению с контролем (Рисунок 5E). В частности, на четвертый день хранения активность CHI на обоих концах и в средних секциях обработанного CAC батата была увеличена на 52,73% и 33,78% соответственно, относительно контроля. Аналогично, активность GLU отражала двухфазную закономерность, достигая пика на четвертый день. В течение периода хранения группа обработки 0,5% в/в CAC стабильно демонстрировала значительно более высокую активность GLU по сравнению с контролем. На четвертый день активность GLU на обоих концах и в средних секциях обработанного CAC батата была в 1,03 и 1,28 раза соответственно выше, чем наблюдаемая в контрольной группе (Рисунок 5F). Это исследование подчеркивает потенциал обработки CAC в укреплении защитных способностей батата против мягкой гнили, вероятно, через модуляцию активности PR-белков, что может иметь значительные последствия для послеуборочного управления и продовольственной безопасности.
В заключение, обработка 0,5% в/в CAC изменила активность ферментов, связанных с защитой, и улучшила устойчивость батата к мягкой гнили.
3.2.4. Влияние предварительной обработки 0,5% в/в CAC на ферменты, разрушающие клеточную стенку, при стрессе мягкой гнили
PG и Cx являются ключевыми ферментами, разрушающими клеточную стенку, которые играют значительную роль в ухудшении состояния растительной ткани [44,45]. Cao и др. [46] установили корреляцию между повышенной активностью этих ферментов клеточной стенки и ускоренной порчей батата. Во время хранения активность PG увеличивалась с периодом хранения в обеих группах обработки. Однако она оставалась стабильно ниже в группе 0,5% CAC по сравнению с контролем (Рисунок 6A). В частности, на 7-й день активность PG на обоих концах и в средних секциях обработанного CAC батата была снижена на 16,62% и 13,49% соответственно, по сравнению с контролем. Активность Cx показала двухфазную закономерность, достигнув своего пика на четвертый день хранения в обеих группах обработки. Тем не менее, активность Cx была стабильно ниже в группе, обработанной 0,5% CA, по сравнению с контрольной группой (Рисунок 6B). На критический четвертый день активность Cx снизилась на 24,47% и 18,61% на концах и в средних секциях обработанного CAC батата соответственно, по сравнению с контролем. Это предполагает, что обработка 0,5% в/в CAC может эффективно ингибировать активность ферментов, разрушающих клеточную стенку, и задерживать порчу батата. Этот вывод отражает результаты, показанные на Рисунке 3. Выдвигается гипотеза, что модуляция активности PG и Cx с помощью обработки CAC может быть стратегическим подходом для повышения долговечности батата против послеуборочных болезней.
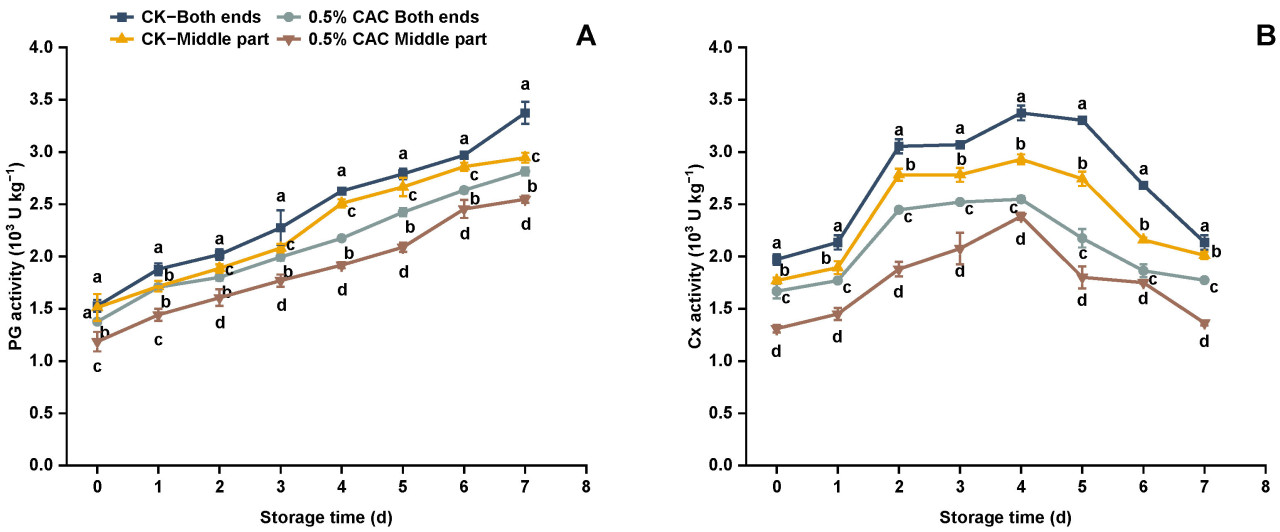
Рисунок 6. Активность PG (A) и Cx (B) в различных группах обработки батата в течение 7 дней хранения. Статистически значимые различия между средними значениями (p < 0,05) обозначены буквами над графиком.
3.2.5. Влияние предварительной обработки 0,5% в/в CAC на путь метаболизма фенилпропана при стрессе мягкой гнили
Метаболический путь фенилпропаноидов играет ключевую роль в защитных механизмах растений [47,48]. PAL является лимитирующим скорость ферментом в пути фенилаланина и катализирует превращение L-фенилаланина в антимикробные соединения, включая фенолы и флавоноиды. Этот путь активируется в ответ на стресс у растений, усиливая устойчивость к болезням [49]. Фенольные соединения и флавоноиды известны своими антиоксидантными и антимикробными свойствами [50,51]. Эти соединения накапливаются в тканях растений при вторжении патогенов, образуя защитный барьер [52]. В этом исследовании было обнаружено, что все части клубня активировали метаболический путь фенилпропаноидов после обработки 0,5% CAC в/в. Активность PAL проявляла двухфазную закономерность, изначально увеличиваясь до достижения пика на четвертый день. Во всех группах обработки активность PAL была ниже на обоих концах батата, чем в средней секции. Обработка 0,5% в/в CAC значительно увеличила активность PAL по сравнению с контролем (Рисунок 7A). В частности, на четвертый день активность PAL в концевой и средней секциях обработанного CAC батата была увеличена на 72,49% и 56,09% соответственно, по сравнению с контролем. Обработка 0,5% в/в CAC значительно увеличила содержание общих фенолов по сравнению с контролем (Рисунок 7B). В частности, на четвертый день содержание общих фенолов на обоих концах и в средних секциях обработанного CA батата было увеличено на 13,21% и 13,08% соответственно. Аналогично, обработка 0,5% в/в CAC также значительно увеличила содержание флавоноидов по сравнению с контролем (Рисунок 7C). На пятый день содержание флавоноидов на обоих концах и в средних секциях обработанного CAC батата было увеличено на 4,43% и 10,56% соответственно, относительно контроля. Эти находки согласуются с Liu и др. [9], которые сообщали о сходных результатах относительно активации пути фенилпропаноидов. Выдвигается гипотеза, что модуляция метаболического пути фенилпропана с помощью обработки CAC может быть стратегическим подходом для повышения долговечности батата против послеуборочных болезней.
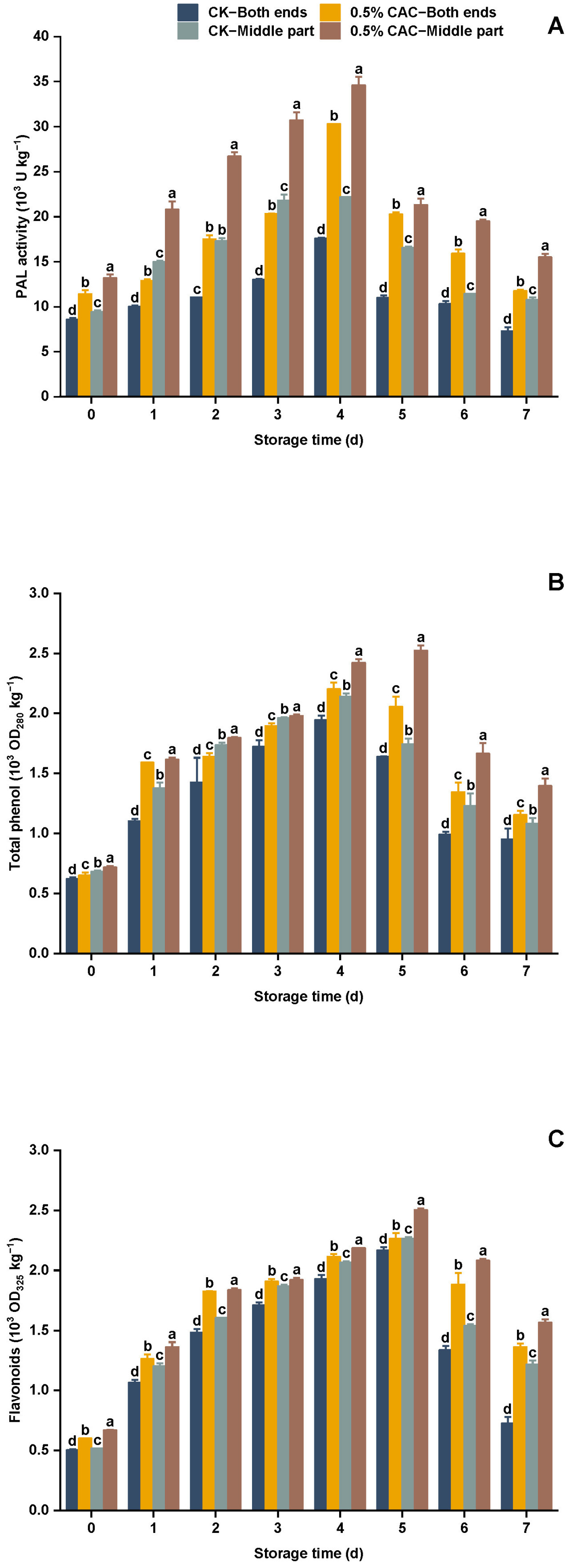
Рисунок 7. Изменения активности PAL (A), содержания общих фенолов (B) и флавоноидов (C) в различных группах обработки батата в течение 7 дней хранения. Статистически значимые различия (p < 0,05) между средними значениями обозначены буквами над столбцами гистограммы.
3.2.6. Влияние предварительной обработки 0,5% в/в CAC на питательное качество при стрессе мягкой гнили
Содержание белка и растворимых сахаров являются критически важными питательными параметрами в батате, значительно влияющими как на пищевую ценность, так и на органолептические свойства [53]. Группа, обработанная 0,5% в/в CAC, стабильно поддерживала более высокие уровни содержания белка и растворимых сахаров, чем контроль (Рисунок 8A,B). На 7-й день содержание белка в группе 0,5% CAC увеличилось на 33,87% в целом по сравнению с контролем, при этом содержание белка в центре увеличилось на 9,45%. На 7-й день содержание растворимых сахаров было на 1,76% выше в целом и на 5,27% выше в средней секции группы 0,5% в/в CAC относительно контроля. Содержание белка показало снижение в течение периода хранения в обеих группах обработки, что может быть связано с ухудшением белка после атаки болезни во время хранения. Содержание растворимых сахаров в обеих группах показало возрастающий тренд во время хранения, что может быть связано с осахаривающей реакцией во время хранения [54]. Опираясь на работу Chandrajith и др. [55], это исследование продемонстрировало, что послеуборочная обработка 0,5% в/в CAC была эффективна в сохранении этих ключевых питательных веществ. Это открытие особенно важно для оптимизации стратегий хранения, чтобы обеспечить сохранение важнейших диетических компонентов.
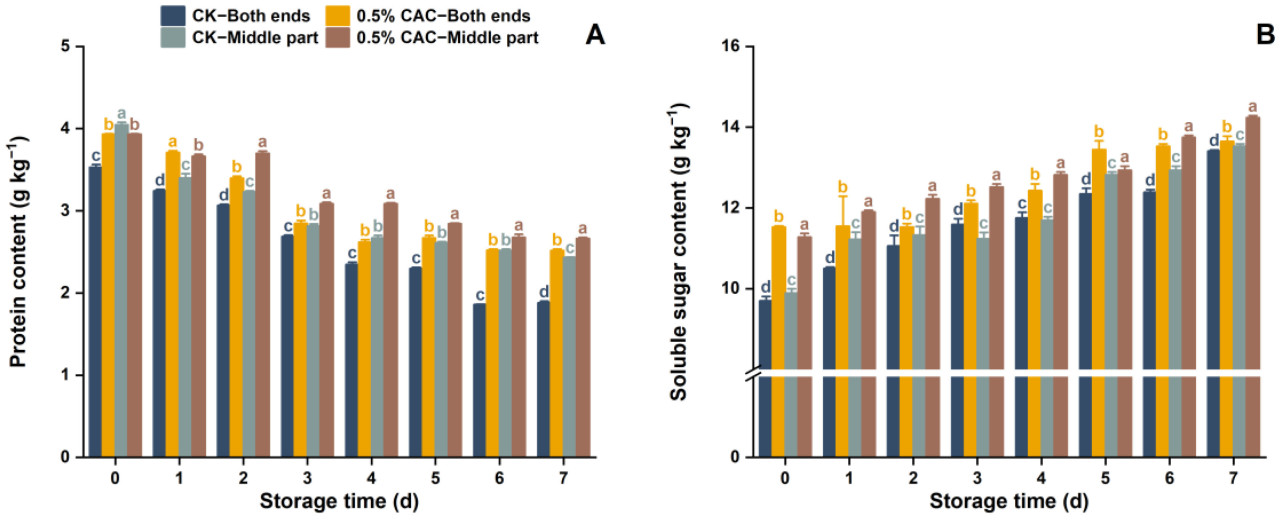
Рисунок 8. Изменения содержания белка (A) и растворимых сахаров (B) в различных участках клубня батата в течение 7 дней хранения в контрольных условиях и при обработке 0,5% в/в CAC. Статистически значимые различия (p < 0,05) между средними значениями обозначены буквами над столбцами гистограммы.
3.3. Анализ главных компонент и комплексная оценка качества хранения батата, обработанного 0,5% в/в CAC
Оценка послеуборочного качества плодов является сложной задачей, которая часто требует оценки множественных параметров, что представляет сложность для комплексных оценок [11]. PCA зарекомендовал себя как ценный инструмент в анализе биологических характеристик и послеуборочных исследований [56]. Chu и др. [57] успешно применили PCA для создания комплексной модели оценки качества лилий, подчеркивая преимущества паровой бланшировки и обработки CAC в предотвращении потемнения и сохранении питательной ценности.
Чтобы определить влияние обработки 0,5% в/в CAC на усиление устойчивости к мягкой гнили у батата, в этом исследовании был применен PCA для оценки качества клубней. Каждый показатель был сведен к трем главным компонентам (Таблица 1). Флавоноиды и общие фенолы показали наибольшие нагрузки на PC1, в основном отражая накопление антимикробных соединений. Твердость, связность и белок показали наибольшие нагрузки на PC2, что позволяет предположить, что этот компонент в основном отражает текстурные свойства образцов. PC3 в наибольшей степени зависел от содержания растворимых сахаров, указывая на его роль в формировании общего питательного качества (Таблица 2).
Характеристический вектор коэффициента главной компоненты был получен путем деления коэффициента нагрузки на соответствующий характеристический корень, как подробно описано в Таблице 3. Линейные дисперсии трех главных компонент и уравнение комплексной оценки получены следующим образом:
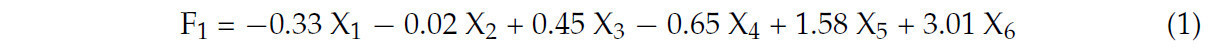
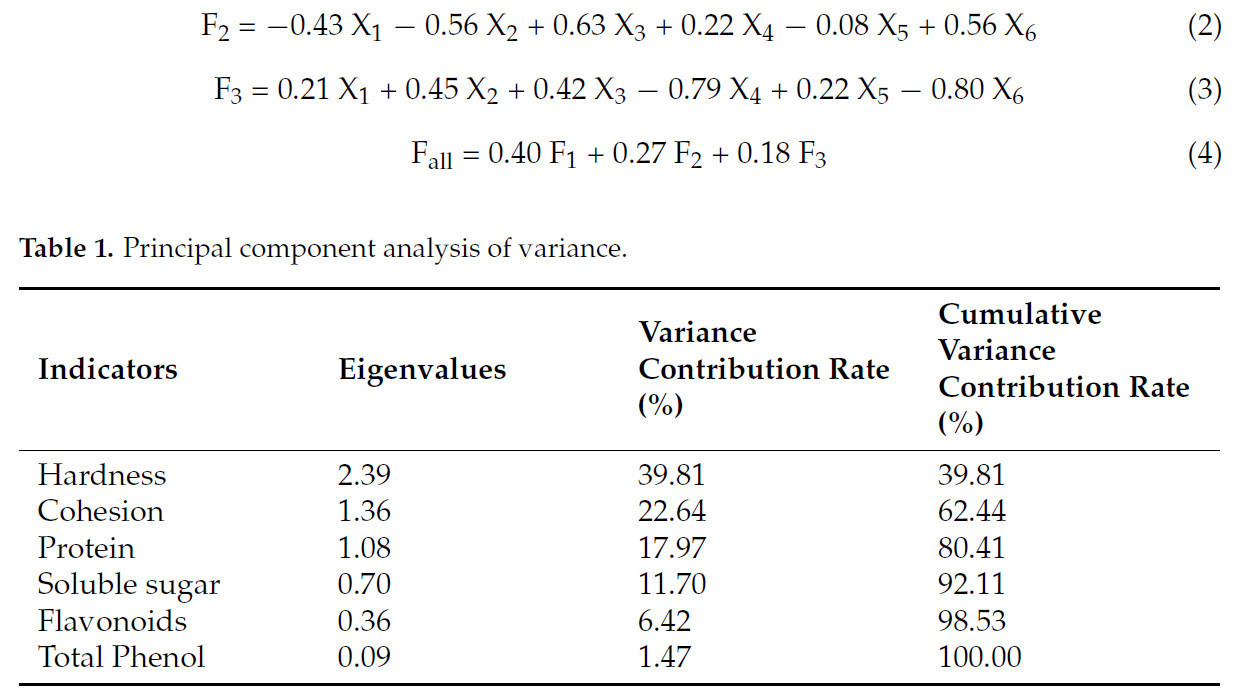
Таблица 1. Анализ главных компонент дисперсии.
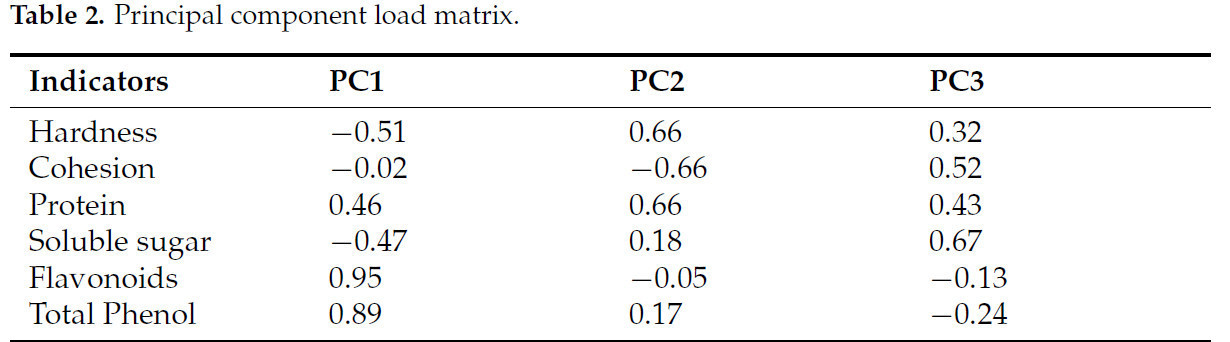
Таблица 2. Матрица нагрузок главных компонент.
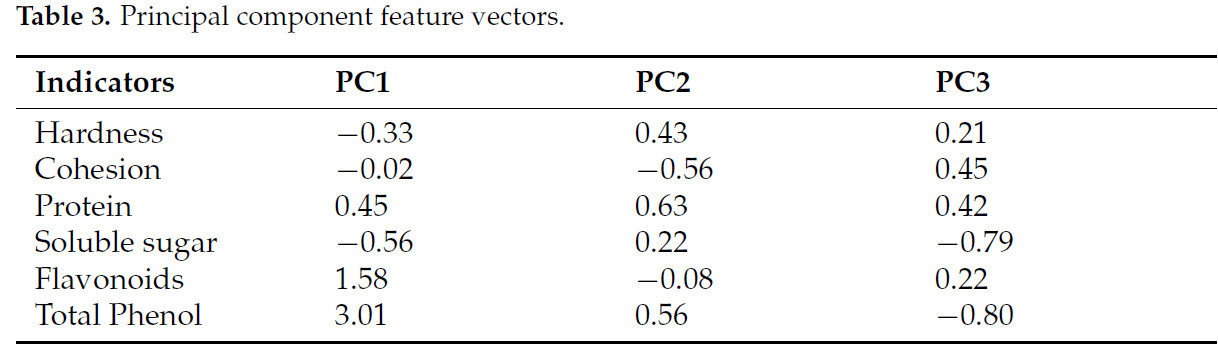
Таблица 3. Характеристические векторы главных компонент.
Комплексный балл каждой обработки был рассчитан согласно Уравнению (4). В течение всего периода хранения баллы были стабильно выше в группе обработки 0,5% в/в CAC по сравнению с контролем. В каждой группе обработки средние участки клубней демонстрировали более высокие баллы, чем концевые участки (Рисунок 9A). Эти результаты указывают, что после заражения Rhizopus stolonifer средняя ткань сохраняла лучшее качество, чем концевые участки, и что обработка 0,5% в/в CAC способствовала поддержанию лучшего качества по сравнению с контролем.
3.4. Корреляционный анализ качества хранения батата, обработанного 0,5% CAC
Устойчивость растений к болезням представляет собой сложный процесс, включающий многочисленные метаболические пути, некоторые из которых могут действовать синергетически, в то время как другие могут оказывать ингибирующее действие [58]. Проведение корреляционного анализа каждого показателя может помочь выяснить механизмы, лежащие в основе устойчивости к мягкой гнили, индуцированной 0,5% в/в CAC.
Корреляционный анализ выявил, что уровни POD были положительно связаны с метаболизмом фенилпропана. Активность APX положительно коррелировала с CAT и отрицательно - с PG. Эти ферменты демонстрировали положительные связи с флавоноидами и растворимыми сахарами, но отрицательные - с белками. Метаболизм фенилпропана положительно коррелировал с Cx, CHI и GLU. Белок отрицательно коррелировал с растворимыми сахарами (Рисунок 9B).
Результаты показали, что во время индуцированной 0,5% в/в CAC устойчивости к мягкой гнили у батата существует синергетическая взаимосвязь между путем метаболизма фенилпропаноидов, путем метаболизма клеточной стенки, белками, связанными с патогенезом (PR), и антиоксидантными ферментами. Эта взаимосвязь предоставляет направление для дальнейшего понимания механизма устойчивости батата к мягкой гнили и имеет важное значение для разработки устойчивых к болезням сортов сельскохозяйственных культур.
4. Выводы
Настоящее исследование убедительно продемонстрировало, что обработка 0,5% в/в CAC значительно ингибирует рост и развитие Rhizopus stolonifer - возбудителя мягкой гнили батата. Примечательно, что средняя часть клубня батата показала более высокую естественную устойчивость к инфекции по сравнению с концевыми участками. Обработка 0,5% в/в CAC усиливает устойчивость батата к мягкой гнили через множественные механизмы, включая активацию защитных ферментов, подавление активности ферментов, разрушающих клеточную стенку, и накопление антимикробных соединений.
Благодаря своей эффективности и безопасности, обработка 0,5% в/в CAC представляет собой перспективную стратегию для продления срока хранения батата и управления мягкой гнилью. Результаты данного исследования предоставляют ценную информацию, которая может быть использована для совершенствования методов сохранения и хранения батата, потенциально влияя на будущие садоводческие и послеуборочные стратегии.
Ссылки
1. Liu, Q. Sweet potato omics and biotechnology in china. Plant Omics J. Plant Mol. Biol. Omics 2011, 4, 295–301. [Google Scholar]
2. Xie, Y.; Guo, X.; Jia, Z.; Ma, P.; Bian, X.; Yu, Y. Progresses and prospects on edible Sweet potato breeding in China. Jiangsu J. Agric. Sci. 2018, 34, 1419–1424. [Google Scholar]
3. Yang, J.; Yu, G.; Nam, S.S.; Lee, S.Y.; Paul, N.C. First report of fusarium rot caused by fusarium circinatum on sweet potato (Ipomoea batatas). J. Plant Pathol. 2019, 101, 407. [Google Scholar] [CrossRef]
4. Qiu, B.; Xu, F.; Sun, J.; Han, Z.; Wang, Z. Review on research progress of sweet potato soft rot and its control methods. Agric. Technol. 2022, 42, 91–94. [Google Scholar]
5. Qi, Y.; Wang, F. Food Additives; Ocean Press: Beijing, China, 2014. [Google Scholar]
6. Cao, J. Effects of SA, ASM, INA and Citric Acid on Postharvest Disease Resistance and Quality of Ypear Fruit. Ph.D. Thesis, China Agricultural University, Beijing, China, 2005. [Google Scholar]
7. Dai, X.; Sun, Z.; Han, L.; Ju, G. Callus Induction and Inhibition Effect of Citric Acid on Browning of Perennial Ryegrass Callus. J. Nucl. Agric. Sci. 2015, 29, 1893–1900. [Google Scholar]
8. Tapia-Rodriguez, M.R.; Bernal-Mercado, A.T.; Palomares-Navarro, J.J.; Sugich-Miranda, R.; Enciso-Martinez, Y.; Cruz-Valenzuela, M.R.; Ayala-Zavala, J.F. Citric acid and CaCl2 extended the shelf life, maintained antioxidant capacity, and improved sensory attributes of fresh-cut kiwifruit. J. Hortic. Postharvest Res. 2021, 4, 67–80. [Google Scholar] [CrossRef]
9. Liu, J.; Zhang, H.; Cheng, S.; He, X.; Zhang, M. Effect of ginger extract on sweet potatoes with soft rot disease in room temperature logistics and its mechanism. Food Ferment. Ind. 2020, 46, 57–65. [Google Scholar]
10. Arrebola, E.; Sivakumar, D.; Bacigalupo, R.; Korsten, L. Combined application of antagonist bacillus amyloliquefaciens and essential oils for the control of peach postharvest diseases. Crop Prot. 2010, 29, 369–377. [Google Scholar] [CrossRef]
11. Li, H.; Ma, Q.; Lin, X.; Zeng, P.; Chen, J. Comprehensively Analyzing the effect of harvest maturity on storage quality of jinsha pomelo based on PCA. Sci. Technol. Food Ind. 2019, 40, 255–262. [Google Scholar]
12. Chai, H.; Jia, J.; Bai, X.; Meng, L.; Zhang, W.; Jin, R.; Wu, H.; Su, Q. Identification of Pathogenic Fusarium spp. Causing Maize Ear Rot and Susceptibility of Some Strains to Fungicides in Jilin Province. Sci. Agric. Sin. 2023, 56, 64–78. [Google Scholar]
13. Yang, D.; Xu, Z.; Zhao, Y.; Zhang, C.; Sun, H.; Xie, Y. Research on the identification method of sweet potato soft rot resistance and its resistance evaluation to sweet potato germplasm resources. Acta Agric. Boreali-Sin. 2014, 29, 54–56. [Google Scholar]
14. Aiessandrini, L.; Balestra, F.; Romani, S.; Rocculi, P.; Rosa, M.D. Physicochemical and sensoryproperties of fresh potato-based pasta (Gnocchi). J. Food Sci. 2020, 75, S542–S547. [Google Scholar] [CrossRef]
15. Wu, J.; Zhang, J.; Ni, W.; Xu, X.; George, M.S.; Lu, G. Effect of Heat Treatment on the Quality and Soft Rot Resistance of Sweet Potato during Long-Term Storage. Foods 2023, 12, 4352. [Google Scholar] [CrossRef] [PubMed]
16. Apiamu, A.; Asagba, S.O. Zinc-cadmium interactions instigated antagonistic alterations in lipid peroxidation, ascorbate peroxidase activity and chlorophyll synthesis in Phaseolus vulgaris leaves. Sci. Afr. 2021, 11, e00688. [Google Scholar] [CrossRef]
17. Dong, H. Effects of chitosan coating on quality and shelf life of peeled litchi fruit. J. Food Eng. 2004, 64, 355–358. [Google Scholar] [CrossRef]
18. Zong, Y.; Liu, J.; Li, B.; Qin, G.; Tian, S. Effects of yeast antagonists in combination with hot water treatment on postharvest diseases of tomato fruit. Biol. Control 2010, 54, 316–321. [Google Scholar] [CrossRef]
19. Zdunek, A.; Kozioł, A.; Pieczywek, P.M.; Cybulska, J. Evaluation of the nanostructure of pectin, hemicellulose and cellulose in the cell walls of pears of different texture and firmness. Food Bioprocess. Technol. 2014, 7, 3525–3535. [Google Scholar] [CrossRef]
20. Abu-Goukh, A.B.; Elhassan, S.Y. Changes in pectic substances and cell wall degrading enzymes during muskmelon fruit ripening. Univ. Khartoum J. Agric. Sci. 2019, 25, 73–93. [Google Scholar] [CrossRef]
21. Lister, C.E.; Lancaster, J.E.; Walker, J.R. Developmental changes in enzymes of flavonoid biosynthesis in the skins of red and green apple cultivars. J. Sci. Food Agric. 1996, 71, 313–320. [Google Scholar] [CrossRef]
22. Zhou, Y.; Deng, L.; Zeng, K. Enhancement of biocontrol efficacy of Pichia membranaefaciens by hot water treatment in postharvest diseases of citrus fruit. Crop Prot. 2014, 63, 89–96. [Google Scholar] [CrossRef]
23. Sun, W.; Zhou, Z.; Li, Y.; Xu, Z.; Xia, W.; Zhong, F. Differentiation of flue-cured tobacco leaves in different positions based on neutral volatiles with principal component analysis (PCA). Eur. Food Res. Technol. 2012, 235, 745–752. [Google Scholar] [CrossRef]
24. Wang, X.; Huang, M.; Wang, Y.; Yang, W.; Shi, J. Effect of 1-octen-3-ol on soft rot of postharvest peach fruit. Acta Microbiol. Sin. 2022, 62, 4878–4893. [Google Scholar]
25. Nickerson, K.W.; Leastman, E. Cerulenin inhibition of spore germination in Rhizopus stolonifer. Exp. Mycol. 1978, 2, 26–31. [Google Scholar] [CrossRef]
26. Zhang, L.; Ma, Y.; Zhang, Y.; Zhou, H.; Li, X. The effect of controlled atmosphere on the storage quality of strawberry. J. Shanxi Agric. Univ. 2019, 39, 41–45. [Google Scholar]
27. Zhao, M.; Pang, L.; Cheng, J.; Lu, G.; Lu, X.; Wang, S. Effects of Ceratocystis Fimbriata Infection on Cell Wall of Different Resistant Sweet Potato. Packag. Eng. 2022, 43, 10–16. [Google Scholar]
28. Passardi, F.; Cosio, C.; Penel, C.; Dunand, C. Peroxidases have more functions than a Swiss army knife. Plant Cell Rep. 2005, 24, 255–265. [Google Scholar] [CrossRef]
29. Wang, R.; Li, J.; Dang, H.; Zhang, X.; Zhang, S.; Zhang, J.; Li, X. Effects of amino-oligosaccharides on induced disease resistance and biochemical indices of Wolfberry. Acta Agric. Univ. Jiangxiensis 2024, 46, 672–681. [Google Scholar] [CrossRef]
30. Kim, Y.H.; Kim, C.Y.; Song, W.K.; Park, D.S.; Kwon, S.Y.; Lee, H.S.; Bang, J.W.; Kwak, S.S. Overexpression of Sweet potato swpa4 peroxidase results in increased hydrogen peroxide production and enhances stress tolerance in tobacco. Planta 2008, 227, 867–881. [Google Scholar] [CrossRef] [PubMed]
31. Lee, D.H.; Lee, C.B. Chilling stress-induced changes of antioxi-dant enzymes in the leaves of cucumber: In gel enzyme activity as-says. Plant Sci. 2000, 159, 75–85. [Google Scholar] [CrossRef] [PubMed]
32. Para-Lobato, M.C. Fernandez-Garcia N, Olmos E, Methyl jasmonate-induced antioxidant defence in root apoplast from sunflower seedlings. Environ. Exper Bot. 2009, 66, 9–17. [Google Scholar] [CrossRef]
33. Zhu, Z.; Tian, S. Resistant responses of tomato fruit treated with exogenous methyl jasmonate toBotrytis cinerea infec-tion. Sci. Horticul 2012, 142, 38–43. [Google Scholar] [CrossRef]
34. Wang, H.; Cao, C.; Kang, J.; Zeng, F. Systemic Resistance of Rice to Bacterial Blight Induced by Salicylic Acid and Changes in Activities of Some Enzymes in Untreated Leaves. Chin. J. Rice Sci. 2002, 16, 252–256. [Google Scholar]
35. Mayer, A.M. Polyphenol oxidases in plants-recent progress. Phytochemistry 1987, 26, 11–20. [Google Scholar] [CrossRef]
36. Feng, L.; Yin, Y.; Yang, X.; Wu, C.; Sun, L. Effects of Citric Acid Treatment on Vitro Browning of Pomegranate Pericarp. Sci. Silvae Sin. 2015, 51, 42–50. [Google Scholar]
37. Lin, Y.; Chen, L.; Chen, J.; Jiang, X.; Zheng, J.; Chen, H. Effect of ε-poly-L-lysine on Postharvest Diseases and Disease—Resistant Substance Metabohsm in Passion Fruits. Food Sci. 2024, 45, 142–149. [Google Scholar]
38. Ji, C.; Kuć, J. Antifungal activity of cucumber β-1,3-glucanase and chitinase. Physiol. Mol. Plant P 1996, 49, 257–265. [Google Scholar] [CrossRef]
39. Tian, S.; Yao, H.; Deng, X.; Xu, X.; Qin, G.; Chan, Z. Characterization and expression of β-1,3-glucanase genes in jujube fruit induced by the microbial biocontrol agent cryptococcus laurentii. Phytopathology 2007, 97, 260–268. [Google Scholar] [CrossRef] [PubMed]
40. Loon, L.C.; Rep, M.; Pieterse, C.M. Significance of inducible defense related proteins in infected plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [PubMed]
41. Zhang, L.; Liu, R.; Han, Y.; Chen, H.; Wu, W.; Fang, X.; Gao, H. Effects of Cuticular Wax on Disease Resistance of Postharvest Blueberry. J. Chin. Inst. Food Sci. Technol. 2021, 21, 205–213. [Google Scholar]
42. Ye, X.; Wu, Q.; Xie, G. Effect of Preharvest Spraying Extract from Camellia oleifera Cake on the Disease Resistance of Postharvest Pitaya Fruit. Chin. J. Trop. Crops 2023, 44, 2093–2101. [Google Scholar]
43. Zhou, J.; Wang, Y.; Liu, D.; Xi, K.; Yang, P.; Cai, X.; Li, G.; Ma, J.; Jia, Q.; Le, Y. Identification and evaluation of resistance to fusarium wilt of ginger varieties and screening of resistance indexes. J. South. Agric. 2022, 53, 2557–2567. [Google Scholar]
44. Dopico, B.; Nicolos, G.; Labrador, E. Cell wall localization of the natural substrate of a β-galactosidase, the main enzyme responsible for the autolytic process of cicer arietinum epicotyl cell walls. Physiol. Plant. 1990, 80, 636–641. [Google Scholar] [CrossRef]
45. Mandels, M.; Reese, E.T. Inhibition of cellulases. Annu. Rev. Phytopathol. 1965, 3, 85–102. [Google Scholar] [CrossRef]
46. Cao, R.; Xu, S.; Li, L. Changes of cell wall components and degradation enzyme activity in sweet potato storage roots during storage. J. China Agric. Univ. 2020, 25, 65–75. [Google Scholar]
47. Stadnik, M.J.; Buchenauer, H. Inhibition of phenylalanine ammonia-lyase suppresses the resistance induced by benzothiadiazole in wheat to Blumeria graminis f. sp. tritici. Physiol. Mol. Plant 2000, 57, 25–34. [Google Scholar] [CrossRef]
48. Bourbonnais, R.; Paice, M.G. Oxidation of non-phenolic substrates. an expanded role for laccase in lignin biodegradation. FEBS Lett. 1990, 267, 99–102. [Google Scholar] [CrossRef] [PubMed]
49. Li, M.; Wang, Y.; Jiao, Y.; Liu, Y.; Qiao, L.; Liu, X. Browning Control of Fresh-Cut fruit and Vegetables by the Combination of Cold Shock and Pomelo Peel Extract Treatment. Food Res. Dev. 2023, 44, 8–14. [Google Scholar]
50. Ignat, I.; Volf, I.; Popa, V.I. A critical review of methods for characterisation of polyphenolic compounds in fruit and vegetables. Food Chem. 2021, 126, 1821–1835. [Google Scholar] [CrossRef] [PubMed]
51. Cueva, C.; Moreno-Arribas, M.V.; Martín-Álvarez, P.J.; Bills, G.; Vicente, M.F.; Rivas, C.L.; Basilio, A.; Requena, T.; Rodríguez, J.M.; Basilio, A.; et al. Antimicrobial activity of phenolic acids against commensal, probiotic and pathogenic bacteria. Res. Microbiol. 2010, 161, 372–382. [Google Scholar] [CrossRef]
52. Reimers, P.J.; Leach, J.E. Race-specific resistance to xanthomonas oryzae pv. oryzae conferred by bacterial blight resistance gene xa-10 in rice (oryza sativa) involves accumulation of a lignin-like substance in host tissues. Physiol. Mol. Plant P 1991, 38, 39–55. [Google Scholar] [CrossRef]
53. Kitahara, K.; Nakamura, Y.; Otani, M.; Hamada, T.; Nakayachi, O.; Takahata, Y. Carbohydrate components in Sweet potato storage roots: Their diversities and genetic improvement. Breed. Sci. 2017, 67, 62–72. [Google Scholar] [CrossRef] [PubMed]
54. Isherwood, F.A. Mechanism of starch-sugar interconversion in Solanum tuberosum. Phytochemistry 1976, 15, 33–41. [Google Scholar] [CrossRef]
55. Chandrajith, U.G.; Wijewardane, R.M.; Wasala, W.M.; Dissanayake, C.A. Combined Effect of Citric Acid and Ascorbic Acid as Chemical PreTreatments to Prevent Enzymatic Browning in Fresh Cut Ambul Banana (Musa spp). In Proceedings of the 8th International Research Conference, Seville, Spain, 18–20 November 2015. KDU, Sri Lanka, 2015. [Google Scholar]
56. Hu, B.; Li, G.; Xie, Z. Effect of different concentration of 1-MCP treatment on quality of Winter Jujube in Northern Hubei under Cold Storage. Food Res. Dev. 2014, 35, 1–3. [Google Scholar]
57. Chu, Q.; Ren, G.; Duan, X.; Li, L.; Zhu, K.; Wang, Z. Effect of Different Pretreatment Methods on Brownning and Drying Characteristics of Hot Air-Dried dlily Buds. Food Sci. 2023, 44, 81–88. [Google Scholar]
58. Ding, L.; Li, Y.; Wu, Y.; Li, T.; Geng, R.; Cao, J.; Zhang, W.; Tan, X. Plant Disease Resistance-Related Signaling Pathways: Recent Progress and Future Prospects. Int. J. Mol. Sci. 2022, 23, 16200. [Google Scholar] [CrossRef]

Chen Y, Pang L, Bai J, Yu W, Zhu Y, George MS, Lv Z, Lu G. Exploring the Impact of Citric Acid on Mitigating Sweet Potato Soft Rot and Enhancing Postharvest Quality. Agriculture. 2025; 15(2):215. https://doi.org/10.3390/agriculture15020215
Перевод статьи «Exploring the Impact of Citric Acid on Mitigating Sweet Potato Soft Rot and Enhancing Postharvest Quality» авторов Chen Y, Pang L, Bai J, Yu W, Zhu Y, George MS, Lv Z, Lu G., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык

















































































































Комментарии (0)