Биорециклинг скорлупы фундука для улучшения антиоксидантных свойств плодовых тел Lentinus tigrinus
Lentinus tigrinus — это гриб, разлагающий древесину, известный своей питательной, кулинарной и лекарственной ценностью. Он содержит биологически активные соединения, такие как полифенолы, терпены и флавоноиды, которые обладают антиоксидантным, противомикробным и противовоспалительным действием. Эти природные антиоксиданты все чаще признаются за их потенциал в предотвращении окислительного повреждения, связанного со старением и хроническими заболеваниями.
Аннотация
В данном исследовании изучается антиоксидантная активность гидроалкогольных экстрактов, полученных из плодовых тел L. tigrinus, выращенных на скорлупе фундука (Lt1_HS) — отходах, богатых фенольными соединениями, и плодовых тел, выращенных на опилках (Lt1_S). Были проведены биоактивностные тесты, включая анализы DPPH, TBARS, MTT и DCFH-DA, чтобы оценить эффективность гидроалкогольных экстрактов. Результаты показали, что все экстракты содержали различные биологически активные соединения, в основном полифенолы. Примечательно, что кофеоилхинные кислоты, присутствующие в HS и Lt1_HS, связаны с антипероксидантным эффектом. Биологический анализ показал, что экстракт Lt1_HS обладает более высокой антипероксидантной активностью (IC50 0,77 ± 0,01 мг/мл) по сравнению с Lt1_S (IC50 1,36 ± 0,01 мг/мл) и снижает накопление активных форм кислорода в клетках HaCaT на 80%. Однако конкретные биологически активные соединения, ответственные за эти антиоксидантные эффекты, все еще не ясны, и дальнейший анализ будет проведен. Кроме того, это исследование способствует рециклингу скорлупы фундука в качестве ценного субстрата для культивирования грибов, поддерживая устойчивое управление отходами.
Конечно, вот перевод продолжения статьи, выполненный в соответствии со всеми вашими требованиями.
1. Введение
Lentinus tigrinus (Bull.) Fr. — это гриб, разлагающий древесину (wood-decay fungus, WDF), принадлежащий к группе грибов белой гнили, который растет как сапрофитный организм, поодиночке или группами, на бревнах и пнях, в основном видов Salix и Populus [1]. Он известен в литературе своей ролью в разложении лигноцеллюлозного материала благодаря продукции лакказ, пероксидаз и других лигноцеллюлозных ферментов [2,3]. L. tigrinus широко распространен от тропических до умеренных и бореальных регионов; он отмечен в 17 странах, принадлежащих к разным регионам, включая Центральную Азию, Южную Америку, Европу и Африку, обычно плодонося с мая по октябрь [2,4]. Морфологически L. tigrinus узнаваем по своему твердому, плотному, округлому шляпкоподобному образованию (pileus), украшенному чешуйками от светло-коричневого до черного цвета и зазубренными краями; ножка (stipe) имеет белый, чешуйчатый вид. Гименофор представлен пластинками (lamellae) с оттенками белого или бело-кремового цвета, а базидиоспоры варьируются от эллипсоидальных до цилиндрических [5]. Этот WDF является съедобным грибом с приятным ароматом и уникальным вкусом, подходящим для изысканных блюд; он также известен своими лекарственными свойствами [6,7,8]. Исследование Dulay и др. [9] подробно описало пищевой состав. Шляпкоподобное образование L. tigrinus содержало большее количество белка (25,9%), жира (2,1%), золы (7,4%) и имело более высокую энергетическую ценность (142,1 ккал/100 г), чем ножка, в то время как ножка содержала большее количество общих углеводов (67,7%), чем шляпкоподобное образование. Что касается полисахаридов, плодовые тела L. tigrinus содержали растворимые полисахариды в диапазоне от 30,1% (в шляпкоподобном образовании) до 38,3% (в ножке) [9,10]. Дополнительно, L. tigrinus содержит различные биологически активные соединения, такие как полифенолы, терпены, флавоноиды и поликетиды [11,12]. Все эти молекулы проявляют различные биологические эффекты, такие как антиоксидантное, противомикробное, противовоспалительное, противоожирительное и гипогликемическое действие [13,14,15]. Более того, эти соединения помогают предотвращать окислительный стресс — патологический процесс у человека, связанный в основном со старением и хроническими заболеваниями [16]. Окислительный стресс вызывается дисбалансом между образованием активных форм кислорода (АФК, ROS), таких как супероксид-анион (O₂⁻), перекись водорода (H₂O₂) и гидроксильные радикалы (OH), и несостоятельностью антиоксидантной защиты. АФК — это высоко реактивные молекулы, обычно нейтрализуемые ферментами, защищающими здоровье клеток [17]. Однако, когда этот естественный защитный механизм нарушается, уровень АФК возрастает, что может вызывать значительное клеточное повреждение, способствуя развитию хронических заболеваний, таких как рак, сердечно-сосудистые заболевания, нейродегенеративные расстройства и болезни, связанные со старением [18,19]. По этой причине в настоящее время ключевое значение имеет поиск биологически активных соединений, экстрагированных из натуральных продуктов, которые могут ингибировать развитие АФК. Биологические тесты, исследующие антиоксидантную активность, имеют ключевое значение для понимания того, как различные вещества могут защищать клетки и ткани от окислительного повреждения. Сегодня множество тестов может быть использовано для оценки антиоксидантной эффективности экстракта; среди них широко используются DPPH (1,1-дифенил-2-пикрилгидразил) и DCFH-DA (2′,7′-дихлордигидрофлуоресцеин диацетат) [20,21,22]. Необходимо подчеркнуть, что тестирование на антиоксидантную активность помогает в разработке функциональных продуктов питания и нутрицевтиков, предназначенных для улучшения здоровья и благополучия, поддержки профилактического здравоохранения и направляет разработку терапевтических агентов, направленных на снижение окислительного повреждения в организме [23]. Продуктами, богатыми антиоксидантными соединениями, являются фундук, в основном богатый полифенолами, и грибы, такие как Hericium erinaceus, которые богаты антиоксидантными соединениями, такими как флавоноиды, витамин Е и эрготионеин [24]. Известно, что полифенолы нейтрализуют АФК и свободные радикалы, отдавая электрон или атом водорода. Дополнительно к их антиоксидантной и радикальной поглощающей способности, полифенолы также известны как хелаторы металлов [25]. Исследователи все больше сосредотачиваются на лекарственных грибах, чтобы изучить их потенциальные профилактические эффекты против окислительного стресса, а также исследовать новые возможности для пищевых добавок и нутрицевтических продуктов [26,27]. Дополнительно, отходы представляют собой перспективный, но недостаточно используемый источник биологически активных молекул. Например, скорлупа фундука (HS) содержит высокую концентрацию фенольных соединений и других биологически активных молекул [28,29]. К сожалению, из-за высокого содержания лигнина эти отходы не перерабатываются, а используются как топливо или био-пеллеты [30,31]. Таким образом, скорлупу фундука не следует рассматривать только как топливо; это ценный ресурс, который также может быть использован в качестве субстрата для выращивания грибов [32].
Данное исследование было направлено на оценку того, является ли антиоксидантная активность плодовых тел L. tigrinus, выращенных на скорлупе фундука, лучше, чем у выращенных на опилках. Для сравнения гидроалкогольных экстрактов плодовых тел были проведены тесты на биоактивность и биологические анализы. Дополнительно, этот проект был направлен на идентификацию молекул, ответственных за антиоксидантные эффекты, с фокусом на фенольные соединения. В заключение, это исследование усиливает потенциал L. tigrinus, выращенного на скорлупе фундука, как инновационного пищевого продукта, полезного для снижения окислительного стресса, и в то же время подчеркивает хорошую практику рециклинга этих трудноразлагаемых отходов.
2. Материалы и методы
2.1. Штамм Lentinus tigrinus
Плодовое тело L. tigrinus было собрано в Сан-Дженезио-эд-Унити (Павия, Италия), и мицелий был выделен в чистую культуру в чашках Петри, содержащих 2% масс./об. агар с солодовым экстрактом (MEA). Штамм был идентифицирован молекулярным анализом. Общая геномная ДНК была экстрагирована из лиофилизированного мицелия с использованием набора Nucleospin Plant II (Macherey-Nagel, Дюрен, Германия). Пара праймеров ITS1-ITS4 была использована для амплификации области внутреннего транскрибируемого спейсера (ITS) с помощью полимеразной цепной реакции (ПЦР), как сообщалось Girometta и др. [33]. Штамм (Lt1) хранится в Исследовательной коллекции культур грибов (MicUNIPV) на кафедре наук о Земле и окружающей среде (Университет Павии, Павия, Италия).
2.2. Выращивание L. tigrinus на опилках (S) и на скорлупе фундука (HS)
Протокол, использованный для выращивания плодовых тел L. tigrinus как на опилках (S), так и на скорлупе фундука (HS), следовал руководствам, предложенным Lechner и Albertò [7] и Stamets [34]. Производство посевного мицелия (spawn) происходило в инокулированной банке (SacO2, Дейнзе, Бельгия), содержащей 150 г стерилизованного проса. Стерильные порции мицелия Lt1, выращенного на MEA, переносили в банку, содержащую зерно, и добавляли 10 мл дистиллированной воды. Банку закрывали фильтрующей крышкой и хранили при 25 °C в темноте до полной колонизации проса (около 15 дней). Выращивание плодовых тел Lt1 проводилось параллельно на опилках (субстрат, наиболее похожий на естественные условия) и на скорлупе фундука (чтобы усилить использование этих трудноразлагаемых сельскохозяйственных отходов). Каждый субстрат помещали в автоклавируемые прозрачные фильтровальные мешки из полипропилена (ПП, 20 × 30, SacO2, Дейнзе, Бельгия) и стерилизовали; этот процесс повторяли дважды. Впоследствии мешки инокулировали 10% посевного мицелия, герметизировали и хранили при 25 °C в темноте до полной колонизации субстрата. Через 1 месяц субстрат был полностью колонизирован мицелием, и мешки перемещали в климатическую камеру при 19–24 °C, влажности 85% и 8 часах освещения. Собранные плодовые тела взвешивали в свежем виде, а затем подвергали медленной сушке при низкой температуре (T = 35 °C). После сушки плодовые тела снова взвешивали и хранили в морозильной камере для дальнейшего анализа.
Субстрат S состоял из 75% дубовых опилок, 10% пшеничной муки, 10% овсяных хлопьев и, наконец, 5% CaCO₃ (Таблица 1). В каждый мешок добавляли 150 г растительного субстрата и 150 мл дистиллированной воды, что давало конечный вес 300 г. Всего было подготовлено 10 фильтровальных мешков. Плодовое тело L. tigrinus, выращенное на опилках, обозначали как Lt1_S.

Таблица 1. Состав двух субстратов для выращивания, опилок и скорлупы фундука, для культивирования плодовых тел L. tigrinus.
Скорлупу фундука предоставила ферма «Le Nocciole di Greta», расположенная в Баскапе, Павия. Изначально была проведена тщательная отбор скорлупы, и впоследствии ее измельчили с помощью щековой дробилки до получения «крупного» размера зерна. Субстрат, использованный для выращивания Lt1, состоял из 100 г сухого веса скорлупы фундука (Таблица 1). Всего было подготовлено 30 фильтровальных мешков. Собранные плодовые тела взвешивали в свежем виде, а затем подвергали медленной сушке при низкой температуре (T = 35 °C). После сушки плодовые тела снова взвешивали и хранили в морозильной камере для дальнейшего анализа. Плодовое тело L. tigrinus, выращенное на скорлупе фундука, обозначали как Lt1_HS.
В соответствии со многими авторами [35,36,37], продуктивность плодовых тел оценивали, вычисляя биологическую эффективность (BE):
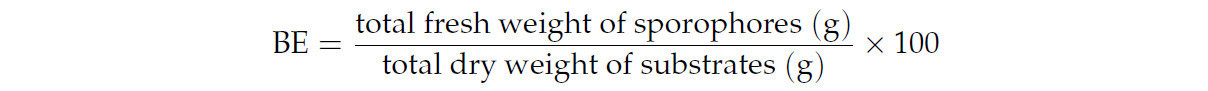
2.3. Биологическая активность
2.3.1. Приготовление гидроалкогольных экстрактов
Экстракты из плодовых тел L. tigrinus получали с помощью процесса динамической мацерации. Вкратце, 1,5 г высушенного образца взвешивали и экстрагировали EtOH/H₂O (80:20, об./об., 30 мл) при магнитном перемешивании и оставляли для мацерации на один час при комнатной температуре на мешалке. После этого экстракт фильтровали через бумажный фильтр (Whatman № 4, Merck KGaA, Дармштадт, Германия), осадок собирали, и тот же процесс повторяли для повторной экстракции. Жидкую часть фильтровали, а осадок удаляли, и экстракт концентрировали с использованием роторного вакуум-испарителя (≤40 °C, 100 об/мин, Büchi, R-210, Флавиль, Швейцария) для обеспечения полного испарения этанола. Водную фазу затем лиофилизировали (FeeeZone 4.5, Labconco, Канзас-Сити, Миссури, США).
2.3.2. Анализ поглощения радикала DPPH• (1,1-Дифенил-2-пикрилгидразил)
Анализ DPPH проводили в соответствии с методикой, описанной Souilem и др. [38]. Эту методику выполняли с использованием ридера для микропланшетов ELX800 (Bio-Tek, Санта-Клара, Калифорния, США). Реакционная смесь в каждой из 48 лунок состояла из 30 мкл последовательных разведений из исходного раствора экстрактов (20 мг/мл) и 270 мкл метанольного раствора радикалов DPPH (6 × 10⁻⁵ моль/л). Смесь оставляли на 60 мин в темноте для протекания реакции. Восстановление радикала DPPH определяли путем измерения поглощения при длине волны 515 нм. Активность по поглощению радикалов (RSA) рассчитывали в процентах обесцвечивания DPPH с использованием следующего уравнения: % RSA = [(A_DPPH − A_S)/A_DPPH] × 100, где A_S — это поглощение раствора, когда добавлен экстракт образца на определенном уровне, а A_DPPH — поглощение раствора DPPH (контроль). Результаты выражали в значениях IC₅₀ (мг/мл). Таким образом можно было рассчитать концентрацию экстракта, необходимую для поглощения свободного радикала и достижения антиоксидантной активности 50% экстракта. Аскорбиновую кислоту (Farmacia Fapa, Павия, Италия) использовали в качестве положительного контроля в диапазоне концентраций от 500 мкМ до 1 мкМ.
2.3.3. Анализ TBARS (Реактивные вещества тиобарбитуровой кислоты)
Использованный протокол для анализа был таким же, как описано в [38]. Гидроэтановый экстракт гриба повторно растворяли в воде и подвергали разведениям от 20 мг/мл до 0,625 мг/мл. Мозги свиней получали от официально забитых животных, рассекали и гомогенизировали с помощью Polytron в охлажденном льдом Tris-HCl буфере (20 мМ, pH 7,4) для получения раствора 1:2 масс./об., который центрифугировали при 3000× g в течение 10 мин. Аликвоту (100 мкл) супернатанта инкубировали с различными концентрациями растворов образцов (200 мкл) в присутствии FeSO₄ (10 мМ; 100 мкл) и аскорбиновой кислоты (0,1 мМ; 100 мкл) при 37,5 °C в течение 1 ч. Реакцию останавливали добавлением трихлоруксусной кислоты (28% масс./об., 500 мкл), затем тиобарбитуровой кислоты (TBA, 2%, масс./об., 380 мкл), и смесь затем инкубировали при 80 °C в течение 20 мин. Затем образцы центрифугировали при 3000× g в течение 10 мин. Интенсивность цвета комплекса малонового диальдегида (MDA)–TBA в супернатанте измеряли по его поглощению при 532 нм. Коэффициент ингибирования (%) рассчитывали по следующей формуле: коэффициент ингибирования (%) = [(A − B)/A] × 100%, где A и B — это поглощение контрольного и опытного растворов соответственно. Результаты выражали в значениях IC₅₀ (мг/мл). Таким образом можно было рассчитать концентрацию экстракта, необходимую для сохранения 50% перекисного окисления липидов. Тролокс (Sigma Chemical Co., Сент-Луис, Миссури, США) использовали в качестве положительного контроля.
2.4. Биологический анализ in vitro
2.4.1. Культура клеток — HaCaT
Клеточная линия HaCaT — это спонтанно иммортализованная линия человеческих кератиноцитов, приобретенная у Cell Line Service (Эппельхайм, Германия). Клетки культивировали в среде Игла, модифицированной Дульбекко (DMEM, Cls), с добавлением 10% эмбриональной бычьей сыворотки (FBS, Gibco — Life Technologies Europe BV, Блейсвейк, Нидерланды) и 1% антибиотиков пенициллин-стрептомицин (Gibco — Life Technologies Europe BV, Блейсвейк, Нидерланды). Клетки HaCaT выращивали в инкубаторе при 37 °C во влажной атмосфере с 5% CO₂. Среду меняли каждые 2–3 дня, и после достижения ~90% конфлюэнтности клетки отделяли с помощью TrypLE Express (Gibco — Life Technologies Europe BV, Блейсвейк, Нидерланды) и ресуспендировали в свежей среде.
2.4.2. Определение жизнеспособности клеток с использованием анализа MTT
Использованный протокол для анализа был таким же, как сообщалось Brandalise и др. [39].
Жизнеспособность клеток человеческой кожи (HaCaT) определяли с использованием анализа MTT [3-(4,5-диметилтиазол-2-ил)-2,5-дифенилтетразолий бромид] через 24–48 ч.
Вкратце, клетки культивировали в колбе 25 см², а затем высевали в 96-луночный планшет с плотностью 10 000 клеток на лунку (0,2 мл на лунку). Планшеты инкубировали при 37 °C в течение 24 ч во влажной атмосфере с 5% CO₂. На следующий день культуральную среду заменяли на свежую среду, дополненную различными концентрациями экстракта (в диапазоне от 0,0125 до 1 мг/мл). Для контрольного условия клетки инкубировали только с культуральной средой. После 24 ч воздействия раствор MTT (20 мкл на лунку) добавляли в темноте, и планшеты subsequently инкубировали примерно три часа при 37 °C. Затем кристаллы формазана растворяли с помощью DMSO. Далее quantification проводили, измеряя поглощение образцов при 550 нм с использованием ридера для микропланшетов ELx808TM Absorbance Microplate Reader (Bio-Tek Instruments, Inc., Уинуски, Вермонт, США). Те же шаги повторяли после 48 ч обработки. Данные выражали в процентах жизнеспособности клеток, и IC₂₅ рассчитывали после 24 и 48 ч обработки. Таким образом можно было рассчитать концентрацию экстракта, необходимую для ингибирования 25% жизнеспособности клеток.
Коэффициент ингибирования (%) рассчитывали по следующей формуле: коэффициент ингибирования (%) = [(A − B)/A] × 100%, где A и B — это поглощение контрольного и опытного растворов соответственно. Результаты выражали в значениях IC₂₅ (мг/мл). Таким образом можно было рассчитать концентрацию экстракта, которая ингибирует 25% жизнеспособности клеток.
2.4.3. Аналис с трипановым синим
Использованный протокол для анализа был таким же, как сообщалось Giammello и др. [40]. Для анализа с трипановым синем 250 000 клеток HaCaT высевали в каждую лунку 6-луночного планшета и инкубировали overnight для прикрепления. Затем клетки обрабатывали концентрациями IC₂₅ экстрактов, предварительно определенными с помощью анализа MTT. После 24 ч обработки клетки промывали 1× фосфатно-солевым буфером (PBS) и отделяли с использованием фермента TrypLE™ Select (1×) (Gibco) (Life Technologies Europe BV, Блейсвейк, Нидерланды) в течение 10 мин.
Клеточную суспензию центрифугировали при 800 об/мин в течение 7 мин, супернатант осторожно удаляли, и клеточный осадок ресуспендировали в 1 мл свежей среды. Для оценки жизнеспособности клеток 10 мкл ресуспендированных клеток смешивали с 10 мкл 0,4% раствора трипанового синего. Смесь загружали в камеру автоматического счетчика клеток Countess™ 3 (Thermo Fisher Scientifics Inc., Life Technologies Corporation, Вашингтон, округ Колумбия, США) для подсчета клеток. Жизнеспособные (неокрашенные) и нежизнеспособные (окрашенные) клетки количественно определяли, и рассчитывали процент жизнеспособных клеток.
2.4.4. Аналис DCFH–DA (2′,7′-Дихлордигидрофлуоресцеин диацетат)
Использованный протокол для анализа был таким же, как сообщалось Kim и Xue [41]. Флуорогенный краситель DCFH-DA использовали для оценки способности экстрактов Lt1_S и Lt1_HS снижать внутриклеточные уровни активных форм кислорода в клетках HaCaT. После пассивной диффузии в клетки DCFH-DA деацетилируется внутриклеточными эстеразами с образованием нефлуоресцирующего соединения, которое впоследствии окисляется АФК до высокофлуоресцентного 2′,7′-дихлорфлуоресцеина (DCF), который можно количественно определить с помощью флуоресцентного детектирования. Всего 150 000 клеток HaCaT высевали на покровные стекла, помещенные в 6-луночные планшеты, инкубировали overnight, при 37 °C с 5% CO₂, чтобы позволить клеткам прикрепиться. После этого клетки обрабатывали концентрациями IC₂₅ экстрактов, рассчитанными на 24 ч.
На следующий день были установлены два экспериментальных условия: базовое условие и условие окислительного стресса. Для базового условия клетки инкубировали с 10 мкМ DCFH-DA, разведенным в среде, в течение 30 мин при 37 °C, в темноте. Для условия окислительного стресса клетки сначала подвергали воздействию 500 мкМ перекиси водорода (H₂O₂) в течение 30 мин для индукции окислительного стресса. После обработки H₂O₂ клетки промывали PBS, а затем инкубировали с 10 мкМ DCFH-DA, как описано выше. После инкубации с DCFH-DA в течение 30 мин в обоих условиях (базовом и индуцированном H₂O₂) клетки дважды промывали 1× PBS для удаления избытка DCFH-DA. Интенсивность флуоресценции, которая коррелирует с продукцией АФК, измеряли с использованием оптического микроскопа Olympus BX51 (Olympus Italia S.r.l., Сограте, Италия), оснащенного ртутной лампой на 100 Вт, при длине волны возбуждения 485 нм и длине волны излучения 530 нм. Изображения записывали с помощью камеры Olympus MagnaFire, и результаты обрабатывали с помощью программного обеспечения Olympus Cell F, версия 3.1. В обоих условиях интенсивность флуоресценции коррелирует с продукцией АФК, и антиоксидантные эффекты экстрактов Lt1_S и Lt1_HS оценивали путем сравнения флуоресценции.
2.5. Характеризация экстрактов: анализ фенольных соединений методом HPLC-DAD-(ESI)MS/MS
Экстракты анализировали с помощью прибора высокоэффективной жидкостной хроматографии (HPLC — Dionex UltiMate 3000 series, Thermo Fisher Scientific — Сан-Хосе, Калифорния, США), оборудованного диодно-матричным детектором (DAD) и соединенного последовательно с масс-спектрометром ионной ловушки (MS, LinearIon Trap LTQ XL, Thermo Fisher Scientific — Сан-Хосе, Калифорния, США). Соединения разделяли в колонке Spherisorb S3 ODS-2 C18 (3 мкм, 4,6 × 150 мм, Waters — Милфорд, Массачусетс, США), поддерживаемой при 35 °C, в градиенте 0,1% (об./об.) муравьиной кислоты в ultrapure воде (A) и ацетонитриле (B). УФ-Видимые спектры регистрировали в диапазоне от 180 до 700 нм, и хроматограммы обрабатывали при 280, 330 и 370 нм для различных классов фенольных соединений. Элюат HPLC анализировали с помощью тандемной масс-спектрометрии высокого разрешения, и соединения ионизировали с использованием электроспрейного ионного источника (ESI), работающего в отрицательном режиме. Полные масс-спектры MS и MS/MS регистрировали в диапазоне от 110 до 1800 отношения массы к заряду (m/z). Полное сканирование MS и сканирование MS n проводили, применяя энергию столкновения 35 (условные единицы). При необходимости использовали стратегию динамического исключения. Сбор и обработку данных проводили с помощью программного обеспечения Xcalibur® 4.3 (Thermo Fisher Scientific, Сан-Хосе, Калифорния, США). Для идентификации соединений интерпретировали порядок элюирования на колонке C 18 и характеристики УФ-Видимых и масс-спектров (молекулярный ион ([M-H]⁻) и фрагменты MS/MS) и сравнивали со стандартами, когда они были доступны, данными литературы и доступными библиотеками. Количественное определение выполняли с использованием 9-точечных калибровочных кривых аутентичных стандартов. Результаты по фенольным соединениям выражали в мг на г лиофилизированного экстракта (мг·г⁻¹, сухой вес, dw).
2.6. Статистический анализ
Статистический анализ выполняли с помощью программного обеспечения GraphPad Prism 10.2 (GraphPad Software Inc., Ла-Хойя, Калифорния, США). Данные представлены как среднее значение ± стандартная ошибка среднего (SEM). Для проверки статистически значимых различий использовали однофакторный дисперсионный анализ (ANOVA) с последующим тестом Бонферрони. Различия считаются статистически значимыми при p < 0,05 (), p < 0,01 (), p < 0,001 () и p < 0,0001 ().
3. Результаты
3.1. Продуктивность плодовых тел, выращенных на опилках и скорлупе фундука
Идентичность штамма L. tigrinus (Lt1) была подтверждена молекулярным анализом. Регистрационный номер последовательностей ITS этого итальянского штамма был депонирован в GenBank [42] как PQ136439. Плодовые тела культивировали на опилках (Lt1_S) или скорлупе фундука (Lt1_HS), как описано в разделе 2 (Рисунок 1). Плодовые тела L. tigrinus, выращенные на опилках (Lt1_S), были собраны (общий урожай свежего веса 621,8 г), что составило 58,81 г сухого веса, с биологической эффективностью (BE) 40,6% (Таблица 2). Плодовые тела L. tigrinus, выращенные на скорлупе фундука (Lt1_HS), были собраны (общий урожай свежего веса 432 г), что составило 34,82 г сухого веса, с BE 14,5% (Таблица 2). BE статистически различалась между двумя образцами (p-значение: 0,0001).

Рисунок 1. Выращивание плодовых тел L. tigrinus на опилках (Lt1_S) и скорлупе фундука (Lt1_HS). Фото A. Desiderio.
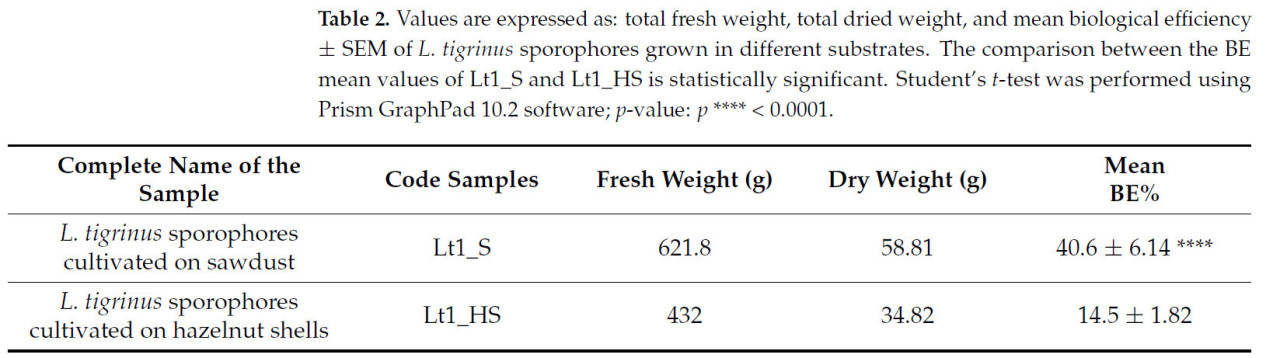
Таблица 2. Значения выражены как: общий свежий вес, общий сухой вес и средняя биологическая эффективность ± SEM плодовых тел L. tigrinus, выращенных на разных субстратах. Сравнение между средними значениями BE Lt1_S и Lt1_HS является статистически значимым. t-критерий Стьюдента был выполнен с использованием программного обеспечения Prism GraphPad 10.2; p-значение: p < 0,0001.
3.2. Схожий антиоксидантный, но различный антипероксидантный эффект гидроалкогольных экстрактов Lt1_S и Lt1_HS
Гидроалкогольные экстракты Lt1_S и Lt1_HS, а также скорлупы фундука (HS) были приготовлены, как описано (см. раздел 2). Антиоксидантный и антипероксидантный эффекты трех экстрактов оценивали с использованием анализов DPPH и TBARS соответственно.
Результаты, полученные в анализе DPPH, показали, что все экстракты проявляли антиоксидантный эффект. В частности, экстракт HS показал IC₅₀ 0,098 ± 0,011 мг/мл, что согласуется с ранее опубликованными данными Esposito и др. [30]. Гидроалкогольные экстракты плодовых тел, выращенных на любом из субстратов, проявляли схожую антиоксидантную активность, более низкую, чем образец HS (IC₅₀ 0,339 ± 0,009 мг/мл и 0,352 ± 0,001 мг/мл для Lt1-S и Lt1-HS соответственно; p < 0,0001 для обоих образцов по сравнению с HS) (Рисунок 2A). Разница между антиоксидантной активностью двух плодовых тел (Lt1_S и Lt1_HS) не была статистически значимой (Рисунок 2A). Этот результат свидетельствует о том, что два субстрата, опилки и фундук, оказывали схожий эффект на антиоксидантные свойства L. tigrinus в анализе DPPH.

Рисунок 2. Экстракты Lt1_S и Lt1_HS имеют схожие антиоксидантные свойства, но различающиеся антипероксидантные эффекты. Значения IC₅₀ (мг/мл), полученные с помощью DPPH (A) и TBARS (B) для трех экстрактов: (HS) скорлупа фундука, (Lt1_S) плодовое тело L. tigrinus, выращенное на опилках, и (Lt1_HS) плодовое тело L. tigrinus, выращенное на скорлупе фундука. Однофакторный ANOVA был выполнен с использованием программного обеспечения Prism GraphPad 10.2; p-значение: p < 0,0001; ns, не статистически значимо.
Напротив, анализ TBARS показал различные значения IC₅₀ среди трех образцов. Более высокая антипероксидантная активность присутствовала в экстракте HS (IC₅₀ 0,36 ± 0,04 мг/мл), но между двумя плодовыми телами экстракт Lt1_HS проявлял более высокую антипероксидантную активность (IC₅₀ 0,77 ± 0,01 мг/мл) по сравнению с экстрактом Lt1_S (IC₅₀ 1,36 ± 0,01 мг/мл) (Рисунок 2B). Экстракт HS статистически значительно отличается по сравнению с Lt1_S и Lt1_HS (Рисунок 2B), но, что интересно, экстракт Lt1_HS проявлял вдвое более высокую антипероксидантную активность по сравнению с Lt1_S. Из результатов однофакторного анализа ANOVA была выявлена статистически значимая разница p < 0,0001 между Lt1_S и Lt1_HS.
3.3. Жизнеспособность клеток под действием экстрактов Lt1_S и Lt1_HS
Чтобы оценить антиоксидантный эффект в биологической модели, мы изучили антиоксидантные эффекты в культуре клеток. В частности, поскольку клетки кожи являются хорошо зарекомендовавшей себя моделью для изучения окислительного стресса, была выбрана спонтанно иммортализованная линия человеческих кератиноцитов, линия клеток кожи (HaCaT). Сначала мы проверили подходящую дозу экстракта, которая не вызывала цитотоксичности в клетках HaCaT, используя анализ MTT. Клетки HaCaT подвергали воздействию в течение 24 ч различным концентрациям экстрактов Lt1_S и Lt1_HS, в диапазоне от 12,5 мкг/мл до 1 мг/мл, и оценивали жизнеспособность клеток (Рисунок 3). Все данные были статистически значимы, за исключением контроля по сравнению со следующими концентрациями: 1,25 × 10⁻² (Lt1_S и Lt1_HS) и 6,25 × 10⁻² Lt1_HS.
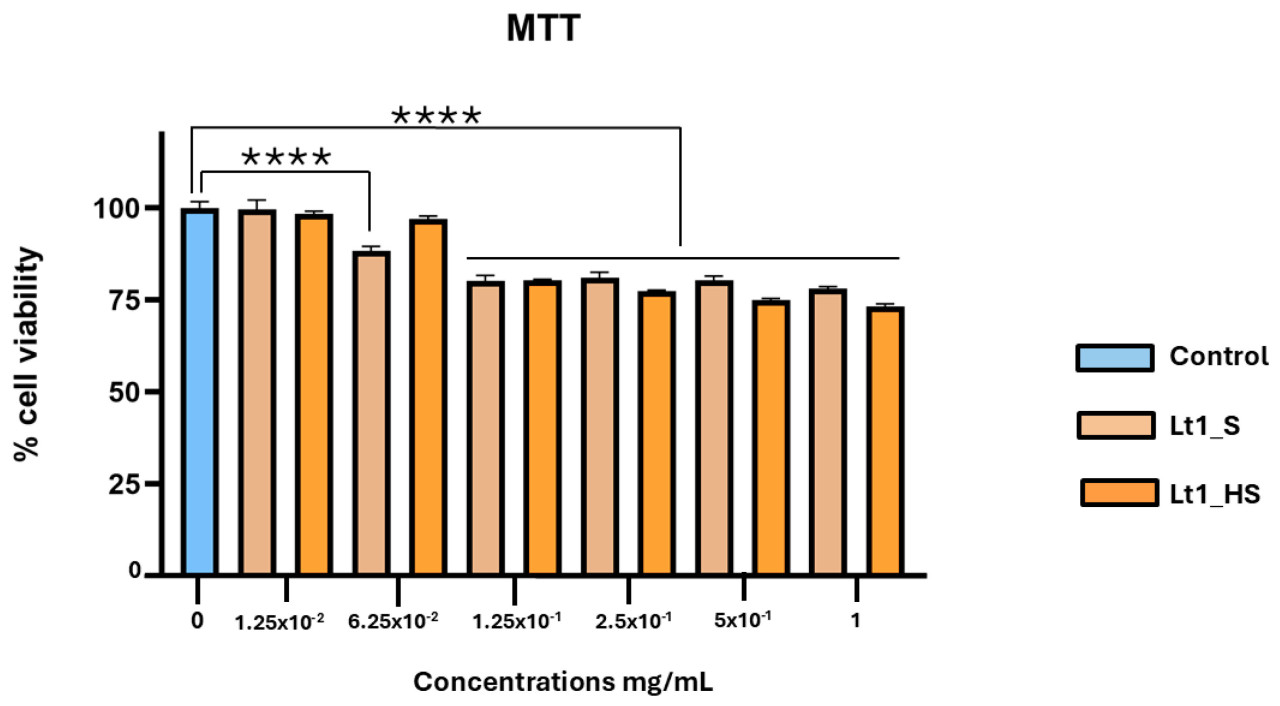
Рисунок 3. Жизнеспособность клеток HaCaT, подвергшихся воздействию различных концентраций экстракта Lt1_S (коричневый) и Lt1_HS (оранжевый) в течение 24 ч (среднее ± SEM, n = 4 повтора), по сравнению с контролем (синий). Однофакторный ANOVA был выполнен с использованием программного обеспечения Prism GraphPad 10.2; p-значение: p < 0,0001.
IC₂₅ был рассчитан для оценки концентрации, при которой экстракт ингибирует жизнеспособность клеток на 25% после 24 ч воздействия. Lt1_S не проявлял жизнеспособности клеток 25% при самой высокой протестированной концентрации (1 мг/мл), тогда как экстракт Lt1_HS ингибирует жизнеспособность клеток на 25% при концентрации 0,531 ± 0,015 мг/мл.
Чтобы подтвердить результаты анализа MTT, мы выполнили анализ с трипановым синим. Этот анализ позволяет определить процент живых клеток после специфической обработки. Основываясь на результатах MTT, протестировали концентрации IC₂₅ Lt1_S (1 мг/мл) и Lt1_HS (0,531 ± 0,01 мг/мл) после 24 ч воздействия. Результаты показали, что жизнеспособность клеток HaCaT после воздействия экстрактов Lt1_S и Lt1_HS не была статистически значимой и была равна 75% при протестированных концентрациях (Рисунок 4). Результаты для экстрактов Lt1_S и Lt1_HS были схожими, и не было разницы между двумя экстрактами.
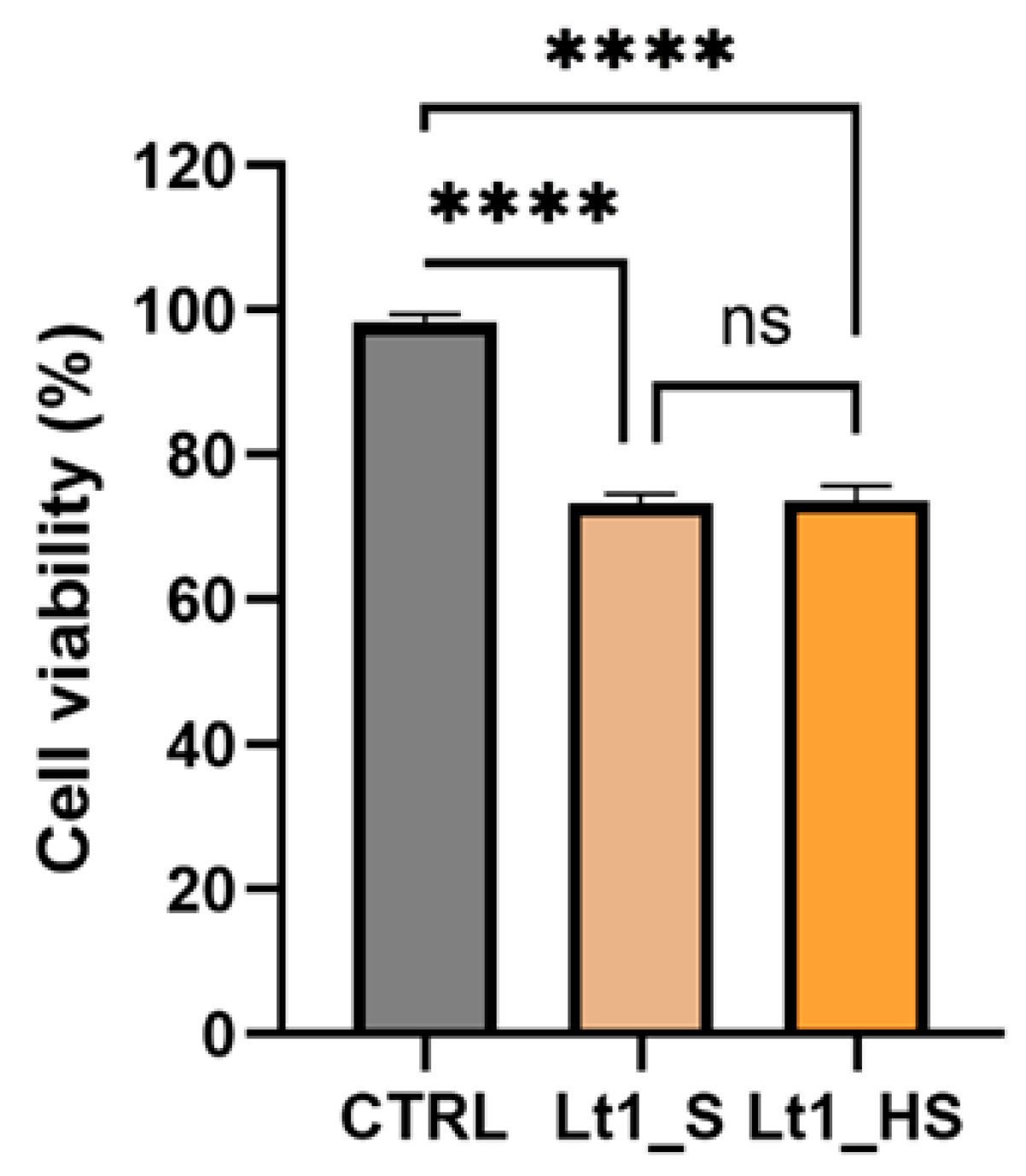
Рисунок 4. Проценты жизнеспособности клеток HaCaT после 24 ч обработки экстрактами плодовых тел Lt1_S и Lt1_HS. Два экстракта не показали статистически значимой разницы. Однофакторный ANOVA был выполнен с использованием программного обеспечения Prism GraphPad 10; p-значение: p < 0,0001; ns, не статистически значимо.
3.4. Скорлупа фундука обогатила антиоксидантную активность плодового тела L. tigrinus в клетках кожи
Анализ DCFH-DA измеряет активные формы кислорода (АФК, ROS) в клетках. DCFH-DA превращается в флуоресцентное соединение (DCF) при реакции с АФК, и интенсивность флуоресценции указывает на уровни окислительного стресса. Основываясь на результатах MTT и трипанового синего через 24 ч, концентрации IC₂₅ экстрактов Lt1_S и Lt1_HS были протестированы на клетках HaCaT. Клетки в контрольном условии (CTRL) демонстрировали базальный окислительный стресс без какого-либо воздействия.
Анализ DCFH-DA выявил, что экстракт Lt1_HS значительно ингибировал окислительный стресс в базовых условиях, достигну reduction 80% по сравнению с контролем. Напротив, экстракт Lt1_S снижал окислительный стресс примерно на 30% по сравнению с контролем. Два экстракта снижают окислительный стреск статистически значимым образом по сравнению с контролем (p < 0,0001). Между двумя экстрактами Lt1_HS показал значительное снижение по сравнению с Lt1_S (p < 0,0001), (Рисунок 5A). Более высокий антиоксидантный эффект Lt1_HS по сравнению с Lt1_S был подтвержден в условии индуцированного окислительного стресса. Клетки HaCaT подвергали воздействию перекиси водорода, которая удвоила продукцию АФК по сравнению с базовым условием. Следует отметить, что экстракт Lt1_HS снизил окислительный стресс со 200% до примерно 30%, достигнув значения окислительного стресса более низкого, чем контрольное условие. Для сравнения, экстракт Lt1_S снизил окислительный стресс до 100%, значения, аналогичного контрольному условию.
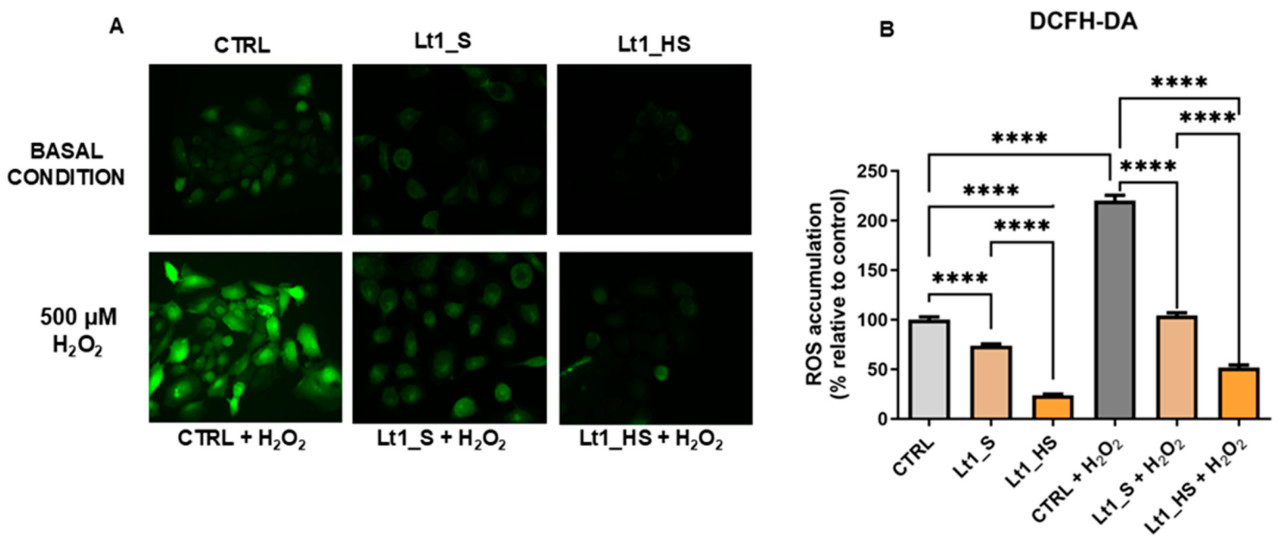
Рисунок 5. Различные антиоксидантные эффекты Lt1_S и Lt1_HS на клетках HaCaT. (A) Верхние панели показывают базовые условия. Нижние панели показывают условие индуцированного окислительного стресса. (B) Гистограмма суммирует данные в двух экспериментальных условиях. Однофакторный ANOVA был выполнен с использованием программного обеспечения Prism GraphPad 10.2; p-значение: p < 0,0001.
Полученные результаты указали, что экстракт Lt1_HS проявлял очень высокую антиоксидантную активность как в базовых, так и в условиях индуцированного окислительного стресса, с очень высокой статистически значимой разницей по сравнению с Lt1_S (Рисунок 5B). Следовательно, субстрат, состоящий из скорлупы фундука, обогатил антиоксидантную активность плодового тела L. tigrinus в клетках кожи.
3.5. Характеризация профилей фенольных соединений
Более высокая антиоксидантная активность Lt1_HS могла быть обусловлена присутствием фенольных соединений, которые содержатся в скорлупе фундука. Мы предполагаем, что полифенольные соединения могли быть поглощены плодовым телом L. tigrinus во время его роста. Богатство полифенолами в скорлупе фундука можно объяснить их естественной ролью в защитной системе растения. Полифенолы действуют как антиоксиданты и защитные агенты, оберегая ядро фундука от окислительного повреждения, микробных атак и стрессоров окружающей среды [43]. Чтобы проверить гипотезу о наличии фенольных соединений, были выполнены хроматограммы HPLC-DAD на гидроалкогольных экстрактах HS, Lt1_S и Lt1_HS (Рисунок 6). Идентифицированные молекулы были представлены (Таблица 3, Таблица 4 и Таблица 5) для каждого образца.
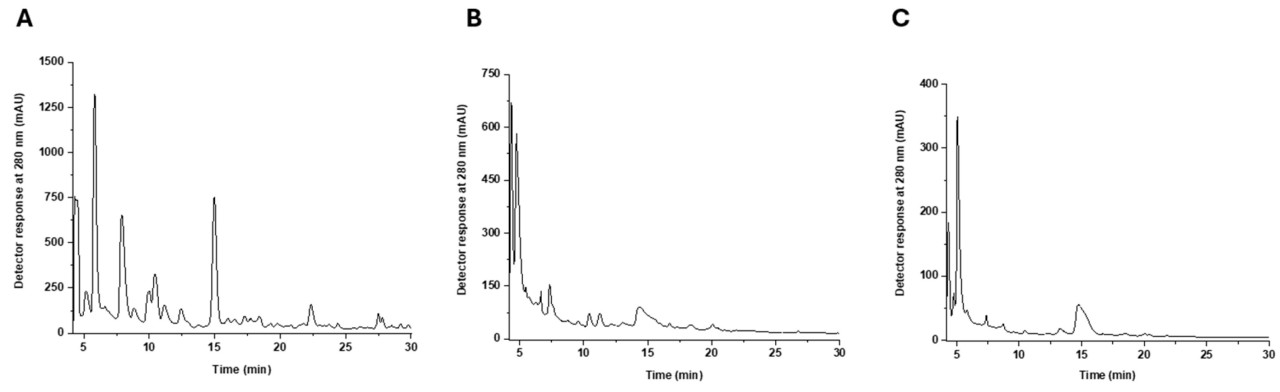
Рисунок 6. Хроматограммы, полученные с помощью HPLC-DAD для (A) HS, (B) Lt1_S, (C) Lt1_HS. Идентификация пиков приведена в Таблице 3, Таблице 4 и Таблице 5.
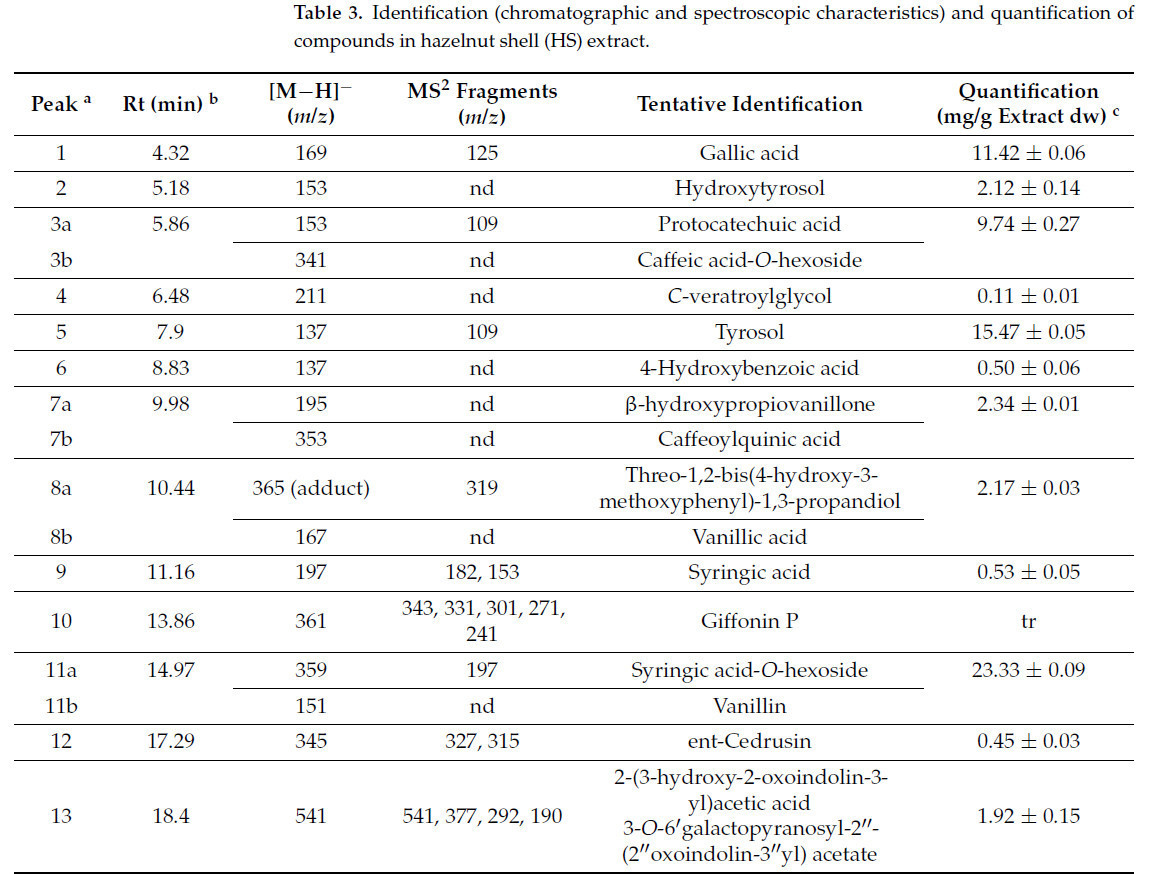
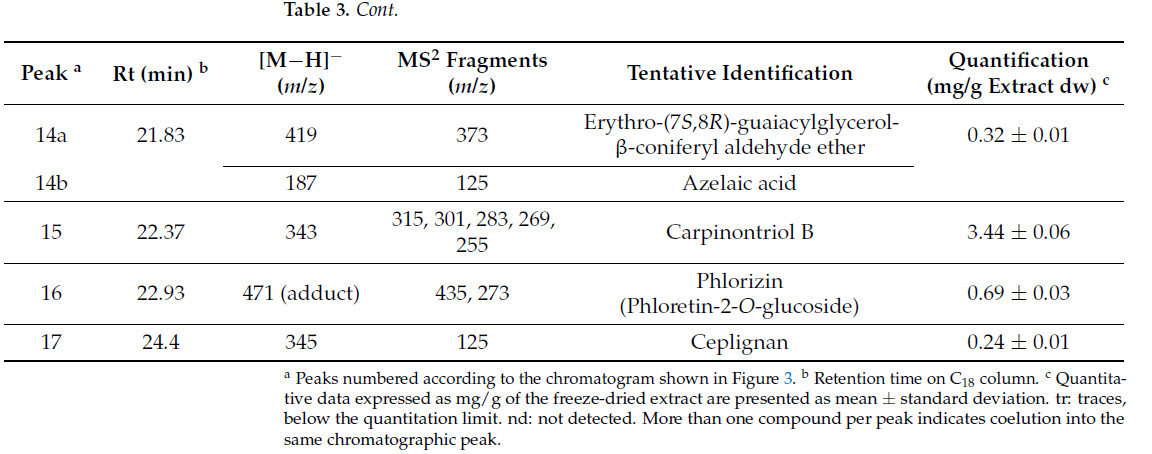
Таблица 3. Идентификация (хроматографические и спектроскопические характеристики) и количественное определение соединений в экстракте скорлупы фундука (HS).

Таблица 4. Идентификация (хроматографические и спектроскопические характеристики) и количественное определение соединений в экстракте L. tigrinus, выращенного на опилках (Lt1_S).
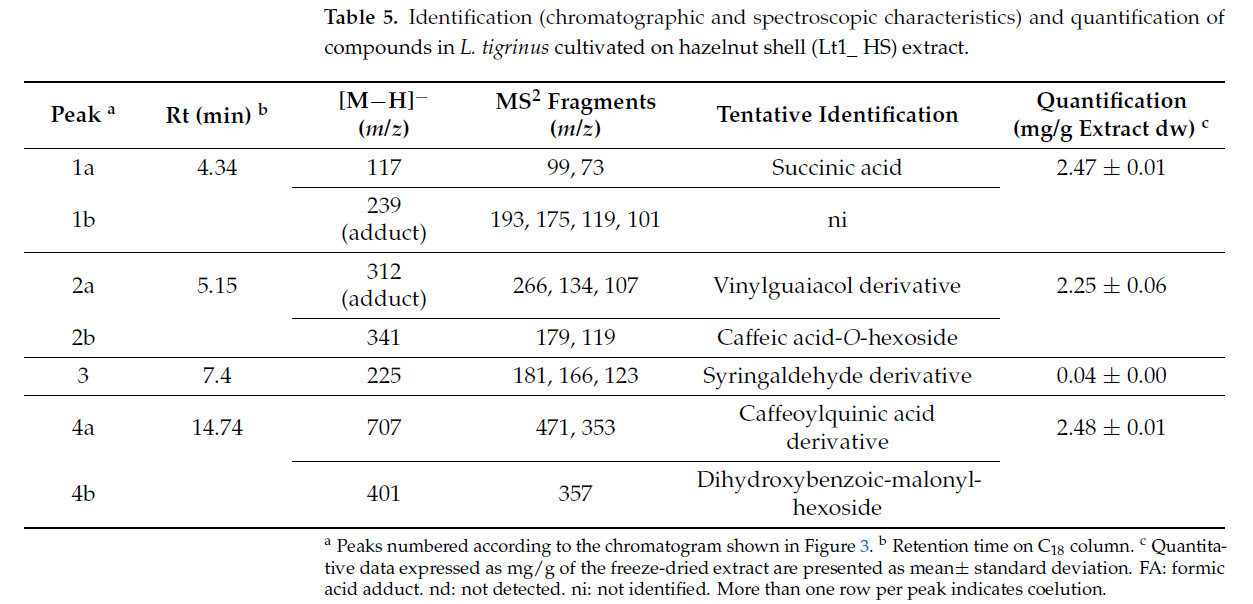
Таблица 5. Идентификация (хроматографические и спектроскопические характеристики) и количественное определение соединений в экстракте L. tigrinus, выращенного на скорлупе фундука (Lt1_ HS).
В экстракте HS присутствовало 17 пиков, и 22 биологически активных соединения были предварительно идентифицированы. Те, которые имели наивысшую концентрацию, были сиринговая кислота-O-гексозид (23,33 мг/г экстракта dw), тирозол (15,47 мг/г экстракта dw), галловая кислота (11,42 мг/г экстракта dw) и протокатеховая кислота (9,74 мг/г экстракта dw). Следует отметить, что большинство биологически активных соединений, идентифицированных в экстракте HS, являются полифенолами (Таблица 3).
В экстракте Lt1_S присутствовало всего шесть пиков, и было идентифицировано 17 биологически активных соединений. Ванилин был основным соединением, предварительно идентифицированным в образце (25,29 мг/г экстракта dw). Следует отметить, что всего 8 из 17 соединений были идентифицированы как фенольные соединения (Таблица 4). Дополнительно, три органические кислоты, яблочная, фумаровая и янтарная, также были идентифицированы.
В экстракте Lt1_HS можно было предварительно идентифицировать только шесть биологически активных соединений. Следует отметить, что пять молекул являются фенольными соединениями, помимо янтарной кислоты, которая является органической кислотой (Таблица 5). Наивысшие концентрации были обнаружены для янтарной кислоты и производного винилгваякола, которые были равны 2,47 ± 0,01 мг/г и 2,25 ± 0,06 мг/г экстракта dw соответственно.
Было проведено сравнение между тремя экстрактами для идентификации молекул, присутствующих во всех образцах (Таблица 6). Сравнение выявляет значительные различия между образцами HS и Lt1_S. Действительно, единственными общими молекулами являются кофейная кислота-O-гексозид и ванилин, причем последний особенно важен, поскольку он является продуктом деградации лигнина, как сообщается в литературе [44].
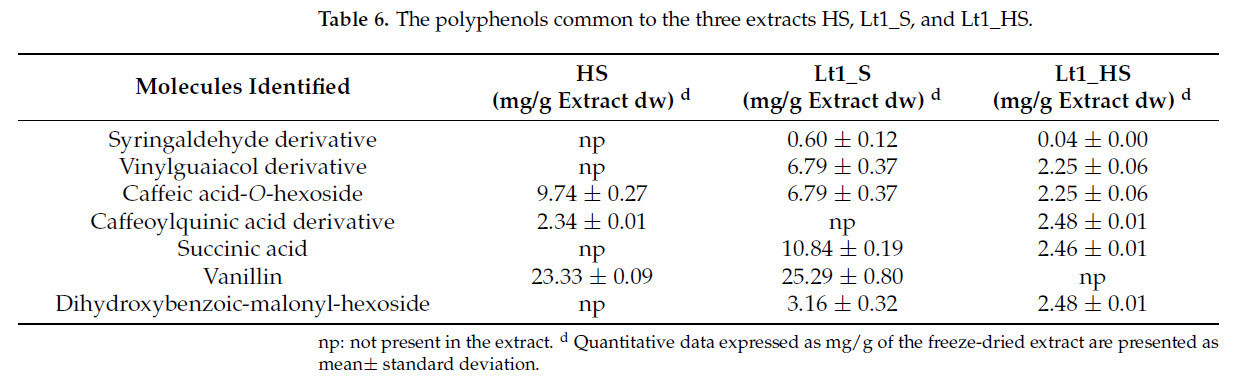
Таблица 6. Полифенолы, общие для трех экстрактов HS, Lt1_S и Lt1_HS.
Интересно, что единственными фенольными соединениями, которые присутствовали в гидроалкогольных экстрактах HS и Lt1_HS в схожих концентрациях, но которые отсутствовали в Lt1_S, были кофейоилхинная кислота и ее производное. Производные сиреневого альдегида и винилгваякола, дигидроксибензой-малонил-гексозид и янтарная кислота присутствовали как в Lt1_HS, так и в Lt1_S, но отсутствовали в экстракте HS, что предполагает, что эти молекулы не зависят от двух различных субстратов.
4. Обсуждение
L. tigrinus — это съедобный гриб белой гнили, способный расти на различных субстратах, таких как скорлупа фундука, которую трудно разлагать из-за высокого содержания лигнина [45,46,47]. L. tigrinus особенно известен своими биологически активными соединениями, такими как полифенолы, токоферол, флавоноиды и терпены [48,49]. Все эти биологически активные соединения обладают различными биологическими свойствами, и их антиоксидантная активность является наиболее известной и глубоко изученной [50,51]. Исследования сосредоточены на изучении натуральных продуктов, богатых антиоксидантными соединениями, в частности грибов, для разработки новых, инновационных и полезных для здоровья пищевых продуктов и внесения вклада в усилия по открытию новых лекарств [52,53,54,55]. Следовательно, благодаря своему потенциалу грибы могут найти применение в различных областях, включая нутрицевтики и космецевтики [56,57]. Этот новый подход согласуется с проблемой циркулярной экономики, которая усиливает переоценку отходов как нового ресурса.
Настоящая работа была направлена на использование скорлупы фундука, сельскохозяйственных отходов, которые трудно перерабатывать, в качестве субстрата для роста L. tigrinus, чтобы исследовать, как антиоксидантные молекулы, присутствующие в скорлупе фундука, могут быть использованы или метаболизированы грибом во время роста плодового тела. Поэтому мы исследовали антиоксидантную активность, выраженную гидроалкогольным экстрактом, полученным из плодовых тел, выращенных на скорлупе фундука, по сравнению с гидроалкогольным экстрактом, полученным из плодовых тел, выращенных на опилках. Выбор использовать скорлупу фундука в качестве субстрата для выращивания L. tigrinus основан на их высоком содержании полифенолов, доказывая, что скорлупа фундука является природным источником антиоксидантов и может усиливать рост грибов и производство ферментов [58]. Это первое исследование, посвященное росту L. tigrinus на скорлупе фундука.
Несмотря на то, что выращивание L. tigrinus на скорлупе фундука показало более низкую урожайность по сравнению с опилками, полученная биологическая эффективность сравнима с той, о которой сообщали Puliga и др. [32]. Первый значительный вывод заключается в том, что L. tigrinus может развиваться и расти на субстрате, полностью состоящем из скорлупы фундука, что указывает на его адаптивность и потенциал использования этого ресурса. L. tigrinus показал большой потенциал в качестве кандидата для выращивания на скорлупе фундука, что подтверждается литературой, подчеркивающей его гидролитическое действие на лигноцеллюлозный и трудноразлагаемый субстрат [59,60]. Рост видов грибов на скорлупе фундука представляет экологическую особенность, представляющую значительный интерес, потому что эти сельскохозяйственные отходы обычно считаются сложными для разложения [29]. Действительно, скорлупа фундука характеризуется высоким содержанием лигнина, сложного биополимера, который трудно разлагать [28], но грибы, разлагающие древесину, в частности белая гниль, действуют как мощные агенты разложения, производя ферменты, такие как лакказы и пероксидазы, которые разрушают лигнин [61,62]. Деградация лигнина высвобождает различные питательные вещества, такие как сахара, целлюлоза и органические соединения, особенно в присутствии субстратов, богатых биологически активными молекулами, таких как скорлупа фундука, создавая благоприятную среду для роста грибов [63]. Мицелий поглощает необходимые питательные вещества и биологически активные соединения, которые включают флавоноиды, большую группу фенолов [64,65]. В результате высокой доступности из-за распада лигнина можно предположить, что грибы обогащены биологически активными молекулами, такими как полифенолы, увеличивая их антиоксидантную активность [66,67]. Другая гипотеза предполагает, что увеличенная доступность этих высвобожденных молекул стимулирует грибной метаболизм, приводя к продукции вторичных метаболитов [68]. Таким образом, субстрат из отходов не только биодеградирует, но и производит высокоценную грибную биомассу с потенциальными антиоксидантными эффектами. Биологические анализы были выполнены для оценки антипероксидантных и антиоксидантных свойств [15] двух экстрактов, Lt1_S и Lt1_HS.
Анализ TBARS выявляет превосходный антипероксидантный эффект, выраженный экстрактом Lt1_HS по сравнению с Lt1_S. Различие в фенольном составе между Lt1_S и Lt1_HS может объяснять вариацию в антипероксидантной активности, поддерживая гипотезу о том, что субстрат HS обогащает плодовое тело биологически активными молекулами [66,69]. Недавние статьи Bouyahya и др. [70] и Deore и др. [71] указывают, что различные фенольные соединения являются ключевыми факторами, способствующими антипероксидантным эффектам, и результаты согласуются с нашими. В частности, из опубликованных результатов, кофейные производные, такие как кофейоилхинные кислоты, присутствующие как в экстрактах Lt1_HS, так и HS, как известно, ингибируют перекисное окисление липидов, и синергия между этими молекулами может усиливать антипероксидантную активность [72,73,74,75].
Анализ DCFH-DA, использованный для оценки внутриклеточных уровней АФК, демонстрирует, что Lt1_HS значительно снижал продукцию АФК как в базовых, так и в условиях индуцированного окислительного стреска в клетках кожи HaCaT, указывая на устойчивую антиоксидантную способность, вероятно, обусловленную синергетической активностью его фенольных компонентов [76]. Примечательно, что анализ DCFH-DA является надежным тестом, широко используемым для измерения окислительного стресса, и он также применялся в многочисленных исследованиях EFSA, что дополнительно поддерживает его достоверность в научных исследованиях [77,78]. Мы предполагаем, что обогащенный антиоксидантами экстракт Lt1_HS может не проявлять каких-либо потенциальных рисков потребления при обычной рекомендуемой дозе (1 г/сутки сушеного гриба). Это соображение основано на двух основных пунктах: (1) анализ MTT in vitro показал, что при протестированных концентрациях экстракты проявляли только снижение жизнеспособности клеток на 25%, что считается низким процентом цитотоксичности; (2) дополнительно, содержание полифенолов, измеренное, значительно ниже по сравнению с верхним пределом, указанным в DRA (Оценка диетического риска). Как сообщают Liu и др. [79], антиоксидантный эффект экстракта значительно усиливается синергетическим взаимодействием его молекул, в частности полифенолов. Эта синергия усиливает общую антиоксидантную способность экстракта, предполагая, что сложные смеси полифенолов и других биологически активных соединений обеспечивают более сильную, более комплексную защиту от окислительного стресса, чем отдельные компоненты.
Из химического анализа трех экстрактов следует, что состав экстракта HS соответствует литературе [80]. Вместо этого для экстракта Lt1_S, хотя библиографическое исследование полифенолов в L. tigrinus ограничено, наши находки поддерживают выводы, полученные Sanchez [15] и Karaman и др. [13]. В частности, Sanchez идентифицировал несколько полифенолов в экстракте плодового тела L. tigrinus, включая n-кумаровую кислоту, кофейную кислоту и протокатеховую кислоту, которые согласуются с нашими находками. Однако, хотя Sanchez не обнаружил ванилин, ванилиновая кислота присутствовала. С другой стороны, в результатах Karaman и др. [13] присутствовала только протокатеховая кислота. Интересно, что ванилиновая кислота и производные кофейной и n-кумароильной кислот, которые отсутствовали в их находках, явно присутствуют в нашем экстракте Lt1_S. Сравнение трех экстрактов, HS, Lt1_S и Lt1_HS, подчеркивает заметное различие в их составах, особенно в отношении ванилина, который присутствует в экстрактах Lt1_S и HS и отсутствует в экстракте Lt1_HS. Присутствие ванилина в экстракте Lt1_S согласуется с исследованиями Deore и др. [71], Zhu и др. [81] и Bogdan и др. [82], которые указывают ванилин как продукт деградации лигнина с различными биологическими свойствами, включая противомикробную и антиоксидантную активность. Известно, что производство ванилина сильно зависит от активности специфических лигнинолитических ферментов, таких как лакказы и пероксидазы, и условий окружающей среды процесса деградации, включая pH, доступность кислорода и состав субстрата [83,84]. Отсутствие ванилина в экстракте Lt1_HS приводит нас к гипотезе, что катаболизм лигнина мог бы выражаться различными ферментами, или что другие метаболические пути могут быть активированы. Это могло бы привести к различиям в вовлеченных грибных или микробных видах, вариациям в составе субстрата или ферментативной неэффективности [85].
Вместо этого, кофейоилхинная кислота и ее производное присутствуют в экстрактах Lt1_HS и HS. Литература показывает, что кофейоилхинная кислота и ее производные являются метаболитами, производимыми растениями, такими как Ipomoea batatas, Coffea arabica, Corylus avellana и другие виды растений [75,86]. Мы предполагаем, что плодовое тело, развитое на скорлупе фундука, могло поглощать молекулы непосредственно из самого субстрата, усиливая свою антиоксидантную биоактивность [87].
Хотя полученные результаты по антиоксидантной активности являются перспективными, остается неясным, какие полифенолы среди обнаруженных или другие молекулы, еще не обнаруженные, вовлечены в антиоксидантную активность. Однако ясно, что использование субстрата, такого как HS, увеличило антиоксидантную активность. Химические анализы указывают, что экстракт Lt1_HS содержит ограниченное число полифенолов. По этой причине будет выполнена глубокая химическая характеризация для исследования того, какие другие молекулы, такие как флавоноиды, терпены или токоферолы, могут вносить вклад в антиоксидантный эффект [88,89,90].
5. Выводы
В заключение, использование скорлупы фундука в качестве субстрата для роста L. tigrinus представляет перспективный подход для обогащения плодовых тел биологически активными соединениями, которые предлагают преимущества для здоровья, в частности в снижении окислительного стресса. Скорлупа фундука, сельскохозяйственные отходы, богатые лигнином и полифенолами, используются не только как источник питательных веществ для грибов, но и как катализаторы в стимулировании производства антиоксидантных молекул внутри грибов.
В частности, плодовые тела L. tigrinus, выращенные на скорлупе фундука, усилили антипероксидантную и антиоксидантную активность, указывая на перспективную стратегию для максимизации биоактивного потенциала этих сельскохозяйственных остатков. Этот вид субстрата из отходов обогащает грибы соединениями, известными своей способностью нейтрализовать свободные радикалы и защищать от клеточного окисления, способствуя общему поддержанию здоровья и потенциально снижая риск заболеваний, связанных с окислительным стрессом. Превращая скорлупу фундука в ценный субстрат, этот подход поддерживает устойчивое сельское хозяйство и предоставляет природный, экологически чистый источник антиоксидантов, демонстрируя потенциал биофортифицированных грибных продуктов как функциональных продуктов питания с полезными для здоровья свойствами. Это исследование было направлено на изучение того, может ли процесс деградации лигнина обогащать плодовые тела L. tigrinus. Дополнительно, важный акцент будет сделан на понимании вовлеченных молекулярных путей. В ближайшем будущем мы стремимся решить этот значительный вопрос.
Ссылки
1. Boccardo, F.; Traverso, M.; Vizzini, A.; Zotti, M. Funghi d’Italia; Zanichelli editore S.p.A.: Bologna, Italy, 2008; pp. 198–200. [Google Scholar]
2. Fabros, J.A.; Dulay, R.M.R.; De Leon, A.; Kalaw, S.; Reyes, R. Distribution, cultivation, nutritional composition, and bioactivities of Lentinus (Polyporaceae, Basidiomycetes): A review. Curr. Res. Environ. Appl. Mycol. 2022, 12, 170–219. [Google Scholar] [CrossRef]
3. Gafforov, Y.; Angelini, P.; Cusumano, G.; Venanzoni, R.; Angeles Flores, G.; Tomšovský, M.; Yarasheva, M.; Reseta, M.W.; Bussman, R.; Rapior, S. Lentinus arcularius (Batsch) Zmitr.; Lentinus brumalis (Pers.) Zmitr.; Lentinus ciliatus (Fr.) Zmitr.; Lentinus squarrosulus Mont.; Lentinus tigrinus (Bull.) Fr.-POLYPORACEAE. In Ethnobiology of Uzbekistan: Ethnomedicinal Knowledge of Mountain Communities; Springer International Publishing: Cham, Switzerland, 2023; pp. 1253–1270. [Google Scholar]
4. Karunarathna, S.C.; Yang, Z.L.; Zhao, R.L.; Vellinga, E.C.; Bahkali, A.; Chukeatirote, E.; Hyde, K.D. Three new species of Lentinus from northern Thailand. Mycol. Prog. 2011, 10, 389–398. [Google Scholar] [CrossRef][Green Version]
5. Dulay, R.M.; Cabrera, E.C.; Kalaw, S.P.; Reyese, R.G.; Hou, C.T. Nutritional requirement for mycelial growth of three Lentinus species from the Philippines. Biocatal. Agric. Biotechnol. 2020, 23, 101506. [Google Scholar] [CrossRef]
6. Dulay, R.M.R.; Cabrera, E.C.; Kalaw, S.P.; Reyes, R.G. Optimal growth conditions for basidiospore germination and morphogenesis of Philippine wild strain of Lentinus tigrinus (Bull.) Fr. Mycosphere 2012, 3, 926–933. [Google Scholar] [CrossRef]
7. Lechner, B.E.; Albertó, E. Optimal conditions for the fruit body production of natural occurring strains of Lentinus tigrinus. Bioresour. Technol. 2007, 98, 1866–1869. [Google Scholar] [CrossRef]
8. Peksen, A.; Yakupoglu, G.; Kibar, B. Influence of various sawdust-based substrates from different wood species and supplementary wheat bran on yield and fruiting body size of Lentinus tigrinus. Acta Hortic. 2009, 830, 319–326. [Google Scholar] [CrossRef]
9. Dulay, R.M.R.; Arenas, M.C.; Kalaw, S.P.; Reyes, R.G.; Cabrera, E.C. Proximate composition and functionality of the culinary-medicinal tiger sawgill mushroom, Lentinus tigrinus (Higher Basidiomycetes), from the Philippines. Int. J. Med. Mushrooms 2014, 16, 85–94. [Google Scholar] [CrossRef]
10. Ragasa, C.Y.; Tan, M.C.S.; De Castro, M.E.; Mariquit, M.; Oyong, G.G.; Shen, C.C. Sterols from Lentinus tigrinus. Pharmacogn. J. 2018, 10, 1079–1081. [Google Scholar] [CrossRef]
11. Reis, F.S.; Pereira, E.; Barros, L.; Sousa, M.J.; Martins, A.; Ferreira, I.C. Biomolecule profiles in inedible wild mushrooms with antioxidant value. Molecules 2011, 16, 4328–4338. [Google Scholar] [CrossRef]
12. Ferreira, I.C.F.R.; Baptista, P.; Boas, V.M.; Barros, L. Free-radical scavenging capacity and reducing power of wild edible mushrooms from northeast Portugal. Food Chem. 2007, 100, 1511–1516. [Google Scholar]
13. Karaman, M.; Jovin, E.; Malbaša, R.; Matavuly, M.; Popović, M. Medicinal and edible lignicolous fungi as natural sources of antioxidative and antibacterial agents. Phytother. Res. 2010, 24, 1473–1481. [Google Scholar] [CrossRef] [PubMed]
14. Barros, L.; Baptista, P.; Correia, D.M.; Casal, S.; Oliveira, B.; Ferreira, I.C. Fatty acid and sugar compositions, and nutritional value of five wild edible mushrooms from Northeast Portugal. Food Chem. 2007, 105, 140–145. [Google Scholar] [CrossRef]
15. Sánchez, C. Reactive oxygen species and antioxidant properties from mushrooms. Synth. Syst. Biotechnol. 2017, 2, 13–22. [Google Scholar] [CrossRef] [PubMed]
16. Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef]
17. Jabeen, S.; Noor, S. Oxidative stress and chronic kidney disease. In Fundamental Principles of Oxidative Stress in Metabolism and Reproduction; Academic Press: Cambridge, MA, USA, 2024; pp. 151–165. [Google Scholar]
18. Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative stress: Harms and benefits for human health. Oxidative Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
19. Kryston, T.B.; Georgiev, A.B.; Pissis, P.; Georgakilas, A.G. Role of oxidative stress and DNA damage in human carcinogenesis. Mutat. Res. 2011, 711, 193–201. [Google Scholar] [CrossRef]
20. Munteanu, I.G.; Apetrei, C. Analytical methods used in determining antioxidant activity: A review. Int. J. Mol. Sci. 2021, 22, 3380. [Google Scholar] [CrossRef]
21. López-Alarcón, C.; Denicola, A. Evaluating the antioxidant capacity of natural products: A review on chemical and cellular-based assays. Anal. Chim. Acta 2013, 763, 1–10. [Google Scholar] [CrossRef]
22. Collins, A.R. Assays for oxidative stress and antioxidant status: Applications to research into the biological effectiveness of polyphenols. Am. J. Clin. Nutr. 2005, 81, 261S–267S. [Google Scholar] [CrossRef]
23. Marra, A.; Manousakis, V.; Zervas, G.P.; Koutis, N.; Finos, M.A.; Adamantidi, T.; Panoutsopoulou, E.; Ofrydopoulou, A.; Tsoupras, A. Avocado and Its By-Products as Natural Sources of Valuable Anti-Inflammatory and Antioxidant Bioactives for Functional Foods and Cosmetics with Health-Promoting Properties. Appl. Sci. 2024, 14, 5978. [Google Scholar] [CrossRef]
24. Roda, E.; Ratto, D.; De Luca, F.; Desiderio, A.; Ramieri, M.; Goppa, L.; Savino, E.; Bottone, M.G.; Locatelli, C.A.; Rossi, P. Searching for a longevity food, we bump into Hericium erinaceus primordium rich in ergothioneine: The “longevity vitamin” improves locomotor performances during aging. Nutrients 2022, 14, 1177. [Google Scholar] [CrossRef] [PubMed]
25. Bertelli, A.; Biagi, M.; Corsini, M.; Baini, G.; Cappellucci, G.; Miraldi, E. Polyphenols: From theory to practice. Foods 2021, 10, 2595. [Google Scholar] [CrossRef] [PubMed]
26. Shaffique, S.; Kang, S.M.; Kim, A.Y.; Imran, M.; Aaqil Khan, M.; Lee, I.J. Current knowledge of medicinal mushrooms related to anti-oxidant properties. Sustainability 2021, 13, 7948. [Google Scholar] [CrossRef]
27. De Silva, D.D.; Rapior, S.; Hyde, K.D.; Bahkali, A. Medicinal mushrooms in prevention and control of diabetes mellitus. Fungal Divers. 2012, 56, 1–29. [Google Scholar] [CrossRef]
28. Di Michele, A.; Pagano, C.; Allegrini, A.; Blasi, F.; Cossignani, L.; Raimo, E.D.; Faieta, M.; Oliva, E.; Pittia, P.; Primavilla, S.; et al. Hazelnut shells as source of active ingredients: Extracts preparation and characterization. Molecules 2021, 26, 6607. [Google Scholar] [CrossRef]
29. Solís, A.; Rocha, S.; König, M.; Adam, R.; Garcés, H.O.; Candia, O.; Munoz, R.; Azòcar, L. Preliminary assessment of hazelnut shell biomass as a raw material for pellet production. Fuel 2023, 333, 126517. [Google Scholar] [CrossRef]
30. Esposito, T.; Sansone, F.; Franceschelli, S.; Del Gaudio, P.; Picerno, P.; Aquino, R.P.; Mencherini, T. Hazelnut (Corylus avellana L.) shells extract: Phenolic composition, antioxidant effect and cytotoxic activity on human cancer cell lines. Int. J. Mol. Sci. 2017, 18, 392. [Google Scholar] [CrossRef]
31. Ceylan, F.D.; Adrar, N.; Bolling, B.W.; Capanoglu, E. Valorisation of hazelnut by-products: Current applications and future potential. Biotechnol. Genet. Eng. Rev. 2023, 39, 586–621. [Google Scholar] [CrossRef]
32. Puliga, F.; Leonardi, P.; Minutella, F.; Zambonelli, A.; Francioso, O. Valorization of hazelnut shells as growing substrate for edible and medicinal mushrooms. Horticulturae 2022, 8, 214. [Google Scholar] [CrossRef]
33. Girometta, C.; Dondi, D.; Baiguera, R.M.; Bracco, F.; Branciforti, D.S.; Buratti, S.; Lazzaroni, S.; Savino, E. Characterization of mycelia from wood-decay species by TGA and IR spectroscopy. Cellulose 2020, 27, 6133–6148. [Google Scholar] [CrossRef]
34. Stamets, P.; Chilton, J.S. The Mushroom Cultivator. A Practical Guide to Growing Mushrooms at Home; Agaricon Press: Washington, DC, USA, 2009; 415p. [Google Scholar]
35. Pattanayak, S.; Das, S. Optimization of Biological efficiency and comparative analysis of nutraceutical properties in paddy straw and oyster mushrooms cultivation: A Review. J. Mycopathol. Res. 2022, 60, 13–21. [Google Scholar]
36. Yang, W.; Guo, F.; Wan, Z. Yield and size of oyster mushroom grown on rice/wheat straw basal substrate supplemented with cotton seed hull. Saudi J. Biol. Sci. 2013, 20, 333–338. [Google Scholar] [CrossRef] [PubMed]
37. Sharma, S.; Yadav, R.K.P.; Pokhrel, C.P. Growth and yield of oyster mushroom (Pleurotus ostreatus) on different substrates. J. New Biol. Rep. 2013, 2, 03–08. [Google Scholar]
38. Souilem, F.; Fernandes, Â.; Calhelha, R.C.; Barreira, J.C.; Barros, L.; Skhiri, F.; Martins, A.; Ferreira, I.C. Wild mushrooms and their mycelia as sources of bioactive compounds: Antioxidant, anti-inflammatory and cytotoxic properties. Food Chem. 2017, 230, 40–48. [Google Scholar] [CrossRef]
39. Brandalise, F.; Ramieri, M.; Pastorelli, E.; Priori, E.C.; Ratto, D.; Venuti, M.T.; Rosa, E.; Talpo, F.; Rossi, P. Role of Na+/Ca2+ exchanger (NCX) in glioblastoma cell migration (in vitro). Int. J. Mol. Sci. 2023, 24, 12673. [Google Scholar] [CrossRef]
40. Giammello, F.; Biella, C.; Priori, E.C.; Filippo, M.A.D.S.; Leone, R.; D’Ambrosio, F.; Paterno, M.; Cassioli, G.; Minetti, A.; Macchi, F.; et al. Modulating voltage-gated sodium channels to enhance differentiation and sensitize glioblastoma cells to chemotherapy. Cell Commun. Signal. 2024, 22, 434. [Google Scholar] [CrossRef]
41. Kim, H.; Xue, X. Detection of total reactive oxygen species in adherent cells by 2′,7′-dichlorodihydrofluorescein diacetate staining. J. Vis. Exp. 2021. [Google Scholar] [CrossRef]
42. GenBank. Available online: https://www.ncbi.nlm.nih.gov/genbank (accessed on 8 January 2025).
43. Sathe, S.K.; Monaghan, E.K.; Kshirsagar, H.H.; Venkatachalam, M. Chemical Composition of Edible Nut Seeds and its Implications in Human Health. In Tree Nuts: Composition, Phytochemicals, and Health Effects; CRC Press: Boca Raton, FL, USA, 2009; pp. 11–35. [Google Scholar]
44. Wang, Y.; Sun, S.; Li, F.; Cao, X.; Sun, R. Production of vanillin from lignin: The relationship between β-O-4 linkages and vanillin yield. Ind. Crops Prod. 2018, 116, 116–121. [Google Scholar] [CrossRef]
45. Dulay, R.M.R.; Rivera, A.G.C.; Garcia, E.J.B. Mycelial growth and basidiocarp production of wild hairy sawgill Lentinus strigosus, a new record of naturally occurring mushroom in the Philippines. Biocatal. Agric. Biotechnol. 2017, 10, 242–246. [Google Scholar] [CrossRef]
46. Ediriweera, S.S.; Wijesundera, R.L.C.; Nanayakkara, C.M.; Weerasena, O.V.D.S.J. Comparative study of growth and yield of edible mushrooms, Schizophyllum commune Fr., Auricularia polytricha (Mont.) Sacc. and Lentinus squarrosulus Mont. on lignocellulosic substrates. Mycosphere 2015, 6, 760–765. [Google Scholar] [CrossRef]
47. Leon, A.D.; Reyes, R.G.; Cruz, T.D. Enriched cultivation of three wild strains of Lentinus tigrinus (Bull.) Fr. using agricultural wastes. Int. J. Agric. Technol. 2013, 9, 1199–1214. [Google Scholar]
48. Yazdi, Y.; Rezaei, M.R.; Pourianfar, H.R. Antibacterial Activity of Culinary-Medicinal Polypore Mushroom Lentinus tigrinus (Agaricomycetes). Int. J. Med. Mushrooms 2023, 25, 21–30. [Google Scholar] [CrossRef] [PubMed]
49. Pourianfar, H.R.; Mohammadnejad, S.; Shahtahmasebi, S.; Ansari, A.M.; Zibaei, S.; Ghadirian, R.; Rezaeian, S.; Dowom, S.A. Toxicity and nutritional assessment of extracts of medicinal Tiger Sawgill mushroom, Lentinus tigrinus (Agaricomycetes), a newly domesticated in Iran. Int. J. Med. Mushrooms 2020, 22, 1079–1088. [Google Scholar] [CrossRef] [PubMed]
50. Liuzzi, G.M.; Petraglia, T.; Latronico, T.; Crescenzi, A.; Rossano, R. Antioxidant compounds from edible mushrooms as potential candidates for treating age-related neurodegenerative diseases. Nutrients 2023, 15, 1913. [Google Scholar] [CrossRef] [PubMed]
51. Venturella, G.; Ferraro, V.; Cirlincione, F.; Gargano, M.L. Medicinal mushrooms: Bioactive compounds, use, and clinical trials. Int. J. Mol. Sci. 2021, 22, 634. [Google Scholar] [CrossRef]
52. EFSA Panel on Nutrition, Novel Foods and Food Allergens (NDA); Turck, D.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Kearney, J.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; et al. Safety of Vitamin D2 mushroom powder (Agaricus bisporus) as a Novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2021, 19, e06516. [Google Scholar]
53. Bell, V.; Silva, C.R.P.G.; Guina, J.; Fernandes, T.H. Mushrooms as future generation healthy foods. Front. Nutr. 2022, 9, 1050099. [Google Scholar] [CrossRef]
54. Chen, C.; Han, Y.; Li, S.; Wang, R.; Tao, C. Nutritional, antioxidant, and quality characteristics of novel cookies enriched with mushroom (Cordyceps militaris) flour. CyTA J. Food 2021, 19, 137–145. [Google Scholar] [CrossRef]
55. Mwangi, R.W.; Macharia, J.M.; Wagara, I.N.; Bence, R.L. The antioxidant potential of different edible and medicinal mushrooms. Biomed. Pharmacother. 2022, 147, 112621. [Google Scholar] [CrossRef]
56. Mukherjee, A.; Das, S.; Chakraborty, D.; Pal, N.; Das, N. Fungi’s treasure in cosmeceuticals-a comprehensive chemical approach. S. Afr. J. Bot. 2024, 166, 311–331. [Google Scholar] [CrossRef]
57. Taofiq, O.; Rodrigues, F.; Barros, L.; Barreiro, M.F.; Ferreira, I.C.; Oliveira, M.B.P. Mushroom ethanolic extracts as cosmeceuticals ingredients: Safety and ex vivo skin permeation studies. Food Chem. Toxicol. 2019, 127, 228–236. [Google Scholar] [CrossRef] [PubMed]
58. Zhao, J.; Wang, X.; Lin, H.; Lin, Z. Hazelnut and its by-products: A comprehensive review of nutrition, phytochemical profile, extraction, bioactivities and applications. Food Chem. 2023, 413, 135576. [Google Scholar] [CrossRef] [PubMed]
59. Dulay, R.M.R.; Andres, S.M.B.; Asuncion, A.F.C.; Calalang, A.S.; Cumbe, A.P. Mycelial biomass production and radical scavenging activity of Lentinus tigrinus in submerged cultivation using selected tropical fruit juice. Int. J. Biol. Pharm. Allied Sci. 2017, 6, 2154–2161. [Google Scholar]
60. Lechner, B.E.; Papinutti, V.L. Production of lignocellulosic enzymes during growth and fruiting of the edible fungus Lentinus tigrinus on wheat straw. Process Biochem. 2006, 41, 594–598. [Google Scholar] [CrossRef]
61. Jasinska, A. Sustainability of Mushroom Cultivation Systems. Horticulturae 2023, 9, 1191. [Google Scholar] [CrossRef]
62. Diamantopoulou, P.; Fourtaka, K.; Melanouri, E.M.; Dedousi, M.; Diamantis, I.; Gardeli, C.; Papanikolaou, S. Examining the impact of substrate composition on the biochemical properties and antioxidant activity of Pleurotus and Agaricus mushrooms. Fermentation 2023, 9, 689. [Google Scholar] [CrossRef]
63. Siwulski, M.; Rzymski, P.; Budka, A.; Kalač, P.; Budzyńska, S.; Dawidowicz, L.; Hajduk, E.; Kozak, L.; Budzulak, J.; Sobieralski, K.; et al. The effect of different substrates on the growth of six cultivated mushroom species and composition of macro and trace elements in their fruiting bodies. Eur. Food Res. Technol. 2019, 245, 419–431. [Google Scholar] [CrossRef]
64. Ab Rhaman, S.M.S.; Naher, L.; Siddiquee, S. Mushroom quality related with various substrates’ bioaccumulation and translocation of heavy metals. J. Fungi 2021, 8, 42. [Google Scholar] [CrossRef]
65. Gupta, S.; Summuna, B.; Gupta, M.; Annepu, S.K. Edible mushrooms: Cultivation, bioactive molecules, and health benefits. Bioact. Mol. Food 2018, 1, 1815–1847. [Google Scholar]
66. Atila, F. A useful way to dispose of phenolic-rich agro-industrial wastes: Mushroom cultivation. Eur. J. Eng. Nat. Sci. 2019, 3, 32–41. [Google Scholar]
67. Attaran Dowom, S.; Rezaeian, S.; Pourianfar, H.R. Agronomic and environmental factors affecting cultivation of the winter mushroom or Enokitake: Achievements and prospects. Appl. Microbiol. Biotechnol. 2019, 103, 2469–2481. [Google Scholar] [CrossRef] [PubMed]
68. Anke, T. Secondary metabolites from mushrooms. J. Antibiot. 2020, 73, 655–656. [Google Scholar] [CrossRef] [PubMed]
69. Kalač, P. A review of chemical composition and nutritional value of wild-growing and cultivated mushrooms. J. Sci. Food Agric. 2013, 93, 209–218. [Google Scholar] [CrossRef] [PubMed]
70. Bouyahya, A.; Lagrouh, F.; El Omari, N.; Bourais, I.; El Jemli, M.; Marmouzi, I.; Salhi, N.; Faouzi, M.E.A.; Belmehdi, O.; Dakka, N.; et al. Essential oils of Mentha viridis rich phenolic compounds show important antioxidant, antidiabetic, dermatoprotective, antidermatophyte and antibacterial properties. Biocatal. Agric. Biotechnol. 2020, 23, 101471. [Google Scholar] [CrossRef]
71. Deore, B.; Vartak, R. Role of Flavonoids in Chronic Metabolic Diseases: From Bench to Clinic, 1st ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA; Scrivener Publishing LLC: Beverly, MA, USA; Wiley-Scrivener: Hoboken, NJ, USA, 2024; pp. 51–66. [Google Scholar]
72. Herrera-Rocha, K.M.; Rocha-Guzmán, N.E.; Gallegos-Infante, J.A.; González-Laredo, R.F.; Larrosa-Pérez, M.; Moreno-Jiménez, M.R. Phenolic acids and flavonoids in acetonic extract from quince (Cydonia oblonga Mill.): Nutraceuticals with antioxidant and anti-inflammatory potential. Molecules 2022, 27, 2462. [Google Scholar] [CrossRef]
73. Woźniak, D.; Nawrot-Hadzik, I.; Kozłowska, W.; Ślusarczyk, S.; Matkowski, A. Caffeoylquinic Acids. In Handbook of Dietary Phytochemicals; Springer: Berlin/Heidelberg, Germany, 2021; pp. 1065–1104. [Google Scholar]
74. Park, H.J. Chemistry and pharmacological action of caffeoylquinic acid derivatives and pharmaceutical utilization of chwinamul (Korean Mountainous vegetable). Arch. Pharmacal Res. 2010, 33, 1703–1720. [Google Scholar] [CrossRef]
75. Alcázar Magaña, A.; Kamimura, N.; Soumyanath, A.; Stevens, J.F.; Maier, C.S. Caffeoylquinic acids: Chemistry, biosynthesis, occurrence, analytical challenges, and bioactivity. Plant J. 2021, 107, 1299–1319. [Google Scholar] [CrossRef]
76. Palacios, I.; Lozano, M.; Moro, C.; D’Arrigo, M.; Rostagno, M.A.; Martínez, J.A.; Garcia-Lafuente, A.; Guillamon, E.; Villares, A. Antioxidant properties of phenolic compounds occurring in edible mushrooms. Food Chem. 2011, 128, 674–678. [Google Scholar] [CrossRef]
77. EFSA Panel on Plant Protection Products and their Residues (PPR); Jerez, A.H.; Adriaanse, P.; Berny, P.; Coja, T.; Duquesne, S.; Focks, A.; Marinovich, M.; Millet, M.; Pelkonen, O.; et al. Statement on the active substance acetamiprid. EFSA J. 2022, 20, e07031. [Google Scholar]
78. EFSA Panel name on Food Additives and Flavourings (FAF); Younes, M.; Aquilina, G.; Castle, L.; Degen, G.; Engel, K.H.; Flower, P.J.; Fernandes, M.J.F.; Furst, P.; Gundert-Remy, U.; et al. Scientific opinion on the renewal of the authorisation of Smoke Concentrate 809045 (SF-003) as a smoke flavouring Primary Product. EFSA J. 2023, 21, e08365. [Google Scholar]
79. Liu, G.; Zhu, W.; Li, S.; Zhou, W.; Zhang, H.; Wang, J.; Liu, X.; Liang, L.; Xu, X. Antioxidant capacity and interaction of endogenous phenolic compounds from tea seed oil. Food Chem. 2022, 376, 131940. [Google Scholar] [CrossRef] [PubMed]
80. Fuso, A.; Risso, D.; Rosso, G.; Rosso, F.; Manini, F.; Manera, I.; Caligiani, A. Potential valorization of hazelnut shells through extraction, purification and structural characterization of prebiotic compounds: A critical review. Foods 2021, 10, 1197. [Google Scholar] [CrossRef] [PubMed]
81. Zhu, Y.; Liu, J.; Liao, Y.; Lv, W.; Ma, L.; Wang, C. Degradation of vanillin during lignin valorization under alkaline oxidation. In Lignin Chemistry; Springer: Berlin/Heidelberg, Germany, 2018; pp. 33–51. [Google Scholar]
82. Bogdan, V.I.; Sergeeva, Y.E.; Lunin, V.V.; Perminova, I.V.; Konstantinov, A.I.; Zinchenko, G.E.; Bogdan, K.V. Bioconversion of phenolic monomers of lignin and Lignin-Containing substrates by the basidiomycete Lentinus tigrinus. Appl. Biochem. Microbiol. 2018, 54, 198–205. [Google Scholar] [CrossRef]
83. D’Arrigo, P.; Rossato, L.A.; Strini, A.; Serra, S. From Waste to Value: Recent Insights into Producing Vanillin from Lignin. Molecules 2024, 29, 442. [Google Scholar] [CrossRef]
84. Khoyratty, S.; Verpoorte, R.; Kodja, H. Vanillin: Biosynthesis, biotechnology, and bioproduction. In Orchids Phytochemistry, Biology and Horticulture: Fundamentals and Applications; Springer International Publishing: Cham, Switzerland, 2022; pp. 341–358. [Google Scholar]
85. Xu, L.; Liaqat, F.; Sun, J.; Khazi, M.I.; Xie, R.; Zhu, D. Advances in the vanillin synthesis and biotransformation: A review. Renew. Sustain. Energy Rev. 2024, 189, 113905. [Google Scholar] [CrossRef]
86. Lattanzio, V. Phenolic compounds: Introduction. In Natural Products; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1543–1580. [Google Scholar]
87. Suwannarach, N.; Kumla, J.; Zhao, Y.; Kakumyan, P. Impact of cultivation substrate and microbial community on improving mushroom productivity: A review. Biology 2022, 11, 569. [Google Scholar] [CrossRef]
88. Naim, M.J. A review on mushrooms as a versatile therapeutic agent with emphasis on its bioactive constituents for anticancer and antioxidant potential. Explor. Med. 2024, 5, 312–330. [Google Scholar] [CrossRef]
89. Cirlincione, F.; Venturella, G.; Gargano, M.L.; Mirabile, G.; Balenzano, G.; Pardi, R.; Ferraro, V. Medicinal wood decay fungi on large old oaks in the Madonie Mountains (Sicily). Ital. J. Mycol. 2024, 53, 160–165. [Google Scholar]
90. Gargano, M.L.; van Griensven, L.J.L.D.; Isikhuemhen, O.S.; Lindequist, U.; Venturella, G.; Wasser, S.P.; Zervakis, G.I. Medicinal mushrooms: Valuable biological resources of high exploitation potential. Plant Biosyst. 2017, 151, 548–565. [Google Scholar] [CrossRef]

Desiderio A, Pedrosa MC, Heleno SA, Carocho M, Rodrigues DB, Buratti S, Soffientini I, Ratto D, Savino E, Rossi P. Bio-Recycling Hazelnut Shells to Improve Antioxidant Properties of Lentinus tigrinus Sporophore. Agriculture. 2025; 15(2):178. https://doi.org/10.3390/agriculture15020178
Перевод статьи «Bio-Recycling Hazelnut Shells to Improve Antioxidant Properties ofLentinus tigrinusSporophore» авторов Desiderio A, Pedrosa MC, Heleno SA, Carocho M, Rodrigues DB, Buratti S, Soffientini I, Ratto D, Savino E, Rossi P., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык

















































































































Комментарии (0)