Влияние концентрации крахмала в рационе на продуктивность молока, усвояемость питательных веществ и выбросы метана у дойных коров в середине лактации
Целью нашего исследования была оценка влияния концентрации крахмала в рационе на продуктивность молока, усвояемость питательных веществ и выбросы метана у лактирующих дойных коров. Тридцать коров в середине лактации после 3-недельного адаптационного периода были случайным образом распределены на две группы: с рационом с высоким содержанием нейтрально-детергентной клетчатки и низким содержанием крахмала (НК; 20,2% крахмала) или с рационом с низким содержанием нейтрально-детергентной клетчатки и высоким содержанием крахмала (ВК; 25,2% крахмала). Исследование продолжалось 8 недель, при этом пробы молока и замеры газов проводились еженедельно в течение адаптационного периода, а также на 2, 4, 6 и 8 неделях. Пробы крови и фекалий собирались в период адаптации и на 8 неделе.
Аннотация
По сравнению с коровами группы НК, коровы группы ВК производили на 1,9 кг/день больше молока, скорректированного по энергии (увеличение на 4,45%), с более высоким выходом истинного белка (+0,13 кг/день), лактозы (+0,10 кг/день) и общих сухих веществ (+0,24 кг/день). Переваримость сухого вещества и органического вещества была на 4,2% и 4,3% выше, соответственно, в группе ВК. Профиль жирных кислот (ЖК) в молоке различался: у коров группы НК было выше содержание смешанных ЖК, а у коров группы ВК наблюдалось более высокое содержание и выход de novo ЖК. Хотя производство метана имело тенденцию к увеличению у коров группы ВК (+25 г/день), выход метана снизился на 8,8%.
В целом, рацион с высоким содержанием крахмала улучшил продуктивность молока, усвояемость питательных веществ и экологическую эффективность за счет снижения выхода метана у дойных коров.
1. Введение
Прогнозируется, что к 2050 году численность населения мира достигнет 9,7 млрд человек [ 1 ], что представляет собой серьезную проблему увеличения поставок продовольствия при минимизации воздействия на окружающую среду. Текущие системы производства и модели потребления считаются неустойчивыми [ 2 ]. Следовательно, сельское хозяйство находится на критическом этапе, поскольку ему необходимо удовлетворять как потребности растущего населения, так и его воздействие на окружающую среду. В животноводческой отрасли особое внимание уделяется сокращению выбросов метана (CH₄) из-за значительного вклада жвачных животных в антропогенные уровни CH₄. Потенциал глобального потепления метана в 27-30 раз превышает потенциал углекислого газа (CO 2 ) в течение 100 лет [ 3 ]. Кроме того, его более короткая продолжительность жизни в атмосфере делает его еще более мощным в течение 20-летнего периода, в 84-86 раз больше, чем CO 2 . Эти факторы предполагают, что нацеливание на CH 4 может быть более эффективной стратегией для краткосрочных усилий по смягчению последствий изменения климата [ 3 ].
Метаногенез у жвачных животных не только представляет собой проблему для окружающей среды, но и представляет собой потерю валовой энергии рациона, отражая неоптимальное использование кормов [ 4 ]. Эта потеря энергии значительна и составляет от 2 до 12% валовой энергии потребления, в среднем от 5 до 6% у молочного скота [ 5 ]. Поэтому исследователи и фермеры молочного скота активно изучают методы снижения энтеральных выбросов CH 4 на фермах при одновременном повышении эффективности коров [ 6 ]. На выработку CH 4 влияют различные диетические факторы, включая тип и количество корма, которые влияют на популяцию микробов рубца и изменяют использование газообразного водорода (H 2 ) и общие закономерности ферментации. Понимание профилей питательных веществ и оптимальных уровней включения в рацион имеет важное значение для снижения энтеральных выбросов CH 4. Например, выбор крахмала вместо клетчатки и увеличение содержания крахмала в концентрированной части рациона являются потенциальными стратегиями для снижения выработки CH 4 в рубце [ 7 ]. Этот подход особенно эффективен, когда концентраты скармливаются вместе с базовым рационом из низкокачественных кормов, что еще больше снижает выбросы CH₄ от крупного рогатого скота [ 8 ].
Крахмал является основным энергетическим компонентом в зернах, играя ключевую роль в качестве первичного источника глюкогенной энергии для высокопродуктивных молочных коров, выступая в качестве ферментируемого субстрата для микроорганизмов рубца и стимулируя синтез микробного белка [ 9 ]. Понимание переваривания крахмала необходимо для оптимизации метаболизируемого белка и снабжения энергией, тем самым повышая эффективность рациона [ 10 ]. Ферментация корма в рубце производит летучие жирные кислоты, CO 2 и H 2 . Метаногенные археи используют этот H 2 для преобразования CO 2 в CH 4 . По сравнению с пищевыми волокнами, ферментация крахмала может снизить выработку кишечного CH 4 , поскольку она генерирует больше пропионата, обеспечивая альтернативный сток H 2 для метаногенеза [ 11 ]. Кроме того, крахмал снижает pH рубца, создавая неблагоприятную среду для метаногенов, простейших и целлюлозолитических бактерий. Эта кислая среда также затрудняет усвояемость клетчатки и снижает доступность H2 для производства CH4 [ 12 , 13 ] . Более того, в отличие от клетчатки и сахара, значительная часть крахмала может обходить рубцовую ферментацию и подвергаться ферментативному перевариванию в тонком кишечнике, способствуя снабжению животного энергией без связанных с производством CH4 потерь [ 14 ] .
Тем не менее, несколько факторов, включая источник крахмала, уровень включения и скорость ферментации, могут влиять на усвояемость крахмала и, следовательно, на производство CH₄. Агуэрре и др. [ 15 ] оценили четыре рациона с различными соотношениями фуража и концентрата и уровнями крахмала от 20,0 до 29,0%. Они обнаружили, что увеличение содержания крахмала снижало выработку CH4 , ее интенсивность и выход, не влияя на потребление сухого вещества (DMI) или надои молока [ 15 ]. Пирондини и др. [ 16 ] сравнили два уровня крахмала (23,8 против 28,0%), изменив состав концентрата при сохранении постоянного включения фуража, и заметили, что группа с более низким содержанием крахмала имела более высокую усвояемость сухого вещества (DM) и органического вещества (OM), без различий в выработке, интенсивности или выходе CH4. Хэтью и др. [ 17 ] исследовали различные скорости ферментации крахмала и уровни включения, обнаружив, что быстро ферментирующий крахмал и более высокие уровни диетического крахмала снижали выход CH4 . Кроме того, повышенное содержание крахмала снижало выработку CH4 из -за снижения потребления сухого вещества [ 17 ]. Непоследовательность результатов исследований с участием молочных коров может быть обусловлена разницей в уровнях крахмала между различными видами кормления, различиями в составе ингредиентов основных рационов или несоответствиями потребления сухого вещества и уровня продуктивности.
Измерение выбросов CH 4 может быть сложной задачей, что побуждает к исследованию альтернативных методов для более практичной и экономичной оценки. Один из многообещающих подходов к прогнозированию продукции CH 4 у дойных молочных коров включает анализ концентрации конкретных жирных кислот (ЖК) в молоке [ 18 ]. Дейкстра и др. [ 19 ] наблюдали положительную связь между продукцией CH 4 и концентрациями C14:0 изо и C15:0 изо в молоке, а также обратную связь с несколькими транс-промежуточными продуктами, в частности C18:1 транс -10 и транс -11. Аналогичным образом, Рико и др. [ 20 ] сообщили об отрицательной корреляции между продукцией CH 4 и различными ненасыщенными жирными кислотами молока с длиной углеродной цепи 16, 18, 20 и 22. Хотя этот подход показывает потенциал, необходимы дальнейшие исследования для подтверждения этих корреляций.
Целью данного исследования была оценка влияния двух различных уровней содержания крахмала в рационе на молочную продуктивность, усвояемость питательных веществ и выбросы CH4 у молочных коров в середине лактации. Мы предположили, что увеличение концентрации крахмала при одновременном снижении содержания клетчатки в рационе приведет к повышению потребления сухого вещества (СВ), надоев молока, концентрации чистого белка в молоке и усвояемости органического вещества (ОВ), одновременно снижая выход CH4 , интенсивность CH4 и усвояемость клетчатки. Целью данного исследования является расширение нашего понимания оптимальных стратегий кормления для повышения молочной продуктивности и снижения выбросов CH4 в глобальном масштабе. Кроме того, мы стремимся к дальнейшему изучению корреляции между выработкой CH4 и удельной жирной кислотностью молока .
2. Материалы и методы
2.1 Экспериментальный дизайн
Все экспериментальные процедуры проводились в соответствии с протоколом Комитета по уходу и использованию животных Корнеллского университета (протокол № 2022-0132). Тридцать коров голштинской породы в середине лактации со средней продолжительностью лактации (± SD) 2,53 ± 1,78, продуктивностью 117 ± 24,9 дней и надоем 38,3 ± 9,13 кг молока в день были включены в исследование с полностью рандомизированным дизайном, проведенное в Центре исследований молочного хозяйства Корнеллского университета в Харфорде, штат Нью-Йорк, США. После 3 недель акклиматизации к коровнику с привязным содержанием и обучения использованию установок GreenFeed (C-Lock, Inc., Rapid City, SD, USA) коровы были распределены в одну из двух групп лечения (15 коров на лечение) следующим образом: диета с высоким содержанием нейтрально-детергентной клетчатки и низким содержанием крахмала (LS; 20,2% крахмала) или диета с низким содержанием нейтрально-детергентной клетчатки и высоким содержанием крахмала (HS; 25,2% крахмала). Коровы были сбалансированы по удою с поправкой на энергию (ECM), дню в молоке и количеству опоросов на момент распределения. Критерии включения включали отсутствие активного или 30-дневного предыдущего случая мастита. Рационы состояли в основном из кукурузного силоса, тритикалевого силоса, молотой кукурузы, соевого шрота и соевой шелухи ( таблица 1 ). Все рационы были составлены с использованием AMTS.Farm.Cattle(Pro) (Agricultural Modeling & Training Systems, LLC, Гротон, Нью-Йорк, США) для удовлетворения потребностей в обменной энергии (ME) и протеине коровы 2-й лактации весом 743 кг, потребляющей 26,0 кг/день сухого вещества и производящей 41,0 кг/день молока с 4,10% жира и 3,40% чистого протеина. Рационы обеспечивали 1,14 г метионина на мегакал ME и поддерживали соотношение лизина к метионину приблизительно 2,7:1. Ни один из рационов не содержал пробиотиков, монензина, дрожжей или их производных. Различия в уровнях крахмала достигались путем корректировки пропорций грубых кормов и концентратов. Концентраты были предоставлены Purina Animal Nutrition (Трумансбург, Нью-Йорк, США). Рационы смешивали и подавали в виде полных смешанных рационов (ПСР) ежедневно примерно в 07:00, при этом количество ПСР ежедневно корректировали для достижения 10% отказов. Коров доили 3 раза в день в 06:00, 14:00 и 22:00. Температура и влажность в коровнике контролировались ежедневно, а индекс температуры и влажности рассчитывался для оценки условий окружающей среды. Вес тела (МТ) и оценка упитанности (УБ; шкала от 1 до 5) [ 21 ] измерялись еженедельно. Руминацию постоянно регистрировали с помощью ошейников Allflex (Allflex Livestock Intelligence Global, Мэдисон, Висконсин, США) на протяжении всего исследования [ 22 ].
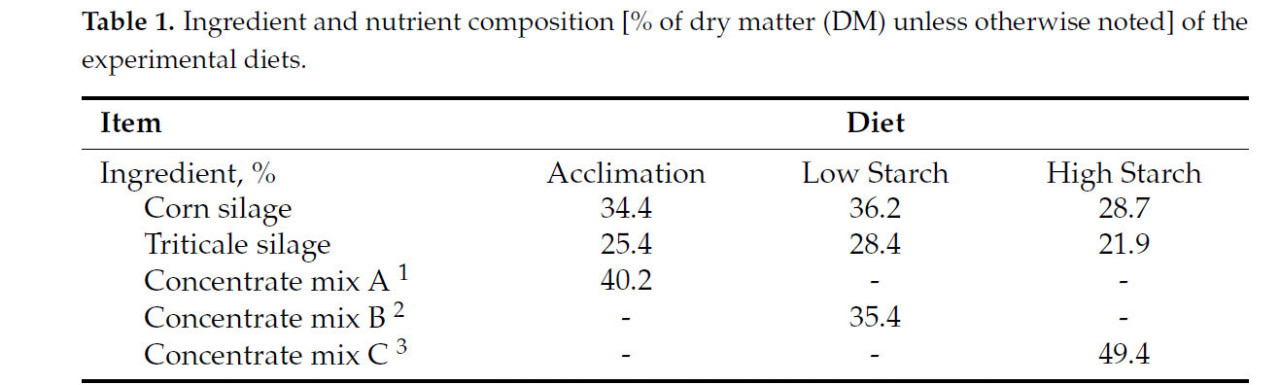
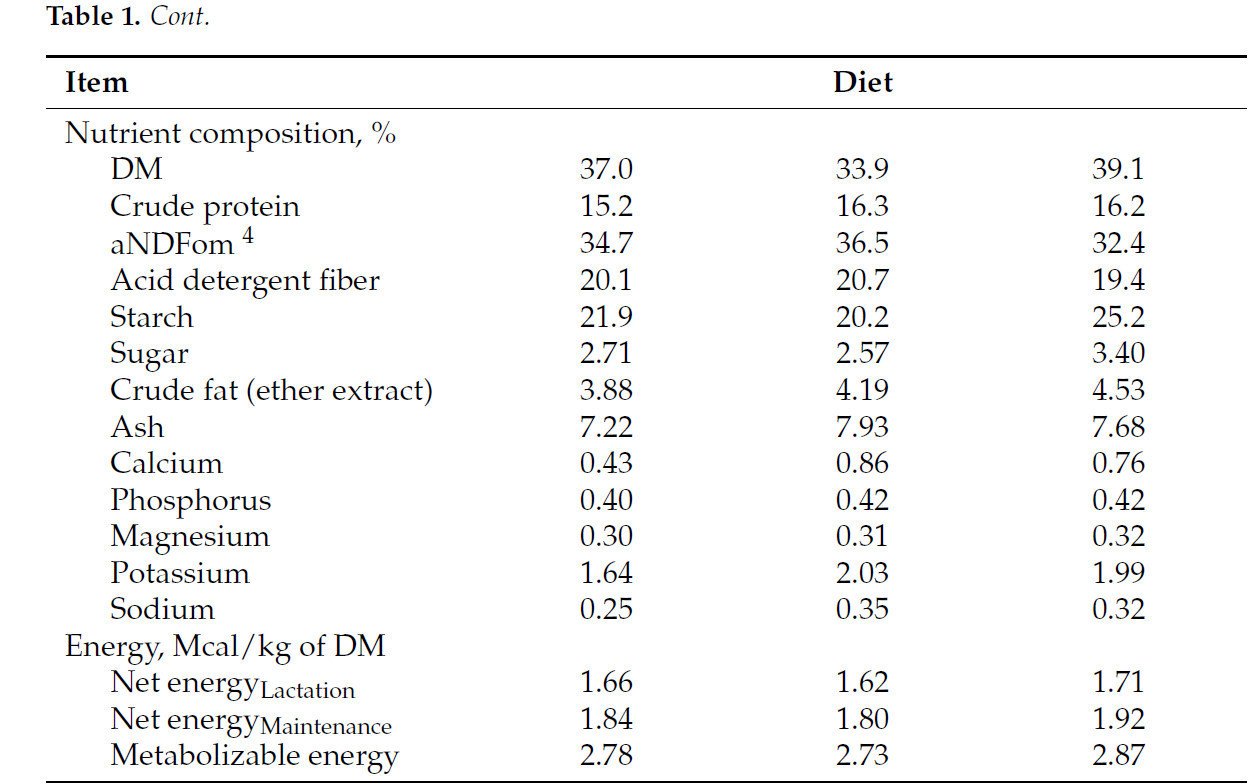
![<b>Таблица 1.</b> Состав ингредиентов и питательных веществ [% от сухого вещества (СВ), если не указано иное] экспериментальных рационов.](https://storage.agriexpert.ru/storage/images/articles/5958/block/89145/model/xl/siocARHk76rq.jpg)
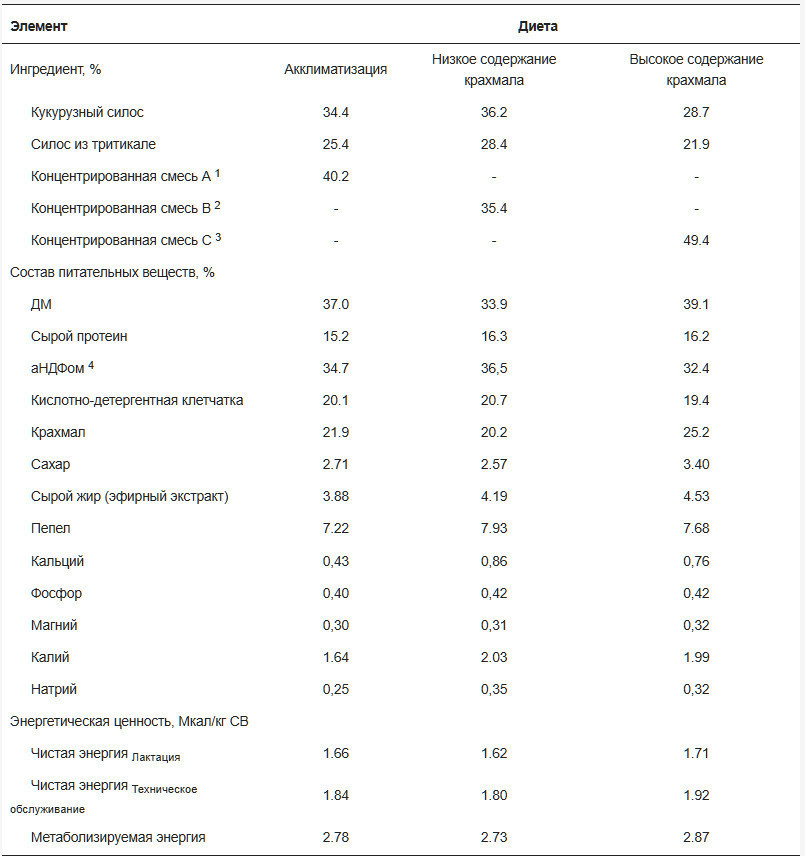
Таблица 1. Состав ингредиентов и питательных веществ [% от сухого вещества (СВ), если не указано иное] экспериментальных рационов.
2.2. Отбор проб кормов и анализы
Образцы отдельных ингредиентов, TMR и отказов собирали еженедельно, высушивали в течение 72 ч при 55 °C в печи с принудительной циркуляцией воздуха (VWR Scientific) для определения DM и измельчали до состояния, пригодного для прохождения через сито 1 мм с использованием мельницы Wiley (AH Thomas Co., Филадельфия, Пенсильвания, США). Измельченные образцы TMR составлялись ежемесячно и анализировались в соответствии с методами AOAC [ 23 ] для определения DM: 934,01, сырого протеина (CP): 990,03, эфирного экстракта (EE): 2003,05, золы: 942,05, крахмала [ 24 ], кислотно-детергентной клетчатки (ADF): 973,18, беззольной нейтрально-детергентной клетчатки (aNDFom) [ 25 ] и нейтрально-детергентной клетчатки после 240 ч ферментации in vitro (iNDF) [ 26 ] компанией Cumberland Valley Analytical Services Inc. (Уэйнсборо, Пенсильвания, США). Дополнительные образцы TMR еженедельно анализировались на распределение размера частиц с помощью сепаратора частиц Penn State [ 27 ]. Два образца каждого рациона анализировались еженедельно, в общей сложности 16 образцов каждого рациона за период исследования.
2.3. Измерения газообразных выбросов
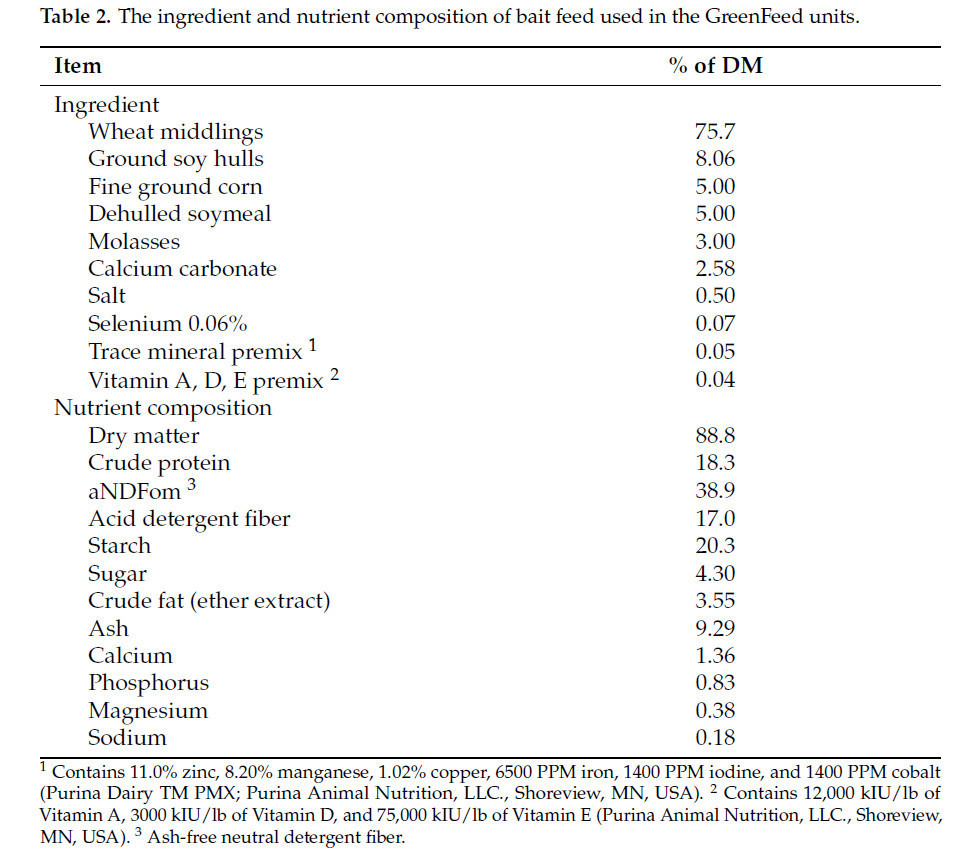
Кишечные выбросы CH 4 , CO 2 и H 2 оценивались с использованием 3 блоков GreenFeed в 02:00, 10:00 и 18:00, 3 дня в неделю (9 измерений в неделю) в течение недели −1, 2, 4, 6 и 8. До начала эксперимента коровы были акклиматизированы к блокам GreenFeed. Однако 1 корова в группе HS не посещала блок постоянно, в результате чего в этой группе лечения образовалась подгруппа из 14 коров. В качестве приманки использовался специально разработанный гранулированный корм (Purina Animal Nutrition, LLC., Shoreview, MN, USA), состоящий в основном из зерна, грубых кормов и мелассы ( таблица 2 ). Каждый сбор образцов длился от 5 до 7 минут с дополнительными 2 минутами для фоновых измерений. Для обеспечения постоянного потока воздуха воздушные фильтры в блоках GreenFeed менялись еженедельно. Кроме того, перед началом исследования, в течение 3-й недели и в конце исследования проводился тест на восстановление CO2. В ходе этого теста на восстановление датчик потока воздуха калибровался путем выпуска известного количества CO2 в каждую систему и сравнения количества выпущенного и собранного CO2. В результате степень восстановления CO2 составила 99,8 ± 2,59% (n = 9).

2.4. Отбор и анализ проб молока
Образцы молока собирали 3 дня в неделю (9 доек в неделю) в течение недель -1, 2, 4, 6 и 8. Образцы хранили в пробирках, содержащих консервант 2-бром-2-нитропропан-1,3-диол, и хранили при температуре 4 °C для анализа состава молока в течение 5 дней после сбора. Образцы анализировали на содержание жира, чистого белка, лактозы, азота мочевины молока и общую концентрацию сухих веществ с помощью инфракрасной спектроскопии с преобразованием Фурье и подсчета соматических клеток (SCC) методом проточной цитометрии (Dairy One, Итака, штат Нью-Йорк, США). Образцы молока для анализа состава жирных кислот составляли на основе выхода молочного жира, чтобы представить недели -1 и 8. Образцы центрифугировали при 17800 × g в течение 30 минут при 4 °C, а жировые осадки собирали и хранили при температуре -80 °C до экстракции липидов. Общие липиды из жировых лепешек (320 ± 10 мг) были извлечены с использованием н- гексана/изопропанола (3:2, об. / об. ) [ 28 ]. Анализ газожидкостной хроматографии (ГХ) был проведен с использованием GC system-8890 (Agilent Technologies, Пало-Альто, Калифорния, США), оснащенной пламенно-ионизационным детектором, автодозатором, инжектором с разделением/без плевания и колонкой CP-Sil 88 (внутренний диаметр 100 м × 0,25 мм, толщина пленки 0,20 мкм; Agilent, Technologies, Пало-Альто, Калифорния, США). Водород использовали в качестве газа-носителя со скоростью потока 1 мл/мин, для ПИД — 40 мл/мин, а подпиточный газ азот — 30 мл/мин. Инжектор и детектор поддерживались при температуре 250 °C. Программа температуры печи была следующей: начальная температура 80 °C поддерживалась в течение 1 мин, затем повышалась до 215 °C со скоростью 2 °C/мин и поддерживалась в течение 21,5 мин [ 29 ]. Каждый анализ ГХ включал инъекцию 1 мкл образца с соотношением разделения 1:100. Отдельные пики идентифицировали с использованием стандартных образцов (стандартный образец ГЖХ 463, стандартный образец ГЖХ 481-B и октадекадиеновая смесь # UC-59 M, Nu-Chek Prep Inc., Элизиан, Миннесота, США). Расхождения в массе метиловых эфиров короткоцепочечных жирных кислот корректировали с использованием факторов отклика, опубликованных Ульбертом и Шраммелем [ 30 ]. Концентрации жирных кислот определяли на основе массы с использованием молекулярной массы каждой жирной кислоты с поправкой на глицерин [ 31 ].
2.5. Забор крови и анализы
Образцы крови собирали путем венепункции копчиковых сосудов один раз в неделю в течение −1 и 8 недель. После сбора образцы крови немедленно помещали на лед примерно на 45 минут, затем центрифугировали при 2171× g в течение 20 минут при 4 °C. Образцы плазмы хранили при температуре −80 °C до анализа. Концентрацию глюкозы в плазме (Autokit Glucose № 997-03001, FUJIFILM Medical Systems USA, Лексингтон, Массачусетс, США) количественно определяли в двух повторностях в соответствии с инструкциями производителя. Концентрацию инсулина в плазме измеряли с помощью радиоиммунного анализа (набор для РИА свиного инсулина #PI-12K; EMD Millipore Corp., Берлингтон, Массачусетс, США) на счетчике LKB-Wallac CliniGamma (Beckman Coulter, Индианаполис, Индиана, США). Внутри- и межанализные коэффициенты вариации составили 4,79 и 1,65% и 4,53 и 2,96% для глюкозы и инсулина в плазме соответственно.
2.6. Отбор проб кала и анализы
Точечные образцы кала собирали непосредственно из прямой кишки или во время добровольной дефекации в течение недели −1 и 8. Образцы собирали каждые 5 часов в течение 3 последовательных дней для учета суточных изменений. В каждой точке отбора проб получали около 200 г образцов кала, которые переносили в 4-литровые пакеты для получения еженедельного составного образца от каждой коровы и хранили при температуре −20 °C до дальнейшей обработки. Образцы размораживали при комнатной температуре, помещали в алюминиевые лотки и лиофилизировали с использованием лиофилизатора Virtis модели 20SRC-X (Гардинер, штат Нью-Йорк, США); образцы выдерживали при температуре −40 °C в течение 3 часов, затем при −20 °C в течение 3 часов, 0 °C в течение 16 часов и 15 °C до высыхания. Затем образцы измельчали до прохождения через сито 1 мм с помощью мельницы Wiley (AH Thomas Co., Филадельфия, штат Пенсильвания, США). Образцы были отправлены в Cumberland Valley Analytical Services, Inc. (Уэйнсборо, Пенсильвания, США) для анализа DM, CP, EE, золы, крахмала, ADF, aNDFom и iNDF, как описано выше.
2.7. Расчеты и статистический анализ
Выход молока с поправкой на жирность 3,5 % (FCM), ECM и компонентов молока рассчитывали на основе надоев и концентраций компонентов от каждой дойки, суммировали для получения суточной суммы и усредняли для каждого периода сбора следующим образом: FCM = [(0,4324 × кг молока) + (16,216 × кг молочного жира)]) и ECM = [(0,327 × кг молока) + (12,95 × кг молочного жира) + (7,65 × кг истинного молочного белка)] [ 32 ]. Соматический балл клеток (SCS) рассчитывали из SCC с использованием логарифмического преобразования, где SCS = log 2 (SCC/100 000) + 3 [ 33 ]. Индивидуальный выход жиров (г/д) определялся с использованием выхода молочного жира и концентрации жирных кислот для определения выхода на основе массы, используя молекулярную массу каждой жирной кислоты с поправкой на глицерин. Эффективность кормления (FE) для надоя молока, FCM и производства ECM рассчитывалась как отношение надоя молока, FCM или ECM к DMI. Кажущаяся общая усвояемость тракта рассчитывалась с использованием следующего уравнения в соответствии с Хисадоми и соавторами [ 34 ]: усвояемость (%) = 100 − {100 × [содержание диетического iNDF (%DM)/содержание фекального iNDF (%DM)] × [содержание питательных веществ в фекалиях (%DM)/содержание питательных веществ в рационе (%DM)]}. Образцы выделения газа, собираемые еженедельно, суммировались для оценки общего выделения газа за период отбора проб. Интенсивность выбросов анализировалась путём расчёта газопродукции на кг надоя (г/кг надоя), а также на кг молока с энергетической коррекцией (г/кг ECM) и молока с жировой коррекцией (г/кг FCM). Газопродукцию определяли как общее газопродукцию на кг сухого вещества (г/кг DMI) и на кг потреблённого органического вещества (г/кг OMI). Выбросы CH4 в кишечнике выражали в эквиваленте CO2 ( CO2 - экв) с использованием коэффициента 100-летнего потенциала глобального потепления, равного 28. Процентная разница рассчитывалась по формуле {|a − b|/[(a + b) ÷ 2]} × 100.
Статистический анализ проводился с использованием процедуры смешанной модели SAS (v9.4, SAS Institute Inc., Кэри, Северная Каролина, США) в соответствии со следующей моделью:
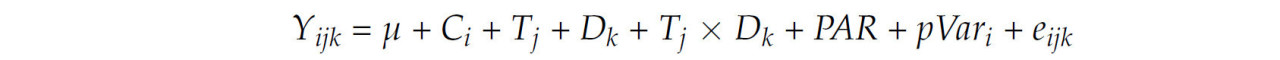
где Y ijk = зависимая переменная; μ = общий средний эффект для меры; C i = случайный эффект коровы (i = от 1 до 30); T j = фиксированный эффект уровня крахмала ( j = низкий или высокий); D k = фиксированный эффект wk (l = от 1 до 8); T j × D k = фиксированный эффект взаимодействия между уровнем крахмала и wk; PAR = паритет, используемый в качестве ковариата; pVar i = базовое измерение для каждой переменной отклика, используемой в качестве ковариата; и e ijk = остаточная ошибка. После оценки пяти различных структур ковариации (компоненты дисперсии, авторегрессия первого порядка, неструктурированная, сложная симметрия и антезависимость первого порядка) была выбрана наиболее подходящая структура ковариации для каждой переменной в анализе повторных измерений. Процесс выбора включал в себя определение структуры с самым низким коэффициентом информационного критерия Акаике для последующего анализа. Путем моделирования структуры ковариации были выявлены паттерны, которые наиболее эффективно характеризуют взаимосвязи между повторными измерениями в модели. Для множественных сравнений применялся апостериорный тест Тьюки, позволяющий выявить различия в каждой временной точке. Модель оценивала реакции продукции, метаболиты крови и показатели газового состава. Корреляция между продукцией CH4 и концентрацией жирных кислот в молоке оценивалась с помощью корреляций Пирсона. Для поддержания контролируемого уровня ложных срабатываний на уровне 5% применялись поправки на множественные сравнения [ 35 ]. Были зарегистрированы только корреляции, попадающие в допустимый диапазон данного теста.
Наблюдения считались выбросами, если стьюдентизированные остатки > 3,0 или < −3,0. Нормальность остатков проверялась с помощью нормального распределения и диаграммы ящиков, а однородность дисперсий – с помощью графиков остатков против прогнозируемых значений, чтобы убедиться в отсутствии нарушений допущений модели. Сравнения средних значений методом наименьших квадратов представлены с использованием скорректированных p -значений. Результаты представлены как средние значения методом наименьших квадратов ± стандартная ошибка среднего, если не указано иное. Основные эффекты считались значимыми при p ≤ 0,05 и имели тенденцию к повышению значимости при 0,05 < p ≤ 0,15.
3. Результаты
Коровы, получавшие рацион HS, производили на 2 кг больше молока (40,6 против 38,6 кг/д; p < 0,01; Рисунок 1 A) и потребляли на 4,2 кг больше DM (28,6 против 24,4 кг/д; p < 0,01; Рисунок 1 B) по сравнению с коровами на рационе LS. Однако FE была ниже у коров HS (1,43 против 1,57; p < 0,01; Рисунок 1 C), при этом взаимодействие крахмала × неделя наблюдалось в течение недель 5, 6, 7 и 8 ( Таблица 3 ). Кроме того, коровы HS имели большую массу тела (696 против 674 кг; p < 0,01) и, как правило, имели более высокий BCS (3,21 против 3,13; p = 0,08; Таблица 3 ) по сравнению с коровами LS. Распределение размеров частиц в HS TMR составило 3,25 ± 0,60% для частиц размером > 19,0 мм, 56,5 ± 2,50% для частиц размером 8,0–19,0 мм, 12,8 ± 1,10% для частиц размером 3,18–8,0 мм и 27,5 ± 2,50% для частиц размером < 3,18 мм. В то же время, в LS TMR доля частиц размером > 19,0 мм составила 4,62 ± 0,83%, частиц размером от 8,0 до 19,0 мм – 63,5 ± 2,52%, частиц размером от 3,18 до 8,0 мм – 12,8 ± 0,60% и частиц размером < 3,18 мм – 19,1 ± 2,33%. Коровы на диете HS имели более высокий надой ECM (44,6 против 42,7 кг/д; p = 0,04); содержание истинного протеина (3,47 против 3,27%; p < 0,01); и выход истинного протеина (1,36 против 1,23 кг/д; p < 0,01), лактозы (1,95 против 1,85 кг/д; p < 0,01) и общего количества сухих веществ (5,40 против 5,16 кг/д; p < 0,01; Таблица 3 ) по сравнению с коровами LS. Однако у коров LS, как правило, было более высокое содержание жира в молоке (4,45 против 4,28%; p = 0,09; Таблица 3 ). Взаимосвязь между обработкой × неделя для СВ, молока, надоев ECM и FCM, жвачки, содержания истинного белка в молоке, выхода молочного жира, выхода лактозы в молоке и FE (кг надоя молока/кг СВ; кг выхода ECM/кг СВ; кг выхода FCM/кг СВ) представлена на рисунках S1–S11 . Различий в концентрации глюкозы в плазме не наблюдалось ( p = 0,86; Таблица 3 ), но у коров HS, как правило, была более высокая концентрация инсулина в плазме по сравнению с коровами LS (1,61 против 1,31 нг/мл, p = 0,12; Таблица 3 ). Содержание крахмала в рационе влияло на профиль жирных кислот молока: у коров с низким содержанием жира (LS) наблюдалось более высокое содержание смешанных жирных кислот (35,2% против 32,8%; p < 0,01), а у коров с высоким содержанием жира (HS) – более высокое содержание и выход жирных кислот de novo (22,0% против 23,6% и 362% против 405 г/д; p < 0,01). Концентрации и выходы C18:2 цис -9 и цис -12 также были выше у коров с высоким содержанием жира (1,41% против 1,75% и 190% против 206 г/д соответственно; p ≤ 0,02; Таблица 4 ). Полный список концентраций жирных кислот и выходов молока представлен в Таблицах S1 и S2.. Видимая общая усвояемость DM и OM была ниже у коров LS по сравнению с коровами HS (69,4 против 73,6% и 70,5 против 74,8% соответственно; p < 0,01; Таблица 5 ). Выработка метана, как правило, была ниже у коров LS по сравнению с коровами HS (386 против 411 г/д, p = 0,08; Таблица 6 ; Рисунок 2 A), показывая разницу в 6,27%. Коровы на рационе LS также имели более низкие выбросы CO2 - эквивалентов на кг произведенного жира (8,34 против 8,99 кг CO2 - экв/кг жира; p = 0,02) и более низкую интенсивность CO2 в терминах FCM по сравнению с коровами HS (315 против 333 г CO2 / кг FCM; p < 0,01; Таблица 6 ). Однако у коров HS наблюдалось снижение выхода CH 4 по сравнению с коровами LS (14,6 против 16,0 г CH 4 /кг DMI; p = 0,03; Таблица 6 ; Рисунок 2 B), что составляет разницу в 9,15%. У коров на диете HS также наблюдалось снижение выхода CH 4 с точки зрения OMI (15,9 против 17,5 г CH 4 /кг OMI; p = 0,03; Таблица 6 ), что демонстрирует разницу в 9,58%. Кроме того, в течение 6-й недели наблюдалось взаимодействие крахмала × неделя ( p = 0,08; Таблица 6 ; Рисунок 2 B). Выработка метана отрицательно коррелировала с антеизо C15:0, C16:1 транс -9, C18:1 цис -9 и C20:1 цис -11 (−0,41, −0,43, −0,38 и −0,41 соответственно; p ≤ 0,05; Таблица 7 ).
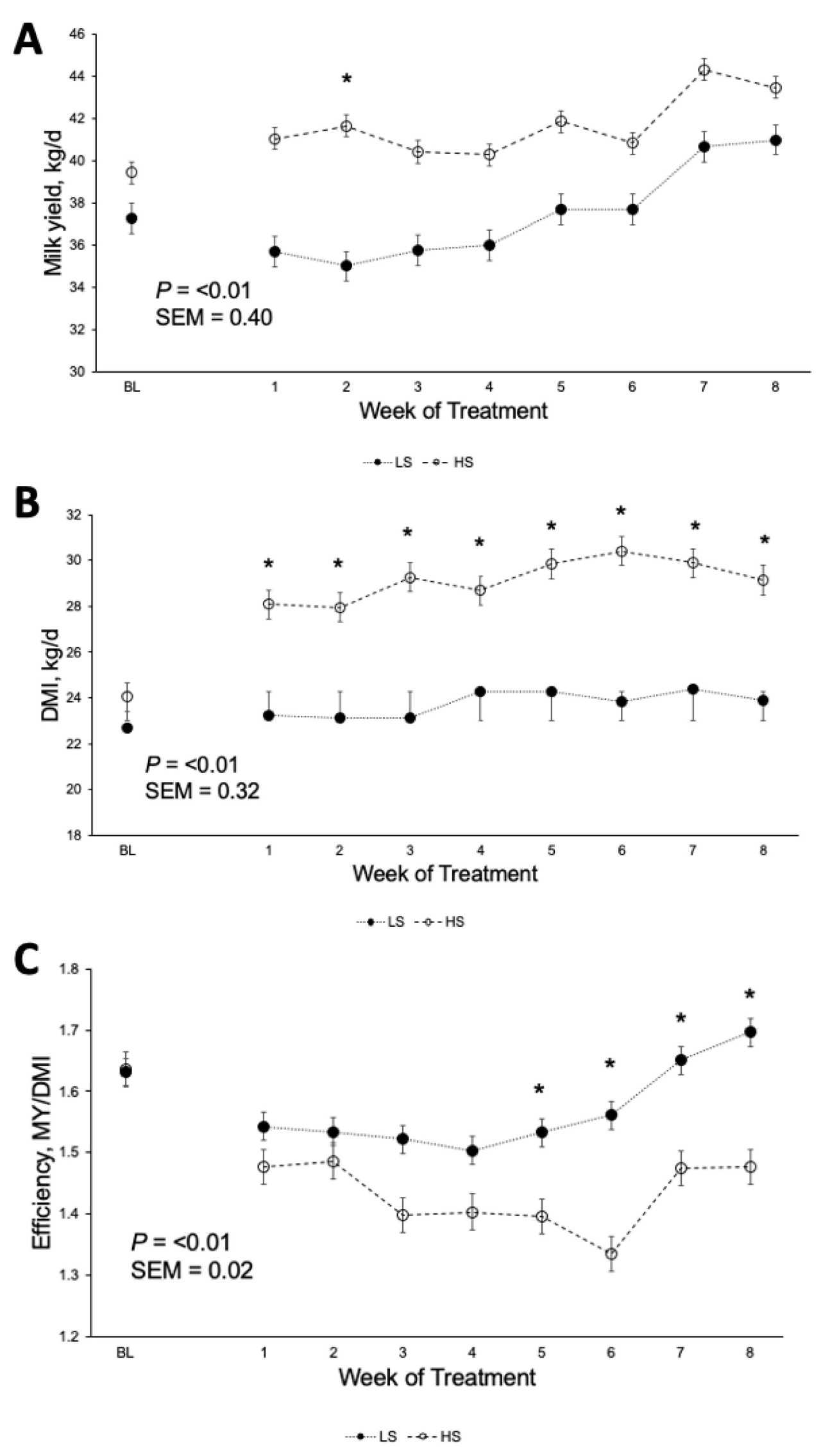
Рисунок 1. Влияние концентрации крахмала в рационе на ( A ) надои молока (MY), ( B ) потребление сухого вещества (DMI) и ( C ) эффективность (MY/DMI). * Указывает на значимое взаимодействие ( p ≤ 0,05) между неделей и лечением.
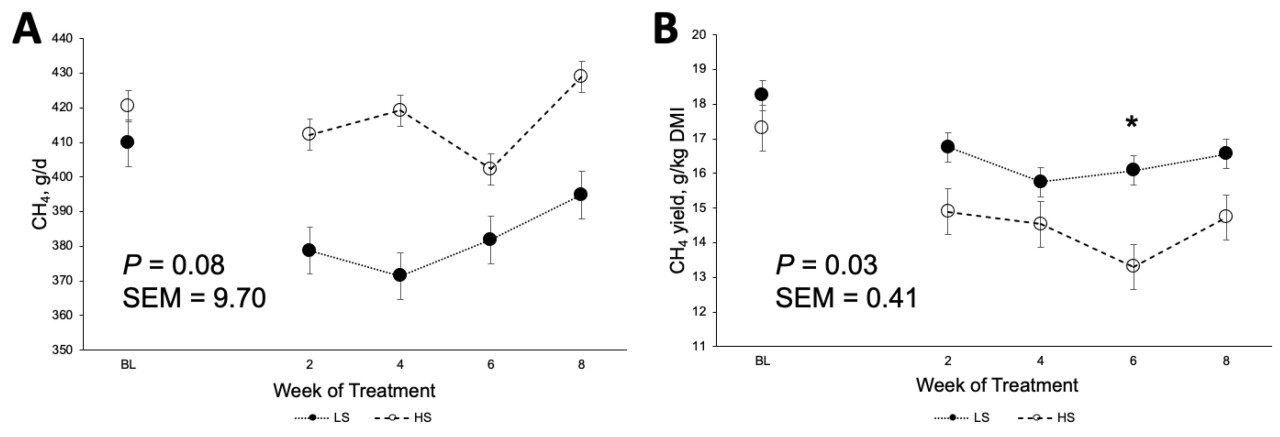
Рисунок 2. Влияние концентрации крахмала в рационе на ( A ) выработку метана (г CH4 / день) и ( B ) выход метана (г CH4 / кг сухого вещества). * Указывает на значимое взаимодействие ( p ≤ 0,05) между неделей и лечением.
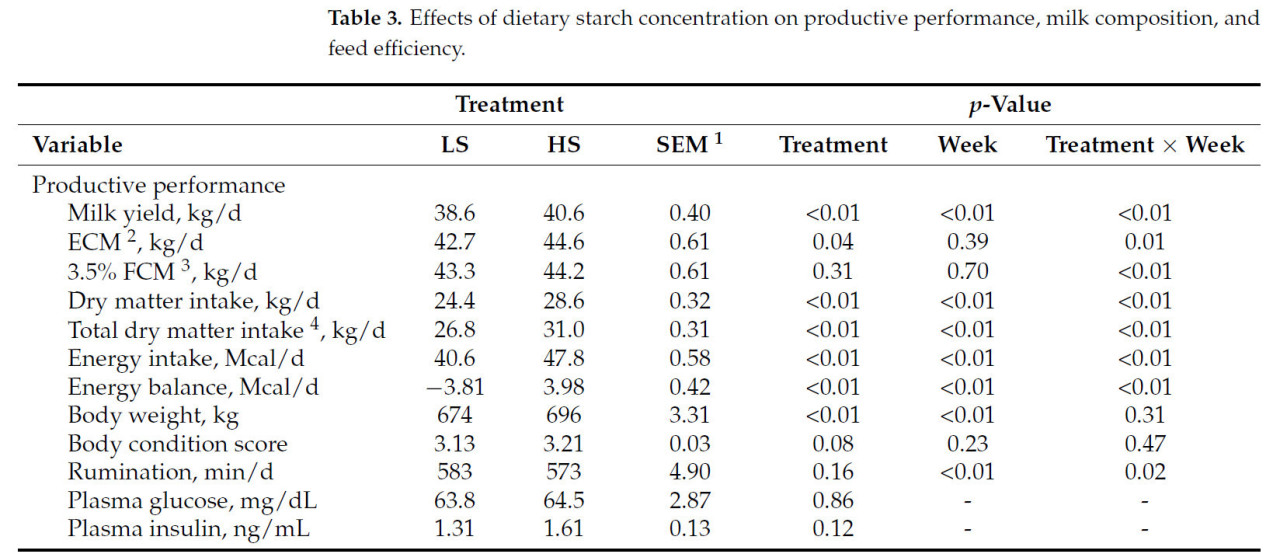
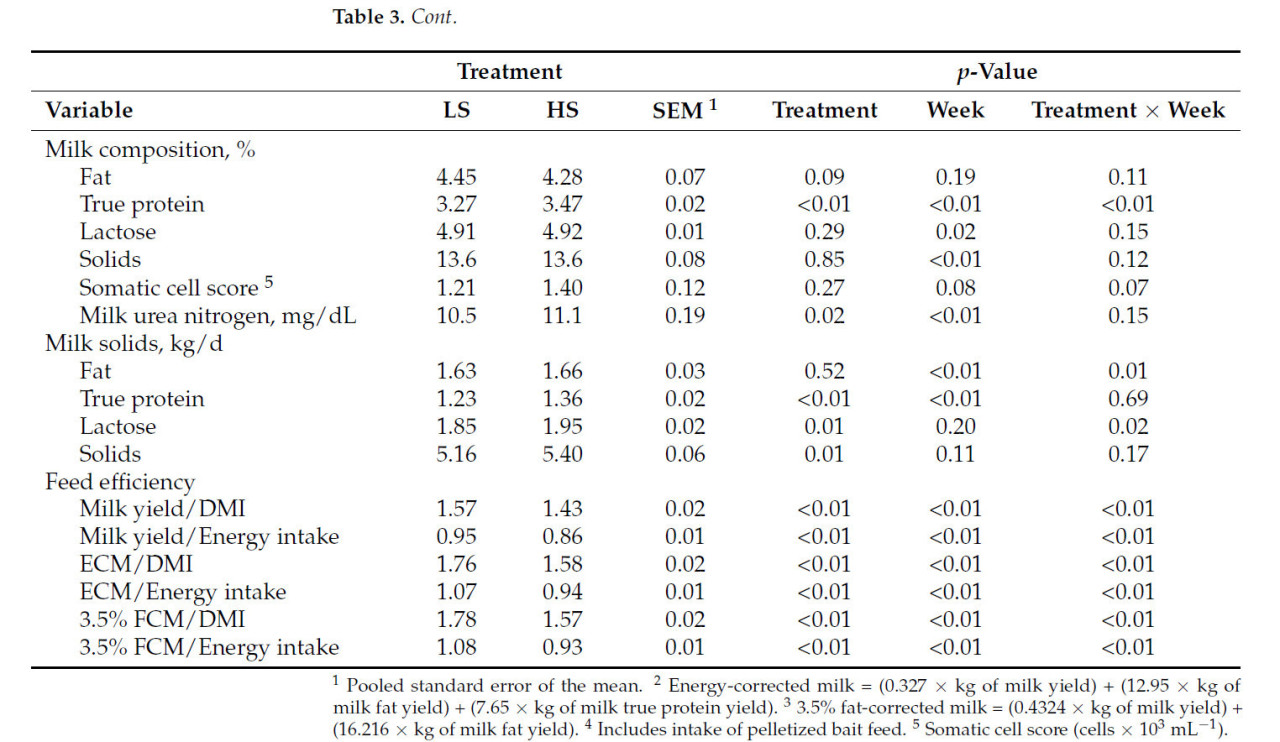
Таблица 3. Влияние концентрации крахмала в рационе на продуктивность, состав молока и эффективность кормления.

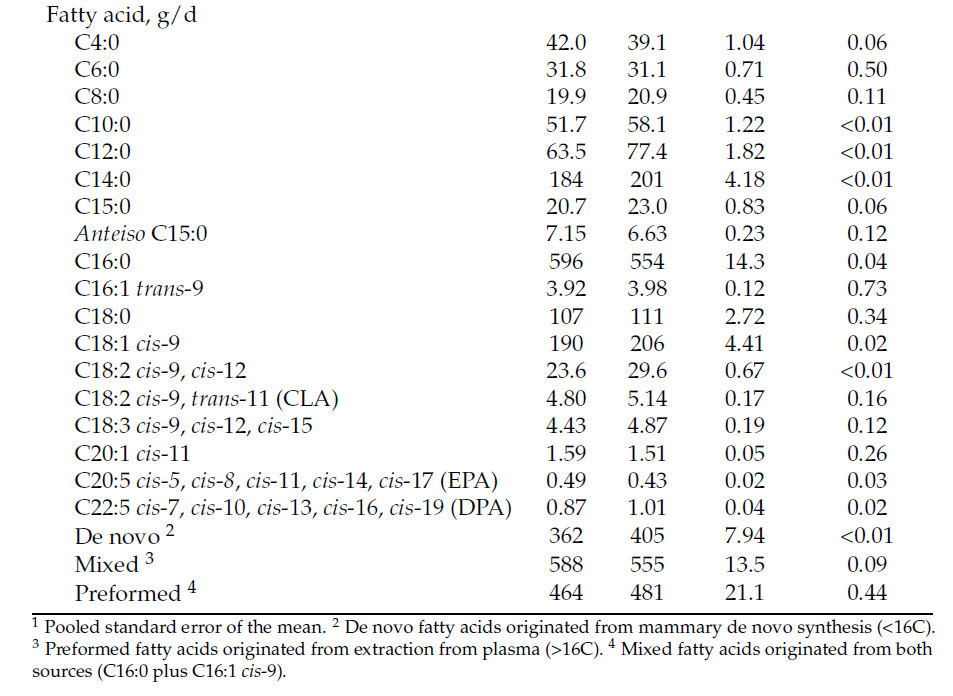
Таблица 4. Влияние концентрации крахмала в рационе на концентрацию жирных кислот в молоке (г/100 г молочного жира) и выход молока (г/д).
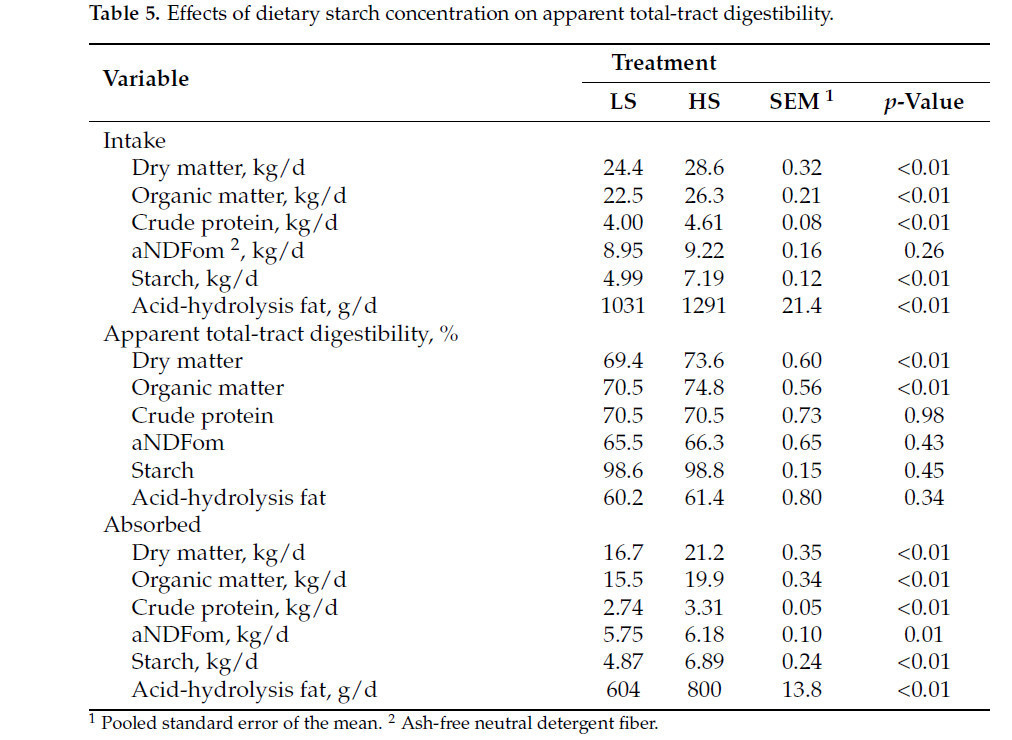
Таблица 5. Влияние концентрации пищевого крахмала на кажущуюся общую усвояемость.
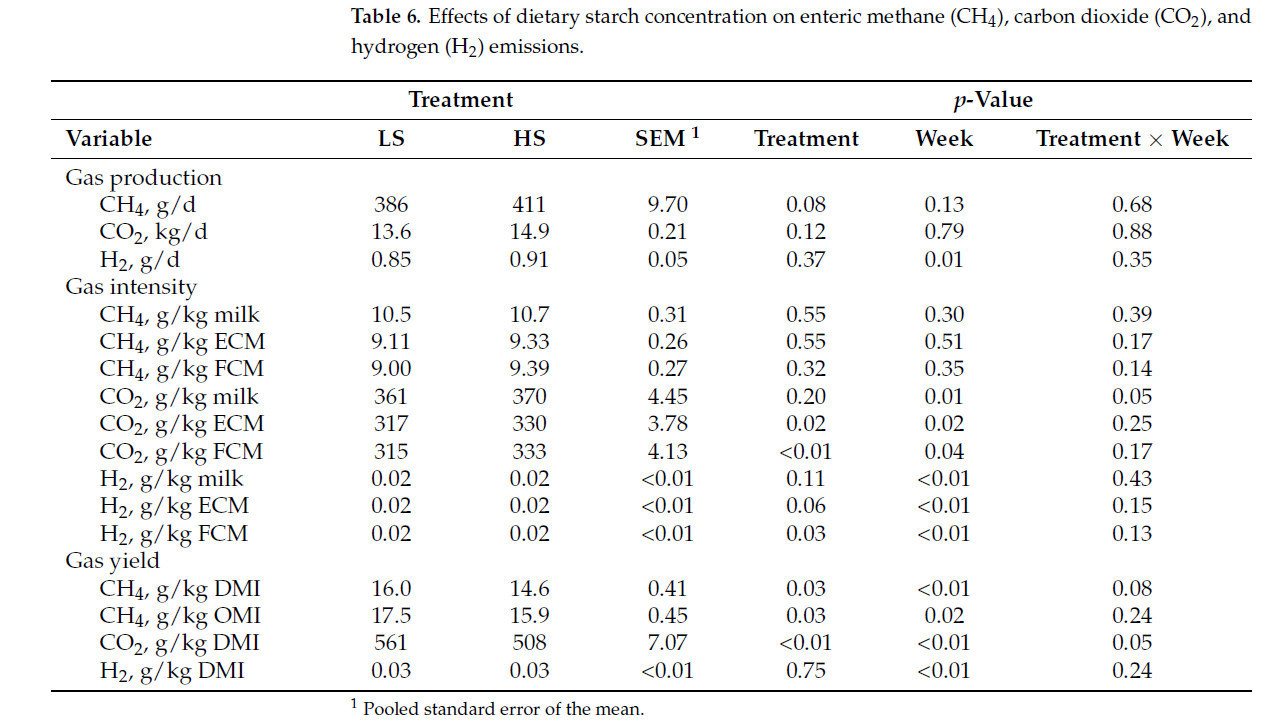
Таблица 6. Влияние концентрации пищевого крахмала на энтеральные выбросы метана (CH 4 ), углекислого газа (CO 2 ) и водорода (H 2 ).

Таблица 7. Корреляция Пирсона между концентрацией жирных кислот в молоке (г/100 г молочного жира) и продукцией CH4 .
4. Обсуждение
Выбросы CH4 в кишечнике вносят значительный вклад в воздействие молочной промышленности на окружающую среду. Исследования показали, что состав углеводов в рационе может влиять на характер брожения в рубце и метаногенез [ 36 ]. Увеличение доли крахмала в молочных рационах было предложено в качестве стратегии для снижения выбросов CH4 за счет стимуляции выработки пропионата в рубце [ 11 ]. Поскольку крахмал и клетчатка являются основными углеводными компонентами, понимание того, как различные уровни включения влияют на выработку CH4, стало критически важным направлением исследований. Целью данного исследования было изучение влияния концентрации крахмала в рационе на продуктивность молока, усвояемость питательных веществ и выбросы CH4 у дойных молочных коров.
Снижение DMI, наблюдаемое у коров, получавших рацион LS, вероятно, объясняется более высоким содержанием фуража (т. е. aNDFom) и более низкой долей концентрата. Фураж способствует большему физическому наполнению кишечника, что может подавлять DMI [ 37 , 38 ]. Следовательно, коровы LS потребляли на 4,2 кг меньше DM и производили на 2 кг меньше молока, чем коровы на рационе HS. По сравнению с другими компонентами TMR было продемонстрировано, что эффект физического наполнения более высокой концентрации aNDFom в корме представляет собой более значительное ограничение DMI по мере увеличения надоев молока [ 39 ]. Кроме того, высокопродуктивные коровы часто испытывают снижение молочной продуктивности при снижении концентрации крахмала в рационе [ 40 ]. Следовательно, замена концентратов фуражом в рационе LS снижала энергию, доступную как микробам рубца, так и животному-хозяину, что приводило к снижению молочной продуктивности у коров LS. Эффективность кормления у коров HS могла снизиться из-за более быстрой скорости прохождения крахмала. Рационы с высоким соотношением концентрата и фуража могут ускорить прохождение крахмала в тонкий кишечник, который имеет ограниченную способность переваривать большое количество крахмала. Это может привести к неэффективному пищеварению и снижению общей FE [ 41 , 42 ]. По мере увеличения производства молока улучшения FE обычно снижаются, отчасти из-за снижения усвояемой энергии, связанного с высокой скоростью прохождения [ 43 ]. И наоборот, более низкий DMI коррелирует с большей FE [ 44 ], а мобилизация тканей тела, как было показано, улучшает FE [ 45 ]. Отрицательный энергетический баланс у коров LS, возможно, способствовал наблюдаемому увеличению FE. Кроме того, более крупные коровы с более высокой BCS генетически предрасположены к более низкой FE [ 46 ], что согласуется с нашими результатами, поскольку у коров HS были более высокая BW и BCS.
Коровы, которых кормили диетой HS, имели более высокое содержание чистого белка в молоке и лактозы и больший выход чистого белка, чем коровы LS, что согласуется с предыдущими исследованиями [ 47 , 48 , 49 ]. Такая реакция на молочный белок, вероятно, связана с более высоким потреблением DM и CP у коров HS, что могло усилить синтез микробного белка и концентрацию пропионата в рубце [ 50 ]. Кроме того, коровы HS имели тенденцию к более высокой концентрации инсулина в плазме, который, как известно, влияет на синтез молочного белка [ 51 ]. Напротив, более низкое содержание крахмала в рационе LS могло снизить выработку микробного белка, ограничив доступный белковый пул для синтеза молочного белка у коров LS [ 52 ]. Тенденция к более высокому содержанию жира в молоке у коров LS по сравнению с коровами HS была ожидаема, поскольку рационы с низким содержанием aNDFom и высоким содержанием крахмала являются известными факторами риска снижения уровня жира в молоке [ 53 ]. Этот эффект можно объяснить улучшенной буферной емкостью рациона LS, который имел более высокую долю aNDFom. Эта буферизация помогает поддерживать более высокий pH в рубце, снижая частоту депрессии молочного жира [ 54 ]. Более низкое содержание молочного жира у коров HS может также быть результатом эффекта разбавления из-за их более высокой молочной продуктивности по сравнению с коровами LS. Кроме того, Рейнольдс и соавторы [ 55 ] связали снижение содержания молочного жира с повышенной концентрацией инсулина в плазме у коров, потребляющих рационы с высоким содержанием крахмала, поскольку инсулин снижает липолиз и способствует липогенезу в жировой ткани, снижая доступность жирных кислот для молочной железы.
Различия в рационе питания также повлияли на усвояемость питательных веществ. Крахмал обычно используется для повышения энергетической плотности рационов, усиления рубцовой ферментации и улучшения усвояемости OM. Более низкая кажущаяся общая усвояемость DM и OM в тракте коров, получавших рацион LS, может быть связана с заменой неволокнистых углеводов (в основном из кукурузного зерна в рационе HS) волокнистыми углеводами (в основном из кукурузного силоса и тритикалевого силоса), что снижает общую усвояемость питательных веществ. Эти результаты согласуются с данными Сильвестра и соавторов [ 56 ], которые сравнивали типичный крахмальный рацион с рационом с пониженным содержанием крахмала (24,8 против 18,4% крахмала). Усвояемость органического вещества, вероятно, была основным фактором, определяющим эффективность усвоения и использования коровами на рационе HS питательных веществ для производства молока. Хотя было обнаружено, что рационы, богатые крахмалом, снижают усвояемость клетчатки [ 57 , 58 ], в нашем исследовании этот эффект не наблюдался. Это может быть связано с относительно небольшой разницей в концентрации крахмала между рационами (5%) и тем фактом, что содержание aNDFom в обоих рационах превышало 32%, что, вероятно, позволяло поддерживать pH рубца достаточно высоким для поддержания активности целлюлозолитических бактерий. Аналогичным образом, исследование Сильвестра и соавторов [ 56 ], используя сопоставимые концентрации крахмала в рационе, не обнаружило существенной разницы в усвояемости aNDFom [ 56 ].
Следует отметить, что источник крахмала, тип зерна и степень обработки являются критическими факторами, влияющими на переваривание крахмала у молочных коров. В данном исследовании в рационе группы HS использовалась более тонкоизмельченная кукуруза, которая известна своей быстрой ферментацией в рубце благодаря увеличенной площади поверхности, что повышает усвояемость крахмала и синтез микробного белка. Такая обработка, вероятно, способствовала повышению энергетической продуктивности и усвояемости молока в группе HS. При сравнении результатов разных исследований важно учитывать различия в источнике и обработке крахмала, поскольку более грубый помол или использование альтернативных зерновых может привести к разной динамике ферментации и продуктивности.
Коровы, которых кормили рационами LS, имели более низкие концентрации C18:2 цис -9, цис -12 и de novo жирных кислот в молоке и более низкий выход de novo жирных кислот. Однако у них было более высокое содержание смешанных жирных кислот, чем у коров HS. Наблюдаемое снижение усвояемости DM и OM у коров, которых кормили рационами LS, могло ограничить доступность субстратов, необходимых для синтеза de novo жирных кислот в молочной железе. Молочные жирные кислоты имеют два различных источника: те, у которых менее 16 атомов углерода, производятся посредством синтеза de novo в молочной железе, в то время как те, у которых более 16 атомов углерода, получаются путем экстракции плазмы. Жирные кислоты, такие как C16:0 и C16:1 цис -9, поступают из смеси этих двух источников [ 59 ]. Учитывая значительное снижение концентраций de novo жирных кислот, увеличение смешанных жирных кислот у коров LS, вероятно, связано с большей мобилизацией жировых резервов организма в результате их более низкого потребления сухого вещества и отрицательного энергетического баланса. Более высокие концентрации и выходы C18:2 цис -9, цис -12 в молоке от коров HS, вероятно, были вызваны повышенным потреблением соевой муки, диетического источника линолевой кислоты [ 60 ].
При учете колебаний в потреблении, коровы на диете HS имели более низкий выход CH 4 относительно как DMI, так и OMI по сравнению с коровами на диете LS. Аналогичным образом, Aguerre et al. [ 15 ] сообщили о последовательном линейном снижении выхода CH 4 , до 19%, в диапазоне соотношений фуража к концентрату от 68:32 до 47:53. Вполне вероятно, что более высокий уровень крахмала в рационе привел к более эффективному пищеварению, что привело к более быстрому прохождению и меньшей степени ферментации в рубце. Аналогичным образом, Boadi и Wittenberg [ 61 ] продемонстрировали, что выбросы CH 4 на единицу OMI имеют тенденцию к снижению с повышением усвояемости рациона. Это согласуется с нашими выводами, поскольку более высокая усвояемость диеты HS привела к более низким выбросам CH 4 на переваренную единицу OMI по сравнению с диетой LS. Интересно, что Olijhoek et al. [ 62 ] наблюдали снижение выхода CH 4 на 27,2% и 13,8% у коров голштинской и джерсейской пород соответственно при увеличении доли концентратов в рационе с 32 до 61%. Это говорит о том, что увеличение количества концентратов, а следовательно, и крахмала, может быть более эффективной стратегией снижения уровня CH 4 для коров голштинской породы, чем для коров джерсейской породы.
Однако это не обязательно подразумевает снижение общего производства CH 4. У коров HS было больше общего DMI, что давало больше субстрата для микробной ферментации. Хотя выход CH 4 на единицу DMI и OMI был ниже у коров HS, коровы LS имели тенденцию производить меньше абсолютного CH 4 , выделяя на 25 г/д меньше. Этот результат был ожидаем, поскольку коровы LS потребляли на 4,2 кг меньше DM, чем коровы HS, что приводило к меньшему количеству субстратов, доступных для микробов рубца. Хорошо известно, что основным фактором метаногенеза является потребление корма сверх энергетических потребностей поддержания [ 63 , 64 ]. Исследования установили сильную положительную корреляцию между суточным производством CH 4 и потреблением рационов на основе кормов, независимо от уровней потребления или типа корма [ 65 ]. Таким образом, разница в 6,27% в суточном производстве CH4, наблюдаемая в настоящем исследовании, вероятно, обусловлена разницей в 15,9% в СВ, а не содержанием крахмала в рационе.
Включение более 35% концентрата в рацион молочных коров было связано с уменьшением производства CH 4 [ 66 ]. В текущем исследовании уровни концентрата составили 35,4 и 49,4% для рационов LS и HS соответственно. Сходное общее производство CH 4 , наблюдаемое в обеих группах, может быть отнесено к обоим рационам, превышающим этот порог. Муньос и др. [ 67 ] исследовали влияние двух уровней диетического концентрата (29 против 46% от сухого вещества рациона) на выбросы CH 4 у молочных коров и обнаружили, что в то время как более высокий уровень концентрата увеличил общее производство CH 4 на 10,7%, он снизил выход CH 4 на 12,7%. В соответствии с нашим исследованием, интенсивность CH 4 осталась неизменной. Напротив, Олийхук и др. [ 62 ] сравнили уровни концентрата 32 против 61% и сообщили, что более высокий уровень снизил производство CH 4 , интенсивность и выход. Разница в концентрации крахмала между их диетами составила 11,3%, что больше, чем разница между диетами LS и HS в нашем исследовании, что потенциально объясняет различные результаты.
Отрицательная корреляция, наблюдаемая между продукцией CH 4 и FA anteiso C15:0, C16:1 trans -9, C18:1 cis -9 и C20:1 cis -11, согласуется с результатами предыдущих исследований [ 19 , 68 , 69 ]. Эту связь можно объяснить ролью бактерий рубца в использовании H 2 для биогидрогенизации ненасыщенных ЖК. Поскольку H 2 потребляется в этом процессе, его становится меньше для гидрогенотрофных метаногенов, что снижает продукцию CH 4 [ 6 ]. Кроме того, ненасыщенные ЖК могут ингибировать метаногенез, оказывая токсическое воздействие на простейших и целлюлозолитические бактерии [ 70 ]. Аналогичным образом, anteiso C15:0, преимущественно продуцируемый амилолитическими бактериями [ 71 ], может способствовать повышенному потреблению H 2 за счет усиления продукции пропионата, что еще больше ограничивает доступность H 2 для метаногенеза.
Более существенное увеличение концентрации крахмала в рационе HS могло привести к снижению продукции CH 4 из-за увеличения продукции пропионата в рубце, который теоретически потребляет H 2 , ингибируя метаногенез. Кроме того, было показано, что рационы с высоким содержанием крахмала изменяют микробный состав рубца, благоприятствуя бактериям, продуцирующим пропионат [ 72 ]. Более низкий pH рубца в результате рациона, богатого крахмалом, также влияет на рост простейших, метаногенов и целлюлозолитических бактерий [ 73 ]. Однако значительное увеличение включения крахмала может снизить потребление сухого вещества (DMI), поскольку пропионат стимулирует окисление в печени, которое подает сигнал мозгу о сытости и уменьшает размер порции [ 74 ]. Если это происходит, наблюдаемое снижение продукции CH 4 можно было бы приписать снижению потребления сухого вещества (DMI), а не сдвигам в путях ферментации. Например, Zang et al. [ 75 ] обнаружили, что увеличение концентрации крахмала в рационе с 12,3 до 34,4% снижает потребление сухого вещества, что приводит к снижению выработки CH 4 на 20% .
Нацеливание на выход CH₄, а не на общую продуктивность или интенсивность, было предложено в качестве наиболее эффективного признака для разведения скота с низким уровнем выбросов. Сокращение выхода CH₄ может снизить отдельные выбросы за счет изменения функции рубца с минимальным влиянием на продуктивность или массу тела [ 76 ]. Однако, хотя снижение выхода CH4 полезно , необходима осторожность при использовании рационов с высоким содержанием крахмала, поскольку чрезмерное включение крахмала может отрицательно повлиять на продуктивность и усвояемость питательных веществ. Было показано, что концентрации крахмала от 28 до 32% снижают pH рубца, увеличивая риск подострого ацидоза рубца и потенциально ставя под угрозу здоровье и продуктивность животных [ 77 ]. Кроме того, необходимо учитывать экологические компромиссы, поскольку более высокие уровни диетических концентратов могут привести к увеличению потерь азота [ 78 , 79 ] и большему потреблению воды [ 80 ], что потенциально усугубляет будущие проблемы с водными ресурсами. Следовательно, балансировка крахмала с другими диетическими компонентами имеет важное значение для разработки эффективных и устойчивых стратегий кормления.
5. Выводы
Наши результаты показывают, что повышение концентрации крахмала в рационе может улучшить продуктивность молока и усвояемость рациона, одновременно снижая выход CH 4. Это имеет важные последствия для методов молочного животноводства, особенно в регионах с ограниченным доступом к передовым технологиям снижения выбросов CH 4 и кормовым добавкам. Будущие исследования должны быть сосредоточены на определении оптимальной концентрации крахмала, которая максимизирует энергетически скорректированный удой молока и минимизирует выход CH 4 без увеличения риска ацидоза рубца или других проблем со здоровьем у молочных пород. После того, как этот порог будет установлен, следует изучить влияние сезонности и климата на уровни крахмала, чтобы гарантировать глобальную применимость этой стратегии питания. Более того, изучение долгосрочных эффектов рационов с высоким содержанием крахмала на разных стадиях лактации имеет решающее значение для понимания их более широкого влияния на здоровье и продуктивность животных.
Дополнительные материалы
Следующая вспомогательная информация может быть загружена по адресу https://www.mdpi.com/article/10.3390/agriculture15020211/s1 , Таблица S1: Концентрация жирных кислот в молоке (г/100 г молочного жира) рационов LS и HS; Таблица S2: Выход жирных кислот в молоке (г/д) рационов LS и HS; Рисунок S1: Влияние концентрации диетического крахмала на надои молока; Рисунок S2: Влияние концентрации диетического крахмала на надои молока с поправкой на энергию; Рисунок S3: Влияние концентрации диетического крахмала на надои молока с поправкой на жир 3,5%; Рисунок S4: Влияние концентрации диетического крахмала на потребление сухого вещества; Рисунок S5: Влияние концентрации диетического крахмала на жвачку; Рисунок S6: Влияние концентрации диетического крахмала на содержание молочного белка; Рисунок S7: Влияние концентрации диетического крахмала на выход молочного жира; Рисунок S8: Влияние концентрации диетического крахмала на выход лактозы молока; Рисунок S9: Влияние концентрации крахмала в рационе на эффективность кормления (кг надоя молока/кг потребления сухого вещества); Рисунок S10: Влияние концентрации крахмала в рационе на эффективность кормления (кг надоя молока с поправкой на энергию/кг потребления сухого вещества); Рисунок S11: Влияние концентрации крахмала в рационе на эффективность кормления (кг надоя молока с поправкой на жирность 3,5%/кг потребления сухого вещества).
Ссылки
1. United Nations Department of Economic and Social Affairs, Population Division. World Population Prospects 2022: Summary of Results; United Nations: New York, NY, USA, 2022. [Google Scholar]
2. Steenson, S.; Buttriss, J.L. The Challenges of Defining a Healthy and ‘Sustainable’ Diet. Nutr. Bull. 2020, 45, 206–222. [Google Scholar] [CrossRef]
3. Intergovernmental Panel on Climate Change. Climate Change 2021—The Physical Science Basis: Working Group I Contribution to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change, 1st ed.; Cambridge University Press: Cambridge, UK, 2023; ISBN 978-1-00-915789-6. [Google Scholar]
4. Chagunda, M.G.G.; Römer, D.A.M.; Roberts, D.J. Effect of Genotype and Feeding Regime on Enteric Methane, Non-Milk Nitrogen and Performance of Dairy Cows during the Winter Feeding Period. Livest. Sci. 2009, 122, 323–332. [Google Scholar] [CrossRef]
5. Johnson, K.A.; Johnson, D.E. Methane Emissions from Cattle. J. Anim. Sci. 1995, 73, 2483–2492. [Google Scholar] [CrossRef] [PubMed]
6. Boadi, D.; Benchaar, C.; Chiquette, J.; Massé, D. Mitigation Strategies to Reduce Enteric Methane Emissions from Dairy Cows: Update Review. Can. J. Anim. Sci. 2004, 84, 319–335. [Google Scholar] [CrossRef]
7. Hristov, A.N.; Oh, J.; Firkins, J.L.; Dijkstra, J.; Kebreab, E.; Waghorn, G.; Makkar, H.P.S.; Adesogan, A.T.; Yang, W.; Lee, C.; et al. SPECIAL TOPICS—Mitigation of Methane and Nitrous Oxide Emissions from Animal Operations: I. A Review of Enteric Methane Mitigation Options1. J. Anim. Sci. 2013, 91, 5045–5069. [Google Scholar] [CrossRef]
8. Zubieta, Á.S.; Savian, J.V.; De Souza Filho, W.; Wallau, M.O.; Gómez, A.M.; Bindelle, J.; Bonnet, O.J.F.; De Faccio Carvalho, P.C. Does Grazing Management Provide Opportunities to Mitigate Methane Emissions by Ruminants in Pastoral Ecosystems? Sci. Total Environ. 2021, 754, 142029. [Google Scholar] [CrossRef] [PubMed]
9. Koenig, K.M.; Beauchemin, K.A.; Rode, L.M. Effect of Grain Processing and Silage on Microbial Protein Synthesis and Nutrient Digestibility in Beef Cattle Fed Barley-Based Diets1,2. J. Anim. Sci. 2003, 81, 1057–1067. [Google Scholar] [CrossRef] [PubMed]
10. Knowlton, K.F. High Grain Diets for Dairy Cattle; University of New England: Armidale, Australia, 2001; ISBN 1-86389-743-7. [Google Scholar]
11. Bannink, A.; Kogut, J.; Dijkstra, J.; France, J.; Kebreab, E.; Van Vuuren, A.M.; Tamminga, S. Estimation of the Stoichiometry of Volatile Fatty Acid Production in the Rumen of Lactating Cows. J. Theor. Biol. 2006, 238, 36–51. [Google Scholar] [CrossRef] [PubMed]
12. Murphy, M.R.; Baldwin, R.L.; Koong, L.J. Estimation of Stoichiometric Parameters for Rumen Fermentation of Roughage and Concentrate Diets. J. Anim. Sci. 1982, 55, 411–421. [Google Scholar] [CrossRef]
13. Van Kessel, J. The Effect of pH on Ruminal Methanogenesis. FEMS Microbiol. Ecol. 1996, 20, 205–210. [Google Scholar] [CrossRef]
14. Dijkstra, J.; Oenema, O.; Bannink, A. Dietary Strategies to Reducing N Excretion from Cattle: Implications for Methane Emissions. Curr. Opin. Environ. Sustain. 2011, 3, 414–422. [Google Scholar] [CrossRef]
15. Aguerre, M.J.; Wattiaux, M.A.; Powell, J.M.; Broderick, G.A.; Arndt, C. Effect of Forage-to-Concentrate Ratio in Dairy Cow Diets on Emission of Methane, Carbon Dioxide, and Ammonia, Lactation Performance, and Manure Excretion. J. Dairy Sci. 2011, 94, 3081–3093. [Google Scholar] [CrossRef] [PubMed]
16. Pirondini, M.; Colombini, S.; Mele, M.; Malagutti, L.; Rapetti, L.; Galassi, G.; Crovetto, G.M. Effect of Dietary Starch Concentration and Fish Oil Supplementation on Milk Yield and Composition, Diet Digestibility, and Methane Emissions in Lactating Dairy Cows. J. Dairy Sci. 2015, 98, 357–372. [Google Scholar] [CrossRef] [PubMed]
17. Hatew, B.; Podesta, S.C.; Van Laar, H.; Pellikaan, W.F.; Ellis, J.L.; Dijkstra, J.; Bannink, A. Effects of Dietary Starch Content and Rate of Fermentation on Methane Production in Lactating Dairy Cows. J. Dairy Sci. 2015, 98, 486–499. [Google Scholar] [CrossRef]
18. Negussie, E.; De Haas, Y.; Dehareng, F.; Dewhurst, R.J.; Dijkstra, J.; Gengler, N.; Morgavi, D.P.; Soyeurt, H.; Van Gastelen, S.; Yan, T.; et al. Invited Review: Large-Scale Indirect Measurements for Enteric Methane Emissions in Dairy Cattle: A Review of Proxies and Their Potential for Use in Management and Breeding Decisions. J. Dairy Sci. 2017, 100, 2433–2453. [Google Scholar] [CrossRef]
19. Dijkstra, J.; Van Zijderveld, S.M.; Apajalahti, J.A.; Bannink, A.; Gerrits, W.J.J.; Newbold, J.R.; Perdok, H.B.; Berends, H. Relationships between Methane Production and Milk Fatty Acid Profiles in Dairy Cattle. Anim. Feed Sci. Technol. 2011, 166–167, 590–595. [Google Scholar] [CrossRef]
20. Rico, D.E.; Chouinard, P.Y.; Hassanat, F.; Benchaar, C.; Gervais, R. Prediction of Enteric Methane Emissions from Holstein Dairy Cows Fed Various Forage Sources. Animal 2016, 10, 203–211. [Google Scholar] [CrossRef] [PubMed]
21. Wildman, E.E.; Jones, G.M.; Wagner, P.E.; Boman, R.L.; Troutt, H.F.; Lesch, T.N. A Dairy Cow Body Condition Scoring System and Its Relationship to Selected Production Characteristics. J. Dairy Sci. 1982, 65, 495–501. [Google Scholar] [CrossRef]
22. Schirmann, K.; Von Keyserlingk, M.A.G.; Weary, D.M.; Veira, D.M.; Heuwieser, W. Technical Note: Validation of a System for Monitoring Rumination in Dairy Cows. J. Dairy Sci. 2009, 92, 6052–6055. [Google Scholar] [CrossRef] [PubMed]
23. AOAC International. Official Methods of Analysis of AOAC International, 17th ed.; Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2000. [Google Scholar]
24. Hall, M.B. Determination of Starch, Including Maltooligosaccharides, in Animal Feeds: Comparison of Methods and a Method Recommended for AOAC Collaborative Study. J. AOAC Int. 2009, 92, 42–49. [Google Scholar] [CrossRef] [PubMed]
25. Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
26. Goering, H.K.; Van Soest, P.J. Forage Fiber Analysis. Agricultural Handbook No. 379; US Department of Agriculture: Washington, DC, USA, 1970; pp. 1–20. [Google Scholar]
27. Heinrichs, J.; Jones, C.M. Penn State Particle Separator; Penn State University: State College, PA, USA, 2022; Volume 2022, Available online: https://extension.psu.edu/penn-state-particle-separator (accessed on 5 May 2024).
28. Lock, A.L.; Preseault, C.L.; Rico, J.E.; DeLand, K.E.; Allen, M.S. Feeding a C16:0-Enriched Fat Supplement Increased the Yield of Milk Fat and Improved Conversion of Feed to Milk. J. Dairy Sci. 2013, 96, 6650–6659. [Google Scholar] [CrossRef] [PubMed]
29. Duplessis, M.; Gervais, R.; Lapierre, H.; Girard, C.L. Combined Biotin, Folic Acid, and Vitamin B12 Supplementation given during the Transition Period to Dairy Cows: Part II. Effects on Energy Balance and Fatty Acid Composition of Colostrum and Milk. J. Dairy Sci. 2022, 105, 7097–7110. [Google Scholar] [CrossRef] [PubMed]
30. Ulberth, F.; Schrammel, F. Accurate Quantitation of Short-, Medium-, and Long-Chain Fatty Acid Methyl Esters by Split-Injection Capillary Gas-Liquid Chromatography. J. Chromatogr. A 1995, 704, 455–463. [Google Scholar] [CrossRef]
31. Schauff, D.J.; Clark, J.H.; Drackley, J.K. Effects of Feeding Lactating Dairy Cows Diets Containing Extruded Soybeans and Calcium Salts of Long-Chain Fatty Acids. J. Dairy Sci. 1992, 75, 3003–3019. [Google Scholar] [CrossRef]
32. Martin, M.J.; Weigel, K.A.; White, H.M. Assessment of the Relationship between Postpartum Health and Mid-Lactation Performance, Behavior, and Feed Efficiency in Holstein Dairy Cows. Animals 2021, 11, 1385. [Google Scholar] [CrossRef] [PubMed]
33. Ali, A.K.A.; Shook, G.E. An Optimum Transformation for Somatic Cell Concentration in Milk. J. Dairy Sci. 1980, 63, 487–490. [Google Scholar] [CrossRef]
34. Hisadomi, S.; Haruno, A.; Fujieda, T.; Sugino, T.; Oba, M. Effects of Rumen-Protected Glutamate Supplementation during the Periparturient Period on Digestibility, Inflammation, Metabolic Responses, and Performance in Dairy Cows. J. Dairy Sci. 2022, 105, 3129–3141. [Google Scholar] [CrossRef] [PubMed]
35. Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Stat. Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
36. Jentsch, W.; Schweigel, M.; Weissbach, F.; Scholze, H.; Pitroff, W.; Derno, M. Methane Production in Cattle Calculated by the Nutrient Composition of the Diet. Arch. Anim. Nutr. 2007, 61, 10–19. [Google Scholar] [CrossRef]
37. Forbes, J.M. Voluntary Food Intake and Diet Selection in Farm Animals, 2nd ed.; CABI: Egham, UK, 2007; ISBN 978-1-84593-279-4. [Google Scholar]
38. Mertens, D.R. Forage Quality, Evaluation, and Utilization; American Society of Agronomy Inc., Crop Science Society of America Inc., Soil Science Society of America Inc.: Madison, WI, USA, 1994. [Google Scholar]
39. Allen, M.S. Effects of Diet on Short-Term Regulation of Feed Intake by Lactating Dairy Cattle. J. Dairy Sci. 2000, 83, 1598–1624. [Google Scholar] [CrossRef] [PubMed]
40. Boerman, J.P.; Potts, S.B.; VandeHaar, M.J.; Allen, M.S.; Lock, A.L. Milk Production Responses to a Change in Dietary Starch Concentration Vary by Production Level in Dairy Cattle. J. Dairy Sci. 2015, 98, 4698–4706. [Google Scholar] [CrossRef]
41. Mills, J.; France, J.; Dijkstra, J. A Review of Starch Digestion in the Lactating Dairycow and Proposals for a Mechanistic Model: 2.Postruminal Starch Digestion and Small Intestinalglucose Absorption. J. Anim. Feed Sci. 1999, 8, 451–481. [Google Scholar] [CrossRef]
42. Reynolds, C.K. Production and Metabolic Effects of Site of Starch Digestion in Dairy Cattle. Anim. Feed Sci. Technol. 2006, 130, 78–94. [Google Scholar] [CrossRef]
43. Nutrient Requirements of Dairy Cattle, Seventh revised ed.; National Research Council, Ed.; Nutrient requirements of domestic animals; National Academy Press: Washington, DC, USA, 2001; ISBN 978-0-309-06997-7. [Google Scholar]
44. Ranathunga, S.D.; Kalscheur, K.F.; Hippen, A.R.; Schingoethe, D.J. Replacement of Starch from Corn with Nonforage Fiber from Distillers Grains and Soyhulls in Diets of Lactating Dairy Cows. J. Dairy Sci. 2010, 93, 1086–1097. [Google Scholar] [CrossRef] [PubMed]
45. Vallimont, J.E.; Dechow, C.D.; Daubert, J.M.; Dekleva, M.W.; Blum, J.W.; Barlieb, C.M.; Liu, W.; Varga, G.A.; Heinrichs, A.J.; Baumrucker, C.R. Short Communication: Heritability of Gross Feed Efficiency and Associations with Yield, Intake, Residual Intake, Body Weight, and Body Condition Score in 11 Commercial Pennsylvania Tie Stalls. J. Dairy Sci. 2011, 94, 2108–2113. [Google Scholar] [CrossRef]
46. Arndt, C.; Powell, J.M.; Aguerre, M.J.; Crump, P.M.; Wattiaux, M.A. Feed Conversion Efficiency in Dairy Cows: Repeatability, Variation in Digestion and Metabolism of Energy and Nitrogen, and Ruminal Methanogens. J. Dairy Sci. 2015, 98, 3938–3950. [Google Scholar] [CrossRef] [PubMed]
47. Dias, A.L.G.; Freitas, J.A.; Micai, B.; Azevedo, R.A.; Greco, L.F.; Santos, J.E.P. Effects of Supplementing Yeast Culture to Diets Differing in Starch Content on Performance and Feeding Behavior of Dairy Cows. J. Dairy Sci. 2018, 101, 186–200. [Google Scholar] [CrossRef]
48. Poorkasegaran, S.; Yansari, A.T. Effects of Different Sources of Carbohydrates on Intake, Digestibility, Chewing, and Performance of Holstein Dairy Cows. J. Anim. Sci. Biotechnol. 2014, 5, 6. [Google Scholar] [CrossRef]
49. Shahmoradi, A.; Alikhani, M.; Riasi, A.; Ghorbani, G.R.; Ghaffari, M.H. Effects of Partial Replacement of Barley Grain with Beet Pulp on Performance, Ruminal Fermentation and Plasma Concentration of Metabolites in Transition Dairy Cows. Anim. Physiol. Nutr. 2016, 100, 178–188. [Google Scholar] [CrossRef] [PubMed]
50. Oba, M.; Allen, M.S. Effects of Diet Fermentability on Efficiency of Microbial Nitrogen Production in Lactating Dairy Cows. J. Dairy Sci. 2003, 86, 195–207. [Google Scholar] [CrossRef]
51. Arriola Apelo, S.I.; Knapp, J.R.; Hanigan, M.D. Invited Review: Current Representation and Future Trends of Predicting Amino Acid Utilization in the Lactating Dairy Cow. J. Dairy Sci. 2014, 97, 4000–4017. [Google Scholar] [CrossRef] [PubMed]
52. Weiss, W.P.; Pinos-Rodríguez, J.M.; Wyatt, D.J. The Value of Different Fat Supplements as Sources of Digestible Energy for Lactating Dairy Cows. J. Dairy Sci. 2011, 94, 931–939. [Google Scholar] [CrossRef] [PubMed]
53. Bauman, D.E.; Harvatine, K.J.; Lock, A.L. Nutrigenomics, Rumen-Derived Bioactive Fatty Acids, and the Regulation of Milk Fat Synthesis. Annu. Rev. Nutr. 2011, 31, 299–319. [Google Scholar] [CrossRef] [PubMed]
54. Martel, C.A.; Titgemeyer, E.C.; Mamedova, L.K.; Bradford, B.J. Dietary Molasses Increases Ruminal pH and Enhances Ruminal Biohydrogenation during Milk Fat Depression. J. Dairy Sci. 2011, 94, 3995–4004. [Google Scholar] [CrossRef] [PubMed]
55. Reynolds, C.K.; Sutton, J.D.; Beever, D.E. Effects of Feeding Starch to Dairy Cattle on Nutrient Availability and Production; Nottingham University Press: Nottingham, UK, 1997; ISBN 1-897676-03-4. [Google Scholar]
56. Silvestre, T.; Fetter, M.; Räisänen, S.E.; Lage, C.F.A.; Stefenoni, H.; Melgar, A.; Cueva, S.F.; Wasson, D.E.; Martins, L.F.; Karnezos, T.P.; et al. Performance of Dairy Cows Fed Normal- or Reduced-Starch Diets Supplemented with an Exogenous Enzyme Preparation. J. Dairy Sci. 2022, 105, 2288–2300. [Google Scholar] [CrossRef]
57. Boerman, J.P.; Potts, S.B.; VandeHaar, M.J.; Lock, A.L. Effects of Partly Replacing Dietary Starch with Fiber and Fat on Milk Production and Energy Partitioning. J. Dairy Sci. 2015, 98, 7264–7276. [Google Scholar] [CrossRef]
58. Ferraretto, L.F.; Crump, P.M.; Shaver, R.D. Effect of Cereal Grain Type and Corn Grain Harvesting and Processing Methods on Intake, Digestion, and Milk Production by Dairy Cows through a Meta-Analysis. J. Dairy Sci. 2013, 96, 533–550. [Google Scholar] [CrossRef] [PubMed]
59. Dos Santos Neto, J.M.; De Souza, J.; Lock, A.L. Nutrient Digetibility and Production Responses of Lactating Dairy Cows When Saturated Free Fatty Acid Supplements Are Included in Diets: A Meta-Analysis. J. Dairy Sci. 2021, 104, 12628–12646. [Google Scholar] [CrossRef]
60. AbuGhazaleh, A.A.; Schingoethe, D.J.; Hippen, A.R.; Kalscheur, K.F. Conjugated Linoleic Acid Increases in Milk When Cows Fed Fish Meal and Extruded Soybeans for an Extended Period of Time. J. Dairy Sci. 2004, 87, 1758–1766. [Google Scholar] [CrossRef] [PubMed]
61. Boadi, D.A.; Wittenberg, K.M. Methane Production from Dairy and Beef Heifers Fed Forages Differing in Nutrient Density Using the Sulphur Hexafluoride (SF6) Tracer Gas Technique. Can. J. Anim. Sci. 2002, 82, 201–206. [Google Scholar] [CrossRef]
62. Olijhoek, D.W.; Løvendahl, P.; Lassen, J.; Hellwing, A.L.F.; Höglund, J.K.; Weisbjerg, M.R.; Noel, S.J.; McLean, F.; Højberg, O.; Lund, P. Methane Production, Rumen Fermentation, and Diet Digestibility of Holstein and Jersey Dairy Cows Being Divergent in Residual Feed Intake and Fed at 2 Forage-to-Concentrate Ratios. J. Dairy Sci. 2018, 101, 9926–9940. [Google Scholar] [CrossRef] [PubMed]
63. Ramin, M.; Huhtanen, P. Development of Equations for Predicting Methane Emissions from Ruminants. J. Dairy Sci. 2013, 96, 2476–2493. [Google Scholar] [CrossRef] [PubMed]
64. Atmospheric Methane: Sources, Sinks, and Role in Global Change; Khalil, M.A.K., North Atlantic Treaty Organization, Eds.; NATO ASI series; Springer: Berlin, Germany; New York, NY, USA, 1993; ISBN 978-3-540-54584-2. [Google Scholar]
65. Charmley, E.; Williams, S.R.O.; Moate, P.J.; Hegarty, R.S.; Herd, R.M.; Oddy, V.H.; Reyenga, P.; Staunton, K.M.; Anderson, A.; Hannah, M.C. A Universal Equation to Predict Methane Production of Forage-Fed Cattle in Australia. Anim. Prod. Sci. 2016, 56, 169. [Google Scholar] [CrossRef]
66. Sauvant, D.; Giger-Reverdin, S. Modélisation Des Interactions Digestives et de La Production de Méthane Chez Les Ruminants. INRA Prod. Anim. 2009, 22, 375–384. [Google Scholar] [CrossRef]
67. Muñoz, C.; Herrera, D.; Hube, S.; Morales, J.; Ungerfeld, E.M. Effects of Dietary Concentrate Supplementation on Enteric Methane Emissions and Performance of Late Lactation Dairy Cows. Chil. J. Agric. Res. 2018, 78, 429–437. [Google Scholar] [CrossRef]
68. Chilliard, Y.; Martin, C.; Rouel, J.; Doreau, M. Milk Fatty Acids in Dairy Cows Fed Whole Crude Linseed, Extruded Linseed, or Linseed Oil, and Their Relationship with Methane Output. J. Dairy Sci. 2009, 92, 5199–5211. [Google Scholar] [CrossRef] [PubMed]
69. De Souza, J.; Leskinen, H.; Lock, A.L.; Shingfield, K.J.; Huhtanen, P. Between-Cow Variation in Milk Fatty Acids Associated with Methane Production. PLoS ONE 2020, 15, e0235357. [Google Scholar] [CrossRef] [PubMed]
70. Doreau, M.; Ferlay, A. Effect of Dietary Lipids on Nitrogen Metabolism in the Rumen: A Review. Livest. Prod. Sci. 1995, 43, 97–110. [Google Scholar] [CrossRef]
71. Fievez, V.; Colman, E.; Castro-Montoya, J.M.; Stefanov, I.; Vlaeminck, B. Milk Odd- and Branched-Chain Fatty Acids as Biomarkers of Rumen Function—An Update. Anim. Feed Sci. Technol. 2012, 172, 51–65. [Google Scholar] [CrossRef]
72. Ellis, J.L.; Dijkstra, J.; Kebreab, E.; Bannink, A.; Odongo, N.E.; McBRIDE, B.W.; France, J. Aspects of Rumen Microbiology Central to Mechanistic Modelling of Methane Production in Cattle. J. Agric. Sci. 2008, 146, 213–233. [Google Scholar] [CrossRef]
73. Hook, S.E.; Steele, M.A.; Northwood, K.S.; Wright, A.-D.G.; McBride, B.W. Impact of High-Concentrate Feeding and Low Ruminal pH on Methanogens and Protozoa in the Rumen of Dairy Cows. Microb. Ecol. 2011, 62, 94–105. [Google Scholar] [CrossRef]
74. Allen, M.S.; Bradford, B.J.; Oba, M. BOARD-INVITED REVIEW: The Hepatic Oxidation Theory of the Control of Feed Intake and Its Application to Ruminants. J. Anim. Sci. 2009, 87, 3317–3334. [Google Scholar] [CrossRef] [PubMed]
75. Zang, Y.; Silva, L.H.P.; Geng, Y.C.; Ghelichkhan, M.; Whitehouse, N.L.; Miura, M.; Brito, A.F. Dietary Starch Level and Rumen-Protected Methionine, Lysine, and Histidine: Effects on Milk Yield, Nitrogen, and Energy Utilization in Dairy Cows Fed Diets Low in Metabolizable Protein. J. Dairy Sci. 2021, 104, 9784–9800. [Google Scholar] [CrossRef]
76. Fresco, S.; Boichard, D.; Fritz, S.; Lefebvre, R.; Barbey, S.; Gaborit, M.; Martin, P. Comparison of Methane Production, Intensity, and Yield throughout Lactation in Holstein Cows. J. Dairy Sci. 2023, 106, 4147–4157. [Google Scholar] [CrossRef]
77. Khafipour, E.; Krause, D.O.; Plaizier, J.C. A Grain-Based Subacute Ruminal Acidosis Challenge Causes Translocation of Lipopolysaccharide and Triggers Inflammation. J. Dairy Sci. 2009, 92, 1060–1070. [Google Scholar] [CrossRef] [PubMed]
78. Condren, S.A.; Kelly, A.K.; Lynch, M.B.; Boland, T.M.; Whelan, S.J.; Grace, C.; Rajauria, G.; Pierce, K.M. The Effect of By-Product Inclusion and Concentrate Feeding Rate on Milk Production and Composition, Pasture Dry Matter Intake, and Nitrogen Excretion of Mid-Late Lactation Spring-Calving Cows Grazing a Perennial Ryegrass-Based Pasture. J. Dairy Sci. 2019, 102, 1247–1256. [Google Scholar] [CrossRef] [PubMed]
79. Molossi, L.; Hoshide, A.K.; Pedrosa, L.M.; Oliveira, A.S.D.; Abreu, D.C.D. Improve Pasture or Feed Grain? Greenhouse Gas Emissions, Profitability, and Resource Use for Nelore Beef Cattle in Brazil’s Cerrado and Amazon Biomes. Animals 2020, 10, 1386. [Google Scholar] [CrossRef] [PubMed]
80. Pereira, C.H.; Patino, H.O.; Hoshide, A.K.; Abreu, D.C.; Alan Rotz, C.; Nabinger, C. Grazing Supplementation and Crop Diversification Benefits for Southern Brazil Beef: A Case Study. Agric. Syst. 2018, 162, 1–9. [Google Scholar] [CrossRef]

Culbertson RL, Gutiérrez-Oviedo FA, Uzun P, Seneviratne N, Fontoura ABP, Yau BK, Judge JL, Davis AN, Reyes DC, McFadden JW. Effects of Dietary Starch Concentration on Milk Production, Nutrient Digestibility, and Methane Emissions in Mid-Lactation Dairy Cows. Agriculture. 2025; 15(2):211. https://doi.org/10.3390/agriculture15020211
Перевод статьи «Effects of Dietary Starch Concentration on Milk Production, Nutrient Digestibility, and Methane Emissions in Mid-Lactation Dairy Cows» авторов Culbertson RL, Gutiérrez-Oviedo FA, Uzun P, Seneviratne N, Fontoura ABP, Yau BK, Judge JL, Davis AN, Reyes DC, McFadden JW., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык

















































































































Комментарии (0)