Дәріс №2. Түйелер популяциясындағы көбею үдерістері түрлеріне байланысты генетикалық құрылымдары
1. Панмиксиялық популяция теориясы
2. Популяциялық генетикадағы негізгі эволюциялық факторлар
3. Популяциялардағы қарапайым эволюциялық факторлар
Популяция құрамындағы даралардың жалпы гендік жиынытығын –генофонд деп атайды. Генофонд – аллельдер мен генотиптердің жиілігін сипаттайды. Аллель жиілігі – бұл белгілі геннің аллельдердер жиынтығындағы алатын үлесі. Харди- Вайнберг заңына сәйкес, ұрпақтан-ұрпаққа берілген кезде популяциадағы аллельдердің салыстырмалы жиліктері өзгеріссіз қалады, яғни популяциялар ішінде кездейсоқ будандасулар жүріп, сыртқы факторлардың әсерлері болмаған жағдайда, популяциадағы аллельдер жиілігі тұрақты деңгейде сақталады.
Популяциалардағы генотиптер мен аллельдер жиіліктері арсындағы математикалық тәуелділікті 1908 жылы, бір-біріне байланыссыз ағылшын математигі Г.Н.Харди (Hardy, 1908; «Адамдағы тұқым қуалауды дәлелдеу туралы») және неміс дәрігері В. Вайнберг (Weinberg, 1908; «Араласу жағдайындағы популяциядағы мендель қатынастары») тапқан болатын. Осы тәуелділікті Харди-Вайнберг тепе-теңдігі деп атайды және ол төмендегіше тұжырылымдалады: "Белгілі популяциадағы доминатты және рецессивті аллельдер жилігі ұрпақтан-ұрпаққа берілу барысында төмендегі жағдайлар сақталса, тұрақты болып қала береді: популяциалар көлемі өте үлкен; шағылысулар кездейсоқ өтеді; жаңа мутациялар пайда болмайды; барлық генотиптер біріңғай фертильді, яғни сұрыпталу жүзеге аспайды; ұрпақтары арасында өзара шағылысулар жүрмейді; эмиграция да, иммиграция да болмайды, яғни берілген популяция мен басқа популяциялар арасында гендер ағысы жүзеге аспайды". Алынған нәтиже «Харди-Вайнберг заңы» аталады. Оны төмендегідей сипаттаймыз.
Аутосомды диаллельді ген (А және а) арқылы тұқым қуалайтын, мейоз үдерісі қалыпты жағдайда өтетін айқас шағылысатын диплоидты организмдердің гипотетикалық популяциясын қарастырайық. Бұл популяцияда төмендегі шарттар орындалады делік:
(1) мутациялық үдеріс жүрмейді;
(2) сұрыптау қысымы орын алады;
(3) популяция саны шексіз үлкен;
(4) ол басқа популяциялардан түгелдей оқшауланған және мұнда панмиксия (даралардың генотипі мен фенотипіне байланыссыз ата-аналық жұптардың кездейсоқ қалыптасуы) орын алады.
Ата-ана популяциясы (нөльдік ұрпақ) кездейсоқ қалыптасады, және жалпы жағдайда ата мен аналарда түрлі деңгейдегі АА, Аа және аа генотиптер жиіліктері арқылы беріледі. Мұндай популяцияда А және а гендердің жиіліктері ұрпақ тізбегінде өзгермейді. Бұл құбылыс популяция беталысын зерттеу кезінде ұсталынатын, жоғарыда көрсетілген шарттар мен тұқым қуалайтын факторлар дискреттігінен шығады. Алайда, генотиптер жиілігінің уақыттағы динамикасы белгісіз болып қалады. Бұл өте маңызды мәселе, себебі тек жоғарыда айтылған қарапайым жағдайда тұрақты генетикалық жүйе ғана эволюциялық мәнге ие болады. Тұрақтылық жағдайының болуы қарапайым жолмен дәлелденеді (Feller, 1957). Аутосомды ген қарастырылғандықтан, генотиптер жиілігі бірінші ұрпақтағы аталықтар мен аналықтардың өзінде бірдей болады. Екінші ұрпақта пайда болған гендер жиіліктері келесі ұрпақтар тізбегінде өзгермейді. Ал осы жағдайда Харди популяциясында ген мен генотиптер жиіліктері араларында қарапайым қатынастар орын алады:
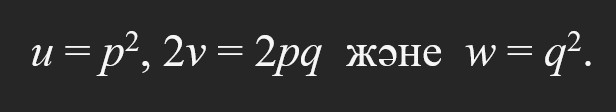
В. Вайнберг (1908) аутосом генде екі аллель емес, көп аллельдердің сериясы болса да генотиптер жиілігінің тепе-теңдігі орын алатынын көрсетті. Ал екі бөлек, мысалы, диаллельді А, а және В, b гендер арқылы тұқым қуалайтын популяцияның беталысы басқа болатынын болжады. Шынымен, жоғарыда көрсетілген қарапайым жағдайдың өзінде генотип құрылымы өте маңызды рөл атқаратыны кейінірек дәлелденді. Мысалы, екі диаллельді тіркелген, немесе тіркелмеген аутосом гендер үшін тепе-теңдік қалыптасу үшін белгілі бастапқы жағдайлар қажет. Жалпы жағдайда тіркелген немесе тіркелмеген гендер үшін тепе-теңдік асимптотикалық қалде орын алады және тіркелу күші төмен болған сайын үдей түседі (Neyman, 1950). Осы сияқты, егер тұқым қуалау жыныспен тіркелген генмен анықталса және екі жыныстың да гендерінің бастапқы жиіліктері әр түрлі мәнде болса ғана, популяция тек шектік жағдайда тепе-теңдікке ұмтылады (Feller, 1957).
Харди- Вайнберг теңдеуінде, популяциадағы рецессивті аллельдердің басым бөлігі гетерозиготалы тасымалдаушыларда болатыны байқалады. Гетерозиготалы геноптиптер генетикалық өзгергіштіктің нақты қоры болып табылады. Бұл, өз кезегінде, әрбір ұрпақта рецессивті аллельдердің тек өте аз бөлігінің ғана популяциядан жойылуына (элиминация) алып келеді. Тек гомозиготалық жағдайда болған кейбір рецессивті алльдер ғана фенотипте көрініс табады және нәтижесінде ол орта факторларының селекциялау әрекетіне ұшырап, жойылып отырылады. Рецесивті аллельдердің көпшілігі, әдетте финотип үшін қолайсыз әрекет етуі себепті жойылады. Мұндай элиминациялану тірілей ағзаның тұқым қалдырып үлгермей жатып ақ жоғалуы нәтижесінде жүзеге асады, яғни "генетикалық өлімнің" жүзеге асуы себепті, көбею мүмкіндігінің жоқтығы орын алады.
Табиғи жағдайларды қаншалықты қарапайымдатса да, Харди- Вайнберг заңының маңызы өте зор. Ол қарама-қарсы ұрықтанатын ағзалар популяцияларында панмиксияның өте кең таралуына байланысты. Панмиксия жағдайында, мутантты генге қарсы сұрыптаудың қатаң қысымына қарамастан, ұрпақтан-ұрпаққа ауысқанда алдыңғы ұрпақтың гендер жиіліктері мен келесі ұрпақтың басындағы гендер жиіліктері арасында жоғарыда көрсетілген (4) қатынастар орын алады.
Харди-Вайнберг заңын тереңірек түсіну үшін жаңа пайда болған мутациялардың басым бөлігі популяцияда кеңінен таралған «жабайы» аллельге қатысты рецессивті болатынын ескеру керек. Сондықтан панмиксия кезінде сұрыптау мутант гендердің тек гомозиготаларда кездесетін аа (q2), аз үлесін ғана тексере алады. Ал сонымен қоса, мутант гендердің басым бөлігі сұрыптау әсерінен фенотип бойынша доминантты «жабайы» гомозиготамен бірдей гетерозиготаларда (2pq) қорғалып қалады. Сондықтан, осы заң нақты популяциялардағы ген мен гетерозиготалардың жиіліктерін бағалауға кеңінен қолданады. Әсіресе бұл медициналық генетикада маңызды. Мысалы, егер рецессив аутосом ген тудыратын әйтеуір бір аурудың кездесу жиілігі 10-6 тең болса, онда қатерлі геннің және табуға қажет гетерозиготалардың жиіліктері, шамамен, 10-3 болады.
Жоғарыда сипатталған «харди» тепе-теңдігі тек идеалды популяцияда орын алады. Сондықтан, ылғи да шектеулі саны бар табиғи популяцияларда ол әрдайым сынамалық қателер әсерінен бұзылып отырады. Популяция өте үлкен болса да, ген жиілігі кездейсоқ флуктуацияларға байланысты р-дан р'- қа дейін үнемі өзгеріп тұрады. Популяцияны бұрынғы р күйіне бағытты түрде қайтып әкелетін ешқандай күш болмағандықтан, эволюция бағыты пайда болған р' жиілігінен ары қарай жүре береді: Харди популяциясының «жады болмайды». Ұрпақтар тізбегінде үзіліссіз жүретін осы үдеріс қайсібір кездейсоқ жағдайларға байланысты А немесе а аллельдерінің біреуінің толық жойылуына алып келеді.
Харди-Вайнберг заңы – популяциялық генетиканың қазіргі математикалық негізі болып есептелінеді. Осы қарапайым модельге генотиптің күрделі құрылымын, генотип пен фенотиптің әрқилы (кейде бір мәнді емес) қатынастарын енгізу, мутация үдерісін, популяция санының динамикасын, панмиксияның бұзылуын, миграцияның енгізілуін, сұрыптау қысымының күрделі архитектоникасын және т.б. енгізу арқылы, қазіргі кездері популяцияда орын алған генетикалық ерекшеліктер мен өзгерістер анықталып тұрады. Әлбетте, бұл есептерде Харди-Вайнберг заңы кейбір шектеулі жағдайлардың дербес шешімі болып табылады.
Екеуден көп аллельдері бар локусқа да Харди–Вайнберг заңын қолдануға болады, мысалы, үш аллельді локус үшін ол төмендегідей жазылады:
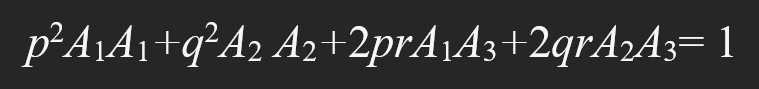
Харди-Вайнберг тепе-теңдігі заңының ұстанымы бойынша, белгілі бір жағдайлар сақталғанда аллельдер жиеліктері ұрпақтан-ұрпақта тұрақты түрде қайталанады. Бұл жағдайларда популяция тепе-теңдік қалыпында болады және ешқандай эволюциялық өзгерістерге ұшырамайды. Дегенменде, Харди-Вайнберг ұстанымы тек теоретикалық сипаттамаға ие екенін айта кетуіміз қажет. Көптеген эволюциялық өзгерістер жаңа аллельдер пайда болғаннан кейін жүзеге асады, ал оның басты көзі – мутациялар болып табылады.
Харди-Вайнберг тепе-теңдігіне қажетті жағдайлар келесі себептерден бұзылады: егер шағылысулар кездесоқ жолмен жүрмесе; популяция көлемі кішкене болса, ол гендердің ығуына (дрейф) алып келеді; генетикалық ауырлыққа ұшырататын әр түрлі гинотиптері бар даралардың тиісінше ұрпақ әкелу мүмкіндігімен (фертильділігі) шектелсе; популяциялар арасында гендер алмасуларының жүзеге асқаны жағдайларында.
Сонымен, Харди-Вайнберг заңы келесідей ұстанымдар сақталғанда ғана дұрыс жүреді: популяция көлемі өте үлкен; популяцияда іріктеу, жұптау жұмыстары жүргізілмей еркін шағылысулар жүзеге асады; жаңа мутациялар пайда болмайды; популяцияға басқа гинотиптердің енуі не одан сыртқа шығуы жүзеге аспайды. Осы талаптарды ұзақ уақыт бойы қанағаттандыра алатын популяциялар табиғатта кездеспейді. Популяцияға, оның генетикалық тепе-теңдігін бұзатын ішкі және сыртқы факторлар әрдайым әсер етіп отырады. Популяцияның генетикалық құрамын, оның генофондын ұзақ мерзімді және мақсатты түрде өзгерту – элементарлы эфолюциялық құбылыс деген атауға ие болады.
Харди-Вайнберг заңын, шындап келгенде, өте қолайлы (идеальды), яғни, еркін шағылысулар жүзеге асатын, көлемі де едәуір үлкен популяцияларда және сыртқы факторлардың әсері де жоқ жағдайларында ғана пайдалануға болады. Мұндай өте қолайлы жағдайлар табиғатта ешқашан кездеспейді. Сондықтан, Харди-Вайнберг заңы пайдаланылуының екі шектелу жағдайларын, яғни, еркін шағылысу мен сыртқы факторлар әсерлерін қарастырайық. Популяциялар гинетикасында шағылысулардың 2 түрі ажыратылады:
1. Панмиксия дегеніміз – еркін шағылысулар, яғни мұнда жұптардың түзілу мүмкіндіктері, олардың бастапқы ата тектік генотиптерінің формаларына байланысты қалыптаспайды. Табиғатта панмиксия тұтас генотип үшін еш уақытта да сақталмаса, ал жекеленген локустар бойынша оны толығымен пайдалануға болады.
2. Ассортативтілік дегеніміз – шағылысулардың іріктеу, талғау арқылы жүруі. Мұнда жұптың екінші серігін таңдауына генотип ықпал жасамайды, басқаша айтқанда, белгігі бір генотиптегі даралардың жұптасулары кездейсоқ шағылысуларға қарағанда жиірек кездеседі. Таңдау арқылы жүрген будандасулар геннің жиеліктерін өзгертпегенмен, генотиптер жиілігін өзгерте алады. Ассортативтіліктің өте шектік түрінің бірі – арнайы мақсатты жүргізілетін инбридинг, яғни, туыс дараларды бір-бірімен шағылыстырулар болып табылады.
Харди-Вайнберг тепе-теңдігінен ауытқу көрінісі популяцияға қандай да бір сыртқы фактор әсер етеді дегенді білдіреді. Гендер жиліктеріндегі өзгерістерге талдау жасау мақсатында қазіргі кездегі күрделі математикалық теңдеулер жүйесі құрастырылған. Бұл, алынатын нәтижеге ықпал ететін өзгермелі факторлардың барлығымен түсіндіріледі. Саны бойынша жеткілікті түрде үлкен, кез-келген популяцияларда ауытқулар көрсеткіштері өте аз мөлшерде болғандықтан, Харди-Вайнберг заңы мұндағы маңызды есептеулерді жасауға мүмкіндік береді және түйелердің популяциялық генетикасының негізі болып табылады. Жоғарыда көрсетілген ауытқулар үдерісі эволюцияның уақыттық ауқымында қарастырғанда ғана үлкен мәнге ие болады. Сайып келгенде, популяция генофондындағы динамикалық өзгерістерінің өзі – генетикалық деңгейдегі эволюция болып табылады.
Популяциялардағы аллельдер жиілігін өзгертетін үдерістер – қарапайым эволюциялық факторлар деген атқа ие болды. Қазіргі кезде популяциялық генетикада келесідей төрт негізгі эволюциялық факторларды ажыратады:
1. Мутациялық үдеріс – жаңа генетикалық нұсқалардың (вариант) пайда болу құбылысы. Мутация сирек кездесетін оқиға болғандықтан, олардың генофондқа әсері мен ықпалы өте аз келеді.
2. Гендер ағысы – әр түрлі популяциялардың бір-бірімен ген алмасулары. Даралардың өзара миграциясы аллельдер жиілігін мутация үдерісіндегіге қарағанда жылдамырақ өзгертуі мүмкін.
3. Гендердің ығуы (дрейф) – популяциядағы аллельдер жиіліктерінің кездейсоқ өзгеруі. Гендердің ығуы математикалық талдаулар жасау кезіндегі дараларды жалпы жиынтыққа алу барысында жол берілген қателіктерден шыққын оқиғалар қатарына жатады. Популяциядағы жиынтыққа енгізілген даралар саны неғұрлым аз болған сайын, іріктеудегі кететін қателік мөлшері соншалықты көп болады, яғни аллельдер жиіліктерінің де ауытқуы көбірек кездеседі. Саны 100-ден асатын даралары бар популяциялар үшін гендер дрейфінің маңызы шамалы. Бірнеше ғана даралардан құралған (мысалы миграция кезінде) жаңа популяцияның пайда болуы кезінде ген дрейфінің маңызы өте елеулі болуы мүмкін, яғни, "негізін қалаушының әсері" байқалады. Дәл осы даралардың генотиптері жаңа популяцияның генофондының негізіне жататын болады. Сыртқы ортаның қолайсыз жағдайы туындаған кезде, популяциядағы даралардың саны төмендейді, яғни "шөлмек эффектісі" байқалады. Популяциядағы санның бұдан бұлайғы көбеюі, яғни оның қайта қалпына келуі, тірі қалған даралар есебінен жүзеге асады.
4. Табиғи сұрыпталу – эволюцияның маңызды факторы. Популяциялық генетикада табиғи сұрыпталудың әртүрлі нұсқаларының математикалық модельдері жасап шығарылған.
Төлшеңдік (көп төлділік, өсімталдық) – белгілі бір мерзім аралығында популяцияда табиғи жолмен туылған жаңа төлдер саны. Төлшеңдікті максимальды және нақты төлшеңдік деп екі түрге бөледі.
Максимальды төлшеңдік – ортаның шектеулі факторлары болмаған кездегі туу мүмкіндігінің барынша толық жүзеге асуы.
Нақты төлшеңдік – туу мүмкіндігінің нақты жүзеге асырылуы.
Өлім-жітімге ұшырауы (шығындалуы) – белгілі бір мерзім аралығындағы популяцияда өлген даралар саны (жыртқыштардың кесірінен, аурулардың, қартайып өлу және т.б. себептерден). Өлім-жітім көрсеткіштері төлшеңдікке кері мән болып табылады.
Популяцияның өсу жылдамдығы – популяция саның белгілі бір мерзім аралығындағы арту қарқыны.
Өмір сүру ұзақтығы – даралар тіршілігінің ұзақтығы. Ол фенотиптік және генотиптік факторларға байланысты болады. Өмір сүру ұзақтығының физиологиялық, максимальды және орташа деген үш түрлері болады.
Өмір сүрудің физиологиялық ұзақтығы – белгілі бір түрдің дараларына, оның тіршілік ету кезеңі барысында шектеу жасайтын факторлардың әсері болмаған жағдайларында, өмір сүруінің теория жүзіндегі мүмкін болу ұзақтығы.
Өмір сүрудің максимальды ұзактығы – ортаның нақты жағдайында даралардың өте аз бөлігінде ғана кездесетін ұзақ ғұмыр сүру көрінісі.
Өмір сүрудің орташа ұзақтығы – популяция құрамындағы барлық даралардың сүрген өмірлерінің арифметикалық орташа мәні.
Өміршеңдік – белгілі бір мерзім аралығында популяция құрамындағы даралардың абсолюттік сақталу саны. Өміршеңдік популяцияның жастық және жыныстық құрамына, қоршаған ортаның әсерлеріне байланысты қалыптасады.
Өміршеңдіктің экологиялық стратегиясы – популяцияның тірілей қалу мен ұрпақ өрбіту мүмкіндігін арттыруға бағытталған қасиеттерінің кешені.
Популяцияның гомеостазы – популяцияның белгілі бір санын (тығыздығын) сақтап тұруы. Популяция санының өзгеруі ортаның абиотикалық, биотикалық және антропогендік сияқты бірқатар факторларына тәуелді келеді.
Популяцияға құрамындағы барлық даралар аллельдерінің кіруі себепті, оның генофонды алуан турлі. Популяцияның генетикалық өзгергіштігінің өлшем бірлігі ретінде оның гетерозиготалығы алынады.
Популяцияның гетерозиготалығы – олардың ішінде белгілі бір локус бойынша даралардың кездесу жиілігін көрсетеді. Гетерозиготалықты есептеп шығару үшін, алдымен әрбір локус бойынша гетерозиготалық жиіліктері анықталынады, сосын алынған нәтиженің орташа мәні табылады. Популяцияның гетерезиготалығына болжамды баға беру үшін 20-дан кем емес локустарға талдау жасау жеткілікті.
Гетерозиготалық – өзгергіштіктің басты сенімді көрсеткішінің бірі ретінде бағаланады. Бұл дегеніміз, популяция генофондынан кездейсоқ алынған қандайда бір аллель локустарының әртүрлі болу мүмкіндігін анықтайды. Д.А. Баймұқанов, А. Баймұқановтардың мәліметтеріне сәйкес, түйелер популяцияларының орташа гетерозиготалығы 5,18% құрайды. Сондықтан, популяцияны көптеген локустар бойынша айырмашылықтары бар генотиптер жиынтығы деп қарастыруға болады. Локустардың көпшілігі аллельдерінің молдығымен сипатталынады. Мұндай құбылысты полиморфизм деп атайды.
Түйелерді таза қанды өсіру барысында олардың шежіресін ескеру, бағалау (бонитировка) нәтижелері бойынша кластылығын анықтаудың маңызы өте зор.
Жануардың шежіресі – асыл тұқымды жануардың ата- енелері және одан бұрынғы бірнеше буын ата-бабалары жөнінде мәліметтер келтірілген шығу тегі туралы дерекнама.
Бағалау (бонитировка) – жануарларға тән белгілер кешендерін (тұқымдық тегін, өнімділік қасиеттерін, түр-сипатының ерекшеліктерін) бағалай отырып оның тұқымдық құндылығының деңгейін анықтап, тиісті кластылығын беру.
Популяциялар санына зерттеулер жүргізу барысында А.С. Северцов келесідей ортақ құбылыстарды анықтады:
1. Популяцияның көбеюге қабілетті бөлігі, яғни жыныстық жағынан жетілген және өзінен кейін ұрпақ қалдыра алатындарының саны, барлық жастағы топтарды (когорталар, генерациялар), оның ішінде жыныстық тұрғыдан жетілмегендерді қоса есептегендегі популяцияның жалпы санынан әрдайым кем болады.
2. Кіші жастағылары арасындағы өлім - жітім әдетте ересектерге қарағанда көбірек кездеседі.
3. Жылдың қолайсыз маусымдарында өлім жітім саны ұлғая түседі.
Яғни, өсімталдық (төлшеңдік), басқа да бейімдеушілік түрлері сияқты, бастапқы эволюция барысында қалыптасқан бейімделушілік түрі болып табылады. Өсімталдық – тіршілік үшін күресудің себебі емес, керісінше – оның нәтижесі.
Жасы бойынша орын алған өлім – жітімдер , әрбір дараның оны қоршаған экологиялық ортамен арасындағы күрделі кешенді қарым-қатынасының, яғни осы түрдің дараларымен және алған экологиялық орнына байланысты қалыптасатын нәтижелер.
Эволюция тұрғысынан қарағанда жасы бойынша орын алған өлім-жітімдер – популяцияның ұрпақ бере алатын бөлігін құрайтын даралардың бейімделушілігін арттыратын қуатты сүзгі болып табылады. Сонымен бірге, ол бүкіл популяцияның қоршаған ортада орын алатын онша-мұнша жағдайларына экологиялық орнықты болуына да ықпал етеді. Даралардың маусымдық опат болуы ең алдымен жылдың қолайсыз мезгілдеріндегі экологиялық жағдайдың нашарлауына байланысты орын алады, мысалы, оған оңтүстік ендіктердегі құрғақшылық маусымы, ал бореальды ендіктердегі – суық қыс маусымы жатады.
Түйе шаруашылығында қазақ бактрианы мен түркмен дромедарын түраралық будандастыруды жаппай пайдалануының салдарынан, бір жағынан будан ұрпақтар мен екінші жағынан бактриандар арасындағы түраралық жанама бәсекелестік күшейе түседі. Мұның барлығы таза қанды қазақ бактрианы санының кемуіне, ал түраралық будан түйелер санының артуына алып келеді. Будан және таза қанды түйелердің, сондай ақ түйе популяциялары арасындағы байқалатын бәсекелестік дивергенцияны күшейте түседі. Дивергенцияның нәтижесінде, түр ішіндегі дифференциялану үдерістері жылдамдайды. Оған әрбір популяцияның тіршілік етудің локальды (оқшалауланған) жағдайларына бейімделгендігі себеп болады. Бәсекелестікте өрбитін дивергенцияға қарағанда, түрішіндік дифференцияланудың өзі әлдеқайда күрделі үдеріс болып табылады.
Нарықтық экономика жағдайында түйелердің түрішіндік қатынастағы шартты бәсекелестік, түйені падаланудан (эксплаутациялық) және интерференциялық бәсекелестіктерімен алмастырылады. Осының салдарынан элиминация таңдалып жасалынады. Шартты бәсекелестік кезінде әрбір ағзаның жеке, ең майда ерекшеліктеріне дейін бағаланатыны белгілі. Ал интерференттік бәсекелестік кезінде нарық сұраныстарына сай келетін белгілер ғана бағаланады. Нақтырақ айтсақ, енжар (пассивный) бәсекелестіктің маңызы арта түседі. Енжар (пассивті) бәсекелестік – ағзаның жыртқыштармен паразиттерге, қатаң абиотикалық әсерлерге, ортаның зақым келтіретін факторларына төтеп берудегі биологиялық сапасының әртүрлілігі. Ортаның тұрақты жағдайында сұрыпталу әрбір түр дараларының қалыптасып қалған бейімделушілік құрылымдарын қолдап тұрады. Бұған мысал ретінде, түйелер (бактриан және дромедарлар) мен олардың популяцияларын азды - көпті тұтас және тұрақты гоместатикалық жүйе есебінде келтіруге болады
Тіршілік ету жағдайларының күшті флуктуациялануы, не болмаса оларды бағытты түрде өзгертулер, бастапқыда ағзаның прогрессивтік эволюцияға қарсылығын тудырып, тіршілікке қабілеттілігіне оң әсер етулер орын алғаннан кейін ғана, жаңа бейімделінушіліктің қалыптасуына алып келуі мүмкін.
Тrade off тиімділігіне негіз беретін ағзаның тұтастығы мен қоршаған орта факторларының кешенді әсерлері, бағытты түрдегі және ұзақ мерзім бойына тіршілік ету жағдайының өзгертілуі нәтижелерінде, жекелеген бейімделушілік белгілері ғана емес, барлық даралар жиынтығы мен популяция құрылымының қайта құралуларына алып келеді. Мысалы, Д.А. Баймұқановтың мәліметтеріне сәйкес, түйелердегі өркештердің пайда болуы – Еуразия және Африканың шөл және шөлейт жағдайларына бейімделуімен байланысты. Сонымен бірге, генетикалық зерттеулер түйелердегі өркештердің пайда болуына хромосомадағы емес, ген деңгейіндегі мутациялардың пайда болуы себеп болатынын анықтаған. Түйелердің өркеші ағзаның қуат қорын жоғарылатады және жылдың қыс және ерте көктем маусымдарында орын алатын табиғи жайылымдардағы азықтың тапшылығына, ауаның жазда өте ыстық, қыста өте суық болған кездерінде дене температурасын бір қалыпты сақтап қалуы үшін де пайдаланылады. Жоғарғы дәрежедегі қоңдылықтағы түйелер (өркештер майға толық толған кезде) өздерінің жақсы өсімталдығымен ерекшеленеді. Қазіргі кезде қоршаған ортаның және молекулярлық-генетикалық деңгейінде жүрген мутациялық өзгерістердің әсерінен, түйелерде ағзаның бейімделушілік гемостазының ығысуы байқалады. Сондықтан да, түйелердегі шөл және шөлейт аймақтардың экологиялық жағдайларына айқын байқалып тұрған бейімделушіліктерін салыстырмалы деуге болады.
Әдебиеттер
1. AWF Edwards. ·Genetics. 2008 Jul;179(3):1143-50. doi: 10.1534/genetics.
2. Айала Ф. Введение в популяционную и эволюционную генетику. – Москва: Мир, 1984. -230с.
3. Баймуканов Д.А. Эволюция и систематическое положение верблюдов //Верблюдоводство Казахстана XXI века. – Алматы
4. Аубакиров Х.А. Биометрия. Оқулық. КР Білім және ғылым министрлігі. ӘОЖ 575 (075) КБЖ 28.04 я7 А91. Алматы, ЖШС РПБК «Дәуір», 2011., -407
5. Аубакиров Х.Ә., Тлепов А.Ә., Кенжеходжаев М.Д. Популяциялар және сапалық белгілер генетикасы. Оқу құралы. ӘОЖ 579 (075) х; КБЖ28.70я7. А 93. ISBN 978-601-329-303-5. Алматы, «Эферо» баспасы, 2022., – 310 бет
6. Баймуканов Д.А., Баймуканов А. Эволюция и систематика верблюдов (2-е издание) //Монография. – Шымкент: Полиграф, 2011. - 88с.
7. Баймуканов Д.А., Баймуканов А. Генетика, эволюция и систематика верблюдов (полное издание). // Монография. – Шымкент: Полиграф, 2011. -117с.
8. Баймуканов Д.А., Юлдашбаев Ю.А., Дошанов Д.А. Верблюдоводство (Бакалавриат): (ISBN 978-5-906818-14-0). Учебное пособие - Москва: Издательство КУРС, НИЦ ИНФРА-Москва, 2016. - 184 с.
9. Баймұқанов Д.А., Исхан К., Жунісов А.Т. Түйе шаруашылығы өнімдерің өндіру технологиясы // Окұ қуралы (ISBN 978-601-241-084-6). – Алматы: «Айтұмар» баспасы, 2021. 164 б.





















































































































Комментарии (0)