Взаимодействия между микроэлементами и макроэлементами у стельных нетелей
Существует недостаток информации о взаимодействиях минералов, происходящих в период стельности нетелей, когда потребность в питательных веществах высока. Целью настоящего исследования было оценить взаимодействия между макроэлементами кальцием (Ca), фосфором (P), магнием (Mg), серой (S), натрием (Na) и калием (K) и микроэлементами медью (Cu), железом (Fe), марганцем (Mn), молибденом (Mo), селеном (Se), цинком (Zn) и кобальтом (Co) у стельных нетелей.
Аннотация
В 7-дневном опыте по переваримости (косвенный метод с использованием лигнина в качестве маркера) использовали 24 стельных нетели (возраст 18.2 ± 0.14 месяцев; 87.5 ± 4.74 дня стельности; 497 ± 8.5 кг живой массы), которых кормили рационом, обеспечивающим минералы в соответствии с нормами NASEM для стельного мясного скота. По окончании опыта по переваримости у каждой нетели были взяты пробы крови (пункция яремной вены) и печени (биопсия). Множественные модели линейной регрессии были разработаны на основе основных корреляций и с учетом эффектов коллинеарности между переменными.
Потребление железа положительно влияло на концентрацию Mo в сыворотке крови, которая у большинства животных составляла >10 мкг/дл. Кажущееся усвоение Fe отрицательно зависело от потребления Mo и соотношения Ca к P в корме, тогда как усвоение Se отрицательно зависело от потребления Fe и положительно — от потребления Mo. Коэффициенты усвоения Mo и Fe были обратно связаны, что подчеркивает взаимодействие между этими элементами. Содержание Fe в сыворотке и Cu в печени положительно коррелировали, в то время как содержание Fe в печени было обратно связано с содержанием Mn в печени. Меньше взаимодействий наблюдалось между макроэлементами.
В заключение, среди наблюдавшихся взаимодействий особого внимания заслуживают взаимодействия между Fe и Mo, поскольку высокое содержание Fe в рационе обычно характерно для типичных рационов жвачных, а высокий уровень Mo в сыворотке может указывать на молибденоз. Кроме того, высокое потребление Fe, по-видимому, влияет на усвоение Se и Mn.
1. Введение
Многие макро- и микроэлементы играют ключевую роль в здоровье жвачных животных, хотя последние признаны более значимыми для репродуктивной функции [1, 2, 3]. К примеру, сообщалось, что адекватное добавление Cu, Zn, Mn [4] и Co улучшает показатели стельности у многорожавших молочных и мясных коров [5, 6], в то время как другие исследователи наблюдали, что добавление Mo задерживает наступление половой зрелости, снижает частоту зачатия и может вызывать ановуляцию и анэструс у нетелей [7]. Селен также играет важную роль в репродукции жвачных животных, защищая ранний плод от окислительного повреждения, тем самым снижая смертность [8]. Микроэлементы также могут влиять на здоровье и зоотехнические показатели как коровы, так и теленка [1, 9] и даже вызывать эпигенетические эффекты у потомства [10].
Усвоение минералов и микроэлементов в кишечнике — сложный процесс, на который влияют многочисленные факторы [11, 12, 13, 14], среди которых синергические или ингибирующие взаимодействия между некоторыми из них имеют ключевое значение [15]. Взаимодействия между определенными минералами хорошо задокументированы [14, 16, 17, 18], особенно между Cu, Mo и S. Взаимодействие Mo и S в рубце приводит к образованию тиомолибдата (ТМ), который связывает Cu, в результате чего образуется нерастворимый комплекс Cu, недоступный для поглощения и метаболизма организмом [19]. Более того, абсорбированный ТМ может связывать Cu, присутствующий в других тканях, вызывая вторичный дефицит Cu, который также может быть инициирован взаимодействием Cu и Fe [20].
Напротив, существуют взаимодействия между минералами у жвачных животных, которые недостаточно изучены и поняты, несмотря на их возможное негативное воздействие на здоровье животных [21, 22, 23, 24]. Например, рационы жвачных животных с высоким содержанием железа не являются редкостью, и его влияние на усвоение других минералов, таких как марганец и молибден, заслуживает большего внимания. Учитывая множественные и сложные взаимодействия минералов, необходимо оценивать усвоение каждого элемента в различных условиях питания, поскольку дефицит этих элементов может негативно сказаться на продуктивности животных [15]. Напротив, когда количество поступающего минерала превышает потребности животного, может возникнуть потенциальный антагонизм и усиленное выделение с навозом минералов, имеющих важное значение для окружающей среды [15]. Более того, негативное воздействие этих взаимодействий, вероятно, будет более выраженным у беременных животных из-сть изменений в их метаболизме и повышенной потребности в питательных веществах [25, 26, 27]. Поэтому важно лучше понимать взаимодействие минералов и то, как это может влиять на их усвоение стельными нетелями. Эта информация может быть использована для обеспечения адекватного уровня минералов в рационе, чтобы гарантировать удовлетворение потребностей потомства в минералах без ущерба для здоровья матери [1, 9, 10, 27]. В этом контексте цель данного исследования была направлена на оценку взаимодействия и усвоения Cu, Fe, Zn, Mn, Mo, Se, Ca, P, Mg, S, Na и K у стельных нетелей, получающих типичный рацион.
2. Материалы и методы
2.1. Животные и диета
Сорок семь нетелей были подвергнуты воздействию быка-пробника и носили ошейник для определения течки (SCR HEATIME®, SCR North America, Мэдисон, Висконсин, США) для выявления охоты и дальнейшего осеменения. После положительных тестов на стельность двадцать четыре стельные нетели в возрасте 18 месяцев (средний породный состав: 63,4% ангус, 24,3% симментальская, 2,9% гельбвийская и 9,4% другие европейские породы) были размещены в шести открытых загонах размером 10,8 × 5,4 м с защищенной зоной (по четыре нетели в каждом загоне) в течение первых трех месяцев стельности. Эти животные получали основной рацион, содержащий сенаж, силос из цельной кукурузы и премикс минералов и витаминов (Таблица 1). Экспериментальные процедуры, проведенные в исследовательском центре Elora Beef Cattle при Университете Гвельфа, соответствовали рекомендациям Канадского совета по уходу за животными [28] и были одобрены Комитетом по уходу за животными Университета Гвельфа (номер протокола использования животных (AUP) 10R093).
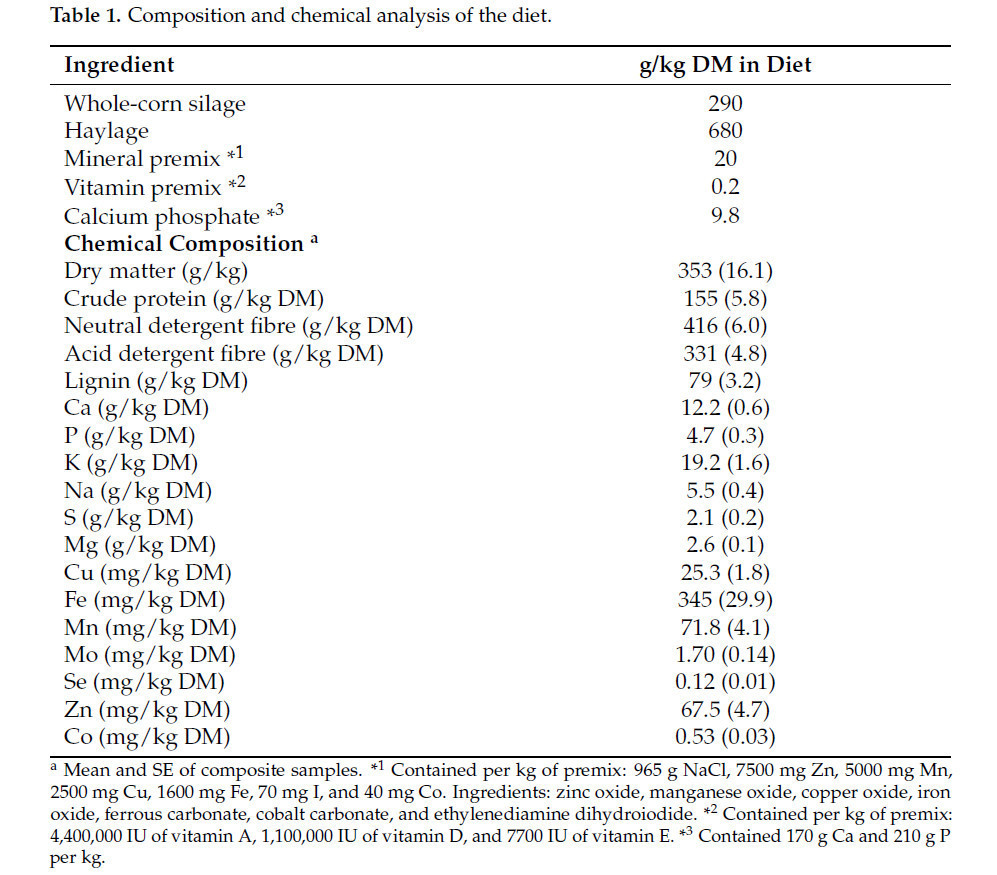
Таблица 1. Состав и химический анализ рациона питания.
2.2. Испытание на переваримость
Примерно на третьем месяце стельности нетелей (возраст 18,2 ± 0,14 месяцев; срок стельности 87,5 ± 4,74 дня; масса тела 497 ± 8,5 кг) содержали в отдельных крытых стойлах размером 10 × 3 м площадью 9 м², застеленных древесной стружкой, для проведения 7-дневного эксперимента по определению переваримости (косвенный метод). Пробы влажного кала (каждая примерно по 100 г) отбирали с пола дважды в день в течение семи дней подряд (с 06:00 до 07:30 и с 18:00 до 19:30). После сбора кала стойла очищали, чтобы избежать загрязнения между пробами. Все пробы кала от каждой нетели за определенный период смешивали для получения единого сводного образца. Ежедневные образцы фекалий (350–400 г) хранились в морозильной камере (−20 °C) до конца недели, после чего все образцы за 7 дней были объединены в один. Нетелей кормили ежедневно, а остатки корма собирали и взвешивали каждые 24 часа для дальнейшего расчета потребления корма. Образцы корма и остатков также объединяли в один еженедельный образец для дальнейшего анализа. После окончания экспериментального периода все образцы высушивали в сушильном шкафу при 60 °C и доставляли на анализ в Отдел лабораторных услуг Университета Гвельфа.
Микроэлементы и некоторые макроэлементы (Na, S) в фекалиях и кормах анализировались с помощью ICP-MS (масс-спектрометрия с индуктивно-связанной плазмой) для анализа металлов (масс-спектрометр Varian 820-MS ICP, Varian Inc., Пало-Альто, Калифорния, США), в то время как макроэлементы анализировались в соответствии с рекомендациями Ассоциации официальных аналитических химиков [29] (метод AOAC 985.01 для Ca, P, K и Mg; метод AOAC 923.01 для S и метод AOAC 969.10 для Na). Кислотно-детергентная клетчатка (КДК) определялась в соответствии с методами Ассоциации официальных аналитических химиков [29], тогда как нейтрально-детергентная клетчатка (НДК) определялась в соответствии с процедурой Ван Соеста и др. [30]. Содержание клетчатки и лигнина в корме анализировали с помощью анализатора клетчатки ANKOM (Ankom Technology Corp., Fairport, NY, USA), а содержание влаги и сырого протеина определяли в соответствии с методами AOAC 930.15 и AOAC 2001.11 соответственно.
Лигнин использовался в качестве внутреннего маркера, а кажущийся коэффициент усвоения (КУ) каждого минерала/элемента рассчитывался следующим образом [31]:
КУ питающего элемента = 100 − 100 × (% лигнина в сухом веществе корма / % лигнина в фекалиях (сухое вещество)) × (% элемента в фекалиях СВ / % элемента в сухом веществе корма)

2.3. Сыворотка крови
Образцы крови отбирали путем венепункции яремной вены в стерильные вакуумные пробирки vacutainer® (BD Biosciences®, Миссиссауга, Онтарио, Канада), надетые на иглу 18 калибра × 2,5 см. Кровь собирали у всех 24 нетелей в последний день эксперимента по определению переваримости корма. Образцы оставляли при комнатной температуре на 25 минут для свертывания крови, после чего сыворотку получали центрифугированием при 3750× g в течение 25 минут при 4 °C и хранили при −80 °C до отправки в Отдел лабораторных услуг Университета Гвельфа для анализа содержания Cu, Fe, Mn, Mo, Zn, Co и Se методом ICP-MS. Содержание макроэлементов Ca, P (неорганический фосфат), K, Na и Mg определяли с помощью анализаторов Roche cobas 6000 c501 (Roche Diagnostics, Лаваль, Квебек, Канада) с использованием протоколов и реагентов, предоставленных Roche/Hitachi.
2.4. Печень
Образцы печени (0,5 г) были взяты у каждой нетели к концу эксперимента по определению переваримости. Ткань печени была извлечена путем биопсии, как описано Гринвудом и др. [32]. Более подробная информация приведена в работе Диаса и др. [33]. Образцы были отправлены в Отдел лабораторных услуг для анализа содержания Cu, Fe, Mn, Mo, Zn, Co и Se во влажной ткани методом ICP-MS.
2.5. Статистический анализ
Все статистические анализы проводились с использованием SAS (версия 9.2; SAS Institute Inc., Кэри, Северная Каролина, США). Все измерения проводились для каждого животного, таким образом, экспериментальной единицей являлась каждая нетель, содержащаяся в отдельном стойле во время эксперимента по определению переваримости (общее количество наблюдений составило 24). Коэффициент корреляции Пирсона (r) был получен с помощью PROC CORR в SAS. Линейная регрессия (PROC REG) использовалась для построения моделей множественной линейной регрессии (МЛР), основанных на значимых корреляциях, обнаруженных между следующими переменными для каждого минерала: потребление, КУ, концентрации в сыворотке и печени, т.е.
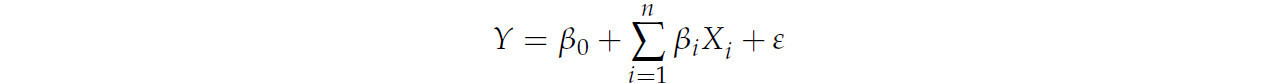
где Y — зависимая переменная, β₀ — свободный член, βᵢ — наклон для независимой переменной Xᵢ, n — количество независимых переменных, включенных в модель множественной линейной регрессии, и ε — ошибка.
Для устранения влияния коллинеарности между переменными в модели множественной линейной регрессии был проведен тест на коллинеарность в SAS (PROC REG, опции VIF, TOL и COLLINOINT). Допуск (запрашиваемый опцией TOL) — это доля дисперсии каждого предиктора, которая не объясняется всеми остальными предикторами, а VIF (или коэффициент инфляции дисперсии) — это просто обратный допуск. VIF представляет собой коэффициент, на который умножается дисперсия оцениваемого коэффициента из-за мультиколлинеарности в модели.
Из модели были удалены переменные с высоким индексом обусловленности (>30) и предикторы с большой долей вариации для очень малых собственных значений (в данном случае, последние три собственных значения). Таким образом, в статистических моделях были сохранены только значимые объясняющие переменные (p < 0,05). Кроме того, в модели были сохранены только переменные, которые либо имели нулевую, либо слабую корреляцию друг с другом и/или те, чей VIF был ниже 10. Когда корреляция между переменными была сильной (≥0,80) и VIF > 10, выбор переменных для включения в модели основывался на (а) предыдущих знаниях о взаимодействиях между объясняющей и зависимой переменными и (б) более высоком коэффициенте корреляции и меньшей среднеквадратичной ошибке модели при замене одной переменной другой. Таким образом, хотя минералы, включенные в модель, влияют на зависимую переменную, возможно, что другие минералы, которые также могут влиять на эту переменную, отсутствуют в модели. Однако в большинстве случаев другие минералы не включаются в модель из-за их слабого влияния на независимую переменную.
3. Результаты
Уравнения, показывающие взаимосвязь между коэффициентами усвоения минералов и их потреблением, представлены в таблице 2. Коэффициенты усвоения всех микроэлементов были тесно связаны (r² > 0,50) с потреблением других минералов, особенно других микроэлементов. В отличие от этого, коэффициенты усвоения некоторых макроэлементов, таких как Na или K, либо не зависели от потребления минералов, либо зависели лишь слабо, что приводило к низкому коэффициенту детерминации (r² < 0,15); другие, такие как P [уравнение (9)] и Mg [уравнение (10)], показали более слабые, хотя и значимые (p < 0,4), значения r² (<0,40). Коэффициенты усвоения микроэлементов и макроэлементов также были связаны с коэффициентами усвоения других минералов (Таблица 3), за исключением коэффициентов усвоения Na и K, для которых значения r² были <0,25. Коэффициенты регрессии иногда были положительными, а иногда отрицательными, что указывает на сложные взаимодействия между минеральными элементами при их абсорбции в кишечнике.
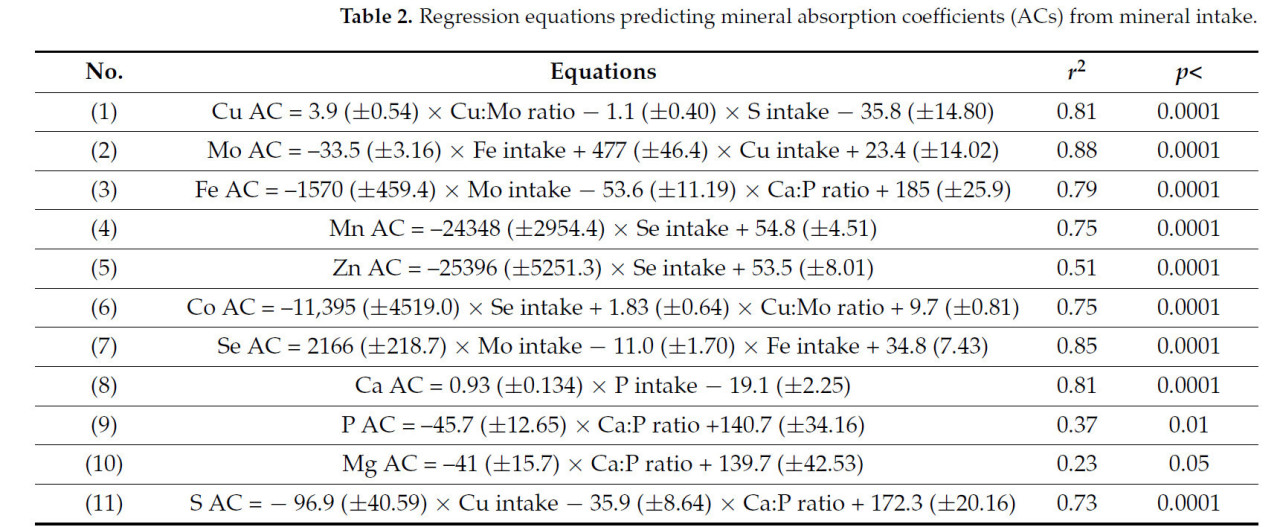
Таблица 2. Регрессионные уравнения, прогнозирующие коэффициенты усвоения минералов (КУ) на основе потребления минералов.
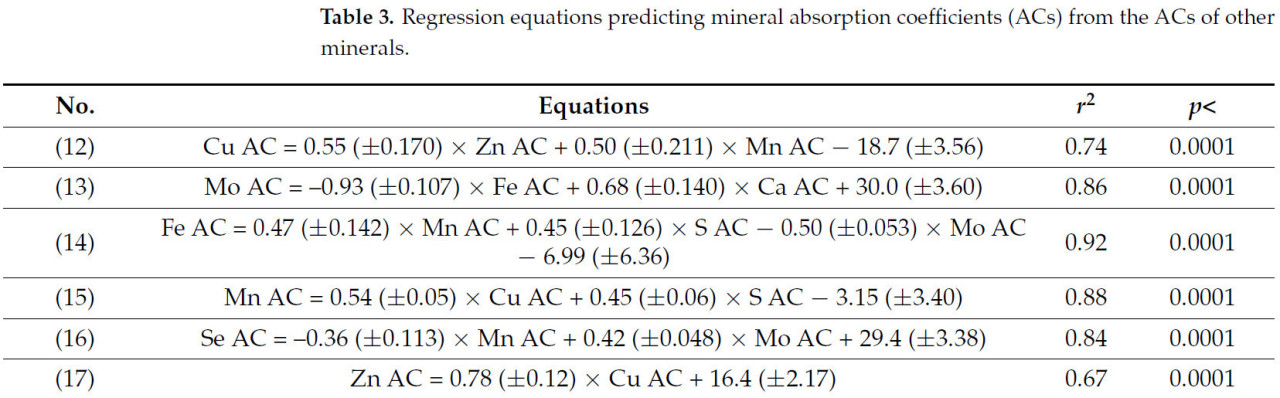
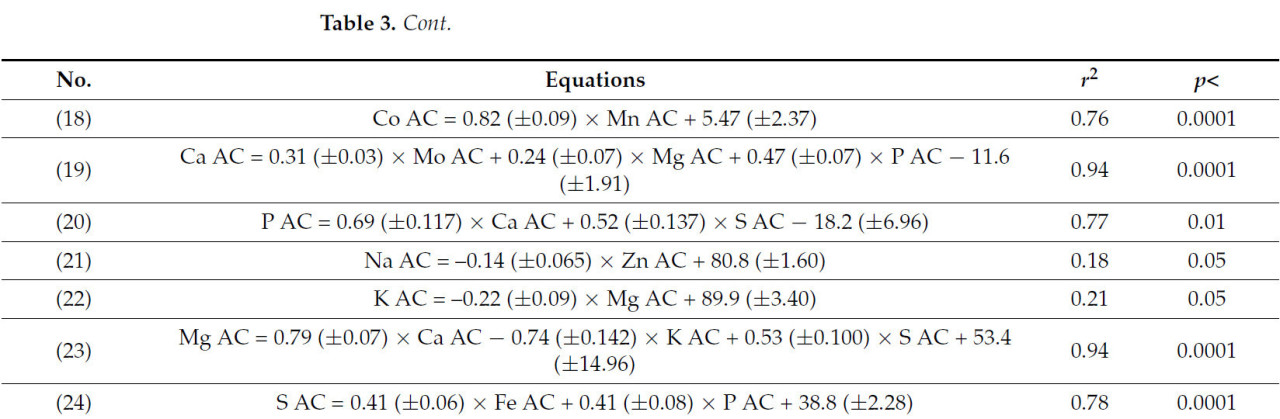
Таблица 3. Уравнения регрессии, прогнозирующие коэффициенты усвоения минералов (КУ) на основе КУ других минералов.
Было обнаружено несколько корреляций между концентрацией минералов в сыворотке и потреблением минералов (Таблица 4), в частности, между концентрациями Cu, Mo, Se и Zn в сыворотке и потреблением других микроэлементов. Корреляции между Cu в сыворотке и потреблением Cu [уравнение (25)], а также между Mo в сыворотке и потреблением Fe [уравнение (26)], хотя и статистически значимые (p < 0,01), были умеренными (r² < 0,40). Хотя потребление минералов показало существенные корреляции с КУ и концентрациями микроэлементов в сыворотке, значимой корреляции (p > 0,05) между потреблением минералов и концентрацией минералов в печени не наблюдалось.
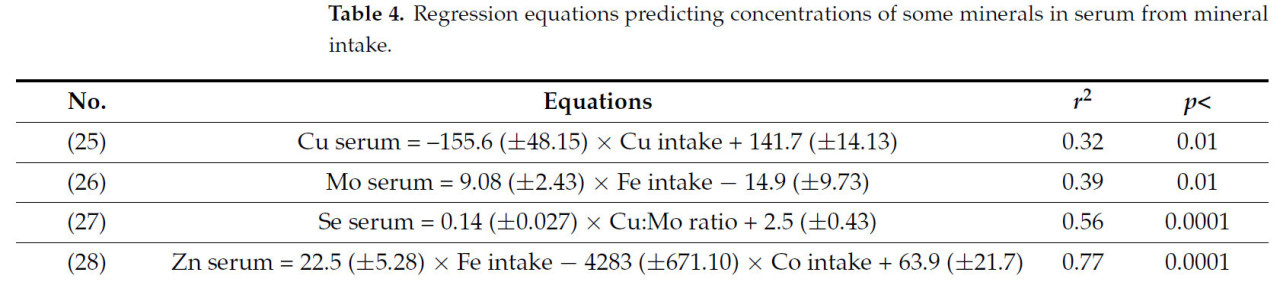
Таблица 4. Регрессионные уравнения, прогнозирующие концентрацию некоторых минералов в сыворотке крови на основе потребления минералов.
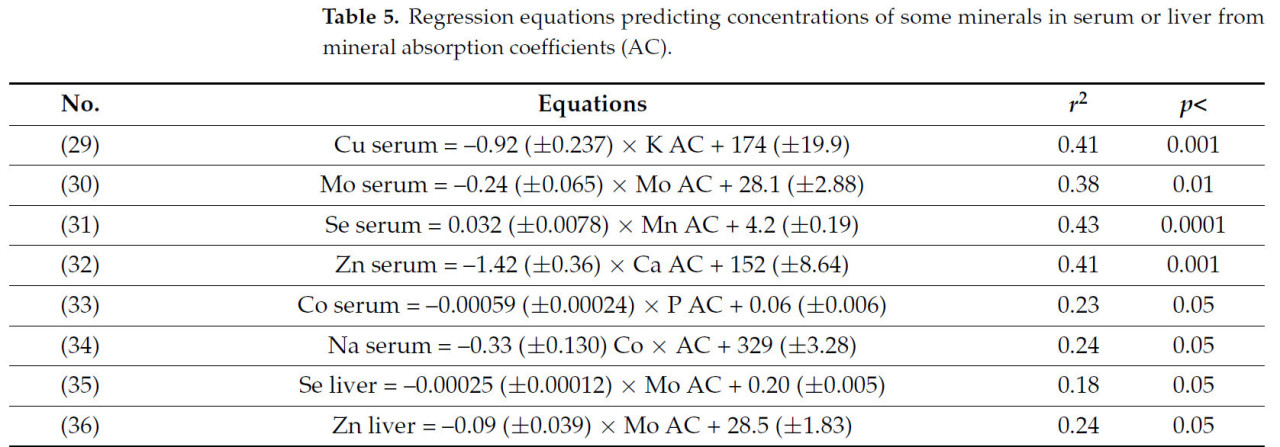
Статистические модели, описывающие взаимосвязь между концентрациями некоторых металлов в сыворотке или печени и минеральными КУ, представлены в таблице 5. Хотя коэффициенты детерминации этих моделей не особенно высоки, эти уравнения указывают на то, что концентрация некоторых микроэлементов и Na в сыворотке может зависеть от минерального КУ (Таблица 5). В меньшей степени уравнения (35) и (36) в таблице 5 показывают, что концентрации Se и Zn в печени показали значительную отрицательную корреляцию с КУ Mo, хотя значения r² были относительно низкими (< 0,25). Другие уравнения были выведены на основе взаимосвязи между концентрациями Mo, Cu или Zn в сыворотке и потреблением или КУ других минералов:
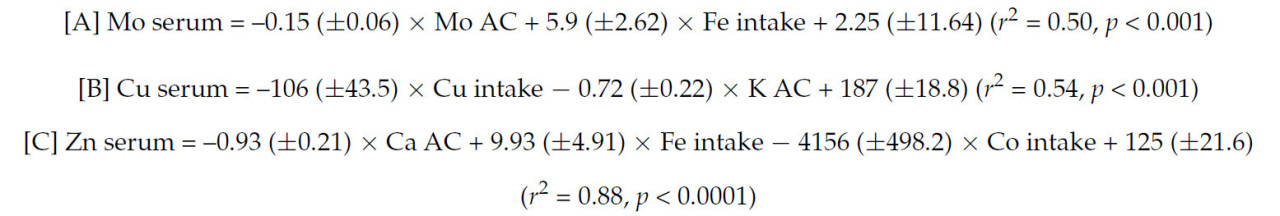
Наконец, концентрации некоторых минералов в сыворотке или печени были связаны с концентрациями других минералов в сыворотке или печени, как показано уравнениями в таблице 6.
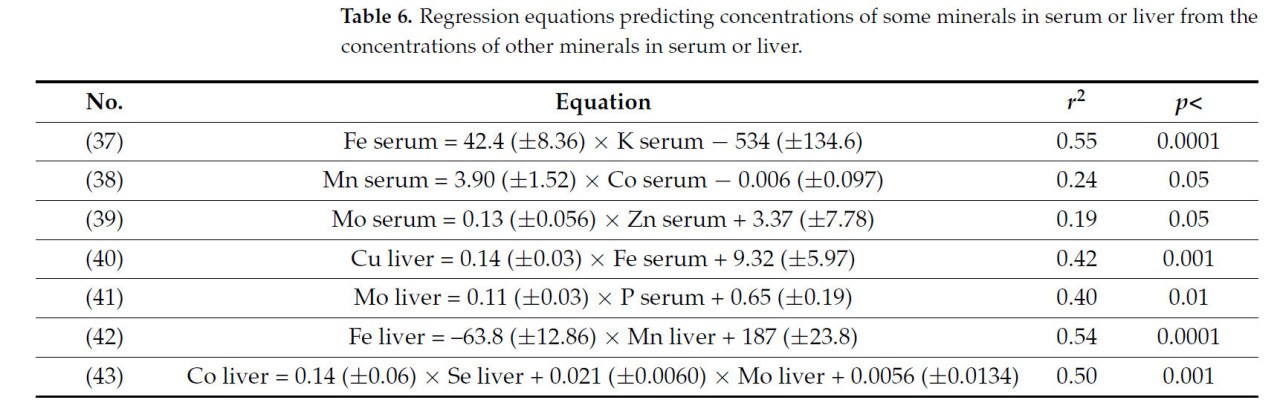
Таблица 6. Уравнения регрессии, прогнозирующие концентрации некоторых минералов в сыворотке или печени на основе концентраций других минералов в сыворотке или печени.
4. Обсуждение
4.1. Коэффициенты усвоения (КУ) минералов в зависимости от потребления или КУ других минералов
Минеральный состав рациона был составлен с учетом требований NASEM для стельных мясных нетелей [34]. Однако, как это часто бывает в животноводческих системах, количество некоторых микроэлементов и минералов превышало минимальные требования, чтобы предотвратить возможный дефицит. Точнее, потребление Fe было почти в 7 раз выше [35] минимальных требований NASEM, тогда как Co, Ca и Na были, соответственно, примерно в 5, 4 и 7 раз выше требований NASEM, а другие минералы были в 1-3 раза выше требований. Содержание Se в рационе находилось в пределах требований NASEM.
Влияние соотношения потребления Cu:Mo и S на КУ Cu (Таблица 2, уравнение (1)) согласуется с известным фактом, что образование тиомолибдатов благоприятствуется низким содержанием Cu:Mo в рационе и высоким содержанием S в рационе, что, в свою очередь, снижает усвоение Cu [19, 36, 37]. Избыточное потребление Fe отрицательно влияло на КУ Mo, тогда как потребление Cu оказывало на нее положительное влияние (Таблица 2, уравнение (2)). Известно, что взаимодействие между Fe и Mo может происходить в результате их участия в ксантиноксидазе [38, 39]. На КУ Mn, Zn и Co отрицательно влияло потребление Se (Таблица 2, уравнения (4)–(6) соответственно). Основная известная связь между Mn, Zn и Se связана с антиоксидантными функциями. В то время как Zn и Mn входят в состав супероксиддисмутазы (СОД), которая восстанавливает свободный радикал супероксида до перекиси водорода, Se входит в состав глутатионпероксидазы, которая превращает перекись водорода в воду. Однако информация о влиянии потребления Se на абсорбцию Mn и Zn скудна. Взаимодействие между Se и Co было описано в эксперименте на крысах, где Гардинер и Никол [40] пришли к выводу, что взаимодействие Se и Co, вероятно, происходит в кишечнике, влияя на абсорбцию. Аммерман [41] предположил, что взаимосвязь между Se и Co может быть связана с присутствием витамина B12 в метаболизме диметилселенида.
В то время как потребление Se отрицательно влияло на КУ Zn, Mn и Co, КУ Se зависел от потребления Mo и Fe (Таблица 2, уравнение (7)). Взаимодействие между Se и Mo было продемонстрировано на бройлерах, получавших разные уровни Se и Mo [42]. Интересно, что было показано, что при токсичных уровнях Se в рационе птицы, по-видимому, могли переносить более высокие уровни Mo, или что токсичность Mo снижалась. В нашем исследовании уровни Se в печени и сыворотке находились в пределах допустимых значений (0,19 мкг/г и 4,86 мкг/дл соответственно), тогда как уровень Mo в сыворотке был выше адекватных уровней для большинства животных (>10 мкг/дл). Таким образом, было бы интересно дополнительно исследовать, влияет ли низкое потребление Se на метаболизм Mo. Обратная зависимость между КУ Se и потреблением высоких уровней Fe согласуется с ингибирующим эффектом избыточного потребления Fe на утилизацию Se у крыс [43].
Как и ожидалось, КУ Ca был связан с потреблением P, тогда как КУ P, Mg и S были выше при снижении соотношения Ca:P в рационе (Таблица 2, Уравнения (8)–(11)). Отрицательные корреляции между соотношением Ca:P в рационе и КУ P (Уравнение (9)) или КУ Mg (Уравнение (10)) были значимыми, но относительно слабыми. Взаимодействие минералов при их всасывании в кишечнике было рассмотрено Гоффом [12]. Среднее значение соотношения Ca:P в рационе в данном исследовании составило 2,6, что выше рекомендации NASEM [34]. Хотя существует общепринятое мнение, что жвачные животные могут переносить широкий диапазон соотношений Ca:P без ущерба для своей продуктивности [34], возможно, что соотношения выше 2:1 могут повлиять на доступность других минералов. Согласно Хёрли и Доану [36], избыток Ca может ухудшить репродуктивную функцию, снижая усвоение P, Mg, Zn и Cu и вызывая вторичный дефицит этих минералов у молочного скота.
Взаимодействие между Cu и Zn, а также Cu и Mn было описано другими исследователями [44, 45]. Можно предположить, что высокие корреляции между КУ Cu, Zn и Mn (Таблица 3, уравнения (12), (15) и (17)) могут быть связаны с их присутствием в таких соединениях, как супероксиддисмутаза (СОД) и металлопротеины [46, 47]. Сообщалось, что экспрессия Cu/Zn-СОД, присутствующей в цитозоле, была повышена в органах развивающихся плодов [48], тогда как более высокая ферментативная активность Cu/Zn-СОД наблюдалась во время стельности овец с 15 по 40 день [49]. С другой стороны, взаимосвязь между КУ Cu и КУ Mn (Уравнение (12)) и между КУ Mn и КУ Fe (Уравнение (14)) может быть связана с общим белком переносчика двухвалентных металлов 1 (DMT1) [50], участвующим в поглощении Cu, Mn и Fe.
Отрицательная взаимосвязь между кажущимся усвоением Fe и Mo (Таблица 3, уравнения (13) и (14)) может быть ответом на присутствие этих минералов в ферментном комплексе ксантиноксидазы [51], хотя механизм, посредством которого происходят эти взаимодействия, неясен. Природа положительной взаимосвязи между кажущимся усвоением Fe и S также неясна. Увеличение кажущегося усвоения Co с увеличением кажущегося усвоения Mn (Таблица 3, уравнение (18)) ранее, насколько нам известно, не сообщалось. Однако возможно, что интерференция в кажущемся усвоении друг друга связана с аналогичным механизмом, посредством которого Co интерферирует в поглощении Fe [52]. Железо может быть замещено Mn в нескольких случаях, и оба минерала, по-видимому, имеют один и тот же механизм переноса [50].
Положительные взаимосвязи между КУ Mo и КУ Ca (Таблица 3, уравнения (13) и (19)) противоречат взаимосвязи, обнаруженной Кэткартом и др. между потреблением Ca (а не усвоением) и КУ Mo у молочного скота [53]. К сожалению, других работ, связывающих КУ Ca и Mo, для получения дополнительной информации о возможном взаимодействии этих минералов не было. Напротив, положительные взаимосвязи между КУ Ca и КУ P и Mg (Таблица 3, уравнения (19), (20) и (23)) легче понять благодаря известным взаимодействиям между этими минералами, молекулярным механизмам кишечной абсорбции и транспортным процессам, регулируемым рядом физиологических медиаторов этих необходимых питательных веществ [54], а также их присутствию в костях и во внутри- и внеклеточных жидкостях организма [55, 56, 57]. Отрицательные корреляции между КУ Mg и КУ K (Таблица 3, уравнения (22) и (23)) согласуются с сообщенными антагонистическими эффектами K на использование Mg у овец [58, 59, 60] и крупного рогатого скота [61, 62].
Корреляция между КУ S и КУ P (Таблица 3, уравнения (20) и (24)) редко встречается в питании жвачных животных. Известно, что P необходим для оптимального синтеза микробного белка и захвата S, и что рубцовые микроорганизмы используют как неорганическую, так и органическую серу для синтеза серосодержащих аминокислот при производстве микробного белка [63]. Наши результаты показывают значительную и положительную корреляцию между КУ обоих питательных веществ, хотя, насколько нам известно, эта прямая взаимосвязь ранее не описывалась. Сообщалось об аддитивном эффекте этих минералов на перевариваемость целлюлозы in vitro [64]. Отсутствие дополнительной информации препятствует дальнейшему обсуждению обнаруженной взаимосвязи между этими минералами.
4.2. Влияние на уровень микроэлементов в сыворотке крови
Концентрации минералов и микроэлементов в крови находились в пределах значений, указанных Аткинсом и др. [65]. Содержание меди в сыворотке отрицательно коррелировало с потреблением Cu, как показано в уравнении (25) (Таблица 4), хотя коэффициент детерминации был не особенно высоким. К сожалению, эта взаимосвязь неясна, поскольку изменения содержания Cu в сыворотке/плазме чувствительны к другим факторам, таким как стресс [66] и накопление Cu в печени [19, 67]. Положительная взаимосвязь между содержанием Mo в сыворотке и потреблением Fe (Таблица 4, уравнение (26)) подтверждает наличие взаимодействия между этими минералами, особенно учитывая, что у 79% животных наблюдался высокий уровень Mo в сыворотке (>10 мкг/дл), что указывает на молибденоз. В уравнении (27) (Таблица 4) содержание Se в сыворотке увеличивается с увеличением соотношения Cu:Mo в рационе. Хотя данных о возможной связи между этими минералами недостаточно, Миллс [68] отметил, что несколько исследований показали, что влияние дефицита Se на перекисное окисление липидов было сильнее при одновременном дефиците Cu и Mn. Возможно, Cu повлиял на метаболизм Se в нашем исследовании. Дальнейшие исследования были бы интересны для оценки того, усугубят ли одновременные дефициты Se и Cu влияние на перекисное окисление липидов. Возможные последствия взаимодействия Se и Mo были упомянуты ранее в этой статье.
Взаимодействие между Zn и Fe хорошо изучено [68, 69] и было обнаружено между Zn в сыворотке и потреблением Fe, как показано в уравнении (28) (Таблица 4). Напротив, хотя взаимодействие между Co и другими минералами, такими как Cu и Fe, было признано [50, 52], обнаруженная здесь взаимосвязь между Zn в сыворотке и потреблением Co, хотя и значительная, не поддается простому объяснению. Одна из возможностей заключается в том, что их взаимосвязь связана с кобаламин-зависимой метионинсинтазой, которая является важным металлоферментом, использующим Zn для активации субстрата гомоцистеина.
В литературе мало информации о корреляциях между усвоением K и Cu в сыворотке (Таблица 5, уравнение (29)). Интересно, что усвоение K не сильно коррелировало ни с усвоением Mg, ни с усвоением S, которые слабо коррелировали с Cu в сыворотке, что исключает эффекты коллинеарности между K и другими минералами, коррелирующими с Cu в сыворотке. Молибден в сыворотке отрицательно коррелировал с усвоением Mo (Таблица 5, уравнение (30)). Высокие концентрации Mo в сыворотке (более 10 мкг/дл), вероятно, являются следствием как усвоения Mo, так и потребления Fe, как показано в объединенном уравнении [A] в разделе 3. Дисбаланс, связанный с метаболизмом Mo, может привести к физиологическому дефициту Cu, что вредно для здоровья матери [70]. В уравнении (31) (Таблица 5) концентрация Se в сыворотке положительно коррелирует с усвоением Mn, что указывает на взаимодействие между этими элементами. Взаимодействие между уровнем Mn и Se в рационе овец, влияющее на прохождение Mg в тонкий кишечник, было отмечено Эрнандесом-Кальвой и др. [71]. Однако трудно установить, связано ли обнаруженное этими исследователями взаимодействие между Se и Mn с обнаруженным здесь взаимодействием. В другом исследовании [72] дефицит Mg у крыс повлиял на распределение Se в их тканях (например, снижение содержания Se в плазме, печени и большеберцовой кости). Таким образом, хотя взаимодействие между этими минералами недостаточно изучено, есть данные о том, что уровень Mg в рационе может влиять на метаболизм Se.
Отрицательная взаимосвязь между кажущимся усвоением кальция и цинком в сыворотке (Таблица 5, уравнение (32)) согласуется с данными Миллса [68], который сообщил о снижении концентрации цинка в плазме у ягнят, получавших повышенное количество кальция с кормом. У нежвачных животных есть данные о том, что высокое содержание кальция в корме снижает абсорбцию цинка в присутствии фитата. Однако у жвачных животных снижение абсорбции цинка из-за высокого содержания кальция в рационе, по-видимому, связано с другими причинами, поскольку фитат расщепляется в рубце ферментами фитазы. Необходимы дальнейшие исследования для уточнения взаимодействия между цинком и кальцием у жвачных животных. Взаимодействия между кобальтом и другими минералами, такими как фосфор и натрий, которые можно вывести из уравнений (33) и (34) (Таблица 5), не были обнаружены в других исследованиях, описанных в литературе. Важно дополнительно изучить взаимосвязь между кобальтом и другими минералами, чтобы понять их последствия для стельности и здоровья плода.
Были обнаружены слабые корреляции между минералами в сыворотке (Таблица 6). Корреляция между Fe и K в сыворотке (уравнение (37)) может быть связана с корреляцией между потреблением этих минералов (r = 0,76). Однако положительная взаимосвязь между Mn и Co в сыворотке (уравнение (38)) может быть следствием сильной корреляции между КУ обоих элементов (уравнение (18)).
4.3. Влияние на уровень микроэлементов в печени
Концентрации минералов и микроэлементов в тканях печени находились в пределах значений, указанных Хёрлбертом и др. [73]. Наблюдалась положительная корреляция между Fe в сыворотке и Cu в печени (Таблица 6, уравнение (40)), и оба считались присутствующими в адекватных количествах [74, 75]. Эта корреляция, возможно, является ответом на взаимодействие Cu и Fe в процессе гемопоэза. Этот процесс особенно важен во время стельности, когда образуются новые клетки крови для компенсации резкого увеличения объема крови. Хостелер и др. [8] предположили, что Cu может быть связан с активацией секреции белка утероферрина, который активен в транспорте трехвалентного железа к зародышу у свиней [76]. Взаимосвязь между P в сыворотке и Mo в печени (Таблица 6, уравнение (41)) нелегко объяснить, поскольку информация о взаимодействии между этими минералами отсутствует.
Содержание Mo в печени отрицательно коррелировало с содержанием Se и Zn (Таблица 5, уравнения (35) и (36)). Несмотря на многочисленные корреляции, обнаруженные между Mo и другими элементами, указывающие на взаимодействие между ними, отсутствие данных о роли Mo в питании жвачных животных и его связи с метаболизмом других минералов подчеркивает необходимость дополнительных исследований по этим темам.
Отрицательная корреляция между содержанием Mn и Fe в печени (Таблица 6, уравнение (42)), вероятно, указывает на то, что Fe, по крайней мере частично, ответственен за пограничные уровни Mn, обнаруженные в печени (в среднем 1,85 ± 0,06 мкг/г). Антагонистические отношения между Fe и Mn в печени могут быть результатом общего пути усвоения, общего для двухвалентного железа и Mn (DMT-1) [35]. У человека этот общий путь обсуждался, где абсорбция этих минералов в кишечнике, по-видимому, не различима [77, 78]. Содержание кобальта в печени положительно коррелировало с содержанием Se в печени (r = 0,47; p < 0,05) и Mo в печени (r = 0,54; p < 0,01) (уравнение (43)). Связь между Co и Se у жвачных животных была описана [79], который предположил участие витамина B12 в метаболизме диметилселенида. Взаимодействие добавок Co и Mo изучалось у растущего крупного рогатого скота Чапманом и Киддером [80]. В этом последнем исследовании было замечено, что диетический Co не полностью компенсировал эффекты диетического Mo на уровень гемоглобина. Насколько нам известно, связь между Co и этими минералами в печени не была описана в более ранних исследованиях.
5. Выводы
Представленные здесь уравнения регрессии предоставляют дополнительную информацию о взаимодействии между микроэлементами и дополнительно демонстрируют корреляции между минералами, которые заслуживают дальнейшего изучения. Например, показано, что потребление железа сверх нормы связано с использованием нескольких минералов и микроэлементов: Mo, Cu, Zn, S и Mn. Результаты показывают, что высокое потребление железа привело к повышению уровня Mo в сыворотке крови выше нормы у большинства животных в эксперименте. Кроме того, возможно, что уровни Se в рационе, близкие к требованиям NASEM, влияют на метаболизм Mo. Тот факт, что нетели были стельными, мог усугубить результаты, поскольку их потребность в определенных минералах, вероятно, была повышена. Таким образом, результаты этого исследования не только демонстрируют взаимодействие и использование Cu, Mo, Fe, Se, Mn, Co, Ca, P, Mg, S, Na и K, но и подчеркивают важность обеспечения адекватного уровня минералов в рационах жвачных животных. Особое внимание следует уделить железу в рационе, которое часто присутствует в кормах для скота в количестве, превышающем рекомендуемую суточную норму содержания железа (NASEM), и селену в рационе, который может влиять на усвоение других минералов, когда его содержание приближается к требуемому уровню.
Ссылки
1. Van Emon, M.; Sanford, C.; McCoski, S. Impacts of Bovine Trace Mineral Supplementation on Maternal and Offspring Production and Health. Animals 2020, 10, 2404. [Google Scholar] [CrossRef] [PubMed]
2. Silva, F.L.; Oliveira-Júnior, E.S.; Silva, M.H.M.E.; López-Alonso, M.; Pierangeli, M.A.P. Trace Elements in Beef Cattle: A Review of the Scientific Approach from One Health Perspective. Animals 2022, 12, 2254. [Google Scholar] [CrossRef] [PubMed]
3. Palomares, R.A. Trace Minerals Supplementation with Great Impact on Beef Cattle Immunity and Health. Animals 2022, 12, 2839. [Google Scholar] [CrossRef] [PubMed]
4. Ahola, J.K.; Baker, D.S.; Burns, P.D.; Mortimer, R.G.; Enns, R.M.; Whittier, J.C.; Geary, T.W.; Engle, T.E. Effect of Copper, Zinc, and Manganese Supplementation and Source on Reproduction, Mineral Status, and Performance in Grazing Beef Cattle over a Two-Year Period123. J. Anim. Sci. 2004, 82, 2375–2383. [Google Scholar] [CrossRef]
5. Uchida, K.; Mandebvu, P.; Ballard, C.S.; Sniffen, C.J.; Carter, M.P. Effect of Feeding a Combination of Zinc, Manganese and Copper Amino Acid Complexes, and Cobalt Glucoheptonate on Performance of Early Lactation High Producing Dairy Cows. Anim. Feed. Sci. Technol. 2001, 93, 193–203. [Google Scholar] [CrossRef]
6. Ballantine, H.T.; Socha, M.T.; Tomlinson, D.A.D.J.; Johnson, A.B.; Fielding, A.S.; Shearer, J.K.; Van Amstel, S.R. Effects of Feeding Complexed Zinc, Manganese, Copper, and Cobalt to Late Gestation and Lactating Dairy Cows on Claw Integrity, Reproduction, and Lactation Performance. Prof. Anim. Sci. 2002, 18, 211–218. [Google Scholar] [CrossRef]
7. Phillippo, M.; Humphries, W.R.; Garthwaite, P.H. The Effect of Dietary Molybdenum and Iron on Copper Status and Growth in Cattle. J. Agric. Sci. 1987, 109, 315–320. [Google Scholar] [CrossRef]
8. Hostetler, C.E.; Kincaid, R.L.; Mirando, M.A. The Role of Essential Trace Elements in Embryonic and Fetal Development in Livestock. Vet. J. 2003, 166, 125–139. [Google Scholar] [CrossRef]
9. Harvey, K.M.; Cooke, R.F.; Marques, R.D.S. Supplementing Trace Minerals to Beef Cows during Gestation to Enhance Productive and Health Responses of the Offspring. Animals 2021, 11, 1159. [Google Scholar] [CrossRef]
10. Anas, M.; Diniz, W.J.S.; Menezes, A.C.B.; Reynolds, L.P.; Caton, J.S.; Dahlen, C.R.; Ward, A.K. Maternal Mineral Nutrition Regulates Fetal Genomic Programming in Cattle: A Review. Metabolites 2023, 13, 593. [Google Scholar] [CrossRef]
11. Hazell, T. Minerals in Foods: Dietary Sources, Chemical Forms, Interactions, Bioavailability. In Minerals in Food and Nutritional Topics (Book Series: World Review of Nutrition and Dietetics); Bourne, G.H., Ed.; S. Karger AG: Basel, Switzerland, 1985; Volume 46, pp. 1–123. [Google Scholar]
12. Goff, J.P. Invited Review: Mineral Absorption Mechanisms, Mineral Interactions That Affect Acid–Base and Antioxidant Status, and Diet Considerations to Improve Mineral Status. J. Dairy Sci. 2018, 101, 2763–2813. [Google Scholar] [CrossRef] [PubMed]
13. Fairweather-Tait, S.; Hurrell, R.F. Bioavailability of Minerals and Trace Elements. Nutr. Res. Rev. 1996, 9, 295–324. [Google Scholar] [CrossRef] [PubMed]
14. Spears, J.W. Trace Mineral Bioavailability in Ruminants. J. Nutr. 2003, 133, 1506S–1509S. [Google Scholar] [CrossRef] [PubMed]
15. Weiss, W.P.; Hansen, S.L. Invited Review: Limitations to Current Mineral Requirement Systems for Cattle and Potential Improvements. J. Dairy Sci. 2024, 107, 10099–10114. [Google Scholar] [CrossRef]
16. Graham, T.W. Trace Element Deficiencies in Cattle. Vet. Clin. N. Am. Food Anim. Pract. 1991, 7, 153–215. [Google Scholar] [CrossRef]
17. Duplessis, M.; Royer, I. Mini-Review: The Importance of an Integrated Approach to Assess Trace Mineral Feeding Practices in Dairy Cows. Front. Anim. Sci. 2023, 4, 1155361. [Google Scholar] [CrossRef]
18. Ledoux, D.R.; Shannon, M.C. Bioavailability and Antagonists of Trace Minerals in Ruminant Metabolism. In Proceedings of the Florida Ruminant Nutrition Symposium, Gainesville, FL, USA, 1–2 February 2005; pp. 23–37. [Google Scholar]
19. López-Alonso, M.; Miranda, M. Copper Supplementation, a Challenge in Cattle. Animals 2020, 10, 1890. [Google Scholar] [CrossRef]
20. Bremner, I.; Humphries, W.R.; Phillippo, M.; Walker, M.J.; Morrice, P.C. Iron-Induced Copper Deficiency in Calves: Dose-Response Relationships and Interactions with Molybdenum and Sulphur. Anim. Sci. 1987, 45, 403–414. [Google Scholar] [CrossRef]
21. Knowles, S.O.; Grace, N.D. A Recent Assessment of the Elemental Composition of New Zealand Pastures in Relation to Meeting the Dietary Requirements of Grazing Livestock. J. Anim. Sci. 2014, 92, 303–310. [Google Scholar] [CrossRef]
22. Spears, J.W.; Brandao, V.L.N.; Heldt, J. Invited Review: Assessing Trace Mineral Status in Ruminants, and Factors That Affect Measurements of Trace Mineral Status. Appl. Anim. Sci. 2022, 38, 252–267. [Google Scholar] [CrossRef]
23. Beck, P.A.; Hall, J.O. Vitamin and Trace Element Nutrition of Stocker Cattle on Small Grain and Winter Annual Pastures. Vet. Clin. N. Am. Food Anim. Pract. 2023, 39, 491–504. [Google Scholar] [CrossRef] [PubMed]
24. Swecker, W.S. Trace Mineral Supplementation of Beef Cattle in Pasture Environments. Vet. Clin. N. Am. Food Anim. Pract. 2023, 39, 459–469. [Google Scholar] [CrossRef] [PubMed]
25. Hidiroglou, M.; Knipfel, J.E. Maternal-Fetal Relationships of Copper, Manganese, and Sulfur in Ruminants. A Review. J. Dairy Sci. 1981, 64, 1637–1647. [Google Scholar] [CrossRef] [PubMed]
26. Barone, A.; Ebesh, O.; Harper, R.G.; Wapnir, R.A. Placental Copper Transport in Rats: Effects of Elevated Dietary Zinc on Fetal Copper, Iron and Metallothionein. J. Nutr. 1998, 128, 1037–1041. [Google Scholar] [CrossRef]
27. Vieira-Neto, A.; Lean, I.J.; Santos, J.E.P. Periparturient Mineral Metabolism: Implications to Health and Productivity. Animals 2024, 14, 1232. [Google Scholar] [CrossRef]
28. Canadian Council on Animal Care. Guide to the Care and Use of Experimental Animals; Canadian Council on Animal Care: Ottawa, ON, Canada, 1993; ISBN 0919087183. [Google Scholar]
29. Association of Official Analytical Chemists. Official Methods of Analysis of AOAC International, 18th ed.; Horwitz, W., Ed.; AOAC International: Gaithersburg, MD, USA, 2005; ISBN 0-935584-77-3. [Google Scholar]
30. Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy. Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
31. Dhanoa, M.S.; López, S.; France, J. Linear Models for Determining Digestibility. In Mathematical Modelling in Animal Nutrition; France, J., Kebreab, E., Eds.; CABI: Wallingford, UK, 2008; pp. 12–46. [Google Scholar]
32. Greenwood, S.L.; Wright, T.C.; Purdie, N.G.; Doelman, J.; Cant, J.P.; McBride, B.W. Lactation Induces Upregulation of the Ubiquitin-Mediated Proteolytic Pathway in Skeletal Muscle of Dairy Cows but Does Not Alter Hepatic Expression. Can. J. Anim. Sci. 2009, 89, 309–313. [Google Scholar] [CrossRef]
33. Dias, R.S.; López, S.; Montanholi, Y.R.; Smith, B.; Haas, L.S.; Miller, S.P.; France, J. A Meta-Analysis of the Effects of Dietary Copper, Molybdenum, and Sulfur on Plasma and Liver Copper, Weight Gain, and Feed Conversion in Growing-Finishing Cattle. J. Anim. Sci. 2013, 91, 5714–5723. [Google Scholar] [CrossRef]
34. National Academies of Sciences, Engineering, and Medicine (NASEM). Nutrient Requirements of Beef Cattle, 8th Revised Edition; The National Academies Press: Washington, DC, USA, 2016; ISBN 978-0-309-31702-3. [Google Scholar]
35. Hansen, S.L. Nutritional Interrelationships between Iron, Copper and Manganese in Domestic Livestock; NC State University: Raleigh, NC, USA, 2008. [Google Scholar]
36. Hurley, W.L.; Doane, R.M. Recent Developments in the Roles of Vitamins and Minerals in Reproduction. J. Dairy Sci. 1989, 72, 784–804. [Google Scholar] [CrossRef]
37. Thorndyke, M.P.; Guimaraes, O.; Tillquist, N.M.; Zervoudakis, J.T.; Engle, T.E. Molybdenum Exposure in Drinking Water vs Feed Impacts Apparent Absorption of Copper Differently in Beef Cattle Consuming a High-Forage Diet. Biol. Trace Elem. Res. 2021, 199, 2913–2918. [Google Scholar] [CrossRef]
38. Seelig, M.S. Review: Relationships of Copper and Molybdenum to Iron Metabolism. Am. J. Clin. Nutr. 1972, 25, 1022–1037. [Google Scholar] [CrossRef] [PubMed]
39. Henry, P.R.; Miles, R.D. Interactions among the Trace Minerals. Ciênc Anim. Bras. 2006, 1, 95–106. Available online: https://revistas.ufg.br/vet/article/view/251 (accessed on 8 January 2025).
40. Gardiner, M.; Nicol, H. Cobalt-selenium Interactions in the Nutrition of the Rat. Aust. J. Exp. Biol. Med. Sci. 1971, 49, 291–296. [Google Scholar] [CrossRef]
41. Ammerman, C.B. Recent Developments in Cobalt and Copper in Ruminant Nutrition: A Review. J. Dairy Sci. 1970, 53, 1097–1107. [Google Scholar] [CrossRef]
42. Weisstock, S.R. The Establishment of a Dietary Interaction between Molybdenum and Selenium Based on Weight Gain and Feed Consumption in Broilers. Ph.D. Thesis, University of British Columbia, Vancouver, BC, Canada, 1980. [Google Scholar]
43. Lee, Y.H.; Layman, D.K.; Bell, R.R.; Norton, H.W. Response of Glutathione Peroxidase and Catalase to Excess Dietary Iron in Rats. J. Nutr. 1981, 111, 2195–2202. [Google Scholar] [CrossRef]
44. Ivan, M.; Grieve, C.M. Effects of Zinc, Copper, and Manganese Supplementation of High-Concentrate Ratio on Gastrointestinal Absorption of Copper and Manganese in Holstein Calves. J. Dairy Sci. 1976, 59, 1764–1768. [Google Scholar] [CrossRef]
45. Suttle, N. Mineral Nutrition of Livestock, 4th ed.; Suttle, N., Ed.; CAB International: Wallingford, UK, 2010; ISBN 9781845934736. [Google Scholar]
46. Abdelrahman, M.M.; Kincaid, R.L. Deposition of Copper, Manganese, Zinc, and Selenium in Bovine Fetal Tissue at Different Stages of Gestation. J. Dairy Sci. 1993, 76, 3588–3593. [Google Scholar] [CrossRef]
47. Graham, T.W.; Thurmond, M.C.; Mohr, F.C.; Holmberg, C.A.; Anderson, M.L.; Keen, C.L. Relationships between Maternal and Fetal Liver Copper, Iron, Manganese, and Zinc Concentrations and Fetal Development in California Holstein Dairy Cows. J. Vet. Diagn. Investig. 1994, 6, 77–87. [Google Scholar] [CrossRef]
48. De Haan, J.B.; Tymms, M.J.; Cristiano, F.; Kola, I. Expression of Copper/Zinc Superoxide Dismutase and Glutathione Peroxidase in Organs of Developing Mouse Embryos, Fetuses, and Neonates. Pediatr. Res. 1994, 35, 188–195. [Google Scholar] [CrossRef][Green Version]
49. Al-Gubory, K.H.; Bolifraud, P.; Germain, G.; Nicole, A.; Ceballos-Bicot, I. Antioxidant Enzymatic Defence Systems in Sheep Corpus Luteum throughout Pregnancy. Reproduction 2004, 128, 767–774. [Google Scholar] [CrossRef]
50. Hansen, S.L.; Trakooljul, N.; Liu, H.-C.S.; Hicks, J.A.; Ashwell, M.S.; Spears, J.W. Proteins Involved in Iron Metabolism in Beef Cattle Are Affected by Copper Deficiency in Combination with High Dietary Manganese, but Not by Copper Deficiency Alone1. J. Anim. Sci. 2010, 88, 275–283. [Google Scholar] [CrossRef] [PubMed]
51. Cammack, R.; Barber, M.J.; Bray, R.C. Oxidation-Reduction Potentials of Molybdenum, Flavin and Iron-Sulphur Centres in Milk Xanthine Oxidase. Biochem. J. 1976, 157, 469–478. [Google Scholar] [CrossRef] [PubMed]
52. Becker, G.; Huebers, H.; Rummel, W. Intestinal Absorption of Cobalt and Iron: Mode of Interaction and Subcellular Distribution. Blut 1979, 38, 397–406. [Google Scholar] [CrossRef] [PubMed]
53. Cathcart, E.B.; Shelford, J.A.; Peterson, R.G. The Effects of Calcium Carbonate on the Apparent Digestibility, Serum Concentration and Apparent Retention of Dietary Minerals in Dairy Cattle. Can. J. Anim. Sci. 1983, 63, 173–180. [Google Scholar] [CrossRef]
54. Kiela, P.R.; Ghishan, F.K. Molecular Mechanisms of Intestinal Transport of Calcium, Phosphate, and Magnesium. In Physiology of the Gastrointestinal Tract, 6th ed.; Said, H.M., Ed.; Academic Press: New York, NY, USA, 2018; pp. 1405–1449. [Google Scholar] [CrossRef]
55. Chicco, C.F.; Ammerman, C.B.; Feaster, J.P.; Dunavant, B.G. Nutritional Interrelationships of Dietary Calcium, Phosphorus and Magnesium in Sheep. J. Anim. Sci. 1973, 36, 986–993. [Google Scholar] [CrossRef]
56. Littledike, E.T.; Goff, J. Interactions of Calcium, Phosphorus, Magnesium and Vitamin D That Influence Their Status in Domestic Meat Animals. J. Anim. Sci. 1987, 65, 1727–1743. [Google Scholar] [CrossRef]
57. Fontenot, J.P.; Allen, V.G.; Bunce, G.E.; Goff, J.P. Factors Influencing Magnesium Absorption and Metabolism in Ruminants. J. Anim. Sci. 1989, 67, 3445. [Google Scholar] [CrossRef]
58. House, W.A.; Van Campen, D. Magnesium Metabolism of Sheep Fed Different Levels of Potassium and Citric Acid. J. Nutr. 1971, 101, 1483–1492. [Google Scholar] [CrossRef]
59. Greene, L.W.; Fontenot, J.P.; Webb, K.E. Site of Magnesium and Other Macromineral Absorption in Steers Fed High Levels of Potassium. J. Anim. Sci. 1983, 57, 503–510. [Google Scholar] [CrossRef]
60. Khorasani, G.R.; Armstrong, D.G. Effect of Sodium and Potassium Level on the Absorption of Magnesium and Other Macrominerals in Sheep. Livest. Prod. Sci. 1990, 24, 223–235. [Google Scholar] [CrossRef]
61. Goff, J.P. Calcium and Magnesium Disorders. Vet. Clin. N. Am. Food Anim. Pract. 2014, 30, 359–381. [Google Scholar] [CrossRef] [PubMed]
62. Khiaosa-ard, R.; Ottoboni, M.; Verstringe, S.; Gruber, T.; Hartinger, T.; Humer, E.; Bruggeman, G.; Zebeli, Q. Magnesium in Dairy Cattle Nutrition: A Meta-Analysis on Magnesium Absorption in Dairy Cattle and Assessment of Simple Solubility Tests to Predict Magnesium Availability from Supplemental Sources. J. Dairy Sci. 2023, 106, 8758–8773. [Google Scholar] [CrossRef] [PubMed]
63. Durand, M.; Komisarczuk, S. Influence of Major Minerals on Rumen Microbiota. J. Nutr. 1988, 118, 249–260. [Google Scholar] [CrossRef] [PubMed]
64. Evans, J.L.; Davis, G.K. Dietary Phosphorus, Sulfur and Molybdenum and Mineral Composition of Rumen Fluid. J. Anim. Sci. 1966, 25, 1010–1013. [Google Scholar] [CrossRef]
65. Atkins, N.E.; Bleach, E.C.L.; Mackenzie, A.M.; Hargreaves, P.R.; Sinclair, L.A. Mineral Status, Metabolism and Performance of Dairy Heifers Receiving a Combined Trace Element Bolus and out-Wintered on Perennial Ryegrass, Kale or Fodder Beet. Livest. Sci. 2020, 231, 103865. [Google Scholar] [CrossRef]
66. Nockels, C.F.; DeBonis, J.; Torrent, J. Stress Induction Affects Copper and Zinc Balance in Calves Fed Organic and Inorganic Copper and Zinc Sources. J. Anim. Sci. 1993, 71, 2539–2545. [Google Scholar] [CrossRef]
67. Minatel, L.; Carfagnini, J.C. Evaluation of the Diagnostic Value of Plasma Copper Levels in Cattle. Prev. Vet. Med. 2002, 53, 1–5. [Google Scholar] [CrossRef]
68. Mills, C.F. Dietary Interactions Involving the Trace Elements. Annu. Rev. Nutr. 1985, 5, 173–193. [Google Scholar] [CrossRef]
69. Standish, J.F.; Ammerman, C.B.; Palmer, A.Z.; Simpson, C.F. Influence of Dietary Iron and Phosphorus on Performance, Tissue Mineral Composition and Mineral Absorption in Steers. J. Anim. Sci. 1971, 33, 171–178. [Google Scholar] [CrossRef]
70. Ward, G.M. Molybdenum Toxicity and Hypocuprosis in Ruminants: A Review. J. Anim. Sci. 1978, 46, 1078–1085. [Google Scholar] [CrossRef]
71. Hernández-Calva, L.M.; Guerrero-Legarreta, M.I.; Pérez-Chabela, M.L.; López-Arellano, R.; Ramírez-Bribiesca, J.E. Interaction of Dietary Selenium and Magnesium Level on Digestive Function in Lambs Fed High-Concentrate Diets. J. Appl. Anim. Res. 2007, 31, 41–46. [Google Scholar] [CrossRef]
72. Jiménez, A.; Planells, E.; Aranda, P.; Sánchez-Viñas, M.; Llopis, J. Changes in Bioavailability and Tissue Distribution of Selenium Caused by Magnesium Deficiency in Rats. J. Am. Coll. Nutr. 1997, 16, 175–180. [Google Scholar] [CrossRef] [PubMed]
73. Hurlbert, J.L.; Baumgaertner, F.; Menezes, A.C.B.; Bochantin, K.A.; Diniz, W.J.S.; Underdahl, S.R.; Dorsam, S.T.; Kirsch, J.D.; Sedivec, K.K.; Dahlen, C.R. Supplementing Vitamins and Minerals to Beef Heifers during Gestation: Impacts on Mineral Status in the Dam and Offspring, and Growth and Physiological Responses of Female Offspring from Birth to Puberty. J. Anim. Sci. 2024, 102, skae002. [Google Scholar] [CrossRef] [PubMed]
74. Puls, R. Mineral Levels in Animal Health: Diagnostic Data; Sherpa International: Abbotsford, BC, USA, 1989. [Google Scholar]
75. Underwood, E.J.; Suttle, N.F. The Mineral Nutrition of Livestock, 3rd ed.; CABI Publishing: Wallingford, UK, 1999; ISBN 9780851991283. [Google Scholar]
76. Raub, T.J.; Bazer, F.W.; Michael Roberts, R. Localization of the Iron Transport Glycoprotein, Uteroferrin, in the Porcine Endometrium and Placenta by Using Immunocolloidal Gold. Anat. Embryol. 1985, 171, 253–258. [Google Scholar] [CrossRef]
77. Rossander-Hultén, L.; Brune, M.; Sandström, B.; Lönnerdal, B.; Hallberg, L. Competitive Inhibition of Iron Absorption by Manganese and Zinc in Humans. Am. J. Clin. Nutr. 1991, 54, 152–156. [Google Scholar] [CrossRef]
78. Sandström, B. Micronutrient Interactions: Effects on Absorption and Bioavailability. Br. J. Nutr. 2001, 85, S181–S185. [Google Scholar] [CrossRef]
79. Wise, W.R.; Weswig, P.H.; Muth, O.H.; Oldfield, J.E. Dietary Interrelationship of Cobalt and Selenium in Lambs. J. Anim. Sci. 1968, 27, 1462. [Google Scholar] [CrossRef]
80. Chapman, H.L.; Kidder, R.W. Oral Administration of Molybdenum and Cobalt to Brahman-Angus Heifers. J. Anim. Sci. 1963, 22, 985–988. [Google Scholar] [CrossRef]

Dias Betini RS, López S, Montanholi YR, France J. Interactions Between Trace Elements and Macro Minerals in Pregnant Heifers. Agriculture. 2025; 15(2):167. https://doi.org/10.3390/agriculture15020167
Фото: freepik
Перевод статьи « Interactions Between Trace Elements and Macro Minerals in Pregnant Heifers» авторов Dias Betini RS, López S, Montanholi YR, France J., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык

















































































































Комментарии (0)