NPK повышают устойчивость промышленной конопли к щелочному стрессу
Глобальное засоление почв становится все более серьезной проблемой для сельскохозяйственного производства, особенно затрагивая выращивание технических культур на маргинальных землях. Промышленная конопля (Cannabis sativa L.), несмотря на свой экономический потенциал, демонстрирует заметную чувствительность к солево-щелочному стрессу, что ограничивает ее распространение в засоленно-щелочных регионах. В данном исследовании изучались регуляторные эффекты азотных (N), фосфорных (P) и калийных (K) удобрений на рост конопли и питательный гомеостаз в условиях щелочного солевого стресса.
Аннотация
Используя ортогональный экспериментальный план «3414», мы оценили четырнадцать комбинаций NPK в условиях стресса 200 мМ NaHCO₃ — концентрации, определенной в предварительных экспериментах для моделирования умеренного щелочного стресса. Параметры роста растений, распределение биомассы и профили минеральных питательных веществ были проанализированы после обработки в трех биологических повторностях.
Обработка N1P2K2 (N 120 мг·л⁻¹, P 238 мг·л⁻¹, K 348 мг·л⁻¹) значительно улучшила показатели растений, увеличив биомассу побегов на 45,3%, а корней — на 38,7% по сравнению с контролем. Эта оптимальная комбинация поддерживала соотношение K⁺/Na⁺ в листьях выше 1,2 и регулировала гомеостаз Ca²⁺/Mg²⁺, сохраняя соотношение в пределах 2,8–3,2, что указывает на повышенную солеустойчивость. Примечательно, что чрезмерное внесение удобрений (суммарно >400 мг·л⁻¹ питательных веществ) усиливало солевое повреждение, снижая накопление биомассы на 25–30% и нарушая ионный гомеостаз.
Наши результаты выявляют критические пороги внесения NPK для конопли в условиях щелочного стресса и предлагают практические стратегии удобрения для устойчивого возделывания конопли в засоленно-щелочных регионах.
1. Введение
В глобальном масштабе засоление почв является все более серьезной проблемой, и последние оценки свидетельствуют о том, что примерно 950 миллионов гектаров земель были сильно затронуты. Это явление деградации земель является не только результатом естественных изменений окружающей среды, но и прямым следствием несоответствующих методов ведения сельского хозяйства человеком. В частности, неподходящие методы обработки почвы и чрезмерные стратегии удобрения в современном сельском хозяйстве не только усугубили развитие существующего засоления почв, но и вызвали крупномасштабное вторичное засоление, при котором изначально незасоленные земли постепенно превращаются в засоленные почвы из-за деятельности человека. Этот порочный круг не только приводит к значительному снижению эффективности использования земель, но и серьезно влияет на здоровый рост и качество продукции сельскохозяйственных культур, тем самым угрожая глобальной безопасности здоровья растений и долгосрочной устойчивости агроэкосистем [1,2,3]. Конопля (Cannabis sativa L.) широко культивируется во всем мире как важная техническая культура с множеством применений благодаря волокну, маслу и лекарственным соединениям [4,5]. Однако из-за ее чувствительности к засоленно-щелочным условиям она сталкивается с серьезными проблемами для продвижения и возделывания в этих районах [6]. Следовательно, существует настоятельная необходимость в разработке эффективных стратегий управления для улучшения роста и производственных показателей конопли в засоленно-щелочных условиях.
В последние годы, с возрастающим воздействием изменения климата и деградации земель, исследователи все больше сосредотачиваются на механизмах роста и стратегиях адаптации растений в неблагоприятных условиях. Солево-щелочной стресс оказывает множественное воздействие на рост растений, включая снижение эффективности фотосинтеза, нарушение поглощения питательных веществ, нарушение внутриклеточного ионного гомеостаза, гормональный дисбаланс, ограничение роста корней и снижение функций антиоксидантной защитной системы. Совокупное действие этих факторов оказывает глубокое влияние на рост растений [7,8]. Для решения этих проблем управление удобрениями, как эффективная мера для улучшения состояния почвы и питания растений, широко изучалось и применялось для различных культур, таких как картофель (Solanum tuberosum), сорго (Sorghum bicolor) и томат (Solanum lycopersicum) [9,10,11].
В частности, применение трех основных элементов, а именно азота (N), фосфора (P) и калия (K), как было показано, значительно улучшает химические свойства засоленно-щелочных почв и повышает приспособляемость растений к неблагоприятным условиям [12]. Например, N-удобрения в основном способствуют росту растений и формированию листьев, поскольку N является ключевым элементом в составе белков, нуклеиновых кислот и других жизненных молекул и особенно важен для роста стеблей и листьев [13]. P-удобрения в основном влияют на развитие корней и формирование цветов и плодов, потому что P играет важную роль в процессах преобразования энергии (таких как образование и использование АТФ) и делении клеток, таким образом помогая в развитии корней растений и накоплении энергии [14]. K-удобрения играют важную роль в регулировании водного баланса растений и повышении устойчивости растений к болезням. Они также обеспечивают стабильность фотосинтеза и транспорта питательных веществ. K может помочь растениям лучше регулировать открытие и закрытие устьиц, тем самым влияя на эффективность использования воды растениями и фотосинтез [15,16,17]. В то же время между ними также существуют определенные взаимодействия и зависимости. Например, адекватное снабжение N может способствовать синтезу большего количества хлорофилла, тем самым улучшая фотосинтез, в то время как улучшенный фотосинтез требует достаточного количества K для поддержания соответствующей высокой эффективности транспорта воды и питательных веществ. Между тем, адекватное снабжение P также необходимо для обеспечения достаточного количества энергии для поддержки всех процессов роста растений, включая процессы утилизации N и K [18,19]. Следовательно, оптимальные пропорции и нормы внесения этих питательных веществ в специфических неблагоприятных условиях, особенно для такой специальной культуры, как конопля, заслуживают дальнейшего изучения.
Несмотря на эти известные преимущества, взаимодействие N, P и K удобрений на коноплю в условиях щелочного стресса остается малоизученным. Предыдущие исследования в основном были направлены на реакцию на отдельные питательные вещества или бинарные взаимодействия [9,10,20,21], но несколько ключевых вопросов остаются без ответа: (1) Каково оптимальное соотношение N, P и K для роста конопли в условиях специфического щелочного стресса? (2) Как различные комбинации N, P и K влияют на распределение питательных веществ и ионный гомеостаз? (3) Выше какого порога концентрации внесение удобрений становится вредным?
Это исследование использовало метод экспериментального плана "3414" для систематической оценки эффектов различных комбинаций N, P и K удобрений на рост конопли, биомассу, распределение питательных веществ и ионный гомеостаз в условиях щелочного солевого стресса. Экспериментальный план "3414", как метод неполного ортогонального регрессионного планирования с тремя факторами и четырьмя уровнями, имеет преимущества комплексности факторов, гибкости уровней, простоты операции и эффективности анализа и широко используется в сельскохозяйственных экспериментах [22,23]. Используя этот метод, данное исследование может оценить основные эффекты трех факторов удобрения на четырех уровнях с меньшим количеством комбинаций обработок, удовлетворяя профессиональным требованиям для принятия решений по управлению удобрениями и обеспечивая сильную поддержку метода эксперимента для углубленного анализа регуляторного механизма воздействия удобрений на сухую биомассу, накопление и распределение питательных веществ, и выявления взаимосвязи между биомассой и питательными веществами. Эти результаты не только заполняют пробел в исследованиях удобрения конопли в условиях щелочного солевого стресса, но также предоставляют важную теоретическую основу и практическое руководство для устойчивого и стрессоустойчивого возделывания конопли на маргинальных землях.
2. Материалы и методы
2.1. Растительный материал и условия выращивания
В данном исследовании использовался промышленный сорт конопли "Конопля №5" (предоставлен Хэйлунцзянской академией сельскохозяйственных наук). Семена были поверхностно стерилизованы 75% (об./об.) этанолом в течение 30 с, затем 10% гипохлоритом натрия в течение 10 мин, а затем тщательно промыты пять раз стерильной дистиллированной водой. Стерилизованные семена проращивали в субстрате, состоящем из почвы, вермикулита и перлита (3:1:1, об./об./об.), который автоклавировали при 121 °C в течение 20 мин. Субстрат имел следующие характеристики: pH 6,5 ± 0,2, электрическая проводимость (ЕС) 0,8 ± 0,1 дСм·м⁻¹ (измерено в суспензии почва-вода 1:5 с использованием портативного кондуктометра, модель DDS-307A, Shanghai Precision Scientific Instrument Co., Ltd., Шанхай, Китай). Через семь дней после прорастания, когда семядоли полностью раскрылись, однородные сеянцы пересаживали в пластиковые горшки (диаметр 16 см × высота 17 см), содержащие стерилизованную смесь почвы и вермикулита (3:1, об./об.). В каждом горшке на дне были дренажные отверстия для предотвращения переувлажнения. Свойства почвы были следующими: pH 7,2 (измерено в суспензии почва-вода 1:25 с использованием pH-метра, модель PHS-3C, Shanghai Precision Scientific Instrument Co., Ltd., Шанхай, Китай), электрическая проводимость (ЕС) 0,5 дСм·м⁻¹, органическое вещество 15,3 г·кг⁻¹ (определено бихроматным объемным методом) [24], общий азот 0,82 г·кг⁻¹ (определен методом Кьельдаля), доступный фосфор 15,6 мг·кг⁻¹ и доступный калий 125 мг·кг⁻¹ (содержание P и K определяли с использованием масс-спектрометрии с индуктивно связанной плазмой (ICP-MS 7850, Agilent Technologies, Санта-Клара, Калифорния, США). После прореживания на стадии двух листьев поддерживали по два сеянца на горшок.
Эксперимент в горшках проводился в теплице Северо-Восточного лесного университета в следующих условиях выращивания рассады: дневная/ночная температура 25/20 ± 1 °C, относительная влажность 60 ± 5%, фотопериод 16 ч и интенсивность света, которая постепенно увеличивалась от 200 мкмоль·м⁻²·с⁻¹ до 800 мкмоль·м⁻²·с⁻¹ в течение первых 2 ч фотопериода и постепенно снижалась в течение последних 2 ч. Влажность почвы управлялась путем взвешивания горшков (глубина: 17 см) и добавления воды для поддержания среднего объемного содержания воды (VWC) на уровне 70–80% во всем объеме почвы.
2.2. Обработка щелочным стрессом
На основе предварительных экспериментов по оценке физиологических реакций на концентрации NaHCO₃ (0, 50, 100, 200 и 300 мМ) в качестве оптимального уровня стресса была выбрана концентрация 200 мМ [25,26]. При этой концентрации уровень малонового диальдегида (МДА) в листьях значительно увеличился [27], оказавшись на 85% выше, чем в контрольной группе, в то время как выживаемость растений оставалась выше 90%. Обработку щелочным стрессом применяли к 25-дневным, равномерно растущим сеянцам (в 25 дней растения прошли раннюю стадию рассады, рост стабилизировался, и размер растений стал подходящим для обработки стрессом и возделывания в горшках). Раствор NaHCO₃ (200 мМ, pH 8,7 ± 0,1, ЕС 21,3 ± 0,2 дСм·м⁻¹) применяли каждые 4 дня по 250 мл за одно применение, всего шесть применений. Раствор применяли утром (с 8:00 до 9:00), чтобы избежать потенциального взаимодействия с тепловым стрессом. ЕС просачивающейся воды контролировали после каждого применения для обеспечения постоянного уровня стресса.
После обработки стрессом, на основе оптимальных концентраций элементов азота (N), фосфора (P) и калия (K) для роста конопли, полученных в предыдущих экспериментах, был приготовлен модифицированный питательный раствор Хогланда [28,29], содержащий 0,24 г·л⁻¹ N, 0,238 г·л⁻¹ P и 0,348 г·л⁻¹ K, и с использованием ортогонального экспериментального плана "3414" были установлены обработки с различными соотношениями N, P и K (Таблица 1). Эксперимент включал три фактора (N, P и K), каждый с четырьмя уровнями: 0 (без удобрения), 1 (50% от оптимального уровня удобрения), 2 (оптимальный уровень удобрения) и 3 (150% от оптимального уровня удобрения). Оптимальный уровень удобрения определялся как уровень внесения N, P и K, который максимизировал биомассу растений в предварительном эксперименте и служил основой для определения различных уровней удобрения в основном эксперименте. Среди этих 14 групп обработок обработки T2, T3, T6 и T11 представляли четыре уровня удобрения N; T4, T5, T6 и T7 представляли четыре уровня удобрения P; и T8, T9, T6 и T10 представляли четыре уровня удобрения K. Концентрации N, P и K контролировались путем регулирования количеств NH₄NO₃, NaH₂PO₄ и K₂SO₄ в питательном растворе (другие компоненты питательного раствора показаны в Таблице 2). Каждая обработка имела три повторности, всего 42 горшка. Питательный раствор меняли каждые 3 дня, всего семь обработок (21 день). Рост и физиологические показатели измеряли в конце обработки.
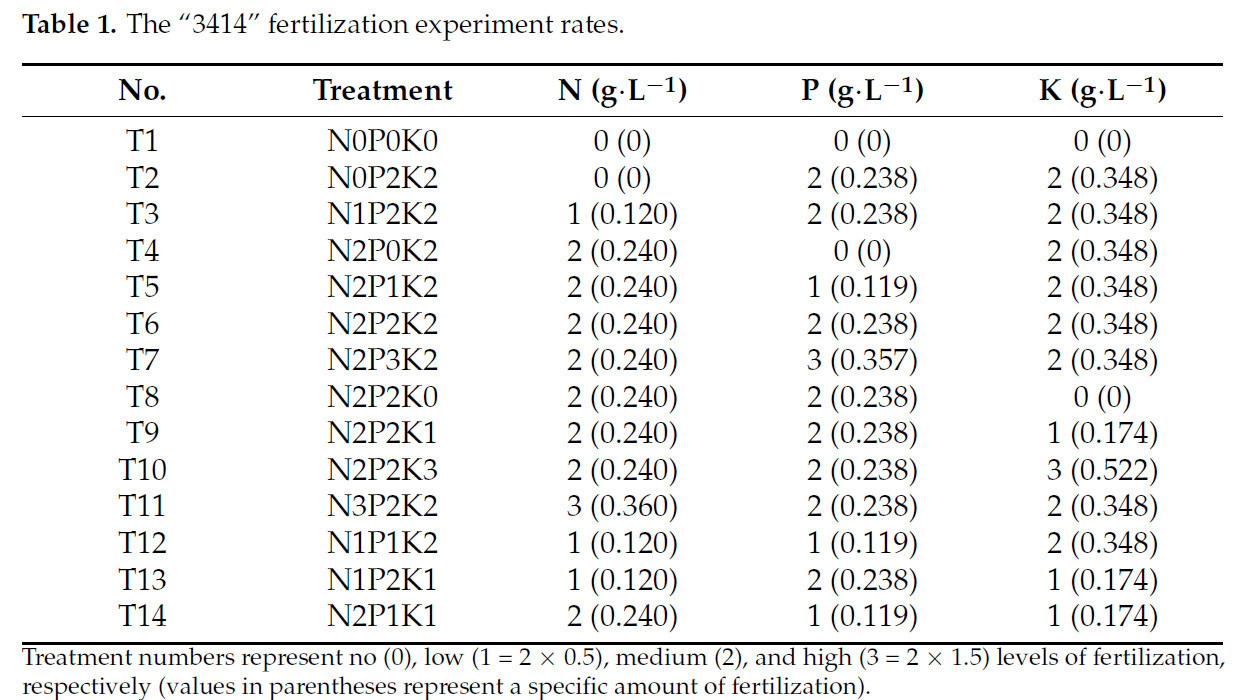
Таблица 1. Нормы эксперимента по удобрению "3414".
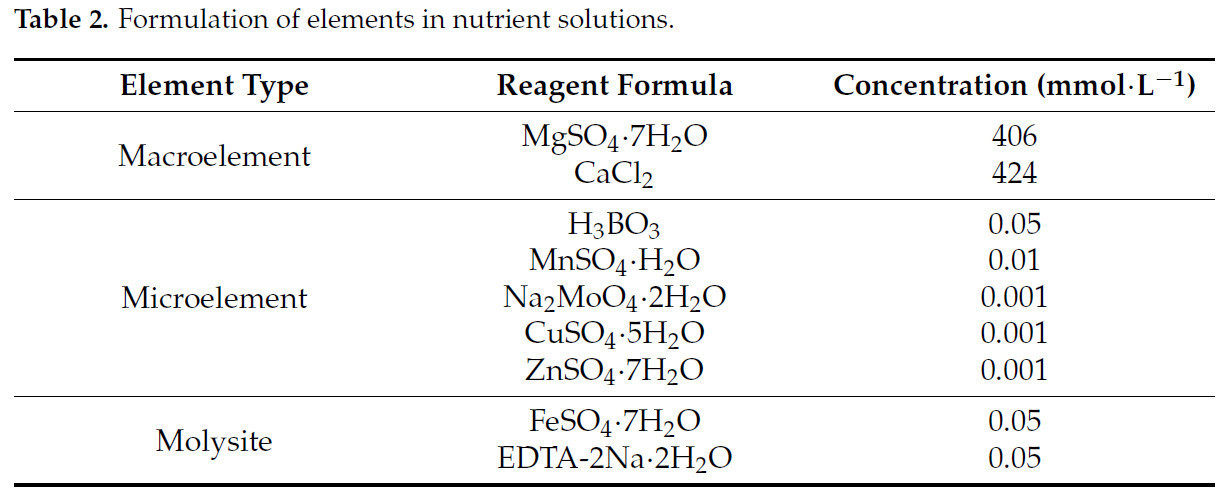
Таблица 2. Состав элементов в питательных растворах.
Весь эксперимент проводился с мая по сентябрь 2023 года, с тремя независимыми биологическими повторностями, выполняемыми с интервалом в 15 дней. Каждая повторность следовала той же схеме стерилизации семян, проращивания и обработки стрессом. Условия окружающей среды постоянно контролировались с использованием автоматической системы контроля климата и поддерживались постоянными в различных повторностях.
2.3. Параметры роста и анализ биомассы
Высоту растений измеряли от поверхности почвы до апикальной точки роста с помощью измерительной ленты, в то время как диаметр стебля определяли на высоте 5 см над поверхностью почвы с помощью цифровых штангенциркулей (CD-15CP, Mitutoyo, Кавасаки, Канагава, Япония). Максимальную длину корня измеряли после тщательной промывки корневых систем от субстрата. Для определения биомассы растения разделяли на корни, стебли и листья. Корневые системы осторожно промывали деионизированной водой для удаления частиц субстрата, слегка промокали бумажными полотенцами и регистрировали их свежий вес. Затем все части растений сушили в сушильном шкафу при 70 °C в течение нескольких часов до достижения постоянного веса (±0,01 г изменение между двумя последовательными измерениями с интервалом в 12 ч). Сухой вес определяли с помощью аналитических весов (BSA124S-CW, Sartorius, Гёттинген, Германия). Соотношение корень/надземная часть = масса сухой биомассы подземной части / масса сухой биомассы надземной части сеянца × 100%.
2.4. Анализ минеральных питательных веществ
Высушенные образцы растений измельчали в мелкий порошок с помощью ступки и пропускали через сито 0,25 мм. Общее содержание азота определяли с использованием автоматического анализатора Кьельдаля (KDN-19K, Shanghai Xianqian Instrument Co., Ltd., Шанхай, Китай) после кислотного разложения. Для каждого образца 0,2 г высушенного материала разлагали смесью H₂SO₄ и H₂O₂ при 260 °C.
Для других минеральных элементов образцы (0,1 г) разлагали в смеси ультрачистой HNO₃ и H₂O₂ (4:1, об./об.) с использованием графитового минерализатора (YXGD-I-36, Yunxiang (Tianjin) Instruments Ltd., Тяньцзинь, Китай). Программа разложения была установлена следующим образом: нагрев до 180 °C за 15 мин, затем выдержка при 180 °C в течение 180 мин. Концентрации Na⁺, K⁺, Ca²⁺, Mg²⁺ и P анализировали с использованием масс-спектрометрии с индуктивно связанной плазмой (ICP-MS 7850, Agilent Technologies, Санта-Клара, Калифорния, США). Контроль качества выполняли с использованием сертифицированного стандартного образца (GBW10015, GSB-6, Национальный институт метрологии, Пекин, Китай), анализируемого вместе с образцами. Процент извлечения для всех элементов варьировался от 95% до 105%.
2.5. Статистический анализ
Все данные представлены как среднее значение ± стандартное отклонение (SD) трех независимых биологических повторностей. Перед анализом данные проверяли на однородность дисперсии с помощью критерия Левена. Однофакторный дисперсионный анализ (ANOVA) выполняли с использованием программного обеспечения SPSS (версия 26.0, IBM, Армонк, Нью-Йорк, США). Когда ANOVA показывал значительные различия, средние значения сравнивали с использованием критерия множественного диапазона Дункана при p < 0,05. Результаты теста на однородность дисперсии и анализа значимости представлены в таблицах S1 и S2, соответственно, в дополнительных материалах. Для анализа ортогонального экспериментального плана использовали метод анализа размаха (Range Analysis) для определения влияния различных уровней факторов на переменную отклика. Этот метод использовали для определения оптимальной комбинации факторов и порядка их влияния. Расчеты анализа размаха выполняли с использованием Excel 2021 (Microsoft, Редмонд, Вашингтон, США). Путем сравнения значений размаха для каждого уровня фактора (т.е. разницы между максимальным и минимальным средними значениями на каждом уровне) количественно оценивали значимость факторов [30,31]. Графики были построены с использованием Origin Pro 2022 (OriginLab Corporation, Нортгемптон, Массачусетс, США).
3. Результаты
3.1. Влияние NPK на рост конопли в условиях стресса NaHCO₃
3.1.1. Влияние различных комбинаций NPK на показатели роста и биомассу конопли в условиях стресса NaHCO₃
Взаимодействие N, P и K значительно повлияло на рост и распределение биомассы конопли в условиях стресса NaHCO₃ (Таблица 3, Рисунок 1A). Среди всех обработок N1P2K2 (3#) увеличил высоту растения, диаметр стебля и длину корня на 22,40%, 18,67% и 33,25% соответственно по сравнению с контролем (1#, N0P0K0), достигнув оптимальных уровней. Обработка N2P0K2 (4#) привела к наибольшему количеству эффективных листьев, которое увеличилось на 48,28% по сравнению с контролем. Обработка 3# увеличила сухой вес корня, листа и всего растения на 1,53%, 190,20% и 177,79% соответственно. Эта обработка показала наилучшие результаты по биомассе.
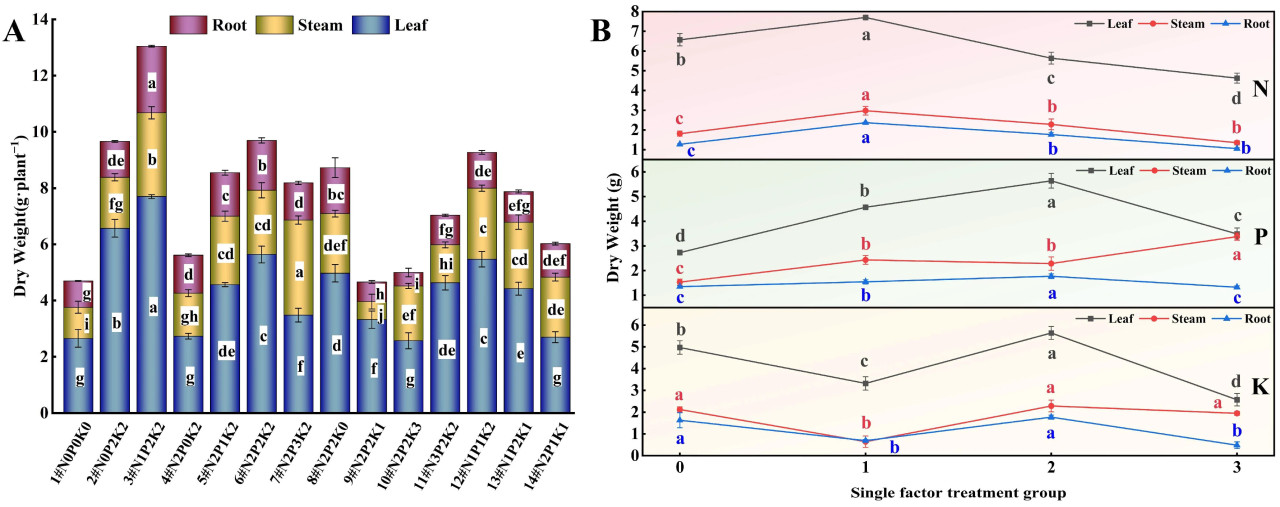
Рисунок 1. Влияние азота (N), фосфора (P) и калия (K) на биомассу конопли в условиях стресса NaHCO₃. (A) Четырнадцать наборов соотношений NPK (#1–#14); (B) Единичный фактор удобрений N, P и K. Разные строчные буквы указывают на значительные различия между обработками для одного и того же органа растения при p < 0,05.
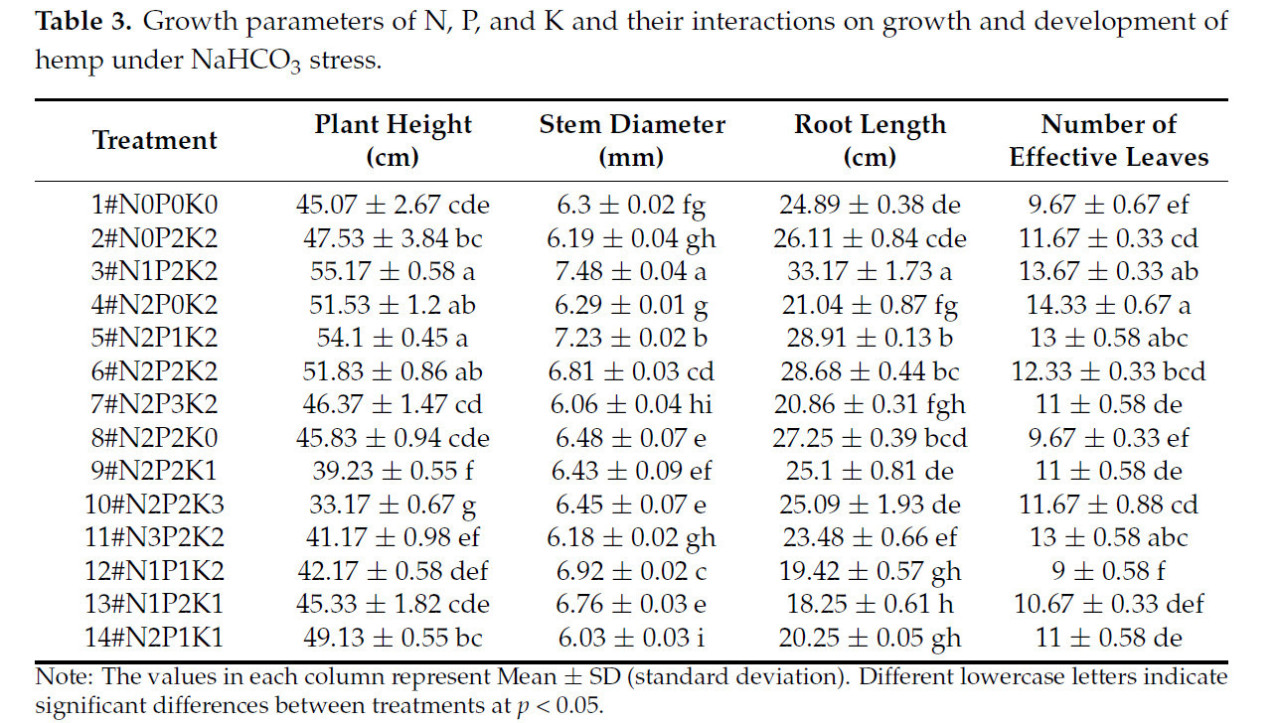
Таблица 3. Параметры роста N, P и K и их взаимодействия на рост и развитие конопли в условиях стресса NaHCO₃.
В эксперименте с единичным фактором N (Таблица 3, Рисунок 1B) высота растения, диаметр стебля, длина корня, а также сухой вес корня, стебля и всего растения показали квадратичный отклик, сначала увеличиваясь, а затем уменьшаясь с увеличением внесения N (N1 > N2 > N0 > N3). Количество и сухой вес листьев были наибольшими при обработке N1, затем N3, N2 и N0. Уровень N1 (120 мг·л⁻¹) увеличил различные параметры роста на 16,06–86,13% по сравнению с контролем без N (2#N0P2K2), в то время как чрезмерное внесение N (N3, 360 мг·л⁻¹) привело к снижению на 0,11–13,39%.
Обработки P вызывали разные реакции в различных органах растений (Таблица 3, Рисунок 1B). С увеличением внесения P высота растения, диаметр стебля и длина корня показали тенденцию сначала к увеличению, а затем к уменьшению (P1 > P2 > P0 > P3), в то время как количество эффективных листьев постоянно уменьшалось (P0 > P1 > P2 > P3). Обработка P1 (119 мг·л⁻¹) увеличила высоту растения, диаметр стебля и длину корня на 4,98%, 14,94% и 37,42% соответственно по сравнению с контролем без P (4#), в то время как обработка P3 (522 мг·л⁻¹) уменьшила их на 10,03%, 3,66% и 0,84% соответственно. Сухой вес корня был наибольшим при обработке P2, затем P1, P0 и P3. Сухой вес листа и общий сухой вес были наибольшими при обработке P2, затем P1, P3 и P0. Сухой вес стебля увеличивался с увеличением концентрации P (P3 > P2 > P1 > P0).
Эксперимент с единичным фактором K показал (Таблица 3, Рисунок 1B), что с увеличением внесения K высота растения, длина корня, а также сухой вес корня и листьев показали тенденцию сначала к увеличению, а затем к уменьшению (K2 > K0 > K1 > K3). Диаметр стебля, сухой вес стебля и общий сухой вес были наибольшими при обработке K2, затем K0, K3 и K1. Обработка K2 (348 мг·л⁻¹) увеличила различные параметры роста на 5,04–13,48% по сравнению с контролем без K (8#N2P2K0), достигнув оптимальных уровней, в то время как чрезмерное внесение K (K3, 522 мг·л⁻¹) ингибировало рост.
Дополнительно, обработки питательными веществами значительно изменили соотношение корень/надземная часть, при этом обработка N1 привела к наибольшему соотношению корень/надземная часть, что является ключевым для усвоения питательных веществ в условиях стресса. За исключением T1, все обработки значительно увеличили сухую биомассу органов и общую, что указывает на то, что сбалансированное удобрение является ключевым для смягчения щелочного стресса.
3.1.2. Анализ размаха влияния удобрения NPK на показатели роста конопли в условиях стресса NaHCO₃
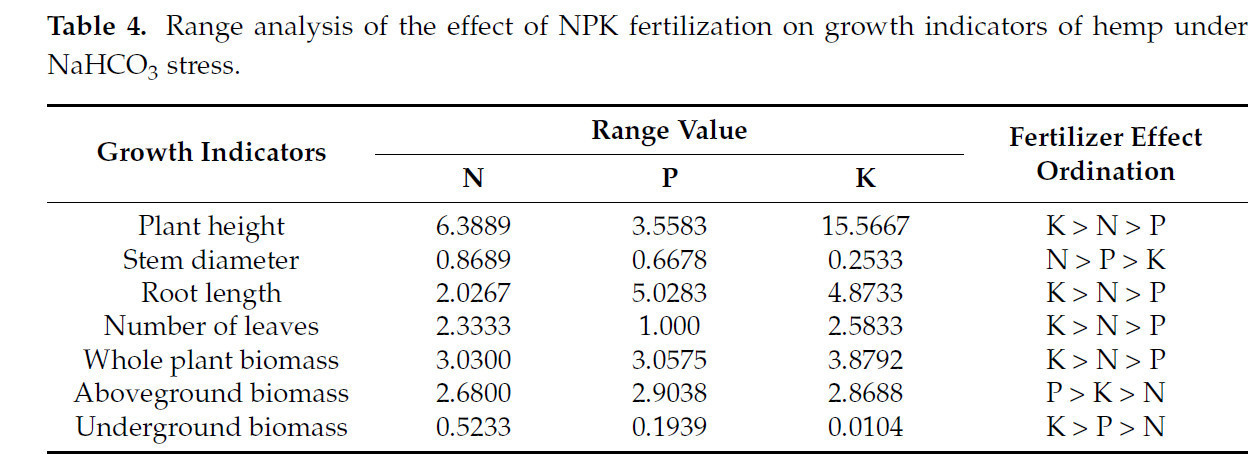
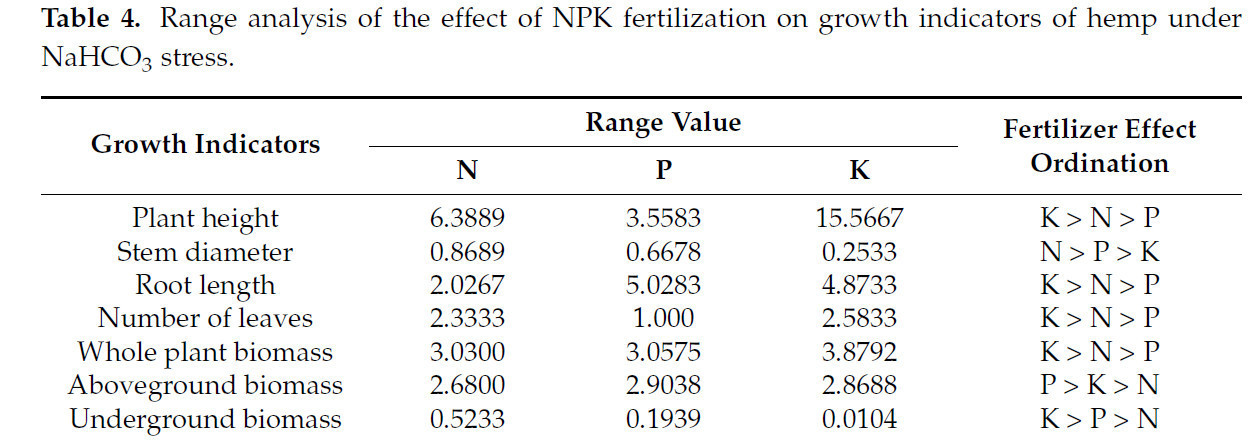
Метод анализа крайних разностей использовали для анализа эффектов различных способов удобрения на показатели роста конопли в условиях щелочного солевого стресса (Таблица 4). Результаты показали, что N оказал наибольшее влияние на толщину стебля, P оказал наибольшее влияние на сухой вес надземной части, а K оказал наибольшее влияние на высоту растения, длину корня, количество листьев, сухой вес подземной части и общий сухой вес растения.
Таблица 4. Анализ размаха влияния удобрения NPK на показатели роста конопли в условиях стресса NaHCO₃.
3.2. Влияние внесения N, P и K на ионное содержание конопли в условиях стресса NaHCO₃
3.2.1. Na⁺
3.2. Влияние внесения N, P и K на ионное содержание конопли в условиях стресса NaHCO₃3.2.1. Na⁺При взаимодействии N, P и K (Рисунок 2A) наблюдались значительные различия в содержании Na⁺ в корнях, стеблях и листьях конопли, подвергнутой щелочному стрессу, среди 14 обработок удобрением. Содержание Na⁺ в корнях было наименьшим при обработке N2P2K3 (10#), что на 82,86% ниже, чем в контрольной группе (1#, N0P0K0). Содержание Na⁺ в стеблях и листьях было наименьшим при обработке N2P2K2 (6#), что на 66,34% и 68,19% ниже, чем в контрольной группе, соответственно.
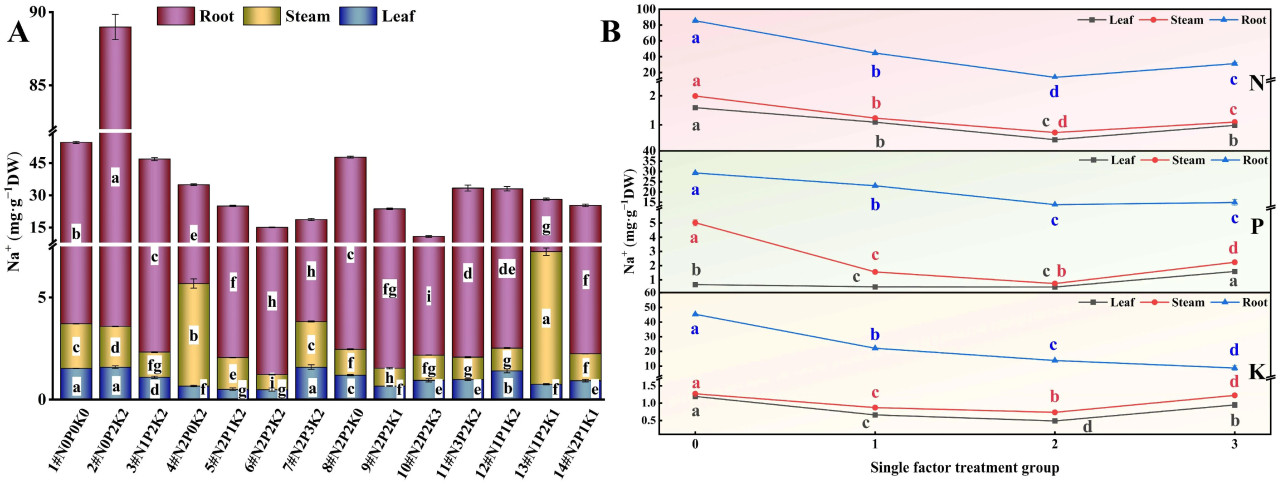
Рисунок 2. Влияние внесения N, P и K на содержание Na⁺ в конопле в условиях стресса NaHCO₃. (A) Четырнадцать наборов соотношений NPK (#1–#14); (B) Единичный фактор удобрений N, P и K. Разные строчные буквы указывают на значительные различия между обработками для одного и того же органа растения при p < 0,05.
При рассмотрении только эффекта N (Рисунок 2B) содержание Na⁺ в различных органах конопли, подвергнутой щелочному стрессу, следовало порядку N0 > N1 > N3 > N2. С увеличением внесения N содержание Na⁺ в корнях, стеблях и листьях постепенно уменьшалось, достигая самого низкого уровня при N2. Это указывает на то, что внесение N может снизить накопление Na⁺ в конопле, подвергнутой щелочному стрессу.
В эксперименте с единичным фактором P (Рисунок 2B) в условиях щелочного стресса содержание Na⁺ в корнях конопли следовало порядку P0 > P1 > P3 > P2, содержание Na⁺ в стеблях следовало P0 > P3 > P1 > P2, а содержание Na⁺ в листьях следовало P3 > P0 > P1 > P2. С увеличением внесения P содержание Na⁺ в корнях и стеблях постепенно уменьшалось, достигая самого низкого уровня при P2. Содержание Na⁺ в листьях сначала уменьшалось, а затем увеличивалось, также достигая самого низкого уровня при P2. Эти результаты предполагают, что умеренный уровень внесения P наиболее благоприятен для снижения содержания Na⁺ в различных органах конопли, подвергнутой щелочному стрессу, но чрезмерное внесение P может увеличить накопление Na⁺ в листьях.
Результаты эксперимента с единичным фактором K (Рисунок 2B) показали, что содержание Na⁺ в корнях конопли, подвергнутой щелочному стрессу, следовало порядку K0 > K1 > K2 > K3, в то время как содержание Na⁺ в стеблях и листьях следовало K0 > K3 > K1 > K2. Содержание Na⁺ в различных органах при внесении K было ниже, чем в контроле без K, при этом содержание Na⁺ в корнях было наименьшим на уровне K3, а содержание Na⁺ в стеблях и листьях было наименьшим на уровне K2. Эти результаты указывают на то, что внесение K помогает снизить уровень Na⁺ в конопле, подвергнутой щелочному стрессу.
В целом, содержание Na⁺ в различных органах конопли следовало порядку корень > стебель > лист.
3.2.2. K⁺
При взаимодействии N, P и K (Рисунок 3A) наблюдались значительные различия в содержании K⁺ в корнях, стеблях и листьях конопли, подвергнутой щелочному стрессу, среди 14 обработок удобрением. Содержание K⁺ в корнях было наибольшим при обработке N3P2K2 (11#), что на 214,33% выше, чем в контроле (1#, N0P0K0). Содержание K⁺ в стеблях было наибольшим при обработке N2P2K2 (6#), что на 69,71% выше, чем в контроле. Содержание K⁺ в листьях достигло максимума при обработке N2P1K2 (5#), что на 293,61% выше, чем в контроле.
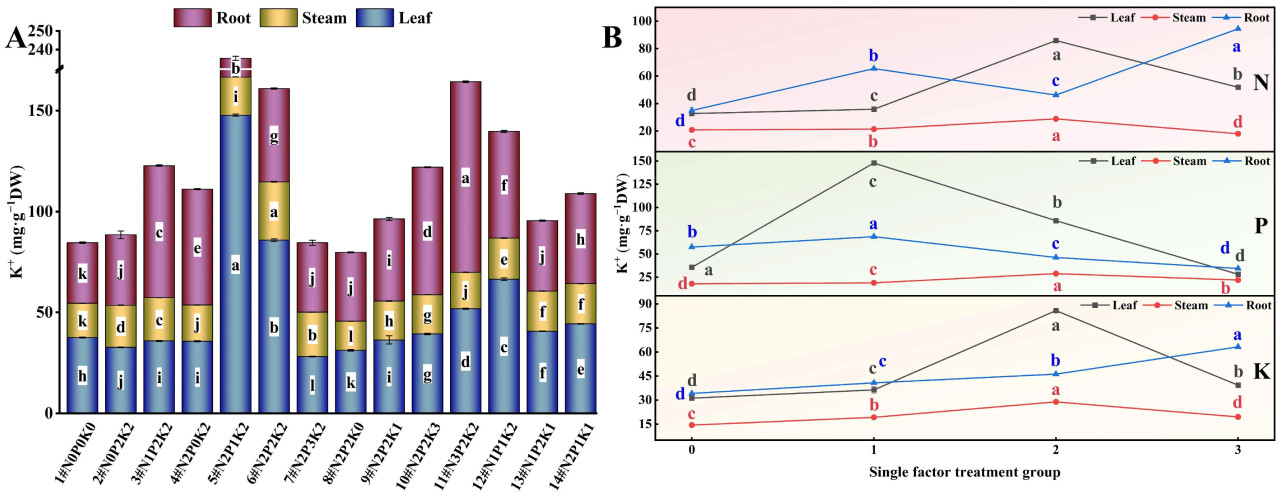
Рисунок 3. Влияние внесения N, P и K на содержание K⁺ в конопле в условиях стресса NaHCO₃. (A) Четырнадцать наборов соотношений NPK (#1–#14); (B) Единичный фактор удобрений N, P и K. Разные строчные буквы указывают на значительные различия между обработками для одного и того же органа растения при p < 0,05.
При рассмотрении только эффекта N (Рисунок 3B) содержание K⁺ в корнях конопли, подвергнутой щелочному стрессу, следовало порядку N3 > N1 > N2 > N0, содержание K⁺ в стеблях следовало N2 > N1 > N0 > N3, а содержание K⁺ в листьях следовало N2 > N3 > N1 > N0. С увеличением внесения N содержание K⁺ в корнях постепенно увеличивалось, достигая максимума на уровне N3, в то время как содержание K⁺ в стеблях и листьях было наибольшим на уровне N2. Эти результаты предполагают, что высокие уровни внесения N наиболее благоприятны для увеличения содержания K⁺ в корнях конопли, подвергнутой щелочному стрессу, в то время как умеренные уровни внесения N более эффективны для увеличения содержания K⁺ в стеблях и листьях.
В эксперименте с единичным фактором P (Рисунок 3B) содержание K⁺ в корнях конопли, подвергнутой щелочному стрессу, следовало порядку P1 > P0 > P2 > P3, содержание K⁺ в стеблях следовало P2 > P3 > P1 > P0, а содержание K⁺ в листьях следовало P1 > P2 > P0 > P3. С увеличением внесения P содержание K⁺ в корнях и листьях показало тенденцию сначала к увеличению, а затем к уменьшению, достигая самого низкого уровня при P3. Содержание K⁺ в стеблях при каждой обработке P было выше, чем в контроле без P, достигая максимума на уровне P2. Эти результаты указывают на то, что чрезмерное внесение фосфорного удобрения снизит уровень K⁺ в корнях и листьях конопли, подвергнутой щелочному стрессу, в то время как умеренное внесение P наиболее благоприятно для увеличения содержания K⁺ в стеблях.
Результаты эксперимента с единичным фактором K (Рисунок 3B) показали, что в условиях щелочного стресса содержание K⁺ в корнях конопли следовало порядку K3 > K2 > K1 > K0, в то время как содержание K⁺ в стеблях и листьях следовало K2 > K3 > K1 > K0. С увеличением внесения K содержание K⁺ в различных органах постепенно увеличивалось, при этом содержание K⁺ в корнях было наибольшим на уровне K3, а содержание K⁺ в стеблях и листьях достигало максимума на уровне K2. Эти результаты демонстрируют, что высокие уровни внесения K наиболее благоприятны для увеличения уровня K⁺ в различных органах конопли, подвергнутой щелочному стрессу.
В целом, в условиях щелочного солевого стресса, при внесении удобрений N, P и K в разных пропорциях, содержание K⁺ в различных органах конопли следовало порядку корень > стебель > лист.
3.2.3. N
При взаимодействии N, P и K (Рисунок 4A) наблюдались значительные различия в содержании N в корнях, стеблях и листьях конопли, подвергнутой щелочному стрессу, среди 14 обработок удобрением. Содержание N в корнях было наибольшим при обработке N3P2K2 (11#), что на 53,34% выше, чем в контрольной группе (1#, N0P0K0). Содержание N в стеблях и листьях было наибольшим при обработке N2P2K2 (6#), что на 53,47% и 103,89% выше, чем в контрольной группе, соответственно.
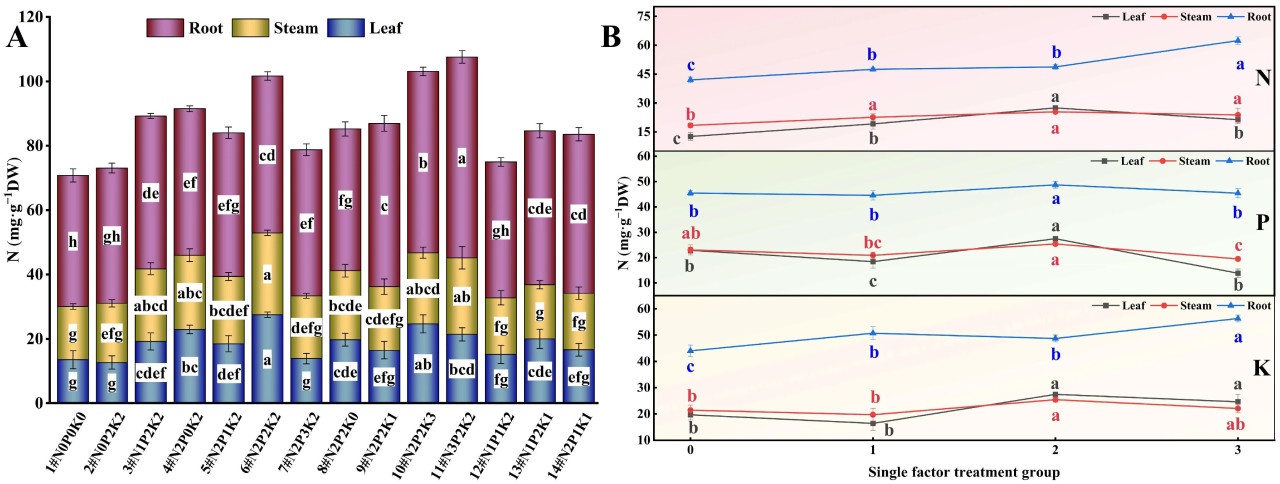
Рисунок 4. Влияние внесения N, P и K на содержание N в конопле в условиях стресса NaHCO₃. (A) Четырнадцать наборов соотношений NPK (#1–#14); (B) Единичный фактор удобрений N, P и K. Разные строчные буквы указывают на значительные различия между обработками для одного и того же органа растения при p < 0,05.
При рассмотрении только эффекта N (Рисунок 4B) содержание N в корнях конопли, подвергнутой щелочному стрессу, следовало порядку N3 > N2 > N1 > N0, в то время как содержание N в стеблях и листьях следовало N2 > N3 > N1 > N0. С увеличением внесения N содержание N в корнях постепенно увеличивалось, достигая максимума на уровне N3. Содержание N в стеблях и листьях показало тенденцию сначала к увеличению, а затем к уменьшению, достигая самого высокого уровня при N2. Эти результаты указывают на то, что внесение N может увеличить уровень N в различных органах конопли, подвергнутой щелочному стрессу, при этом высокие уровни N благоприятны для накопления N в корнях, а умеренные уровни N более эффективны для увеличения содержания азота в стеблях и листьях.
В эксперименте с единичным фактором P (Рисунок 4B) содержание N в корнях конопли, подвергнутой щелочному стрессу, следовало порядку P2 > P0 ≈ P3 > P1, в то время как содержание азота в стеблях и листьях следовало P2 > P0 > P1 > P3. С увеличением внесения P содержание азота в различных органах показало тенденцию сначала к уменьшению, а затем к увеличению, достигая самого высокого уровня при P2. Эти результаты предполагают, что умеренное внесение P наиболее благоприятно для увеличения уровня N в корнях, стеблях и листьях конопли, подвергнутой щелочному стрессу.
Результаты эксперимента с единичным фактором K (Рисунок 4B) показали, что в условиях щелочного стресса содержание N в корнях конопли следовало порядку K3 > K1 > K2 > K0, в то время как содержание N в стеблях и листьях следовало K2 > K3 > K0 > K1. Содержание N в корнях, стеблях и листьях при каждой обработке K было выше, чем в контроле без K, при этом содержание N в корнях было наибольшим на уровне K3, а содержание N в стеблях и листьях достигало максимума на уровне K2. Эти результаты демонстрируют, что высокие уровни K благоприятны для накопления N в корнях, в то время как умеренные уровни K более эффективны для увеличения уровня N в стеблях и листьях.
В целом, в условиях щелочного солевого стресса содержание N в различных органах конопли следовало порядку корень > стебель > лист.
3.2.4. P
При взаимодействии N, P и K (Рисунок 5A) наблюдались значительные различия в содержании P в корнях, стеблях и листьях конопли, подвергнутой щелочному стрессу, среди 14 обработок удобрением. Содержание P в корнях и листьях было наибольшим при обработке N2P3K2 (7#), что на 178,34% и 130,09% выше, чем в контрольной группе (1#, N0P0K0), соответственно. Содержание P в стеблях достигло максимума при обработке N3P2K2 (11#), что на 122,51% выше, чем в контрольной группе.
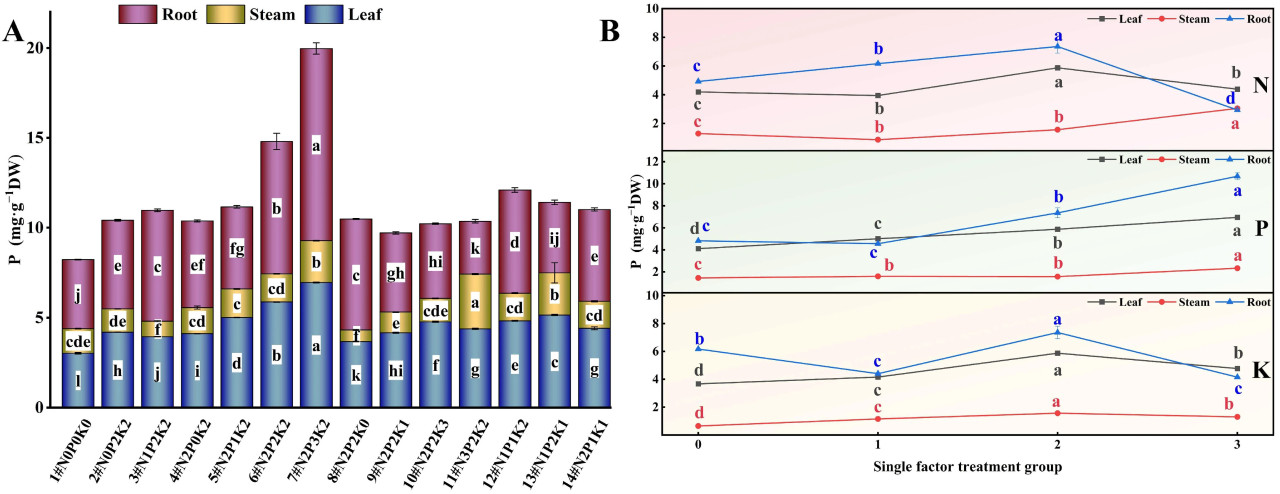
Рисунок 5. Влияние внесения N, P и K на содержание P в конопле в условиях стресса NaHCO₃. (A) Четырнадцать наборов соотношений NPK (#1–#14); (B) Единичный фактор удобрений N, P и K. Разные строчные буквы указывают на значительные различия между обработками для одного и того же органа растения при p < 0,05.
При рассмотрении только эффекта N (Рисунок 5B) содержание P в корнях конопли, подвергнутой щелочному стрессу, следовало порядку N2 > N1 > N0 > N3, содержание P в стеблях следовало N3 > N2 > N0 > N1, а содержание P в листьях следовало N2 > N3 > N0 > N1. С увеличением внесения N содержание P в корнях и листьях показало тенденцию сначала к увеличению, а затем к уменьшению, достигая самого низкого уровня при N3, в то время как содержание P в стеблях показало тенденцию сначала к уменьшению, а затем к увеличению, достигая самого низкого уровня при N1. Эти результаты предполагают, что низкие и умеренные уровни N благоприятны для увеличения уровня P в корнях конопли, подвергнутой щелочному стрессу, в то время как умеренные и высокие уровни N более эффективны для накопления P в стеблях и листьях.
В эксперименте с единичным фактором P (Рисунок 5B) содержание P в корнях конопли, подвергнутой щелочному стрессу, следовало порядку P3 > P2 > P0 > P1, содержание фосфора в стеблях следовало P3 > P1 > P2 > P0, а содержание P в листьях следовало P3 > P2 > P1 > P0. С увеличением внесения P содержание P в корнях показало тенденцию сначала к уменьшению, а затем к увеличению, содержание P в листьях постепенно увеличивалось, и уровень P в различных органах достиг своего максимума при обработке P3. Эти результаты указывают на то, что высокие условия P наиболее благоприятны для увеличения содержания P в корнях, стеблях и листьях конопли, подвергнутой щелочному стрессу.
Результаты эксперимента с единичным фактором K (Рисунок 5B) показали, что в условиях щелочного стресса содержание P в корнях конопли следовало порядку K2 > K0 > K1 > K3, эффект каждой обработки K на содержание P в стеблях не был значительным, и содержание фосфора в стеблях и листьях оба следовали порядку K2 > K3 > K1 > K0. С увеличением внесения K содержание фосфора в корнях сначала уменьшалось, а затем увеличивалось, достигая своего максимума на уровне K2. Хотя содержание P в стеблях и листьях оба были выше, чем в контроле без K, они также достигли своего максимума при обработке K2. Эти результаты демонстрируют, что умеренное внесение K наиболее благоприятно для увеличения уровня P в различных органах конопли, подвергнутой щелочному стрессу.
В целом, в условиях щелочного солевого стресса содержание P в различных органах конопли следовало порядку корень > лист > стебель.
3.2.5. Ca²⁺ и Mg²⁺
При интерактивных эффектах N, P и K (Рисунок 6A и Рисунок 7A) содержание Ca²⁺ и Mg²⁺ в корнях, стеблях и листьях конопли, подвергнутой щелочному стрессу, значительно варьировало среди 14 обработок удобрением. Обработка N2P3K2 (7#) привела к максимальному содержанию Ca²⁺ и Mg²⁺ в корнях, что было на 361,31% и 265,97% выше, соответственно, чем в контроле (1#, N0P0K0). Стебли имели наибольшее содержание Ca²⁺ при обработке N3P2K2 (11#), на 59,84% выше, чем в контроле, в то время как обработка N2P2K2 (6#) привела к наибольшему содержанию Mg²⁺ в стеблях, на 66,70% выше, чем в контроле. Листья показали максимальное содержание Ca²⁺ и Mg²⁺ при обработке N2P2K2 (6#), превысив контроль на 106,94% и 178,32% соответственно.
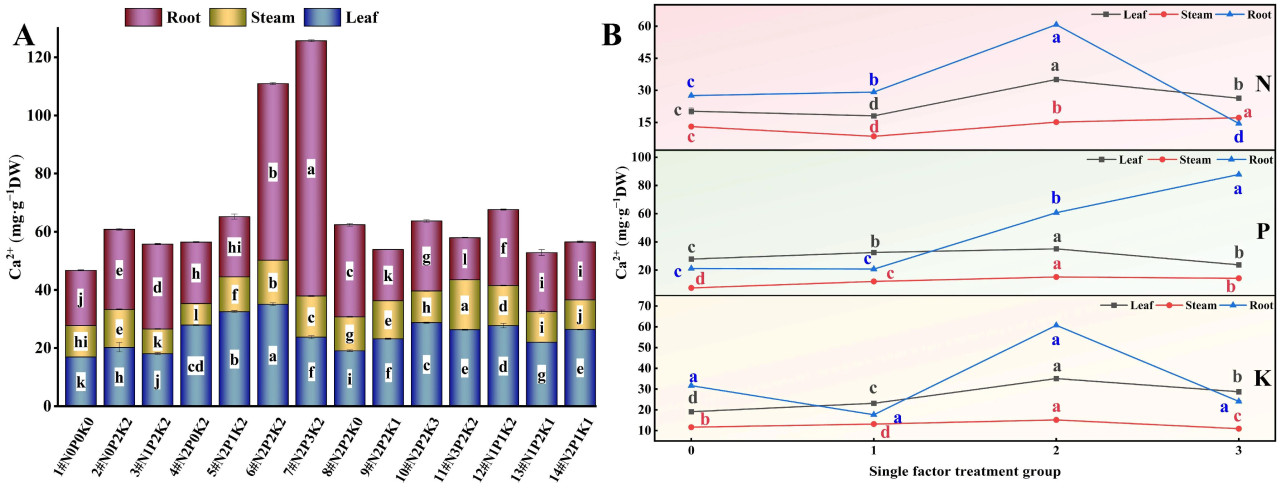
Рисунок 6. Влияние внесения N, P и K на содержание Ca²⁺ в конопле в условиях стресса NaHCO₃. (A) Четырнадцать наборов соотношений NPK (#1–#14); (B) Единичный фактор удобрений N, P и K. Разные строчные буквы указывают на значительные различия между обработками для одного и того же органа растения при p < 0,05.
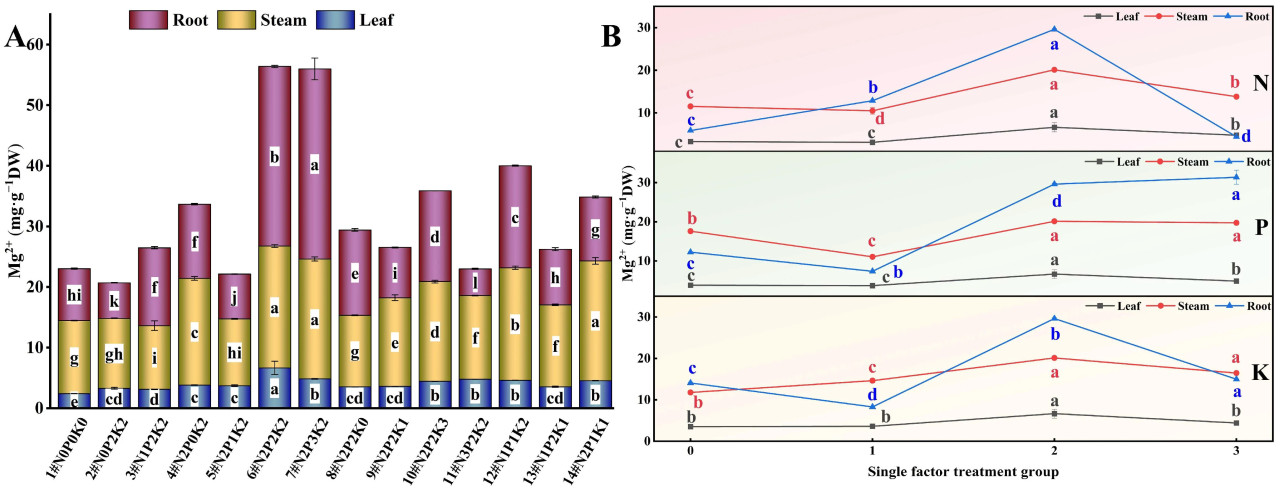
Рисунок 7. Влияние внесения N, P и K на содержание Mg²⁺ в конопле в условиях стресса NaHCO₃. (A) Четырнадцать наборов соотношений NPK (#1–#14); (B) Единичный фактор удобрений N, P и K. Разные строчные буквы указывают на значительные различия между обработками для одного и того же органа растения при p < 0,05.
Рассматривая эффект только N (Рисунок 6B и Рисунок 7B), содержание Ca²⁺ и Mg²⁺ в корнях конопли, подвергнутой щелочному стрессу, уменьшалось в порядке N2 > N1 > N0 > N3. Содержание Ca²⁺ в стеблях уменьшалось в порядке N3 > N2 > N0 > N1, в то время как содержание Ca²⁺ в листьях и содержание Mg²⁺ в стеблях и листьях следовало порядку N2 > N3 > N0 > N1. С увеличением внесения N содержание Ca²⁺ и Mg²⁺ в корнях сначала увеличивалось, а затем уменьшалось, с самыми низкими значениями на уровне N3. Напротив, содержание Ca²⁺ и Mg²⁺ в стеблях и листьях сначала уменьшалось, а затем увеличивалось, показывая минимальные значения на уровне N1. Обработка N2 привела к наибольшему содержанию Mg²⁺ во всех органах. Эти результаты предполагают, что низкие и умеренные уровни N благоприятствуют накоплению Ca²⁺ в корнях конопли, подвергнутой щелочному стрессу, в то время как средние и высокие уровни N благоприятствуют накоплению Ca²⁺ в стеблях и листьях. Умеренное внесение N наиболее эффективно для одновременного увеличения уровней Mg²⁺ в корнях, стеблях и листьях конопли, подвергнутой щелочному стрессу.
В эксперименте с единичным фактором P содержание Ca²⁺ и Mg²⁺ в корнях конопли, подвергнутой щелочному стрессу, уменьшалось в порядке P3 > P2 > P0 > P1. Содержание Ca²⁺ в стеблях следовало порядку P2 > P3 > P1 > P0, в то время как в листьях оно уменьшалось в порядке P2 > P1 > P0 > P3. С увеличением внесения P содержание Ca²⁺ и Mg²⁺ в корнях сначала уменьшалось, а затем увеличивалось, достигая пика на уровне P3. Однако содержание Ca²⁺ в листьях сначала увеличивалось, а затем уменьшалось, с самым низким значением при обработке P3. Эти результаты указывают на то, что средние и высокие уровни P увеличивают накопление Ca²⁺ в корнях конопли, подвергнутой щелочному стрессу, но чрезмерное P может снизить содержание Ca²⁺ в листьях. Содержание Mg²⁺ в стеблях и листьях уменьшалось в порядке P2 > P3 > P0 > P1. С увеличением P содержание Mg²⁺ в стеблях и листьях сначала увеличивалось, а затем уменьшалось, достигая максимума на уровне P2, что предполагает, что средние и высокие уровни P увеличивают содержание Mg²⁺ в различных органах конопли, подвергнутой щелочному стрессу.
Эксперимент с единичным фактором K показал, что в условиях щелочного стресса содержание Ca²⁺ в корнях конопли уменьшалось в порядке K2 > K0 > K3 > K1, в то время как в стеблях оно следовало порядку K2 > K1 > K3 > K0. Содержание Ca²⁺ и Mg²⁺ в листьях и содержание Mg²⁺ в корнях и стеблях уменьшалось в порядке K2 > K3 > K1 > K0. Обработка K2 привела к наибольшему содержанию Ca²⁺ и Mg²⁺ во всех органах, при этом содержание Ca²⁺ в стеблях и листьях превышало таковое в контроле без K. Уровни Mg²⁺ в корнях, стеблях и листьях превышали уровни контроля без K во всех обработках K. Эти результаты демонстрируют, что умеренное внесение K является наиболее эффективным способом увеличения уровней Ca²⁺ и Mg²⁺ в различных органах конопли, подвергнутой щелочному стрессу.
В целом, в условиях щелочного солевого стресса содержание Ca²⁺ в различных органах конопли уменьшалось в порядке корни > листья > стебли, в то время как содержание Mg²⁺ следовало порядку корни > стебли > листья.
3.2.6. Анализ размаха влияния удобрений N, P и K на содержание питательных веществ в конопле в условиях стресса NaHCO₃
Анализ крайних разностей (Таблица 5) способов удобрения выявил значительные эффекты на элементы питания в конопле в условиях щелочного солевого стресса. Элемент N оказал наиболее значительное влияние на N, P и Ca²⁺ в стеблях; N, K и Na⁺ в корнях; и N и Mg²⁺ в листьях. Элемент P в основном влиял на P, Ca²⁺ и Mg²⁺ в корнях; Na⁺, P и K⁺ в листьях; и Na⁺ и Mg²⁺ в стеблях. Элемент K в основном влиял на K⁺ в стеблях и Ca²⁺ в листьях.
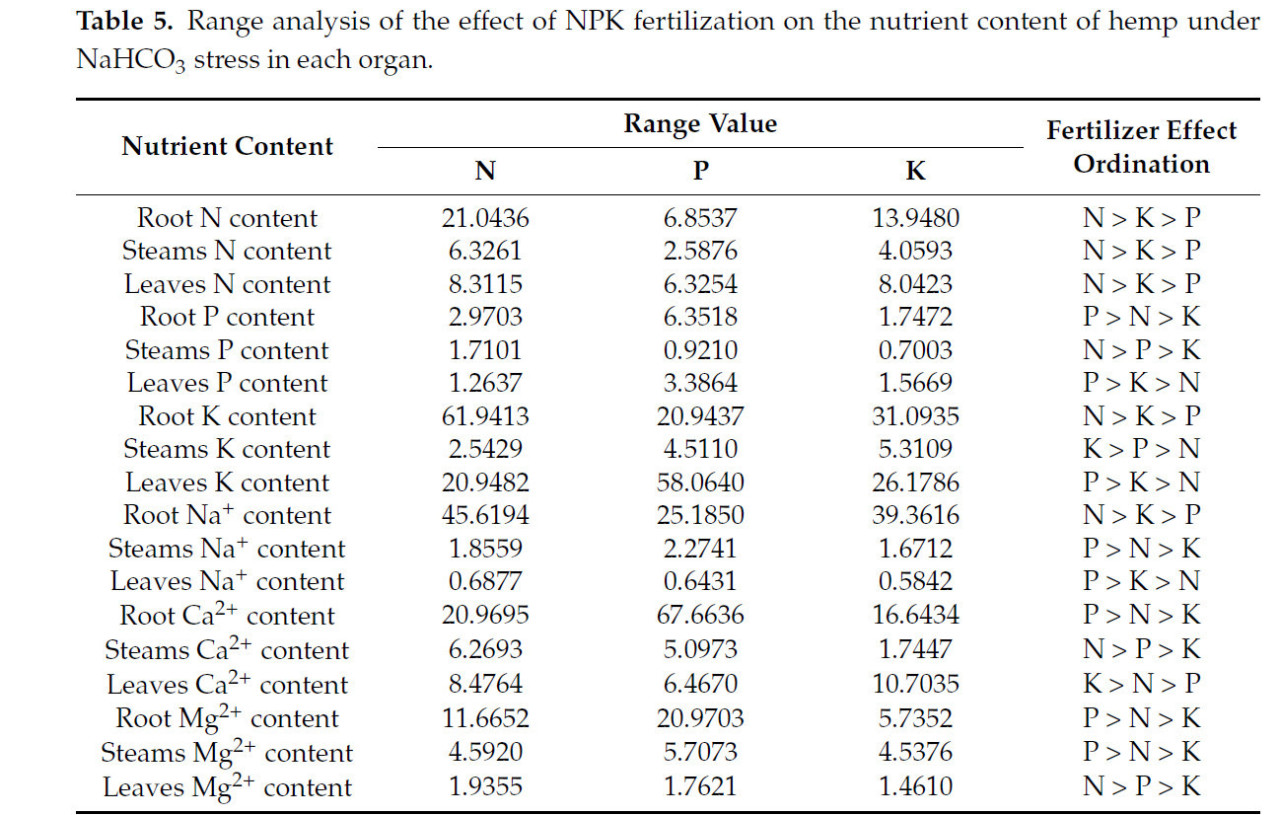
Таблица 5. Анализ размаха влияния удобрения NPK на содержание питательных веществ в конопле в условиях стресса NaHCO₃ в каждом органе.
4. Обсуждение
4.1. Оптимальные соотношения N, P и K для роста конопли в условиях щелочного стресса
Это исследование продемонстрировало, что в условиях специфического щелочного стресса (бикарбонат натрия, NaHCO₃, 200 мМ) оптимальное соотношение NPK для роста конопли составляет N1P2K2, что соответствует азоту (N) 120 мг·л⁻¹, фосфору (P) 238 мг·л⁻¹ и калию (K) 348 мг·л⁻¹. При этих условиях биомасса стеблей и корней конопли показала увеличение на 45% и 39% соответственно по сравнению с контрольной группой. Одновременно соотношение K⁺/Na⁺ в листьях показало благоприятный уровень > 1,2. Эти результаты предполагают, что сбалансированное снабжение NPK является ключевым для повышения солеустойчивости и продуктивности конопли. В соответствии с этими наблюдениями аналогичные результаты исследований были задокументированы у других видов растений, включая райграс многоукосный (Lolium multiflorum), сорго двуцветное (Sorghum bicolor) и Sulla carnosa [10,32,33].
N является значительным компонентом хлорофилла, белков и ферментов, играя жизненно важную роль в росте растений и адаптации к стрессу. Это исследование показало, что умеренный уровень снабжения N (N1, 120 мг·л⁻¹) значительно способствовал увеличению высоты растения, диаметра стебля, длины корня и накоплению биомассы в различных органах конопли в условиях щелочного стресса. Однако высокое снабжение N (N3, 360 мг·л⁻¹) проявляло некоторые ингибирующие эффекты. Этот результат согласуется с результатами связанных исследований пшеницы и томата (Solanum lycopersicum), где соответствующее снабжение N может улучшить солеустойчивость растений путем стимулирования роста и накопления биомассы [34,35]. Усиливающий эффект N на солеустойчивость и рост растений может быть тесно связан с его способствованием фотосинтетической способности, осмотической регуляции и антиоксидантным защитным механизмам [9,19].
P также является важным элементом, который влияет на рост конопли в условиях щелочного стресса. В этом исследовании снабжение P оказало значительное влияние на рост конопли, при этом низкие и умеренные уровни снабжения P (P1-P2, 119–238 мг·л⁻¹) оказывали стимулирующее действие на развитие растений. Среди обработок уровень P2 (238 мг·л⁻¹) привел к наибольшим значениям высоты растения, диаметра стебля, длины корня и сухого веса корня у конопли. Этот результат согласуется с выводами в исследованиях нута, где условия низкого P приводили к повышению солеустойчивости через стимулированное развитие корней и улучшенное усвоение питательных веществ [21]. Наблюдаемое стимулирование роста корней P может быть связано с его ролью в усилении деления клеток, удлинении клеток и формировании боковых корней [36,37].
K, наиболее распространенный катион в клетках растений, играет ключевую роль в активации ферментов. В контексте щелочного стресса адекватное снабжение K (K2, 348 мг·л⁻¹), как было показано, способствует значительному увеличению высоты растения, диаметра стебля, длины корня, сухого веса органов и количества листьев у конопли. Это K-индуцированное явление усиления роста также было задокументировано в исследованиях солевого стресса на арахисе, пшенице и других культурах [38,39,40].
4.2. Влияние комбинаций питания N, P и K на распределение питательных веществ и ионный гомеостаз
Исследования показали, что разнообразные комбинации N, P и K оказывают существенное влияние на модели поглощения и распределения питательных веществ в конопле в условиях щелочного стресса. Например, обработки N2P2K3 и N2P2K2 эффективно снижали содержание Na⁺ в корнях и увеличивали содержание K⁺ в стеблях соответственно, значительно улучшая соотношение K⁺/Na⁺. Эти результаты подразумевают, что путем оптимизации питания NPK можно регулировать поглощение и транспорт Na⁺, а также повышать способность усвоения и распределения K⁺, тем самым улучшая ионный гомеостаз в условиях солевого стресса [41]. Дополнительно, было показано, что сбалансированное удобрение усиливает поглощение других минеральных питательных веществ (например, N, P, Ca²⁺ и Mg²⁺), которые являются ключевыми для поддержания общего нутритивного статуса и солеустойчивости конопли [42,43].
С точки зрения специфических питательных веществ, снабжение N оказывает значительное влияние на модели накопления Na⁺ и K⁺ в различных органах конопли. Исследование продемонстрировало, что на уровне N2 содержание Na⁺ в корнях может быть эффективно снижено, в то время как содержание K⁺ в стеблях было увеличено, значительно улучшая соотношение K⁺/Na⁺. Этот результат предполагает, что эффективные методы управления N могут улучшить ионный баланс в условиях солевого стресса путем ограничения процессов поглощения и транспорта Na⁺ и путем стимулирования усвоения и распределения K⁺. Потенциальные механизмы, лежащие в основе этого явления, включают регуляцию активности транспортеров Na⁺/K⁺ с помощью N, стимулирование синтеза совместимых растворенных веществ и усиление прочности клеточной стенки [44,45,46,47].
P играет ключевую роль в различных аспектах биологии растений, включая энергетический метаболизм, передачу сигналов и производство биомассы. Было продемонстрировано, что P оказывает существенное влияние на поглощение и распределение минеральных питательных веществ в конопле в условиях щелочного стресса. Исследование продемонстрировало, что обработка P2 увеличила содержание N и P в корнях, одновременно уменьшая накопление Na⁺ и поддерживая более высокое соотношение K⁺/Na⁺ в стеблях и листьях. Было задокументировано, что адекватное снабжение P может усилить селективное поглощение K⁺ и Na⁺ корнями и ограничить приток Na⁺ путем модуляции активности плазматической мембраны H⁺-АТФазы и свойств клеточной стенки [14]. Более того, применение P, как было показано, повышает уровни Ca²⁺ и Mg²⁺ в органах конопли, тем самым способствуя стабилизации структуры клеточной мембраны и смягчению окислительного повреждения, вызванного солевым стрессом [48].
K играет ключевую роль в поддержании тургора клеток, усилении фотосинтеза и активации генов ответа на стресс [49]. Исследование показало, что по мере увеличения снабжения K содержание Na⁺ в корнях, стеблях и листьях постепенно уменьшалось, в то время как содержание K⁺ и соотношение K⁺/Na⁺ значительно увеличивались. Среди различных оцененных обработок обработка K3 (522 мг·л⁻¹) оказалась наиболее эффективной в снижении накопления Na⁺ в корнях и значительном усилении селективного поглощения K⁺/Na⁺ корнями. Этот результат указывает на то, что достаточное питание K может препятствовать поглощению Na⁺, способствовать исключению Na⁺ и усиливать поглощение K⁺ путем регулирования множественных ионных транспортеров и каналов [50]. Более того, применение K, как было показано, увеличивает содержание N, P, Ca²⁺ и Mg²⁺ в органах конопли, тем самым подчеркивая ключевую роль K в улучшении общего нутритивного статуса и повышении солеустойчивости конопли [15,16].
4.3. Вредные эффекты и пороговые концентрации чрезмерного удобрения
Это исследование определило критические пороги для внесения NPK в коноплю в условиях щелочного стресса. Чрезмерно высокие концентрации удобрений (общие питательные вещества > 400 мг·л⁻¹), как было показано, усугубляют солевое повреждение, снижая накопление биомассы на 25–30% и нарушая ионный гомеостаз. Этот вредный эффект может быть связан с повышенной засоленностью почвы, нутритивным дисбалансом и окислительным стрессом [51]. Более того, повышенные уровни азота (N3, 360 мг·л⁻¹), как было отмечено, препятствуют высоте растения, диаметру стебля, длине корня и биомассе органов у конопли. Этот эффект, возможно, обусловлен токсичностью аммония, нарушением внутриклеточного pH и накоплением активных форм кислорода (АФК) в щелочных условиях [52]. Аналогичное явление наблюдалось при избытке фосфора (P3, 357 мг·л⁻¹), который, как было обнаружено, снижает рост корней и содержание кальция в листьях. Этот эффект может быть связан с вызванным фосфором дефицитом цинка, изменениями структуры корней и нарушением кальциевой сигнализации [53]. Более того, чрезмерное внесение K (K3, 522 мг·л⁻¹), как было показано, ингибирует рост, возможно, из-за ионного антагонизма, осмотического стресса и нарушения клеточных метаболических процессов [54]. Следовательно, при возделывании конопли в засоленно-щелочной почве необходимо тщательно регулировать внесение удобрений NPK, чтобы избежать превышения порога толерантности культуры и обеспечить эффективное использование питательных веществ и устойчивое производство.
В целом, это исследование предоставляет систематическое понимание влияния управления удобрениями N, P и K на рост и минеральное питание конопли в условиях щелочного солевого стресса. Однако исследование не лишено ограничений. Например, оптимальное соотношение NPK может варьироваться в зависимости от генотипа конопли, стадии роста и специфических условий засоленно-щелочного стресса [11,55]. Следовательно, необходимо дальнейшее изучение для определения оптимального соотношения NPK для возделывания конопли в различных засоленно-щелочных почвах и условиях окружающей среды. Будущие исследовательские усилия должны уделять приоритетное внимание всестороннему изучению комбинированных эффектов удобрений NPK и вспомогательных агрономических мер, таких как орошение, мульчирование и улучшение почвы, на различные генотипы конопли на протяжении всего их цикла роста в условиях подлинной засоленно-щелочной почвы. Разработка более точных, эффективных и устойчивых интегрированных методов управления для конопли на засоленно-щелочных землях, как ожидается, будет способствовать восстановлению и использованию засоленно-щелочных земель и здоровому развитию индустрии конопли.
5. Выводы
Это исследование предоставляет новые идеи о влиянии удобрения NPK на рост, ионный гомеостаз и минеральное питание конопли в условиях щелочного солевого стресса. Результаты подчеркивают ключевую роль сбалансированного питания NPK, в частности обработки N1P2K2, в повышении солеустойчивости и производства биомассы конопли. Потенциальные физиологические механизмы включают скоординированную регуляцию поглощения, транспорта и компартментализации ионов, а также улучшение общего нутритивного статуса и антиоксидантной защиты. Эти результаты могут помочь в разработке целевых стратегий удобрения для выращивания конопли в засоленно-щелочных районах. Сочетание сбалансированного удобрения с другими агрономическими практиками может быть хорошим способом повышения устойчивости и рентабельности производства конопли в агроэкосистемах, подверженных засолению.
Дополнительные материалы
Следующая вспомогательная информация может быть загружена по адресу: https://www.mdpi.com/article/10.3390/agriculture15020125/s1.
Ссылки
1. Jones, A.D.; Ejeta, G. A new global agenda for nutrition and health: The importance of agriculture and food systems. Bull. World Health Organ. 2016, 94, 228–229. [Google Scholar] [CrossRef] [PubMed]
2. Zhou, B.; Liang, C.; Chen, X.; Ye, S.; Peng, Y.; Yang, L.; Duan, M.; Wang, X. Magnetically-treated brackish water affects soil water-salt distribution and the growth of cotton with film mulch drip irrigation in Xinjiang, China. Agric. Water Manag. 2022, 263, 107487. [Google Scholar] [CrossRef]
3. Hu, H.; Liu, H.; Liu, F. Seed germination of hemp (Cannabis sativa L.) cultivars responds differently to the stress of salt type and concentration. Ind. Crop. Prod. 2018, 123, 254–261. [Google Scholar] [CrossRef]
4. Manaia, J.P.; Manaia, A.T.; Rodriges, L. Industrial hemp fibers: An overview. Fibers 2019, 7, 106. [Google Scholar] [CrossRef]
5. Amaducci, S.; Scordia, D.; Liu, F.; Zhang, Q.; Guo, H.; Testa, G.; Cosentino, S. Key cultivation techniques for hemp in Europe and China. Ind. Crop. Prod. 2015, 68, 2. [Google Scholar] [CrossRef]
6. Liu, J.; Qiao, Q.; Cheng, X.; Du, G.; Deng, G.; Zhao, M.; Liu, F. Transcriptome differences between fiber-type and seed-type Cannabis sativa variety exposed to salinity. Physiol. Mol. Biol. Plants 2016, 22, 429–443. [Google Scholar] [CrossRef] [PubMed]
7. Jia, X.; Zhu, Y.; Zhang, R.; Zhu, Z.; Zhao, T.; Cheng, L.; Gao, L.; Liu, B.; Zhang, X.; Wang, Y. Ionomic and metabolomic analyses reveal the resistance response mechanism to saline-alkali stress in Malus halliana seedlings. Plant Physiol. Biochem. 2020, 147, 77–90. [Google Scholar] [CrossRef] [PubMed]
8. Yang, C.; Shi, D.; Wang, D. Comparative effects of salt and alkali stresses on growth, osmotic adjustment and ionic balance of an alkali-resistant halophyte Suaeda glauca (Bge.). Plant Growth Regul. 2008, 56, 179–190. [Google Scholar] [CrossRef]
9. Michalska, A.; Wojdyło, A.; Bogucka, B. The influence of nitrogen and potassium fertilisation on the content of polyphenolic compounds and antioxidant capacity of coloured potato. J. Food Compos. Anal. 2016, 47, 69–75. [Google Scholar] [CrossRef]
10. Guo, X.; Wu, Q.; Zhang, Z.; Zhu, G.; Zhou, G. Nitrogen and phosphorus counteracted the adverse effects of salt on sorghum by improving ROS scavenging and osmotic regulation. Agronomy 2023, 13, 1020. [Google Scholar] [CrossRef]
11. Singh, M.; Singh, V.P.; Prasad, S.M. Responses of photosynthesis, nitrogen and proline metabolism to salinity stress in Solanum lycopersicum under different levels of nitrogen supplementation. Plant Physiol. Biochem. 2016, 109, 72–83. [Google Scholar] [CrossRef]
12. Bouras, H.; Choukr-Allah, R.; Amouaouch, Y.; Bouaziz, A.; Devkota, K.P.; El Mouttaqi, A.; Bouazzama, B.; Hirich, A. How does quinoa (Chenopodium quinoa Willd.) respond to phosphorus fertilization and irrigation water salinity? Plants 2022, 11, 216. [Google Scholar] [CrossRef]
13. Iqbal, N.; Umar, S.; Khan, N.A. Nitrogen availability regulates proline and ethylene production and alleviates salinity stress in mustard (Brassica juncea). J. Plant Physiol. 2015, 178, 84–91. [Google Scholar] [CrossRef] [PubMed]
14. Talbi Zribi, O.; Abdelly, C.; Debez, A. Interactive effects of salinity and phosphorus availability on growth, water relations, nutritional status and photosynthetic activity of barley (Hordeum vulgare L.). Plant Biol. 2011, 13, 872–880. [Google Scholar] [CrossRef]
15. Huang, W.; Lin, M.; Liao, J.; Li, A.; Tsewang, W.; Chen, X.; Sun, B.; Liu, S.; Zheng, P. Effects of potassium deficiency on the growth of tea (Camelia sinensis) and strategies for optimizing potassium levels in soil: A critical review. Horticulturae 2022, 8, 660. [Google Scholar] [CrossRef]
16. Munir, A.; Shehzad, M.T.; Qadir, A.A.; Murtaza, G.; Khalid, H.I. Use of potassium fertilization to ameliorate the adverse effects of saline-sodic stress condition (ECw: SARw Levels) in rice (Oryza sativa L.). Commun. Soil Sci. Plant Anal. 2019, 50, 1975–1985. [Google Scholar] [CrossRef]
17. Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The critical role of potassium in plant stress response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef]
18. Zhao, D.-Y.; Zhang, X.-L.; Zhao, S.-P.; Liu, G.-L.; Zhang, Z.-W.; Zhao, W.-F.; Li, X.-P.; Khan, S.A.; Siddique, K.H.M. Biomass allocation and nutrients utilization in wheat as affected by phosphorus placement and salt stress. Agronomy 2023, 13, 1570. [Google Scholar] [CrossRef]
19. Reinbott, T.M.; Blevins, D.G. Phosphorus and temperature effects on magnesium, calcium, and potassium in wheat and tall fescueleaves. Agron. J. 1994, 86, 523–529. [Google Scholar] [CrossRef]
20. Liu, K.; Zhang, T.Q.; Tan, C.S.; Astatkie, T. Responses of fruit yield and quality of processing tomato todrip rrigation and ferilizers phosphorus and potassium. Agron. J. 2011, 103, 1339–1345. [Google Scholar] [CrossRef]
21. Kaci, H.S.-A.; Haddadj, A.C.; Aid, F. Enhancing of symbiotic efficiency and salinity tolerance of chickpea by phosphorus supply. Acta Agric. Scand. Sect. B-Soil Plant Sci. 2018, 68, 534–540. [Google Scholar] [CrossRef]
22. Chen, Z.; Ma, H.; Xia, J.; Hou, F.; Shi, X.; Hao, X.; Hafeez, A.; Han, H.; Luo, H. Optimal pre-plant irrigation and fertilization canimprove biomass accumulation by maintaining the root and leaf productive capacity of cotton crop. Sci. Rep. 2017, 7, 17168. [Google Scholar] [CrossRef] [PubMed]
23. Chen, B.; Wang, Q.; Ye, Z.; Stiles, S.; Feng, G. Optimisation of phosphorus fertilisation promotes biomass and phosphorus nutrientaccumulation, partitioning and translocation in three cotton (Gossypium hirsutum) genotypes. Crop Pasture Sci. 2020, 71, 56–69. [Google Scholar] [CrossRef]
24. GB 9834-1988; Standardization Administration of China. Method for Determination of Soil Organic Matter. China Standard Press: Beijing, China, 1988.
25. Cao, K.; Sun, Y.; Zhang, X.; Zhao, Y.; Bian, J.; Zhu, H.; Wang, P.; Gao, B.; Sun, X.; Hu, M. The miRNA–mRNA regulatory networks of the response to NaHCO3 stress in industrial hemp (Cannabis sativa L.). BMC Plant Biol. 2023, 23, 509. [Google Scholar] [CrossRef] [PubMed]
26. Yuan, H.; Si, H.; Ye, Y.; Ji, Q.; Wang, H.; Zhang, Y. Arbuscular Mycorrhizal Fungi-Mediated Modulation of Physiological, Biochemical, and Secondary Metabolite Responses in Hemp (Cannabis sativa L.) under Salt and Drought Stress. J. Fungi 2024, 10, 283. [Google Scholar] [CrossRef] [PubMed]
27. Wei, T.J.; Li, G.; Wang, M.M.; Jin, Y.Y.; Zhang, G.H.; Liu, M.; Yang, H.Y.; Jiang, C.J.; Liang, Z.-W. Physiological and transcriptomic analyses reveal novel insights into the cultivar-specific response to alkaline stress in alfalfa (Medicago sativa L.). Ecotoxicol. Environ. Saf. 2021, 228, 113017. [Google Scholar] [CrossRef]
28. Wei, X.; Zhou, W.; Long, S.; Guo, Y.; Qiu, C.; Zhao, X.; Wang, Y. Effects of Different N, P, and K Rates on the Growth and Cannabinoid Content of Industrial Hemp. J. Nat. Fibers 2023, 20, 2159605. [Google Scholar] [CrossRef]
29. Liu, J.; Chen, W.; Wang, H.; Peng, F.; Chen, M.; Liu, S.; Chu, G. Effects of NPK fertilization on photosynthetic characteristics and nutrients of pecan at the seedling stage. J. Soil Sci. Plant Nutr. 2021, 21, 2425–2435. [Google Scholar] [CrossRef]
30. Chen, K.; Ma, L.; Chen, C.; Liu, N.; Wang, B.; Bao, Y.; Liu, Z.; Zhou, G. Long-term impact of N, P, K fertilizers in different rates on yield and quality of Anisodus tanguticus (Maxinowicz) Pascher. Plants 2023, 12, 2102. [Google Scholar] [CrossRef]
31. Yang, Z.-J.; Wu, X.-H.; Chen, L.-H.; Huang, L.-M.; Chen, Y.; Wu, J.; El-Kassaby, Y.A.; Grossnickle, S.C.; Feng, J.-L. Fertilization regulates accumulation and allocation of biomass and nutrients in Phoebe bournei seedlings. Agriculture 2021, 11, 1187. [Google Scholar] [CrossRef]
32. Shao, A.; Sun, Z.; Fan, S.; Xu, X.; Wang, W.; Amombo, E.; Yin, Y.; Li, X.; Wang, G.; Wang, H. Moderately low nitrogen application mitigate the negative effects of salt stress on annual ryegrass seedlings. PeerJ 2020, 8, e10427. [Google Scholar] [CrossRef] [PubMed]
33. Hafsi, C.; Falleh, H.; Saada, M.; Ksouri, R.; Abdelly, C. Potassium deficiency alters growth, photosynthetic performance, secondary metabolites content, and related antioxidant capacity in Sulla carnosa grown under moderate salinity. Plant Physiol.Biochem. 2017, 118, 609–617. [Google Scholar] [CrossRef] [PubMed]
34. Ahanger, M.A.; Qin, C.; Begum, N.; Maodong, Q.; Dong, X.X.; El-Esawi, M.; El-Sheikh, M.A.; Alatar, A.A.; Zhang, L. Nitrogen availability prevents oxidative effects of salinity on wheat growth and photosynthesis by up-regulating the antioxidants and osmolytes metabolism, and secondary metabolite accumulation. BMC Plant Biol. 2019, 19, 479. [Google Scholar] [CrossRef]
35. Siddiqui, M.H.; Mohammad, F.; Khan, M.N.; Al-Whaibi, M.H.; Bahkali, A.H. Nitrogen in Relation to Photosynthetic Capa city and Accumulation of Osmo protectant and Nutrients in Brassica Genotypes Grown Under Salt Stress. Agric. Sci. China 2010, 9, 671–680. [Google Scholar] [CrossRef]
36. Xia, Z.; Zhang, S.; Wang, Q.; Zhang, G.; Fu, Y.; Fu, Y.; Lu, H.; Lu, H. Effects of root zone warming on maize seedling growth and photosynthetic characteristics under different phosphorus levels. Front. Plant Sci. 2021, 12, 746152. [Google Scholar] [CrossRef] [PubMed]
37. Zhang, D.; Zhang, C.; Tang, X.; Li, H.; Zhang, F.; Rengel, Z.; Whalley, W.R.; Davies, W.J.; Shen, J. Increased soil phosphorus availability induced by faba bean root exudation stimulates root growth and phosphorus uptake in neighbouring maize. New Phytol. 2016, 209, 823–831. [Google Scholar] [CrossRef]
38. Shi, X.; Zhou, D.; Guo, P.; Zhang, H.; Dong, J.; Ren, J.; Jiang, C.; Zhong, C.; Zhao, X.; Yu, H. External potassium mediates the response and tolerance to salt stress in peanut at the flowering and needling stages. Photosynthetica 2020, 58, 1141–1149. [Google Scholar] [CrossRef]
39. Gul, M.; Wakeel, A.; Steffens, D.; Lindberg, S. Potassium-induced decrease in cytosolic Na+ alleviates deleterious effects of salt stress on wheat (Triticum aestivum L.). Plant Biol. 2019, 21, 825–831. [Google Scholar] [CrossRef]
40. Rady, M.M.; Mossa, A.-T.H.; Youssof, A.M.; Osman, A.S.; Ahmed, S.M.; Mohamed, I.A. Exploring the reinforcing effect of nano-potassium on the antioxidant defense system reflecting the increased yield and quality of salt-stressed squash plants. Sci. Hortic. 2023, 308, 111609. [Google Scholar] [CrossRef]
41. Chen, W.; Hou, Z.; Wu, L.; Liang, Y.; Wei, C. Effects of salinity and nitrogen on cotton growth in arid environment. Plant Soil 2010, 326, 61–73. [Google Scholar] [CrossRef]
42. Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef] [PubMed]
43. Liu, L.; Miao, Q.; Wang, H.; Xue, Y.; Qi, S.; Zhang, J.; Li, J.; Meng, Q.; Cui, Z. Optimizing phosphorus application for winter wheat production in the coastal saline area. Agronomy 2022, 12, 2966. [Google Scholar] [CrossRef]
44. Chen, Z.; Pottosin, I.I.; Cuin, T.A.; Fuglsang, A.T.; Tester, M.; Jha, D.; Zepeda-Jazo, I.; Zhou, M.; Palmgren, M.G.; Newman, I.A. Root plasma membrane transporters controlling K+/Na+ homeostasis in salt-stressed barley. Plant Physiol. 2007, 145, 1714–1725. [Google Scholar] [CrossRef] [PubMed]
45. Tester, M.; Davenport, R. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef] [PubMed]
46. Yang, Z.; Tan, S.; Yang, Q.; Chen, S.; Qi, C.; Liu, X.; Liang, J.; Wang, H. Nitrogen Application Alleviates Impairments for Jatropha curcas L. Seedling Growth under Salinity Stress by Regulating Photosynthesis and Antioxidant Enzyme Activity. Agronomy 2023, 13, 1749. [Google Scholar] [CrossRef]
47. Giambalvo, D.; Amato, G.; Borgia, D.; Ingraffia, R.; Librici, C.; Lo Porto, A.; Puccio, G.; Ruisi, P.; Frenda, A.S. Nitrogen availability drives mycorrhizal effects on wheat growth, nitrogen uptake and recovery under salt stress. Agronomy 2022, 12, 2823. [Google Scholar] [CrossRef]
48. Li, L.; Tang, C.; Rengel, Z.; Zhang, F.S. Calcium, magnesium and microelement uptake as affected by phosphorus sources andinterspecific root interactions between wheat and chickpea. Plant Soil 2004, 261, 29–37. [Google Scholar] [CrossRef]
49. Zhang, L.; Jiang, Q.; Zong, J.; Guo, H.; Liu, J.; Chen, J. Effects of Supplemental Potassium on the Growth, Photosynthetic Characteristics, and Ion Content of Zoysia matrella under Salt Stress. Horticulturae 2023, 10, 31. [Google Scholar] [CrossRef]
50. Gao, Y.; Li, D. Assessing leaf senescence in tall fescue (Festuca arundinacea Schreb.) under salinity stress using leaf spectrum. Eur. J. Hortic. Sci 2015, 80, 170–176. [Google Scholar] [CrossRef]
51. Mauchamp, A.; Mésleard, F. Salt tolerance in Phragmites australis populations from coastal Mediterranean marshes. Aquat. Bot. 2001, 70, 39–52. [Google Scholar] [CrossRef]
52. Elhanafi, L.; Houhou, M.; Rais, C.; Mansouri, I.; Elghadraoui, L.; Greche, H. Impact of excessive nitrogen fertilization on the biochemical quality, phenolic compounds, and antioxidant power of Sesamum indicum L. seeds. J. Food Qual. 2019, 2019, 9428092. [Google Scholar] [CrossRef]
53. Tang, H.; Niu, L.; Wei, J.; Chen, X.; Chen, Y. Phosphorus limitation improved salt tolerance in maize through tissue mass density increase, osmolytes accumulation, and Na+ uptake inhibition. Front. Plant Sci. 2019, 10, 856. [Google Scholar] [CrossRef] [PubMed]
54. Daoud, B.; Pawelzik, E.; Naumann, M. Different potassium fertilization levels influence water-use efficiency, yield, and fruit quality attributes of cocktail tomato—A comparative study of deficient-to-excessive supply. Sci. Hortic. 2020, 272, 109562. [Google Scholar] [CrossRef]
55. Duan, M.; Chang, S.X. Nitrogen fertilization improves the growth of lodgepole pine and white spruce seedlings under low salt stress through enhancing photosynthesis and plant nutrition. For. Ecol. Manag. 2017, 404, 197–204. [Google Scholar] [CrossRef]

Ye Y, Wang H, Zhang P, Zhang Y. Adaptation Strategies for Hemp in Alkaline Salt Environments: Fertilizer Management for Nutrient Uptake and Optimizing Growth. Agriculture. 2025; 15(2):125. https://doi.org/10.3390/agriculture15020125
Перевод статьи «Adaptation Strategies for Hemp in Alkaline Salt Environments: Fertilizer Management for Nutrient Uptake and Optimizing Growth» авторов Ye Y, Wang H, Zhang P, Zhang Y., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык

















































































































Комментарии (0)