Влияние опрыскивания селеном на рост и устойчивость к полеганию проростков ароматного риса
Предыдущие исследования показали, что селен (Se) может влиять на рост и урожайность риса. Однако влияние селена на полегание риса остается неизученным. Целью данного исследования было изучение воздействия различных обработок селеном на рост проростков и твердость оболочки стебля у ароматного риса.
Аннотация
Был проведен гидропонный эксперимент с использованием двух сортов ароматного риса — Юйсяньючжань и Сянъясянчжань — в качестве объектов исследования. Применялись две формы селеновых удобрений (хелатированный аминокислотами селен и селенит натрия). Было проведено пять вариантов внекорневой обработки селеновыми удобрениями (CK: без селеновых удобрений; T1: 4 мкмоль·л⁻¹ хелатированного аминокислотами селена; T2: 8 мкмоль·л⁻¹ хелатированного аминокислотами селена; T3: 4 мкмоль·л⁻¹ селенита натрия; и T4: 8 мкмоль·л⁻¹ селенита натрия), и изучалось влияние различных обработок селеновыми удобрениями на рост проростков и твердость оболочки стебля у ароматного риса.
Было обнаружено значительное влияние обработок селеном на сырую массу корней, сухую массу проростков, высоту растений, длину оболочки стебля, количество листьев, содержание хлорофилла, твердость оболочки стебля, активность пероксидазы в листьях и оболочках стеблей, а также содержание лигнина в корнях. Наблюдалось значительное взаимодействие между обработкой селеном и сортом в отношении твердости оболочки стебля. Различные формы и уровни селеновых удобрений по-разному влияли на рост проростков и твердость оболочки стебля. Обработки селеном улучшили рост проростков и оказали значительное влияние на сухую массу, содержание хлорофилла, твердость оболочки стебля и активность пероксидазы в листьях и оболочках стеблей.
По сравнению с контролем (CK), обработки селеном увеличили общую сухую массу проростков у сортов Сянъясянчжань и Юйсяньючжань в диапазонах 25,43–52,77% и 18,97–30,09% соответственно. Обработки T2–T4 повысили показатели твердости оболочки стебля у Сянъясянчжань и Юйсяньючжань на 21,6–54,7% и 38,3–146,6% соответственно по сравнению с контролем. Обработки селеном оказали стимулирующее воздействие на такие физиологические показатели, как длина оболочки стебля, содержание лигнина в оболочке стебля и накопление сухого вещества в различных тканях растения, что привело к увеличению общей сухой массы. Обработка селеном оказала ингибирующее влияние на содержание хлорофилла b и общее содержание хлорофилла, в то же время увеличив содержание хлорофилла a и соотношение хлорофилла a/b, что, в свою очередь, повлияло на фотосинтез риса.
Следовательно, соответствующие обработки селеном (применение 8 мкмоль·л⁻¹ хелатированного аминокислотами селена, 4 мкмоль·л⁻¹ селенита натрия и 8 мкмоль·л⁻¹ селенита натрия) могут улучшить рост проростков и твердость оболочки стебля, что было связано с изменениями таких параметров, как сухая масса, фотосинтетические пигменты и активность пероксидазы. Эти данные позволяют предположить, что различные селеновые удобрения могут положительно влиять на устойчивость риса к полеганию и его рост. Данное исследование может служить теоретической основой для применения селеновых удобрений.
1. Введение
Рис является важной зерновой культурой, и развитие рисоводства необходимо для обеспечения продовольственной безопасности [1]. Ароматный рис — это особый тип возделываемого риса, обладающий высокой экономической ценностью, но низкой урожайностью и склонностью к полеганию [2]. Полегание может приводить к проблемам низкой урожайности и низкого качества в рисоводстве. Основным типом полегания риса является стеблевое полегание, возникающее из-за слабого стебля [3]. Полегание риса может приводить к значительным потерям урожая и существенному снижению качества, а также вызывает значительное увеличение затрат на уборку урожая, что в конечном итоге приводит к экономическим потерям [4,5,6]. Следовательно, повышение устойчивости риса к полеганию имеет большое значение для рисоводства.
Селен (Se) является эссенциальным микроэлементом и важен для здоровья человека [7]. Производство селенобогащенного зерна пользуется популярностью, поскольку оно обеспечивает более безопасный и эффективный способ удовлетворения потребностей организма в селене [8,9,10]. Селен приносит пользу не только организму человека, но и росту риса, и предыдущее исследование показало, что умеренное добавление селена может способствовать повышению урожайности риса, увеличению качества, регулировать фотосинтез, а также повышать антиоксидантную способность и устойчивость [8]. Таким образом, исследование применения селеновых удобрений на рисе имеет большую практическую ценность.
Исследования показали, что концентрация селенового удобрения и способ его внесения являются двумя ключевыми факторами, влияющими на ростовые характеристики растений и эффективность накопления селена. Предыдущее исследование показало, что внесение низких концентраций селенового удобрения способствует росту и развитию растений, увеличивая урожайность, качество и антиоксидантную способность [11]. Что касается способа внесения селеновых удобрений, исследования показали, что внесение селеновых удобрений в почву, внекорневые опрыскивания селеном и обработка семян селеном являются популярными способами повышения уровня селена в растениях [12]. Исследования показали, что эффект обогащения селеном в растениях в результате внекорневого опрыскивания селеновыми удобрениями в восемь раз выше, чем при внесении в почву, а также что внекорневое опрыскивание селеновыми удобрениями является более экологически безопасным [13,14]. Кроме того, форма селена существенно влияет на эффективность селеновых удобрений [15]. Органический селен обладает низкой токсичностью и высокой эффективностью усвоения, в то время как неорганический селен имеет специфическую физиологическую токсичность и низкую эффективность усвоения, но хорошую биодоступность [16]. Другие исследования показали, что селенит натрия оказывает двойственное влияние на рост проростков риса в гидропонных условиях. При низких концентрациях обработка селеном способствовала росту проростков риса за счет увеличения биомассы, длины корней и антиоксидантной способности. Напротив, высокие концентрации селенита натрия могут наносить вред рису, приводя к снижению содержания хлорофилла, уменьшению биомассы и замедлению роста [17]. Следовательно, эффекты обработки селеном на рис можно изучать путем создания различных градиентов концентраций. Предыдущее исследование показало, что селен может увеличивать содержание лигнина и толщину клеточных стенок в растениях на клеточном уровне, и что применение селена может повышать устойчивость растений к полеганию [18]. Предыдущее исследование показало, что внесение селенового удобрения в почву увеличивало высоту растений и повышало индекс полегания [19]. Этот результат противоречит выводам предыдущих исследований. Таким образом, до сих пор ведутся споры о том, связан ли селен с полеганием риса положительно или отрицательно.
Можно заключить, что концентрация, способ внесения и форма селена, применяемого на рисовых растениях, являются важными факторами для регулирования полегания риса. В данном исследовании выдвигается гипотеза, что внекорневое опрыскивание селеном благотворно влияет на устойчивость риса к полеганию, и что различные формы селена будут оказывать различное влияние на устойчивость риса к полеганию. Поэтому был проведен гидропонный эксперимент с использованием двух сортов ароматного риса, выращиваемых при различных формах и уровнях селенового удобрения, и были исследованы рост проростков, твердость оболочки стебля, а также синтез целлюлозы и лигнина. Данное исследование было направлено на изучение способности различных форм и уровней селеновых удобрений положительно регулировать устойчивость риса к полеганию и предоставить теоретическую поддержку для применения селеновых удобрений и обеспечения продовольственной безопасности.
2. Материалы и методы
2.1. Описание эксперимента и обработки
Вегетационный эксперимент был проведен в октябре 2020 года на агрономическом факультете Южно-Китайского сельскохозяйственного университета, Гуанчжоу, провинция Гуандун, Китай. Эксперимент проводился при комнатной температуре и в естественных условиях. Температура колебалась от 17 до 29 °C, средняя температура составила 24,15 °C. Влажность колебалась от 52% до 81%, средняя влажность составила 66,86%. Сорта ароматного риса, использованные в эксперименте, — Сянъясянчжань и Юйсяньючжань, которые были предоставлены лабораторией рисоводства Южно-Китайского сельскохозяйственного университета. Сянъясянчжань и Юйсяньючжань являются популярными высококачественными сортами ароматного риса в Южном Китае. Сянъясянчжань — это сорт обычного индийского риса. Он характеризуется слабой агрономической устойчивостью, относительно низкой урожайностью и высокой степенью чувствительности к условиям окружающей среды [20]. Юйсяньючжань — это суперсорт риса с большим потенциалом высокой продуктивности и широкой адаптивностью [21]. Между двумя сортами риса существуют различия. Следовательно, можно изучить влияние обработки селеном на разные сорта риса, используя Сянъясянчжань и Юйсяньючжань в качестве экспериментальных материалов.
Два сорта ароматного риса (Сянъясянчжань и Юйсяньючжань) были использованы в качестве экспериментальных материалов. Два сорта ароматного риса обрабатывали двумя формами селеновых удобрений: неорганическим селеном (селенит натрия) и органическим селеном (хелатированный аминокислотами селен). Обработки селеновыми удобрениями проводились путем внекорневого опрыскивания, и пять вариантов внекорневого опрыскивания селеновыми удобрениями были следующими: без обработки селеновым удобрением (CK); внекорневое опрыскивание 4 мкмоль·л⁻¹ хелатированного аминокислотами селена (T1); внекорневое опрыскивание 8 мкмоль·л⁻¹ хелатированного аминокислотами селена (T2); внекорневое опрыскивание 4 мкмоль·л⁻¹ селенита натрия (T3); и внекорневое опрыскивание 8 мкмоль·л⁻¹ селенита натрия (T4). Перед посевом пустые зерна Юйсяньючжань и Сянъясянчжань были удалены методом водной сепарации. Проростки выращивали в субстрате, и пересадку проводили, когда проростки достигали фазы трех листьев. Однородно развитые проростки были пересажены в гидропонные контейнеры, содержащие 15 л питательной среды. Через три дня было проведено первое внекорневое опрыскивание, и опрыскивание селеновым удобрением проводилось всего 4 раза. Питательную среду меняли каждые 7 дней. Питательный раствор для роста проростков в контейнерах представлял собой питательный раствор Кимуры B, значение pH было доведено до 4,7–4,9. Питательный раствор включал следующее: маточный раствор 1 объемом 500 мл с KNO₃ (9,25 г) и Ca(NO₃)₂·4H₂O (43,16 г); маточный раствор 2 объемом 500 мл с MgSO₄·7H₂O (67,50 г), K₂SO₄ (7,95 г) и (NH₄)₂SO₄ (24,1 г); маточный раствор 3 объемом 500 мл с KH₂PO₄ (12,4 г); маточный раствор 4 объемом 1000 мл с FeSO₄·7H₂O (5,57 г) и Na₂EDTA (7,45 г); и маточный раствор 5 объемом 500 мл с MnCl₂·4H₂O (0,9 г), ZnSO₄·7H₂O (0,11 г), CuSO₄·5H₂O (0,04 г), H₃BO₃ (1,43 г) и (NH₄)₆Mo₇O₂₄·4H₂O (0,09 г). Маточный раствор в соотношении 1:1000 являлся окончательным питательным раствором для роста проростков. Размер каждого гидропонного контейнера составлял 65,0 см × 41,0 см × 15,5 см, с тремя повторными контейнерами для каждой обработки. Проростки в каждом контейнере высаживались с густотой посадки 2,0 см × 2,0 см.
2.2. Отбор образцов и измерения
Отбор образцов проводился через одну неделю после обработки. Сорок проростков были отобраны для измерения морфологических показателей. Восемнадцать проростков были собраны для измерения твердости оболочки стебля. Девяносто проростков риса были случайным образом отобраны из каждой обработки и разделены на различные части растений, а затем сразу после отбора погружены в жидкий азот. Затем образцы были помещены в холодильник при температуре −80 °C для последующего измерения физиологических показателей.
2.2.1. Определение морфологических показателей
Измерения сырой массы, сухой массы и морфологических показателей проводились в шести повторностях. Высота растения — это средняя длина каждого растения от нижней точки основания стебля до самого высокого конца листа. Длина корня — это расстояние от окончания корневой системы до нижней части основания стебля. Листовой возраст определяется количеством листьев, появившихся на главном стебле, и длиной листьев, которые не полностью развернулись. Грубое сравнение проводилось с соседним листом в процентах от вытянутой длины. Корни, стебли и листья были разделены, и для всех трех растений была измерена сырая масса каждой части. Сумма представляет общую сырую массу проростков. Каждая часть была высушена в сушильном шкафу при 80 °C до достижения постоянной массы, после чего была измерена сухая масса каждой части и общая сухая масса.
2.2.2. Определение содержания хлорофилла
Свежие образцы листьев (0,100 г) были взяты и растерты с 7,5 мл 95% этанола и помещены в центрифужную пробирку объемом 10 мл. Их поместили в темное место на 24 ч, а затем центрифугировали при 5000 об/мин в течение 10 мин при 4 °C. Оптическая плотность супернатанта была зарегистрирована при длинах волн 665 нм, 649 нм и 470 нм. Содержание хлорофилла a, содержание хлорофилла b и общее содержание хлорофилла были оценены [22].
2.2.3. Определение содержания лигнина и целлюлозы
Для определения содержания целлюлозы и лигнина в различных частях проростков риса использовали набор для обнаружения содержания лигнина и набор для обнаружения содержания целлюлозы (Beijing Suolaibao Technology Co. Ltd., Пекин, Китай) [23].
2.2.4. Определение активности пероксидазы (POD)
Для определения активности POD использовали метод гваякола. 100 мкл 0,3% H₂O₂, 285 мкл 0,2% гваякола и 300 мкл 0,05 моль·л⁻¹ натрий-фосфатного буфера смешивали с 15 мкл неочищенного ферментного экстракта. Значение оптической плотности регистрировали каждые 30 с при длине волны 470 нм, общая продолжительность измерения составляла 2 мин. Активность POD была определена как U·г⁻¹ сырой массы [24].
2.2.5. Определение твердости оболочки стебля
Для измерения твердости оболочки стебля проростков риса использовали измеритель устойчивости к полеганию (YYD-1A, Topyunong Science and Technology Co., Ханчжоу, Чжэцзян, Китай) [19,25]. Единица измерения твердости оболочки стебля выражается в Н.
2.3. Анализ данных
Данные были зарегистрированы в Microsoft Excel 2019 и проанализированы с помощью Statistics version 8.0 для дисперсионного анализа и множественных сравнений. Средние значения каждой обработки сравнивались с помощью теста наименьшей существенной разности (НСР) при 5% уровне вероятности. Корреляционный анализ Пирсона был выполнен для данных с использованием программного обеспечения IBM SPSS Statistics 21. Рисунки были созданы с помощью Origin 2021.
3. Результаты
3.1. Дисперсионный анализ
Сорт существенно влиял на сырую массу корней, общую сырую массу, высоту растений, длину оболочки стебля, площадь листьев на растение, твердость оболочки стебля, активность пероксидазы в листьях, содержание лигнина в листьях и содержание лигнина в корнях (p < 0,05). Обработка селеном существенно влияла на сырую массу корней, сухую массу, высоту растений, содержание длины оболочки стебля, количество листьев, общее содержание хлорофилла, твердость оболочки стебля, активность пероксидазы в листьях, активность пероксидазы в оболочке стебля и содержание лигнина в корнях (p < 0,05). Взаимодействие сорта и обработки селеном существенно влияло на сырую массу листьев, высоту растений, длину оболочки стебля, количество листьев, общее содержание хлорофилла, твердость оболочки стебля, активность пероксидазы в оболочке стебля, содержание целлюлозы в корнях и содержание лигнина в листьях (p < 0,05) (Таблица 1).
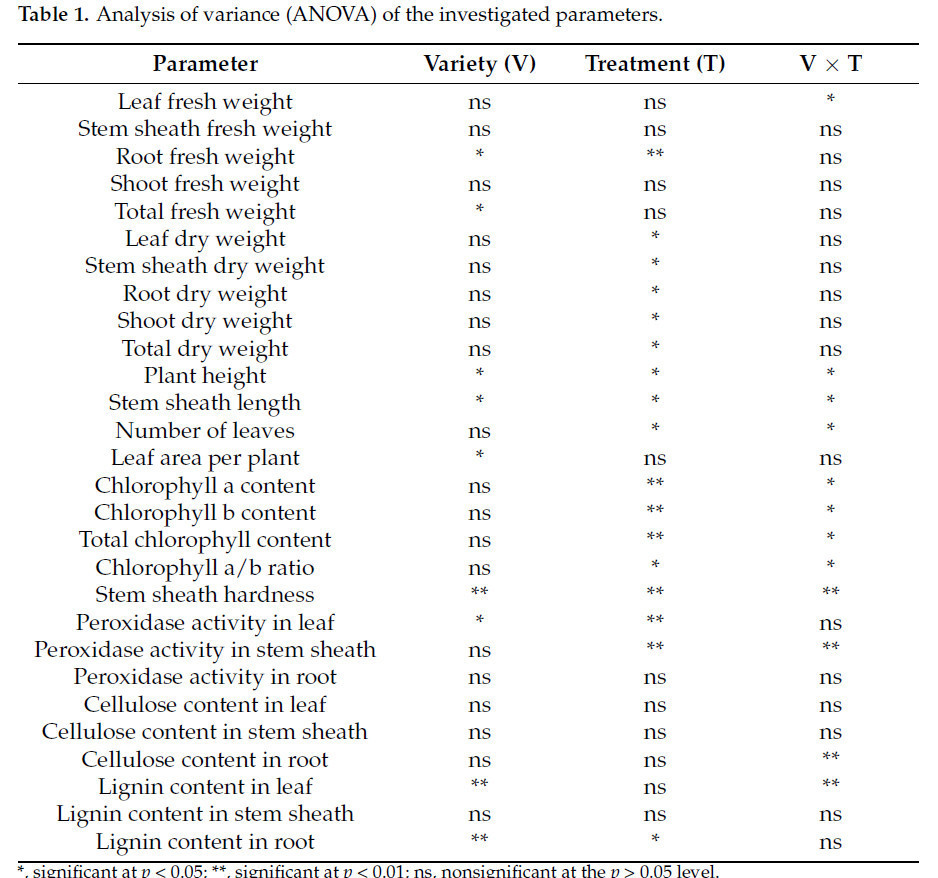
Таблица 1. Дисперсионный анализ (ANOVA) исследуемых параметров.
3.2. Сырая масса
Для сорта Сянъясянчжань по сравнению с контролем (CK) обработки T2 и T4 значительно увеличили сырую массу листьев на 33,88% и 32,09% соответственно. Сырая масса листьев Юйсяньючжань немного снизилась (Рисунок 1А). Сырая масса оболочки стебля Сянъясянчжань значительно увеличилась на 37,51%, 39,66% и 45,65% при обработках T1, T2 и T4 соответственно (Рисунок 1B). Для Сянъясянчжань по сравнению с контролем (CK) обработка T4 значительно увеличила сырую массу корней. Сырая масса корней Юйсяньючжань увеличилась на 40,10% и 47,23% при обработках T3 и T4 соответственно (Рисунок 1C). По сравнению с контролем (CK), обработки T1, T2 и T4 увеличили сырую массу побегов у Сянъясянчжань на 30,49%, 37,12% и 39,44% соответственно (Рисунок 1D). По сравнению с контролем (CK), обработки T2 и T4 значительно увеличили общую сырую массу у Сянъясянчжань на 36,31% и 44,87% соответственно (Рисунок 1E).
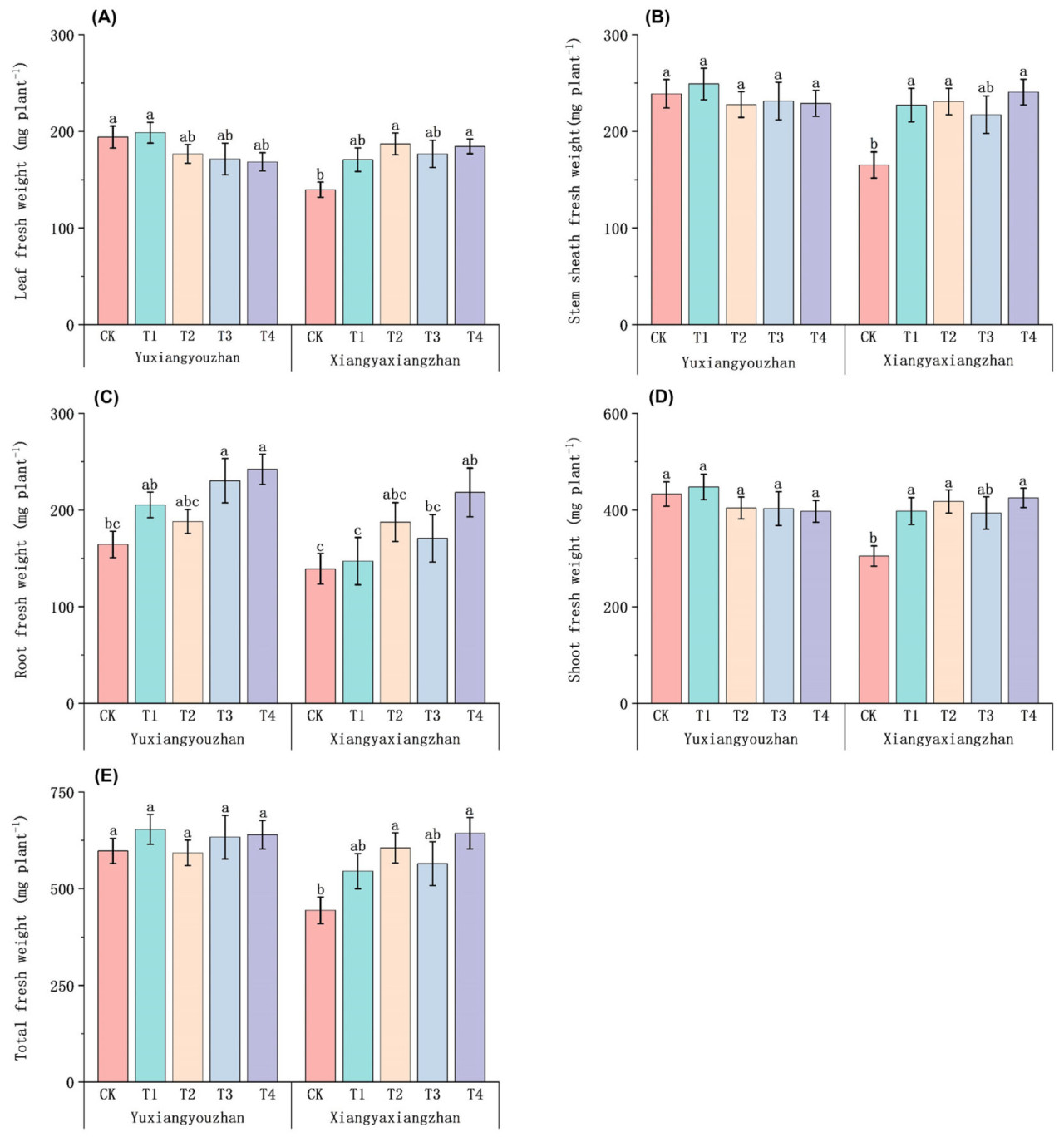
Рисунок 1. Влияние применения селена на сырую массу ароматного риса. Сырая масса листьев (A), сырая масса оболочки стебля (B), сырая масса корней (C), сырая масса побегов (D) и общая сырая масса (E). CK: без селенового удобрения; T1: 4 мкмоль·л⁻¹ хелатированного аминокислотами селена; T2: 8 мкмоль·л⁻¹ хелатированного аминокислотами селена; T3: 4 мкмоль·л⁻¹ селенита натрия; T4: 8 мкмоль·л⁻¹ селенита натрия. Строчные буквы обозначают значимые различия между обработками (тест НСР, p < 0,05).
3.3. Сухая масса
По сравнению с контролем (CK), обработка Se увеличила сухую массу листьев. Сухая масса листьев Юйсяньючжань значительно увеличилась на 31,61% при обработке T1. Сухая масса листьев Сянъясянчжань значительно увеличилась при обработке T4 (Рисунок 2A). По сравнению с контролем (CK), обработки T2 и T4 значительно увеличили сухую массу оболочки стебля у Сянъясянчжань на 43,94% и 70,38% соответственно (Рисунок 2B). По сравнению с контролем (CK), сухая масса корней Сянъясянчжань и Юйсяньючжань значительно увеличилась при обработке T4 (Рисунок 2C). По сравнению с контролем (CK), обработки T2 и T4 значительно увеличили сухую массу побегов у Сянъясянчжань на 36,14% и 47,08% соответственно. Сухая масса побегов Юйсяньючжань значительно увеличилась на 32,05% при обработке T1 (Рисунок 2D). Общая сухая масса была улучшена при обработках Se. Для Сянъясянчжань по сравнению с контролем (CK) обработки T2 и T4 значительно увеличили общую сухую массу на 35,41% и 52,78% соответственно (Рисунок 2E).
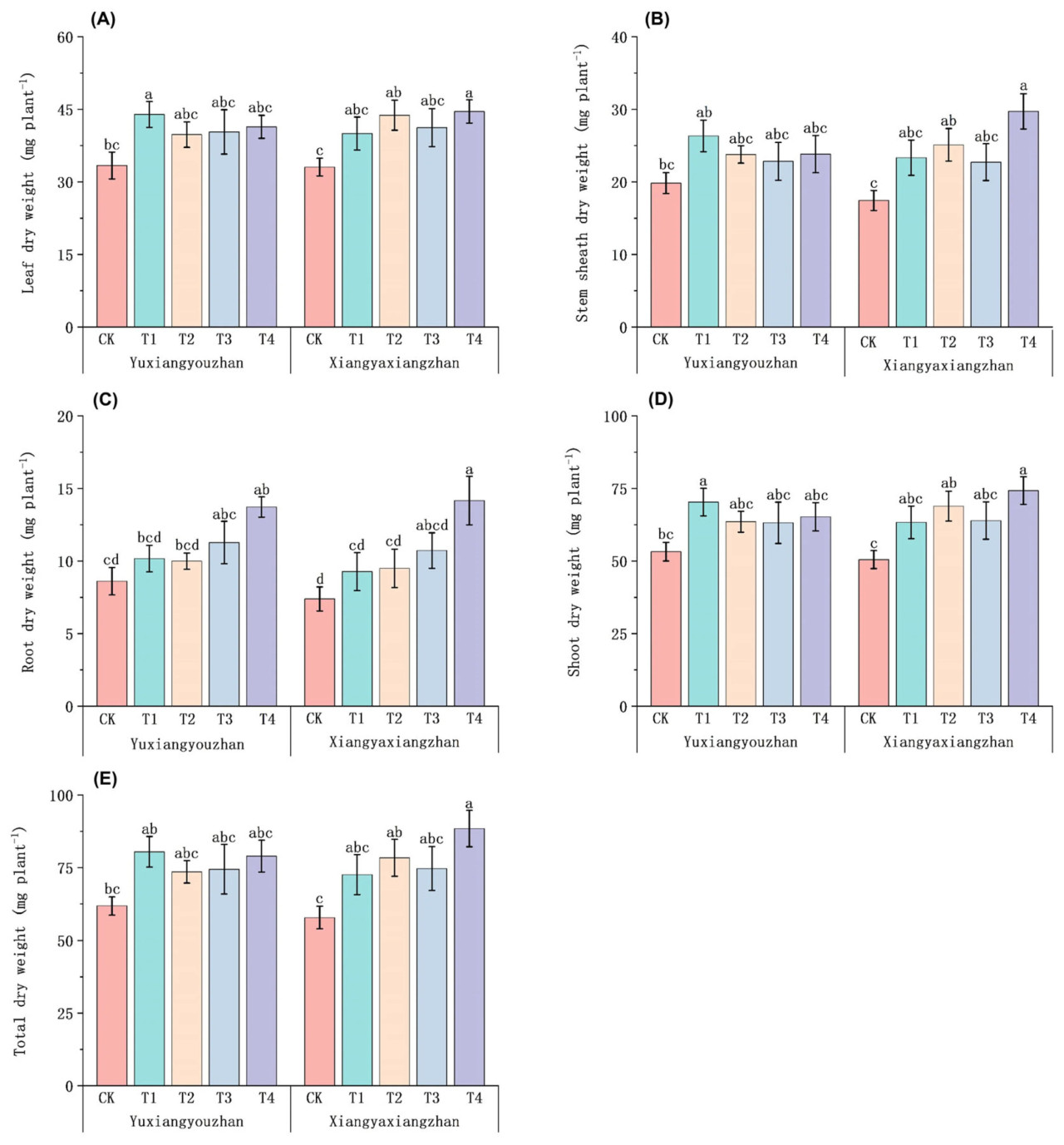
Рисунок 2. Влияние применения селена на сухую массу ароматного риса. Сухая масса листьев (A), сухая масса оболочки стебля (B), сухая масса корней (C), сухая масса побегов (D) и общая сухая масса (E). CK: без селенового удобрения; T1: 4 мкмоль·л⁻¹ хелатированного аминокислотами селена; T2: 8 мкмоль·л⁻¹ хелатированного аминокислотами селена; T3: 4 мкмоль·л⁻¹ селенита натрия; T4: 8 мкмоль·л⁻¹ селенита натрия. Строчные буквы обозначают значимые различия между обработками (тест НСР, p < 0,05).
3.4. Высота растений, длина стебля, листовой возраст и площадь листьев проростков риса
Для Сянъясянчжань по сравнению с контролем (CK) применение Se немного увеличило высоту растений. Высота растений Юйсяньючжань значительно увеличилась при обработке T4 по сравнению с контролем (CK) (Рисунок 3A). По сравнению с контролем (CK), длина оболочки стебля Юйсяньючжань значительно увеличилась при обработках T3 и T4 соответственно (Рисунок 3B). По сравнению с контролем (CK), обработки T1, T3 и T4 значительно увеличили количество листьев у Сянъясянчжань (Рисунок 3C). По сравнению с контролем (CK), обработки Se не привели к изменениям площади листьев на растение (Рисунок 3D).
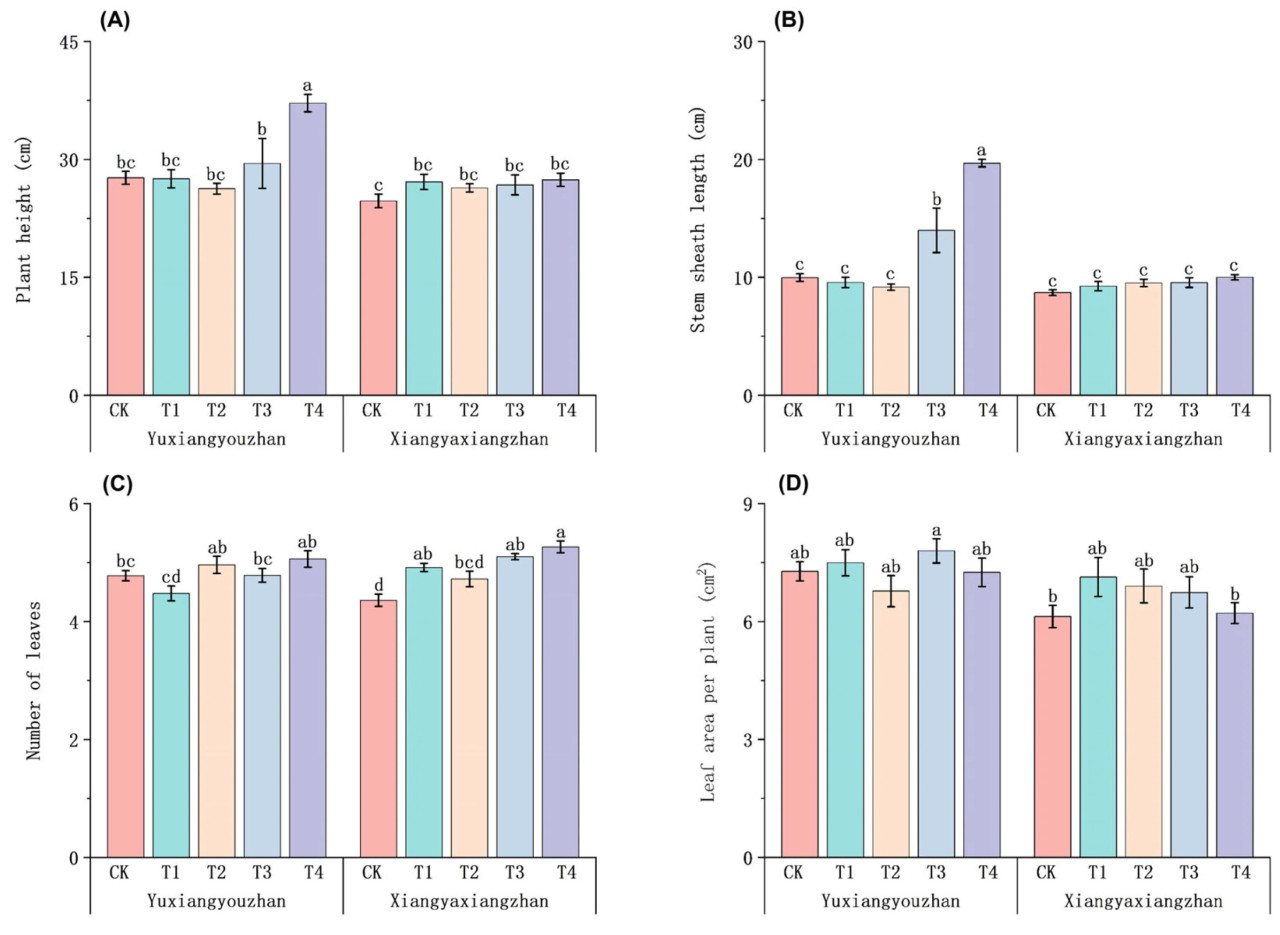
Рисунок 3. Влияние применения Se на морфологические показатели ароматного риса. Высота растений (A), длина оболочки стебля (B), количество листьев (C) и площадь листьев на растение (D). CK: без селенового удобрения; T1: 4 мкмоль·л⁻¹ хелатированного аминокислотами селена; T2: 8 мкмоль·л⁻¹ хелатированного аминокислотами селена; T3: 4 мкмоль·л⁻¹ селенита натрия; T4: 8 мкмоль·л⁻¹ селенита натрия. Строчные буквы обозначают значимые различия между обработками (тест НСР, p < 0,05).
3.5. Содержание хлорофилла в проростках риса
По сравнению с контролем (CK), обработки Se способствовали увеличению содержания хлорофилла a, в то время как они ингибировали содержание хлорофилла b и регулировали соотношение хлорофилла a/b. Для Сянъясянчжань по сравнению с контролем (CK) обработки T1, T2, T3 и T4 значительно увеличили содержание хлорофилла a. Содержание хлорофилла a у Юйсяньючжань значительно увеличилось при обработках T1, T2 и T3 (Рисунок 4A). По сравнению с контролем (CK), обработки Se привели к снижению содержания хлорофилла b (Рисунок 4B). В целом, обработки Se ингибировали общее содержание хлорофилла у Сянъясянчжань и Юйсяньючжань (Рисунок 4C). По сравнению с контролем (CK), обработки T1 и T4 увеличили соотношение хлорофилла a/b у Сянъясянчжань на 111,05% и 198,13% соответственно. Соотношение хлорофилла a/b у Юйсяньючжань значительно увеличилось на 100,16% и 123,79% при обработках T2 и T3 соответственно (Рисунок 4D).
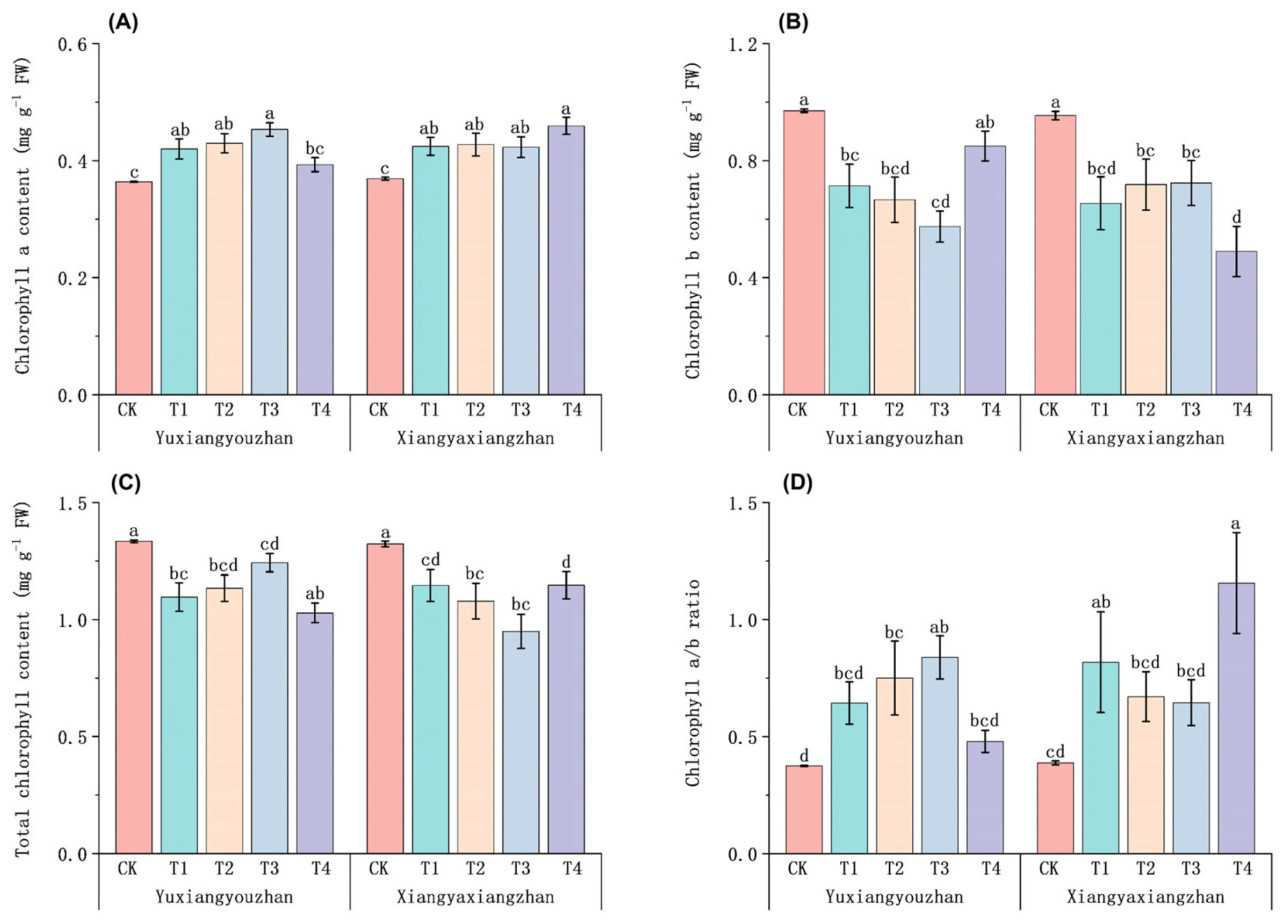
Рисунок 4. Влияние применения селена на содержание хлорофилла в ароматном рисе. Содержание хлорофилла a (A), содержание хлорофилла b (B), общее содержание хлорофилла (C) и соотношение хлорофилла a/b (D). CK: без селенового удобрения; T1: 4 мкмоль·л⁻¹ хелатированного аминокислотами селена; T2: 8 мкмоль·л⁻¹ хелатированного аминокислотами селена; T3: 4 мкмоль·л⁻¹ селенита натрия; T4: 8 мкмоль·л⁻¹ селенита натрия. Строчные буквы обозначают значимые различия между обработками (тест НСР, p < 0,05).
3.6. Твердость оболочки стебля проростков риса
По сравнению с контролем (CK), обработки T2, T3 и T4 значительно увеличили твердость оболочки стебля у Сянъясянчжань на 38,31%, 84,86% и 146,62% соответственно. Твердость оболочки стебля Юйсяньючжань значительно увеличилась на 38,77%, 21,57% и 54,73% при обработках T2, T3 и T4 соответственно (Рисунок 5).
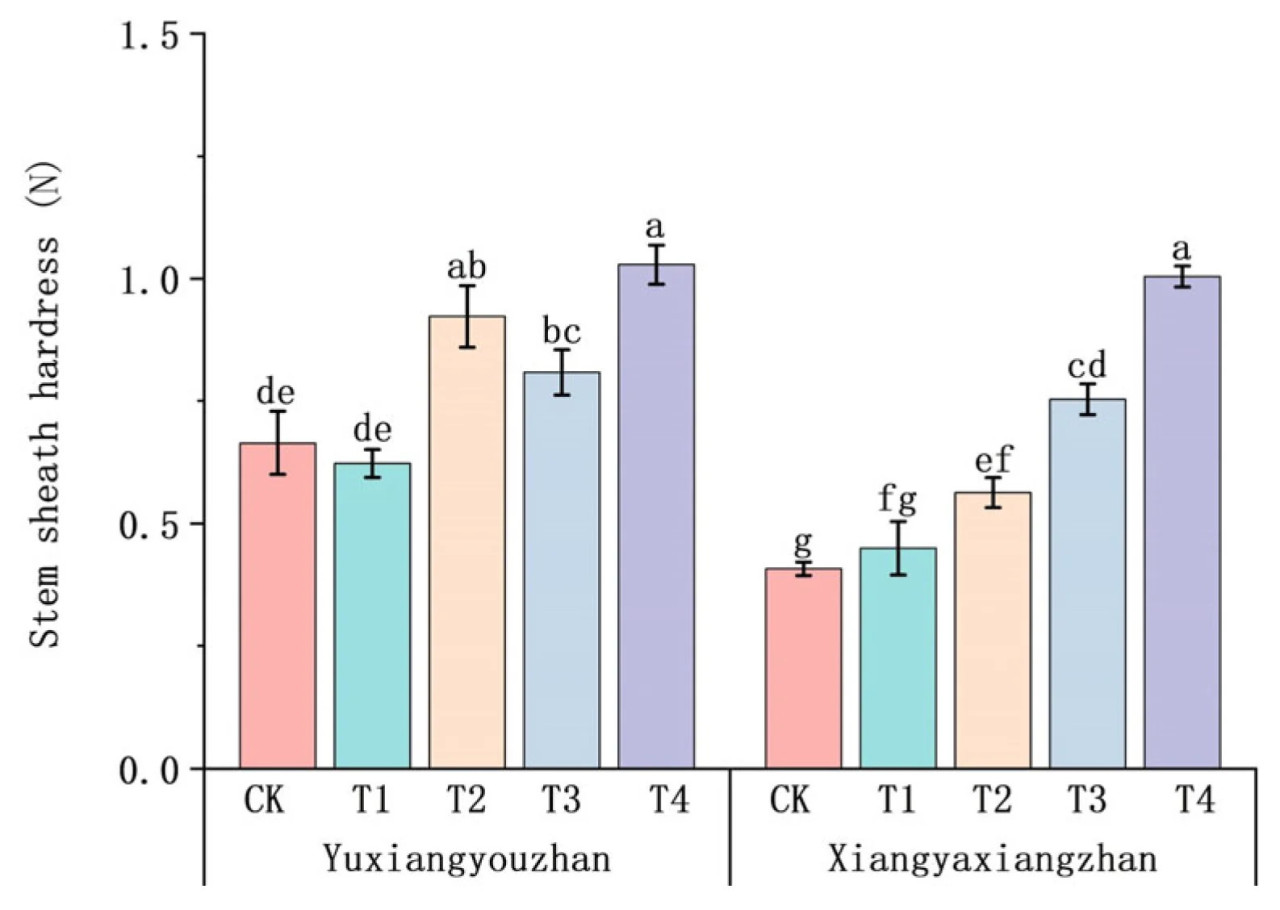
Рисунок 5. Влияние применения селена на твердость оболочки стебля ароматного риса. Твердость оболочки стебля. CK: без селенового удобрения; T1: 4 мкмоль·л⁻¹ хелатированного аминокислотами селена; T2: 8 мкмоль·л⁻¹ хелатированного аминокислотами селена; T3: 4 мкмоль·л⁻¹ селенита натрия; T4: 8 мкмоль·л⁻¹ селенита натрия. Строчные буквы обозначают значимые различия между обработками (тест НСР, p < 0,05).
3.7. Содержание лигнина в проростках риса
По сравнению с контролем (CK), обработка T4 значительно снизила содержание лигнина в листьях у Сянъясянчжань, в то время как содержание лигнина в листьях у Юйсяньючжань значительно увеличилось при обработке T3 (Рисунок 6A). По сравнению с контролем (CK), содержание лигнина в оболочке стебля у Сянъясянчжань не показало значительных изменений, в то время как содержание лигнина в оболочке стебля у Юйсяньючжань значительно увеличилось при обработках T3 и T4 (Рисунок 6B). По сравнению с контролем (CK), обработка T2 значительно снизила содержание лигнина в корнях у Юйсяньючжань. Значительных изменений содержания лигнина в корнях у Сянъясянчжань обнаружено не было (Рисунок 6C).
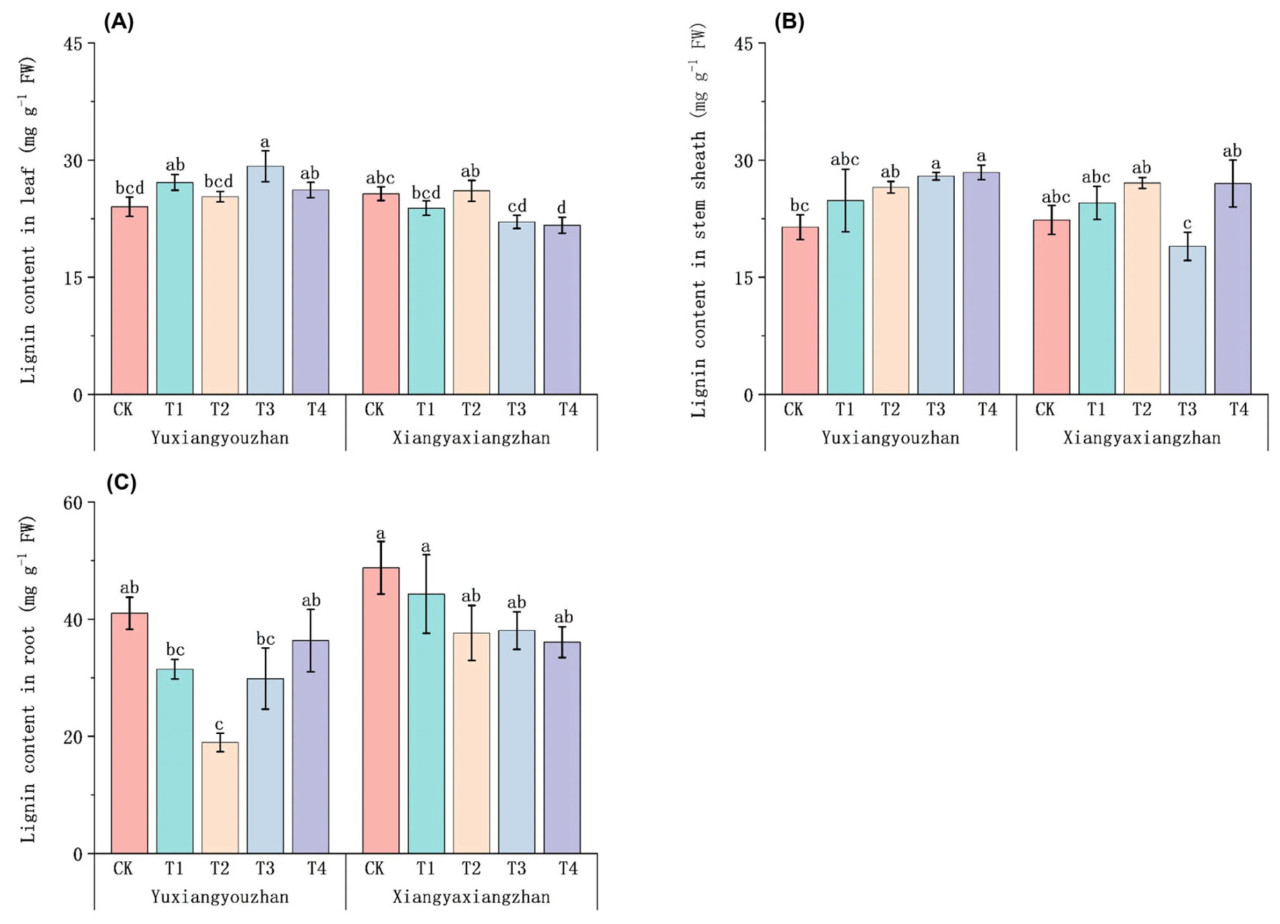
Рисунок 6. Влияние применения селена на лигнин в ароматном рисе. Содержание лигнина в листьях (A), содержание лигнина в оболочках стеблей (B) и содержание лигнина в корнях (C). CK: без селенового удобрения; T1: 4 мкмоль·л⁻¹ хелатированного аминокислотами селена; T2: 8 мкмоль·л⁻¹ хелатированного аминокислотами селена; T3: 4 мкмоль·л⁻¹ селенита натрия; T4: 8 мкмоль·л⁻¹ селенита натрия. Строчные буквы обозначают значимые различия между обработками (тест НСР, p < 0,05).
3.8. Содержание целлюлозы в проростках риса
По сравнению с контролем (CK), обработки селеном не оказали значительного влияния на содержание целлюлозы в листьях, оболочке стебля и корнях у Сянъясянчжань и Юйсяньючжань соответственно. Содержание целлюлозы в листьях Юйсяньючжань при T1 было значительно выше, чем у Сянъясянчжань при T2. Содержание целлюлозы в корнях Юйсяньючжань при T2 и T3 было значительно выше, чем у Сянъясянчжань при T3 (Рисунок 7).
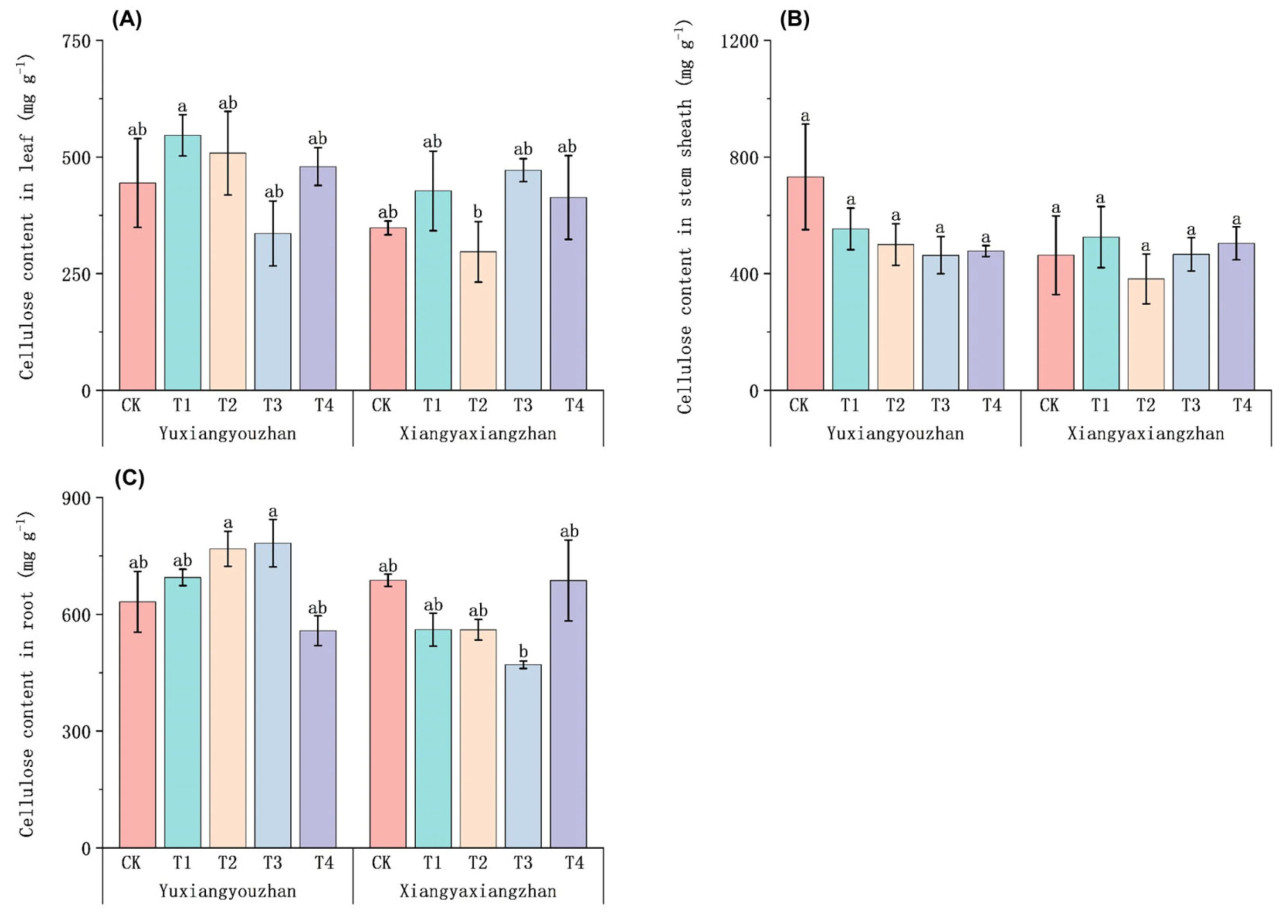
Рисунок 7. Влияние регулирования селеном на целлюлозу в ароматном рисе. Содержание целлюлозы в листьях (A), содержание целлюлозы в оболочках стеблей (B) и содержание целлюлозы в корнях (C). CK: без селенового удобрения; T1: 4 мкмоль·л⁻¹ хелатированного аминокислотами селена; T2: 8 мкмоль·л⁻¹ хелатированного аминокислотами селена; T3: 4 мкмоль·л⁻¹ селенита натрия; T4: 8 мкмоль·л⁻¹ селенита натрия. Строчные буквы обозначают значимые различия между обработками (тест НСР, p < 0,05).
3.9. Активность POD в проростках риса
По сравнению с контролем (CK), обработка T4 привела к более низкой активности POD в листьях у Юйсяньючжань (Рисунок 8A). По сравнению с контролем (CK), обработки T1 и T2 увеличили активность пероксидазы в оболочке стебля на 7,71% и 15,68% соответственно. Активность пероксидазы в стебле Юйсяньючжань значительно увеличилась на 43,63%, 30,25% и 35,03% при обработках T1, T3 и T4 соответственно (Рисунок 8B). Для Сянъясянчжань по сравнению с контролем (CK) обработки T3 и T4 увеличили активность пероксидазы в корнях на 38,10% и 41,27% соответственно. Значительных изменений активности пероксидазы в корнях у Юйсяньючжань обнаружено не было (Рисунок 8C).
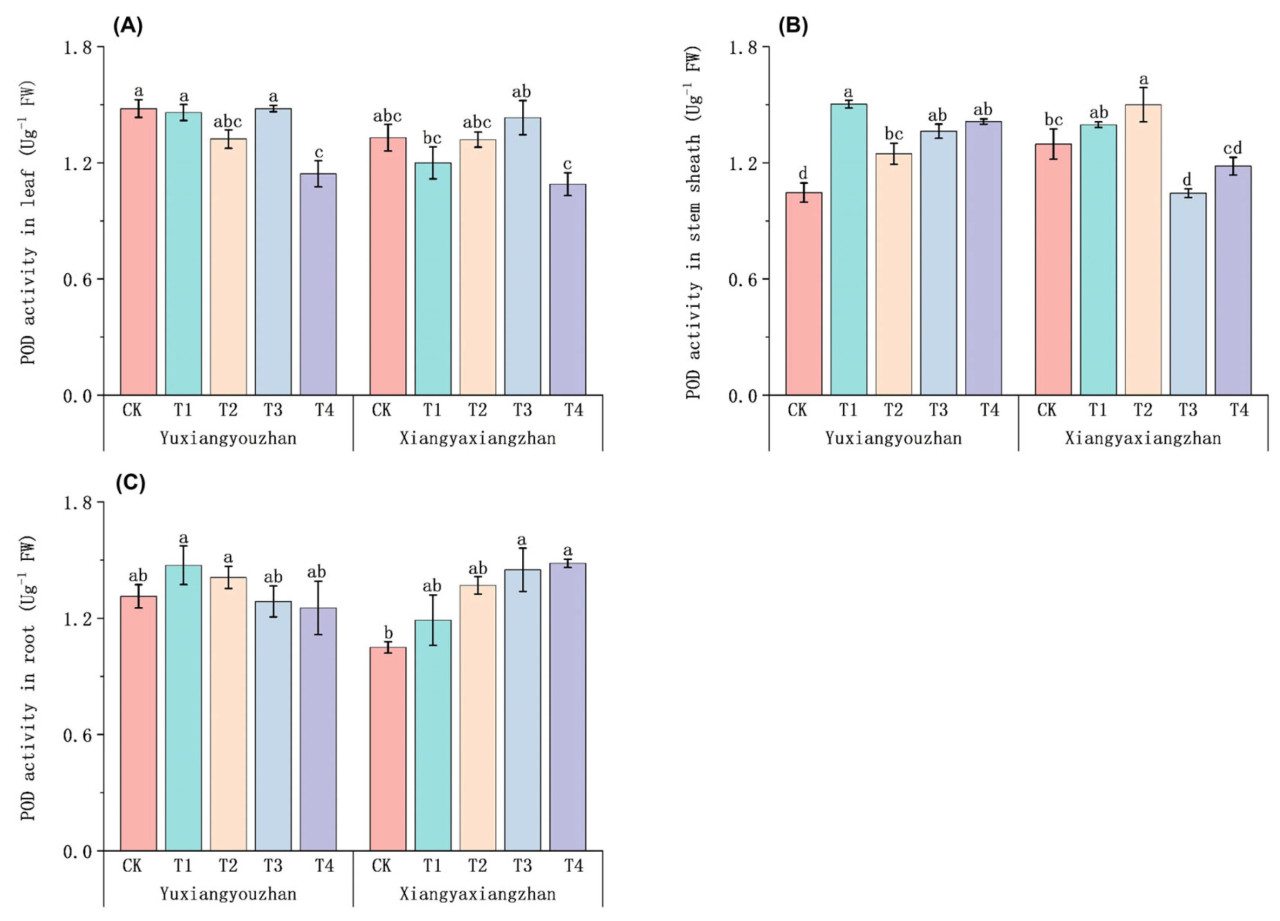
Рисунок 8. Влияние регулирования селеном на активность POD в ароматном рисе. Активность POD в листьях (A), активность POD в оболочке стебля (B) и активность POD в корнях (C). CK: без селенового удобрения; T1: 4 мкмоль·л⁻¹ хелатированного аминокислотами селена; T2: 8 мкмоль·л⁻¹ хелатированного аминокислотами селена; T3: 4 мкмоль·л⁻¹ селенита натрия; T4: 8 мкмоль·л⁻¹ селенита натрия. Строчные буквы обозначают значимые различия между обработками (тест НСР, p < 0,05).
3.10. Корреляционный анализ
Чтобы лучше понять взаимосвязь между улучшенным ростом проростков и твердостью оболочки стебля с другими исследуемыми параметрами, был проведен корреляционный анализ. Результаты показали, что общая сухая масса положительно коррелировала с сырой массой проростков, сухой массой проростков, количеством листьев, содержанием хлорофилла a, соотношением хлорофилла a/b, содержанием лигнина в оболочке стебля и активностью пероксидазы в корнях, в то время как она отрицательно коррелировала с содержанием хлорофилла b, общим содержанием хлорофилла, содержанием целлюлозы в оболочке стебля и активностью пероксидазы в листьях. Твердость оболочки стебля положительно коррелировала с сырой массой проростков, сухой массой проростков, содержанием лигнина в оболочке стебля и активностью пероксидазы в корнях, в то время как она отрицательно коррелировала с содержанием лигнина в корнях. Кроме того, этот результат показал взаимосвязь между улучшенным ростом проростков и хорошей твердостью оболочки стебля (Рисунок 9).
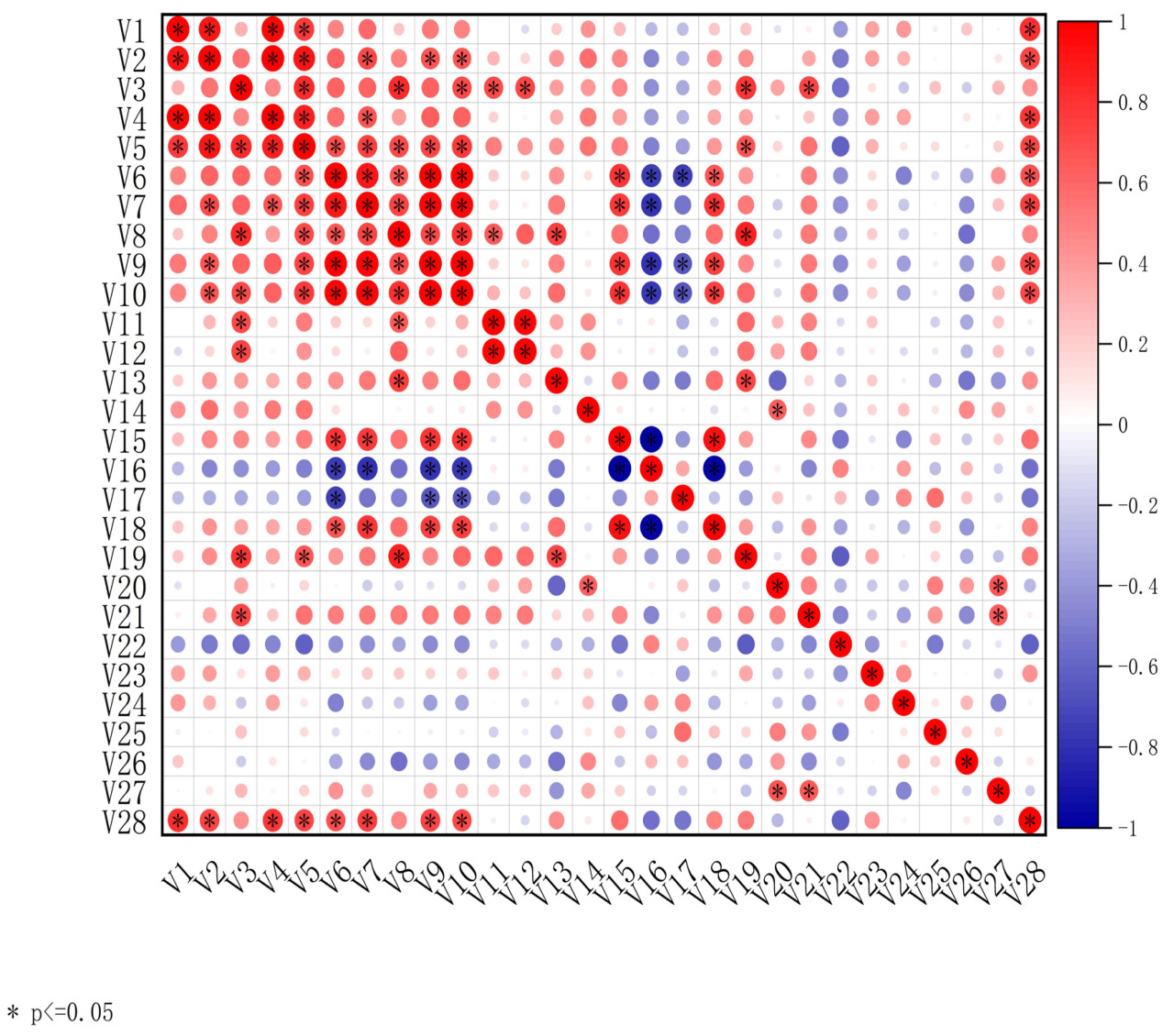
Рисунок 9. Корреляционный анализ между исследуемыми параметрами. V1, Сырая масса листьев; V2, Сырая масса оболочки стебля; V3, Сырая масса корней; V4, Сырая масса побегов; V5, Общая сырая масса; V6, Сухая масса листьев; V7, Сухая масса оболочки стебля; V8, Сухая масса корней; V9, Сухая масса побегов; V10, Общая сухая масса; V11, Высота растений; V12, Длина оболочки стебля; V13, Количество листьев; V14, Площадь листьев на растение; V15, Содержание хлорофилла a; V16, Содержание хлорофилла b; V17, Общее содержание хлорофилла; V18, Соотношение хлорофилла a/b; V19, Твердость оболочки стебля; V20, Содержание лигнина в листьях; V21, Содержание лигнина в оболочке стебля; V22, Содержание лигнина в корнях; V23, Содержание целлюлозы в листьях; V24, Содержание целлюлозы в оболочке стебля; V25, Содержание целлюлозы в корнях; V26, Активность пероксидазы в листьях; V27, Активность пероксидазы в оболочке стебля; V28, Активность пероксидазы в корнях. обозначает значимые различия при p < 0,05. Красный цвет указывает на положительную корреляцию, а синий — на отрицательную корреляцию. Чем глубже цвет, тем выше корреляция.
4. Обсуждение
Полегание является важным ограничивающим фактором в производстве зерновых культур, поскольку оно ограничивает урожайность и качество за счет изгибания или ломки стеблей [26]. Согласно многочисленным исследованиям, такие характеристики, как сорт риса, высота растений, длина стебля и толщина стенок стебля, являются основными факторами, влияющими на полегание риса [27,28,29]. Исследования показали, что индекс полегания связан с высотой растений и толщиной стебля, толщиной стенок и площадью поперечного сечения [30,31,32]. Данное исследование показало, что применение Se удобрения приводило к изменениям высоты или длины стебля проростков риса, и изменения варьировались в зависимости от форм и уровней Se удобрения (Рисунок 3). Это согласуется с результатами предыдущих исследований [33].
Согласно предыдущим исследованиям, добавление селена может способствовать синтезу лигнина в суспензионных клетках риса в условиях кадмиевого стресса [18] и в растениях перца в условиях кадмиевого стресса [34]. В настоящем исследовании было обнаружено, что применение селенового удобрения оказало положительное, но незначительное влияние на содержание лигнина в стебле (Рисунок 6B). Исследования показали, что применение селена может способствовать увеличению биомассы корней, а также синтезу целлюлозы и лигнина [35,36]. Как показано на Рисунке 1C и Рисунке 2C, применение селена увеличило сырую и сухую массу корней. Этот результат согласуется с предыдущими исследованиями. Как показано на Рисунке 7C, обработка низкими концентрациями селенита натрия способствовала синтезу целлюлозы в корнях. Все обработки оказали ингибирующее влияние на содержание лигнина в корнях. Сравнительный анализ двух исследований показал, что причина вышеуказанной ситуации в основном зависит от различной дозы селена, полученной растениями риса. Доза удобрения, использованная в предыдущих исследованиях, была ниже, чем в данном исследовании. Можно заключить, что применение селенового удобрения может способствовать синтезу целлюлозы и лигнина в корнях проростков риса. Согласно предыдущим исследованиям, полегание может быть разделено на стеблевой и корневой типы [37]. Таким образом, показано, что применение селеновых удобрений в правильной концентрации может способствовать росту корней, что в конечном итоге улучшает устойчивость риса к полеганию.
Предыдущие исследования показали, что применение селенового удобрения может увеличить активность POD в стеблях проростков риса [38]; в данном исследовании были обнаружены аналогичные результаты (Рисунок 8B). POD является важным антиоксидантным ферментом в растительных клетках, обладающим различными биологическими функциями, включая антиоксидантную активность, устойчивость к стрессу, клеточную сигнализацию и биосинтез лигнина [39,40]. Данное исследование показало, что применение Se удобрения увеличивало активность POD в стебле и увеличивало содержание лигнина и целлюлозы в стебле (Рисунок 6, Рисунок 7 и Рисунок 8). Структура стебля в основном состоит из нескольких сосудистых пучков, слоев толстостенных клеток ткани, степени их лигнификации и толщины корковой волокнистой ткани. Когда сосудистые пучки стебля толще и многочисленнее, толстостенные ткани имеют больше слоев клеток и большую лигнификацию. Чем толще корковая волокнистая ткань, тем выше устойчивость стеблей риса к изгибанию и полеганию [41]. Рисунки 5, 6, 7 и 8 показывают, что применение селенового удобрения способствовало активности стеблевой POD и увеличивало содержание целлюлозы, содержание лигнина и твердость оболочки стебля. Эти данные свидетельствуют о том, что применение селенового удобрения может увеличить активность стеблевой POD, способствуя увеличению содержания целлюлозы и лигнина в стебле. Таким образом, можно предварительно заключить, что применение Se удобрения может увеличить твердость оболочки стебля, что приводит к высокой устойчивости риса к полеганию. Конкретные причины и механизмы все еще требуют дальнейших исследований.
Юйсяньючжань — это суперсорт риса, подходящий для широких регионов Южного Китая [42]. Сянъясянчжань — это традиционно возделываемый сорт, чувствительный к температуре [43]. Эти два сорта являются разными сортами риса. Рисунки 1, 2, 3, 4, 5, 6, 7 и 8 показывают, что высота растений, длина стебля, сырая масса корней, общая сырая масса, твердость оболочки стебля, содержание лигнина в листьях и активность POD в листьях значительно различались между сортами. Более того, содержание лигнина в корнях и целлюлозы в стеблях двух сортов риса имело тенденцию к увеличению или уменьшению при одинаковом градиенте концентрации Se удобрения. На основе различных характеристик двух сортов риса можно предварительно заключить, что разница обусловлена различной чувствительностью двух сортов риса к селеновому удобрению. Этот вывод аналогичен предыдущему заключению [44]. В целом, под действием селенового удобрения показатели твердости оболочки стебля и содержания лигнина в стебле у Юйсяньючжань были выше, чем у Сянъясянчжань.
Селен существует в различных формах, таких как неорганический, органический и наноселен. Селенит натрия является типом селенитного селенового удобрения, а хелатированный аминокислотами селен является органическим селеном. Согласно предыдущим исследованиям, растения более склонны к поглощению селенита и селената. Однако неорганический селен обычно эффективно не усваивается организмом человека и может даже оказывать специфическое биотоксическое действие. Он может быть преобразован в органический селен посредством биохимических реакций внутри растения [45]. Анализ Рисунков 1, 2, 3, 4, 5, 6, 7 и 8 показал, что по сравнению с хелатированным аминокислотами селеном селенит натрия в целом оказывает более существенное стимулирующее влияние на стебель, сухое вещество корней и твердость оболочки стебля у двух сортов проростков риса. Эти данные свидетельствуют о том, что селенит натрия более эффективен, чем хелатированный аминокислотами селен, в повышении устойчивости риса к полеганию.
Предыдущие исследования показали, что подкормка Se способствует накоплению хлорофилла у риса [46,47]. В данном исследовании обработки Se увеличивали содержание хлорофилла a, ингибировали содержание хлорофилла b и общее содержание хлорофилла и регулировали соотношение хлорофилла a/b (Рисунок 4). Согласно предыдущим исследованиям [48], хлорофилл a предшествует хлорофиллу b, когда начинается синтез хлорофилла. Превращение хлорофилла приводит к образованию хлорофилла b, который является наиболее важным элементом в формировании хлорофилла. Механизм, с помощью которого селен влияет на превращение хлорофилла a в хлорофилл b, все еще требует дальнейших исследований. Рисунок 4 также показывает, что применение селенового удобрения увеличивало значение соотношения хлорофилла a/b. Согласно предыдущим исследованиям [49], низкое соотношение хлорофилла a/b благоприятствует поглощению световой энергии, тогда как высокое соотношение хлорофилла a/b обладает большей способностью противостоять фотоингибированию. Соотношение хлорофилла a/b было значительно и отрицательно коррелировано со скоростью фотосинтеза. Обработка Se повлияла на сырую/сухую массу проростков риса (Рисунок 1 и Рисунок 2). Можно заключить, что оптимизация применения селенового удобрения может в некоторой степени способствовать росту растений.
5. Выводы
Таким образом, применение селенового удобрения влияло на рост и физиологические показатели риса. Эти показатели различаются в зависимости от взаимодействия между сортами и обработками, например, регулирование активности пероксидазы путем внекорневого опрыскивания селеновым удобрением. Было обнаружено значительное увеличение твердости оболочки стебля у риса. После обработки хелатированным аминокислотами селеном и селенитом натрия в концентрации 8 мкмоль·л⁻¹ значения твердости оболочки у сортов Юйсяньючжань и Сянъясянчжань значительно увеличились — на 38,8% и 38,3% и на 54,7% и 146,6% соответственно. Кроме того, применение Se удобрения в соответствующей концентрации также способствует таким показателям, как целлюлоза стебля, целлюлоза корней и лигнин. Однако внекорневое опрыскивание селеновым удобрением может в некоторой степени ингибировать фотосинтез риса. Следовательно, возможно изучение новых способов снижения негативных последствий применения селеновых удобрений. Кроме того, в данном исследовании не измерялись уровни антиоксидантных ферментов, таких как супероксиддисмутаза и каталаза. Будущие исследования могут более глубоко изучить эти антиоксидантные ферменты. В целом, соответствующая обработка селеновым удобрением может повысить устойчивость риса к полеганию. Это связано с воздействием на твердость оболочки стебля, содержание лигнина в стебле, содержание лигнина в корнях и т.д., которые регулируют распределение накопления сухого вещества.
Ссылки
1. Shi, N. Analysis and development strategy on rice production & marketing trend in China. Chin. Rice 2015, 21, 1–5. [Google Scholar]
2. You, Q.; Huang, T. Research and breeding utilization of rice aromas. Fujian Rice Wheat Sci. Technol. 2002, 20, 30–33. [Google Scholar]
3. Zhang, Z. Causes and prevention strategies for rice lodging. New Agric. 2021, 21, 11. [Google Scholar]
4. Zhou, P.; Zhou, K.; Liu, T.; Wu, W.; Sun, C. Progress in monitoring research on rice lodging. J. Chin. Agric. Mech. 2019, 40, 162–168. [Google Scholar]
5. Zhao, X.; Shao, Z.; Wu, Y.; Zhao, Y.; Wang, Y.; Wang, Y.; Yang, L. Influence of artificial lodging at the grain-filling stage on plant growth, yield, and quality of super rice. Chin. J. Eco-Agric. 2018, 26, 980–989. [Google Scholar]
6. Wang, X.; Lu, Z.; Liu, W.; Lu, D.; Wang, S.; Wu, H.; Fang, Z.; He, X. Advances in lodging resistance of rice since the “green revolution”. Guangdong Agric. Sci. 2022, 49, 1–13. [Google Scholar]
7. Li, L.; Dong, J.; Li, L.; Wang, Z.; Cheng, S. Research progress of organic selenium-rich biological resources. Food Sci. Technol. 2021, 46, 61–66. [Google Scholar]
8. Jin, X.; Zhu, X.; Huang, J.; Du, L.; Wang, L.; Chen, J. Effects of Selenium on Chloroplast and Photosynthesis. Mol. Plant Breed. 2019, 17, 288–294. [Google Scholar]
9. Zhang, Y.; Wang, M.; Zhou, W.; Wang, D.; Yan, J. Improvement effect of different conditioners on acidic selenium-rich soil. Resour. Environ. Eng. 2024, 38, 34–39. [Google Scholar]
10. Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Raza, A.; Hawrylak-Nowak, B.; Matraszek-Gawron, R.; Mahmud, J.A.; Nahar, K.; Fujita, M. Selenium in plants: Boon or bane. Environ. Exp. Bot. 2020, 178, 104–170. [Google Scholar] [CrossRef]
11. Rizwan, M.; Ali, S.; Rehman MZ, U.; Rinklebe, J.; Tsang DC, W.; Tack FM, G. Effects of selenium on the uptake of toxic trace elements by crop plants: A review. Crit. Rev. Environ. Sci. Technol. 2021, 51, 2531–2566. [Google Scholar] [CrossRef]
12. Ebrahimi, N.; Stoddard, F.L.; Hartikainen, H.; Seppanen, M.M. Plant species and growing season weather influence the efficiency of selenium biofortification. Nutr. Cycl Agroecosyst. 2019, 114, 111–124. [Google Scholar] [CrossRef]
13. Ros, G.H.; Rotterdam, A.V.; Bussink, D.; Bindraban, P.S. Selenium fertilization strategies for biofortification of food: An agro-ecosystem approach. Plant Soil 2016, 404, 99–112. [Google Scholar] [CrossRef]
14. Deng, X.; Liu, K.; Li, M.; Zhang, W.; Zhao, X.; Zhao, Z.; Liu, X. Difference of selenium uptake and distribution in the plant and selenium form in the grains of rice with foliar spray of selenite or selenate at different stages. Field Crops Res. 2017, 211, 165–171. [Google Scholar] [CrossRef]
15. Sun, X.; Zhang, X.; Luo, Y.; Ye, S.; Zhuang, W.; Zhou, X. Effects of Four Valence state of Selenium on Part of Root Physiological Characteristics of Plum Seedlings. North. Hortic. 2021, 3, 41–46. [Google Scholar]
16. Xu, H.; Yan, J.; Qin, Y.; Xu, J.; Shohang MJ, I.; Wei, Y.; Gu, M. Effect of different forms of selenium on the physiological response and the cadmium uptake by rice under cadmium stress. Int. J. Environ. Res. Public Health 2020, 17, 6991. [Google Scholar] [CrossRef]
17. Liu, Y.; Ma, J.; Li, F.; Zeng, X.; Wu, Z.; Huang, Y.; Xue, Y.; Wang, Y. High Concentrations of Se Inhibited the Growth of Rice Seedlings. Plant 2024, 12, 1580. [Google Scholar] [CrossRef]
18. Cui, J.; Liu, T.; Li, Y.; Li, F. Selenium reduces cadmium uptake into rice suspension cells by regulating the expression of lignin synthesis and cadmium-related genes. Sci. Total Environ. 2018, 644, 602–610. [Google Scholar] [CrossRef]
19. Liu, X.; Huang, Z.; Li, Y.; Xie, W.; Li, W.; Tang, X.; Ashraf, U.; Kong, L.; Wu, L.; Wang, S.; et al. Selenium-silicon (Se-Si) induced modulations in physio-biochemical responses, grain yield, quality, aroma formation and lodging in fragrant rice. Ecotoxicol. Environ. Saf. 2020, 196, 110525. [Google Scholar] [CrossRef]
20. Liang, J.; Kong, L.; Hu, X.; Fu, C.; Bai, S. Chromosomal-level genome assembly of the high-quality Xian/Indica rice (Oryza sativa L.) Xiangyaxiangzhan. BMC Plant Biol. 2023, 23, 94. [Google Scholar] [CrossRef]
21. Chen, W.; Liao, G.; Sun, F.; Ma, Y.; Chen, Z.; Chen, H.; Tang, X.; Mo, Z. Foliar spray of La2O3 nanoparticles regulates the growth, antioxidant parameters, and nitrogen metabolism of fragrant rice seedlings in wet and dry nurseries. Environ. Sci. Pollut. Res. 2023, 30, 80349–80363. [Google Scholar] [CrossRef] [PubMed]
22. Li, Y.; Liang, L.; Li, W.; Ashraf, U.; Ma, L.; Tang, X.; Pan, S.; Tian, H.; Mo, Z. ZnO nanoparticle-based seed priming modulates early growth and enhances physio-biochemical and metabolic profiles of fragrant rice against cadmium toxicity. J. Nanobiotechnol. 2021, 19, 75. [Google Scholar] [CrossRef]
23. Deng, H.; Li, Y.; Ashraf, U.; Gui, R.; Wang, Z.; Nawaz, H.; Tang, X.; Duan, M.; Mo, Z. The application of liquid fertilizer with reduced nitrogen rate improves the lodging resistance in fragrant rice. J. Soil Sci. Plant Nutr. 2023, 23, 6071–6087. [Google Scholar] [CrossRef]
24. Lin, T.; Chen, X.; Ren, Y.; Qing, B.; Zhang, M.; Mo, Z.; Wang, S. Effect of iron oxide nanocoating on the seed germination, seedling growth, and antioxidant response of aromatic rice grown in the presence of different concentrations of rice straw extracts. J. Nanopart. Res. 2024, 26, 78. [Google Scholar] [CrossRef]
25. Hong, W.; Chen, Y.; Huang, S.; Li, Y.; Wang, Z.; Tang, X.; Pan, S.; Tian, H.; Mo, Z. Optimization of nitrogen-silicon (N-Si) fertilization for grain yield and lodging resistance of early-season indica fragrant rice under different planting methods. Eur. J. Agron. 2022, 136, 126508. [Google Scholar] [CrossRef]
26. Acreche, M.M.; Slafer, G.A. Lodging yield penalties as affected by breeding in Mediterranean wheats. Field Crops Res. 2011, 122, 40–48. [Google Scholar] [CrossRef]
27. Liu, Q.; Ma, J.; Zhao, Q.; Zhou, X. Physical traits related to rice lodging resistance under different simplified- cultivation methods. Agron. J. 2018, 110, 127–132. [Google Scholar] [CrossRef]
28. Jiang, M.; Yamamoto, E.; Yamamoto, T.; Matsubara, K.; Kato, H.; Adachi, S.; Nomura, T.; Kamahora, E.; Ma, J.; Ookawa, T. Mapping of QTLs associated with lodging resistance in rice (Oryza sativa L.) using the recombinant inbred lines derived from two high yielding cultrivars, Tachisugata and Hokuriku 193. Plant Growth Regul. 2019, 87, 267–276. [Google Scholar] [CrossRef]
29. Tsugawa, S.; Shima, H.; Ishimoto, Y.; Ishikawa, K. Thickness-stiffness trade-off improves lodging resistance in rice. Sci. Rep. 2023, 13, 10828. [Google Scholar] [CrossRef]
30. Yang, Y.; Zhu, Z.; Zhang, Y.; Chen, T.; Zhao, Q.; Zhou, L.; Yao, S.; Zhang, Y.; Dong, S.; Wang, C. Relationships between lodging resistance and stem morphological traits in different rice varieties (lines). Jiangsu Agric. Sci. 2011, 27, 231–235. [Google Scholar]
31. Lai, S.; Chen, C.; Lai, S.; Wang, L.; Chen, W. Genotypic differences and correlations between rice main agronomic traits and lodging-resistance. J. Nucl. Agric. Sci. 2018, 32, 1256–1266. [Google Scholar]
32. Zou, D.; Tang, Q.; Huang, Y.; Liu, L.; Fang, S.; Lv, G.; Kuang, N.; Luo, Y.; Mao, R.; Zhang, M. Effects of anti-pour agent on yield and lodging resistance of high-quality rice. Chin. Agric. Sci. Bull. 2023, 39, 8–13. [Google Scholar]
33. Zhou, K. The Effects of Nanoselenium and Sodium Selenite on Rice Seed Germination and Seedling Growth. Master’s Thesis, Heilongjiang University, Heilongjiang, China, 2023. [Google Scholar]
34. Li, D.; Zhou, C.; Ma, J.; Wu, Y.; Kang, L.; An, S.; Zhang, J.; Deng, K.; Li, J.; Pan, C. Nanoselenium transformation and inhibition of cadmium accumulation by regulating the lignin biosynthetic pathway and plant hormone signal transduction in pepper plants. J. Nanobiotechnol. 2021, 19, 316. [Google Scholar] [CrossRef] [PubMed]
35. Liu, Y.; Lv, H.; Yang, N.; Li, Y.; Liu, B.; Rensing, C.; Dai, J.; Fekih, I.B.; Wang, L.; Mazhar, S.H.; et al. Roles of root cell wall components and root plaques in regulating elemental uptake in rice subjected to selenite and different speciation of antimony. Environ. Exp. Bot. 2019, 163, 36–44. [Google Scholar] [CrossRef]
36. Wang, L.; Wu, K.; Liu, Z.; Li, Z.; Shen, J.; Wu, Z.; Liu, H.; You, L.; Yang, G.; Rensing, C.; et al. Selenite reduced uptake/translocation of cadmium via regulation of assembles and interactions of pectins, hemicelluloses, lignins, callose and casparian 15 strips in rice roots. J. Hazard. Mater. 2023, 448, 130812. [Google Scholar] [CrossRef]
37. Sun, Y.; Zeng, X.; Wang, Q.; Song, Q.; Wang, M.; Li, X.; Feng, Y. Research progress on influencing factors and solving strategies of lodging resistance in rice. Heilongjiang Agric. Sci. 2023, 9, 137–142. [Google Scholar]
38. Duan, M.; Cheng, S.; Lu, R.; Lai, R.; Zheng, A.; Ashraf, U.; Fan, P.; Du, B.; Luo, H.; Tang, X. Effect of foliar sodium selenate on leaf senescence of fragrant rice in south China. Appl. Ecol. Environ. Res. 2019, 19, 3343–3351. [Google Scholar] [CrossRef]
39. Wan, Y.; Camara, A.Y.; Yu, Y.; Wang, Q.; Guo, T.; Zhu, L.; Li, H. Cadmium dynamics in soil pore water and uptake by rice: Influences of soil-applied selenite with different water managements. Environ. Pollut. 2018, 240, 523–533. [Google Scholar] [CrossRef]
40. Mehrabanjoubani, P.; Abdolzadeh, A.; Sadeghipour, H.R.; Aghdasi, M.; Bagherieh-Najjar, M.B.; Barzegargolchini, B. Silicon increases cell wall thickening and lignification in rice (Oryza sativa) root tip under excess Fe nutrition. Plant Physiol. Biochem. 2019, 144, 264–273. [Google Scholar] [CrossRef]
41. Liu, H. Effects of Silicon on Rice Resistance Against Lodging and Bacterial Blight; Chinese Academy of Agricultural Sciences: Beijing, China, 2015. [Google Scholar]
42. Luo, Y.; Tian, J.; Lin, Q.; Jiang, Y.; Xiao, L.; Tang, X. Effects of cultivation techniques characterized by enhancing sources, activating sinks and improving quality on the yield of extensive-adaptation super rice in South China. J. Northwest AF Univ. 2014, 42, 55–60. [Google Scholar]
43. Wang, H. Cultivation techniques for ivory fragrance with resistance to lodging. Rural Econ. Technol. 2019, 30, 40–41. [Google Scholar]
44. Wan, Y.; Wang, K.; Liu, Z.; Yu, Y.; Wang, Q.; Li, H. Effect of selenium on the subcellular distribution of cadmium and oxidative stress induced by cadmium in rice (Oryza sativa L.). Environ. Sci. Pollut. Res. 2019, 26, 16220–16228. [Google Scholar] [CrossRef] [PubMed]
45. Yi, T.; Yao, J. Development status and countermeasures of organic selenium agricultural products industry in hubei province. South-Cent. Agric. Sci. Technol. 2023, 44, 95–97. [Google Scholar]
46. David, O.A.; Akomolafe, G.F.; Jolayemi, O.L.; Olawumo, I.J.; Awoyemi, O.O. Investigating the effectiveness of selenite on drought stressed upland rice. Indian J. Agric. Res. 2020, 54, 168–174. [Google Scholar] [CrossRef]
47. Luo, H.; Xing, P.; Liu, J.; Pan, S.; Tang, X.; Duan, M. Selenium improved antioxidant response and photosynthesis in fragrant rice (Oryza sativa L.) seedlings during drought stress. Physiol. Mol. Biol. Plants 2021, 27, 2849–2858. [Google Scholar] [CrossRef]
48. Guo, C.; Fang, L.; Xu, X. Chlorophyll-b deficient and photosynthesis in plants. Plant Physiol. Commun. 2006, 42, 967–973. [Google Scholar]
49. Zhou, H.; Huang, S. Effects of sink source relationship on chlorophyll content and photosynthetic characteristics of rice. J. Green Sci. Technol. 2017, 24, 147–149. [Google Scholar]

Zhuang J, Fang Y, Zheng J, Duan Y, Liu X, Mo Z. Effects of Selenium Foliar Spraying on Seedling Growth and Stem Sheath Hardness in Fragrant Rice. Agriculture. 2025; 15(3):335. https://doi.org/10.3390/agriculture15030335
Перевод статьи «Effects of Selenium Foliar Spraying on Seedling Growth and Stem Sheath Hardness in Fragrant Rice» авторов Zhuang J, Fang Y, Zheng J, Duan Y, Liu X, Mo Z., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык
Фото: freepik





















































































































