Пшеница ранних сроков созревания как высокоценное кормовое сырье с пребиотической активностью
Работа посвящена изучению динамики изменения состава биомассы зерновых культур в процессе созревания и выявлению оптимальной стадии, на которой зерновая масса обладает высокой кормовой ценностью. Исследован урожай озимой пшеницы сорта Адмирал, озимой многолетней пшеницы (трититригия) сорта Памяти Любимовой и пырея сизого сорта Сова по аминокислотному составу, содержанию белка, влаги, железа, фосфора, селена, цинка, крахмала и витамина Е.
Аннотация
Зерновые культуры, убранные в фазу твердой восковой спелости, показали содержание белка выше на 3–4%, а также повышенный уровень некоторых аминокислот и минеральных веществ, таких как железо и селен. Зерновая масса пшеницы твердой восковой спелости была исследована на предмет пребиотических свойств. В результате исследования установлено, что она увеличивает количество молочнокислых бактерий в микробиоте кишечника и, следовательно, является перспективным пребиотиком для сельского хозяйства. Основываясь на данном исследовании, рекомендуемая концентрация зерновой массы пшеницы восковой спелости в качестве кормовой добавки составляет 1%.
1. Введение
Зерновые культуры играют важную роль в кормовой промышленности. Они являются источником углеводов; в некоторых рецептурах кормов их доля составляет более 50% от всех компонентов. Благодаря своим свойствам [1] пшеница используется в рецептурах кормов чаще других: белок пшеницы обладает высокой перевариваемостью по сравнению с другими зерновыми культурами (ячменем, рожью и овсом); клейковина, содержащаяся в пшенице, выступает не только источником белка, но и связующим компонентом, что особенно важно при производстве гранулированных кормов для рыб; экструзия пшеницы приводит к гидролизу крахмала, в результате чего образуются сахара и декстрины, которые легко усваиваются организмом животного; кроме того, экструзия улучшает органолептические свойства продукта. По словам Генерального директора ФАО д-ра Цюй Дунъюя [2], 40% пахотных земель используется для кормления скота. Это повышает устойчивость агропромышленного комплекса за счет оптимизации землепользования. Таким образом, совершенствование технологий возделывания сельскохозяйственных культур и использование новых культур будут способствовать устойчивому развитию агропромышленного комплекса в мире.
В качестве фуража используется низкобелковое зерно пшеницы. Предыдущие исследования ученых из разных стран [3,4,5,6,7,8,9,10,11,12,13,14,15] показали, что зерно пшеницы на ранних стадиях спелости (от ранней молочной до конца восковой спелости) содержит более высокий уровень белка и клейковины — в среднем на 3–4%. Эти данные позволяют утверждать, что использование зерна до наступления полной спелости для производства комбикормов является целесообразным.
В работе [3] авторы представляют результаты своего исследования по динамике изменения натуры зерна, стекловидности, содержания белка и клейковины у сорта яровой мягкой пшеницы Баганская 85 (среднеспелый сорт). Авторы заключают, что уборка зерна пшеницы раньше или позже стадии конца восковой спелости ухудшает качество урожая. Проведя исследования в 2010 и 2011 годах, авторы определили, что, несмотря на изменения качества зерна в зависимости от метеорологических условий, самое высокое содержание питательных компонентов наблюдается на стадии конца восковой спелости.
В другом исследовании [4] ученые сравнили химический состав трех сортов полбы на стадии молочной спелости и стадии полной спелости. Они исследовали полбу на содержание минеральных веществ (микроэлементов (Cu, Zn, Mn, Fe, Na, B) и макроэлементов (N, P, K, Mg, Ca)), заменимых и незаменимых аминокислот, а также жирнокислотный состав. Результаты исследования показали, что на стадии молочной спелости содержание полезных питательных веществ было выше, чем на стадии полной спелости. Изучая полбу на стадиях молочной и полной спелости, Berihuete-Azorín и др. [5] выявили, что на стадии молочной спелости зерно полбы имеет более высокое содержание белка и более низкое содержание углеводов, чем на стадии полной спелости. Исследования показали, что на стадии молочной спелости зерно полбы имеет более высокое содержание белка и более низкое содержание углеводов, чем на стадии полной спелости.
В результате изменений в составе зерна пшеницы в процессе созревания Özkaya и др. [6] сделали следующие выводы: накопление белка и функциональных соединений (белков, клетчатки, фруктанов, фенольных веществ и антиоксидантов) происходит до стадии молочной спелости, после чего они расходуются на рост и созревание зерна и пропорционально уменьшаются к стадии полной спелости.
Результаты исследований не позволяют сделать надежных выводов о динамике изменения состава зерна в процессе созревания, а также не могут выявить оптимальную стадию, на которой происходит самое высокое содержание питательных веществ.
− В одних исследованиях указываются более высокие показатели качества зерна на стадии молочной спелости, в то время как в других исследованиях приводятся более высокие показатели качества на стадии конца восковой спелости;
− В некоторых исследованиях анализировались только две стадии — ранняя (молочная) и полная спелость. Это довольно широкий диапазон исследований, который не в полной мере отражает изменение зерна на ранних стадиях: фаза начала молочной спелости, фаза конца молочной спелости, стадия начала восковой спелости, а также середина и конец восковой спелости. Более детальное изучение позволит точнее определить стадию, на которой зерно имеет более высокое качество, и получить максимальную выгоду от его использования;
− Исследования проводились в различные годы и в различных регионах, и в них использовались разные шкалы для обозначения фаз спелости. Большое количество авторов использовали шкалу BBCH для анализа изменений состава зерна в процессе созревания [7,8,9,10,11,12,13,14,15]. Использование этой шкалы в настоящем исследовании позволит унифицировать данные об изменениях состава зерна в процессе созревания и обеспечит более четкую общую картину.
Помимо зерна, дополнительным источником кормового сырья может служить незерновая часть урожая — полова. Ранее этот вид кормового сырья активно использовался в комбикормовой промышленности. Полова была вторичным сырьем, которое получали после очистки зернового вороха на стационарных зерноочистительных пунктах. В настоящее время очистка скошенной зерновой массы осуществляется непосредственно в зерноуборочных комбайнах, и незерновая часть остается в поле. Использование зерна, предназначенного для кормовых целей, вместе с половой без ее отделения является целесообразным. Это позволит увеличить количество заготавливаемого кормового сырья и снизить себестоимость за счет исключения процесса очистки зерна. Ранее ученые [3,4,5,6,7,8,9,10,11,12,13,14,15] изучали динамику изменения зерна. Исследований по изменению показателей качества зернового вороха (зерна и половы) не проводилось.
Многолетние зерновые культуры (например, озимая многолетняя пшеница (трититригия) сорта Памяти Любимовой и пырей сизый сорта Сова) являются перспективным кормовым и пищевым сырьем, так как они засухоустойчивы, устойчивы ко многим заболеваниям и положительно влияют на состояние почвы, повышая ее плодородие. Более подробная информация о многолетних культурах представлена в предыдущих исследованиях [16]. Исследование изменения качества этих культур в процессе созревания также ранее не проводилось.
Помимо питательной ценности, такое кормовое сырье может обладать пребиотическими свойствами. Сельскохозяйственное сырье растительного происхождения содержит полисахариды, которые могут быть природными пребиотиками [17]. Согласно данным [18,19,20,21], пребиотики должны быть устойчивы к перевариванию в верхних отделах желудочно-кишечного тракта, стимулировать рост полезных бактерий и оказывать положительное влияние на здоровье потребителя. Эфирные масла и лекарственные растения [22,23], грибы [24], водоросли [25] и др. чаще всего рассматриваются как источник пребиотиков. Мы исследовали зерновой ворох пшеницы ранней восковой спелости на предмет пребиотических свойств. Мы обнаружили, что он может быть доступным и относительно дешевым сырьем для производства пребиотических препаратов.
Основываясь на вышеизложенном, целью данного исследования является выявление оптимальной стадии созревания зерна, на которой наблюдается самое высокое содержание полезных питательных веществ, и проведение анализа на наличие пребиотических свойств у такого потенциально ценного кормового сырья.
2. Материалы и методы
2.1. Посев и уборка зерновых культур
Для изучения изменения состава зерна в процессе созревания были взяты три следующих вида зерна: озимая пшеница сорта Адмирал (Triticum aestivum), озимая многолетняя пшеница (трититригия) сорта Памяти Любимовой (Trititrigia cziczinii Tzvel.) и пырей сизый сорта Сова (Thinopyrum intermedium). Озимая пшеница — классический вид зерновой культуры, используемый повсеместно. Выбор сорта пшеницы Адмирал обусловлен его распространенностью на Юге России.
Посев исследуемых культур проводился на полях учебно-опытного участка Донского государственного технического университета в поселке Рассвет Ростовской области Российской Федерации. Опытные участки представлены черноземом обыкновенным карбонатным тяжелосуглинистым.
Анализ почвы проводился по следующим параметрам:
- Содержание гумуса (%) определяли по методу И.В. Тюрина. Метод основан на окислении органического вещества хромовой кислотой до образования диоксида углерода;
- Общий азот (мг/кг) определяли ионометрическим методом. Метод включает извлечение нитратов из почвы 1% раствором алюмокалиевых квасцов и измерение активности нитрат-ионов с помощью ионоселективного электрода;
- Для определения подвижного фосфора и калия (мг/кг) использовали метод Кирсанова для определения подвижных соединений фосфора и калия. Метод основан на извлечении подвижных соединений фосфора и калия из почвы раствором соляной кислоты молярной концентрации 0,2 моль/дм3 с последующим определением фосфора на фотоэлектроколориметре и калия на пламенном фотометре;
- pH солевой вытяжки. Метод основан на получении водной вытяжки путем извлечения из почвы катионов, нитратов и подвижной серы раствором хлористого калия и потенциометрическом определении pH с помощью стеклянного электрода.
В течение всего вегетационного периода были проведены три подкормки почвы: при посеве семян использовали аммофос в количестве 100 кг/га (12 кг N/га и 52 кг Р/га); в фазе кущения и в фазе выхода в трубку использовали аммиачную селитру в количестве 70 кг/га (24 кг N/га). Уборка урожая проводилась в 5 этапов (фазы роста представлены по шкале BBCH [7,8,9,10,11,12,13,14,15]): 77 — конец молочной спелости (Рисунок 1), 83 — начало восковой спелости, 87 — твердая восковая спелость, 89 — полная спелость, 92 — перестой. Выбор этих стадий был основан на предыдущих исследованиях [3,4,5,6,7,8,9,10,11,12,13,14,15], где ученые рассматривали динамику изменения состава зерна от молочной спелости до полной спелости и перестоя на корню. Выбор шкалы BBCH обусловлен ее широкой популярностью во всем мире; она используется как в научной сфере, так и на практике в области сельского хозяйства и садоводства, а также в фенологии как комплексной науке об окружающей среде, метеорологии и климатологии [15]. Уборка зерновых культур на ранних стадиях спелости не может быть выполнена классическим зерноуборочным комбайном. Для этой цели использовалась новая очесывающе-молотильная установка для уборки зерновых культур в различных фазах спелости [26,27].

Рисунок 1. Уборка зерновых культур на ранних стадиях спелости: (a) очесывающе-молотильная установка для уборки зерновых культур; (b) зерновой ворох пшеницы на ранних стадиях спелости.
В результате уборки был получен ворох зерна пшеницы (Рисунок 1b), который измельчали и подвергали физико-химическому анализу.
2.2. Физико-химический анализ убранного зернового вороха
Для получения однородной массы для проведения физико-химического анализа полученные пробы зернового вороха тщательно перемешивали и измельчали на лабораторной мельнице LZM-M2 (Лабораторофф, Воронеж, Россия). Полученные пробы были протестированы на следующие показатели качества:
- Массовую долю влаги (%) определяли гравиметрическим методом (высушиванием). Сущность метода заключается в высушивании навески пробы до постоянной массы при температуре 105 °C;
- Массовую долю белка (%) определяли методом Кьельдаля. Содержание белка рассчитывали на основе общего содержания азота с использованием коэффициента пересчета азота в белок, равного 6,25;
- Массовую долю фосфора (%) определяли фотометрическим методом. Сущность метода заключается в минерализации пробы мокрым или сухим озолением с образованием солей ортофосфорной кислоты и последующем фотометрическом определении фосфора в виде желтого окрашенного соединения — гетерополикислоты, образующейся в кислой среде в присутствии ванадатных и молибдатных ионов;
- Массовую долю протеиногенных аминокислот (лизина, аргинина, тирозина, фенилаланина, гистидина, лейцина, изолейцина, метионина, валина, пролина, треонина, серина, аланина, цистина, аспарагиновой кислоты, глутаминовой кислоты и триптофана) (%) определяли методом капиллярного электрофореза. Сущность метода заключается в разложении пробы для анализа путем кислотного (для всех аминокислот, кроме триптофана) или щелочного (для триптофана) гидролиза с переводом аминокислот в свободные формы, получении фенилтиокарбамильных производных аминокислот, их дальнейшем разделении и количественном определении методом капиллярного электрофореза;
- Содержание железа и цинка (мг/кг) определяли атомно-абсорбционным методом. Метод основан на минерализации продукта сухим или мокрым озолением и определении концентрации элемента в растворе минерализата методом пламенной атомно-абсорбционной спектрометрии;
- Содержание селена (мг/кг) определяли флуориметрическим методом. Сущность метода заключается в минерализации анализируемой пробы, переводе селена из органических и неорганических форм в селенит-ион, реакции селенит-иона с реагентом 2,3-диаминонафталином в кислой среде с образованием 4,5-бензопиазоселенола, его экстракции гексаном и измерении интенсивности флуоресценции полученного экстракта.
- Массовую долю крахмала (%) определяли методом Бертрана. Сущность метода заключается в способности редуцирующих сахаров восстанавливать двухвалентную медь в щелочной среде до оксида меди (I), который окисляется железоаммонийными квасцами с последующим титрованием восстановленного двухвалентного железа раствором перманганата калия;
- Содержание витамина Е (мг/кг) определяли методом высокоэффективной жидкостной хроматографии с флуориметрическим детектированием. Метод основан на выделении полициклических ароматических углеводородов из анализируемой пробы с их последующим количественным определением методом высокоэффективной жидкостной хроматографии с флуориметрическим детектированием.
2.3. Исследование зернового вороха пшеницы ранних стадий спелости на пребиотическую активность
После физико-химического анализа образцы зернового вороха пшеницы различных стадий спелости были заморожены для проведения исследований пребиотической активности.
Влияние зернового вороха пшеницы ранних стадий спелости на микробиоту птиц изучали в искусственной кишечной среде. Искусственная кишечная среда была ранее разработана нашей командой [22,28]. Состав среды на 1 л следующий: соевый белок (Kompanioncity, Санкт-Петербург, Россия) — 20 г; водорастворимый крахмал (Химреактивснаб, Уфа, Россия) — 0,5 г; нерафинированное подсолнечное масло (Юг Руси, Ростов-на-Дону, Россия) — 30 мл; Твин 80 (Химфармпродукт, Санкт-Петербург, Россия) — 1,5 мл; MgSO4 — 0,5 г; NaCl — 5 г; K2HPO4 — 0,5 г; MnSO4 — 0,05 г; FeSO4 — 0,05 г (все соли: Химфармпродукт, Санкт-Петербург, Россия).
Метод приготовления включает нагревание воды и растительного масла в сушильном шкафу до 80 °C. Затем масло выливают в колбу, где будет готовиться среда, добавляют Твин 80 и смесь энергично встряхивают. Затем постепенно (по 5–10 мл) добавляют горячую воду при непрерывном встряхивании. Следующую порцию воды добавляют только после гомогенизации суспензии. Количество добавляемой воды каждый раз затем увеличивают до 30–50 мл. После этого добавляют остальные компоненты и смесь автоклавируют. Этот метод приготовления обеспечивает сохранение мицеллярной структуры масла после автоклавирования.
Жидкую искусственную кишечную среду разливали в стерильные колбы объемом 250 мл по 100 мл для обеспечения минимального контакта с воздухом и высоты столба жидкости. В каждую из колб, за исключением контроля, добавляли зерновой ворох пшеницы ранних стадий спелости в концентрациях 0,1%, 0,25%, 0,5%, 0,75%, 1%, 2% и 5% (в зависимости от варианта эксперимента). Затем в колбы вносили кишечные бактерии в виде суспензии, приготовленной из стартера. Стартер включал содержимое слепой кишки нескольких птиц, не получавших про- и пребиотики, а также антибиотики, объемом около 1 см3, хранившееся при температуре -80 °C. Для каждого эксперимента использовали слепую кишку новой птицы. Содержимое разбавляли 10 мл хлорида натрия в концентрации 0,9% и вносили в колбы порциями по 1 мл. Затем колбы инкубировали при 42 °C в течение 3 дней, и данные о росте микробиоты регистрировали на 3-й день. Также измеряли уровень pH.
Количество кишечных бактерий определяли методом глубинного посева бактериальной суспензии в количестве 3 повторностей для каждой питательной среды для каждого исследуемого разведения. Количество бифидобактерий изучали путем посева суспензии в полужидкую питательную среду.
Для определения количества бактерий Lactobacillus использовали среду MRS (ЛенРеактив), Bifidobacterium — среду для выделения бифидобактерий (HiMedia), Enterococcus — среду для выделения энтерококков (HiMedia), E. coli — среду Эндо (HiMedia), Staphylococcus — среду № 10 (ЛенРеактив). Подсчет микроорганизмов проводили на второй день.
Для того чтобы оценить влияние пшеницы молочной спелости на микробиоту других птиц, была изучена модель микробиоты слепой кишки перепела. Для определения влияния зернового вороха пшеницы ранней спелости на лактобациллы в искусственную кишечную питательную среду перепела добавляли зерновой ворох пшеницы ранней спелости с фактической влажностью в концентрациях 0,5%, 0,75% и 1%.
С целью проверки, изменяется ли активность пшеницы молочной спелости после сушки в печи SSh-200 Bossert (Боссерт, Москва, Россия), пшеницу в количествах 1 г, 0,75 г и 0,5 г (что соответствует концентрациям 1%, 0,75% и 0,5%) высушивали в сухожаровом шкафу при температуре 80 °C. Также была приготовлена среда MRS. Затем добавляли суточную культуру Limosilactobacillus frumenti KL31 и инкубировали в течение 2 дней при температуре 42 °C. После этого определяли уровень pH.
2.4. Статистический анализ
Каждый образец анализировали в течение 24 ч после уборки. Каждый образец анализировали в трех повторностях. За окончательный результат принимали среднеарифметическое трех повторных определений. Расхождение между параллельными измерениями не превышало значений, характерных для каждого метода. Полученные результаты были статистически проанализированы с использованием однофакторного дисперсионного анализа (ANOVA), а наименьшие существенные разницы были рассчитаны с использованием критерия Тьюки при уровне ошибки 5%.
Корреляция между свойствами различных штаммов была рассчитана с использованием коэффициента корреляции Пирсона.
Для расчетов использовали программное обеспечение Statistica 10.0 и Microsoft Excel 2016.
3. Результаты
3.1. Результаты исследования изменения состава массы зерновых культур в процессе созревания
Результаты исследования аминокислотного состава массы зерновых культур представлены на Рисунках 2, 3 и 4.
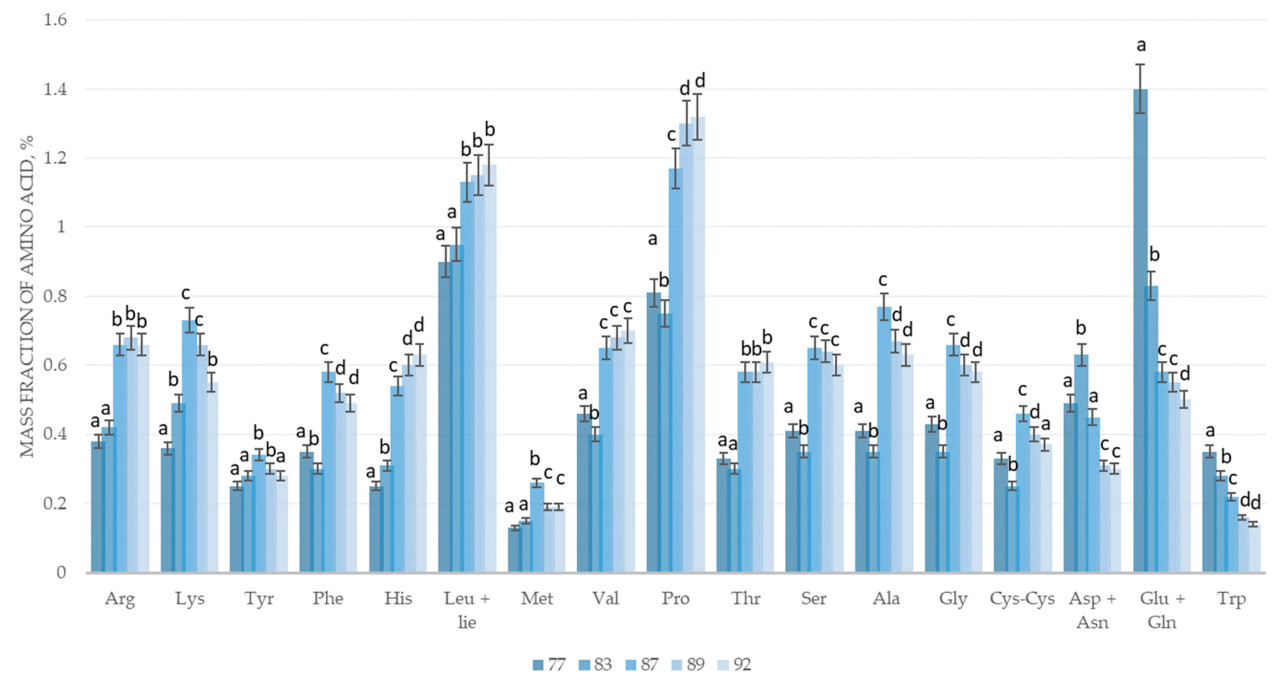
Рисунок 2. Изменение аминокислотного состава зернового вороха пшеницы сорта Адмирал в процессе созревания: 77 — конец молочной спелости, 83 — начало восковой спелости, 87 — твердая восковая спелость, 89 — полная спелость, 92 — перестой. Arg — аргинин, Lys — лизин, Tyr — тирозин, Phe — фенилаланин, His — гистидин, Leu+Ile — лейцин + изолейцин, Met — метионин, Val — валин, Pro — пролин, Thr — треонин, Ser — серин, Ala — аланин, Gly — глицин, Cys-Cys — цистеин, Asp+Asn — аспарагин + аспарагиновая кислота, Glu+Gln — глутамин + глутаминовая кислота, Trp — триптофан; a–d — различные буквы указывают на значительные различия между фазами созревания (p < 0,05).
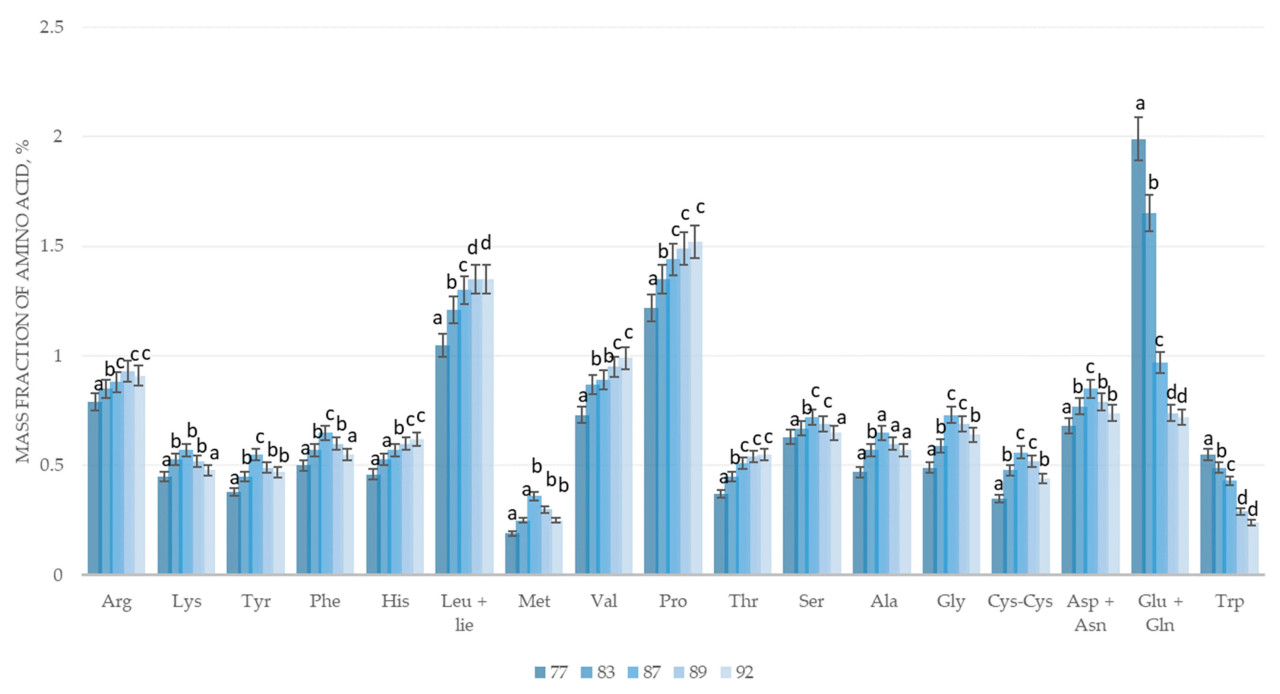
Рисунок 3. Изменение аминокислотного состава зернового вороха озимой многолетней пшеницы (трититригия) сорта Памяти Любимовой в процессе созревания: 77 — конец молочной спелости, 83 — начало восковой спелости, 87 — твердая восковая спелость, 89 — полная спелость, 92 — перестой. Arg — аргинин, Lys — лизин, Tyr — тирозин, Phe — фенилаланин, His — гистидин, Leu+Ile — лейцин + изолейцин, Met — метионин, Val — валин, Pro — пролин, Thr — треонин, Ser — серин, Ala — аланин, Gly — глицин, Cys-Cys — цистеин, Asp+Asn — аспарагин + аспарагиновая кислота, Glu+Gln — глутамин + глутаминовая кислота, Trp — триптофан; a–d — различные буквы указывают на значительные различия между фазами созревания (p < 0,05).
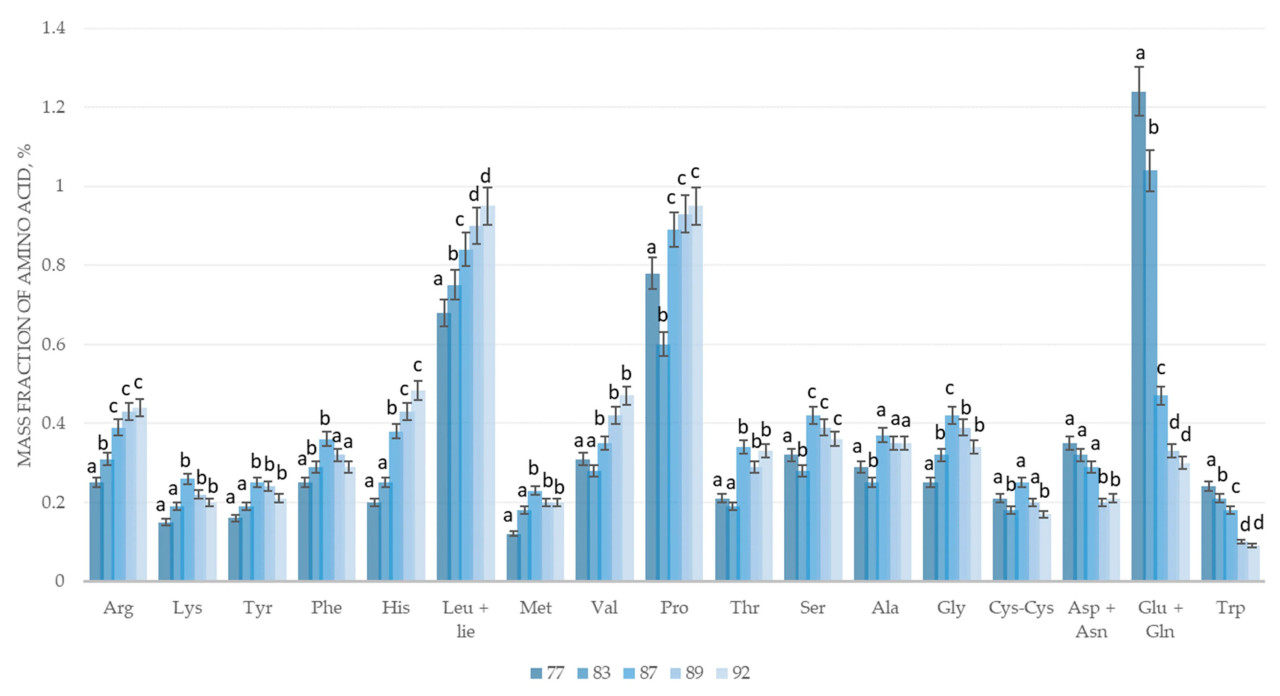
Рисунок 4. Изменение аминокислотного состава зернового вороха пырея сизого сорта Сова в процессе созревания: 77 — конец молочной спелости, 83 — начало восковой спелости, 87 — твердая восковая спелость, 89 — полная спелость, 92 — перестой. Arg — аргинин, Lys — лизин, Tyr — тирозин, Phe — фенилаланин, His — гистидин, Leu+Ile — лейцин + изолейцин, Met — метионин, Val — валин, Pro — пролин, Thr — треонин, Ser — серин, Ala — аланин, Gly — глицин, Cys-Cys — цистеин, Asp+Asn — аспарагин + аспарагиновая кислота, Glu+Gln — глутамин + глутаминовая кислота, Trp — триптофан; a–d — различные буквы указывают на значительные различия между фазами созревания (p < 0,05).
Результаты исследования показывают, что стадия молочной спелости (стадия 77 по шкале BBCH) имеет самое высокое содержание глутаминовой кислоты, глутамина и триптофана, а при переходе к восковой спелости происходит резкое снижение, которое продолжается и после полной спелости.
Содержание аспарагиновой кислоты и аспарагина в фазе начала восковой спелости (83 по шкале BBCH) достигает своих пиковых значений, после чего, как и в случае с глутаминовой кислотой, глутамином и триптофаном, они снижаются до и после полной спелости.
Наибольшее количество аминокислот лизина, тирозина, фенилаланина, метионина, аланина, глицина и цистина в зерновом ворохе пшеницы сорта Адмирал наблюдается на стадии твердой восковой спелости (стадия 87 по шкале BBCH), после чего происходит плавное снижение до и после полной спелости. Содержание гистидина, лейцина, изолейцина, валина, пролина, треонина и серина увеличивается на протяжении всего вегетационного периода.
Динамика изменения аминокислотного состава зернового вороха озимой многолетней пшеницы (трититригия) сорта Памяти Любимовой аналогична динамике озимой пшеницы сорта Адмирал. Аминокислотный состав многолетней пшеницы выше, чем у озимой пшеницы сорта Адмирал, в среднем в 1,5–2,0 раза. Кроме того, накопление аминокислот у многолетней пшеницы происходит более плавно, чем у однолетней.
По содержанию и динамике изменения аминокислот пырей сизый сорта Сова сходен с озимой пшеницей сорта Адмирал. Пырей сизый сорта Сова, по сравнению с двумя предыдущими проанализированными культурами, имеет более низкое содержание аминокислот. Он содержит низкое количество лизина и тирозина — более чем в два раза. Динамика изменения аминокислотного состава пырея сизого сорта Сова сходна с предыдущими культурами, озимой пшеницей сорта Адмирал и озимой многолетней пшеницей (трититригия) сорта Памяти Любимовой, за исключением аспарагиновой кислоты и аспарагина. Их количество достигает пиковых значений на стадии конца молочной спелости и продолжает снижаться до полной спелости. В Таблицах 1–8 представлены результаты исследования содержания белка, влаги, железа, фосфора, селена, цинка, крахмала и витамина Е в массе зерновых культур.
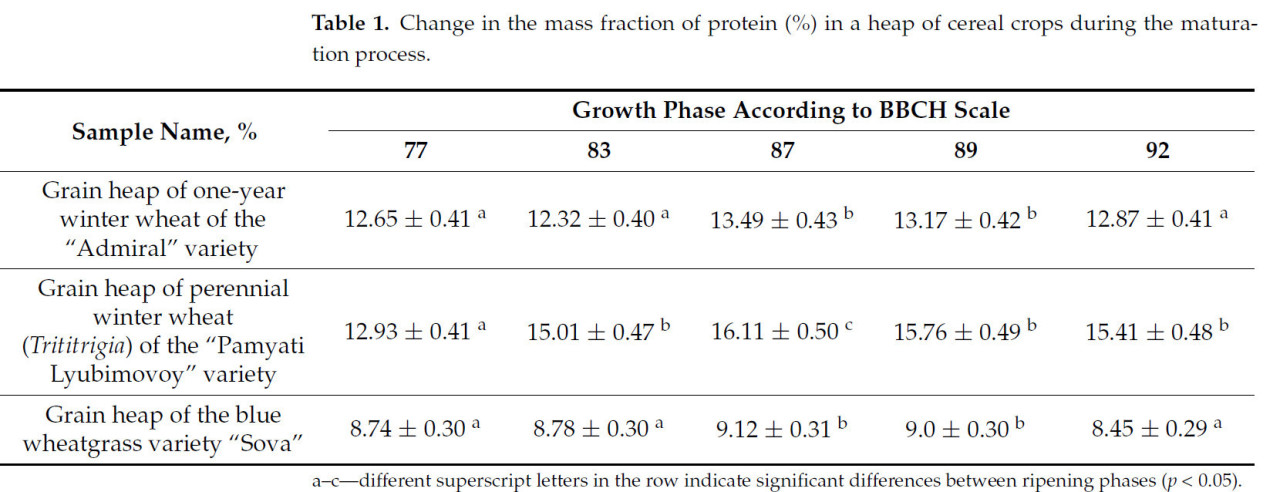
Таблица 1. Изменение массовой доли белка (%) в массе зерновых культур в процессе созревания.
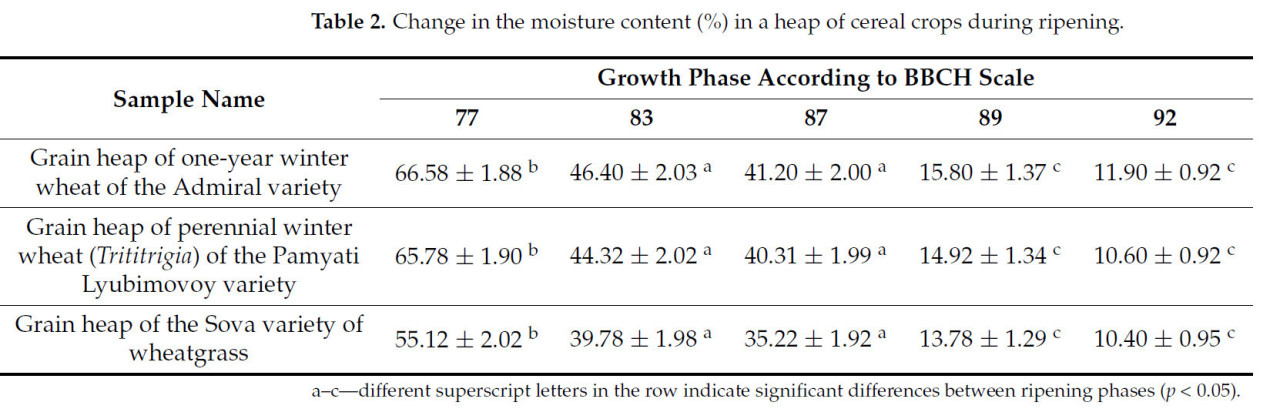
Таблица 2. Изменение влажности (%) в массе зерновых культур в процессе созревания.
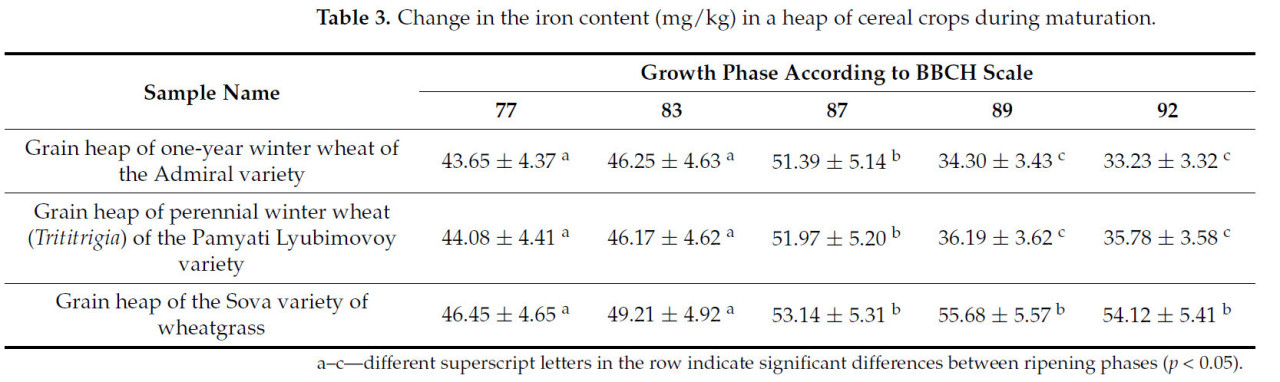
Таблица 3. Изменение содержания железа (мг/кг) в массе зерновых культур в процессе созревания.
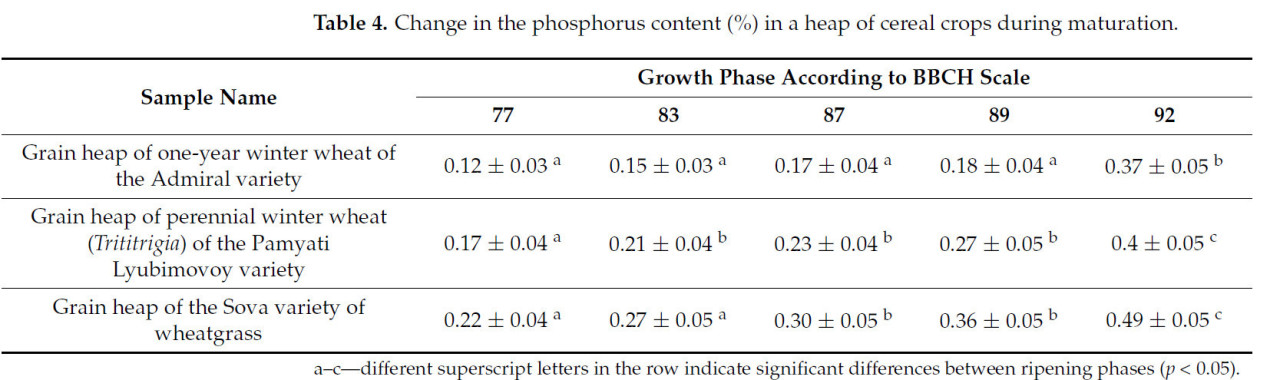
Таблица 4. Изменение содержания фосфора (%) в массе зерновых культур в процессе созревания.
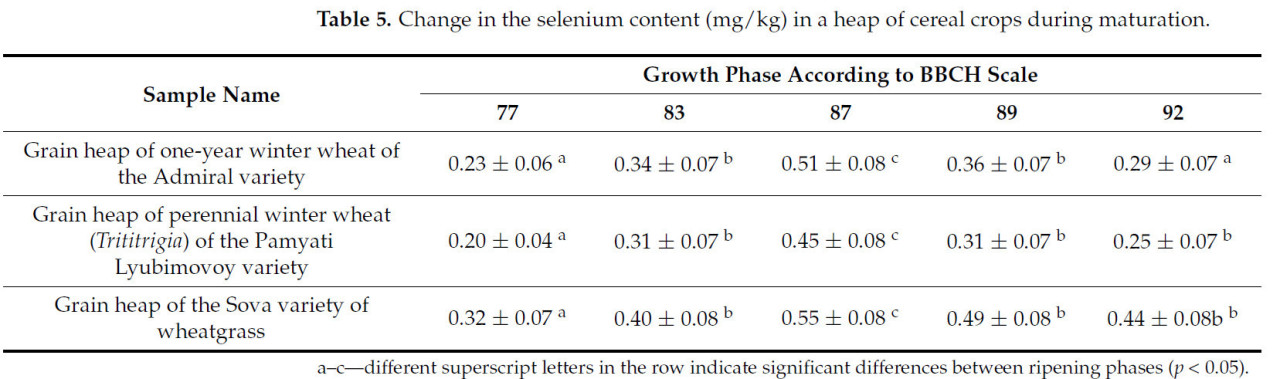
Таблица 5. Изменение содержания селена (мг/кг) в массе зерновых культур в процессе созревания.
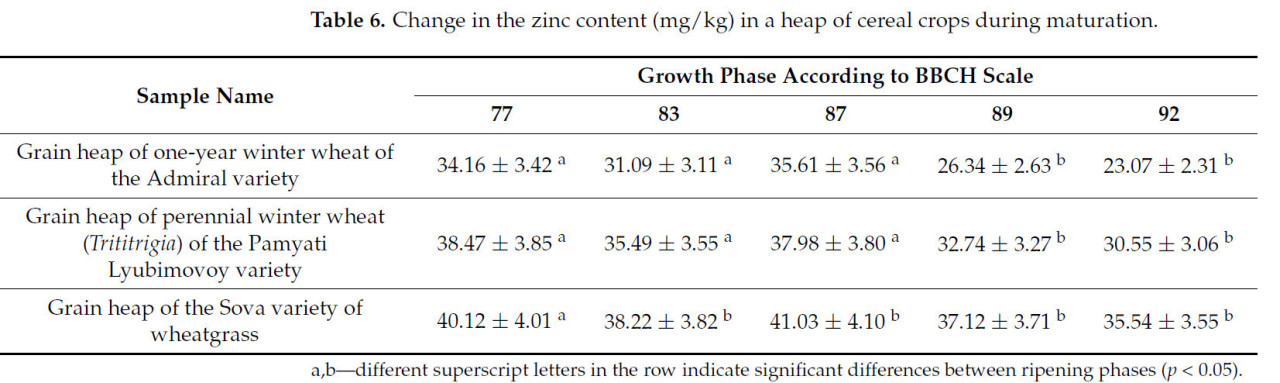
Таблица 6. Изменение содержания цинка (мг/кг) в массе зерновых культур в процессе созревания.
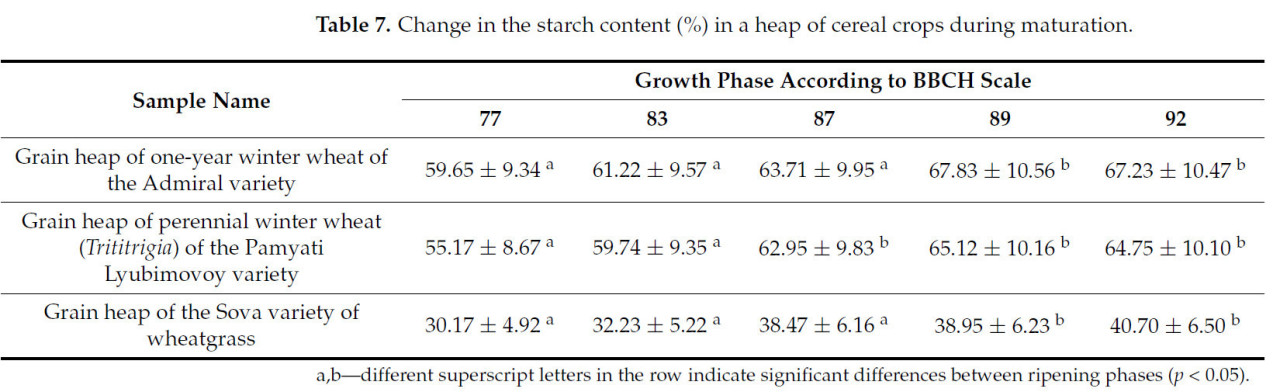
Таблица 7. Изменение содержания крахмала (%) в массе зерновых культур в процессе созревания.
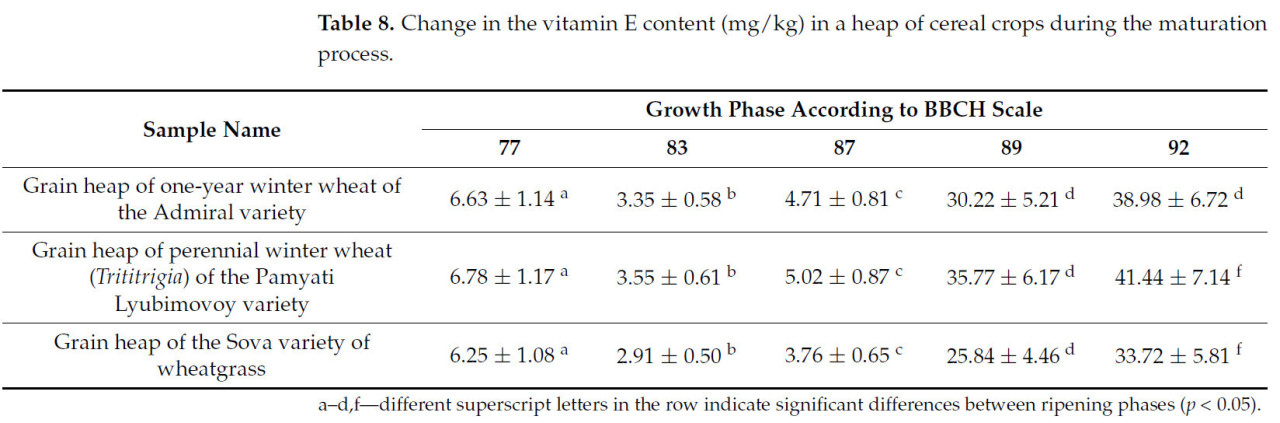
Таблица 8. Изменение содержания витамина Е (мг/кг) в массе зерновых культур в процессе созревания.
Таблица 1 показывает, что все три исследуемые культуры имеют более высокие пиковые значения белка на стадии твердой восковой спелости (стадия 87 по шкале BBCH), после чего они снижаются на 0,1–0,5%. Также после полной спелости содержание белка в зерновой массе снижается на 0,3–0,4%.
Динамика изменения влажности в массе зерновых культур демонстрирует плавное снижение к стадии восковой спелости и резкое снижение влажности перед наступлением полной спелости. Зерно озимой пшеницы сорта Адмирал крупнее, чем у двух других исследуемых культур, в результате чего зерновой ворох пшеницы сорта Адмирал имеет более высокую влажность.
Содержание железа плавно увеличивается до стадии твердой восковой спелости. Затем, достигнув пиковых значений в этой фазе 51–53 мг/кг, происходит резкое снижение в зерновом ворохе озимой пшеницы сорта Адмирал и озимой многолетней пшеницы (трититригия) сорта Памяти Любимовой. У этих культур содержание железа в фазе полной спелости ниже, чем на стадии конца молочной спелости. В зерновом ворохе пырея сизого сорта Сова железо накапливается вплоть до наступления полной спелости и лишь немного снижается после наступления полной спелости (стадия 92 по шкале BBCH).
Содержание фосфора медленно увеличивается на всех исследуемых стадиях. Резкое увеличение (почти в два раза) происходит после полной спелости и достигает 0,37–0,49%.
Содержание селена достигает пиковых значений на стадии твердой восковой спелости. После полной спелости содержание селена уменьшается почти в два раза в двух исследуемых образцах — зерновом ворохе озимой пшеницы сорта Адмирал и зерновом ворохе озимой многолетней пшеницы (трититригия) сорта Памяти Любимовой. В зерновом ворохе пырея сизого сорта Сова содержание селена практически не изменяется после полной спелости. Вероятно, это связано с наличием большого количества незерновой части в зерновом ворохе пырея сорта Сова, что характерно для этого вида культуры (пырей сизый сорта Сова имеет большое количество зеленой массы по сравнению с другими зерновыми культурами [21]).
Содержание цинка на ранних стадиях спелости (молочная и восковая 77–87) остается практически неизменным. После полной спелости количество немного уменьшается на 4–9 мг/кг.
Накопление крахмала в зерне происходит на протяжении всего вегетационного периода и достигает пиковых значений при полной спелости. Зерновой ворох пырея сизого сорта Сова содержит в 1,6 раза меньше крахмала, чем два других проанализированных образца.
Содержание витамина Е также увеличивается после полной спелости. Витамин Е является жирорастворимым и содержится в основном в зародыше, поэтому его количество резко возрастает после полной спелости, когда зародыш полностью сформирован (увеличение с 4,71 до 30,22 мг/кг). Изменение показателей качества зерна аналогично для всех трех образцов.
3.2. Результаты исследования пребиотической активности зернового вороха пшеницы ранних стадий спелости
Для изучения пребиотической активности использовали зерновой ворох пшеницы сорта Адмирал стадии твердой восковой спелости (87 по шкале BBCH), так как именно на этой стадии наблюдается самое высокое содержание всех изученных в данной работе питательных веществ.
3.2.1. Изучение влияния высоких концентраций пшеницы молочной спелости на модели микробиоты цыпленка
В качестве модельной среды использовали искусственную кишечную среду цыпленка. Использовали концентрации 5%, 2% и 1%, так как концентрации 2% были наиболее эффективны в наших предыдущих исследованиях со жмыхами эфиромасличных растений. Результаты представлены в Таблице 9.
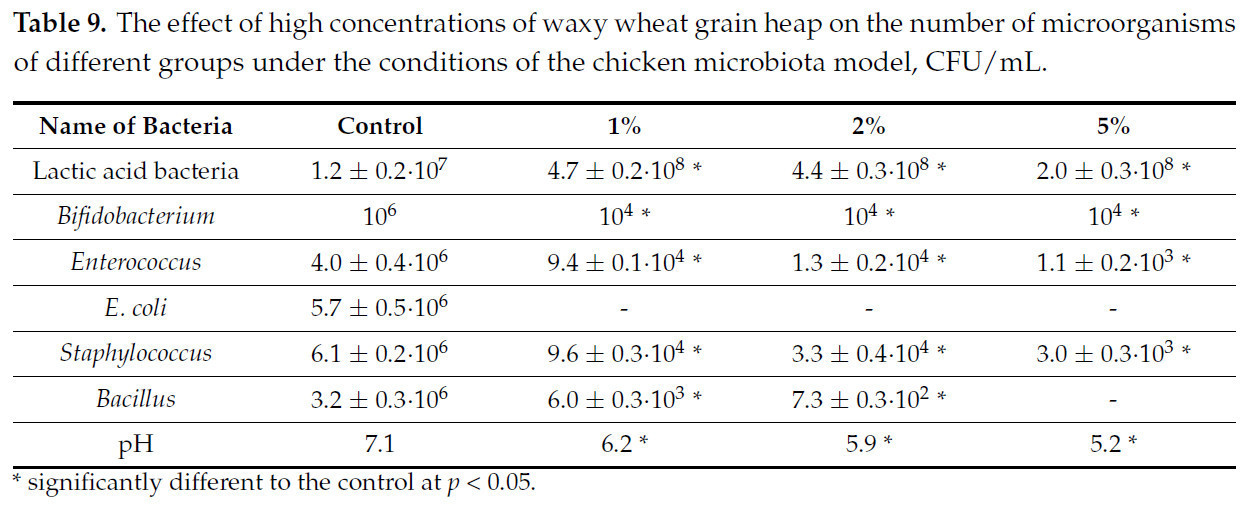
Таблица 9. Влияние высоких концентраций зернового вороха пшеницы восковой спелости на численность микроорганизмов различных групп в условиях модели микробиоты цыпленка, КОЕ/мл.
Представленные данные показывают, что зерновой ворох пшеницы восковой спелости оказал значительное влияние на микробиоту цыпленка. Количество молочнокислых бактерий увеличилось, в то время как уровень pH среды снизился. Количество условно-патогенных микроорганизмов снизилось, а количество E. coli упало до значений ниже порога чувствительности для данной методики. С другой стороны, концентрация бифидобактерий и бацилл также снизилась.
В целом, чрезмерно высокая концентрация зернового вороха пшеницы восковой спелости, несмотря на увеличение количества молочнокислых бактерий, оказывает отрицательное влияние на микробиоту, снижая разнообразие и полностью подавляя некоторые группы микроорганизмов. Тем не менее, такие сдвиги показывают высокий потенциал зернового вороха пшеницы восковой спелости в качестве пребиотика.
3.2.2. Изучение низких концентраций пшеницы молочной спелости на модели микробиоты цыпленка
В качестве модельной среды использовали искусственную кишечную среду цыпленка. Использовали концентрации 0,1%, 0,25% и 0,5%. Добавление сахара в концентрации 0,1% также использовалось в качестве положительного контроля. Необходимо было определить, основан ли наблюдаемый эффект на содержании пребиотических компонентов или только на содержании простых сахаров в составе зернового вороха пшеницы. Полученные результаты представлены в Таблице 10.
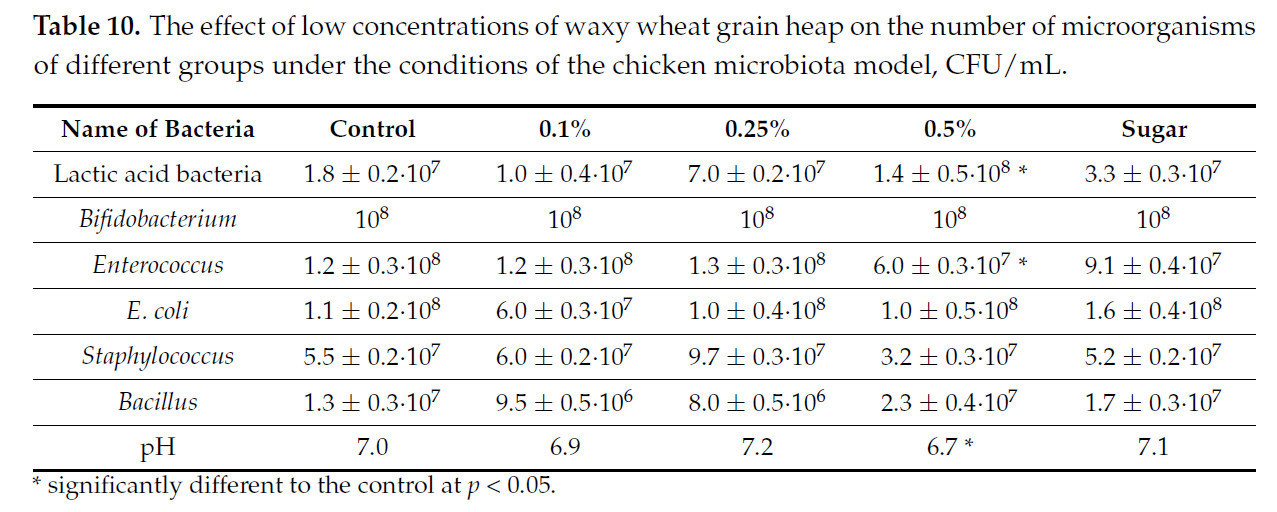
Таблица 10. Влияние низких концентраций зернового вороха пшеницы восковой спелости на численность микроорганизмов различных групп в условиях модели микробиоты цыпленка, КОЕ/мл.
Представленные данные показывают, что микробиота слабее реагировала на введение зернового вороха пшеницы восковой спелости, при этом концентрации 0,1% и 0,25% не оказали достоверного влияния на количество и соотношение групп микроорганизмов в микробиоте цыпленка. Концентрация 0,5% привела к увеличению количества лактобацилл и снижению количества энтерококков.
Сахар, даже в заведомо завышенной концентрации, не оказал достоверного влияния на микробиоту цыпленка, что означает, что наблюдаемый нами эффект не связан с наличием простых сахаров в пшенице.
3.2.3. Изучение средних концентраций пшеницы молочной спелости на модели микробиоты цыпленка
На данном этапе были оценены концентрации, которые показались наиболее перспективными после первых двух этапов: 0,5%, 0,75% и 1%. Результаты представлены в Таблице 11.
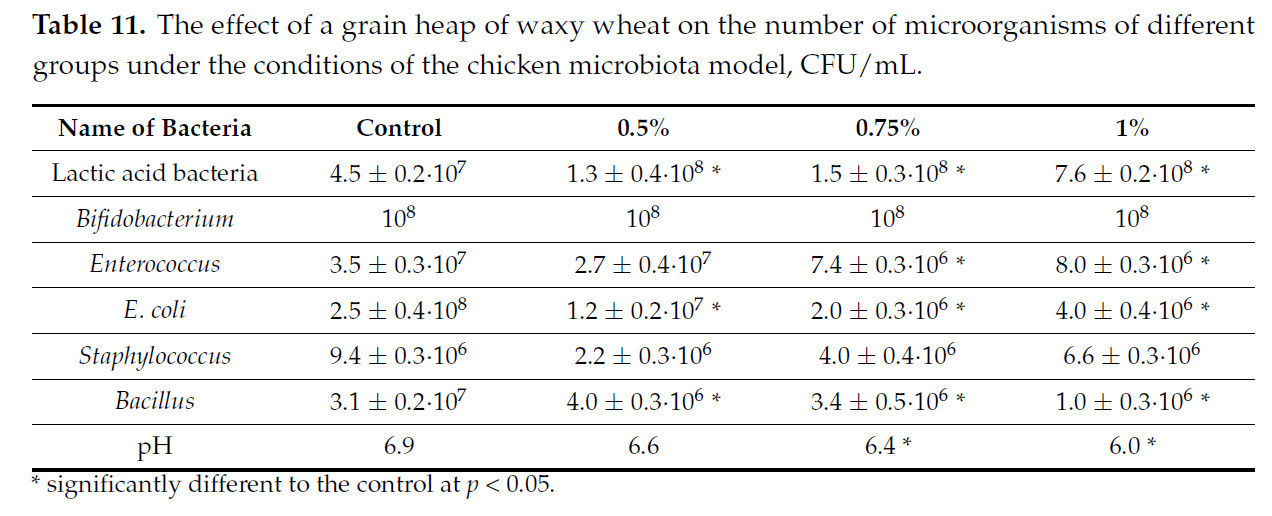
Таблица 11. Влияние зернового вороха пшеницы восковой спелости на численность микроорганизмов различных групп в условиях модели микробиоты цыпленка, КОЕ/мл.
Представленные данные показывают, что эффект от введения зернового вороха пшеницы восковой спелости слабее, чем в первом эксперименте, и сопоставим со вторым. Все три концентрации вызвали увеличение количества лактобацилл. При концентрации 0,5% отмечается снижение количества E. coli и бацилл; при концентрациях 0,75% и 1% количество E. coli, энтерококков и бацилл увеличилось.
3.2.4. Изучение зернового вороха пшеницы восковой спелости на модели микробиоты перепела
Для того чтобы оценить влияние зернового вороха пшеницы восковой спелости на микробиоту других птиц, была изучена модель микробиоты слепой кишки перепела. Пшеницу молочной спелости использовали в концентрациях 0,5%, 0,75% и 1%. Полученные данные представлены в Таблице 12.
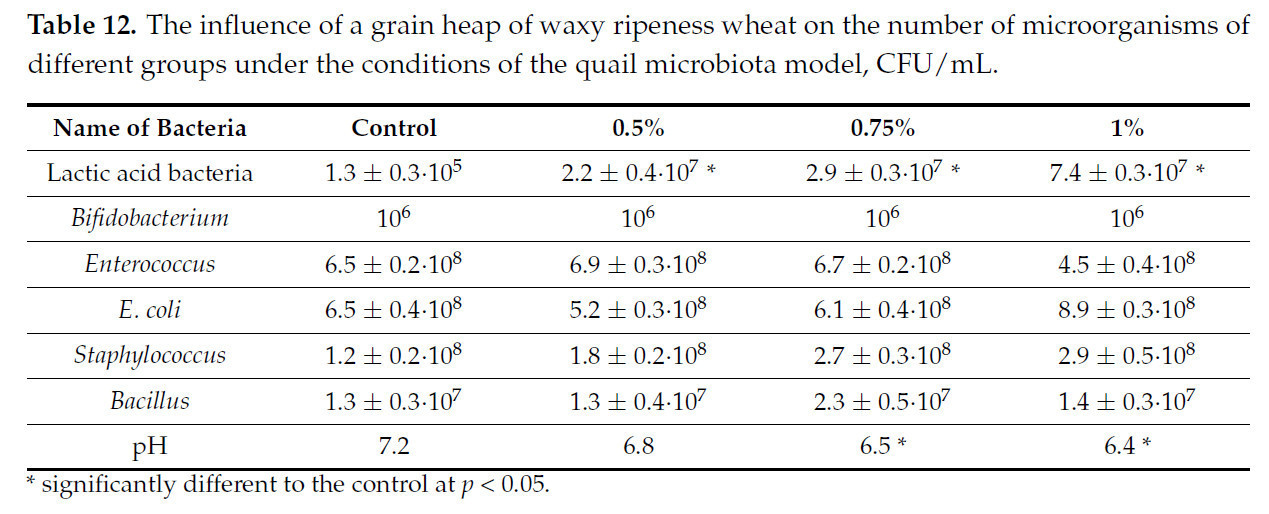
Таблица 12. Влияние зернового вороха пшеницы восковой спелости на численность микроорганизмов различных групп в условиях модели микробиоты перепела, КОЕ/мл.
Представленные данные показывают, что влияние зернового вороха пшеницы восковой спелости на микробиоту перепелов слабее, чем на микробиоту цыплят. Среди групп, представляющих условно-патогенные микроорганизмы, не отмечено достоверного изменения численности при введении зернового вороха пшеницы восковой спелости. Также не отмечено изменения количества бифидобактерий. В то же время количество лактобацилл и других молочнокислых бактерий значительно увеличилось, более чем на два порядка, что повторяет тенденцию, отмеченную в экспериментах на модельной среде цыпленка.
3.2.5. Изучение сухого зернового вороха пшеницы восковой спелости на численность микроорганизмов различных групп в условиях модели микробиоты перепела
С целью проверки, изменяется ли активность зернового вороха пшеницы восковой спелости после сушки, пшеницу в количествах 1 г, 0,75 г и 0,5 г (что соответствует концентрациям 1%, 0,75% и 0,5%) высушивали в сухожаровом шкафу при температуре 80 °C. Влажность пшеницы составляла 32%. Полученные результаты представлены в Таблице 13. Концентрации были пересчитаны на фактическую влажность зернового вороха пшеницы восковой спелости.
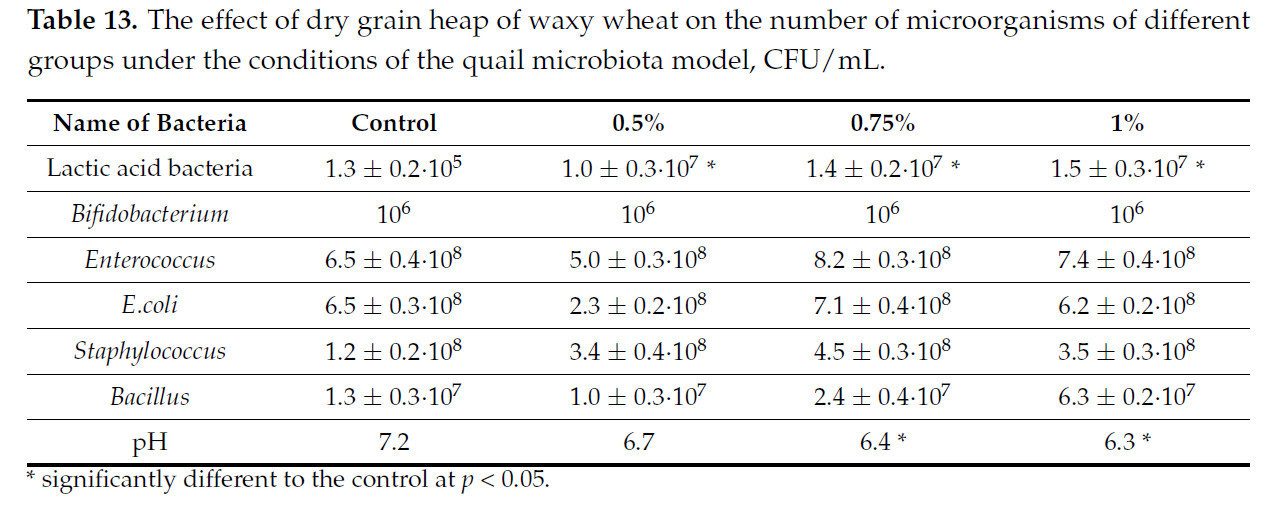
Таблица 13. Влияние сухого зернового вороха пшеницы восковой спелости на численность микроорганизмов различных групп в условиях модели микробиоты перепела, КОЕ/мл.
Как и в предыдущем случае, достоверных различий в количестве групп микроорганизмов от контрольных значений не отмечено, за исключением количества молочнокислых бактерий, которое увеличилось на два порядка.
3.2.6. Влияние зернового вороха пшеницы восковой спелости на лактобациллы
Данные о влиянии зернового вороха пшеницы восковой спелости на pH среды после инкубации с Limosilactobacillus frumenti KL31 представлены в Таблице 14.

Таблица 14. Значения pH сред с добавлением и без добавления зернового вороха пшеницы восковой спелости при инкубации Limosilactobacillus frumenti KL31 в искусственной кишечной среде (ИКС).
Можно отметить, что на питательной среде, богатой сахарами (MRS), наблюдается значительное снижение уровня pH во время инкубации Limosilactobacillus frumenti KL31. На искусственной кишечной среде, содержащей следовые количества простых сахаров, pH остается нейтральным. В то же время введение как влажного, так и сухого зернового вороха пшеницы восковой спелости приводит к увеличению продукции молочной кислоты Limosilactobacillus frumenti KL31. Это означает, что зерновой ворох пшеницы восковой спелости содержит углеводы, которые могут служить источником энергии для лактобацилл.
4. Обсуждение
Полученные данные подтвердили результаты предыдущих исследований [3,4,5,6,7,8,9,10,11,12,13,14,15]: содержание белка выше на ранних стадиях спелости, в среднем на 3–4%. Кроме того, были получены данные, подтверждающие отрицательное влияние увеличения сроков уборки на объем и качество урожая: помимо проблемы осыпания, которое является естественным процессом для сельскохозяйственных культур и приводит к значительным потерям, качество такого зерна также ухудшается. Следовательно, своевременная уборка зерновых культур позволит получить качественный урожай с минимальными потерями [27]. Именно на стадии твердой восковой спелости зерно накапливает максимальное количество белка, что согласуется с результатами исследований [3]. Уборка зерна на этой стадии позволит улучшить качество кормового сырья и повысить фактическую урожайность зерна за счет своевременной уборки и минимальных потерь от осыпания, что будет способствовать устойчивому развитию агропромышленного комплекса (в частности, комбикормовой промышленности) в мире [2]. С экономической точки зрения, снижаются затраты энергии на уборку методом очесывания на корню очесывающе-молотильной установкой [27], что, в свою очередь, положительно сказывается на себестоимости продукции; снижение потерь от осыпания также положительно влияет на себестоимость получаемого урожая.
Анализ изменения аминокислотного состава зерновых культур показывает сходную динамику: на стадии молочной спелости (стадия 77 по шкале BBCH) наблюдается максимальное содержание глутаминовой кислоты, глутамина и триптофана; содержание аспарагиновой кислоты и аспарагина на стадии начала восковой спелости (83 по шкале BBCH) достигает своих пиковых значений; наибольшее количество аминокислот, включая лизин, тирозин, фенилаланин, метионин, аланин, глицин и цистин, в массе зерновых культур наблюдается на стадии твердой восковой спелости (стадия 87 по шкале BBCH); содержание гистидина, лейцина, изолейцина, валина, пролина, треонина и серина увеличивается на протяжении всего вегетационного периода. Это согласуется с данными [4], где ученые изучали полбу на стадиях молочной и полной спелости. Они обнаружили, что в зерне на стадии молочной спелости содержание лизина, метионина, аланина и тирозина было выше, чем на стадии полной спелости. В наших исследованиях полученные данные были уточнены, в результате чего было выявлено, что эти аминокислоты продолжают увеличиваться до твердой восковой спелости (стадия 87 по шкале BBCH).
Лизин и метионин являются важнейшими незаменимыми аминокислотами, которые участвуют в процессах снижения окисления в организме животных, способствуя повышению продуктивности и укреплению иммунной системы [27,28]. Лимитирующие (критические) аминокислоты играют важную роль в питании животных и рыб, и их дефицит или избыток влияет на усвоение других аминокислот. Лимитирующие аминокислоты различаются у различных видов животных. Так, для птиц лимитирующими аминокислотами являются метионин и цистин; для свиней — лизин. Эти аминокислоты достигают пиковых значений при восковой спелости (стадия 87 по шкале BBCH). В растительном сырье триптофан, метионин и лизин чаще всего выступают в качестве лимитирующих аминокислот. Дефицит лимитирующих аминокислот в кормах может привести к нарушениям обмена веществ, снижению иммунитета, ожирению и большему потреблению корма [27,29]. Поэтому целесообразно использовать массу зерновых культур на стадии твердой восковой спелости, когда их количество на этой стадии максимально.
Содержание железа в зерновом ворохе озимой пшеницы сорта Адмирал и озимой многолетней пшеницы (трититригия) сорта Памяти Любимовой достигает пиковых значений на стадии твердой восковой спелости. В зерновом ворохе пырея сизого сорта Сова концентрация железа увеличивается к стадии полной спелости (стадия 89 по шкале BBCH), что согласуется с данными об изменении количества железа в полбе [4].
У двух из трех сортов полбы содержание фосфора выше на стадии молочной спелости [4]. Согласно нашим данным, содержание фосфора медленно увеличивается на всех изучаемых стадиях. Резкое увеличение (почти в два раза) происходит после полной спелости и достигает 0,37–0,49%.
Содержание цинка на ранних стадиях спелости (молочная и восковая стадии 77–87) остается практически неизменным. После полной спелости количество немного уменьшается на 4–9 мг/кг. Это согласуется с данными об изменении содержания цинка в полбе [4].
Таким образом, динамика изменений основных питательных веществ зерновых культур, таких как крахмал и белок, сходна как для пшеницы (однолетней и многолетней), так и для полбы [3,4,5,6,7,8,9,10,11,12,13,14,15], независимо от условий и зоны возделывания.
Динамика изменений минеральных веществ в разной степени варьирует, что может быть связано с погодными условиями, составом почвы и удобрениями [30,31,32,33]. Введение зернового вороха пшеницы восковой спелости стабильно приводило к увеличению количества молочнокислых бактерий. Количество условно-патогенных микроорганизмов снижалось, при этом количество E. coli падало до значений ниже порога чувствительности данного метода. С другой стороны, концентрация бифидобактерий и бацилл также снижалась. Эти результаты сходны с результатами [23], где Panax ginseng использовали в качестве пребиотика. Также, как и при использовании зернового вороха, наблюдается увеличение количества лактобацилл. Отмечается, что при использовании зернового вороха происходит снижение количества E. coli и бацилл, что согласуется с данными [22,23,24,25], где использование водорослей, лекарственных растений и грибов в качестве пребиотической кормовой добавки также подавляет патогенную микрофлору и позволяет восстановить микробиоту после воздействия негативных факторов.
5. Выводы
Масса зерновых культур на стадии твердой восковой спелости является ценным и экономически доступным новым кормовым сырьем. Благодаря повышенному содержанию ряда протеиногенных аминокислот, включая лизин, метионин и цистин — важных лимитирующих аминокислот, доля аминокислот, вводимых в виде кормовой добавки, сокращается, что будет способствовать снижению стоимости корма. Кроме того, такое сырье содержит больше железа и селена, чем полностью созревшее зерно. Отличительной особенностью данного кормового сырья является его высокая пребиотическая активность в отношении лактобацилл и других молочнокислых бактерий. Уборка зерна на этой стадии спелости позволяет проводить уборку в агротехнологические сроки и сокращает потери зерна от осыпания. Таким образом, уборка зерна до наступления полной спелости и его использование в комбикормовой промышленности соответствует принципам экономики замкнутого цикла, а также принципам устойчивого развития сельского хозяйства. В дальнейших исследованиях мы планируем провести серию экспериментов, направленных на разработку метода определения назначения зерна, не достигшего полной спелости (фуражное или продовольственное), в полевых условиях, а также провести экономическую оценку применения данного способа уборки и использования сырья при производстве комбикормов.
Ссылки
1. Ponomarev, S.V.; Grozescu, Y.N.; Bakhareva, A.A. Industrial Fish Farming: Textbook, 2nd ed.; revised. and additional; Publishing House “Lan”: St. Petersburg, Russia, 2022; 448p, Available online: https://e.lanbook.com/book/211118 (accessed on 27 November 2024). (In Russian)
2. FAO Global Forum for Animal Feed and Feed Regulators Opening Remarks by Dr QU Dongyu, FAO Director-General 14/11/2023. Available online: https://www.fao.org/director-general/speeches/details/FAO-Global-Forum-for-Animal-Feed-and-Feed-Regulators-Opening-Remarks/en (accessed on 13 January 2025).
3. Sandrykin, D.V.; Kondratenko, E.P.; Egushova, E.A.; Pinchuk, L.G. Dynamics of dry matter accumulation and changes in the chemical composition of grain during ripening. Achiev. Sci. Technol. Agro-Ind. Complex 2011, 12, 32–33. (In Russian) [Google Scholar]
4. Kraska, P.; Andruszczak, S.; Dziki, D.; Stocki, M.; Stocka, N.; Kwiecińska-Poppe, E.; Różyło, K.; Gierasimiuk, P. Green grain of spelt (Triticum aestivum ssp. spelta) harvested at the stage of milk-dough as a rich source of valuable nutrients. Emir. J. Food Agric. 2019, 31, 263–270. [Google Scholar] [CrossRef]
5. Berihuete-Azorín, M.; Stika, H.-P.; Hallama, M.; Valamoti, S.M. Distinguishing ripe spelt from processed green spelt (Grünkern) grains: Methodological aspects and the case of early La Tène Hochdorf (Vaihingen a.d. Enz, Germany). J. Archaeol. Sci. 2020, 118, 105143. [Google Scholar] [CrossRef]
6. Özkaya, B.; Turksoy, S.; Özkaya, H.; Baumgartner, B.; Özkeser, İ.; Köksel, H. Changes in the functional constituents and phytic acid contents of firiks produced from wheats at different maturation stages. Food Chem. 2018, 246, 150–155. [Google Scholar] [CrossRef] [PubMed]
7. Wollmer, A.C.; Pitann, B.; Mühling, K.H. Grain storage protein concentration and composition of winter wheat (Triticum aestivum L.) as affected by waterlogging events during stem elongation or ear emergence. J. Cereal Sci. 2018, 83, 9–15. [Google Scholar] [CrossRef]
8. Martínez-Núñez, M.; Ruiz-Rivas, M.; Vera-Hernández, P.F.; Bernal-Muñoz, R.; Luna-Suárez, S.; Rosas-Cárdenas, F.F. The phenological growth stages of different amaranth species grown in restricted spaces based in BBCH code. S. Afr. J. Bot. 2019, 124, 436–443. [Google Scholar] [CrossRef]
9. Brandán, J.P.; Curti, R.N.; Acreche, M.M. Phenological growth stages in chia (Salvia hispanica L.) according to the BBCH scale. Sci. Hortic. 2019, 255, 292–297. [Google Scholar] [CrossRef]
10. Hernández, D.P.M.; Aranguren, M.; Reig, C.; Galván, D.F.; Mesejo, C.; Fuentes, A.M.; Saúco, V.G.; Agustí, M. Phenological growth stages of mango (Mangifera indica L.) according to the BBCH scale. Sci. Hortic. 2011, 130, 536–540. [Google Scholar] [CrossRef]
11. Miedaner, T.; Haffke, S.; Siekmann, D.; Fromme, F.J.; Roux, S.R.; Hackauf, B. Dynamic quantitative trait loci (QTL) for plant height predict biomass yield in hybrid rye (Secale cereale L.). Biomass Bioenergy 2018, 115, 10–18. [Google Scholar] [CrossRef]
12. Cutignano, A.; Mamone, G.; Boscaino, F.; Ceriotti, A.; Maccaferri, M.; Picariello, G. Monitoring changes of lipid composition in durum wheat during grain development. J. Cereal Sci. 2021, 97, 103131. [Google Scholar] [CrossRef]
13. Li, G.; Ren, X.; Pang, S.; Feng, C.; Niu, Y.; Qu, Y.; Liu, C.; Lin, X.; Wang, D. Nitrogen redistribution during the grain-filling stage and its correlation with senescence and TaATG8 expression in leaves of winter wheat. J. Integr. Agric. 2024; In Press, Journal Pre-proof. [Google Scholar] [CrossRef]
14. Ma, J.; Wang, K.; Zheng, B.; Xu, Y.; He, Y. Early estimation of glutelin to gliadin ratio in wheat grain using high-dimensional and hyperspectral reflectance. Comput. Electron. Agric. 2024, 227, 109542. [Google Scholar] [CrossRef]
15. Meier, U.; Bleiholder, H.; Buhr, L.; Feller, C.; Hack, H.; Heß, M.; Lancashire, P.D.; Schnock, U.; Stauß, R.; van den Boom, T.; et al. The BBCH system to coding the phenological growth stages of plants-history and publications. J. Kult. 2009, 61, 41–52. [Google Scholar] [CrossRef]
16. Rudoy, D.; Pakhomov, V.; Olshevskaya, A.; Maltseva, T.; Ugrekhelidze, N.; Zhuravleva, A.; Babajanyan, A. Review and analysis of perennial cereal crops at different maturity stages. IOP Conf. Ser. Earth Environ. Sci. 2021, 937, 022111. [Google Scholar] [CrossRef]
17. Zhang, C.; Pi, X.; Li, X.; Huo, J.; Wang, W. Edible herbal source-derived polysaccharides as potential prebiotics: Composition, structure, gut microbiota regulation, and its related health effects. Food Chem. 2024, 458, 140267. [Google Scholar] [CrossRef]
18. Guerreiro, I.; Oliva-Teles, A.; Enes, P. Prebiotics as functional ingredients: Focus on Mediterranean fish aquaculture. Rev. Aquac. 2018, 10, 800–832. [Google Scholar] [CrossRef]
19. Khanjani, M.H.; Mozanzadeh, M.T.; Gisbert, E.; Hoseinifar, S.H. Probiotics, prebiotics, and synbiotics in shrimp aquaculture: Their effects on growth performance, immune responses, and gut microbiome. Aquac. Rep. 2024, 38, 102362. [Google Scholar] [CrossRef]
20. Fangueiro, J.F.; de Carvalho, N.M.; Antunes, F.; Mota, I.F.; Pintado, M.E.; Madureira, A.R.; Costa, P.S. Lignin from sugarcane bagasse as a prebiotic additive for poultry feed. Int. J. Biol. Macromol. 2023, 239, 124262. [Google Scholar] [CrossRef]
21. Serradell, A.; Torrecillas, S.; Soares, F.; Silva, T.; Montero, D. Modelling the effect of prebiotics, probiotics and other functional additives on the growth, feed intake and feed conversion of European sea bass (Dicentrarchus labrax) juveniles. Aquac. Rep. 2023, 32, 101729. [Google Scholar] [CrossRef]
22. Mazanko, M.; Prazdnova, E.; Statsenko, V.; Bren, A.; Rudoy, D.; Maltseva, T.; Chistyakov, V.; Chikindas, M. Oil Cakes of Essential Oil Plants as a Source of Prebiotics for Poultry Production. Agriculture 2023, 13, 591. [Google Scholar] [CrossRef]
23. Li, S.; Qi, Y.; Chen, L.; Qu, D.; Li, Z.; Gao, K.; Chen, J.; Sun, Y. Effects of Panax ginseng polysaccharides on the gut microbiota in mice with antibiotic-associated diarrhea. Int. J. Biol. Macromol. 2019, 124, 931–937. [Google Scholar] [CrossRef] [PubMed]
24. Liu, Y.; Li, Y.; Ke, Y.; Li, C.; Zhang, Z.; Wu, Y.; Hu, B.; Liu, A.; Luo, Q.; Wu, W. In vitro saliva-gastrointestinal digestion and fecal fermentation of Oudemansiella radicata olysaccharides reveal its digestion profile and effect on the modulation of the gut microbiota. Carbohydr. Polym. 2021, 251, 117041. [Google Scholar] [CrossRef] [PubMed]
25. Liu, X.; Xi, X.; Jia, A.; Zhang, M.; Cui, T.; Bai, X.; Liu, C. A fucoidan from Sargassum fusiforme with novel structure and its regulatory effects on intestinal microbiota in high-fat diet-fed mice. Food Chem. 2021, 358, 129908. [Google Scholar] [CrossRef] [PubMed]
26. Lachuga, Y.F.; Meskhi, B.C.; Pakhomov, V.I.; Rudoy, D.V. Harvesting Machine. Patent of the Russian Federation No 206314 U1, 6 September 2021. [Google Scholar]
27. Pakhomov, V.; Rudoy, D.; Kambulov, S.; Maltseva, T. Research on Energy Intensity of Wheat Harvesting at Different Ripeness Phases with a New Stripping–Threshing Unit. AgriEngineering 2024, 6, 3159–3173. [Google Scholar] [CrossRef]
28. Mazanko, M.S.; Prazdnova, E.V.; Kulikov, M.P.; Maltseva, T.A.; Rudoy, D.V.; Chikindas, M.L. Antioxidant and Antimutagenic Properties of Probiotic Lactobacilli Determined Using LUX-Biosensors. Enzym. Microb. Technol. 2022, 155, 109980. [Google Scholar] [CrossRef]
29. Kolmakov, V.I.; Kolmakova, A.A. Amino acids in promising feeds for fish aquaculture: A review of experimental data. J. Sib. Fed. University. Biol. 2020, 13, 424–442. [Google Scholar] [CrossRef]
30. Abrosimova, N.A.; Abrosimov, S.S.; Saenko, E.M. Feed Raw Materials and Additives for Aquaculture Objects; Publishing House “Media-Polis”: Rostov-on-Don, Russia, 2006; 147p, Available online: http://dspace.vniro.ru/handle/123456789/1645?show=full (accessed on 27 November 2024). (In Russian)
31. Aleshin, M.A.; Mikhailova, L.A. Changes in the yield and biochemical composition of grain of field crops in mixed crops when using mineral fertilizers. Fertility 2020, 2, 9–13. (In Russian) [Google Scholar] [CrossRef]
32. Gao, L.; Haesaert, G.; Bockstaele, F.V.; Vermeir, P.; Skirtach, A.; Eeckhout, M. Combined effects of nitrogen and sulfur fertilizers on chemical composition, structure and physicochemical properties of buckwheat starch. Food Chem. 2024, 459, 140351. [Google Scholar] [CrossRef]
33. Lu, T.; Shi, J.; Lu, Z.; Wu, Z.; Wang, Y.; Luo, P.; Han, X. Appropriate application of organic fertilizer enhanced yield, microelement content, and quality of maize grain under a rotation system. Ann. Agric. Sci. 2024, 69, 19–32. [Google Scholar] [CrossRef]

Meskhi B, Pakhomov V, Rudoy D, Maltseva T, Olshevskaya A, Mazanko M. Early-Maturity Wheat as a Highly Valuable Feed Raw Material with Prebiotic Activity. Agriculture. 2025; 15(3):317. https://doi.org/10.3390/agriculture15030317
Перевод статьи «Early-Maturity Wheat as a Highly Valuable Feed Raw Material with Prebiotic Activity» авторов Meskhi B, Pakhomov V, Rudoy D, Maltseva T, Olshevskaya A, Mazanko M., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык



















































































































Комментарии (0)