Генная инженерия против аллергии: как редактируют растения
Растущее потребление растительных белков, призванное смягчить экологические проблемы, связанные с производством животных белков, ведёт к увеличению числа случаев пищевых аллергических реакций. Одним из главных драйверов сельскохозяйственной науки XXI века служит разработка и апробация конкретных подходов к модулированию экспрессии аллергенных белков в культурах ещё до сбора урожая. Увеличение числа случаев аллергии на растительные продукты обусловлено прежде всего аллергенами запасных белков семян, с чем врачи всё чаще сталкиваются в последнее время из-за более активного использования белков растительного происхождения в пищевой промышленности.
Аннотация
Возросшая доступность высокопроизводительных технологий породила постоянно растущий массив омиксных данных, что позволяет лучше понять структурные и молекулярные свойства этих белков, а значит — точнее оценивать их аллергенный потенциал. Современные системы направленного редактирования генома, не вызывающие двуцепочечных разрывов ДНК, дают возможность вносить точные модификации непосредственно в коммерческие сорта растений. Искусственный интеллект коренным образом трансформирует научные исследования на каждом этапе, помогая учёным, обрабатывая масштабные массивы данных, делая прогнозы и автоматизируя рутинные задачи. В этот эпохальный момент, отмеченный пересечением искусственного интеллекта и синтетической биологии, на сцену выходит научный ассистент нового поколения (NGA). В данной работе мы предлагаем новую концептуальную основу, которая призвана облегчить и ускорить редактирование сайтов перекрёстной реактивности для получения гипоаллергенных сортов и предотвращения плейотропных эффектов. Наконец, мы обсуждаем возможные применения этого нового подхода к организации исследований. NGA, без сомнения, способен взять под контроль эволюцию аллергии на запасные белки семян — как за счёт предсказания новых эпитопов, так и благодаря прогнозированию механизмов иммунного ответа.
Введение
Пищевая аллергия (ПА) — это IgE-опосредованный ответ иммунной системы чувствительных лиц на специфические пищевые белки, как описано в руководствах Национального института аллергии и инфекционных заболеваний (NIAID). Пищевые аллергены вызывают огромную озабоченность у потребителей и пищевой промышленности во всём мире, в частности потому, что надёжной профилактики при случайном употреблении не существует, за исключением применения инъекций адреналина для предотвращения тяжёлых анафилактических реакций (Du Toit и др., 2015; Loh и Tang, 2018). Симптомы могут варьировать от лёгких до тяжёлых в зависимости от экологических и генетических факторов. Распространённость пищевой аллергии растёт из-за сложности и глобального характера современного рациона (Liu и др., 2022), а также появления на рынке новых пищевых ингредиентов (Nitride и др., 2023; Quintieri и др., 2023). Антигенпрезентирующие клетки (АПК) распознают и процессируют аллергены, которые сначала проникают в организм через эпителиальный барьер, индуцируя выработку иммуноглобулина Е (IgE) в В-клетках. Затем IgE связывается с тучными клетками и базофилами, сенсибилизируя организм. Последующее воздействие того же антигена активирует тучные клетки и базофилы и высвобождает мощные медиаторы воспаления. Вещества, высвобождаемые этими клетками (такие как гистамин, простагландины и лейкотриены), являются основными факторами, ответственными за такие симптомы, как кожная сыпь или анафилаксия (Sathe и др., 2016; Dong и др., 2021). По данным Всемирной организации здравоохранения (ВОЗ), заболеваемость ПА в развитых странах растёт на протяжении последних двух десятилетий. Европейская академия аллергологии и клинической иммунологии оценивает распространённость аллергии среди европейских детей в пределах от 1,7% до 5%. Исследователи наблюдали аналогичные тенденции в развивающихся странах по мере их экономического роста (Leung и др., 2018; Prescott SL и др., 2013). Действительно, быстрый рост ПА очевиден в странах Азии и в Африке (Arakali и др., 2017). Gupta и др. (2013) подсчитали, что пищевые аллергии обходятся экономике в общей сложности в 24,8 миллиарда долларов в год. Среди аллергенных белков растительные аллергены поражают 1 из 100 взрослых и 1 из 10 детей (Shahali и Dadar, 2018). Согласно классификации ВОЗ/IUIS (Международный союз иммунологических обществ) и курируемым базам данных, таким как COMPARE (https://db.comparedatabase.org/) и FARRP AllergenOnline (http://www.allergenonline.org/), ряд продуктов растительного происхождения способен вызывать тяжёлые анафилактические реакции у восприимчивых лиц (Shahali и Dadar, 2018). Распространённость сенсибилизации к данному аллергенному ингредиенту значительно варьирует в зависимости от географического региона и возраста субъектов. Недавняя экспертная консультация ВОЗ/ФАО включает только глютен, арахис и отдельные древесные орехи (миндаль, кешью, фундук, пекан, фисташку и грецкий орех) в список глобально приоритетных аллергенных ингредиентов, ограничивая ряд других отдельными странами (Datema и др., 2015; Datema и др., 2018). Большинство растительных аллергенов представляют собой белки, связанные с запасающими белками семян (Breiteneder и Radauer, 2004). Запасающие белки семян (ЗБС) включают проламины и купины, которые являются наиболее распространёнными группами растительных аллергенов (Таблица 1). Суперсемейства купинов и проламинов представляют собой пример пищевых аллергенов I класса (Moreno, 2007; Moreno и Clemente, 2008). Эти белки обычно считаются высокостабильными к термической денатурации, кислой среде и желудочно-кишечным протеазам и могут вызывать аллергические реакции от умеренных до тяжёлых (Harrer и др., 2010; Mendes и др., 2019). Члены суперсемейства купинов характеризуются двумя короткими консервативными консенсусными последовательностями и коровым структурным доменом в форме бета-бочонка, что и послужило источником их обозначения «купин» (Breiteneder и Mills, 2005). Вицилины и легумины представляют собой 7S- и 11S-глобулины соответственно, с двумя коровыми доменами (бикупины), и считаются преобладающими ЗБС в ряде орехов и бобовых (Breiteneder и Mills, 2005). Учёные разделили суперсемейство проламинов на различные семейства. Неспецифические липид-транспортирующие белки (nsLTP), как правило, располагаются в различных частях растения, включая семена, листья, корни и плоды. 2S-альбумины, α-амилазы и проламины злаков обычно находятся в семенах (Breiteneder и Mills, 2005). Эти три белковых суперсемейства включают почти 65% идентифицированных на сегодняшний день растительных пищевых аллергенов. nsLTP и 2S-альбумин являются основными триггерами аллергических реакций при аллергии на арахис и сою (Ara h 2, 6 и 7 и Gly m 8 соответственно) (Taylor и др., 2021). В орехе кешью Ana o 1 и Ana o 2 (суперсемейство купинов) были классифицированы как главные аллергены, согласно Mendes и др. (2019). Radauer и др. (2017) сообщили, что в арахисе (Arachis hypogaea) 2S-альбумины Ara h 2 и Ara h 6 являются наиболее мощными аллергенами, за которыми следуют купины Ara h 1 и Ara h 3, 7S-вицилин и 11S-глобулины соответственно. Gly m 5 и Gly m 6 были идентифицированы в сое (Glycine max), Cor a 11 и Cor a 9 — в фундуке (Corylus avellana), Jug r 2 — в грецком орехе, а также некоторые другие ЗБС, принадлежащие к суперсемейству купинов, были идентифицированы как важные аллергены в бобовых, орехах и других семенах фруктов (Radauer и др., 2017; Barre и др., 2018). Запасающие белки, такие как Act d 12, 11S-глобулин, или Fra a 3, ns-LTP, являются главными аллергенными белками в таких фруктах, как киви и клубника соответственно, чьи семена употребляются вместе с мякотью (Sirvent и др., 2014; Sirvent и др., 2014). Комитет экспертов ВОЗ/ФАО по оценке риска пищевых аллергенов недавно включил киви в список наблюдения CODEX в качестве появляющегося аллергена (Risk Assessment of Food Allergens, 2022). Недавно Barre и др. (2018) представили подробную таблицу фруктов, содержащих аллергены-купины (легумин, вицилин, гермин и гермин-подобный белок). Высокая гомология последовательностей и структур часто приводит к перекрёстной аллергенности, что часто объясняется сходными эпитопными последовательностями (Shahali и Dadar, 2018). Таким образом, аллергики, чувствительные к специфическим ЗБС определённого растения, могут проявлять перекрёстную реактивность IgE к гомологичным белкам, обнаруженным в других растениях.
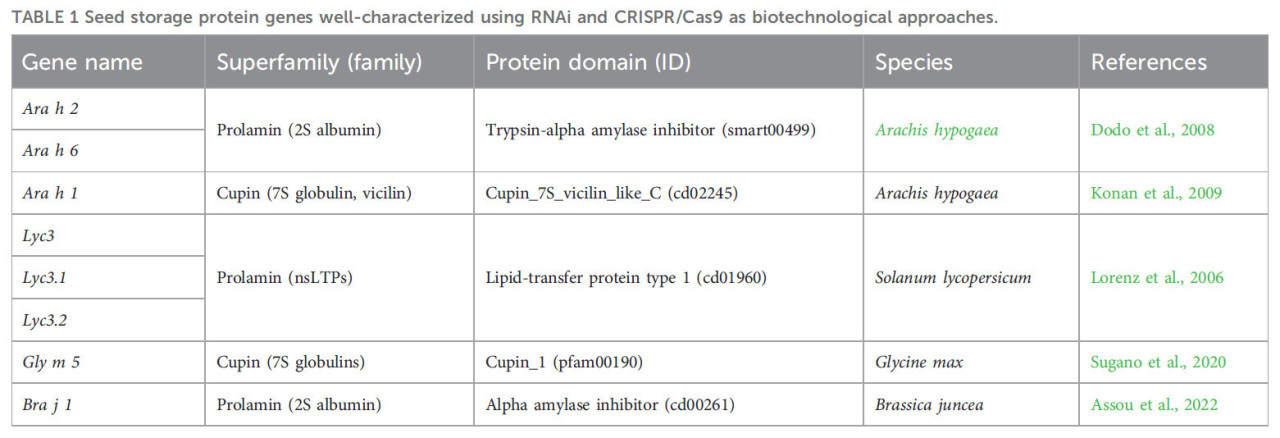
ТАБЛИЦА 1. Гены запасающих белков семян, хорошо охарактеризованные с помощью РНК-интерференции и CRISPR/Cas9 в качестве биотехнологических подходов.
Разработка генетических подходов для получения гипоаллергенных культур насчитывает более 25 лет. Эти подходы основывались на классической селекции и мутагенезе (Takahashi и др., 1996; Hajika и др., 1998). Чтобы углубить понимание ЗБС и смягчить пищевую аллергию, исследователи применяли РНК-интерференцию (РНКи) для изменения экспрессии ЗБС. Примечательно, что подавление экспрессии специфических аллергенных белков в растениях дало многообещающие результаты, что было подтверждено в исследованиях с участием арахиса и томатов (Singh и Bhalla, 2008; Scheurer, 2009). На растениях арахиса исследователи применяли РНКи-опосредованное сайленсирование генов, чтобы убрать иммунодоминантные белки Ara h 1 и Ara h 2 (Таблица 1). Chu и др. (2008) использовали биолистический метод, который привёл к стабильной трансформации гена Ara h 2 в растениях арахиса. Поскольку Ara h 2 обладает гомологией последовательности с Ara h 6, сообщалось, что РНКи-плазмида, нацеленная на Ara h 2, подавляла экспрессию белков как Ara h 2, так и Ara h 6. Масса семян и всхожесть трансгенных растений не показали значительных различий. Используя сыворотки пациентов в качестве источника IgE-антител, они наблюдали пониженный уровень иммуносвязывания, что демонстрировало заметное снижение уровня аллергенности. Аналогичным образом, метод трансформации, опосредованной Agrobacterium, использовался для сайленсинга аллергена Ara h 2 (Dodo и др., 2008). Однако несколько факторов могут препятствовать накоплению молекул РНКи в цитоплазме клетки, из-за чего РНКи может не полностью подавлять гены (Tseng и др., 2009; Ulrich и др., 2015). В связи с этим стоит отметить, что доза яблока, которая, по прогнозам, вызовет реакцию у 50% аллергической популяции, составляет 150 мг белка. Следовательно, частичное подавление генов всё ещё приемлемо, и исследователи могли бы определить подходящий уровень сайленсинга для каждого аллергенного продукта. Le и др. (2006) достигли существенного подавления Lyc e 3, аллергенного nsLTP, в трансгенных растениях томата, с пониженным накоплением по сравнению с растениями дикого типа (Таблица 1). Интересно, что одна конструкция РНКи эффективно подавляла обе изоформы Lyc e 3 — Lyc e 3.01 и Lyc e 3.02, которые показали 76,5% идентичности пар оснований. Используя две сыворотки пациентов со специфической IgE-реактивностью к Lyc e 3, удалось показать, что способность распознавать nsLTP Lyc e 3 в дефицитных томатах была снижена в 10–100 раз. В то время как тестирование in vitro с использованием сывороток аллергиков достаточно информативно в отношении IgE-связывающей способности белковых экстрактов из трансгенных растений, тестирование in vivo (кожные пробы и пищевые провокационные тесты) является золотым стандартом для оценки аллергенного потенциала. Lorenz Y и др. (2006) протестировали генетически модифицированные продукты со сниженным аллергенным потенциалом на людях с заметным снижением кожной реактивности. В дополнение к nsLTP, исследователи нацелились на профилин томата Lyc e 1 для сайленсинга генов посредством РНКи в трансгенных растениях томата (Le и др., 2006). Однако, несмотря на использование РНКи в качестве инструмента для снижения экспрессии аллергенных белков, подавление одного аллергена не отменит автоматически риск пищевой аллергии в популяции, однако оно может способствовать его смягчению (Peters и др., 2011). Часто даже мельчайшие количества аллергенных пищевых белков могут спровоцировать пищевые аллергические реакции, и множество генных продуктов могут обусловливать эти реакции (Knoll и др., 2011; Graham и др., 2020). Следовательно, исследователи могут достичь одновременного ингибирования членов семейства генов, выбирая нуклеотидные области с высокой гомологией последовательностей, которые часто находятся в кодирующих областях (Chu и др., 2008). Таким образом, у учёных есть возможность принять другие стратегии для преодоления проблем, возникающих при использовании РНКи.
В период Неолитической революции, начавшейся примерно 10 000–13 000 лет назад (Stitzer и Ross-Ibarra, 2018), нокаут генов (или аллели с потерей функции) играл заметную роль в эволюции сельскохозяйственных культур в ходе одомашнивания, диверсификации и улучшения (Monroe и др., 2020; Zhang, 2023). Современные биотехнологические подходы, как неотъемлемое дополнение к традиционной селекции растений, попытались имитировать этот сложный природный процесс, оказавшись технологиями вспомогательной эволюции для улучшения признаков культур (Zsögön и др., 2018). Система CRISPR/Cas9 часто приводит к двуцепочечным разрывам ДНК, делеции нуклеотидов и сдвигу рамки считывания с последующей потерей функции гена и, таким образом, оказывается одним из самых популярных инструментов редактирования генома (GE) для селекции растений (Aksoy и др., 2022; Li и др., 2022). Разработка и внедрение этой технологии привели к получению культур с повышенной устойчивостью к биотическим и абиотическим стрессам и сокращёнными циклами селекции (Yıldırım и др., 2023). Однако полные нокауты или нокдауны генов, релевантных для приспособленности, и не только, могут индуцировать плейотропные фенотипы (Bauer и др., 2015; Auge и др., 2019; Hanika и др., 2021), поскольку один ген может прямо или косвенно влиять на множество признаков (Burstin и др., 2007; Nguyen и др., 2015). Более того, ограниченная геномная информация может приводить к непреднамеренным нецелевым эффектам или порождать новые мутантные варианты из-за репарации после расщепления (Yıldırım и др., 2023).
CRISPR/Cas9-опосредованный нокаут генов также использовался для смягчения ПА путём нацеливания на гены, кодирующие ЗБС. Например, в недавнем исследовании были внесены крупные делеции и сдвиги рамки считывания в оба гомолога Bra j 1, главного аллергена в растениях горчицы сарептской (Brassica juncea) (Таблица 1). Мутации были унаследованы поколением T1, которое показало пониженные количества или отсутствие Bra j 1 в экстрактах семян (Assou и др., 2022). Аналогичным образом, система CRISPR-Cas9 была использована для индукции мутаций в генах, кодирующих 7S-альфа- и альфа-штрих-субъединицы бета-конглицинина, аллерген Gly m 5 в соевых бобах (Таблица 1). Этот подход привёл к созданию потомства, свободного от cas9, лишённого этих главных аллергенных белков в семенах, при сохранении технологических свойств (Sugano и др., 2020). Однако для обнаружения нежелательных эффектов, которые могли ускользнуть от визуальной оценки, обычно необходима глубокая фенотипическая характеристика. Инновационная работа была также проведена на зёрнах итальянской твёрдой пшеницы Свево. Camerlengo и др. (2020) применили мультиплексный подход CRISPR-Cas9, свободный от маркерных генов, для индукции мутаций в субъединицах ингибиторов α-амилазы/трипсина (ATI) WTAI-CM3 и WTAI-CM16, которые признаны ключевыми белками, вовлечёнными в развитие астмы пекарей. Хотя они успешно достигли полного нокаута генов, они отметили появление плейотропных эффектов, приписываемых активации псевдогена ATI 0.28 в редактированных линиях. Вмешиваясь в хорошо налаженные генные сети выше и ниже по течению, классический нокаут генов может приводить к непреднамеренным плейотропным эффектам, что согласуется с этими представлениями. Учёные углубили наше понимание, показав, что системная биология растений, включая развитие и созревание семян, включает сложные и запутанные генные сети (Duan и др., 2023). Хотя регуляторная сеть, контролирующая ЗБС в развивающихся семенах, проясняется, мутации с потерей функции в главных регуляторах созревания семян порождают нежелательные фенотипы семян, включая значительное снижение содержания ЗБС (Tang и др., 2008). Выбор мишеней среди ЗБС для получения свободных от аллергенов культур требует всестороннего понимания регуляции генов в росте и развитии растений (Ojolo и др., 2018). Современные и инновационные подходы точного редактирования генома (PGE), хорошо рассмотренные Yıldırım и др. (2023), позволяют модифицировать определённые участки генов с помощью каталитически нарушенного Cas9, обеспечивая прямую запись новой генетической информации в указанный сайт ДНК (Andolfo и др., 2016). Недавние достижения, включая идентификацию новых вариантов Cas и разработку новых стратегий редактирования, продемонстрировали эволюционирующий ландшафт методов PGE в растениях (Yanagawa и др., 2023; Yıldırım и др., 2023). Это позволило бы нам специфически модифицировать эпитопную последовательность вместо подавления белков, тем самым избегая риска плейотропных эффектов, связанных с традиционными методами нокаута генов. Кроме того, это можно было бы использовать для модификации сайтов перекрёстной реактивности в генах, кодирующих ЗБС. В частности, огромный объём информации, полученной на сегодняшний день об использовании методов PGE в растениях с помощью комплекса CRISPR-Cas (Рисунок 1), включая направленную гомологичную репарацию (HDR), базовое редактирование, прайм-редактирование, РНК-направляемые транспозазы и последний CRISPR-инструмент под названием «программируемое добавление через сайт-специфичный нацеливающий элемент» (PASTE) (Strecker и др., 2019; Anzalone и др., 2020; Anzalone и др., 2022), мог бы контекстуально оцениваться программным обеспечением ИИ для определения наилучших методов для конкретной цели (например, замена конкретного нуклеотида, введение или замена специфических нуклеотидных фрагментов).
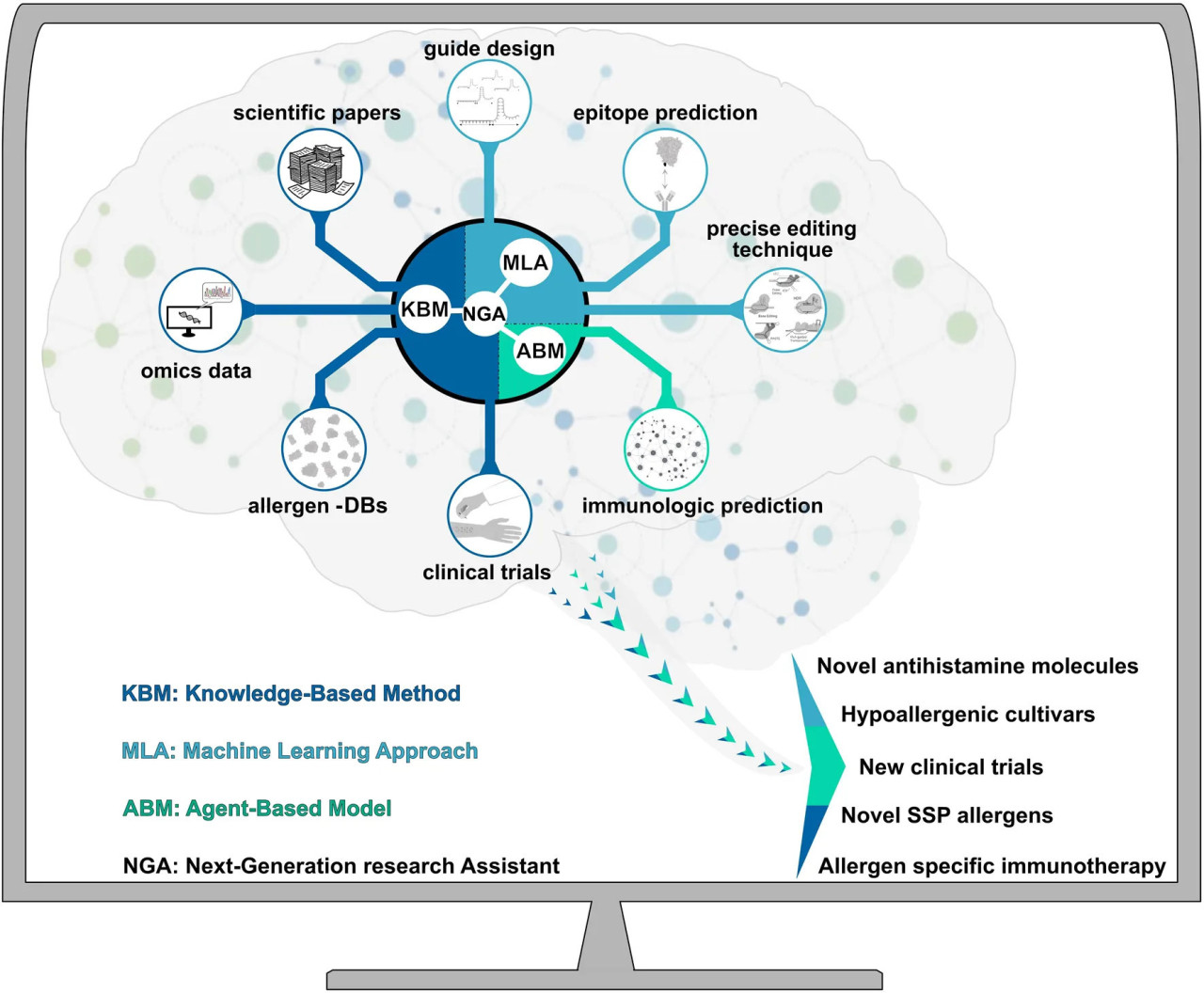
РИСУНОК 1. Научный ассистент нового поколения (NGA) для синтетической биологии с поддержкой искусственного интеллекта. Интегрируя методы, основанные на знаниях (KBM), подходы машинного обучения (MLA) и агент-ориентированные модели (ABM), NGA может значительно расширить наши представления о точном редактировании генов ЗБС и способствовать разработке терапевтических стратегий против аллергенов запасающих белков семян. KBM может определять цели по характеристикам из публичных и частных аллергенных мульти-омиксных (геномика, транскриптомика, протеомика, метаболомика и феномика) и цифровых (базы данных аллергенов) данных, клинических испытаний и научных знаний. MLA могут идентифицировать новые эпитопы (антиген-связывающий сайт), нецелевые и целевые участки и наиболее подходящий метод точного редактирования. ABM могут предсказывать реакции врождённого иммунитета, связанные с конкретным аллергеном ЗБС. Стрелка показывает различные результаты (новые антигистаминные молекулы, гипоаллергенные сорта, новые клинические испытания, новые пищевые аллергены и аллерген-специфическую иммунотерапию), генерируемые исследовательской деятельностью искусственного интеллекта.
Обсуждение
В этом сценарии подходы синтетической биологии (SynBio), которые встречаются с методами искусственного интеллекта (ИИ), стали реальной возможностью для борьбы с пищевой аллергией, связанной с ЗБС (MacMath и др., 2023). Научный ассистент нового поколения (NGA) на основе комбинации ИИ-SynBio может быть conceived как лабораторное оборудование, поддерживающее исследовательскую деятельность (Рисунок 1). ИИ включает методы, основанные на знаниях (KBM), подходы машинного обучения (MLA) и агент-ориентированные модели (ABM) (Maserat, 2022). KBM могут определять цели отбора признаков из аллергенных омиксных данных (геномика, транскриптомика, протеомика, метаболомика и феномика), биологических и гиперчувствительных реакционных сущностей. Исследователи имеют доступ к огромному объёму информации, полученной с помощью различных платформ, таких как омиксные данные, полученные при анализе отдельных клеток или многоклеточных тканей (Haque и др., 2019). Инновации высокопроизводительных биотехнологий позволили накапливать омиксные данные с угрожающей скоростью, тем самым открыв эру «больших данных». Обнаружение и выяснение сложных взаимодействий между различными данными может быть сложным и трудоёмким процессом (Amoroso и др., 2023). Опубликованные и частные данные, сгенерированные различными омиксными платформами и приборами (например, омиксные данные, научные доказательства, наборы данных о клинических тестах), могут одновременно анализироваться NGA. Интересно, что KBM использовались для облегчения интерпретации и понимания сложных омиксных данных. Например, Türei и др. (2021) провели увлекательное исследование, проанализировав несколько омиксных данных для изучения межклеточных взаимодействий и нижестоящих внутриклеточных сигнальных процессов у людей, мышей и крыс. В последние годы вычислительные методы также использовались для создания доступных баз данных аллергенов, содержащих большой объём информации об эпитопах аллергенов или антителах (Kadam и др., 2017). Таким образом, для обработки биологических больших данных современные прогностические модели значительно расширили наши представления о лечении аллергии, в том числе путём обнаружения растительных генетических паттернов, которые потенциально могут быть модифицированы методами точного редактирования (Haque и др., 2019; Li и др., 2022). Разработка и применение MLA и ABM могут выполнять анализы для предсказания эпитопов и иммунологических прогнозов (Jumper и др., 2021). Действительно, подходы машинного обучения использовались для прогнозирования потенциальной аллергенности новых белков (Jumper и др., 2021; Nedyalkova и др., 2023). Методы МО внесли вклад в понимание аллергенов и их потенциальных эпитопов в растениях через предсказание эпитопов и иммунологический анализ (Wang и др., 2013). Поэтому мы предполагаем, что в будущем NGA сможет направлять учёных к более точным и эффективным исследованиям в области синтетической биологии для разработки новых гипоаллергенных культур, в том числе через идентификацию новых пищевых аллергенов, антигистаминных молекул и проведение клинических испытаний (Рисунок 1) (Garcia-Moreno и Gutiérrez-Naranjo, 2022). Глубокие нейронные сети использовались для предсказания активности и специфичности направляющих последовательностей РНК, используемых PGE (Luo и др., 2019). ИИ уже помог в прогнозировании наилучших условий in vitro для роста растений (Malabadi и др., 2023). Аналогично, Akin и др. (2020) применили метод интеллектуального анализа данных в исследованиях тканевых культур, который оказался многообещающим инструментом для анализа непараметрических данных. Биологические наборы данных о растениях часто искажены, высокоразмерны, динамичны и гетерогенны (Ben Ayed и др., 2022). NGA также может поддержать выбор параметрических или непараметрических алгоритмов машинного обучения, анализируя свойства данных, подлежащих обработке. Действительно, наиболее ключевыми этапами для эффективного использования MLA являются проектирование процесса трансформации данных и предобработки (Booij и др., 2022). NGA, интегрируя различные системы ИИ, может служить научным инструментом для анализа информации, полученной с помощью разных методов, облегчая идентификацию наилучшего метода PGE, процедуры трансформации и регенерации для конкретной цели (Рисунок 1). Следовательно, в этом глобальном видении PGE становится инструментом, в котором синтез фрагментов ДНК/РНК, сборка вектора и выбор метода трансформации становятся непосредственно доступными для исследователя, в зависимости от конечной цели исследования. На сегодняшний день МО уже доказало, что является революционным инструментом в поддержке клинических исследований, медицинской и точной диагностики (Mansour и др., 2021; Fang и др., 2022) с применением в медицинской диагностике и для обнаружения, прогнозирования и предсказания иммунных ответов (Nawaz и др., 2021; Buendgens и др., 2022). В этом контексте использование NGA для всестороннего анализа аллергенов может быть полностью революционным для здоровья человека. NGA мог бы работать как «полноценный исследователь» (Рисунок 1), предоставляя новые идеи для разработки гипоаллергенных культур. В ближайшем будущем исследователи должны рассматривать NGA как полезное лабораторное оборудование для ускорения и улучшения их исследовательской работы. Однако оценка результатов по-прежнему останется фундаментальной частью процесса.
Ссылки
1. Akin M., Eyduran S. P., Eyduran E., Reed B. M. (2020). Analysis of macro nutrient related growth responses using multivariate adaptive regression splines. Plant Cell, Tissue Organ Cult. (PCTOC) 140 (3), 661–670. doi: 10.1007/s11240-019-01763-8. CrossRef Google Scholar
2. Aksoy E., Yildirim K., Kavas M., Kayihan C., Yerlikaya B. A., Çalik I., et al (2022). General guidelines for CRISPR/Cas-based genome editing in plants. Mol. Biol. Rep. 49 (12), 12151–12164. doi: 10.1007/s11033-022-07773-8. CrossRef Google Scholar
3. Amoroso C. G., D’Esposito D., Aiese Cigliano R., Ercolano M. R. (2023). Comparison of tomato transcriptomic profiles reveals overlapping patterns in abiotic and biotic stress responses. Int. J. Mol. Sci. 24 (4), 4061. doi: 10.3390/ijms24044061. CrossRef Google Scholar
4. Andolfo G., Iovieno P., Frusciante L., Ercolano M. R. (2016). Genome-editing technologies for enhancing plant disease resistance. Front. Plant Sci. 7, 1813. doi: 10.3389/fpls.2016.01813. CrossRef Google Scholar
5. Anzalone A. V., Gao X. D., Podracky C. J., Nelson A. T., Koblan L. W., Raguram A., et al (2022). Programmable deletion, replacement, integration and inversion of large DNA sequences with twin prime editing. Nat. Biotechnol. 40 (5), 731–740. doi: 10.1038/s41587-021-01133-w. CrossRef Google Scholar
6. Anzalone A. V., Koblan L. W., Liu D. R. (2020). Genome editing with CRISPR–Cas nucleases, base editors, transposases and prime editors. Nat. Biotechnol. 38 (7), 824–844. doi: 10.1038/s41587-020-0561-9. CrossRef Google Scholar
7. Arakali S. R., Green T. D., Dinakar C. (2017). Prevalence of food allergies in south asia. Ann. Allergy, Asthma & Immunol. 118 (1), 16–20. doi: 10.1016/j.anai.2016.09.441. CrossRef Google Scholar
8. Assou J., Zhang D., Roth K. D. R., Steinke S., Hust M., Reinard T., et al (2022). Removing the major allergen Bra j I from brown mustard (Brassica juncea) by CRISPR/Cas9. Plant J. 109 (3), 649–663. doi: 10.1111/tpj.15584. CrossRef Google Scholar
9. Auge G. A., Penfield S., Donohue K. (2019). Pleiotropy in developmental regulation by flowering‐ pathway genes: is it an evolutionary constraint? New Phytol. 224 (1), 55–70. doi: 10.1111/nph.15901. CrossRef Google Scholar
10. Barre A., Simplicien M., Benoist H., Rougé P. (2018). Fruit allergies: beware of the seed allergens. Rev. Française d'Allergologie 58 (4), 308–317. doi: 10.1016/j.reval.2018.01.009. CrossRef Google Scholar
11. Bauer C. R., Li S., Siegal M. L. (2015). Essential gene disruptions reveal complex relationships between phenotypic robustness, pleiotropy, and fitness. Mol. Syst. Biol. 11 (1), 773. doi: 10.15252/msb.20145264. CrossRef Google Scholar
12. Ben Ayed R., Hanana M., Ercisli S., Karunakaran R., Rebai A., Moreau F. (2022). Integration of innovative technologies in the agri-food sector: the fundamentals and practical case of DNA-based traceability of olives from fruit to oil. Plants 2022, 11, 1230. doi: 10.3390/plants11091230. CrossRef Google Scholar
13. Booij T. M., Chiscop I., Meeuwissen E., Moustafa N., Hartog F. T. H. d. (2022). ToN_IoT: the role of heterogeneity and the need for standardization of features and attack types in IoT network intrusion data sets. IEEE Internet Things J. 9 (1), 485–496. doi: 10.1109/jiot.2021.3085194. CrossRef Google Scholar
14. Breiteneder H., Mills E. N. C. (2005). Plant food allergens — structural and functional aspects of allergenicity. Biotechnol. Adv. 23 (6), 395–399. doi: 10.1016/j.biotechadv.2005.05.004. CrossRef Google Scholar
15. Breiteneder H., Radauer C. (2004). A classification of plant food allergens☆. J. Allergy Clin. Immunol. 113(5): 821–830. doi: 10.1016/j.jaci.2004.01.779. CrossRef Google Scholar
16. Buendgens L., Cifci D., Ghaffari Laleh N., van Treeck M., Koenen M. T., Zimmermann H. W., et al (2022). Weakly supervised end-to-end artificial intelligence in gastrointestinal endoscopy. Sci. Rep. 12 (1), 4829. doi: 10.1038/s41598-022-08773-1. CrossRef Google Scholar
17. Burstin J., Marget P., Huart M., Moessner A., Mangin B., Duchene C., et al (2007). Developmental genes have pleiotropic effects on plant morphology and source capacity, eventually impacting on seed protein content and productivity in pea. Plant Physiol. 144 (2), 768–781. doi: 10.1104/pp.107.096966. CrossRef Google Scholar
18. Camerlengo F., Frittelli A., Sparks C., Doherty A., Martignago D., Larré C., et al (2020). CRISPR-Cas9 multiplex editing of the α-amylase/trypsin inhibitor genes to reduce allergen proteins in durum wheat. Front. Sustain. Food Syst. 4, 104. doi: 10.3389/fsufs.2020.00104. CrossRef Google Scholar
19. Chu Y., Faustinelli P., Ramos M. L., Hajduch M., Stevenson S., Thelen J. J., et al (2008). Reduction of IgE binding and nonpromotion of Aspergillus flavus fungal growth by simultaneously silencing Ara h 2 and Ara h 6 in peanut. J. Agric. Food Chem. 56 (23), 11225–11233. doi: 10.1021/jf802600r. CrossRef Google Scholar
20. Datema M. R., Van Ree R., Asero R., Barreales L., Belohlavkova S., de Blay F., et al (2018). Component‐ resolved diagnosis and beyond: multivariable regression models to predict severity of hazelnut allergy. Allergy 73 (3), 549–559. doi: 10.1111/all.13328. CrossRef Google Scholar
21. Datema M. R., Zuidmeer-Jongejan L., Asero R., Barreales L., Belohlavkova S., de Blay F., et al (2015). Hazelnut allergy across Europe dissected molecularly: a EuroPrevall outpatient clinic survey. J. Allergy Clin. Immunol. 136 (2), 382–391. doi: 10.1016/j.jaci.2014.12.1949. CrossRef Google Scholar
22. Dodo H. W., Konan K. N., Chen F. C., Egnin M., Viquez O. M. (2008). Alleviating peanut allergy using genetic engineering: the silencing of the immunodominant allergen Ara h 2 leads to its significant reduction and a decrease in peanut allergenicity. Plant Biotechnol. J. 6 (2), 135–145. doi: 10.1111/j.1467-7652.2007.00292.x. CrossRef Google Scholar
23. Dong X., Wang J., Raghavan V. (2021). Critical reviews and recent advances of novel non-thermal processing techniques on the modification of food allergens. Crit. Rev. Food Sci. Nutr. 61 (2), 196–210. doi: 10.1080/10408398.2020.1722942. CrossRef Google Scholar
24. Duan Z., Li Q., Wang H., He X., Zhang M. (2023). Genetic regulatory networks of soybean seed size, oil and protein contents. Front. Plant Sci. 14, 1160418. doi: 10.3389/fpls.2023.1160418. CrossRef Google Scholar
25. Du Toit G., Roberts G., Sayre P. H., Bahnson H. T., Radulovic S., Santos A. F., et al (2015). Randomized trial of peanut consumption in infants at risk for peanut allergy. N. Engl. J. Med. 372 (9), 803–813. doi: 10.1056/NEJMoa1414850. CrossRef Google Scholar
26. Fang Y., Idnay B., Sun Y., Liu H., Chen Z., Marder K., et al (2022). Combining human and machine intelligence for clinical trial eligibility querying. J. Am. Med. Inf. Assoc. 29 (7), 1161–1171. doi: 10.1093/jamia/ocac051. CrossRef Google Scholar
27. Garcia-Moreno F. M., Gutiérrez-Naranjo M. A. (2022). ALLERDET: a novel web app for prediction of protein allergenicity. J. Biomed. Inf. 135, 104217. doi: 10.1016/j.jbi.2022.104217. CrossRef Google Scholar
28. Graham F., Caubet J., Eigenmann P. A. (2020). Can my child with IgE‐ mediated peanut allergy introduce foods labeled with "may contain traces"? Pediatr. Allergy Immunol. Santos A 31 (6), 601–607. doi: 10.1111/pai.13244. CrossRef Google Scholar
29. Gupta R., Holdford D., Bilaver L., Dyer A., Holl J. L., Meltzer D. (2013). The economic impact of childhood food allergy in the United States. JAMA Pediatr. 167 (11), 1026–1031. doi: 10.1001/jamapediatrics.2013.2376. CrossRef Google Scholar
30. Hajika M., Takahashi M., Sakai S., Matsunaga R. (1998). Dominant inheritance of a trait lacking β-conglycinin detected in a wild soybean line. Jpn. J. Breed. 48 (4), 383–386. doi: 10.1270/jsbbs1951.48.383. CrossRef Google Scholar
31. Hanika K., Schipper D., Chinnappa S., Oortwijn M., Schouten H. J., Thomma B. P. H. J., et al (2021). Impairment of tomato WAT1 enhances resistance to vascular wilt fungi despite severe growth defects. Front. Plant Sci. 12, 721674. doi: 10.3389/fpls.2021.721674. CrossRef Google Scholar
32. Haque S., Ahmad J. S., Clark N. M., Williams C. M., Sozzani R. (2019). Computational prediction of gene regulatory networks in plant growth and development. Curr. Opin. Plant Biol. 47, 96–105. doi: 10.1016/j.pbi.2018.10.005. CrossRef Google Scholar
33. Harrer A., Egger M., Gadermaier G., Erler A., Hauser M., Ferreira F., et al (2010). Characterization of plant food allergens: an overview on physicochemical and immunological techniques. Mol. Nutr. Food Res. 54 (1), 93–112. doi: 10.1002/mnfr.200900096. CrossRef Google Scholar
34. Jumper J., Evans R., Pritzel A., Green T., Figurnov M., Ronneberger O., et al (2021). Highly accurate protein structure prediction with AlphaFold. Nature 596 (7873), 583–589. doi: 10.1038/s41586-021-03819-2. CrossRef Google Scholar
35. Kadam K., Karbhal R., Jayaraman V. K., Sawant S., Kulkarni-Kale U. (2017). AllerBase: a comprehensive allergen knowledgebase. Database 2017, bax066. doi: 10.1093/database/bax066. CrossRef Google Scholar
36. Knoll J. E., Ramos M. L., Zeng Y., Holbrook C. C., Chow M., Chen S., et al (2011). TILLING for allergen reduction and improvement of quality traits in peanut (Arachis hypogaea L.). BMC Plant Biol. 11 (1), 81. doi: 10.1186/1471-2229-11-81. CrossRef Google Scholar
37. Konan K. N., Ananga A. O., Dodo H. (2009). "Reducing peanut allergy risks by means of genetic modification," in Modification of seed composition to promote health and nutrition. Editor Krishnan, H. (Madison, WI: Agronomy Monograph) 51, 289–302. Google Scholar
38. Le L. Q., Lorenz Y., Scheurer S., Fötisch K., Enrique E., Bartra J., et al (2006). Design of tomato fruits with reduced allergenicity by dsRNAi‐ mediated inhibition of ns‐ LTP (Lyc e 3) expression. Plant Biotechnol. J. 4 (2), 231–242. doi: 10.1111/j.1467-7652.2005.00175.x. CrossRef Google Scholar
39. Leung A. S. Y., Wong G. W. K., Tang M. L. K. (2018). Food allergy in the developing world. J. Allergy Clin. Immunol. 141 (1), 76–78. doi: 10.1016/j.jaci.2017.11.008. CrossRef Google Scholar
40. Li R., Li L., Xu Y., Yang J. (2022). Machine learning meets omics: applications and perspectives. Briefings Bioinforma. 23 (1), bbab460. doi: 10.1093/bib/bbab460. CrossRef Google Scholar
41. Liu Q., Lin S., Sun N. (2022). How does food matrix components affect food allergies, food allergens and the detection of food allergens? A systematic review. Trends Food Sci. Technol. 127, 280–290. doi: 10.1016/j.tifs.2022.07.009. CrossRef Google Scholar
42. Loh W., Tang M. (2018). Adjuvant therapies in food immunotherapy. Immunol. Allergy Clin. N. Am. 38 (1), 89–101. doi: 10.1016/j.iac.2017.09.008. CrossRef Google Scholar
43. Lorenz Y., Enrique E., Lequynh L., Fötisch K., Retzek M., Biemelt S., et al (2006). Skin prick tests reveal stable and heritable reduction of allergenic potency of gene-silenced tomato fruits. J. Allergy Clin. Immunol. 118 (3), 711–718. doi: 10.1016/j.jaci.2006.05.014. CrossRef Google Scholar
44. Luo J., Chen W., Xue L., Tang B. (2019). Prediction of activity and specificity of CRISPR-Cpf1 using convolutional deep learning neural networks. BMC Bioinforma. 20 (1), 332. doi: 10.1186/s12859-019-2939-6. CrossRef Google Scholar
45. MacMath D., Chen M., Khoury P. (2023). Artificial intelligence: exploring the future of innovation in allergy Immunology. Curr. Allergy Asthma Rep. 23 (6), 351–362. doi: 10.1007/s11882-023-01084-z. CrossRef Google Scholar
46. Malabadi R. B., Tl N., Kolkar K. P., Chalannavar R. K., Mudigoudra B., Lakshminarayana L., et al (2023). Cannabis sativa: applications of artificial intelligence AI and plant tissue culture for micropropagation. Int. J. Res. Innovation Appl. Sci. 8, 117–142. doi: 10.51584/ijrias.2023.8614. CrossRef Google Scholar
47. Mansour R. F., Escorcia-Gutierrez J., Gamarra M., Gupta D., Castillo O., Kumar S. (2021). Unsupervised deep learning based variational autoencoder model for COVID-19 diagnosis and classification. Pattern Recognit. Lett. 151, 267–274. doi: 10.1016/j.patrec.2021.08.018. CrossRef Google Scholar
48. Maserat E. (2022). Integration of artificial intelligence and CRISPR/Cas9 system for vaccine design. Cancer Inf. 21, 11769351221140102. doi: 10.1177/11769351221140102. CrossRef Google Scholar
49. Mendes C., Costa J., Vicente A. A., Oliveira M. B. P. P., Mafra I. (2019). Cashew nut allergy: clinical relevance and allergen characterisation. Clin. Rev. Allergy & Immunol. 57 (1), 1–22. doi: 10.1007/s12016-016-8580-5. CrossRef Google Scholar
50. Monroe J. G., Arciniegas J. P., Moreno J. L., Sánchez F., Sierra S., Valdes S., et al (2020). The lowest hanging fruit: beneficial gene knockouts in past, present, and future crop evolution. Curr. Plant Biol. 24, 100185. doi: 10.1016/j.cpb.2020.100185. CrossRef Google Scholar
51. Moreno F. J. (2007). Gastrointestinal digestion of food allergens: effect on their allergenicity. Biomed. Pharmacother. 61 (1), 50–60. doi: 10.1016/j.biopha.2006.10.005. CrossRef Google Scholar
52. Moreno F. J., Clemente A. (2008). 2S albumin storage proteins: what makes them food allergens? Open Biochem. J. 2, 16–28. doi: 10.2174/1874091X00802010016. CrossRef Google Scholar
53. Nawaz M. S., Fournier-Viger P., Shojaee A., Fujita H. (2021). Using artificial intelligence techniques for COVID-19 genome analysis. Appl. Intell. 51 (5), 3086–3103. doi: 10.1007/s10489-021-02193-w. CrossRef Google Scholar
54. Nedyalkova M., Vasighi M., Azmoon A., Naneva L., Simeonov V. (2023). Sequence-based prediction of plant allergenic proteins: machine learning classification approach. ACS Omega 8 (4), 3698–3704. doi: 10.1021/acsomega.2c02842. CrossRef Google Scholar
55. Nguyen T.-P., Cueff G., Hegedus D. D., Rajjou L., Bentsink L. (2015). A role for seed storage proteins in Arabidopsis seed longevity. J. Exp. Bot. 66 (20), 6399–6413. doi: 10.1093/jxb/erv348. CrossRef Google Scholar
56. Nitride C., D’Auria G., Ferrara A. (2023). "Environmental, nutritional, and cultural sustainability of novel food protein sources," in Sustainable food science - a comprehensive approach (Elsevier), 199–215. Google Scholar
57. Ojolo S. P., Cao S., Priyadarshani SVGN, Li W., Yan M., Aslam M., et al (2018). Regulation of plant growth and development: a review from a chromatin remodeling perspective. Front. Plant Sci. 9, 1232. doi: 10.3389/fpls.2018.01232. CrossRef Google Scholar
58. Peters S., Imani J., Mahler V., Foetisch K., Kaul S., Paulus K. E., et al (2011). Dau c 1.01 and Dau c 1.02-silenced transgenic carrot plants show reduced allergenicity to patients with carrot allergy. Transgenic Res. 20 (3), 547–556. doi: 10.1007/s11248-010-9435-0. CrossRef Google Scholar
59. Prescott S. L., Pawankar R., Allen K. J., Campbell D. E., Sinn J. K., Fiocchi A., et al (2013). A global survey of changing patterns of food allergy burden in children. World Allergy Organ. J. 6, 21. doi: 10.1186/1939-4551-6-21. CrossRef Google Scholar
60. Quintieri L., Nitride C., De Angelis E., Lamonaca A., Pilolli R., Russo F., et al (2023). Alternative protein sources and novel foods: benefits, food applications and safety issues. Nutrients 15 (6), 1509. doi: 10.3390/nu15061509. CrossRef Google Scholar
61. Radauer C., Kleine-Tebbe J., Beyer K. (2017). "Stable plant food allergens II: storage proteins," in Molecular allergy diagnostics. Editors Kleine-Tebbe J., Jakob T. (Cham: Springer International Publishing), 77–90. Google Scholar
62. Risk Assessment of Food Allergens (2022). Part 1: Review and Validation of codex alimentarius priority allergen List through risk assessment (2022) FAO. Google Scholar
63. Sathe S. K., Liu C., Zaffran V. D. (2016). Food allergy. Annu. Rev. Food Sci. Technol. 7 (1), 191–220. doi: 10.1146/annurev-food-041715-033308. CrossRef Google Scholar
64. Scheurer S., Sonnewald S. (2009). Genetic engineering of plant food with reduced allergenicity. Front. Biosci. Volume (14), 59–71. doi: 10.2741/3231. CrossRef Google Scholar
65. Shahali Y., Dadar M. (2018). Plant food allergy: influence of chemicals on plant allergens. Food Chem. Toxicol. 115, 365–374. doi: 10.1016/j.fct.2018.03.032. CrossRef Google Scholar
66. Singh M. B., Bhalla P. L. (2008). Genetic engineering for removing food allergens from plants. Trends Plant Sci. 13 (6), 257–260. doi: 10.1016/j.tplants.2008.04.004. CrossRef Google Scholar
67. Sirvent S., Cantó B., Gómez F., Blanca N., Cuesta-Herranz J., Canto G., et al (2014). Detailed characterization of Act d 12 and Act d 13 from kiwi seeds: implication in IgE cross-reactivity with peanut and tree nuts. Allergy 69 (11), 1481–1488. doi: 10.1111/all.12486. CrossRef Google Scholar
68. Stitzer M. C., Ross‐Ibarra J. (2018). Maize domestication and gene interaction. New Phytol. 220 (2), 395–408. doi: 10.1111/nph.15350. CrossRef Google Scholar
69. Strecker J., Ladha A., Gardner Z., Schmid-Burgk J. L., Makarova K. S., Koonin E. V., et al (2019). RNA-guided DNA insertion with CRISPR-associated transposases. Science 365 (6448), 48–53. doi: 10.1126/science.aax9181. CrossRef Google Scholar
70. Sugano S., Hirose A., Kanazashi Y., Adachi K., Hibara M., Itoh T., et al (2020). Simultaneous induction of mutant alleles of two allergenic genes in soybean by using site-directed mutagenesis. BMC Plant Biol. 20 (1), 513. doi: 10.1186/s12870-020-02708-6. CrossRef Google Scholar
71. Takahashi K., Mizuno Y., Yumoto S., Kitamura K., Nakamura S. (1996). Inheritance of the α-subunit deficiency of β-conglycinin in soybean (Glycine max L. MERRILL) line induced by γ-ray irradiation. Jpn. J. Breed. 46 (3), 251–255. doi: 10.1270/jsbbs1951.46.251. CrossRef Google Scholar
72. Tang X., Hou A., Babu M., Nguyen V., Hurtado L., Lu Q., et al (2008). The arabidopsis BRAHMA chromatin-remodeling ATPase is involved in repression of seed maturation genes in leaves. Plant Physiol. 147 (3), 1143–1157. doi: 10.1104/pp.108.121996. CrossRef Google Scholar
73. Taylor S. L., Marsh J. T., Koppelman S. J., Kabourek J. L., Johnson P. E., Baumert J. L. (2021). A perspective on pea allergy and pea allergens. Trends Food Sci. Technol. 116, 186–198. doi: 10.1016/j.tifs.2021.07.017. CrossRef Google Scholar
74. Tseng Y.-C., Mozumdar S., Huang L. (2009). Lipid-based systemic delivery of siRNA. Adv. Drug Deliv. Rev. 61 (9), 721–731. doi: 10.1016/j.addr.2009.03.003. CrossRef Google Scholar
75. Türei D., Valdeolivas A., Gul L., Palacio-Escat N., Klein M., Ivanova O., et al (2021). Integrated intra‐ and intercellular signaling knowledge for multicellular omics analysis. Mol. Syst. Biol. 17 (3), e9923. doi: 10.15252/msb.20209923. CrossRef Google Scholar
76. Ulrich J., Dao V. A., Majumdar U., Schmitt-Engel C., Schwirz J., Schultheis D., et al (2015). Large scale RNAi screen in Tribolium reveals novel target genes for pest control and the proteasome as prime target. BMC Genomics 16 (1), 674. doi: 10.1186/s12864-015-1880-y. CrossRef Google Scholar
77. Wang J., Yu Y., Zhao Y., Zhang D., Li J. (2013). Evaluation and integration of existing methods for computational prediction of allergens. BMC Bioinforma. 14 (S4), S1. doi: 10.1186/1471-2105-14-S4-S1. CrossRef Google Scholar
78. Yanagawa Y., Suenaga Y., Iijima Y., Endo M., Sanada N., Katoh E., et al (2023). Genome editing by introduction of Cas9/sgRNA into plant cells using temperature-controlled atmospheric pressure plasma. PLOS ONE Ezura H. 18 (2), e0281767. doi: 10.1371/journal.pone.0281767. CrossRef Google Scholar
79. Yıldırım K., Miladinović D., Sweet J., Akin M., Galović V., Kavas M., et al (2023). Genome editing for healthy crops: traits, tools and impacts. Front. Plant Sci. 14, 1231013. doi: 10.3389/fpls.2023.1231013. CrossRef Google Scholar
80. Zhang J. (2023). Patterns and evolutionary consequences of pleiotropy. Annu. Rev. Ecol. Evol. Syst. 54 (1), 1–19. doi: 10.1146/annurev-ecolsys-022323-083451. CrossRef Google Scholar
81. Zsögön A., Čermák T., Naves E. R., Notini M. M., Edel K. H., Weinl S., et al (2018). De novo domestication of wild tomato using genome editing. Nat. Biotechnol. 36 (12), 1211–1216. doi: 10.1038/nbt.4272. CrossRef Google Scholar

Evangelista AR, Amoroso CG, Nitride C and Andolfo G (2024) Seed storage allergens tackled via next-generation research assistant. Front. Food. Sci. Technol. 4:1372770. doi: 10.3389/frfst.2024.1372770
Перевод статьи «Seed storage allergens tackled via next-generation research assistant» авторов Evangelista AR, Amoroso CG, Nitride C and Andolfo G., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык
Фото: magnific

















































































































Комментарии (0)