Как сохранить красную икру без лишней соли и консервантов?
Сразу после сбора икры пинагора (Cyclopterus lumpus) в арктических регионах распространенной практикой является консервирование свежей икры в рассоле для получения соленого промежуточного продукта (СПП), который можно транспортировать и хранить в охлажденном состоянии до 1 года перед розничной обработкой. Поскольку икра подвержена росту микроорганизмов и окислению липидов, рассолы составляют из смесей соли (>10%) и бензойной кислоты для обеспечения качества и безопасности СПП. Однако на европейском рынке появляется спрос на снижение использования соли и бензойной кислоты. Поэтому целью данного исследования было оценить влияние пяти различных рассолов, состоящих из комбинаций соли, органических кислот и антиоксидантов, на контроль микробиологических и химических изменений СПП, чтобы иметь возможность отказаться от использования бензойной кислоты и снизить содержание соли.
Аннотация
Свежесобранную икру смешивали с рассолами для получения 7 или 10% (мас./мас.) NaCl и комбинаций уксусной, бензойной и/или молочной кислот с добавлением VivOx 7.5 (IFF Inc.) или Herbalox D-450 (Kalsec Inc.) в качестве антиоксидантов или без них. Соленую икру хранили в охлажденном состоянии (1,94°C ± 0,56°C) с отбором проб через 50, 80, 120, 210, 288 и 353 дня. Независимо от применяемой обработки, количество жизнеспособных аэробных микроорганизмов увеличилось до >6,1 log КОЕ/г через 120 дней хранения, и микробиота во всех вариантах обработки стала доминироваться Debaromyces hansenii, идентифицированной с помощью MALDI-TOF и метагеномного секвенирования. Добавление антиоксидантов снизило образование липидных гидропероксидов и веществ, реагирующих с тиобарбитуровой кислотой. Оба препарата, Herbalox и VivOx, снизили образование 1-пентен-3-ола, тогда как VivOx увеличил образование пентаналя в икре. Концентрации 2-метилбутанола и 3-метилбутанола были значительно снижены при обработках, включавших бензойную кислоту, что может быть связано с различиями в активности микробиоты. Herbalox был более эффективен, чем VivOx, в снижении окислительных изменений, тогда как влияние органических кислот на окислительные изменения осталось неопределенным. В заключение, можно снизить содержание соли и отказаться от использования бензойной кислоты без ущерба для микробиологической и окислительной стабильности СПП, а также его безопасности.
1 Введение
Икра пинагора (Cyclopterus lumpus) считается деликатесом во многих странах, а в Скандинавии существуют давние традиции производства различных соленых продуктов из икры пинагора (Basby et al., 1998a; Johannesson, 2006). Икра составляет примерно 25% веса самки рыбы в момент сбора. В Северной Атлантике ежегодно собирается от 2000 до 8000 тонн икры, причем основные производители расположены в Исландии, Гренландии, Норвегии и Канаде (Kennedy et al., 2019). Меньшие объемы свежей икры пинагора производятся в других странах, включая Данию и Швецию (Kennedy et al., 2019).
Основной сезон промысла икры пинагора в Северной Атлантике обычно ограничен несколькими месяцами весной (Johannesson, 2006; Kennedy et al., 2019). Распределение икры пинагора потребителям в течение всего года, следовательно, представляет собой проблему, в частности, потому что значительные количества икры пинагора выгружаются в регионах, где транспортировка производителям, например, в Европу и США, требует недель/месяцев. Эти логистические проблемы очень важны для Гренландии, где пинагора вылавливают вдоль более чем 2000-километровой береговой линии Западной Гренландии, но они также могут быть актуальны и для других икропроизводящих регионов, таких как Канада, Исландия и Норвегия. Свежая икра пинагора имеет короткий срок годности, и в местах выгрузки традиционно используется простой процесс, включающий смешивание икры с рассолами (состоящими из соли и органических кислот), для консервации (Johannesson, 2006). Этот охлажденный и соленый промежуточный продукт (СПП) с содержанием 10%–14% NaCl и 0,2%–0,4% бензоата натрия впоследствии транспортируется на фабрику, где хранится до 1 года до дальнейшей обработки (Рисунок 1). Например, производство готовой к употреблению «икры» пинагора включает обессоливание СПП, добавление нового рассола, красных или черных натуральных красителей, упаковку в стеклянные контейнеры и пастеризацию продукта для окончательной продажи в розничную сеть (Bledsoe et al., 2003; Kennedy et al., 2019). Ожидается, что в качестве сырья качественные характеристики СПП будут влиять на сенсорные свойства конечных икорных продуктов. Несмотря на ценность как СПП, так и конечных продуктов из икры пинагора, мы не нашли опубликованных исследований изменений качества СПП во время хранения.
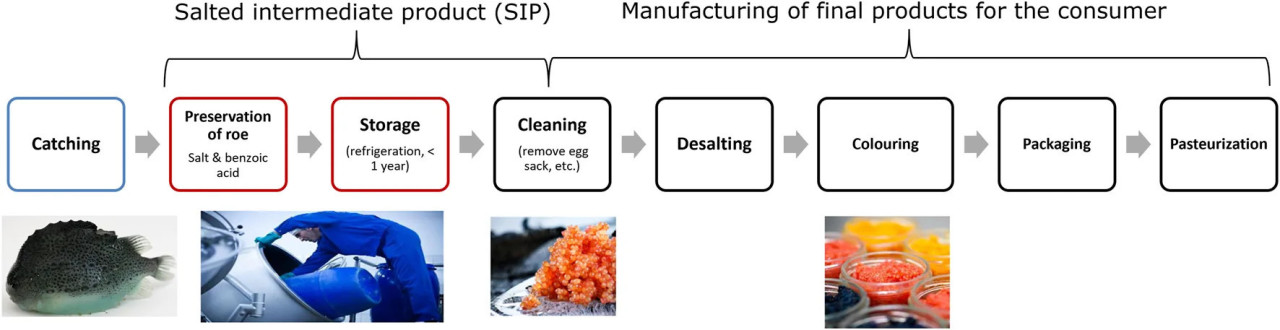
РИСУНОК 1 Иллюстрация вертикального производственного процесса икры пинагора от вылова в Гренландии до готового пастеризованного потребительского продукта.
Как микробиологические, так и окислительные изменения могут быть актуальны для изменения качества икры, консервированной традиционными составами СПП с высоким содержанием соли и бензойной кислоты, хотя это еще предстоит задокументировать. Более того, новые составы СПП с пониженным содержанием соли и бензойной кислоты желательны в соответствии с запросами заказчиков. Однако неизвестно, как они должны быть составлены, чтобы обеспечить достаточную сохранность в отношении микробиологических изменений. Поэтому требуются исследования способности других органических кислот сохранять СПП. СПП с пониженным содержанием соли по сравнению с традиционным рецептом также представляет интерес для потенциального сохранения розового цвета свежей икры, ограничения потери продукта за счет обезвоживания и снижения транспортировки соли в отдаленные регионы, например, Гренландию (Albarracín et al., 2011). Ранее сообщалось, что бензойная кислота повышает риск пищевой аллергии у чувствительных людей (Chipley, 2020), следовательно, замена бензойной кислоты в СПП снизит риск возникновения пищевой аллергии на икорные продукты.
Икра пинагора содержит примерно 4,5% (мас./мас.) липидов и высокий процент ненасыщенных жирных кислот (Basby et al., 1998b; Vasconi et al., 2020; Pountney et al., 2022). Эти жирные кислоты склонны к окислению, что может вызывать нежелательные изменения во время хранения, например, прогорклый запах и неприятный привкус. Ранее было показано, что экстракты розмарина для предотвращения окисления других рыбных продуктов эффективны для широкого разнообразия пищевых продуктов (Ozogul et al., 2010) и могут снижать окислительные изменения в СПП. Кроме того, экстракты розмарина могут обладать противомикробными свойствами в пищевых продуктах (Campo et al., 2000), потенциально обеспечивая двойной защитный эффект на икру пинагора. Насколько нам известно, действие экстрактов розмарина еще не исследовалось для соленой икры пинагора.
Цель настоящего исследования была направлена на изучение влияния рассолов с пониженным содержанием соли и различными комбинациями органических кислот и антиоксидантов на основе экстрактов розмарина на микробиологическую и окислительную стабильность СПП. Во-первых, в Гренландии был произведен СПП с пятью различными рассолами. Во-вторых, продукты из икры были транспортированы в Датский технический университет (DTU, Дания), химически охарактеризованы и хранились при +2°C в течение 1 года с отбором проб через регулярные интервалы для оценки микробиологических и окислительных изменений.
2 Материалы и методы
2.1 Икра пинагора, ингредиенты рассола и обработка икры
Пинагор (Cyclopterus lumpus) был выловлен у берегов Паамиута, Гренландия (ФАО 21, 1E) в апреле 2022 года. Икра пинагора была вручную отделена от тушки, и оболочки икры были удалены. Это было сделано для облегчения смешивания икры с рассолами, что является несколько иной процедурой, чем промышленная обработка (Рисунок 1). Свежая икра хранилась в охлажденном состоянии (3,5°C) до 4 часов до дальнейшей обработки.
Было изучено пять способов обработки для производства СПП с использованием различных составов рассолов (Таблица 1). Рассолы были приготовлены в соответствии с рецептами с использованием различных концентраций соли, бензойной кислоты (бензоат натрия, BAC, Sigma-Aldrich, STL, США), уксусной кислоты (ацетат натрия, AAC, Macco Organiques, Валлифилд, Квебек, Канада) и молочной кислоты (LAC, Purac Powder 60, Corbion, Нидерланды). Кроме того, для трех способов обработки один из двух антиоксидантов, а именно Herbalox® D-450 (0,019%, HER, Kalsec Inc., Калифорния, США), содержащий экстракт розмарина (Rosmarinus officinalis), и VivOx 7.5 (0,046%, VIV, IFF, Нью-Йорк, США), содержащий смесь экстракта розмарина (Rosmarinus officinalis) и мальтодекстрина, был также добавлен в рассолы.
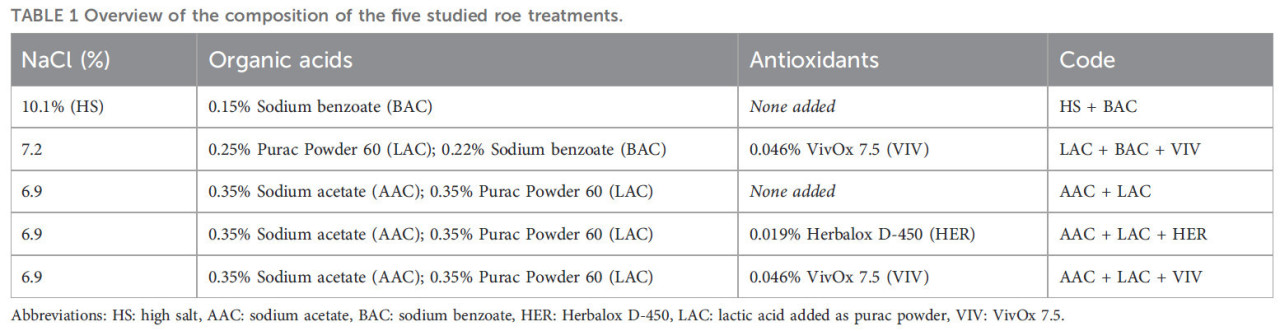
ТАБЛИЦА 1 Обзор состава пяти изученных способов обработки икры.
Ингредиенты, используемые для каждого способа обработки (Таблица 1), были взвешены, добавлены к икре и смешаны с водопроводной водой. Используемая водопроводная вода поступает с муниципальной водоочистной станции, управляемой Гренландской коммунальной компанией (Nukissiorfiit). Вода очищается с помощью песочного фильтра и УФ-обработки, и ее химическое и микробиологическое качество регулярно проверяется для обеспечения соответствия гренландским нормам питьевой воды. Для каждого способа обработки два полиэтиленовых ведра пищевого качества были наполнены 10 кг икры и 2 л рассола для окончательного хранения. В каждое ведро была добавлена сетка для погружения икры в рассол во время хранения. Антиоксиданты были добавлены таким образом, чтобы содержание карнозовой кислоты и карнозола было одинаковым для всех обработок, содержащих антиоксиданты (LAC + BAC + VIV, AAC + LAC + HER и AAC + LAC + VIV).
2.2 Хранение и отбор проб
Ведра с соленой в рассоле икрой были транспортированы в охлажденном состоянии (3,5°C ± 0,9°C) в DTU (Конгенс Люнгбю, Дания) и хранились при температуре 1,94°C ± 0,56°C. Пробы отбирались из каждого ведра для дальнейшего анализа на 50, 80, 120, 210, 288 и 353 день после первоначального процесса засолки в рассоле в Гренландии. Микробиологический подсчет проводился в день отбора проб, тогда как образцы для химического анализа хранились при -80°C до дальнейшего анализа.
2.3 Характеристики продукта
Содержание сухого вещества (СВ) и pH были измерены, как описано Dalgaard et al. (1993). Содержание соли было определено с использованием автоматического потенциометрического титрования (785 DMP Titrino titrator, Methrom, Херизау, Швейцария) (метод AOAC 976.18 (AOAC, 2000), метод AOAC 937.07 (AOAC, 1996) и метод AOAC 971.27 (AOAC, 1976)). Органические кислоты были определены с использованием ВЭЖХ-ДМД (Agilent 1,200 series, Agilent Technologies, Калифорния, США), как описано Dalgaard и Jørgensen (2000). Внешние стандарты были использованы для идентификации и количественного определения бензойной и молочной кислот. Ферментный анализ уксусной кислоты (Megazyme, Уиклоу, Ирландия) был использован для количественного определения концентрации уксусной кислоты. Все анализы были проведены в биологических дубликатах с техническими повторами (n = 2 × 2).
2.4 Микробиологический анализ
2.4.1 Микробиологические изменения
Во время отбора проб 20 г икры были асептически извлечены из каждого пробного ведра и разведены в 10 раз в охлажденном (5°C) пептонном солевом буферном растворе (PS, 0,85% NaCl и 0,1% пептона) и гомогенизированы в течение 60 с в Stomacher 400 (Seward Medical, Лондон, Великобритания). Дальнейшие соответствующие 10-кратные разведения гомогенатов были сделаны в охлажденном PS. Количество жизнеспособных аэробных микроорганизмов (AVC) было определено методом поверхностного посева (15°C, 7 дней) на агар Long и Hammer (LH) с 1% NaCl (NMKL, 2006). Молочнокислые бактерии (LAB) были подсчитаны методом глубинного посева (25°C, 3 дня) в нитрит-актидион-полимиксиновый агар (NAP) с pH 6,2 (Davidson и Cronin, 1973). Дрожжи и плесени были подсчитаны методом поверхностного посева (25°C, 3 дня) на агар Oxytetracycline Glucose Yeast Extract (OGYE) (CM0545, Oxoid, Бейзингсток, Великобритания) с коммерческой добавкой окситетрациклина (SR0073A, Oxoid). Предположительные Pseudomonas были подсчитаны методом поверхностного посева (25°C, 2 дня) на агар CFC/Pseudomonas (CM0559, Oxoid) с селективной добавкой для Pseudomonas (SR0103, Oxoid). Все анализы были проведены в виде единичных определений биологических дубликатов (n = 2), и результаты были представлены как log КОЕ/г икры.
2.4.2 Идентификация бактериальных изолятов с использованием матрично-активированной лазерной десорбции/ионизации — времяпролетной масс-спектрометрии (MALDI-TOF)
Для идентификации доминирующей микробиоты в каждой обработке СПП после 50, 120 и 353 дней хранения были изолированы репрезентативные колонии на чашках с агаром LH. Для каждой из пяти обработок и в каждый из трех моментов отбора проб было изолировано восемь колоний. Однако больше изолятов отбиралось, когда колонии казались более разнообразными. Колонии выращивали в бульоне BHI (CM1135B, Oxoid) и чисто культивировали с использованием LH, оба инкубировали при 15°C в течение 3–7 дней. Белки из колоний на свежих агаровых пластинах были экстрагированы с помощью протокола этанол/муравьиная кислота/ацетонитрил, используемого Nonnemann et al. (2019), как описано Bizzini et al. (2010). Масс-спектры были получены с помощью матрично-активированной лазерной десорбции/ионизации времяпролетной масс-спектрометрии (MALDI-TOF) с использованием прибора Maldi-Biotyper® Sirius one RUO Instrument (Bruker Daltonics, Биллерика, Массачусетс, США) с библиотекой MBT Compass. Лог-баллы от 0 до 3 были рассчитаны алгоритмом Biotyper (Bruker Daltonics, Бремен, Германия). Лог-баллы <1,7 не приводят к идентификации. Лог-баллы 1,7 ≤ x < 2,0 обеспечивают идентификацию рода, тогда как лог-баллы ≥2,0 обеспечивают идентификацию вида. Все изоляты были проанализированы в трех повторностях.
2.4.3 Независимый от культивирования анализ образцов СПП с использованием метагеномного дробовичного секвенирования
ДНК была экстрагирована из дублированных образцов СПП от каждой из пяти обработок на 50 и 353 день эксперимента по хранению, используя 10-1 разведение в мешках для Stomacher (см. раздел 2.4.1), которые хранились замороженными (-80°C) после микробиологического анализа. Короче говоря, аликвоты по 10 мл из каждого образца были центрифугированы при 8 960 g при 5°C в течение 10 минут для получения осадка, содержащего микробные клетки. ДНК была экстрагирована из осадка с помощью набора DNeasy PowerFood (Qiagen, Хильден, Германия) в соответствии с протоколом производителя. Экстрагированная ДНК была количественно определена с использованием Qubit 3.0 (Invitrogen, Карлсбад, Калифорния, США) с набором Qubit DNA HS Assay Kit (Invitrogen). Образцы ДНК были отправлены со льдом в Eurofins Genomics (Констанц, Германия) для контроля качества и метагеномного дробовичного секвенирования Illumina с парными концами по 150 п.н. Один образец (День 50, AAC + LAC) не был секвенирован. Сырые метагеномные данные были переданы в Европейский архив нуклеотидов (ENA) под номером проекта: PRJEB78984. Сырые данные секвенирования 19 образцов были обработаны с помощью CLC Genomics Workbench 24.0. (Qiagen, Орхус, Дания) и CLC Genomics Microbial Module 24.0 для удаления адаптеров и низкокачественных ридов с показателями качества ниже 0,05. После обрезки для каждого образца осталось от 22,6 до 37,4 миллионов ридов, с медианой 26,9 миллиона ридов/образец. Для таксономического профилирования была загружена база данных, содержащая все геномы Fungal NCBI RefSeq (RefSeq Fungal, 584 генома, 30 января 2024 г., ncbi.nlm.nih.gov/refseq/), вместе с QIAGEN Microbial Insights–Prokaryotic Taxonomy Database Family (QMI-PTDB, версия 2.0). Последняя курируется и основана на референсных геномах с представлением всех прокариотических семейств на основе аннотаций из Базы данных таксономии геномов (GTDB, gtdp.ecogenomic.org). Обе базы данных RefSeq Fungal и QMI-PTDB family были использованы для создания комбинированного референсного индекса таксономического профилирования, который был применен для таксономического профилирования в CLC Genomics каждого образца с минимальной длиной семени 30 п.н. и с поправкой на вариацию длины ридов. Таксономическая классификация привела к получению от 0,4 до 20,3 миллиона назначенных ридов для каждого образца с медианой 6,6 миллиона классифицированных ридов и в общей сложности 153,4 миллиона назначенных ридов. Результаты таксономической классификации были отфильтрованы для удаления шума секвенирования и видов с чрезвычайно низкой обильностью путем удаления таксонов с менее чем 1000 классифицированных ридов в совокупности по всем 19 образцам. Отфильтрованные таблицы таксономической обильности были объединены, экспортированы и использованы в качестве входных данных для таксономических визуализаций в GraphPad Prism 10 (GraphPad Software, Бостон, Массачусетс, США).
2.5 Химический анализ
Все анализы образцов икры (пять обработок, шесть временных точек) были проведены в биологических дубликатах с аналитическими повторами (n = 2 × 2), за исключением анализа летучих продуктов окисления, которые были проведены в трипликатах (n = 2 × 3). Если были какие-либо исключения, они отмечены в описании метода.
2.5.1 Экстракция липидов
Липиды экстрагировали с использованием хлороформа и метанола согласно методу Bligh и Dyer (B&D) с уменьшенным количеством растворителя (Bligh и Dyer, 1959; Iverson et al., 2001). После экстракции масла и выпаривания хлороформа содержание липидов определяли гравиметрически. Результаты представлены в процентах от веса образца.
2.5.2 Определение свободных жирных кислот (FFA)
FFA в липидных экстрактах титровали NaOH с использованием фенолфталеина в качестве индикатора в соответствии с методом AOCS Ca 5a-40 (AOCS, 2004). Аликвоты 10–15 г липидного экстракта смешивали с этанолом (25 мл) и добавляли несколько капель индикатора. Затем образец титровали 0,1M NaOH до появления бледно-розового цвета. Объем NaOH, использованный для титрования, использовали для расчета содержания FFA (%). Результаты представлены как количество FFA (%) по содержанию олеиновой кислоты в масле.
2.5.3 Определение содержания токоферолов
Содержание токоферолов определяли в соответствии с методом AOCS Ce 8–89 (AOCS, 1998). Приблизительно 1 г липидного экстракта выпаривали досуха под азотом и повторно растворяли в гептане (1 мл). Экстракт анализировали с помощью ВЭЖХ-ФЛД (Agilent 1,100 series, Agilent Technologies), как описано Lu et al. (2015). Результаты были количественно определены как мкг/г образца.
2.5.4 Содержание карнозовой кислоты и карнозола
Основными антиоксидантными соединениями в экстрактах розмарина были фенольные соединения карнозовая кислота и карнозол. Образцы анализировали на содержание карнозовой кислоты и карнозола на 50 и 353 дни. Икру (0,5 г) отвешивали в центрифужную пробирку и добавляли 2,5 мл растворителя (ацетон или этилацетат). Образец встряхивали (20 с) и обрабатывали ультразвуком в течение 30 минут. Образец центрифугировали (3 500 об/мин, ≈2 602 × g, 10 минут), верхнюю фазу переносили в другую центрифужную пробирку и растворитель выпаривали. Процедуру повторяли дважды, и объединенную выпаренную верхнюю фазу повторно растворяли в 1 мл MeOH с изоаскорбиновой кислотой (1 мг/мл) и фильтровали (0,2 мкм). Экстракцию проводили в дубликатах (n = 2). Поток на ВЭЖХ-ДМД (Agilent 1,200 series, Agilent Technologies) составлял 0,9 мл, и применялся градиентный метод с двумя растворителями, A) Вода (pH 3, доведенный фосфорной кислотой) и B) Ацетонитрил. Градиентный метод был следующим: 0–20 мин 5% Растворителя B, 20–25 мин 100% Растворителя B, 25–27 мин 40% Растворителя B. Для разделения использовали колонку Zorbax ® Eclipse XDB-C8 (5 мкм 4,6 × 150 мм, Agilent Technologies). Соединения детектировали при 280 нм на УФ-детекторе. Для количественного определения карнозовой кислоты и карнозола была приготовлена калибровочная кривая в MeOH, и результаты количественно определены как мкг/г образца.
2.5.5 Определение перекисного числа (PV)
PV определяли в липидных экстрактах с использованием колориметрического ферритиоцианатного метода и измеряли спектрофотометрически при 500 нм, как описано Shantha и Decker (1994), с небольшими модификациями, поскольку объем растворителя был увеличен на 2%. Калибровочная кривая была приготовлена с хлоридом железа, и PV было количественно определено как мэкв. ROOH/кг масла.
2.5.6 Определение веществ, реагирующих с тиобарбитуровой кислотой (TBARS)
Образцы анализировали на изменения TBARS в течение периода хранения в соответствии с Vyncke (1970). Аликвоты по 15 г образцов смешивали с 30 мл 7,5% трихлоруксусной кислоты с использованием Ultra Thurrax (10 000 об/мин, 15 с) и фильтровали. 5 мл фильтрованного экстракта переносили в две пробирки. 5 мл 0,02M 2-тиобарбитуровой кислоты добавляли в одну пробирку, тогда как 5 мл воды добавляли в другую пробирку в качестве холостой пробы. Затем пробирки нагревали (90°C, 40 мин) на водяной бане и охлаждали в воде. Поглощение образцов измеряли спектроскопически при 530 нм (Shimadzu UV-1280, Holm и Halby, Брённбю, Дания). Внешняя калибровочная кривая была выполнена с использованием 1,1,3,3-тетраэтоксипропана (TEP, малоновый диальдегид бис(диэтилацеталь)), растворенного в 7,5% трихлоруксусной кислоте (TCA), в качестве стандарта для расчета концентрации TBARS. Результаты были количественно определены как мкмоль/кг образца.
2.5.7 Определение летучих соединений методом динамической парофазной ГХ-МС
Приблизительно 10 г икры отвешивали и смешивали с 25 мл воды и 30 мг внутреннего стандартного раствора (30 мкг/г 4-метил-1-пентанола в рапсовом масле). Образцы продували в течение 30 минут при 37°C с потоком азота 340 мл/мин, и летучие соединения собирали на трубках Tenax® GR. После сбора трубки Tenax® продували потоком азота 50 мл/мин в течение 20 минут. Собранные летучие вещества десорбировали с трубок Tenax® с помощью автоматического термодесорбера (ATD Turbomatrix 450, PerkinElmer, Норуолк, Коннектикут, США) при 230°C в сочетании с ГХ Agilent 6,890 (Пало-Альто, Калифорния, США), соединенным с масс-спектрометром (HP 5973 Network Mass Selective Detector, Agilent Technologies). Начальная температура печи ГХ составляла 55°C в течение 1,5 мин, с увеличением на 2,0°C/мин до 90°C, затем увеличением на 8,0°C/мин до 230°C, при которой температуру поддерживали в течение 8 мин. Условия МС были: режим электронной ионизации, 70 эВ, сканирование отношения массы к заряду от 30 до 250). Внешние стандарты (2-этилфуран, пентаналь, 1-пентен-3-ол, 3-метилбутанол, 2-метилбутанол, 2-пентеналь, гексеналь, 2-пентен-2-ол, 2-гексеналь, 4-гептаналь, 1-октен-3-ол, октаналь, 2,4-гептадиеналь, нонаналь и 2,6-нонадиеналь (Sigma Aldrich, Миссури, США)) использовали для калибровочных кривых. Стандарты растворяли в этаноле, наносили на трубки Tenax® в концентрациях от 5 до 1500 нг/г и анализировали тем же методом, что описан выше. Результаты представлены в нг/г икры. LOQ был определен как самая низкая концентрация, использованная для стандартной кривой.
2.6 Анализ данных
Графические представления и статистический анализ были выполнены с использованием программного обеспечения GraphPad Prism 10. Анализ увеличения микробных концентраций, PV и TBARS был выполнен с использованием встроенного инструмента для простой линейной регрессии, включая F-отношение и P-значение, чтобы проверить, был ли наклон значительно отличен от нуля. В случае наклона, значительно отличающегося от нуля, среднее значение анализировали с помощью двухфакторного дисперсионного анализа (ANOVA), используя post hoc анализ в форме t-теста Бонферрони, чтобы проверить, какие обработки были значительно (p < 0,05) различными во времени. Корреляции между микробными концентрациями, FFA, PV, TBARS и концентрациями летучих соединений были исследованы с помощью анализа главных компонент (PCA) с использованием RStudio версии 2023.03.1 + 446 (R-Core Team, 2023).
3 Результаты
3.1 Характеристики продукта
Как и ожидалось, в образцах икры из обработок LAC + BAC + VIV, AAC + LAC, AAC + LAC + HER и AAC + LAC + VIV были достигнуты сходные значения pH и содержания соли. Более того, сходное содержание уксусной кислоты и молочной кислоты было обнаружено для икры в обработках AAC + LAC, AAC + LAC + HER и AAC + LAC + VIV (Таблица 2).
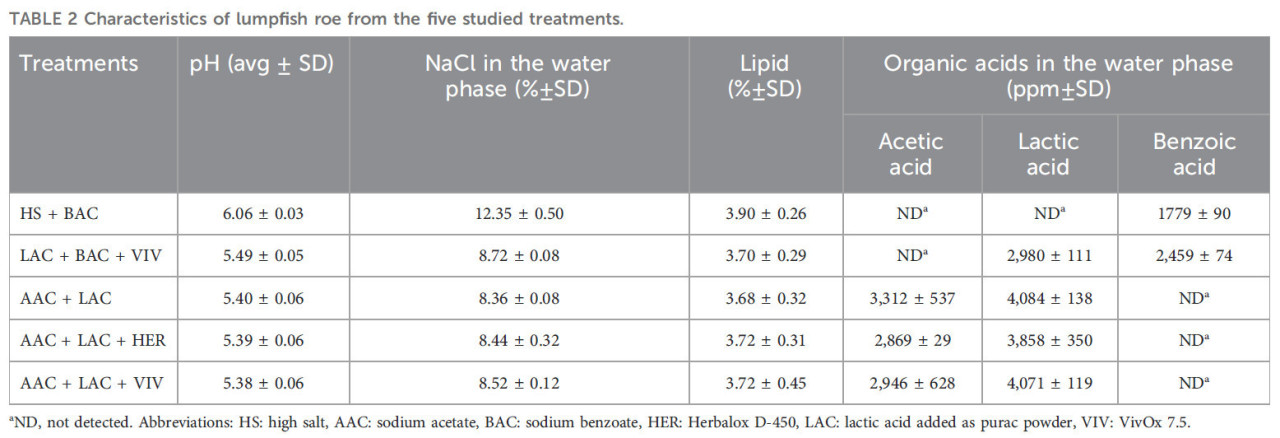
ТАБЛИЦА 2 Характеристики икры пинагора из пяти изученных обработок.
3.2 Микробиологические изменения и идентификация микробиоты
AVC увеличилось с 5,5-6 log КОЕ/г на 50 день до 6,1–6,5 log КОЕ/г после 120 дней хранения и оставалось довольно стабильным в течение оставшегося периода хранения для всех обработок (Рисунок 2A). Значительные (p < 0,05) различия между концентрациями AVC и дрожжей и плесеней наблюдались только на 50 день для обработки HS + BAC и на 288 и 353 дни для обработки AAC + LAC + HER с более высокими концентрациями AVC, что указывает на то, что микробиота была доминирована дрожжами и плесенями (Рисунки 2A, B). После 210 дней хранения значительно (p < 0,05) более высокие концентрации дрожжей и плесеней наблюдались в HS + BAC по сравнению с образцами икры из обработок с уксусной и молочной кислотами (AAC + LAC, AAC + LAC + HER и AAC + LAC + VIV). Не было обнаружено значительного (p > 0,05) увеличения роста на среде CFC для любой из пяти обработок, и средняя концентрация клеток составила 4,76 ± 0,68 log КОЕ/г (Рисунок 2C). Концентрации молочнокислых бактерий на 50 день были значительно (p < 0,05) выше в икре из обработок без бензойной кислоты (4,4–5,4 log КОЕ/г, AAC + LAC, AAC + LAC + HER и AAC + LAC + VIV), чем в икре из обработок с бензойной кислотой (1-2 log КОЕ/г, HS + BAC и LAC + BAC + VIV). Между 50 и 353 днями значительный (p < 0,05) рост молочнокислых бактерий наблюдался исключительно в икре из обработки HS + BAC с увеличением на 1,17 log КОЕ/г (Рисунок 2D).
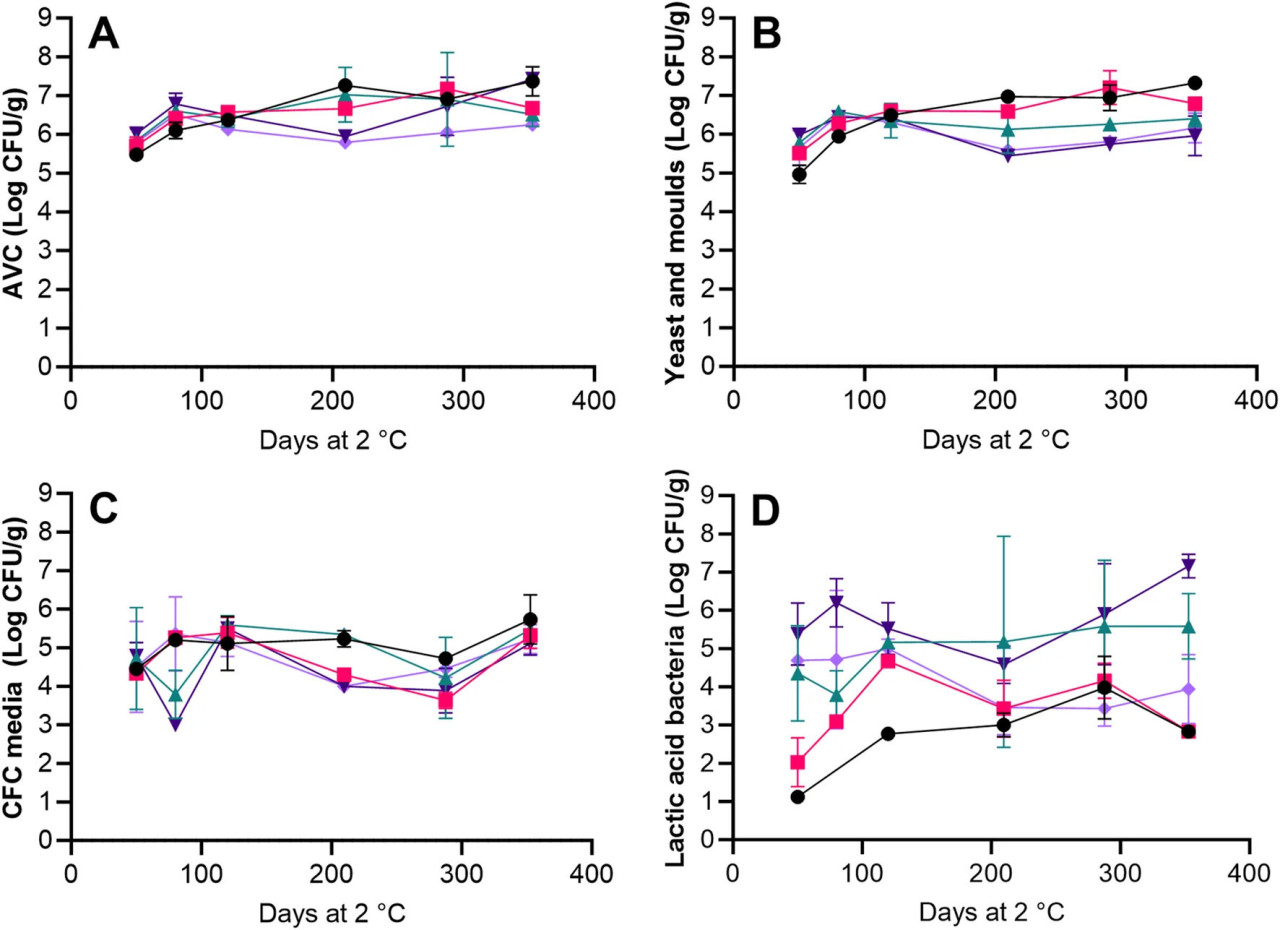

РИСУНОК 2 Микробиологические концентрации образцов икры во время холодильного хранения. AVC (A), определенные на агаре Long и Hammer, концентрации дрожжей и плесеней (B), определенные на агаре OYGE, концентрации предполагаемых Pseudomonas spp. на агаре CFC (C) и концентрации молочнокислых бактерий (D), определенные на агаре NAP, для обработок HS + BAC (●) LAC + BAC + VIV ( ), AAC + LAC ( ), AAC + LAC + HER ( ) и AAC + LAC + VIV ( ). Символы и планки погрешностей указывают среднее значение ± СО.
Для всех обработок микробиота икры, наблюдаемая на 50 день и идентифицированная с помощью MALDI-TOF, была доминирована дрожжами (Debaryomyces hansenii и Candida zeylanoides) и неидентифицированными микроорганизмами, как показано на примере обработки HS + BAC, где 75% микробиоты на 50 день было неидентифицировано, а остальные 25% состояли из D. hansenii (Таблица 3). После 353 дней хранения микробиота обработок HS + BAC и LAC + BAC + VIV была доминирована D. hansenii (>88% микробиоты), тогда как AAC + LAC и AAC + LAC + VIV имели более разнообразную микробиоту, включая виды Psychrobacter и Latilactobacillus. Икра из обработки AAC + LAC + HER была доминирована D. hansenii, но также содержала C. zeylanoides и Psychrobacter (Таблица 3).
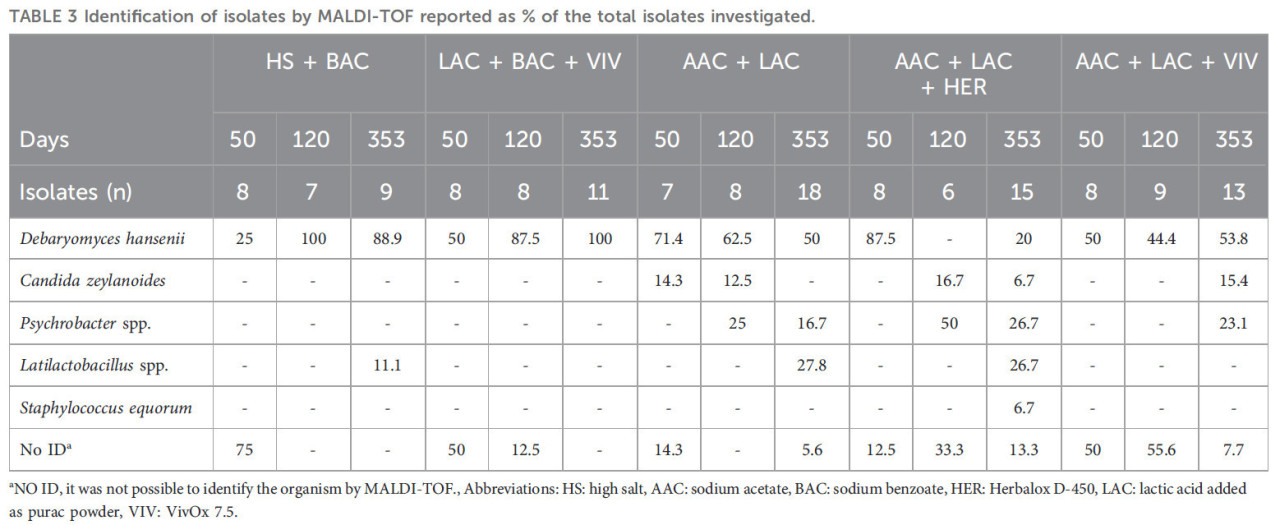
ТАБЛИЦА 3 Идентификация изолятов с помощью MALDI-TOF, представленная как % от общего числа исследованных изолятов.
Сходные результаты были получены с помощью метагеномного секвенирования, однако с еще более выраженным доминированием D. hansenii во всех обработках во время хранения (Рисунок 3). В целом, на 50 день наблюдалось более высокое разнообразие с различными Ascomycota spp., тогда как D. hansenii составляла 96% ± 3% от метагеномной обильности в образцах на 353 день (Вспомогательная таблица S1). Бактерии обычно обнаруживались в низкой обильности, составляя только 1,5% ± 1,7% метагенома. Только два бактериальных семейства были обнаружены с относительной обильностью >1%: Catellicoccaceae, обнаруженные исключительно в икре из обработки HS + BAC через 50 дней, и Lactobacillaceae, имеющие самую высокую обильность (2%–5%) в образцах икры из обработок AAC + LAC и AAC + LAC + HER через 353 дня (Рисунок 3). Хотя это не отражает обильность, обнаруженную с помощью идентификации MALDI-TOF, последовательности Psychrobacter spp. были найдены только в образцах икры, сохраненных с обработками, содержащими AAC, через 353 дня (данные не показаны). Аналогично, C. zeylanoides, принадлежащая к классу Saccharomycetes, была обнаружена во всех метагеномных образцах на 50 и 353 дни, хотя и с обильностью ниже 0,01% (данные не показаны).
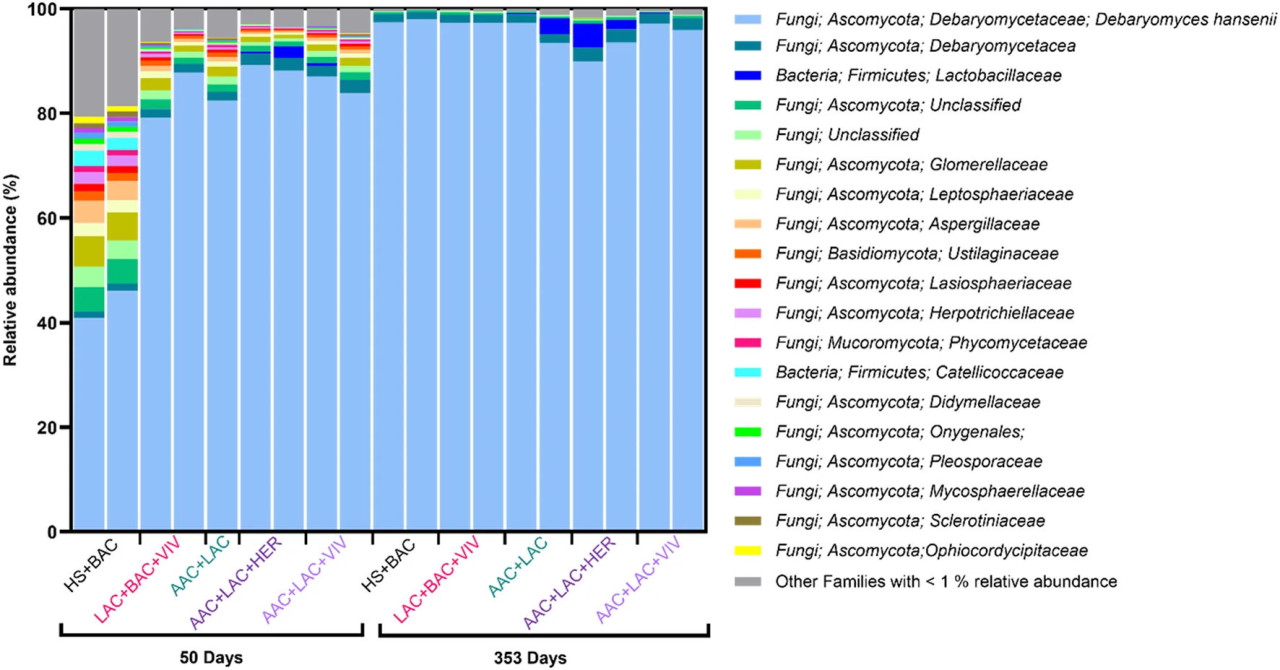
РИСУНОК 3 Состав микробиома обработанной рассолом икры пинагора, определенный с помощью метагеномного секвенирования образцов икры (n = 2 для каждой из пяти обработок) после 50 и 353 дней хранения. Один образец икры (день 50, AAC + LAC) не удалось секвенировать. Исключительно таксоны, присутствующие с относительной обильностью не менее 1% в одном образце, показаны в легенде.
3.3 Химические изменения
Содержание свободных жирных кислот (FFA) увеличилось на 8%–10% во время хранения икры из всех обработок, хотя более низкое содержание FFA наблюдалось в икре из обработки HS + BAC на протяжении всего периода хранения (Рисунок 4).
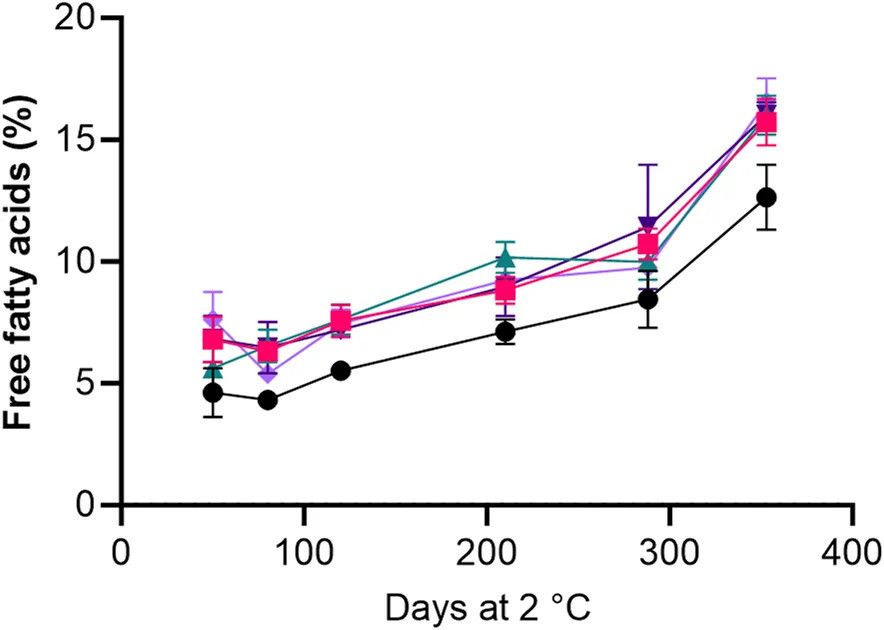

РИСУНОК 4 Влияние холодильного хранения на содержание % свободных жирных кислот (FFA) в образцах икры обработок HS + BAC (●) LAC + BAC + VIV ( ), AAC + LAC ( ), AAC + LAC + HER ( ) и AAC + LAC + VIV ( ). Символы и планки погрешностей указывают среднее значение ± СО.
Токоферолы обладают антиоксидантными свойствами и действуют либо как поглотители свободных радикалов, либо реагируя с синглетным кислородом, следовательно, снижение содержания токоферолов часто наблюдается в окисленных продуктах (Pazos et al., 2005). Из четырех проанализированных гомологов токоферолов был обнаружен только α-токоферол, и более высокие концентрации были найдены в икре из обработок с добавленными антиоксидантами (∼12,7 мкг/г) по сравнению с обработками без антиоксидантов (∼7,2 мкг/г). Значительного (p > 0,05) потребления α-токоферола во время хранения не наблюдалось ни для одной из пяти обработок (данные не показаны).
Сходное содержание карнозовой кислоты и карнозола было обнаружено в образцах икры, обработанных антиоксидантами. Содержание карнозовой кислоты снизилось, хотя и не значительно (p > 0,05) для LAC + BAC + VIV и AAC + LAC + HER во время хранения (снижение на 0,65 мкг/г). Значительное (p < 0,05) снижение на 1,29 мкг/г наблюдалось для AAC + LAC + VIV (Рисунок 5A). Во время хранения начальное содержание карнозола в LAC + BAC + VIV снизилось до неопределяемых уровней, тогда как начальное содержание в AAC + LAC + HER и AAC + LAC + VIV значительно (p < 0,05) снизилось во время хранения на 2,83 и 3,97 мкг/г, соответственно (Рисунок 5B). Как и ожидалось, ни карнозовая кислота, ни карнозол не были обнаружены в икре из обработок HS + BAC или AAC + LAC (данные не показаны).

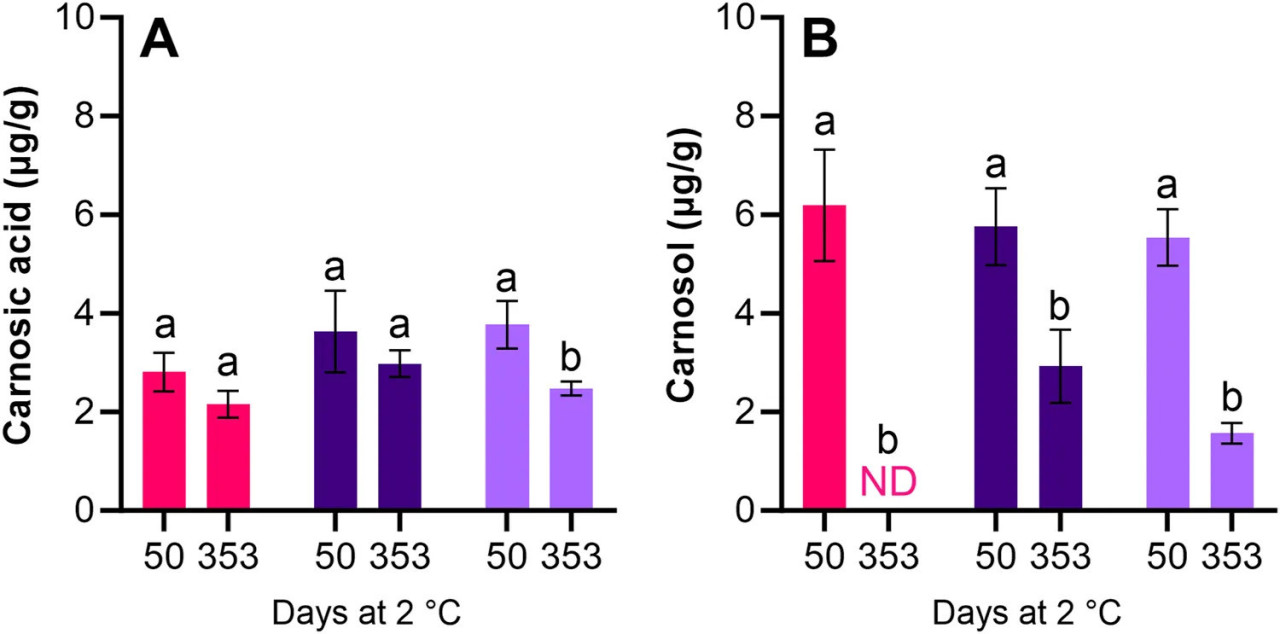
РИСУНОК 5 Содержание карнозовой кислоты (A) и карнозола (B) в образцах икры обработок LAC + BAC + VIV ( ), AAC + LAC + HER ( ) и AAC + LAC + VIV ( ) после 50 и 353 дней холодильного хранения. HS + BAC и AAC + LAC были проанализированы, но соединения не были обнаружены. ND, измерялось, но не обнаружено. Разные буквы над столбцами одного цвета указывают на значительные различия, т.е. значительное снижение во времени в пределах конкретной обработки.
Продукты первичного окисления можно измерить с помощью определений PV, которое измеряет количество липидных гидропероксидов в образце. Обработки с антиоксидантами (LAC + BAC + VIV, AAC + LAC + HER и AAC + LAC + VIV) показали более низкие начальные PV (<2,04 мэкв. ROOH/кг масла) в образцах икры по сравнению с обработками без добавления антиоксидантов (>4 мэкв. ROOH/кг масла, HS + BAC и AAC + LAC; Рисунок 6A). Значительные (p < 0,05) увеличения наблюдались в течение 288 дней хранения икры из всех обработок, за исключением AAC + LAC, что связано с более высокими уровнями PV на 80 и 120 дни. Важно отметить, что наименьшее увеличение во времени наблюдалось в икре из обработки AAC + LAC + HER, которое увеличилось только с 0,99 мэкв. ROOH/кг масла до 2,79 мэкв. ROOH/кг масла после 210 дней хранения (Рисунок 6A). PV остальных четырех обработок увеличились до 7–8 мэкв ROOH/кг масла после 210 дней хранения. Это указывает на то, что добавление Herbalox было более эффективным в подавлении образования гидропероксидов по сравнению с VivOx 7.5. Значительные (p < 0,05) снижения PV наблюдались после 288 дней хранения для образцов, содержащих уксусную и молочную кислоты (AAC + LAC, AAC + LAC + VIV и AAC + LAC + HER), причем AAC + LAC показал наибольшее снижение на 6,3 мэкв. ROOH/кг масла.

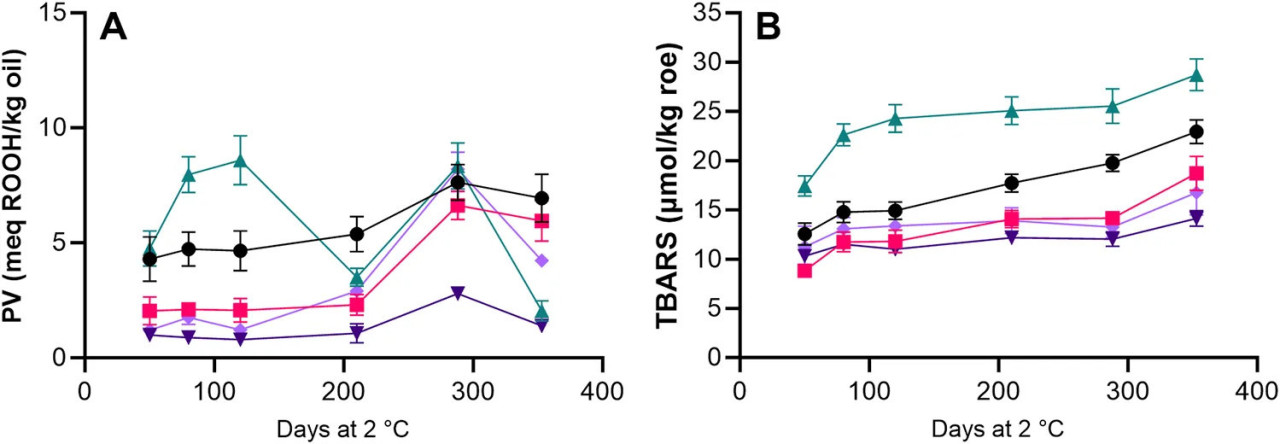
РИСУНОК 6 PV (A) и TBARS (B) в образцах икры обработок HS + BAC (●) LAC + BAC + VIV ( ), AAC + LAC ( ), AAC + LAC + HER ( ) и AAC + LAC + VIV ( ) во время холодильного хранения. Символы и планки погрешностей указывают среднее значение ± СО.
Продукты вторичного окисления можно измерить с помощью TBARS. Значительное (p < 0,05) увеличение TBARS во времени наблюдалось в икре из всех обработок. TBARS были значительно (p < 0,05) выше для AAC + LAC на протяжении всего хранения по сравнению с другими обработками, хотя TBARS в образцах икры HS + BAC также имели тенденцию быть выше, чем TBARS образцов икры из обработок с антиоксидантами. Значительно (p < 0,05) более низкие TBARS наблюдались после 220 дней хранения для образцов икры с добавленными антиоксидантами (LAC + BAC + VIV, AAC + LAC + HER и AAC + LAC + VIV) по сравнению с образцами икры без добавленных антиоксидантов (HS + BAC и AAC + LAC; Рисунок 6B).
Продукты вторичного окисления также можно измерить с помощью анализа летучих продуктов окисления. Количественное определение летучих соединений с помощью ГХ-МС дает более нюансированное понимание вторичного окисления, происходящего во время хранения, по сравнению с TBARS, поскольку образование конкретных летучих соединений часто может быть связано с окислением их предшественников, в основном жирных кислот. Всего было количественно определено 15 летучих соединений, но только 2-этилфуран, 2-пентен-2-ол, 1-пентен-3-ол, 3-метилбутанол, 2-метилбутанол, пентаналь и нонаналь имели концентрации выше LOQ (приблизительно 10 нг/г) метода или увеличились до выше LOQ в течение периода хранения.
Содержание пентаналя (Рисунок 7A) в икре увеличилось выше LOQ для обработок LAC + BAC + VIV и AAC + LAC + VIV после 288 дней хранения, тогда как содержание в икре из обработок HS + BAC, AAC + LAC и AAC + LAC + HER оставалось ниже LOQ на протяжении всего хранения. Содержание 1-пентен-3-ола (Рисунок 7B) увеличилось до более чем 400 нг/г в икре из обработки HS + BAC и до более чем 200 нг/г в икре из AAC + LAC, тогда как содержание в образцах икры с антиоксидантами (LAC + BAC + VIV, AAC + LAC + HER и AAC + LAC + VIV) оставалось на уровне или ниже 100 нг/г на протяжении всего периода хранения, с самым высоким конечным содержанием, наблюдаемым в LAC + BAC + VIV (123,4 ± 8,4 нг/г), и самым низким в AAC + LAC + HER (57 ± 6,7 нг/г). Содержание 2-метилбутанола (Рисунок 7C) и 3-метилбутанола (Рисунок 7D) увеличивалось во время хранения для образцов икры, приготовленных в рассолах, содержащих как уксусную, так и молочную кислоты (AAC + LAC, AAC + LAC + HER и AAC + LAC + VIV), тогда как образцы икры, консервированные бензойной кислотой (HS + BAC и LAC + BAC + VIV), показали значительно (p < 0,05) более низкие увеличения во время хранения. Из оставшихся летучих соединений с концентрациями выше LOQ содержание 2-этилфурана увеличилось до ∼95 нг/г в икре из AAC + LAC и до 38 нг/г в HS + BAC, тогда как образцы икры с антиоксидантами оставались ниже 25 нг/г на протяжении всего хранения, что снова показывает эффект добавленных антиоксидантов и более высокого содержания соли. 2-пентен-2-ол показал наибольшее увеличение во время хранения икры в HS + BAC с конечным содержанием ∼18 нг/г, AAC + LAC увеличился до ∼11 нг/г, тогда как образцы икры, приготовленные с антиоксидантами, оставались на уровне 5–7 нг/г. Содержание нонаналя не показало последовательной тенденции, колеблясь как с увеличением, так и с уменьшением в образцах икры на протяжении периода хранения. Снижение PV в образцах икры после 288 дней хранения, как показано на Рисунке 6A, не могло быть объяснено изменениями продуктов вторичного окисления, определяемыми изменениями TBARS (Рисунок 6B) или летучих соединений (Рисунки 7A–D).
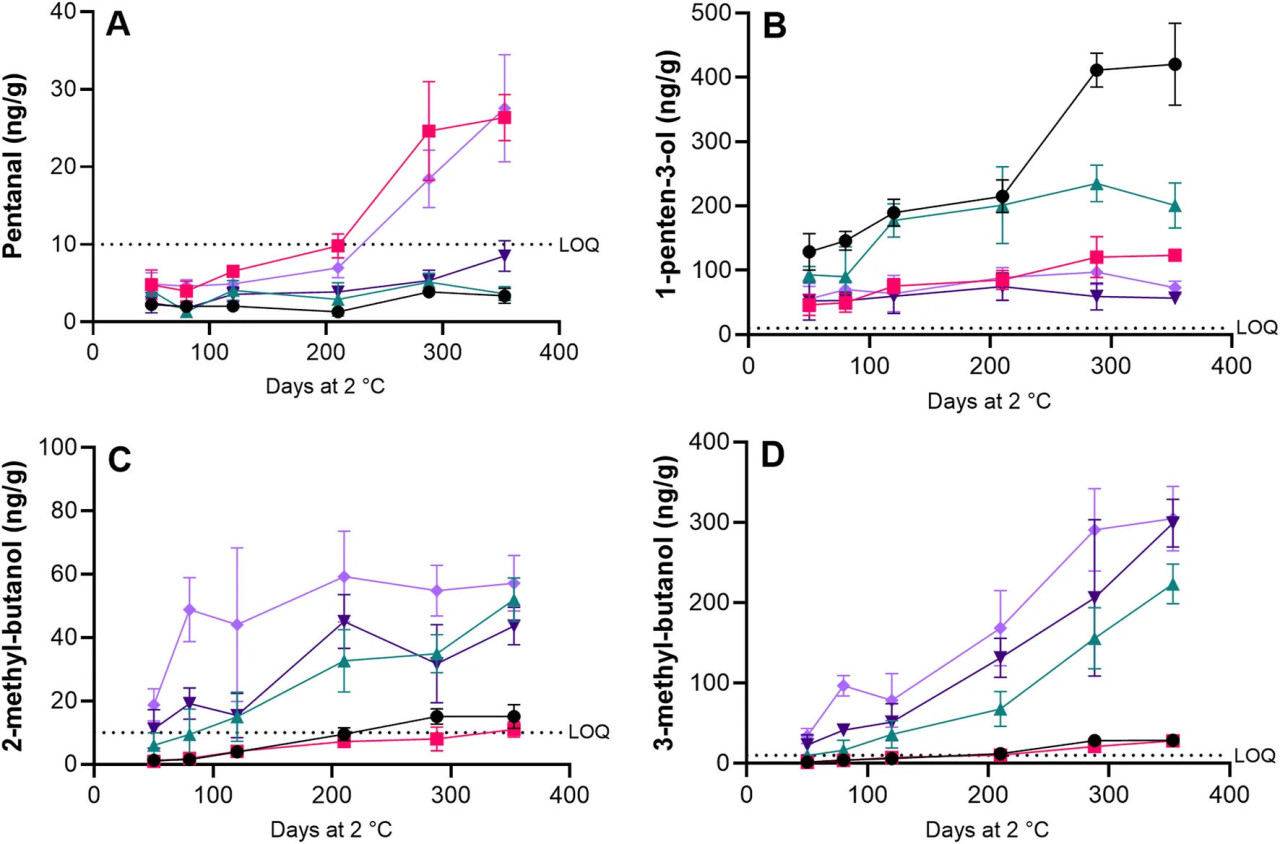

РИСУНОК 7 Развитие выбранных летучих продуктов вторичного окисления во время холодильного хранения образцов икры: пентаналь (A), 1-пентен-3-ол (B), 2-метилбутанол (C) и 3-метилбутанол (D) для обработок HS + BAC (●) LAC + BAC + VIV ( ), AAC + LAC ( ), AAC + LAC + HER ( ) и AAC + LAC + VIV ( ). Символы и планки погрешностей указывают среднее значение ± СО. Пунктирные линии указывают LOQ метода, составляющий приблизительно 10 нг/г.
График PCA, как показано на Рисунке 8, ясно демонстрирует, как различные обработки влияют на образцы икры, и подчеркивает, какие из проанализированных химических и/или микробиологических показателей качества важны для вариации в каждой обработке. Этот анализ позволяет оценить корреляции (положительные или отрицательные) между обработками и измеренными параметрами в икре.
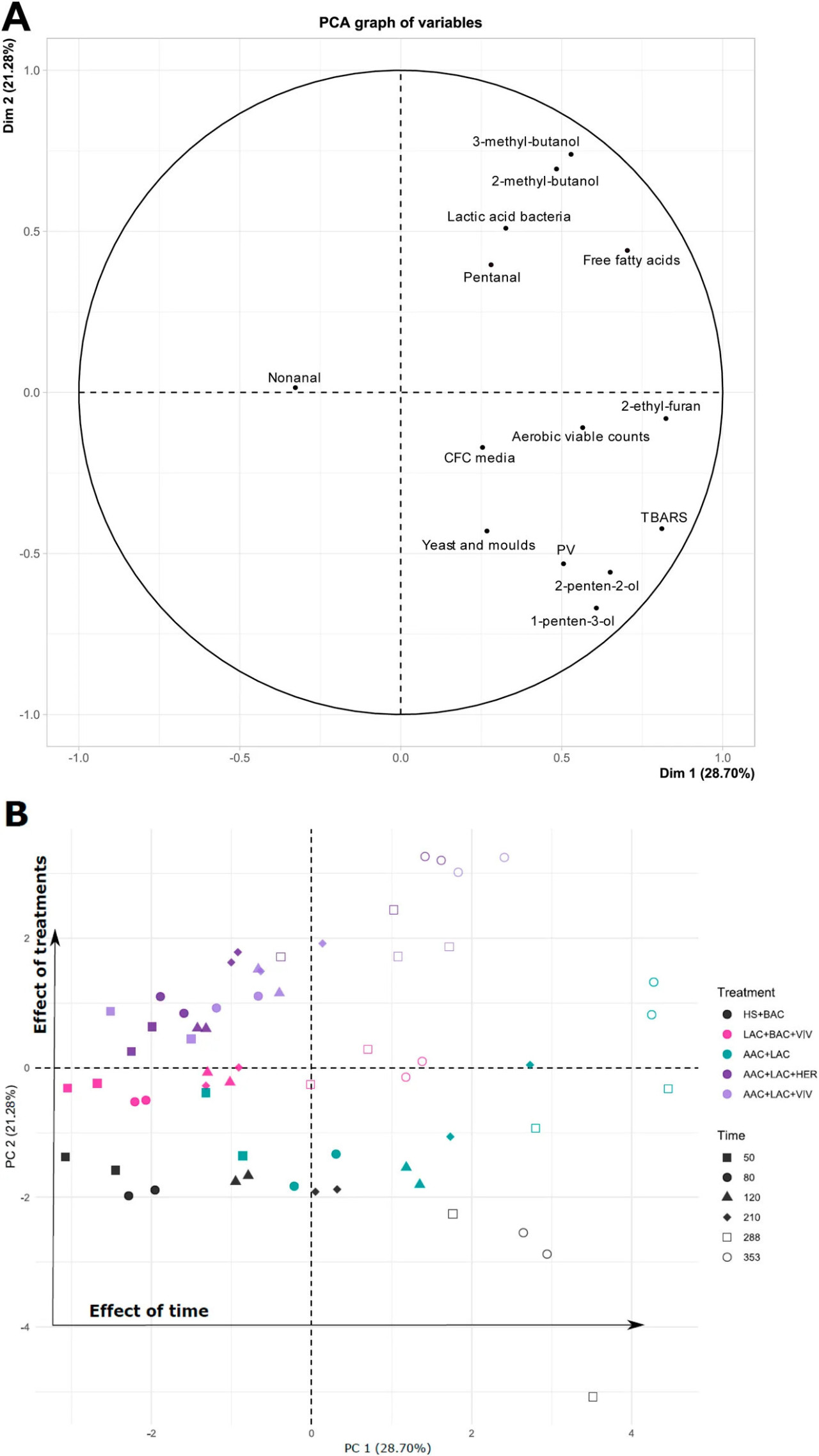
РИСУНОК 8 Анализ главных компонент (PCA) на основе микробных концентраций, FFA, PV, TBARS и летучих соединений образцов икры: Нагрузки (A) и Счета (B). Обработки образцов икры: HS + BAC (черный), LAC + BAC + VIV (розовый), AAC + LAC (зеленый), AAC + LAC + HER (темно-фиолетовый) и AAC + LAC + VIV (светло-фиолетовый), отобранные через 50 (■), 80 (●), 120 (▲), 210 (♦), 288 (□) и 353 (○) дней при 2°C.
Главный компонент 1 (PC1) и PC2 объясняют 30,64% и 21,24% вариации данных, соответственно. Графики показывают четкую временную тенденцию вдоль PC1, тогда как PC2, по-видимому, объясняет дисперсию между обработками (Рисунок 8B). Основываясь на расположении счетов по оси PC1, образцы икры, консервированные с антиоксидантами, по-видимому, варьируются во времени меньше по сравнению с образцами без антиоксидантов. Образцы икры из дней 288 и 353 всех обработок, за исключением HS + BAC, находятся в 1-м квадранте (Рисунок 8B) и коррелируют как со свободными жирными кислотами, молочнокислыми бактериями, так и с тремя летучими соединениями: пентаналем, 2-метилбутанолом и 3-метилбутанолом, т.е. более высоким содержанием (Рисунок 8A). Это хорошо соответствует наблюдаемым более высоким содержаниям FFA (Рисунок 4), 2-метилбутанола, 3-метилбутанола и пентаналя в образцах икры из обработок AAC + LAC + VIV и LAC + BAC + VIV (Рисунок 7). Положение обработанной AAC + LAC + HER икры может быть объяснено изменениями концентраций FFA, 2-метилбутанола и 3-метилбутанола. Образцы HS + BAC из дней 288 и 353, по-видимому, коррелируют с PV, дрожжами и плесенями и двумя летучими спиртами 2-пентен-2-олом и 1-пентен-3-олом. Более высокие TBARS, PV и 1-пентен-3-ол объясняют расположение образцов икры обработок HS + BAC и AAC + LAC. Кроме того, общее увеличение значений большинства параметров в течение периода хранения объясняет дисперсию вдоль PC1.
4 Обсуждение
4.1 Микробиологические изменения
AVC показали уровни 6,1–7,4 log КОЕ/г в течение заключительного периода хранения (>120 дней) в икре из всех обработок. Высокие уровни AVC сходны со значениями, сообщенными в литературе для хранения других икорных продуктов, стабилизированных с заметно более низкими концентрациями соли 3%–5% с органическими кислотами и без них (Basby et al., 1998c; Panjaitan et al., 2020; Lopez et al., 2021). Отсутствие влияния добавления органических кислот на AVC в икре было неожиданным, поскольку предыдущие исследования показали их способность снижать микробный рост в икорных продуктах (Lopez et al., 2021), в креветках в рассоле (Mejlholm et al., 2008) и в икре белого осетра во время длительного хранения при -2°C (Lopez et al., 2021). В отличие от исследований других рыбных продуктов (Campo et al., 2000; Feng et al., 2022), явного противомикробного действия экстракта розмарина в нашем исследовании не наблюдалось, что потенциально связано с составом микробиоты, низким pH продукта или высоким содержанием соли в икре.
В течение периода хранения микробиота всех образцов стала доминироваться дрожжами D. hansenii (Рисунки 2, 3; Таблица 3). Это было неожиданно для образцов икры, обработанных рассолом, содержащим бензойную кислоту, поскольку эта органическая кислота обычно ингибирует дрожжи (Chipley, 2020). Однако эффективность органических кислот зависит от pH и типа продукта. Например, минимальная ингибирующая концентрация (МИК) бензойной кислоты против D. hansenii варьируется в зависимости от pH: 100–200 ppm при pH 3,5, 250–500 ppm при pH 5 и 1500 ppm при pH 7 (Fleet, 2011; Chipley, 2020). Несмотря на >1500 ppm бензойной кислоты (Таблица 2), рост D. hansenii не предотвращался во время хранения изученных обработок СПП. Концентрации AVC и дрожжей в свежеприготовленном СПП из Паамиута составляли, соответственно, 3,2 ± 0,5 log КОЕ/г и 2,1 ± 0,2 log КОЕ/г в течение сезона 2022 года (n = 36; результаты не показаны). Дрожжи, присутствующие после обработки, росли в СПП, как наблюдалось в настоящем исследовании. Микробиота на 50 день для HS + BAC была менее доминирована дрожжами, чем другие обработки, что могло быть результатом >12% соли в водной фазе, в некоторой степени снижающей рост D. hansenii. Bagge-Ravn et al. (2003) обнаружили, что дрожжи составляют 21% микробиоты, извлеченной из оборудования, используемого при дальнейшей обработке СПП в розничные продукты из икры пинагора. Это может предполагать, что дрожжи из СПП загрязнили технологическое оборудование. Более высокие концентрации молочнокислых бактерий на 50 день наблюдались в образцах икры, стабилизированных в рассолах без бензойной кислоты (Рисунок 2C), что указывает на начальный ингибирующий эффект бензойной кислоты на молочнокислые бактерии.
Во время хранения микробиота стала менее разнообразной, поскольку микроорганизмы, которые были лучше всего адаптированы к среде продукта, стали доминировать в образцах икры. Это видно по тому, что в икре в конце хранения присутствует меньше таксонов (Рисунок 3). Исключение бензойной кислоты из рецептов рассолов AAC + LAC, AAC + LAC + HER и AAC + LAC + VIV, по-видимому, также повлияло на разнообразие микробиоты, поскольку несколько бактериальных видов были обнаружены и идентифицированы с помощью MALDI-TOF в образцах икры без бензойной кислоты. Однако в метагеномном анализе Lactobacillales были единственным бактериальным семейством с относительной обильностью >1% в икре. Ранее было показано, что молочнокислые бактерии портят слабосоленую икру пинагора (Basby et al., 1998a; Basby et al., 1998c) и также были выделены из оборудования для переработки морепродуктов (Bagge-Ravn et al., 2003). В частности, Latilactobacillus sakei ранее был выделен из морепродуктов в рассоле из арктических вод (Mejlholm et al., 2008), что может объяснять его присутствие в образцах икры. Ранее было показано, что молочнокислые бактерии способны портить морепродукты, производя сернистые и кислые запахи (Leroi, 2010). D. hansenii была доминирующими дрожжами, обнаруженными во всех образцах как культурально-зависимыми (MALDI-TOF), так и культурально-независимыми (метагеномика) определениями, и была особенно доминирующей после 353 дней хранения. Она была первоначально выделена из морской воды (Norkrans, 1966), что могло привести к естественному заражению пинагора до сбора и, следовательно, к ее присутствию в образцах икры. Дрожжи неоднократно выделялись из продуктов, хранящихся в соленых рассолах, таких как сыр и мясные продукты (Haastrup et al., 2018; Hernández et al., 2018). Известно, что эти дрожжи являются галоторантными, способными расти при низком pH и в присутствии органических кислот (Breuer и Harms, 2006), что, вероятно, заставляет их становиться доминантными во время хранения. В литературе сообщается, что дрожжи действуют и как агент порчи, и как улучшитель вкуса (Breuer и Harms, 2006; Ashaolu et al., 2023), вероятно, в зависимости от пищевой матрицы, в которой они растут, и от микробных концентраций. В будущем следует провести исследования для выяснения их роли в порче икры пинагора. Идентификация единственной колонии Staphylococcus equorum в икре обработки AAC + LAC + HER на 353 день хранения интересна, поскольку эта бактерия обычно не растет при температурах ниже 6°C (Vos et al., 2009). Метагеномный анализ также обнаружил ДНК S. equorum, но с обильностью ниже 0,001%, что показывает его присутствие, хотя и в очень низких количествах. Ранее было обнаружено, что Staphylococcus spp. присутствуют на технологическом оборудовании для икры пинагора (Bagge-Ravn et al., 2003) и были выделены из морских солей (Cordero и Zumalacárregui, 2000), что предполагает, что род может быть внесен через добавление соли в продукты или через окружающую морскую или технологическую среду.
Довольно большое количество неидентифицированных изолятов показывает ограничения использования MALDI-TOF для исследования микробного сообщества СПП. Однако это, скорее всего, можно объяснить размером и типом библиотеки, применяемой для идентификации изолятов. Библиотека, использованная в этом исследовании, состоит из приблизительно 4000 видов, и библиотека в основном нацелена на клинические и наземные изоляты (Han et al., 2021). Ограничением независимого от культивирования метагеномного анализа является неразборчивое секвенирование ДНК из всех жизнеспособных, некультивируемых и мертвых микроорганизмов. Чтобы противодействовать этому ограничению, недавние исследования с использованием моноазида пропедия пытались, с ограниченной эффективностью, уменьшить смещение даже в сложных сообществах (Yap et al., 2022; Liu et al., 2023; Yang et al., 2023). Несмотря на ограничения как культурально-зависимых, так и независимых методов, это исследование показывает, что оба подхода дали одинаковый результат. Преимущество культурального метода заключается в том, что он предоставил нам изоляты D. hansenii, которые могут быть использованы в будущих исследованиях.
При хранении пищевых продуктов в охлажденном состоянии в течение длительных периодов времени необходимо также учитывать и обеспечивать микробиологическую безопасность пищи. Для этой цели полезно прогностическое моделирование (Ross et al., 2000). Прогнозы от Food Safety and Spoilage Predictor (FSSP, www.fssp.dtu.dk) предсказывают, что комбинации соли, pH и органических кислот в экспериментальной обработке рассолом будут ингибировать рост Listeria monocytogenes, который является важнейшим психротолерантным патогеном пищевого происхождения, связанным со слабосолеными рыбными продуктами, во время хранения соленых промежуточных икорных продуктов (Mejlholm и Dalgaard, 2009). Кроме того, характеристики продукта и условия хранения будут эффективно предотвращать рост и потенциальное образование токсинов не протеолитическим Clostridium botulinum (Koukou et al., 2021).
Таким образом, микробиологические результаты показывают, что снижение содержания соли в водной фазе с >12% до ∼8% и отказ от использования бензойной кислоты в рассолах заметно не повлияли на микробный рост в промежуточных икорных продуктах. Также не было очевидного влияния добавления антиоксидантов на микробные изменения. Это указывает на то, что действительно возможно изменить рецептуры рассолов для снижения содержания соли и исключения бензойной кислоты без ущерба для микробиологического качества промежуточного продукта. D. hansenii доминировала в микробиоте изученного продукта СПП, и дрожжи должны учитываться при оценке и управлении его микробиологическим качеством. В частности, для ограничения роста дрожжей предполагается снизить pH продукта, так как изменение с 5,5 до 5,2 будет иметь тот же стабилизирующий эффект, что и удвоение концентраций органических кислот.
4.2 Химические изменения
FFA в морепродуктах образуются в результате липолиза триглицеридов, обычно под действием липолитических ферментов, происходящих из продуктов или микробиоты, но также может происходить термически индуцированный липолиз. FFA более склонны к окислению во время хранения, чем жирные кислоты, связанные с триацилглицеридами или фосфолипидами. Содержание FFA сходным образом увеличилось во всех образцах икры во время хранения, независимо от добавления антиоксидантов в рассолы, что указывает на то, что микробные или эндогенные ферменты, вероятно, были ответственны за образование FFA. Это открытие согласуется с исследованием Ozogul et al. (2010), которое показало, что добавление экстракта розмарина в вакуумированные сардины, хранящиеся при 4°C, не оказало существенного влияния на снижение образования FFA по сравнению с контролем без экстракта розмарина. Более низкое содержание FFA на 50 день было определено в икре из обработки HS + BAC, что может быть связано с более высокой концентрацией соли по сравнению с другими обработками (Рисунок 4). Это, вероятно, связано с ингибирующим действием соли на липазы (Albarracín et al., 2011). Ранее увеличение FFA связывали с прогорклым запахом морепродуктов (Refsgaard et al., 1998; Refsgaard et al., 2000), но необходимо провести больше исследований, чтобы определить их сенсорное влияние в соленой икре пинагора.
Добавление антиоксидантов ингибировало образование гидропероксидов по сравнению с образцами без антиоксидантов. Более того, Herbalox D-450, по-видимому, был более эффективным, чем VivOx 7.5. Причина этого различия между двумя антиоксидантами на основе розмарина неясна, так как единственное различие между ними заключается в добавлении мальтодекстрина в VivOx 7.5. Необходимы дальнейшие исследования для изучения этого различия. В конечной точке отбора проб через 353 дня PV снизилось для всех образцов (Рисунок 6A), что предполагает, что гидропероксиды разлагались на продукты вторичного окисления с большей скоростью, чем они образовывались. Это снижение PV во время хранения также наблюдалось в других рыбных продуктах (Larsson et al., 2007; Elavarasan и Shamasundar, 2022; Jensen et al., 2024).
Более низкие увеличения значений TBARS во время хранения наблюдались в образцах икры, консервированных с антиоксидантами, независимо от различий в составе органических кислот (Рисунок 6B), что показывает полезный эффект добавления антиоксидантов для поддержания окислительной стабильности икры. Аналогичный положительный эффект был показан для добавления экстракта розмарина в другие рыбные продукты, такие как охлажденные вакуумированные сардины, хранящиеся при 4°C (Ozogul et al., 2010), и охлажденный морской окунь (Feng et al., 2022). Результаты указывают на то, что высокое содержание соли в HS + BAC (∼12% NaCl) по сравнению с AAC + LAC (∼8% NaCl) может снижать образование TBARS. Аналогичный эффект высокого содержания соли в снижении TBARS также был сообщен İnanli et al. (2010) и Alak et al. (2021) для соленой икры радужной форели (Oncorhynchus mykiss), хранящейся при 4°C. Последнее исследование предположило, что снижение образования TBARS было связано с обезвоживанием икры, вызванным добавлением соли, которое изменило относительные концентрации липидов и белков в образцах. Ранее сообщалось, что TBARS коррелируют с сенсорными изменениями рыбных продуктов (Larsson et al., 2007). Однако из-за сложности реакций окисления липидов найти универсальный уровень TBARS для определения срока годности и/или сенсорных изменений не просто.
Альдегид пентаналь связан с окислением линолевой кислоты (Gorji et al., 2019). Образование пентаналя, по-видимому, связано с присутствием антиоксиданта VivOx 7.5, поскольку увеличение выше LOQ наблюдалось только в образцах икры, консервированных с этим антиоксидантом, то есть LAC + BAC + VIV и AAC + LAC + VIV (Рисунок 7A). Это может быть связано с содержанием мальтодекстрина в VivOx 7.5, который отсутствует в Herbalox D-450. Следует провести больше исследований для изучения потенциала мальтодекстрина влиять на образование пентаналя. 1-пентен-3-ол может образовываться при окислении n-3 полиненасыщенных жирных кислот (например, докозагексаеновой кислоты (ДГК) и эйкозапентаеновой кислоты (ЭПК)) (Lee et al., 2003). Добавление антиоксидантов привело к тому, что образование этого летучего соединения было заметно снижено по сравнению с образцами икры без антиоксидантов (Рисунок 7B). Кроме того, наблюдалось двукратное увеличение образцов икры HS + BAC между 210 и 288 днями хранения, и концентрация была в 2 раза выше, чем в икре из AAC + LAC после 353 дней хранения. У нас нет четкого объяснения внезапного и заметного увеличения этого летучего соединения в образцах HS + BAC по сравнению с образцами AAC + LAC. 2-метилбутанол и 3-метилбутанол, вероятно, происходят из аминокислот изолейцина и лейцина, соответственно. Промежуточный этап в этом метаболическом пути может включать ферментативное降解 2-метилбутаналя и 3-метилбутаналя с помощью алкогольдегидрогеназ (ADH), происходящих из микроорганизмов, присутствующих в продукте (Zhuang et al., 2021). Из результатов видно, что присутствие бензойной кислоты ограничивало образование этих двух летучих соединений в икре (Рисунки 7C, D), что указывает на влияние органической кислоты на микроорганизмы, которые производят эти метаболиты во время хранения. Глядя на идентификацию микробиоты, представляется вероятным, что члены Lactobacillales могут вносить вклад в образование этих двух летучих соединений, поскольку они были обнаружены в более высокой обильности в образцах икры без бензойной кислоты на протяжении всего хранения. Также сообщалось, что D. hansenii и Psychrobacter spp. продуцируют 2-метилбутанол и/или 3-метилбутанол в пищевых продуктах и модельных системах (Freiding et al., 2011), что подтверждает вероятность того, что микроорганизмы икры способствовали образованию этих двух летучих соединений во время хранения. Подобно результатам этого исследования, было также показано, что содержание 1-пентен-3-ола и 3-метилбутанола увеличивается в соленой икре белого осетра во время хранения при -2°C (Lopez et al., 2021). Другие авторы описали 3-метилбутанол как важный вкладчик в запах перезревшей икры трески, и было высказано предположение, что это соединение может быть потенциальным индикатором порчи (Jonsdottir et al., 2004), но должны быть проведены дальнейшие исследования, чтобы определить важность летучих веществ для сенсорной порчи икры пинагора.
В целом, результаты PV, TBARS и летучих соединений показывают ценность добавления антиоксидантов для снижения окислительных изменений в промежуточном икорном продукте во время охлажденного хранения. Особенно добавление Herbalox D-450 было признано благоприятным для снижения образования как первичных, так и вторичных продуктов окисления по сравнению с использованием VivOx 7.5 (Рисунки 6, 7). Поскольку сходные концентрации активных соединений — карнозовой кислоты и карнозола — были добавлены в рассолы (Рисунок 4), различие может быть связано с добавлением мальтодекстрина в VivOx 7.5, но ранее было показано, что мальтодекстрин улучшает окислительную стабильность, а не способствует окислению. Таким образом, причины различий в действии двух антиоксидантов должны быть далее исследованы.
Различия в цвете икры в зависимости от обработки были отмечены в течение периода хранения; однако изменения не были количественно определены. Дальнейшая работа должна быть предпринята для характеристики сенсорных характеристик икры во время хранения, а также для определения того, влияют ли применяемые антиоксиданты на сенсорные свойства икры пинагора.
5 Заключение
Настоящее исследование является, насколько нам известно, первым, которое оценило микробиологические и окислительные изменения для промежуточного продукта из икры пинагора в рассоле. Результаты этого исследования показывают, насколько полезно исследовать как микробиологические, так и химические изменения продуктов, чтобы иметь возможность понять изменения, потенциальные причины этих изменений и влияние различных обработок на стабильность продукта при хранении. Что касается микробиологической стабильности, не было обнаружено явного влияния различных комбинаций органических кислот в рассолах на микробные концентрации, поскольку все обработки достигли уровней AVC 6,1–7,4 Log КОЕ/г после 120 дней хранения. Образование двух летучих соединений, 2-метилбутанола и 3-метилбутанола, вероятно, было связано с микробиотой промежуточной икры, включая D. hansenii и молочнокислые бактерии, которые встречались в более высоких концентрациях в обработках без бензойной кислоты. Кроме того, не было обнаружено явного противомикробного действия экстрактов розмарина. Дрожжи D. hansenii, идентифицированные с помощью MALDI-TOF и метагеномного секвенирования, росли до высоких концентраций как в обработке с высоким содержанием соли (10% NaCl, pH 6,1 и с бензойной кислотой), так и в изученных альтернативных обработках с пониженным содержанием соли (8%), pH ∼5,4 и где бензойная кислота была заменена комбинацией уксусной и молочной кислот. Чтобы иметь возможность снизить содержание соли и бензойной кислоты в соленом промежуточном икорном продукте и по-прежнему контролировать микробный рост, это исследование предполагает, что pH должен быть снижен дальше, чем изученный уровень ∼5,4, поскольку это усилит ингибирующий эффект добавленных органических кислот. Что касается окислительной стабильности, добавление антиоксидантов снижало окислительные реакции с точки зрения образования гидропероксидов, TBARS и летучих соединений по сравнению с образцами без добавленных антиоксидантов. Не было обнаружено явного влияния различных органических кислот на окислительные изменения. Herbalox D-450 был лучше, чем VivOx 7.5, в ингибировании образования гидропероксидов, TBARS и двух летучих соединений — пентаналя и 1-пентен-3-ола. В заключение, из результатов очевидно, что можно снизить содержание соли и использовать альтернативы потенциальному пищевому аллергену — бензойной кислоте — без ущерба для качества продукта.
Вспомогательные материалы
Вспомогательные материалы к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/frfst.2024.1498035/full#supplementary-material
Ссылки
1. Alak G., Kaynar Ö., Atamanalp M. (2021). The impact of salt concentrations on the physicochemical and microbiological changes of rainbow trout caviar. Food Biosci. 41, 100976. doi: 10.1016/j.fbio.2021.100976. CrossRef Google Scholar
2. Albarracín W., Sánchez I. C., Grau R., Barat J. M. (2011). Salt in food processing; usage and reduction: a review. Int J Food Sci Tech 46, 1329–1336. doi: 10.1111/j.1365-2621.2010.02492.x. CrossRef Google Scholar
3. AOAC (1976). Sodium chloride in canned vegetables. Potentiometric method. Arlington, VA, USA: AOAC. Official method 971.27. Google Scholar
4. AOAC (1996). Fish and marine products. Treatment and preparation of sample procedure. Arlington, VA, USA: AOAC. Official method 937.07. Google Scholar
5. AOAC (2000). Salt (chlorine as sodium chloride) in seafood. Potentiometric method. Arlington, VA, USA: AOAC. Official method 976.18. Google Scholar
6. AOCS (1998). Tocopherols and tocotrienols in vegetable oils and fats by HPLC. IL, USA: AOCS: Campaign. Official method Ce 8-89. Google Scholar
7. AOCS (2004). Free fatty acids in crude and refined fats and oils. IL, USA: AOCS: Campaign. Official method Ca 5a-40. Google Scholar
8. Ashaolu T. J., Khalifa I., Mesak M. A., Lorenzo J. M., Farag M. A. (2023). A comprehensive review of the role of microorganisms on texture change, flavor and biogenic amines formation in fermented meat with their action mechanisms and safety. Crit. Rev. Food Sci. Nutr. 63, 3538–3555. doi: 10.1080/10408398.2021.1929059. CrossRef Google Scholar
9. Bagge-Ravn D., Ng Y., Hjelm M., Christiansen J. N., Johansen C., Gram L. (2003). The microbial ecology of processing equipment in different fish industries - analysis of the microflora during processing and following cleaning and disinfection. Int. J. Food Microbiol. 87, 239–250. doi: 10.1016/S0168-1605(03)00067-9. CrossRef Google Scholar
10. Basby M., Jeppesen V. F., Huss H. H. (1998a). Spoilage of lightly salted Lumpfish (Cyclopterus lumpus) roe at 5 °C. J. Aquat. Food Prod. 7, 23–34. doi: 10.1300/J030v07n04_04. CrossRef Google Scholar
11. Basby M., Jeppesen V. F., Huss H. H. (1998b). Chemical composition of fresh and salted Lumpfish (Cyclopterus lumpus) roe. J. Aquat. Food Prod. 7, 7–21. doi: 10.1300/J030v07n04_03. CrossRef Google Scholar
12. Basby M., Jeppesen V. F., Huss H. H. (1998c). Characterization of the microflora of lightly salted Lumpfish (Cyclopterus lumpus) roe stored at 5 °C. J. Aquat. Food Prod. 7, 35–51. doi: 10.1300/J030v07n04_05. CrossRef Google Scholar
13. Bizzini A., Durussel C., Bille J., Greub G., Prod'hom G. (2010). Performance of matrix-assisted laser desorption ionization-time of flight mass spectrometry for identification of bacterial strains routinely isolated in a clinical microbiology laboratory. J. Clin. Microbiol. 48, 1549–1554. doi: 10.1128/JCM.01794-09. CrossRef Google Scholar
14. Bledsoe G. E., Bledsoe C. D., Rasco B. (2003). Caviars and fish roe products. Crit. Rev. Food Sci. Nutr. 43, 317–356. doi: 10.1080/10408690390826545. CrossRef Google Scholar
15. Bligh E. G., Dyer W. J. (1959). A rapid method of total lipid extraction and purification. J. Biochem. Physiol. 37, 911–917. doi: 10.1139/o59-099. CrossRef Google Scholar
16. Breuer U., Harms H. (2006). Debaryomyces hansenii — an extremophilic yeast with biotechnological potential. Yeast 23, 415–437. doi: 10.1002/yea.1374. CrossRef Google Scholar
17. Campo J. D., Amiot M.-J., Nguyen-The C. (2000). Antimicrobial effect of rosemary extracts. J. Food Prot. 63, 1359–1368. doi: 10.4315/0362-028X-63.10.1359. CrossRef Google Scholar
18. Chipley J. R. (2020). "Sodium benzoate and benzoic acid," in Antimicrobials in food. Editors Davidson P. M., Taylor T. M., David J. R. D. Fourth edition (Boca Raton: CRC Press), 41–88. doi: 10.1201/9780429058196-3. CrossRef Google Scholar
19. Cordero M. R., Zumalacárregui J. M. (2000). Characterization of Micrococcaceae isolated from salt used for Spanish dry-cured ham. Lett. Appl. Microbiol. 31, 303–306. doi: 10.1046/j.1472-765x.2000.00818.x. CrossRef Google Scholar
20. Dalgaard P., Gram L., Huss H. H. (1993). Spoilage and shelf-life of cod fillets packed in vacuum or modified atmospheres. Int. J. Food Microbiol. 19, 283–294. doi: 10.1016/0168-1605(93)90020-H. CrossRef Google Scholar
21. Dalgaard P., Jørgensen L. V. (2000). Cooked and brined shrimps packed in a modified atmosphere have a shelf‐life of >7 months at 0°C, but spoil in 4–6 days at 25°C. Int. J. Food Sci. Technol. 35, 431–442. doi: 10.1046/j.1365-2621.2000.00402.x. CrossRef Google Scholar
22. Davidson C. M., Cronin F. (1973). Medium for the selective enumeration of lactic acid bacteria from foods. Appl. Microbiol. 26, 439–440. doi: 10.1128/am.26.3.439-440.1973. CrossRef Google Scholar
23. Elavarasan K., Shamasundar B. A. (2022). Antioxidant properties of papain mediated protein hydrolysates from fresh water carps (Catla catla, Labeo rohita and Cirrhinus mrigala) and its application on inhibition of lipid oxidation in oil sardine mince during ice storage. J. Food Sci. Technol. 59, 636–645. doi: 10.1007/s13197-021-05053-0. CrossRef Google Scholar
24. Feng H., Liu J., Timira V., Lin H., Wang H., Li Z. (2022). Effects of Plant extracts (Clove, Fennel, Rosemary, Cinnamon, and Tea Polyphenols) on microbiological, chemical, and sensory quality of smoke-flavored sea bass during chilled storage. ACS Food Sci. Technol. 2, 1692–1700. doi: 10.1021/acsfoodscitech.2c00131. CrossRef Google Scholar
25. Fleet G. H. (2011). "Yeast spoilage of foods and beverages," in The yeasts (Elsevier), 53–63. doi: 10.1016/B978-0-444-52149-1.00005-7. CrossRef Google Scholar
26. Freiding S., Gutsche K. A., Ehrmann M. A., Vogel R. F. (2011). Genetic screening of Lactobacillus sakei and Lactobacillus curvatus strains for their peptidolytic system and amino acid metabolism, and comparison of their volatilomes in a model system. Syst. Appl. Microbiol. 34, 311–320. doi: 10.1016/j.syapm.2010.12.006. CrossRef Google Scholar
27. Gorji S. G., Calingacion M., Smyth H. E., Fitzgerald M. (2019). Comprehensive profiling of lipid oxidation volatile compounds during storage of mayonnaise. J. Food Sci. Technol. 56, 4076–4090. doi: 10.1007/s13197-019-03876-6. CrossRef Google Scholar
28. Haastrup M. K., Johansen P., Malskær A. H., Castro-Mejía J. L., Kot W., Krych L., et al (2018). Cheese brines from Danish dairies reveal a complex microbiota comprising several halotolerant bacteria and yeasts. Int. J. Food Microbiol. 285, 173–187. doi: 10.1016/j.ijfoodmicro.2018.08.015. CrossRef Google Scholar
29. Han S.-S., Jeong Y.-S., Choi S.-K. (2021). Current scenario and challenges in the direct identification of microorganisms using MALDI TOF MS. Microorganisms 9, 1917. doi: 10.3390/microorganisms9091917. CrossRef Google Scholar
30. Hernández A., Pérez-Nevado F., Ruiz-Moyano S., Serradilla M. J., Villalobos M. C., Martín A., et al (2018). Spoilage yeasts: what are the sources of contamination of foods and beverages? Int. J. Food Microbiol. 286, 98–110. doi: 10.1016/j.ijfoodmicro.2018.07.031. CrossRef Google Scholar
31. İnanli A. G., Coban Ö. E., Dartay M. (2010). The chemical and sensorial changes in rainbow trout caviar salted in different ratios during storage. Fish. Sci. 76, 879–883. doi: 10.1007/s12562-010-0279-6. CrossRef Google Scholar
32. Iverson S. J., Lang S. L. C., Cooper M. H. (2001). Comparison of the Bligh and Dyer and Folch methods for total lipid determination in a broad range of marine tissue. Lipids 36, 1283–1287. doi: 10.1007/s11745-001-0843-0. CrossRef Google Scholar
33. Jensen H. A., Sørensen A.-D. M., Mejlholm O., Bøknæs N., Jacobsen C. (2024). Effect of different antioxidants and application methods on the shelf-life of whole, cooked, frozen Northern shrimp (Pandalus borealis). Food Chem. Adv. 5, 100764. doi: 10.1016/j.focha.2024.100764. CrossRef Google Scholar
34. Johannesson J. (2006). Lumpfish caviar - from vessel to consumer. Rome: FAO, 60. FAO Fisheries Technical Paper. No. 485. Google Scholar
35. Jonsdottir R., Olafsdottir G., Martinsdottir E., Stefansson G. (2004). Flavor characterization of ripened cod roe by gas chromatography, sensory analysis, and electronic nose. J. Agric. Food Chem. 52, 6250–6256. doi: 10.1021/jf049456g. CrossRef Google Scholar
36. Kennedy J., Durif C. M. F., Florin A.-B., Fréchet A., Gauthier J., Hüssy K., et al (2019). A brief history of lumpfishing, assessment, and management across the North Atlantic. ICES J. Mar. Sci. 76, 181–191. doi: 10.1093/icesjms/fsy146. CrossRef Google Scholar
37. Koukou I., Mejlholm O., Dalgaard P. (2021). Cardinal parameter growth and growth boundary model for non-proteolytic Clostridium botulinum -- effect of eight environmental factors. Int. J. Food Microbiol. 346, 109162. doi: 10.1016/j.ijfoodmicro.2021.109162. CrossRef Google Scholar
38. Larsson K., Almgren A., Undeland I. (2007). Hemoglobin-mediated lipid oxidation and compositional characteristics of washed fish mince model systems made from cod (Gadus morhua), herring (Clupea harengus), and salmon (Salmo salar) muscle. J. Agric. Food Chem. 55, 9027–9035. doi: 10.1021/jf070522z. CrossRef Google Scholar
39. Lee H., Kizito S. A., Weese S. J., Craig-Schmidt M. C., Lee Y., Wei C.-I., et al (2003). Analysis of headspace volatile and oxidized volatile compounds in DHA-enriched fish oil on accelerated oxidative storage. J. Food Sci. 68, 2169–2177. doi: 10.1111/j.1365-2621.2003.tb05742.x. CrossRef Google Scholar
40. Leroi F. (2010). Occurrence and role of lactic acid bacteria in seafood products. Food Microbiol. 27, 698–709. doi: 10.1016/j.fm.2010.05.016. CrossRef Google Scholar
41. Liu F., Lu H., Dong B., Huang X., Cheng H., Qu R., et al (2023). Systematic evaluation of the viable microbiome in the human oral and gut samples with spike-in Gram+/-- bacteria. mSystems 8, e0073822–22. doi: 10.1128/msystems.00738-22. CrossRef Google Scholar
42. Lopez A., Bellagamba F., Tirloni E., Vasconi M., Stella S., Bernardi C., et al (2021). Evolution of food safety features and volatile profile in white sturgeon caviar treated with different formulations of salt and preservatives during a long-term storage time. Foods 10, 850. doi: 10.3390/foods10040850. CrossRef Google Scholar
43. Lu F. S. H., Bruheim I., Ale M. T., Jacobsen C. (2015). The effect of thermal treatment on the quality changes of Antartic krill meal during the manufacturing process: high processing temperatures decrease product quality. Eur. J. Lipid Sci. Technol. 117, 411–420. doi: 10.1002/ejlt.201400280. CrossRef Google Scholar
44. Mejlholm O., Dalgaard P. (2009). Development and validation of an extensive growth and growth boundary model for Listeria monocytogenes in lightly preserved and ready-to-eat shrimp. J. Food Prot. 72, 2132–2143. doi: 10.4315/0362-028X-72.10.2132. CrossRef Google Scholar
45. Mejlholm O., Kjeldgaard J., Modberg A., Vest M. B., Bøknæs N., Koort J., et al (2008). Microbial changes and growth of Listeria monocytogenes during chilled storage of brined shrimp (Pandalus borealis). Int. J. Food Microbiol. 124, 250–259. doi: 10.1016/j.ijfoodmicro.2008.03.022. CrossRef Google Scholar
46. NMKL (2006). Aerobic count and specific spoilage organisms in fish and fish products No. 184. Bergen, Norway: Nordic Committee on Food Analysis, 2–7. Google Scholar
47. Nonnemann B., Lyhs U., Svennesen L., Kristensen K. A., Klaas I. C., Pedersen K. (2019). Bovine mastitis bacteria resolved by MALDI-TOF mass spectrometry. J. Dairy Sci. 102, 2515–2524. doi: 10.3168/jds.2018-15424. CrossRef Google Scholar
48. Norkrans B. (1966). Studies on marine occurring yeasts: growth related to pH, NaCl concentration and temperature. Arch. Mikrobiol. 54, 374–392. doi: 10.1007/BF00406719. CrossRef Google Scholar
49. Ozogul Y., Ayas D., Yazgan H., Ozogul F., Boga E. K., Ozyurt G. (2010). The capability of rosemary extract in preventing oxidation of fish lipid. Int. J. Food. Sci. Tech. 45, 1717–1723. doi: 10.1111/j.1365-2621.2010.02326.x. CrossRef Google Scholar
50. Panjaitan F. C. A., Yeh N., Chai H. (2020). Effect of different processing on quality changes of barramundi (Lates calcarifer) caviar during chilled storage (4°C). J. Food Process. Preserv. 44. doi: 10.1111/jfpp.14790. CrossRef Google Scholar
51. Pazos M., Sánchez L., Medina I. (2005). Alpha-tocopherol oxidation in fish muscle during chilling and frozen storage. J. Agric. Food Chem. 53, 4000–4005. doi: 10.1021/jf047843k. CrossRef Google Scholar
52. Pountney S. M., Lein I., Selly S.-L. C., Migaud H., Davie A. (2022). Comparative proximate analysis of wild and captive lumpfish (Cyclopterus lumpus) eggs show deficiencies in captive eggs and possible egg quality determinants. Aquaculture 557, 738356. doi: 10.1016/j.aquaculture.2022.738356. CrossRef Google Scholar
53. Refsgaard H. H. F., Brockhoff P. B., Jensen B. (1998). Sensory and chemical changes in farmed Atlantic salmon (Salmo salar) during frozen storage. J. Agric. Food Chem. 46, 3473–3479. doi: 10.1021/jf980309q. CrossRef Google Scholar
54. Refsgaard H. H. F., Brockhoff P. M. B., Jensen B. (2000). Free polyunsaturated fatty acids cause taste deterioration of Salmon during frozen storage. J. Agric. Food Chem. 48, 3280–3285. doi: 10.1021/jf000021c. CrossRef Google Scholar
55. Ross T., Dalgaard P., Tienungoon S. (2000). Predictive modelling of the growth and survival of Listeria in fishery products. Int. J. Food Microbiol. 62, 231–245. doi: 10.1016/S0168-1605(00)00340-8. CrossRef Google Scholar
56. Shantha N. C., Decker E. A. (1994). Rapid, sensitive, iron-based spectrophotometric methods for determination of peroxide values of food lipids. J. AOAC Int. 77, 421–424. doi: 10.1093/jaoac/77.2.421. CrossRef Google Scholar
57. Vasconi M., Tirloni E., Stella S., Coppola C., Lopez A., Bellagamba F., et al (2020). Comparison of chemical composition and safety issues in fish roe products: application of chemometrics to chemical data. Foods 9, 540. doi: 10.3390/foods9050540. CrossRef Google Scholar
58. Vos P., Garrity G. M., Jones D., Krieg N. R., Ludwig W., Rainey F. A., et al (2009). Bergey's manual of systematic bacteriology. Volume 3: the firmicutes. 2nd Edn. (NY: Springer New York). doi: 10.1007/978-0-387-68489-5. CrossRef Google Scholar
59. Vyncke W. (1970). Direct determination of the thiobarbituric acid value in trichloracetic acid extracts of fish as a measure of oxidative rancidity. Fette, Seifen, Anstrichm. 72, 1084–1087. doi: 10.1002/lipi.19700721218. CrossRef Google Scholar
60. Yang Y., Deng Y., Shi X., Liu L., Yin X., Zhao W., et al (2023). QMRA of beach water by Nanopore sequencing-based viability-metagenomics absolute quantification. Water Res. 235, 119858. doi: 10.1016/j.watres.2023.119858. CrossRef Google Scholar
61. Yap M., O'Sullivan O., O'Toole P. W., Cotter P. D. (2022). Development of sequencing-based methodologies to distinguish viable from non-viable cells in a bovine milk matrix: a pilot study. Front. Microbiol. 13, 1036643. doi: 10.3389/fmicb.2022.1036643. CrossRef Google Scholar
62. Zhuang S., Hong H., Zhang L., Luo Y. (2021). Spoilage-related microbiota in fish and crustaceans during storage: research progress and future trends. Compr. Rev. Food Sci. Food Saf. 20, 252–288. doi: 10.1111/1541-4337.12659. CrossRef Google Scholar

Jensen HA, Plaza LG, Houmann RH, Andreasen PB, Bøknæs N, Kragh ML, Hansen LT, Mejlholm O, Jacobsen C, Dalgaard P and Sørensen A-DM (2024) Microbial and chemical stability of lumpfish (Cyclopterus lumpus) roe during refrigerated storage in acid-salt brines with or without antioxidants. Front. Food. Sci. Technol. 4:1498035. doi: 10.3389/frfst.2024.1498035
Перевод статьи «Microbial and chemical stability of lumpfish (Cyclopterus lumpus) roe during refrigerated storage in acid-salt brines with or without antioxidants» авторов Jensen HA, Plaza LG, Houmann RH, Andreasen PB, Bøknæs N, Kragh ML, Hansen LT, Mejlholm O, Jacobsen C, Dalgaard P and Sørensen A-DM., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык
Фото: magnific

















































































































Комментарии (0)