Бактериоцины молочнокислых бактерий: стратегии биозащиты молочных продуктов
Молочнокислые бактерии (МКБ), играющие ключевую роль в ферментации молока, продуцируют целый ряд антимикробных соединений, особенно бактериоцины, которые способствуют увеличению срока хранения молочных продуктов. Бактериоцины, рибосомально синтезируемые пептиды, обладают широким или узким спектром антимикробной активности, что делает их перспективными для консервации пищевых продуктов. Классификация бактериоцинов МКБ сложна и отражает постоянно развивающиеся геномные данные и механизмы биосинтеза.
Аннотация
Стратегии внедрения бактериоцинов в молочные продукты включают использование очищенных форм, бактериоциногенных штаммов МКБ и ферментатов, содержащих бактериоцины, каждая из которых имеет свои преимущества и особенности. Оптимизация условий ферментации (времени, температуры, pH и питательной среды) необходима для максимальной продукции бактериоцинов. Такая оптимизация способствует повышению качества и безопасности ферментированных молочных продуктов, что соответствует растущим потребительским предпочтениям в пользу натуральных продуктов с минимальной обработкой.
Кроме того, включение бактериоцинов в комплексный (барьерный) подход наряду с термическими и нетермическими методами обработки открывает перспективы для усиления биозащиты продуктов при одновременном снижении зависимости от химических консервантов. Данный всесторонний обзор подчеркивает потенциал бактериоцинов МКБ как натуральной и эффективной альтернативы традиционным консервантам, предлагая понимание их применения и оптимизации при сохранении молочных продуктов.
1 Введение
Молочнокислые бактерии (LAB) представляют собой гетерогенную группу не образующих спор, грамположительных, каталазоотрицательных без цитохромов, анаэробных или аэротолерантных, требовательных к питательным веществам, кислотоустойчивых и строго бродильных бактерий, продуцирующих молочную кислоту в качестве основного продукта при сбраживании сахаров. LAB издавна ценятся за их ключевую роль в ферментации молочных продуктов, способствуя не только вкусу и текстуре, но и сохранности этих продуктов (Deegan et al., 2006; Favaro et al., 2015). Последняя характеристика в первую очередь обусловлена продукцией LAB антимикробных соединений, включающих главным образом органические кислоты, перекись водорода, диацетил, ацетойн, этанол, антигрибковые пептиды и бактериоцины. Среди них бактериоцины выделяются как мощные антимикробные пептиды с разнообразными применениями в консервации пищевых продуктов.
Бактериоцины представляют собой рибосомально синтезируемые антимикробные пептиды, способные уничтожать микроорганизмы, которые являются близкородственными или нет (соответственно узкий или широкий спектр действия) (Alvarez-Sieiro et al., 2016; O’Connor et al., 2020). Многие LAB и их продукты соответствуют статусу «Общепризнанно безопасные» (GRAS) Управления по санитарному надзору за качеством пищевых продуктов и медикаментов США (FDA) или «Квалифицированной презумпции безопасности» (QPS) Европейского управления по безопасности пищевых продуктов (EFSA) для микроорганизмов, преднамеренно вводимых в пищевую цепочку. Следовательно, LAB могут использоваться в пищевой промышленности для производства ферментированных продуктов благодаря их способности к биоконсервации. Особенно в ферментированных молочных продуктах бактериоцины предлагают привлекательное решение. Эти пептиды не только ингибируют рост патогенов после обработки, но и регулируют посторонние микроорганизмы, а также усиливают процесс ферментации, в конечном итоге обогащая сенсорные характеристики сыров и аналогичных продуктов (O’Connor et al., 2020; Simons et al., 2020). Бактериоцины LAB проявляют замечательную термостойкость, выдерживают широкий диапазон pH, не имеют различимых цвета, запаха или вкуса и уязвимы для деградации пищеварительными протеазами человека, что сохраняет баланс микробиоты кишечника. Эти атрибуты делают бактериоцины хорошо пригодными для использования в консервации пищевых продуктов (Daba and Elkhateeb, 2020). Использование бактериоцинов в молочной промышленности отражает более широкий общественный сдвиг в сторону натуральных продуктов с минимальной обработкой, не содержащих синтетических консервантов.
В свете растущего потребительского спроса на натуральные продукты без добавок включение бактериоцинов представляет собой перспективную возможность для обеспечения целостности и полезности молочных продуктов.
2 Бактериоцины LAB
2.1 Классификация
Классификация бактериоцинов LAB представляет собой сложную область, обогащенную недавними геномными исследованиями и развивающимся пониманием этих антимикробных пептидов. Начиная с первоначальной классификации Klaenhammer в 1993 году, было предложено несколько схем, сосредоточенных на механизмах биосинтеза и биологической активности (Simons et al., 2020).
Хотя полный консенсус все еще отсутствует, некоторые авторы (Arnison et al., 2013; Cotter et al., 2013; Alvarez-Sieiro et al., 2016) предложили классификацию, основанную на биологической активности и механизмах биосинтеза. Здесь мы представляем краткий обзор:
Класс I, лантибиотики, или RiPPs (рибосомально продуцируемые и посттрансляционно модифицированные пептиды, термостойкие) бактериоцины, который включает: класс Ia — пептиды, подвергающиеся посттрансляционным модификациям с добавлением лантионина или (метил)лантионина (например, низин); класс Ib (циклические пептиды «голова-к-хвосту»), образованные циклическими пептидами, связанными N- и C-концами. Они образуют поры в бактериальных мембранах; и класс Ic — пептиды, содержащие связь сера-альфа-углерод, со структурой «шпильки». Класс II, не-лантибиотики (или немодифицированные бактериоцины, термостойкие), включает класс IIa или педиоциноподобные, которые представляют собой бактериоцины широкого спектра действия, активные главным образом против видов Listeria (например, педиоцин PA-1); класс IIb состоит из двух различных пептидов (например, лактицин F); класс IIc — циклические пептиды (например, энтероцин AS-48); и класс IId или не-педиоциноподобные бактериоцины — линейные пептиды. Класс III — крупные, термочувствительные белки, которые включают класс IIIa или бактериолизины (например, энтеролизин A), а класс IIIb — нелитические белки. Класс IV — сложные, термочувствительные белки с липидными или углеводными остатками (липо- или гликопротеины; например, лактоцин 27) (Рисунок 1).
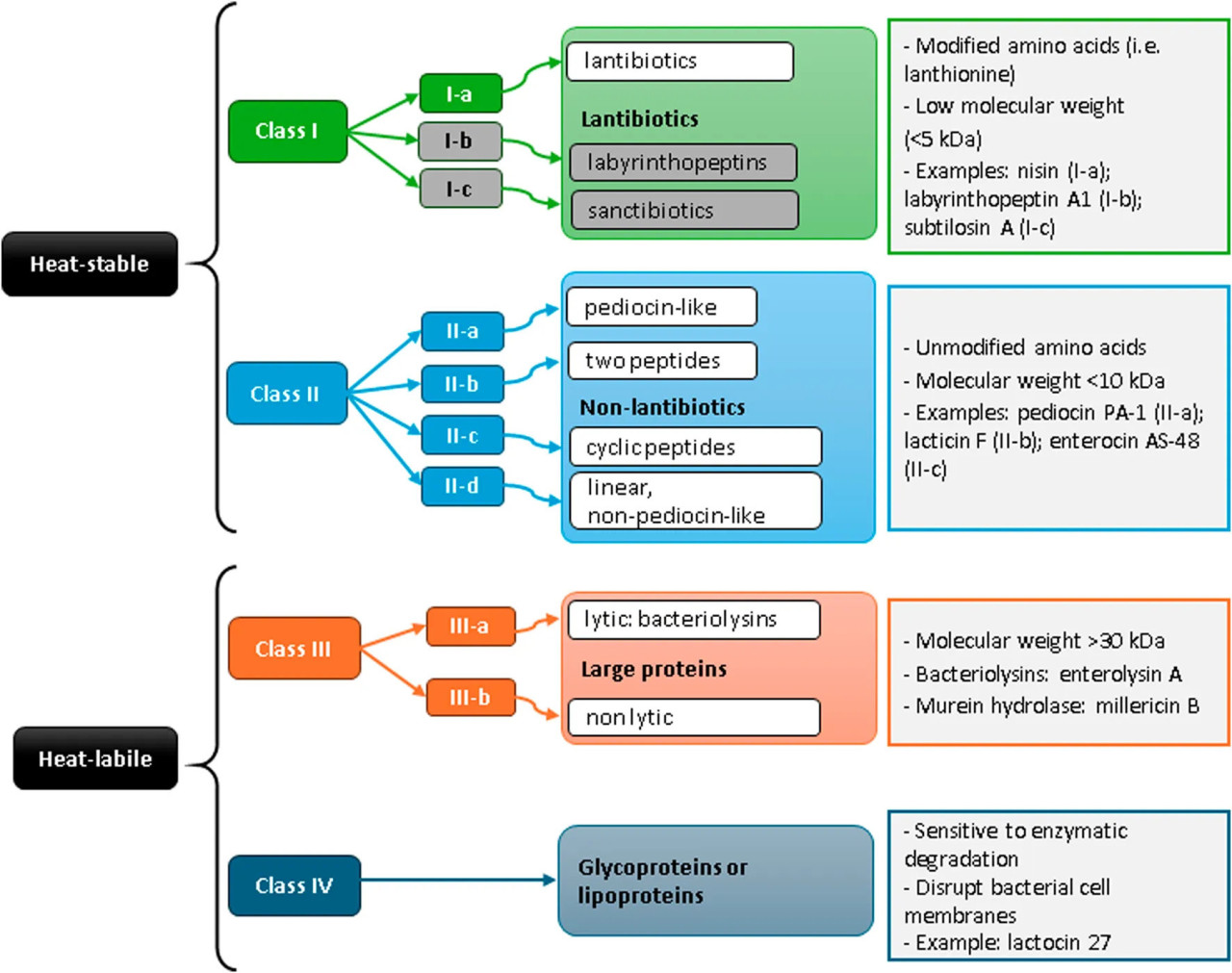
РИСУНОК 1 Общая классификация бактериоцинов молочнокислых бактерий. Подклассы I-b (лабиринтопептины) и I-c (сактибиотики) были включены в схему (серым цветом), а также примеры молекул, хотя они не были зарегистрированы у молочнокислых бактерий на сегодняшний день.
2.2 Использование бактериоцинов в молочных продуктах
Бактериоцины могут использоваться для биоконсервации пищевых продуктов как отдельно, так и в комбинации с другими методами защиты. Однако результаты in vitro должны быть подтверждены в пищевой матрице, чтобы гарантировать эффективность бактериоцинов. Для применения бактериоцинов в пищевых продуктах необходимо соблюдение определенных важных критериев. Продуцирующие штаммы должны быть пищевыми (GRAS или QPS), проявлять широкий спектр ингибирования, обладать высокой удельной активностью, не представлять риска для здоровья, оказывать благотворное воздействие (такое как повышение безопасности, качества и вкуса пищевых продуктов), а также демонстрировать стабильность при нагревании и pH, а также оптимальную растворимость и стабильность для конкретного пищевого применения (Silva et al., 2018).
Было проведено множество исследований по активности, характеристике, организмам-мишеням и применению бактериоцинов; однако лишь несколько были коммерциализированы. Прежде чем достичь этой стадии, необходимо решить различные проблемы. Бактериоцины, используемые в пищевых продуктах, должны быть безопасными и не обладать токсичностью для клеток человека. Некоторые исследования сообщали о низкой цитотоксичности как в экспериментах in vitro, так и in vivo, но только после использования высоких количеств бактериоцинов (превышающих требуемые минимальные ингибирующие концентрации для патогенов или организмов, вызывающих порчу) и длительной инкубации. Кроме того, присутствие некоторых соединений (таких как протеазы) в пищевой матрице может быстро разрушать бактериоцины, препятствуя их биозащитной активности (Soltani et al., 2021). Наконец, коммерческое производство бактериоцина будет зависеть от баланса затрат и выгод при производстве больших количеств очищенного бактериоцина. Это включает крупномасштабное производство, соответствующие процессы очистки и надлежащее хранение для обеспечения стабильности бактериоцина (Reuben et al., 2024).
На протяжении веков бактериоцины потреблялись в ферментированных и неферментированных продуктах. В последнее время все больше людей ищут натуральные продукты без искусственных консервантов. Эта тенденция особенно заметна в молочной промышленности, где растет потребление натуральных ферментированных молок, что стимулирует переход к изучению новых и более натуральных методов консервации.
Ферментированные молочные продукты требуют хранения в охлажденном состоянии, что оптимально для выживания и пролиферации психротрофных бактерий, таких как L. monocytogenes. Следовательно, заражение может произойти на более поздних стадиях переработки молочных продуктов (Carpentier and Cerf, 2011). Таким образом, бактериоцины становятся оптимальным решением для ингибирования роста этого патогена после тепловой обработки (Cotter et al., 2005). Кроме того, бактериоцины могут служить для регулирования посторонних микроорганизмов, таких как нестартерные молочнокислые бактерии (NSLAB) в сыре. Более того, бактериоцины могут быть полезны для ускорения ферментации пищевых продуктов, ускорения созревания сыра и даже улучшения его вкусового профиля (O’Sullivan et al., 2002).
Бактериоцины могут быть включены в молочные продукты в виде очищенных соединений, путем добавления продуцирующей бактериоцины LAB (в качестве вспомогательной культуры) или ферментата, содержащего бактериоцин (в качестве добавки или ингредиента) (Silva et al., 2018; O’Connor et al., 2020) (Рисунок 2; Таблица 1).
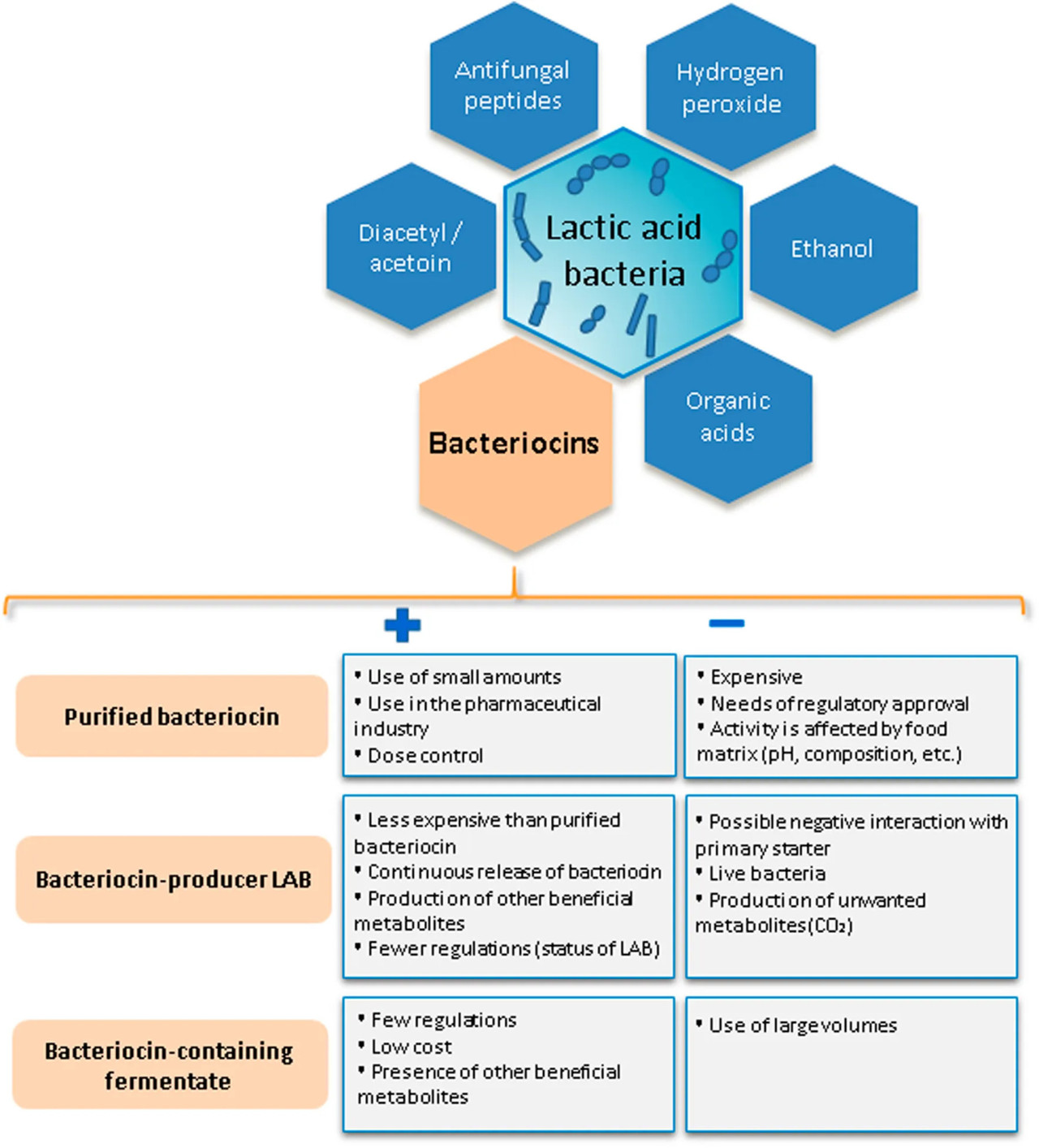
РИСУНОК 2 Антимикробные соединения, продуцируемые молочнокислыми бактериями. Преимущества и недостатки различных подходов к включению бактериоцинов в молочные продукты.
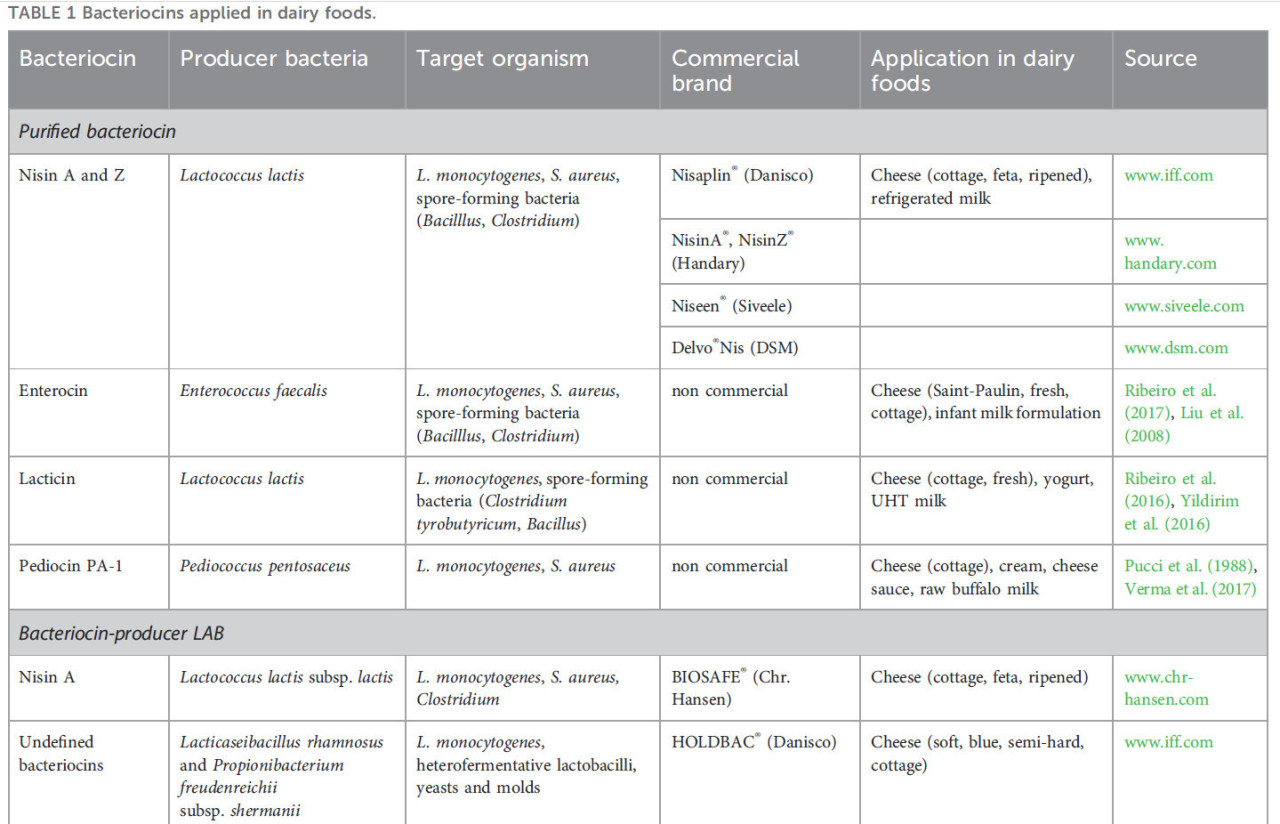
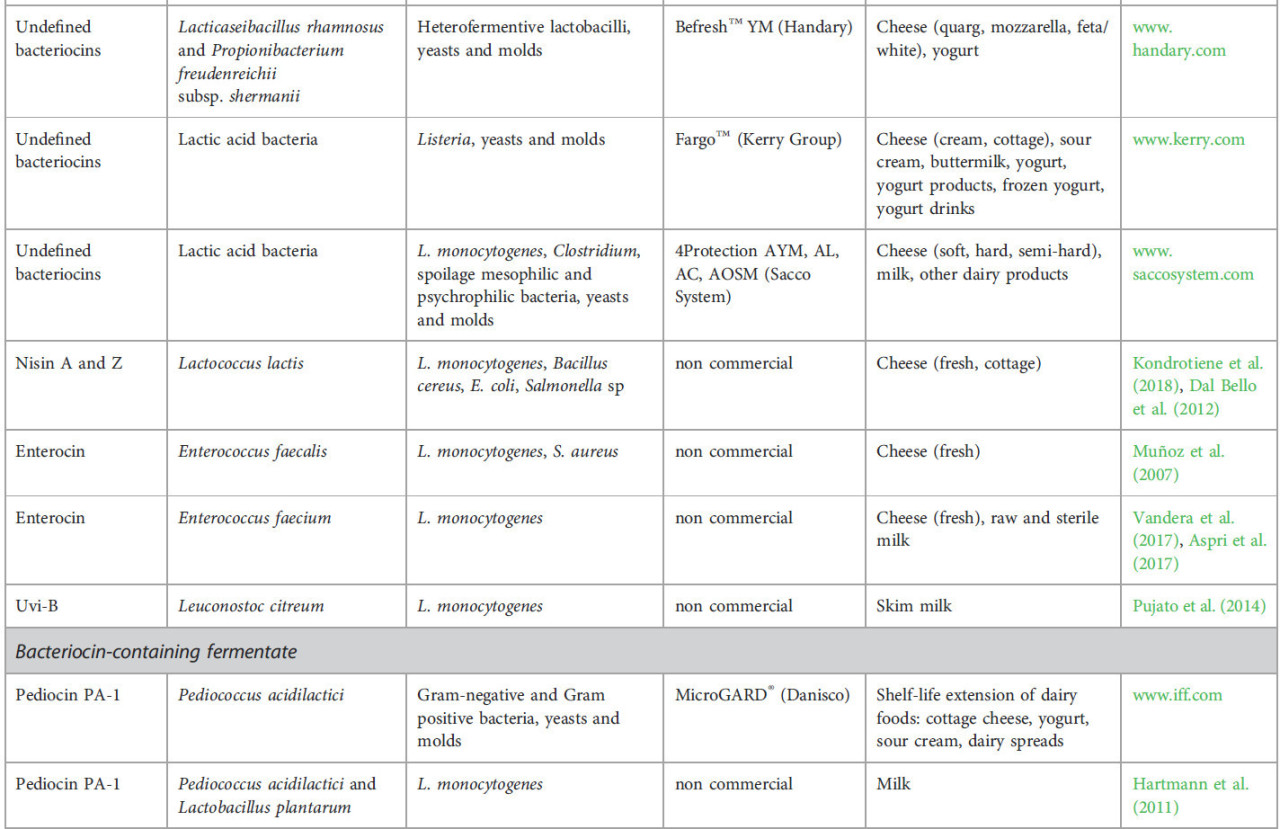

ТАБЛИЦА 1 Бактериоцины, применяемые в молочных продуктах.
2.2.1 Очищенные бактериоцины
Очищенные бактериоцины, используемые в молочных продуктах, предлагают некоторые преимущества по сравнению с использованием ферментированного продукта, содержащего бактериоцин, или продуцирующей бактериоцин LAB. Во-первых, процессы очистки обеспечивают удаление примесей, что приводит к более высокой концентрации активного бактериоцина. Эта повышенная активность позволяет более эффективно ингибировать патогенные и вызывающие порчу бактерии, тем самым увеличивая срок хранения и безопасность молочных продуктов. Более того, очищенные бактериоцины проявляют улучшенную стабильность и последовательность в своей антимикробной активности, что делает их надежными и предсказуемыми агентами для консервации пищевых продуктов (Leroy and De Vuyst, 2010). Кроме того, очистка облегчает удаление любого нежелательного привкуса или запаха, который может быть связан с неочищенными бактериоцинами, тем самым сохраняя аутентичный вкусовой профиль молочного продукта. В целом, использование очищенных бактериоцинов представляет собой превосходный подход к улучшению качества, безопасности и сенсорных характеристик молочных продуктов по сравнению с их неочищенными аналогами. Однако они требуют дорогостоящего одобрения регулирующих органов. На сегодняшний день низин является единственным коммерческим очищенным бактериоцином, который используется. Низин представляет собой бактериоцин типа IA, продуцируемый Lactococcus lactis (Egan et al., 2016; O’Connor et al., 2020). Было обнаружено более десяти вариантов низина. Низин A, F, Q и Z продуцируются L. lactis, низин U/U2, H, P и G продуцируются видами Streptococcus, тогда как низин J, O и кункецин A продуцируются другими бактериями (Sugrue et al., 2023). Низин имеет одобрение на коммерческое использование в более чем 60 странах в качестве пищевой добавки. Его коммерческое применение подпадает под регулирование Европейского управления по безопасности пищевых продуктов (EFSA), где он лицензирован как пищевой консервант (E234). Более того, низин признан безопасным биоконсервантом в пищевых продуктах Объединенным комитетом экспертов ФАО/ВОЗ по пищевым добавкам. В настоящее время коммерческий низин предлагается несколькими компаниями: Nisaplin® (Danisco), NisinA® (Handary) или Niseen® (Siveele) — вот некоторые примеры (Таблица 1). Низин эффективен против многочисленных грамположительных бактерий (включая LAB), патогенов, таких как Staphylococcus и Listeria, и спорообразующих бактерий, таких как Bacillus и Clostridium. Использование низина широко распространено: от охлажденного молока для борьбы с ростом патогенных Listeria monocytogenes и Staphylococcus aureus до различных сортов сыра для предотвращения позднего вспучивания — состояния, вызываемого видами Clostridium (Silva et al., 2018; O’Connor et al., 2020).
2.2.2 Продуцирующая бактериоцин LAB
Добавление LAB, продуцирующих бактериоцины, в пищевую матрицу не имеет нормативных ограничений и предлагает преимущество локализованного и непрерывного высвобождения антибактериальных агентов по мере размножения штамма. Кроме того, это менее дорогостояще, чем очищенные бактериоцины, поскольку нет необходимости в дорогостоящих процессах очистки (Pujato et al., 2014). Синтез бактериоцина зависит от различных факторов, включая условия окружающей среды и фазу роста бактерий. Эти переменные могут влиять на последовательность и эффективность активности бактериоцина в молочных продуктах. В совокупности бактерии, продуцирующие бактериоцины, могут также генерировать нежелательные метаболиты, которые могут повлиять на органолептические характеристики конечного продукта, такие как вкус, аромат и текстура. Хотя компании, производящие коммерческие закваски, гарантируют, что защитные культуры не оказывают негативного влияния на сенсорные характеристики продукта.
Существует несколько коммерчески доступных культур-продуцентов бактериоцинов (Таблица 1). BIOSAFE® (Chr. Hansen) содержит штамм, продуцирующий низин A, L. lactis subsp. lactis BS-10, который используется при производстве твердых и полутвердых сыров для предотвращения дефектов позднего вспучивания. HOLDBAC® (Danisco) — это серия защитных культур, содержащих неопределенные бактериоцины из Propionibacterium freudenreichii subsp. shermanii DSM 706 и Lacticaseibacillus rhamnosus DSM 7061. Они могут использоваться в различных продуктах, включая сыры, мясо, ферментированные овощи и напитки (O’Connor et al., 2020), генерируя различные метаболиты, включая бактериоцины, кислоты и другие полезные соединения, которые усиливают их способности к биоконтролю. Стоит отметить, что более 230 бактерий-продуцентов бактериоцинов были выделены и изучены в молочных продуктах (Silva et al., 2018).
2.2.3 Ферментаты, содержащие бактериоцины
Некоторые коммерческие пищевые ферментаты, содержащие бактериоцины, доступны для использования в молочной промышленности (Таблица 1). Этот инновационный антимикробный раствор получается путем культивирования штамма-продуцента бактериоцина в пищевой среде. Они предлагают многочисленные преимущества по сравнению с добавлением бактериоциногенных штаммов, такие как экономическая эффективность, минимальные нормативные препятствия и наличие других полезных метаболитов в ферментированном продукте. Кроме того, это позволяет регулировать наилучшие условия для продукции бактериоцина и минимизировать нежелательные побочные продукты (например, CO2). Более того, термическая обработка может деактивировать бактерии-продуценты бактериоцинов, не влияя на термостойкие бактериоцины, таким образом предотвращая конкуренцию со стартерными штаммами (Pujato et al., 2022). Однако требуется дополнительное время для производства ферментата отдельно. Несмотря на преимущества, этот дополнительный шаг может удлинить производственный процесс. Некоторыми примерами коммерчески доступных вариантов являются MicroGARD® (Danisco) и DuraFresh™ (Kerry Group P.L.C., Керри, Ирландия). MicroGARD® — это линейка ферментатов, содержащих бактериоцины и другие метаболиты, продуцируемые Pediococcus acidilactici, которая предлагает удобный способ введения антимикробных соединений в пищевые продукты, помогая продлить срок хранения молочных продуктов и шоколадных конфет (Makhal et al., 2015; Mills et al., 2017).
2.3 Оптимизация продукции бактериоцинов
Оптимальная продукция бактериоцинов зависит от тщательного контроля нескольких физических и питательных условий. Во-первых, время ферментации является ключевым для максимизации продукции бактериоцинов. Бактериоцины, обычно считающиеся первичными метаболитами, продуцируются главным образом во время экспоненциальной фазы роста бактерий. Скорость продукции этих метаболитов часто коррелирует со скоростью роста бактериальных клеток. Многочисленные исследования документируют бактериоцины, которые следуют этой закономерности, такие как низин A/Z и лактококцин 140 (Parente and Ricciardi, 1994), педиоцин PA1/SA-1 (Anastasiadou et al., 2008), мезентероцин 5 (Daba et al., 1993), лактоцин 705 (Vignolo et al., 1995), лактострепцин (De Vuyst, 1994) и лейкоцин (Geisen et al., 1993). Однако некоторые LAB продуцируют бактериоцины во время стационарной фазы, когда преобладают такие условия, как более низкий pH и сниженная доступность питательных веществ. Примеры включают Lactobacillus plantarum LPCO10 (Jimenez-Díaz et al., 1993), L. lactis subsp. lactis (Cheigh et al., 2002), Lactobacillus pentosus B96 (Delgado et al., 2005). Эта вариативность требует понимания кинетики роста и метаболических процессов для оптимизации (Sidooski et al., 2019).
Во-вторых, продукция бактериоцинов высокочувствительна к температуре. Температура напрямую влияет на ферментативную активность и скорость роста клеток микроорганизмов. Исследования предполагают, что оптимальная продукция бактериоцинов часто происходит при температурах ниже оптимальной температуры роста. Например, Leuconostoc mesenteroides MB1 (Pujato et al., 2022) и Streptococcus macedonicus ACA-DC 198 (Van den Berghe et al., 2006) лучше всего росли при 30°C и 42°C соответственно, но их продукция бактериоцинов достигала пика при 25°C. Однако это не универсальное правило. Некоторые штаммы LAB, такие как Enterococcus faecium RZS C5 (Leroy and De Vuyst, 2002), P. acidilactici 13 (Altuntas et al., 2010), не показали значительных различий в продукции бактериоцинов между оптимальными и субоптимальными температурами роста. Поэтому необходимо исследовать наилучшие температурные условия для каждого случая, чтобы максимизировать продукцию бактериоцинов.
Кроме того, pH влияет на ферментативную активность, скорость роста и продукцию метаболитов, включая бактериоцины. Высокая кислотность может ухудшать транспорт питательных веществ и основные цитоплазматические процессы, приводя к снижению роста клеток и синтеза бактериоцинов (Guerra, 2014). Чрезмерное подкисление, часто вызванное высокими уровнями молочной кислоты, отрицательно влияет на продукцию бактериоцинов, уменьшая высвобождение иммунных пептидов в растущих клетках. Иногда требуется подкисление среды для стимуляции выработки бактериоцина. В этих случаях буферизованный процесс отрицательно влиял бы на их продукцию (Guerra et al., 2008). Например, у Leuconostoc citreum MB1 оптимальный pH для продукции бактериоцинов был ниже (pH 5 и 6), чем тот, который требовался для максимального бактериального роста (pH 7) (Pujato et al., 2022). Поэтому для оптимизации продукции бактериоцинов крайне важно контролировать pH во время ферментации (Sidooski et al., 2019).
Одним из наиболее влиятельных факторов в продукции бактериоцинов LAB является культуральная среда. Добавление в среду ключевых питательных веществ, таких как углеводы, источники азота, витамины и соли, может значительно усилить продукцию бактериоцинов. Хотя сложные среды, такие как MRS, эффективны, они дорогостоящи и содержат белки, которые усложняют последующую обработку. Поэтому более простые среды с добавками предлагают экономически эффективную альтернативу (Garsa et al., 2014). Более того, оптимизация источников углерода, таких как глюкоза и лактоза, крайне важна, поскольку они влияют на продукцию ферментов и метаболическую активность. Однако слишком высокие концентрации могут ингибировать продукцию бактериоцинов из-за повышенных уровней молочной кислоты. Источники азота, такие как дрожжевой экстракт и триптон, также необходимы, обеспечивая аминокислотами и пептидами, которые способствуют росту клеток и синтезу бактериоцинов (Leães et al., 2011). Такие минералы, как магний и марганец, жизненно важны для ферментативной активности и общего метаболизма LAB, поддерживая структурную и функциональную целостность бактериоцинов. Определенные аминокислоты и витамины также играют роль в усилении продукции, действуя как предшественники и кофакторы в биосинтетических путях бактериоцинов (Moretro et al., 2000; Sidooski et al., 2019).
Условия ферментации значительно влияют на продукцию бактериоцинов. Периодическая ферментация, при которой все питательные вещества предоставляются в начале, проста, но может приводить к более низким выходам из-за истощения питательных веществ. Непрерывная ферментация, которая поддерживает постоянную подачу питательных веществ и удаляет отходы, часто приводит к более последовательному и высокому синтезу бактериоцинов (Guerra, 2014).
Оптимизация физических и питательных условий позволяет максимизировать продукцию бактериоцинов LAB, делая процесс более эффективным и экономически жизнеспособным для применений в консервации пищевых продуктов.
3 Заключение
Уже некоторое время наблюдается повышенный интерес к возврату к менее обработанным, не содержащим химикатов и более натуральным продуктам питания. В этом смысле использование бактериоцинов в пищевых продуктах может быть эффективной альтернативой химическим консервантам, которая может соответствовать современным стандартам здорового образа жизни. В зависимости от потребности, стоимости и типа продукта бактериоцины могут быть включены в пищевую матрицу в виде очищенного соединения, путем добавления бактериоциногенных LAB — с in situ и непрерывным высвобождением антимикробного соединения — или путем добавления ферментата, содержащего бактериоцин. Кроме того, оптимизация условий ферментации, включая время, температуру, pH и культуральную среду, крайне важна для максимизации продукции бактериоцинов, тем самым повышая качество и безопасность ферментированных молочных продуктов. Использование бактериоцинов в рамках барьерного подхода, включающего термические и нетермические обработки, может усилить биозащиту пищевых продуктов и уменьшить или устранить использование потенциально опасных химических консервантов.
Ссылки
1. Altuntas E. G., Cosansu S., Ayhan K. (2010). Some growth parameters and antimicrobial activity of a bacteriocin-producing strain Pediococcus acidilactici 13. Int. J. Food Microbiol. 141 (1–2), 28–31. doi: 10.1016/j.ijfoodmicro.2010.04.024. CrossRef. Google Scholar.
2. Alvarez-Sieiro P., Montalbán-López M., Mu D., Kuipers O. P. (2016). Bacteriocins of lactic acid bacteria: extending the family. Appl. Microbiol. Biotechnol. 100 (7), 2939–2951. doi: 10.1007/s00253-016-7343-9. CrossRef. Google Scholar.
3. Anastasiadou S., Papagianni M., Filiousis G., Ambrosiadis I., Koidis P. (2008). Growth and metabolism of a meat isolated strain of Pediococcus pentosaceus in submerged fermentation. Enzyme Microb. Technol. 43 (6), 448–454. doi: 10.1016/j.enzmictec.2008.05.007. CrossRef. Google Scholar.
4. Arnison P. G., Bibb M. J., Bierbaum G., Bowers A. A., Bugni T. S., Bulaj G., et al. (2013). Ribosomally synthesized and post-translationally modified peptide natural products: overview and recommendations for a universal nomenclature. Nat. Prod. Rep. 30, 108–160. doi: 10.1039/c2np20085f. CrossRef. Google Scholar.
5. Aspri M., O’Connor P. M., Field D., Cotter P. D., Ross P., Hill C., et al. (2017). Application of bacteriocin-producing Enterococcus faecium isolated from donkey milk, in the bio-control of Listeria monocytogenes in fresh whey cheese. Int. Dairy J. 73, 1–9. doi: 10.1016/j.idairyj.2017.04.008. CrossRef. Google Scholar.
6. Carpentier B., Cerf O. (2011). Persistence of Listeria monocytogenes in food industry equipment and premises. Int. J. Food Microbiol. 145 (1), 1–8. doi: 10.1016/j.ijfoodmicro.2011.01.005. CrossRef. Google Scholar.
7. Cheigh C.-I., Choi H.-J., Park H., Kim S.-B., Kook M.-C., Kim T.-S., et al. (2002). Influence of growth conditions on the production of a nisin-like bacteriocin by Lactococcus lactis subsp. lactis A164 isolated from kimchi. J. Biotechnol. 95 (3), 225–235. doi: 10.1016/S0168-1656(02)00010-X. CrossRef. Google Scholar.
8. Cotter P. D., Hill C., Ross R. P. (2005). Bacteriocins: developing innate immunity for food. Nat. Rev. Microbiol. 3, 777–788. doi: 10.1038/nrmicro1273. CrossRef. Google Scholar.
9. Cotter P. D., Ross R. P., Hill C. (2013). Bacteriocins - a viable alternative to antibiotics? Nat. Rev. Microbiol. 11, 95–105. doi: 10.1038/nrmicro2937. CrossRef. Google Scholar.
10. Daba G. M., Elkhateeb W. A. (2020). Bacteriocins of lactic acid bacteria as biotechnological tools in food and pharmaceuticals: current applications and future prospects. Biocatal. Agric. Biotechnol. 28, 101750. doi: 10.1016/j.bcab.2020.101750. CrossRef. Google Scholar.
11. Daba H., Lacroix C., Huang J., Simard R. (1993). Influence of growth conditions on production and activity of mesenterocin 5 by a strain of Leuconostoc mesenteroides. Appl. Microbiol. Biotechnol. 39 (2), 166–173. doi: 10.1007/BF00228601. CrossRef. Google Scholar.
12. Dal Bello B., Cocolin L., Zeppa G., Field D., Cotter P. D., Hill C. (2012). Technological characterization of bacteriocin producing Lactococcus lactis strains employed to control Listeria monocytogenes in cottage cheese. Int. J. Food Microbiol. 153, 58–65. doi: 10.1016/j.ijfoodmicro.2011.10.016. CrossRef. Google Scholar.
13. Deegan L. H., Cotter P. D., Hill C., Ross P. (2006). Bacteriocins: biological tools for biopreservation and shelf-life extension. Int. Dairy J. 16 (9), 1058–1071. doi: 10.1016/j.idairyj.2005.10.026. CrossRef. Google Scholar.
14. Delgado A., Brito D., Peres C., Noé-Arroyo F., Garrido-Fernández A. (2005). Bacteriocin production by Lactobacillus pentosus B96 can be expressed as a function of temperature and NaCl concentration. Food Microbiol. 22 (6), 521–528. doi: 10.1016/j.fm.2004.11.015. CrossRef. Google Scholar.
15. De Vuyst L. (1994). Lactostrepcins, bacteriocins produced by Lactococcus lactis strains. In De Vuyst L., Vandamme E. J. (Eds.), Bacteriocins of lactic acid bacteria (pp. 291–299). Boston, MA: Springer. doi: 10.1007/978-1-4615-2668-1_9. CrossRef. Google Scholar.
16. Egan K., Field D., Rea M. C., Ross R. P., Hill C., Cotter P. D. (2016). Bacteriocins: novel solutions to age old spore-related problems? Front. Microbiol. 7, 461. doi: 10.3389/fmicb.2016.00461. CrossRef. Google Scholar.
17. Favaro L., Barretto Penna A. L., Todorov S. D. (2015). Bacteriocinogenic LAB from cheeses -- application in biopreservation? Trends Food Sci. Technol. 41 (1), 37–48. doi: 10.1016/j.tifs.2014.09.001. CrossRef. Google Scholar.
18. Garsa A. K., Kumariya R., Sood S. K., Kumar A., Kapila S. (2014). Bacteriocin production and different strategies for their recovery and purification. Probiotics Antimicrob. Proteins 6 (1), 47–58. doi: 10.1007/s12602-013-9153-z. CrossRef. Google Scholar.
19. Geisen R., Becker B., Holzapfel W. H. (1993). Bacteriocin production of Leuconostoc carnosum LA54A at different combinations of pH and temperature. J. Ind. Microbiol. 12 (3–5), 337–340. doi: 10.1007/BF01584211. CrossRef. Google Scholar.
20. Guerra N. P. (2014). Modeling the batch bacteriocin production system by lactic acid bacteria by using modified three-dimensional Lotka-Volterra equations. Biochem. Eng. J. 88, 115–130. doi: 10.1016/j.bej.2014.04.010. CrossRef. Google Scholar.
21. Guerra N. P., Bernárdez P. F., Castro L. P. (2008). Modelling the stress inducing biphasic growth and pediocin production by Pediococcus acidilactici NRRL B-5627 in re-alkalized fed-batch cultures. Biochem. Eng. J. 40 (3), 465–472. doi: 10.1016/j.bej.2008.02.001. CrossRef. Google Scholar.
22. Hartmann H., Wilke T., Erdmann R. (2011). Efficacy of bacteriocin-containing cell-free culture supernatants from lactic acid bacteria to control Listeria monocytogenes in food. Int. J. Food Microbiol. 146, 192–199. doi: 10.1016/j.ijfoodmicro.2011.02.031. CrossRef. Google Scholar.
23. Jimenez-Díaz R., Rios-Sánchez R. M., Desmazeaud M., Ruiz-Barba J. L., Piard J. C. (1993). Plantaricins S and T, two new bacteriocins produced by Lactobacillus plantarum LPCO10 isolated from a green olive fermentation. Appl. Environ. Microbiol. 59 (5), 1416–1424. doi: 10.1128/aem.59.5.1416-1424.1993. CrossRef. Google Scholar.
24. Kondrotiene K., Kasnauskyte N., Serniene L., Gölz G., Alter T., Kaskoniene V., et al. (2018). Characterization and application of newly isolated nisin producing Lactococcus lactis strains for control of Listeria monocytogenes growth in fresh cheese. LWT Food Sci. Technol. 87, 507–514. doi: 10.1016/j.lwt.2017.09.021. CrossRef. Google Scholar.
25. Leães F. L., Vanin N. G., Sant’anna V., Brandelli A. (2011). Use of byproducts of food industry for production of antimicrobial activity by Bacillus sp. P11. Food Bioprocess Tech. 4, 822–828. doi: 10.1007/s11947-010-0410-9. CrossRef. Google Scholar.
26. Leroy F., De Vuyst L. (2002). Bacteriocin production by Enterococcus faecium RZS C5 is cell density limited and occurs in the very early growth phase. Int. J. Food Microbiol. 72 (1–2), 155–164. doi: 10.1016/S0168-1605(01)00635-3. CrossRef. Google Scholar.
27. Leroy F., De Vuyst L. (2010). Bacteriocins of lactic acid bacteria to combat undesirable bacteria in dairy products. Aust. J. Dairy Technol. 65, 143–149. Google Scholar.
28. Liu L., O’Conner P., Cotter P. D., Hill C., Ross R. P. (2008). Controlling Listeria monocytogenes in Cottage cheese through heterologous production of enterocin A by Lactococcus lactis. J. Appl. Microbiol. 104 (4), 1059–1066. doi: 10.1111/j.1365-2672.2007.03640.x. CrossRef. Google Scholar.
29. Makhal S., Kanawjia S. K., Giri A. (2015). Effect of microGARD on keeping quality of direct acidified Cottage cheese. J. Food Sci. Technol. 52 (2), 936–943. doi: 10.1007/s13197-013-1055-2. CrossRef. Google Scholar.
30. Mills S., Ross R. P., Hill C. (2017). Bacteriocins and bacteriophage; a narrow-minded approach to food and gut microbiology. FEMS Microbiol. Rev. 41, S129-S153. doi: 10.1093/femsre/fux022. CrossRef. Google Scholar.
31. Moretro T., Aasen I. M., Storro I., Axelsson L. (2000). Production of sakacin P by Lactobacillus sakei in a completely defined medium. J. Appl. Microbiol. 88 (3), 536–545. doi: 10.1046/j.1365-2672.2000.00994.x. CrossRef. Google Scholar.
32. Muñoz A., Ananou S., Gálvez A., Martínez-Bueno M., Rodríguez A., Maqueda M., et al. (2007). Inhibition of Staphylococcus aureus in dairy products by enterocin AS-48 produced in situ and ex situ: bactericidal synergism with heat. Int. Dairy J. 17, 760–769. doi: 10.1016/j.idairyj.2006.09.006. CrossRef. Google Scholar.
33. O’Connor P. M., Kuniyoshi T. M., Oliveira R. P. S., Hill C., Ross R. P., Cotter P. D. (2020). Antimicrobials for food and feed; a bacteriocin perspective. Curr. Opin. Biotechnol. 61, 160–167. doi: 10.1016/j.copbio.2019.12.023. CrossRef. Google Scholar.
34. O’Sullivan L., Ross R. P., Hill C. (2002). Potential of bacteriocin-producing lactic acid bacteria for improvements in food safety and quality. Biochimie 84, 593–604. doi: 10.1016/s0300-9084(02)01457-8. CrossRef. Google Scholar.
35. Parente E., Ricciardi A. (1994). Influence of pH on the production of enterocin 1146 during batch fermentation. Lett. Appl. Microbiol. 19 (1), 12–15. doi: 10.1111/j.1472-765X.1994.tb00891.x. CrossRef. Google Scholar.
36. Pucci M. J., Vedamuthu E. R., Kunka B. S., Vandenbergh P. A. (1988). Inhibition of Listeria monocytogenes by using bacteriocin PA-1 produced by Pediococcus acidilactici PAC 1.0. Appl. Environ. Microbiol. 54, 2349–2353. doi: 10.1128/AEM.54.10.2349-2353.1988. CrossRef. Google Scholar.
37. Pujato S. A., Quiberoni A. del L., Candioti M. C., Reinheimer J. A., Guglielmotti D. M. (2014). Leuconostoc citreum MB1 as biocontrol agent of Listeria monocytogenes in milk. J. Dairy Res. 81, 137–145. doi: 10.1017/S002202991300068X. CrossRef. Google Scholar.
38. Pujato S. A., Quiberoni A. del L., Guglielmotti D. M. (2022). Characterization of bacteriocins produced by lactic acid bacteria of industrial interest. In Gupta V. K., Sarker S. D., Sharma M., Pirovani M. E., Usmani Z., Jayabaskaran C. (Eds.), Biomolecules from natural sources (pp. 458–469). John Wiley and Sons, Inc. doi: 10.1002/9781119769620.ch16. CrossRef. Google Scholar.
39. Ribeiro S. C., O’Connor P. M., Ross R. P., Stanton C., Silva C. C. (2016). An anti-listerial Lactococcus lactis strain isolated from Azorean Pico cheese produces lacticin 481. Int. Dairy J. 63, 18–28. doi: 10.1016/j.idairyj.2016.07.017. CrossRef. Google Scholar.
40. Ribeiro S. C., Ross R. P., Stanton C., Silva C. C. (2017). Characterization and application of antilisterial enterocins on model fresh cheese. J. Food Prot. 80, 1303–1316. doi: 10.4315/0362-028X.JFP-17-031. CrossRef. Google Scholar.
41. Reuben R. C., Torres C., Yildirim (2024). Bacteriocins: potentials and prospects in health and agrifood systems. Arch. Microbiol. 206 (5), 233. doi: 10.1007/s00203-024-03948-y. CrossRef. Google Scholar.
42. Sidooski T., Brandelli A., Bertoli S. L., Krebs de Souza C., Fernandes de Carvalho L. (2019). Physical and nutritional conditions for optimized production of bacteriocins by lactic acid bacteria -- a review. Crit. Rev. Food Sci. Nutr. 59 (17), 2839–2849. doi: 10.1080/10408398.2018.1474852. CrossRef. Google Scholar.
43. Silva C. C. G., Silva S. P. M., Ribeiro S. C. (2018). Application of bacteriocins and protective cultures in dairy food preservation. Front. Microbiol. 9, 594. doi: 10.3389/fmicb.2018.00594. CrossRef. Google Scholar.
44. Simons A., Alhanout K., Duval R. E. (2020). Bacteriocins, antimicrobial peptides from bacterial origin: overview of their biology and their impact against multidrug-resistant bacteria. Microorganisms 8 (5), 639. doi: 10.3390/microorganisms8050639. CrossRef. Google Scholar.
45. Soltani S., Hammami R., Cotter P. D., Rebuffat S., Said L. B., Gaudreau H., et al. (2021). Bacteriocins as a new generation of antimicrobials: toxicity aspects and regulations. FEMS Microbiol. Rev. 8 (1), fuaa039. doi: 10.1093/femsre/fuaa039. CrossRef. Google Scholar.
46. Sugrue I., Hill D., O’Connor P. M., Day L., Stanton C., Hill C., et al. (2023). Nisin E is a novel nisin variant produced by multiple Streptococcus equinus strains. Microorganisms 11 (2), 427. doi: 10.3390/microorganisms11020427. CrossRef. Google Scholar.
47. Van den Berghe E., Skourtas G., Tsakalidou E., De Vuyst L. (2006). Streptococcus macedonicus ACA-DC 198 produces the lantibiotic, macedocin at temperature and pH conditions that prevail during cheese manufacture. Int. J. Food Microbiol. 107 (2), 138–147. doi: 10.1016/j.ijfoodmicro.2005.08.023. CrossRef. Google Scholar.
48. Vandera E., Lianou A., Kakouri A., Feng J., Koukkou A.-I., Samelis J. (2017). Enhanced control of Listeria monocytogenes by Enterococcus faecium KE82, a multiple enterocin-producing strain, in different milk environments. J. Food Prot. 80, 74–85. doi: 10.4315/0362-028X.JFP-16-082. CrossRef. Google Scholar.
49. Verma S. K., Sood S. K., Saini R. K., Saini N. (2017). Pediocin PA-1 containing fermented cheese whey reduces total viable count of raw buffalo (Bubalis bubalus) milk. LWT Food Sci. Technol. 83, 193–200. doi: 10.1016/j.lwt.2017.02.031. CrossRef. Google Scholar.
50. Vignolo G. M., de Kairuz M. N., de Ruiz Holgado A. A. P., Oliver G. (1995). Influence of growth conditions on the production of lactocin 705, a bacteriocin produced by Lactobacillus casei CRL 705. J. Appl. Bacteriol. 78 (1), 5–10. doi: 10.1111/j.1365-2672.1995.tb01665.x. CrossRef. Google Scholar.
51. Yildirim Z., Öncül N., Yildirim M., Karabiyikli S. (2016). Application of lactococcin BZ and enterocin KP against Listeria monocytogenes in milk as biopreservation agents. Acta Aliment. 45, 486–492. doi: 10.1556/066.2016.45.4.4. CrossRef. Google Scholar.

Pujato SA, Mercanti DJ, Briggiler Marcó M, Capra ML, Quiberoni A and Guglielmotti DM (2024) Bacteriocins from lactic acid bacteria: strategies for the bioprotection of dairy foods. Front. Food. Sci. Technol. 4:1439891. doi: 10.3389/frfst.2024.1439891
Перевод статьи «Bacteriocins from lactic acid bacteria: strategies for the bioprotection of dairy foods» авторов Pujato SA, Mercanti DJ, Briggiler Marcó M, Capra ML, Quiberoni A and Guglielmotti DM., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык
Фото: magnific

















































































































Комментарии (0)