Растительный напиток с уваи: как пробиотик выживает в кишечнике
За последние годы вырос спрос на растительные продукты в качестве альтернативы традиционному питанию. В соответствии с этим, в данной работе исследуется производство и переваривание in vitro ферментированного напитка на растительной основе (ФНРО), приготовленного из белков гороха и риса, с добавлением 0% (ФНРО-К), 5% (ФНРО-5) и 10% (ФНРО-10) мякоти уваи путем молочнокислого брожения с использованием Lacticaseibacillus rhamnosus GG. Процесс моделирования желудочно-кишечного пищеварения in vitro был проведен для оценки биодоступности L. rhamnosus GG, общего содержания фенольных соединений (ОСФ) и антиоксидантной активности до и после воздействия условий желудочно-кишечного тракта.
Аннотация
После 48 часов пищеварения в желудочно-кишечном тракте сохранялось высокое количество жизнеспособных клеток L. rhamnosus GG. Образцы ФНРО-К (106,89%) и ФНРО-5 (109,38%) показали более высокую выживаемость, чем ФНРО-10 (102,20%), что указывает на более высокий потенциальный пребиотический эффект этих напитков. По сравнению с непереваренными образцами, после 48 часов пищеварения во всех образцах наблюдалось значительное увеличение ОСФ. Аналогичное поведение отмечено и для антиоксидантной активности ФНРО-К, ФНРО-5 и ФНРО-10 по методам DPPH (4,06; 3,96 и 8,44 мг TEAC мл⁻¹), ABTS (10,28; 11,06; 11,97 мг TEAC мл⁻¹) и FRAP (917,02; 863,87 и 1983,23 мг TEAC мл⁻¹). С помощью ВЭЖХ-ДМД-ИЭР-МС в мякоти уваи было идентифицировано и количественно определено тринадцать соединений, особенно эпигаллокатехин галлат, кверцетин-3-рамноза и кверцетин-3-глюкозид. Изорамнетин был основным фенольным соединением, обнаруженным в толстой кишке, предположительно вследствие конверсии кверцетин-3-глюкозида клетками пробиотика.
В заключение, поскольку все показатели жизнеспособности превышали 9 log КОЕ г⁻¹, составы ФНРО, содержащие белок гороха, риса и мякоть уваи, становятся перспективным носителем для L. rhamnosus GG.
1 Введение
Во всем мире растущий интерес к здоровому образу жизни стимулирует рынок функциональных продуктов питания (Camelo-Silva et al., 2024; Galanakis, 2021; Sorita et al., 2022). В этом контексте растительные напитки, такие как напитки из злаков, орехов и бобовых, ферментированные с пробиотиками, исследуются для производства безмолочных напитков, становясь значительным сегментом в рамках функциональных продуктов питания (Popova et al., 2023). Благодаря своим полезным для здоровья свойствам, предпочтение растительных диет не ограничивается вегетарианцами, людьми с непереносимостью лактозы или аллергией на молочный белок, но распространяется и на других потребителей, включая флекситарианцев (лица, чей рацион в основном состоит из растительной пищи с эпизодическим потреблением продуктов животного происхождения) (Sheen et al., 2023). Эта тенденция вывела рынок безмолочных альтернатив на уровень 29,18 млрд долларов США в 2023 году с прогнозируемым ежегодным ростом в 12,6% к 2030 году. Этот рост направлен на проведение исследований и инноваций в секторе для удовлетворения растущего потребительского спроса (Grand View Research, 2024).
Наиболее потребляемыми растительными белками и продуктами их переработки являются белки пшеницы и сои. Однако оба обладают аллергенным потенциалом, что стимулирует поиск других растительных матриц со сниженными гипоаллергенными характеристиками (Reese et al., 2023). Такие злаки, как сорго и кукуруза, а также бобовые, например фасоль и чечевица, широко используются в качестве альтернатив для создания носителей пробиотиков благодаря их низкой аллергенности и приемлемому аминокислотному составу (Popova et al., 2023; Sheen et al., 2023; Reese et al., 2023; Zhang et al., 2023). Эти богатые белком продукты служат обильным источником макро- и микронутриентов, способствуя росту и значительно повышая жизнеспособность пробиотических микроорганизмов. Белки гороха и риса также являются отличными заменителями аллергенных белков и сохраняют свою доступность и адекватную пищевую ценность. Комбинация белка гороха и риса обеспечивает поступление adequate количеств всех незаменимых аминокислот в рацион человека (Lu et al., 2020). Эти белки являются текущей тенденцией на рынке растительных продуктов, что побуждает исследователей проводить частые исследования (Lu et al., 2020; Manus et al., 2021).
Кроме того, добавление фруктовой мякоти в ферментированные растительные напитки (ФРН) может быть интересной альтернативой для улучшения сенсорных и питательных свойств. Фруктовые мякоти обычно добавляют в напитки, однако native бразильские фрукты все еще мало изучены (Bianchini et al., 2020). Бразилия имеет несколько биомов, включая Атлантический лес, который предоставляет питательные нетрадиционные фрукты, такие как уваи (Eugenia pyriformis), с мало изученным экономическим потенциалом (Sganzerla et al., 2021). Уваи принадлежит к семейству Миртовые (Myrtaceae), имеет округлую, сплюснутую и грушевидную форму с мягкой или твердой текстурой и окраской от желтых до оранжевых тонов; ее аромат приятен со сладкими и кислыми нотами (Bianchini et al., 2020; Spricigo et al., 2023; Rodrigues et al., 2021). Что касается питательных аспектов, уваи признана за ее высокую пищевую ценность и содержание биоактивных соединений (включая галловую, хлорогеновую, п-кумаровую и феруловую кислоты, среди прочих) (Farias et al., 2021). Этот фрукт содержит высокое содержание витамина С и А (Farias et al., 2021), а также аминокислоты: аланин, аргинин, аспарагин, холин, глутамин, изолейцин, лейцин, фенилаланин, пролин, серин, треонин, тирозин и валин (Spricigo et al., 2023). Благодаря такому широкому спектру полезных соединений, исследование in vivo показало, что употребление сока уваи самками крыс показало снижение окислительного повреждения, улучшение антиоксидантной эффективности и ее полезного потенциала для здоровья (Lopes et al., 2018). Другое исследование in vivo также продемонстрировало противовоспалительную и антиоксидантную активность сока мякоти уваи на самцах крыс линии Wistar (Ramirez et al., 2012). Учитывая эти высокие полезные для здоровья свойства, мякоть уваи представляет собой интересный и инновационный пищевой продукт для обогащения сенсорных и функциональных свойств ФРН из гороха и риса.
Ферментация является ключевым этапом в производстве ФРН. В процессе ферментации молочнокислыми бактериями (МКБ) белки гороха и риса, обогащенные мякотью уваи, могут продуцировать различные вторичные метаболиты, которые могут оказывать пребиотическое действие, положительно изменяя в различной степени другие питательные и биоактивные профили напитков посредством ферментативной и микробной активности (Masiá et al., 2021; Cuvas-Limón et al., 2022). Некоторые МКБ могут продуцировать экзополисахариды, которые улучшают стабильность напитков, позволяя заменить загустители. Более того, при использовании пробиотических микроорганизмов можно создать функциональный напиток за счет подкисления и генерировать антимикробные соединения, которые конкурируют с нежелательными микроорганизмами (Alcorta et al., 2021). L. rhamnosus GG был использован для приготовления различных напитков, таких как соевый, овсяный и кокосовый (Masiá et al., 2021), миндальный с инулином (Muncey and Hekmat, 2021), пшеничные хлебобулочные изделия (Sigüenza-Andrés et al., 2023) и других. Недавнее исследование было направлено на разработку и оценку in vitro перевариваемости белка и физико-химических свойств функционального напитка, обогащенного белками риса и гороха (Manus et al., 2021). Авторы наблюдали снижение вязкости и уменьшение количества пептидов с высокой молекулярной массой, связанное с увеличением количества пептидов с низкой молекулярной массой, что привело к заметному улучшению перевариваемости. Однако в литературных данных не было найдено ни одного исследования, касающегося производства ферментированных напитков на основе белка риса и гороха, обогащенных мякотью уваи.
Тем не менее, чтобы биоактивные соединения и пробиотическая культура в ФРН могли проявлять свою биологическую активность и предоставлять пользу для здоровья, они должны высвобождаться из пищевой матрицы и оставаться стабильными в желудочно-кишечных условиях (Camelo-Silva et al., 2024; Tang et al., 2023). Следовательно, оценка биоактивности обогащенных пищевых продуктов после прохождения желудочно-кишечного пищеварения является ключевой. Соответственно, цель настоящего исследования была направлена на разработку ферментированного напитка на растительной основе с использованием белков гороха и риса посредством процесса молочнокислой ферментации с L. rhamnosus GG, обогащенного мякотью уваи, и анализ его пробиотических и пребиотических эффектов. Для этого выживаемость L. rhamnosus GG, общее содержание фенольных соединений, антиоксидантная активность и химический состав (с помощью ВЭЖХ-ИЭР-МС/МС) были оценены до и во время моделирования in vitro желудочно-кишечного пищеварения.
2 Материалы и методы
2.1 Материалы
Для приготовления ферментированного напитка использовали изолированный рисовый белок (80 г белка на 100 г⁻¹, VeganWay, Флорианополис, SC, Бразилия), изолированный гороховый белок (80 г белка на 100 г⁻¹, VeganWay, Флорианополис, SC, Бразилия), лиофилизированную пробиотическую культуру, состоящую из L. rhamnosus LGG® (Lr) (Chr. Hansen, Хёнсхольм, Дания), и мякоть уваи (Eugenia pyriformis) (влажность 89,4%, белок 1,0%, жир 0,25%, углеводы 9,05% и зола 0,30%). Для микробиологического анализа использовали бульон MRS (Man, Rogosa, Sharpe) и агар MRS (Kasvi, Сан-Жозе-дус-Пиньяйс, PR, Бразилия). Для моделирования in vitro желудочно-кишечного тракта использовали реагенты: ферменты α-амилазу, пепсин из слизистой оболочки желудка свиньи, панкреатин свиньи и бычьи соли желчных кислот, все приобретенные у Sigma-Aldrich (Сан-Паулу, SP, Бразилия). Для определения жизнеспособности пробиотических бактерий использовали набор BacTiter-Glo™ Viability Kit (Promega, Мэдисон, Висконсин, США). Реактив Фолина-Чокальтеу использовали в анализе общего содержания фенольных соединений (ОСФ). Для антиоксидантной активности по захвату радикалов DPPH и ABTS использовали радикалы DPPH (2,2-дифенил-1-пикрилгидразил) и ABTS (диаммониевая соль 2,2-азино-бис(3-этилбензотиазолин-6-сульфоновой кислоты)) соответственно. TPTZ (2,4,6-Трис(2-пиридил)-s-триазин) и хлорид железа использовали для антиоксидантной способности по методу восстановления железа FRAP. Галловая кислота была использована в качестве стандартного образца для ОСФ, в то время как Trolox (6-Гидрокси-2,5,7,8-тетраметилхроман-2-карбоновая кислота) был использован в качестве стандартного образца для всех анализов антиоксидантной активности. Все реагенты для ОСФ и антиоксидантной активности были приобретены у Sigma-Aldrich (Сан-Паулу, SP, Бразилия). Все использованные химические вещества были аналитической чистоты.
2.2 Методы
2.2.1 Приготовление ферментированного напитка на растительной основе (ФРН)
Пробиотический ФРН готовили в соответствии с методологией, описанной Deziderio et al. (2023), с модификациями. Сначала 1,83% рисового белка, 1,83% горохового белка, 10% сахарозы, 0,32% соевого масла и 0,2% разведенного соевого лецитина растворяли в дистиллированной воде и гомогенизировали при 13 000 об/мин (Ultra-Turrax, IKA™ T25 digital) в течение 3 мин. Затем инокулировали лиофилизированную пробиотическую культуру, содержащую 10⁹ колониеобразующих единиц (КОЕ) на грамм L. rhamnosus LGG® (Chr. Hansen, Хёнсхольм, Дания), начиная стадию ферментации. Ферментацию проводили при 37°C ± 2°C до достижения pH 4,8. После этого стадию ферментации останавливали охлаждением среды до 4°C ± 1°C (в холодильнике, Consul, Бразилия). Мякоть уваи добавляли в составы после процесса ферментации. Было приготовлено три состава ферментированного напитка: i) без добавления мякоти уваи – контрольный образец (ФРН-К), и с добавлением ii) 5% (ФРН-5) и iii) 10% (ФРН-10) пастеризованной мякоти уваи (80°C в течение 3 мин) соответственно. Образцы хранили в охлажденном состоянии (4°C ± 1°C) до анализа. Образцы были приготовлены в трех повторностях.
2.2.2 Моделирование in vitro желудочно-кишечного пищеварения
Моделирование in vitro желудочно-кишечного пищеварения проводили в соответствии с протоколом INFOGEST (Verruck et al., 2020; Brodkorb et al., 2019) с модификациями. Протокол моделирует пищеварение последовательно в полости рта, желудке, тонком и толстом кишечнике, как показано на Рисунке 1.
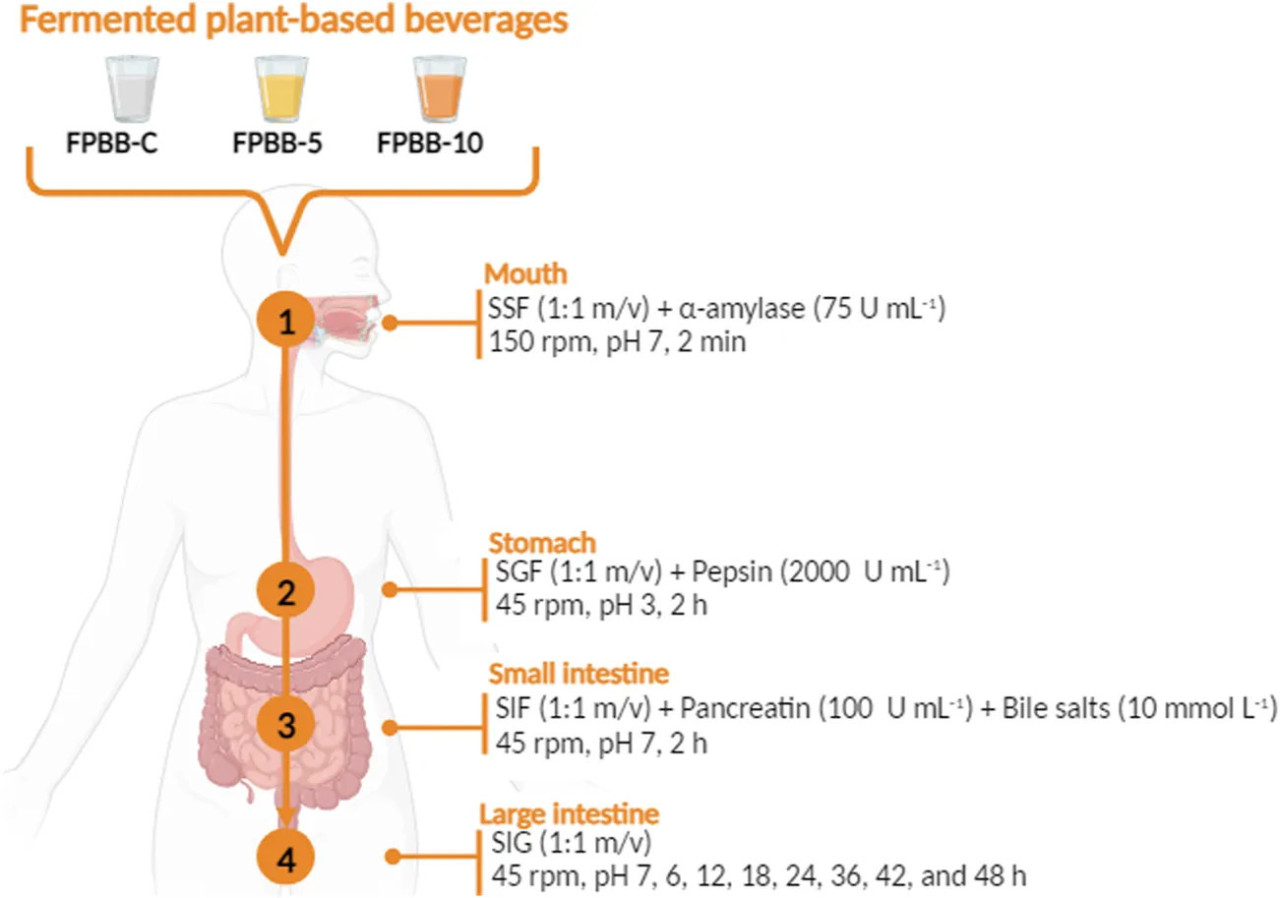
РИСУНОК 1 Блок-схема протокола моделирования in vitro желудочно-кишечного пищеварения, использованного для оценки биодоступности L. rhamnosus GG, ОСФ и антиоксидантной активности ферментированных напитков на растительной основе (ФРН-К, ФРН-5 и ФРН-10). ФРН-К: ферментированный напиток без мякоти уваи; ФРН-5: ферментированный напиток, обогащенный 5% мякоти уваи; ФРН-10: ферментированный напиток, обогащенный 10% мякоти уваи).
Составы моделируемых жидкостей (SSF — моделируемая слюнная жидкость, SGF — моделируемая желудочная жидкость, SIF — моделируемая кишечная жидкость и SFL — моделируемая жидкость толстой кишки), использованных в этом протоколе, описаны в Таблице 1.
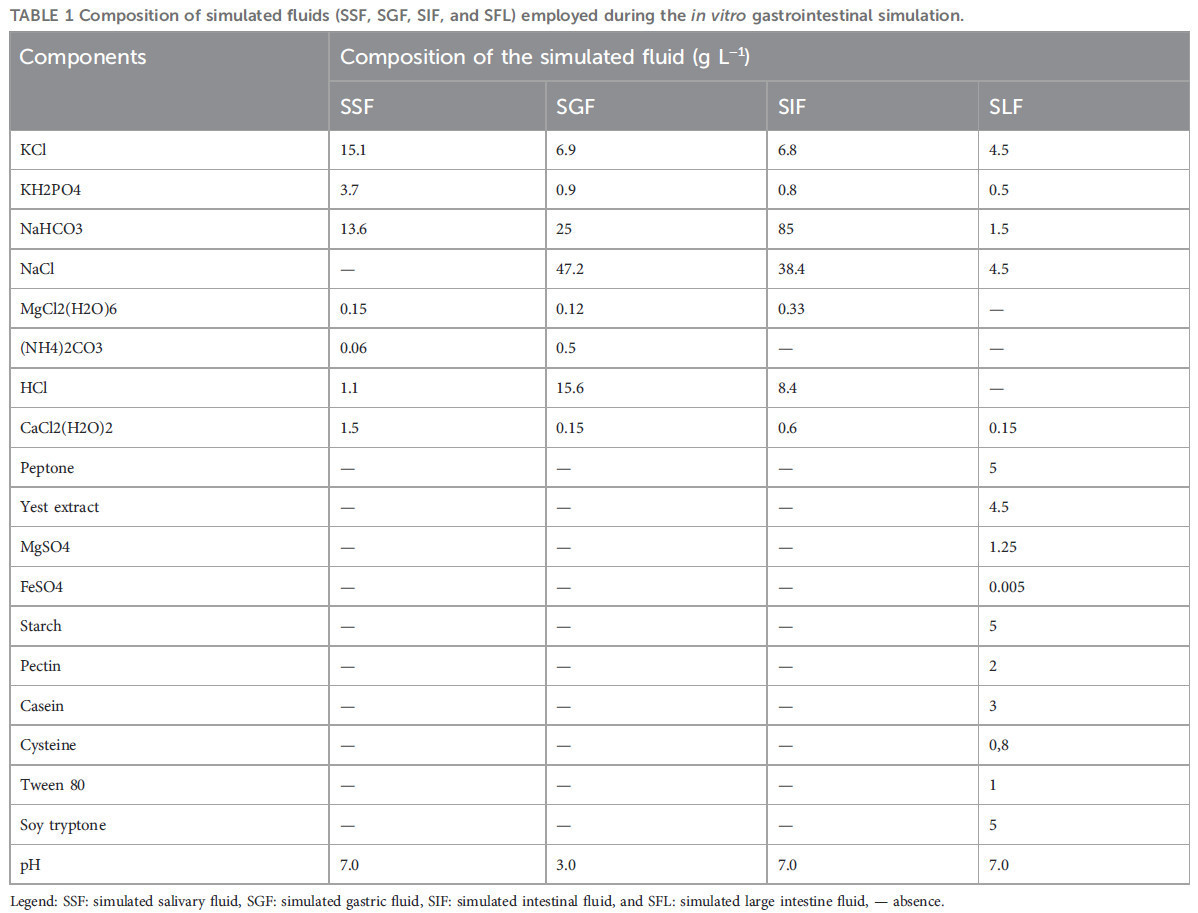
ТАБЛИЦА 1 Состав моделируемых жидкостей (SSF, SGF, SIF и SFL), использованных во время моделирования in vitro желудочно-кишечного пищеварения.
Легенда: SSF: моделируемая слюнная жидкость, SGF: моделируемая желудочная жидкость, SIF: моделируемая кишечная жидкость и SFL: моделируемая жидкость толстой кишки, — отсутствие.
Для первого этапа желудочно-кишечного тракта (моделирование полости рта) по 5 г каждого образца (ФРН-К, ФРН-5 и ФРН-10) сначала гомогенизировали с SSF в пропорции 1:1 (м/о). Затем добавляли раствор человеческой слюнной α-амилазы (75 Ед мл⁻¹), и смесь инкубировали в водяной бане при 37°C при 150 об/мин в течение 2 мин.
После этого для моделирования желудочной фазы пероральный болюс смешивали с SGF 1:1 (об/об) (состав представлен в Таблице 1) и свиным пепсином (2000 Ед мл⁻¹), и pH доводили с помощью HCl (1 моль л⁻¹) до pH 3,0 (желудочный болюс). Затем смесь инкубировали при 37°C при осторожном перемешивании (100 об/мин) в течение 2 ч.
Впоследствии моделировали пищеварение в тонком кишечнике. Для этого желудочный болюс смешивали с SIF 1:1 (об/об) (с составом, описанным в Таблице 1), панкреатином (100 Ед мл⁻¹, исходя из активности трипсина) и солями желчных кислот (10 ммоль л⁻¹), предварительно нагретыми в водяной бане при 37°C. Затем pH доводили до 7,0 добавлением NaOH (1 моль л⁻¹). Раствор инкубировали при 37°C при скорости перемешивания 100 об/мин в течение 2 ч.
После прохождения через тонкий кишечник ферментированный напиток на растительной основе достигал стадии толстой кишки. Для этого этапа эксперимент был адаптирован для моделирования активности кишечной микробиоты в соответствии с протоколом, описанным Verruck et al. (2020). Моделирование толстой кишки проводили путем добавления SLF (Таблица 1) к болюсу тонкой кишки в соотношении 1:1 (об/об). Затем pH доводили до 7,0 раствором NaOH (1 моль л⁻¹) на протяжении всего анализа в условиях толстой кишки. Во время этого этапа температуру поддерживали на уровне 37°C ± 1°C, а начальную скорость перемешивания поддерживали на уровне 45 об/мин для моделирования интенсивности перистальтических движений. Образцы отбирали через 6, 12, 18, 24, 36, 42 и 48 ч.
До и во время моделирования растворы ферментов хранили в охлажденном состоянии и добавляли постепенно на каждом этапе пищеварения. После каждого этапа желудочно-кишечного тракта оставшееся содержание фенольных соединений экстрагировали с использованием 80% раствора ацетона с 0,01% HCl перед проведением анализов ОСФ и АА. Количественное определение L. rhamnosus GG, ОСФ, антиоксидантной активности (методами DPPH, ABTS и FRAP) и химическую характеристику с помощью ЖХ-ИЭР-МС/МС проводили до моделирования желудочно-кишечного пищеварения, во время фазы полости рта, желудка, тонкого кишечника и во время фазы толстой кишки, как показано на Рисунке 1. Каждый этап желудочно-кишечного тракта моделировали в трех повторностях.
2.2.3 Жизнеспособность клеток Lacticaseibacillus rhamnosus GG во время моделирования in vitro желудочно-кишечного пищеварения
Количество жизнеспособных клеток L. rhamnosus GG в конце каждого этапа моделирования желудочно-кишечного пищеварения оценивали с использованием метода биолюминесценции, следуя протоколу, описанному Camelo-Silva et al. (Camelo-Silva et al., 2024). Вкратце, 100 мкл каждого образца, полученного на этапе моделирования желудочно-кишечного пищеварения, смешивали с 50 мкл реагента BacTiter-Glo™ и подвергали измерению люминесценции в планшетном ридере (GloMax®Explorer System модель GM3500, Promega, США). Количество жизнеспособных клеток L. rhamnosus GG на каждом этапе желудочно-кишечного пищеварения получали путем корреляции поглощения образцов с калибровочной стандартной кривой (R² = 0,9925), построенной путем корреляции значений люминесценции и чашечных подсчетов. Чашечные подсчеты были получены с использованием метода глубинного посева, который состоит из выполнения серийных разведений (физиологический раствор, 0,85 г 100 мл⁻¹) чистой культуры L. rhamnosus GG, с последующей инкубацией в агаре MRS (Man, Rogosa и Sharp) при 37°C ± 1°C в течение 48 ч в бактериологическом термостате. Результаты были выражены как логарифм колониеобразующих единиц на грамм (log КОЕ г⁻¹). Анализ проводили в трех повторностях, включая размножение пробиотических бактерий, использованных для построения кривой.
2.2.4 Уровень выживаемости Lacticaseibacillus rhamnosus GG
После моделирования желудочно-кишечных условий уровень выживаемости (%) L. rhamnosus GG оценивали в соответствии с Уравнением 1 (Verruck et al., 2015).
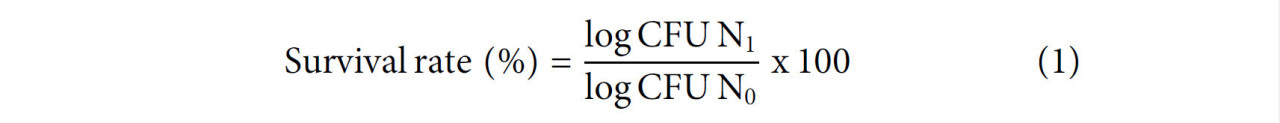
Где N₁ представляет общее количество жизнеспособных клеток L. rhamnosus GG после каждого этапа моделирования желудочно-кишечного пищеварения, а N₀ представляет количество жизнеспособных клеток до воздействия моделирования желудочно-кишечного пищеварения.
2.2.5 Общее содержание фенольных соединений (ОСФ)
ОСФ переваренных и непереваренных ФРН оценивали в соответствии с колориметрическим методом Фолина-Чокальтеу, описанным Singleton и Rossi (Singleton and Rossi, 1965). Сначала 100 мкл каждого образца смешивали с 0,5 мл реактива Фолина-Чокальтеу, 7 мл дистиллированной воды и 1,5 мл 20% (масса/объем) раствора карбоната натрия. Затем растворы выдерживали в темноте в течение 2 ч при комнатной температуре (24°C). После этого измеряли поглощение образцов при 765 нм (УФ-Вид mini-1240, Токио, Япония), используя дистиллированную воду в качестве холостой пробы. Галловую кислоту использовали в качестве стандартного образца, и результаты были выражены в мг эквивалента галловой кислоты (ЭГК) на мл (мг ЭГК мл⁻¹) по результатам трех измерений.
2.2.6 Высвобожденная фракция фенольных соединений
Высвобожденную фракцию (%) содержания фенольных соединений на каждом этапе моделирования желудочно-кишечного пищеварения (рот, желудок, тонкий кишечник и толстая кишка) определяли с помощью Уравнения 2, как описано Arend et al. (Arend et al., 2022).
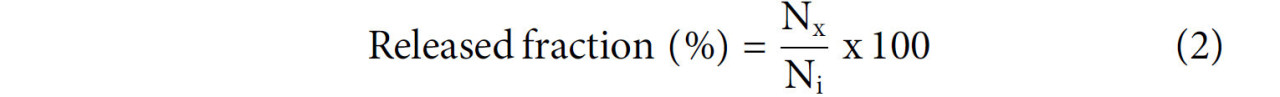
Где: Nₓ представляет ОСФ, количественно определенное после каждого этапа моделируемых желудочно-кишечных условий (мг г⁻¹), а Nᵢ представляет исходное ОСФ, количественно определенное до моделирования.
2.2.7 Антиоксидантная активность (АА) по методам DPPH, ABTS и FRAP
Анализы по захвату радикалов DPPH и ABTS проводили, как описано Rufino et al. (Rufino et al., 2007). Анализ DPPH проводили путем реакции 0,1 мл каждого разбавленного образца с 3,9 мл радикала DPPH (2,2-дифенил-1-пикрилгидразил, 0,06 мМ раствор DPPH). Образцы гомогенизировали и выдерживали в темноте в течение 30 мин. Затем измеряли поглощение образцов на спектрофотометре (УФ-Вид mini-1240, Токио, Япония) при 515 нм. Метиловый спирт использовали в качестве холостой пробы. Для ABTS (Rufino et al.) 30 мкл каждого разбавленного образца смешивали с 3 мл радикала ABTS (диаммониевая соль 2,2-азино-бис(3-этилбензотиазолин-6-сульфоновой кислоты)). Образцы гомогенизировали и выдерживали в темноте при комнатной температуре (24°C) в течение 6 мин. После этого измеряли поглощение (УФ-Вид mini-1240, Токио, Япония) при 734 нм. Этиловый спирт использовали в качестве холостой пробы. Наконец, в методе FRAP (Ferric ion Reducing Antioxidant Power — восстановительная антиоксидантная способность железа) (Rufino et al.) аликвоту 90 мкл каждого разбавленного образца гомогенизировали с 2,7 мл реактива FRAP (25 мл 0,3 М ацетатного буфера, 2,5 мл раствора TPTZ (2,4,6-Трис(2-пиридил)-s-триазина) 10 мМ и 2,5 мл 20 мМ водного раствора хлорида железа). Раствор выдерживали в темноте в течение 30 мин, а затем измеряли поглощение (УФ-Вид mini-1240, Токио, Япония) при 595 нм. Реактив FRAP использовали в качестве холостой пробы.
Для всех методов антиоксидантной активности Trolox использовали в качестве стандартного образца (DPPH — R² = 0,9861, ABTS — R² = 0,9944 и FRAP — R² = 0,9962), и все результаты были выражены в мкМ эквивалента Trolox (ЕТ) мл⁻¹ образца по результатам трех измерений.
2.2.8 Идентификация и количественное определение фенольных соединений с помощью ЖХ-ИЭР-МС/МС
Фенольные соединения анализировали с помощью жидкостной хроматографии с электроспрейной ионизацией и тандемной масс-спектрометрией (ЖХ-ИЭР-МС/МС) для идентификации и количественного определения отдельных соединений. Система ЖХ-ИЭР-МС/МС включала жидкостный хроматограф Agilent 1290 series, сопряженный с гибридным квадрупольным линейным ионным ловушечным масс-спектрометром QTRAP® 5500, оснащенным источником электроспрейной ионизации (ESI). Идентификацию и количественное определение фенольных соединений осуществляли путем сравнения площади пика образца с коммерческими аналитическими стандартами с использованием времени удерживания, иона-предшественника, основного фрагмента (ион для количественного определения) и вторичного фрагмента (ион для подтверждения). Режим множественного мониторинга реакций (MRM), калибровочные кривые, а также пределы обнаружения и количественного определения для 33 протестированных стандартов фенольных соединений можно найти в дополнительной Таблице S1 (Дополнительный материал).
Колонка, использованная для хроматографического разделения, представляла собой Zorbax Eclipse Plus C18 (3,0 мм × 100 мм, 3,5 мкм) от Agilent. Подвижная фаза состояла из воды (A) и метанола (B), оба с 0,1% муравьиной кислоты. Использовали градиентный режим, при котором использовали следующие пропорции B: 2% для 0–3 мин, 20% для 3–10 мин, 90% для 10–11 мин и 2% для 11–13 мин. Скорость потока была установлена на 0,3 мл мин⁻¹, и время уравновешивания 2 мин использовалось между анализами. Объем инъекции составлял 5 мкл. Сбор и анализ данных выполняли с использованием программного обеспечения Analyst и MultiQuant соответственно от Sciex.
2.2.9 Статистический анализ
Результаты этой работы были выражены как среднее значение ± стандартное отклонение по результатам трех измерений. Однофакторный дисперсионный анализ (ANOVA) и критерий Тьюки использовали для определения значительных различий (p < 0,05) между результатами. Весь анализ данных был выполнен с использованием программного обеспечения STATISTICA версии 13.3 (TIBCO Software Inc., Пало-Альто, Калифорния).
3 Результаты и обсуждение
3.1 Жизнеспособность Lacticaseibacillus rhamnosus GG во время моделирования желудочно-кишечного пищеварения
Выживаемость пробиотических микроорганизмов во время их воздействия в суровых желудочно-кишечных средах играет ключевую роль в выполнении их функциональных свойств (Castro-López et al., 2023). Учитывая чувствительность пробиотиков к желудочно-кишечному тракту человека, мониторинг субиетального повреждения пробиотических клеток по всему желудочно-кишечному тракту является ключевым для понимания биологической активности новых функциональных продуктов питания, таких как ФРН.
Количество жизнеспособных клеток и уровень выживаемости клеток L. rhamnosus GG из составов ФРН (ФРН-К, ФРН-5 и ФРН-10) после воздействия моделирования полости рта, желудка и кишечника (подвздошная и толстая кишка) показаны в дополнительной Таблице S2 (Дополнительный материал) и на Рисунке 2 соответственно.
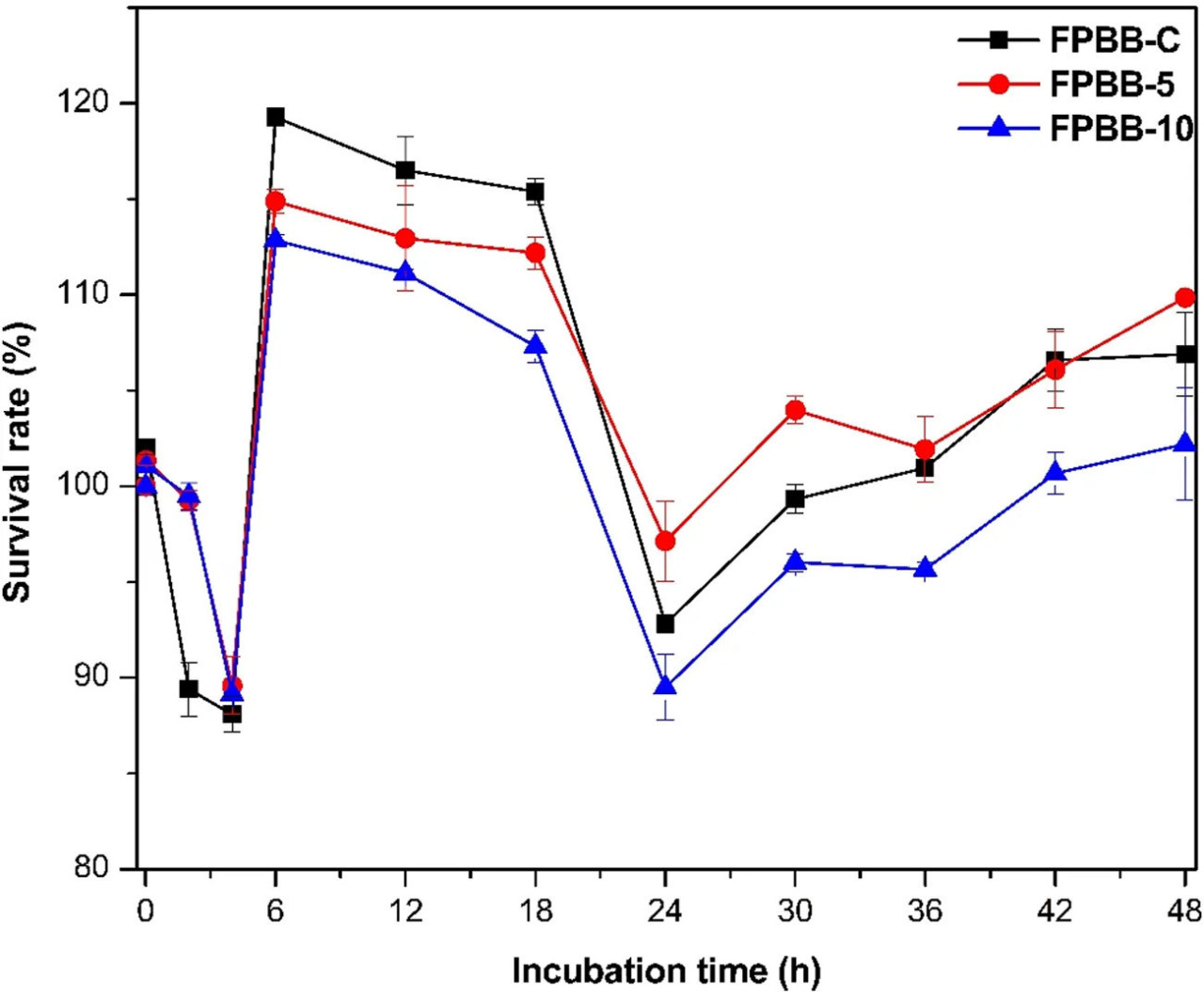
РИСУНОК 2 Уровень выживаемости L. rhamnosus GG после воздействия моделируемых желудочно-кишечных жидкостей. (ФРН-К: ферментированный напиток без мякоти уваи; ФРН-5: ферментированный напиток, обогащенный 5% мякоти уваи; ФРН-10: ферментированный напиток, обогащенный 10% мякоти уваи). Время инкубации относится к этапам желудочно-кишечной системы. Линии используются для лучшей визуализации.
Согласно Prestes et al. (2021), пробиотические продукты должны содержать по крайней мере 6 log КОЕ г⁻¹ жизнеспособных клеток, чтобы предоставлять пользу для здоровья. Соответственно, все образцы ФРН (дополнительная Таблица S2) показали исходное количество жизнеспособных клеток выше 9,30 log КОЕ г⁻¹, превышая рекомендуемый порог для потенциальной пользы для здоровья и, следовательно, классифицируя три ферментированных напитка как пробиотические продукты.
После воздействия условий полости рта не наблюдалось статистических различий (p < 0,05) в количестве жизнеспособных клеток L. rhamnosus GG для всех напитков (ФРН-К: 9,49, ФРН-5: 9,57 и ФРН-10: 9,55 КОЕ г⁻¹) по сравнению с исходными (непереваренными) образцами. Во время оральной фазы жидкая и низковязкая природа растительного напитка приводит к кратковременному контакту с моделируемой слюнной жидкостью (SSF), что не оказывает неблагоприятного воздействия на целостность клеток L. rhamnosus GG.
При моделировании желудка контрольный напиток (ФРН-К) показал статистическое снижение (8,19 log КОЕ г⁻¹) количества клеток L. rhamnosus GG, в то время как количество в составах, обогащенных мякотью уваи (желудок: ФРН-5: 9,37 и ФРН-10: 9,40 log КОЕ г⁻¹), осталось равным (p < 0,05). На этом этапе моделирования желудочно-кишечного пищеварения напитки показали уровни выживаемости от 89,35% до 99,47% (для ФРН-К и ФРН-10 соответственно, Рисунок 2). Важно подчеркнуть, что пищевые матрицы могут защищать пробиотические микроорганизмы во время прохождения через желудочно-кишечный тракт (Tang et al., 2023). Защитные механизмы, предоставляемые мякотью уваи в составах ФРН-5 и ФРН-10 для L. rhamnosus GG во время желудочной фазы, различны и синергичны. Во-первых, высокие уровни выживаемости составов ФРН во время желудочного прохождения (SGF) могут быть частично объяснены образованием нерастворимых комплексов между белками (из гороха и риса) и полисахаридами (такими как пектин из мякоти уваи) (Spricigo et al., 2023). Эти комплексы образуют гелеобразную матрицу вокруг пробиотических клеток, создавая защитный барьер, который защищает L. rhamnosus GG от пепсина и желудочных жидкостей. Этот барьер помогает поддерживать высокую жизнеспособность пробиотической культуры. Полисахариды также способствуют контролируемому высвобождению пробиотиков в кишечник, задерживая воздействие суровых желудочных условий (Mohamadzadeh et al., 2024). Фенольные соединения, включая флавоноиды и фенольные кислоты (Таблица 2; Раздел 3.4), и не фенольные соединения, такие как витамины С и А (идентифицированные в мякоти уваи (Silva et al., 2022)), играют ключевую роль как мощные антиоксиданты. Они нейтрализуют свободные радикалы и снижают окислительный стресс, вызванный кислой средой желудка и пищеварительными процессами (Mohamadzadeh et al., 2024; Wu C. et al., 2023; Ma et al., 2022). Эти соединения также помогают прерывать перекисное окисление липидов, дополнительно защищая пробиотические клетки. Комбинация этих биоактивных компонентов в мякоти уваи работает синергически для усиления защиты L. rhamnosus GG, обеспечивая более высокую выживаемость и эффективность пробиотиков.
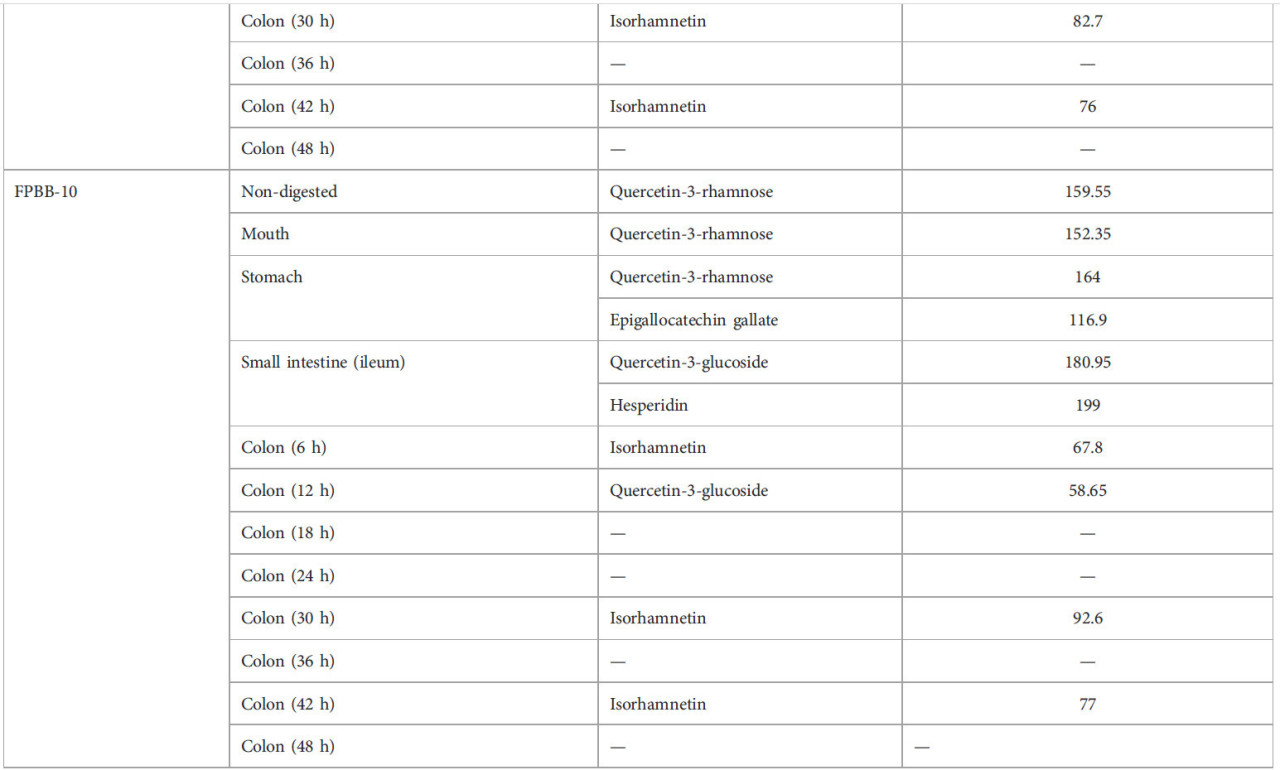
—: none compounds detecte
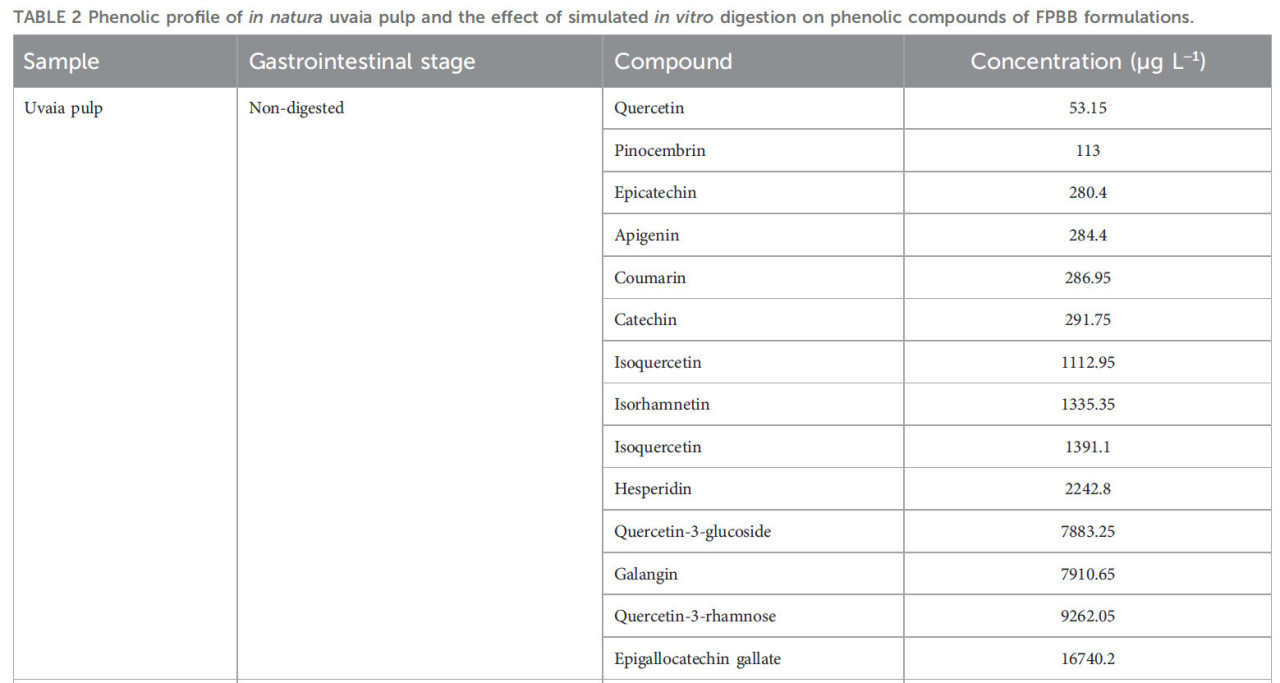
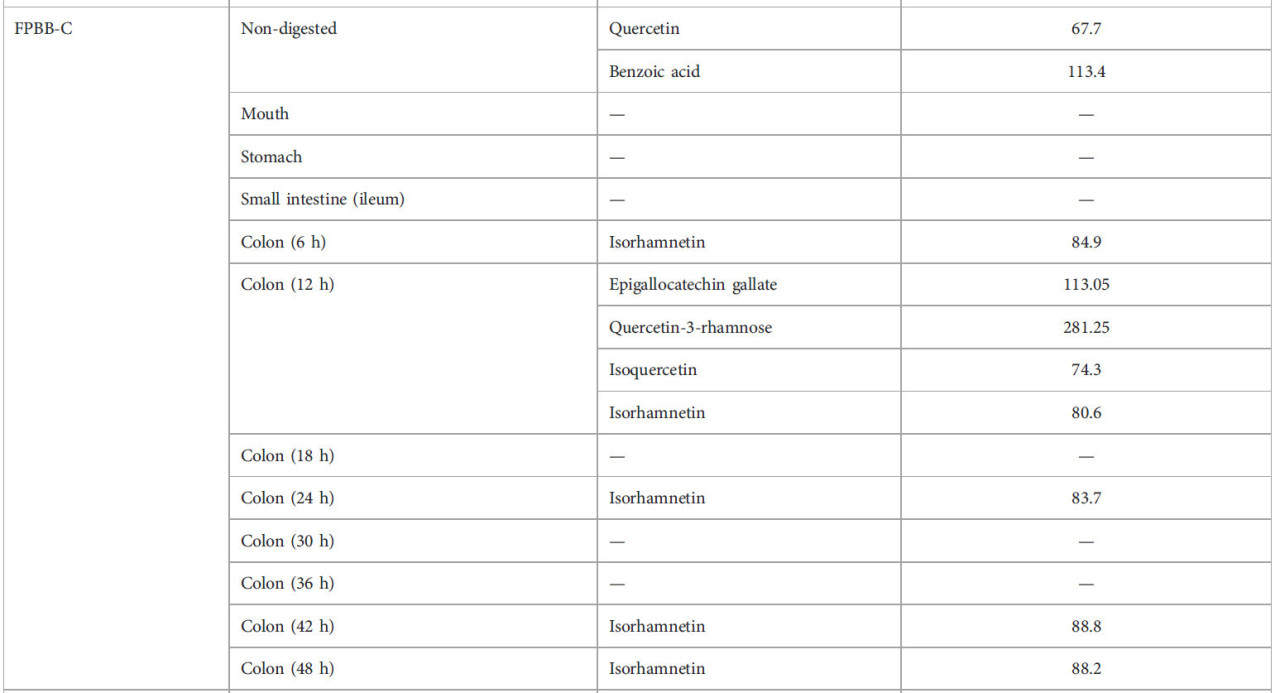
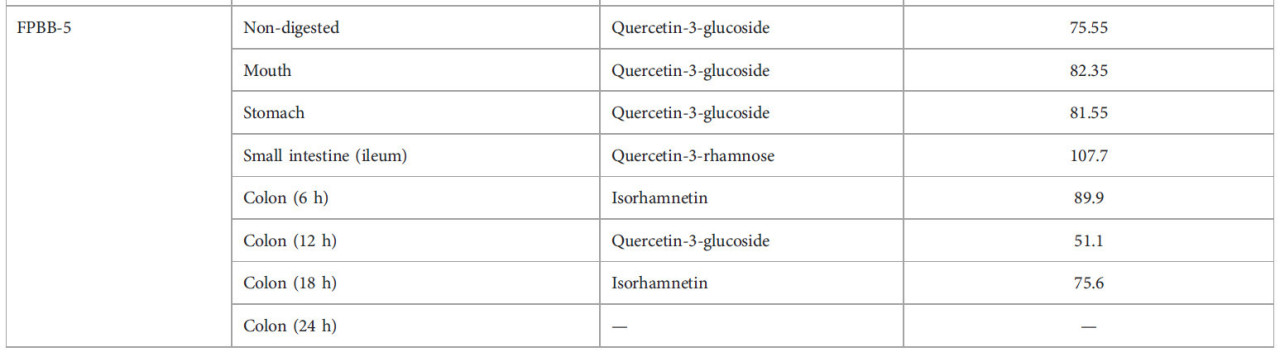
ТАБЛИЦА 2 Фенольный профиль нативной мякоти уваи и влияние моделируемого in vitro пищеварения на фенольные соединения составов ФРН.
—: соединения не обнаружены.
Кроме того, эти комплексы могут контролировать высвобождение пробиотиков в желудочно-кишечном тракте, обеспечивая их выживание и доставку к целевому месту. Более того, эти комплексы могут повышать окислительную стабильность, контролировать высвобождение пробиотиков в желудочно-кишечном тракте и вносить вклад в сенсорные свойства напитка. Ключевые взаимодействия между белками и полисахаридами, такие как водородные связи, гидрофобные взаимодействия и ионные связи, играют ключевую роль в формировании этих комплексов и их защитных свойств (Mohamadzadeh et al., 2024). С точки зрения стабильности во время хранения, эти комплексы могут положительно влиять на срок годности напитков, предотвращая деградацию как пробиотиков, так и биоактивных компонентов (Gentile, 2020). Гелеобразная структура помогает поддерживать целостность составов, снижая вероятность разделения или седиментации (Kupikowska-Stobba et al., 2024). С точки зрения сенсорных свойств, образование комплекса может способствовать желаемой текстуре напитка, повышая его сенсорную привлекательность. Кроме того, комплекс может помочь замаскировать нежелательные вкусы, улучшая общий вкус ФРН (Gentile, 2020; Kupikowska-Stobba et al., 2024).
При моделировании тонкого кишечника было отмечено более выраженное снижение (p < 0,05) (ФРН-К: 8,19, ФРН-5: 8,49 и ФРН-10: 8,42 КОЕ г⁻¹) по сравнению с предыдущими этапами. Уровни выживаемости, наблюдаемые на этом этапе, составили 88,06, 89,94 и 89,10% для ФРН-К, ФРН-5 и ФРН-10 соответственно. Такое поведение может быть связано с влиянием солей желчных кислот в моделируемой кишечной жидкости (SIF). Соли желчных кислот, известные своими детергентообразными характеристиками, как сообщается, взаимодействуют с микробными мембранами, вызывают повреждение ДНК и вызывают гибель бактерий (Chai et al., 2023), снижая скорость восстановления на этом этапе желудочно-кишечного тракта.
Наконец, при моделировании толстой кишки благоприятные условия, характеризующиеся низкой кислотностью и наличием ранее неабсорбированных субстратов (Verruck et al., 2020), способствовали пролиферации L. rhamnosus GG (до 18 ч), что можно подтвердить высоким количеством L. rhamnosus GG (ФРН-К: 10,73, ФРН-5: 10,59 и ФРН-10: 10,50). Однако после 18 ч инкубации ФРН-10 показал значительно более низкую скорость восстановления (p < 0,05), чем ФРН-К и ФРН-5, что может быть связано с микробной адаптацией к менее легкодоступным субстратам, таким как белки с высокой молекулярной массой и олигосахариды (McCoubrey et al., 2023). Кроме того, широко признано, что микроорганизмы, образующие микробиоту кишечника, и фенольные соединения оказывают взаимное влияние. Пищевые полифенолы могут модулировать состав кишечной микробиоты (как пребиотик); в свою очередь, кишечная микробиота катаболизирует полифенолы с высвобождением более активных и лучше всасываемых биоактивных метаболитов (Wang et al., 2022). Тем не менее, некоторые исследования предполагают, что избыточные количества фенольных соединений, достигающих толстой кишки, могут ингибировать рост полезных кишечных микроорганизмов. Таким образом, высокая концентрация мякоти уваи в ФРН-10, вероятно, способствовала высвобождению значительного количества фенольных соединений на этом этапе, ингибируя рост L. rhamnosus GG после 18 ч моделирования желудочно-кишечного пищеварения. Liu и Firrman (2016) сообщили о сходных результатах, показав, что кверцетин (фенольное соединение, также количественно определенное в этом исследовании в образцах толстой кишки, кверцетин-3-рамноза, Раздел 3.4) и нарингенин показали ингибирование роста L. rhamnosus GG в зависимости от дозы через 8 ч после инокуляции. Те же фитохимические вещества также показали дозозависимые ингибирующие эффекты на рост других пробиотических клеток, включая Lactobacillus sp. и Bifidobacterium catenulatum (Duda-Chodak, 2012).
В заключение, после in vitro перевариваемости количество бактерий стабильно оставалось высоким (>9 log КОЕ г⁻¹), что свидетельствует о перспективной жизнеспособности матрицы ФРН в качестве потенциального альтернативного носителя для L. rhamnosus GG. Однако важно отметить, что в этом исследовании состав ФРН-10 нельзя считать оптимальным вариантом для носительства L. rhamnosus GG из-за сниженного уровня выживаемости в толстой кишке (colon), по сравнению с составами ФРН-К и ФРН-5.
3.2 Влияние моделируемого in vitro желудочно-кишечного пищеварения на общее содержание фенольных соединений
Хорошо известно, что желудочно-кишечное пищеварение может изменять фенольный состав пищевых продуктов. В этой связи влияние моделируемого in vitro желудочно-кишечного пищеварения на фенольные соединения определяли путем измерения значений ОСФ до и после каждого этапа желудочно-кишечного тракта. Результаты ОСФ и высвобожденные фракции фенолов (%) показаны в дополнительной Таблице S3 (Дополнительный материал) и на Рисунке 3 соответственно.
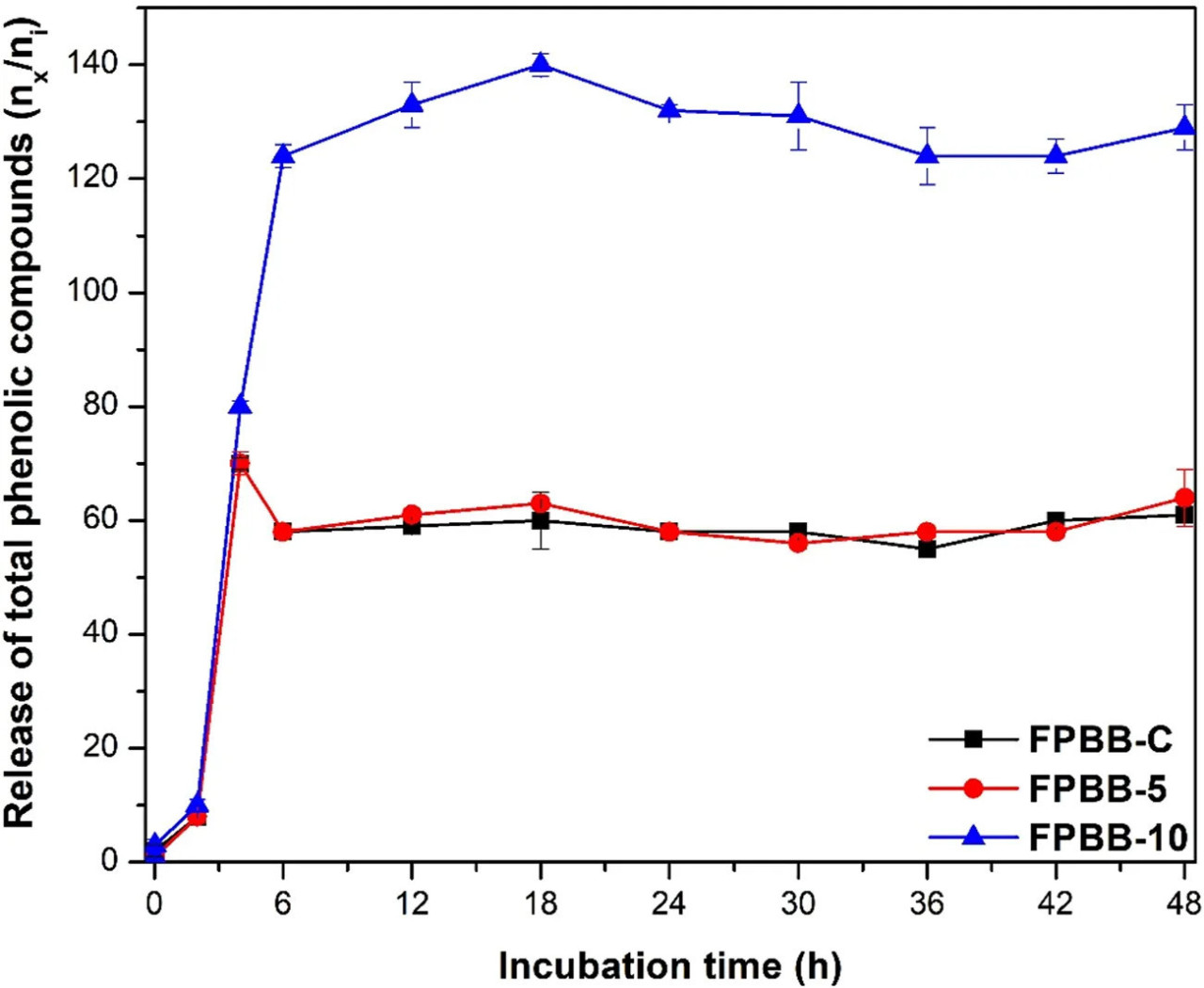
РИСУНОК 3 Высвобождение общих фенольных соединений после воздействия моделируемых желудочно-кишечных условий. (ФРН-К: ферментированный напиток без мякоти уваи; ФРН-5: ферментированный напиток, обогащенный 5% мякоти уваи; ФРН-10: ферментированный напиток, обогащенный 10% мякоти уваи). Время инкубации относится к этапам желудочно-кишечной системы. Линии используются для лучшей визуализации.
Все образцы показали значительное увеличение при p < 0,05 (ФРН-К: 61%, ФРН-5: 64% и ФРН-10: 129%) после 48 ч пищеварения по сравнению с непереваренными образцами. Как показано на Рисунке 3, составы ФРН-К и ФРН-5 показали схожую модель поведения, несмотря на статистическую разницу в конечных значениях ОСФ. Во-первых, в начале желудочно-кишечного анализа (после полости рта) не было обнаружено существенных изменений в результатах ОСФ. Заметное увеличение значений ОСФ наблюдалось после стадии подвздошной кишки, 0,70 мг GAE мл⁻¹ (дополнительная Таблица S3), что отразилось в увеличении на 70% (для обоих составов) по сравнению с количеством, определенным в непереваренных образцах. Такое поведение наблюдалось для ферментированных напитков с соком алоэ вера (Cuvas-Limón et al., 2022) и драконовым фруктом-киви (Tang et al., 2023) и может быть объяснено наличием кислой среды в желудочной и кишечной стадиях, которая может влиять на структурную матрицу ФРН через гидролиз комплексов полифенол-белок (связанных водородными или амидными связями) или полифенол-углевод (связанных гидрофобными связями) (Tang et al., 2023). Аналогично, увеличение значений ОСФ также может быть объяснено действием пищеварительных ферментов (α-амилазы, пепсина и панкреатина) на связь между фенольными соединениями и клеточной стенкой мякоти уваи, в частности через сложноэфирные связи, тем самым высвобождая фенольные кислоты (Wu C. et al., 2023). Высвобождение интактных или гидролизованных фенольных соединений делает их более склонными к реакции с реактивом Фолина-Чокальтеу, увеличивая значения ОСФ.
После in vitro пищеварения наблюдалось небольшое снижение до состояния толстой кишки через 36 ч (55% и 58% для ФРН-К и ФРН-5 соответственно) и снова увеличение до достижения максимального ОСФ в толстой кишке через 48 ч. Для состава ФРН-10 наиболее высокое значение ОСФ было обнаружено в толстой кишке через 18 ч (1,40 мг GAE мл⁻¹) с небольшим снижением до состояния толстой кишки через 48 ч (1,29 мг GAE мл⁻¹), сохраняя общее содержание фенольных соединений значительно выше (на всех этапах желудочно-кишечного тракта) по сравнению с ФРН-К и ФРН-5. Поскольку толстая кишка показывает заметное увеличение размножения L. rhamnosus GG, на что указывает количество жизнеспособных клеток после 24 ч (дополнительная Таблица S3), кишечная микрофлора, вероятно, облегчает структурную трансформацию и метаболизм фенольных соединений (Zhang et al., 2023; Cattivelli et al., 2023), объясняя прогрессивное увеличение на протяжении in vitro перевариваемости, Рисунок 3, что, фактически, является положительным эффектом, поскольку фенольные метаболиты, генерируемые действием кишечной микробиоты, обычно считаются лучше абсорбируемыми и ответственными за полезные эффекты ФРН, как сообщалось ранее (Раздел 3.1).
В отличие от результатов этой работы, Farias et al. (2021) обнаружили значительное снижение (41%) значений ОСФ и увеличение общего содержания флавоноидов (109%) после кишечной фазы мякоти уваи. Те же авторы предположили, что фенольные кислоты (идентифицированные и количественно определенные в мякоти уваи с помощью ВЭЖХ) более восприимчивы к пищеварительной деградации, вероятно, из-за меньшей структурной сложности по сравнению с флавоноидами (в основном встречающимися в гликозилированной форме — связанной с молекулами углеводов). Важно подчеркнуть, что ФРН представляет собой сложную пищевую матрицу. Взаимодействие между химическими соединениями, присутствующими в компонентах напитка (рисовый и гороховый белки, сахароза, соевое масло и соевый лецитин), и фенольными соединениями из мякоти уваи может защищать фенольные соединения (подобно агентам инкапсуляции), способствуя более медленному высвобождению и, следовательно, способствуя высоким результатам ОСФ после пищеварения по сравнению с исследованиями Farias et al. (2021), которые оценивали нативный фрукт.
Таким образом, наши результаты подчеркивают, что ФРН является отличной пищевой матрицей для доставки полифенолов из мякоти уваи в концентрации 5%, поскольку фенольные соединения увеличивались во время in vitro желудочно-кишечного пищеварения без ущерба для жизнеспособности клеток L. rhamnosus GG, что определяет ФРН как пробиотический и пребиотический продукт.
3.3 Изменения антиоксидантной активности (АА) во время моделирования желудочно-кишечного пищеварения
Природные антиоксиданты играют ключевую роль в здоровье человека. Эти соединения могут ингибировать или задерживать нежелательные реакции окисления и, таким образом, предотвращать окислительный стресс, связанный с широким спектром заболеваний организма человека, таких как высокое кровяное давление, нейродегенеративные расстройства или рак (Rumpf et al., 2023). Оценка их активности во время in vitro желудочно-кишечного пищеварения является важным подходом для скрининга их биодоступности и эффективности в предоставлении антиоксидантных преимуществ организму. Эта оценка также является ключевой при доработке новых пищевых составов для максимизации их эффективности после проглатывания. Изучение более чем одного метода АА повышает квалификацию и количественное определение антиоксидантной активности продукта. Этот подход ценен, поскольку антиоксидантная способность зависит от полярности многочисленных соединений в пищевой матрице и их различных взаимодействий и синергических эффектов с конкретным антиоксидантным радикалом (Rumpf et al., 2023).
Влияние моделируемого желудочно-кишечного пищеварения на антиоксидантную способность составов ФРН, оцененное с помощью методов DPPH, ABTS и FRAP, представлено на Рисунке 4.
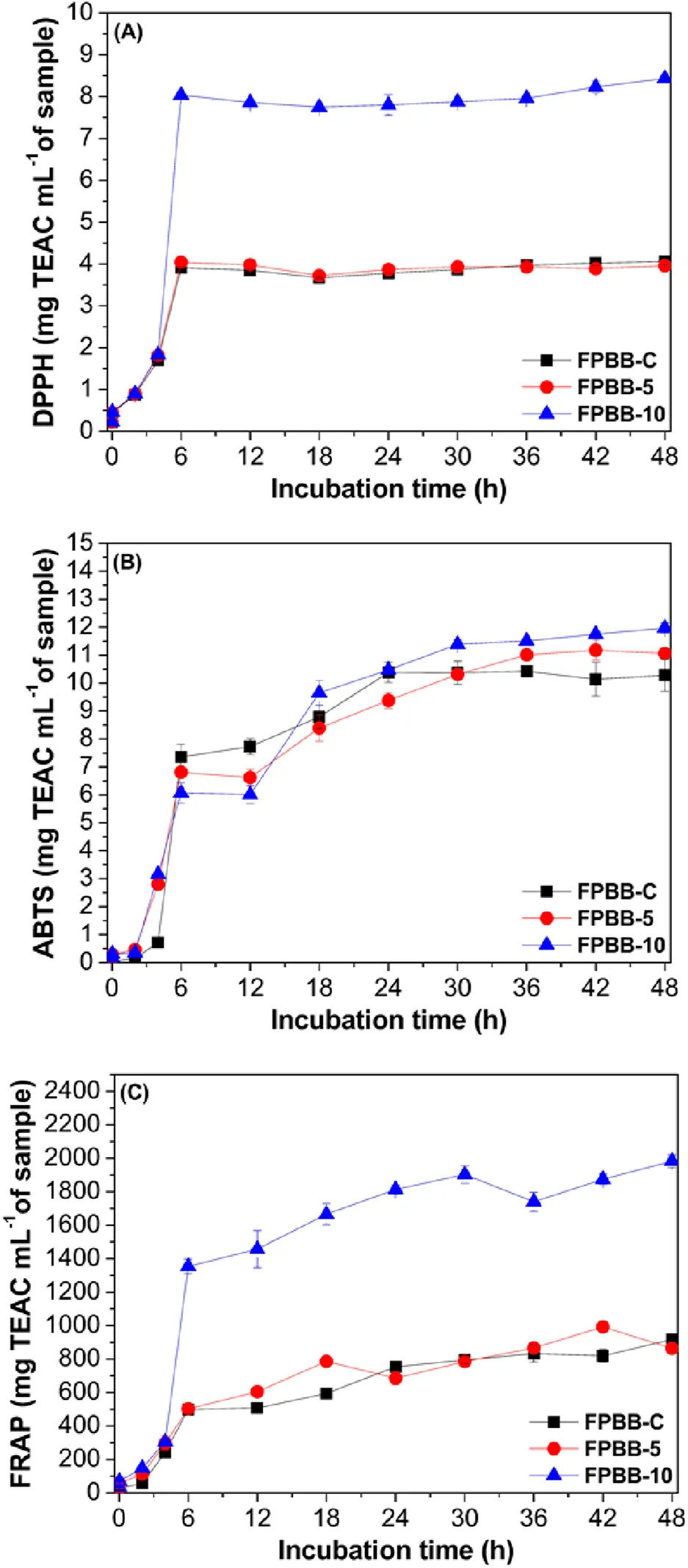
РИСУНОК 4 Антиоксидантная активность (мг TEAC мл⁻¹ образца) по методам DPPH (A), ABTS (B) и FRAP (C) составов ФРН после моделирования желудочно-кишечного пищеварения. Время инкубации относится к этапам желудочно-кишечной системы. Линии используются для лучшей визуализации. ФРН-К: ферментированный напиток без мякоти уваи; ФРН-5: ферментированный напиток, обогащенный 5% мякоти уваи; ФРН-10: ферментированный напиток, обогащенный 10% мякоти уваи. DPPH (2,2-дифенил-1-пикрилгидразил); ABTS (диаммониевая соль 2,2-азино-бис(3-этилбензотиазолин-6-сульфоновой кислоты)); FRAP (восстановительная антиоксидантная способность железа).
Согласно методу DPPH (Рисунок 4A), все образцы показали значительное увеличение АА (p < 0,05) после in vitro перевариваемости. Кроме того, наблюдаемое поведение антиоксидантной активности (АА) оставалось стабильным для всех составов. Первоначально наблюдалось постепенное увеличение АА до фазы подвздошной кишки, за которым следовало значительное увеличение через 6 ч в толстой кишке. Впоследствии наблюдалось снижение после 30 ч (ФРН-К), 42 ч (ФРН-5) и 36 ч (ФРН-10). После этого АА снова увеличилась, достигнув своих максимальных значений в толстой кишке через 48 ч, достигнув максимальных значений (ФРН-К: 4,06, ФРН-5: 3,96 и ФРН-10: 8,44 мг TEAC мл⁻¹ образца). Тем не менее, для метода DPPH не наблюдалось значительных различий (p < 0,05) между ФРН-К и ФРН-5. Однако состав ФРН-10 показал более высокую АА (p < 0,05), чем ФРН-К и ФРН-5, что подтверждает результаты ОСФ (Раздел 3.2).
Такое же поведение наблюдалось для метода ABTS (Рисунок 4B). Все образцы показали увеличение АА после in vitro перевариваемости (p < 0,05), причем максимальное значение было получено в толстой кишке через 48 ч (ФРН-К: 10,28, ФРН-5: 11,06 и ФРН-10: 11,97 мг TEAC мл⁻¹ образца). При анализе максимальной АА между составами (ФРН-К, ФРН-5 и ФРН-10), ФРН-5 и ФРН-10 показали более высокие значения, чем ФРН-К. Следовательно, результаты показали, что мякоть уваи обладает антиоксидантным потенциалом при добавлении в количествах, равных или превышающих 10%, в ферментированный растительный напиток по активности захвата радикала ABTS.
Для метода FRAP (Рисунок 4C) все образцы также показали постепенное увеличение с начала in vitro перевариваемости до достижения своих максимальных значений антиоксидантной активности в толстой кишке через 48 ч, с АА в 58, 31 и 54 раза выше, чем в непереваренных образцах ФРН-К, ФРН-5 и ФРН-10 соответственно.
Увеличение АА (для всех методов), наблюдаемое, когда образцы достигают нисходящей толстой кишки, может быть связано с взаимодействиями фенольных соединений с кишечной микробиотой. Как сообщалось ранее в Разделах 3.1, 3.2, эти взаимодействия, возможно, включают структурные трансформации и высвобождение более реакционноспособных фенольных соединений с более высокой АА. Флавонолы, высвобождаемые из ФРН (Таблица 2), в основном (изокверцетин и кверцетин-3-глюкозидаза), присутствующие в более высоких концентрациях на стадии толстой кишки, вероятно, вносят вклад в его антиоксидантную активность. Исследование Rodrigues et al. (2021) также идентифицировало кверцетин в экстрактах остатков уваи (кожура и семена), приписывая высокую антиоксидантную активность (ABTS: 14,22, DPPH: 9,60 и FRAP: 20,79 ммоль TE г⁻¹ экстракта) присутствию кверцетина и других фенольных соединений.
Помимо присутствия фенольных соединений в мякоти уваи, гидролиз белков гороха и риса под действием пепсина в желудочной фазе может привести к образованию и высвобождению более коротких биоактивных пептидов с улучшенными антиоксидантными свойствами во время пищеварения в подвздошной кишке и толстой кишке ферментированного напитка (Manus et al., 2021), способствуя увеличению АА образцов. Исследование Olagunju et al. (2018) показало, что гидролизованный панкреатином белок гороха с низкой молекулярной массой (1–5 кДа) показал лучшую антиоксидантную активность по методу DPPH (EC50 1,02–1,26 мг мл⁻¹), чем фракции с более высокой молекулярной массой (значение EC50 2,44 мг мл⁻¹). Другое исследование Chen et al. (2023) также показало, что рисовые пептиды с низкой молекулярной массой (<1 кДа, IC50 26,7 мг мл⁻¹) обладают более высокой антиоксидантной способностью, чем пептиды с высокой молекулярной массой (>10 кДа, IC50 39,4 мг мл⁻¹) по методу DPPH.
Другие биоактивные соединения также могут способствовать АА ФРН. Плоды уваи обильны витамином С, ключевой антиоксидантной молекулой, с концентрациями в диапазоне от 9,45 до 122,51 мг на 100 г (Silva et al., 2022), и каротиноидами, с концентрациями в диапазоне от 1,3 до 441,26 мг на 100 г (Sganzerla et al., 2021), взаимодействие этих соединений с радикалами также может увеличивать АА ФРН.
Таким образом, состав ФРН-10 показал повышенную антиоксидантную активность для переваренных и непереваренных образцов по сравнению с ФРН-К и ФРН-5. Повышенная АА в составе с добавлением 10% мякоти уваи может быть связана с различными факторами. Как сообщалось в Разделе 3.2, ФРН-10 содержит значительно более высокое количество фенольных соединений, чем ФРН-К и ФРН-5. Эта более высокая концентрация напрямую повышает антиоксидантную активность. Кроме того, синергические эффекты между различными фенольными соединениями дополнительно усиливают эту активность. Нефенольные соединения, такие как витамины С и А (ранее идентифицированные в мякоти уваи (Sganzerla et al., 2021; Spricigo et al., 2023; Silva et al., 2022)), также присутствующие в мякоти уваи, способствуют общему антиоксидантному эффекту. Более того, полисахариды и пептиды в составе поддерживают антиоксидантную активность за счет удаления свободных радикалов, хелатирования ионов металлов и стабилизации других антиоксидантных соединений (Zhang et al., 2024; Fernandes and Coimbra, 2023). Следовательно, превосходная антиоксидантная активность ФРН-10 является результатом комбинированных эффектов, таких как более высокая концентрация фенольных соединений, синергические взаимодействия и вклад нефенольных соединений, полисахаридов и пептидов.
Учитывая наблюдаемое снижение выживаемости пробиотика в ФРН-10 во время фазы толстой кишки, является ключевым определение оптимальной концентрации мякоти уваи, которая уравновешивает жизнеспособность пробиотика с антиоксидантными преимуществами. Концентрация мякоти уваи между 5% и 10% выглядит перспективной. Концентрация 5% (ФРН-5) поддерживает более высокую выживаемость пробиотика, предоставляя при этом существенные антиоксидантные преимущества. Напротив, концентрация 10% (ФРН-10) значительно усиливает антиоксидантные свойства, но ценой сниженной жизнеспособности пробиотика. Таким образом, концентрация, близкая к 5%, может предоставить лучший баланс между поддержанием жизнеспособности пробиотика и предоставлением антиоксидантных преимуществ.
Анализируя изменения АА составов ФРН, наблюдалась значительная разница в увеличении антиоксидантной активности, что свидетельствует о том, что добавление мякоти уваи в ферментированные растительные напитки может быть ценной стратегией для повышения функциональности таких напитков.
3.4 Фенольные профили мякоти уваи и переваренных и непереваренных составов ФРН
Как сообщалось ранее (Раздел 3.2), включение мякоти уваи в состав ФРН привело к увеличению значений ОСФ. Следовательно, был проведен ВЭЖХ-анализ для идентификации фенольных соединений, потенциально вносимых в препарат за счет добавления мякоти уваи, и для оценки любой биотрансформации этих соединений во время in vitro желудочно-кишечного анализа. Химический состав (фенольные соединения) нативной мякоти уваи (непереваренной) и составов ФРН был оценен качественно и количественно с помощью ВЭЖХ-ДМД-ИЭР-МС. Соединения были идентифицированы на основе их времени удерживания, УФ-Вид спектров поглощения и масс-спектров. Результаты ВЭЖХ показаны в Таблице 2.
Четырнадцать соединений были идентифицированы и количественно определены в мякоти уваи (непереваренный образец), достигнув общего содержания фенольных соединений 49188,00 мкг л⁻¹. Большинство соединений были флавоноидами, в частности отличительным было присутствие эпигаллокатехин галлата (16740,2 мкг л⁻¹), кверцетин-3-рамнозы (9262,05 мкг л⁻¹) и кверцетин-3-глюкозида (7883,25 мкг л⁻¹). Кроме того, значительное количество галангина (7910,65 мкг л⁻¹), гесперидина (2242,8 мкг л⁻¹) и изокверцетина (1112,95 мкг л⁻¹) было обнаружено в нативной мякоти уваи. Например, Wanderley et al. (Wanderley et al., 2022) оценивали фенольный профиль мякоти уваи из Уруземы, Санта-Катарина, Бразилия, идентифицируя (+)-катехин (6,54 мг 100 г⁻¹) в качестве основного фенольного соединения. Кроме того, эпикатехин (4,43 мг 100 г⁻¹) и кверцетин (0,38 мг 100 г⁻¹) также были обнаружены авторами, что подтверждает результаты ВЭЖХ данной работы. Другое исследование Rodrigues et al. (2021) также идентифицировало изокверцитрин, кверцетин и эпикатехин в экстрактах остатков уваи из Параибуны, SP, Бразилия. Тем не менее, эти авторы не обнаружили пиноцембрин, апигенин, галангин и кумарин; насколько нам известно, это первое сообщение об обнаружении этих соединений в мякоти уваи. Известно, что эти соединения широко распространены во фруктах и овощах, и их потребление способствует многочисленным преимуществам для здоровья человека, таким как противовоспалительные, антибактериальные, противораковые свойства и свойства против болезни Альцгеймера, среди прочих, благодаря их антиоксидантной активности против свободных радикалов (Elbatreek et al., 2023; Hoff et al., 2022). Однако их концентрация может сильно варьироваться из-за видов растений, генетики, среды выращивания, стадии развития и части растения (Spricigo et al., 2023; Rodrigues et al., 2021). Этот факт также объясняет различные фенольные профили и концентрации продуктов уваи среди этих работ.
Только два фенольных соединения были количественно определены в непереваренном контрольном напитке (ФРН-К): кверцетин (67,7 мкг л⁻¹) и бензойная кислота (113,4 мкг л⁻¹). Тем не менее, с прогрессом пищеварения (в частности в толстой кишке) другие соединения высвобождались из матрицы ФРН-К. Например, изорамнетин (84,9 мкг л⁻¹) был единственным соединением, обнаруженным в толстой кишке (6 ч). После 12 ч ферментации в толстой кишке высвобождались другие соединения, включая эпигаллокатехин галлат (113,05 мкг л⁻¹), кверцетин-3-рамнозу (281,25 мкг л⁻¹), изокверцетин (74,3 мкг л⁻¹), а также изорамнетин (80,6 мкг л⁻¹). После этого этапа желудочно-кишечного тракта (толстая кишка 12 ч) изорамнетин был единственным соединением, обнаруженным на конечной стадии ЖКТ (толстая кишка 48 ч), будучи снова обнаруженным в толстой кишке (24 ч) (83,7 мкг л⁻¹), толстой кишке (42 ч) (88,8 мкг л⁻¹) и толстой кишке (48 ч) (88,2 мкг л⁻¹). Изорамнетин-3-О-глюкозид, флавоноловое соединение, связанное с углеводом, является одним из наиболее распространенных флавоноидов, широко распространенных в сортах риса (Wang et al., 2023). Кроме того, изорамнетин-3-рутинозид был обнаружен в семенах, стручках, ростках и листьях гороха (Wu D. T. et al., 2023). Как сообщалось ранее, пищеварительные ферменты и моделируемые жидкости воздействовали на белки гороха и риса (не гидролизованные клетками L. rhamnosus GG во время ферментации) из ФРН-К, превращая их в пептиды и аминокислоты, которые высвобождали и метаболизировали изорамнетин. Согласно результатам ВЭЖХ-анализа, вероятно, что изорамнетин-3-О-глюкозид способствовал высокой антиоксидантной активности, наблюдаемой методами DPPH, ABTS и FRAP через 42 и 48 ч в толстой кишке.
Что касается образца ФРН-5, Таблица 2 показала значительное высвобождение кверцетин-3-глюкозида во время оральной и желудочной фаз, 82,35 и 81,55 мкг л⁻¹ соответственно, и кверцетин-3-рамнозы в тонком кишечнике (107,7 мкг л⁻¹). В нашем исследовании изорамнетин в основном обнаруживался на более поздних стадиях толстой кишки с заметными концентрациями 89,9 мкг л⁻¹ в толстой кишке через 6 ч, 82,7 мкг л⁻¹ после 30 ч и 76 мкг л⁻¹ через 42 ч (Рисунок 5). Это интересное открытие. Согласно Olthof et al. (2000), примерно 50% глюкозидов кверцетина всасывается в тонком кишечнике. Кроме того, по мнению тех же авторов, неабсорбированный в тонком кишечнике кверцетин-3-глюкозид может метаболизироваться кишечной микробиотой (в этом исследовании представленной L. rhamnosus GG) в агликон кверцетина и фенольные кислоты (например, в изорамнетин), которые могут всасываться из толстой кишки; действие L. rhamnosus GG на кверцетин-3-глюкозид (наблюдаемое в начале пищеварения, в полости рта и желудке) во время пищеварения в толстой кишке может объяснять существование этого соединения в толстой кишке через 30 и 42 ч.
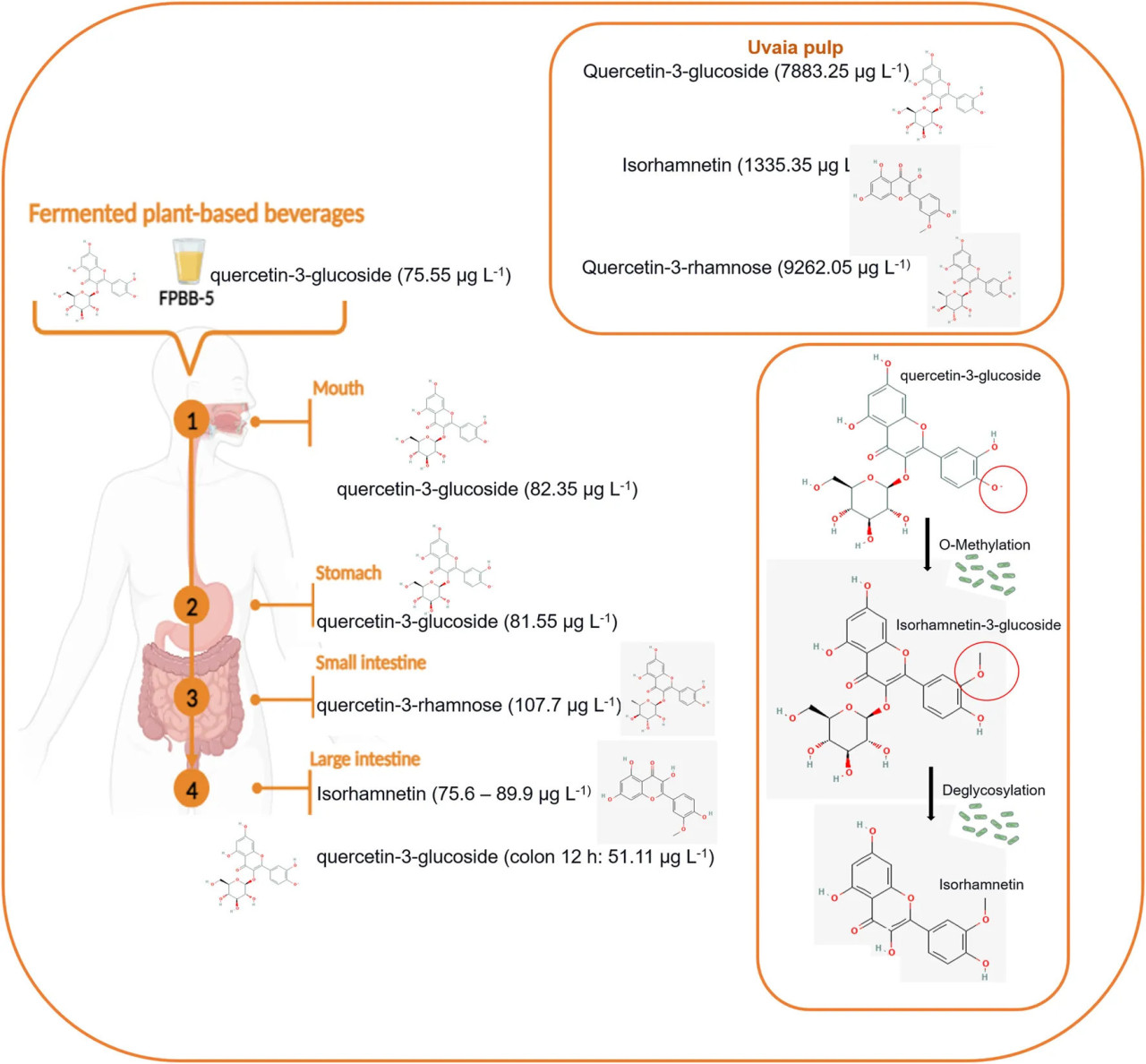
РИСУНОК 5 Трансформация кверцетин-3-глюкозида в изорамнетин во время пищеварения с использованием образца ФРН-5 в качестве модели.
Данные из Таблицы 2 о составе ФРН-10 показывают непрерывное присутствие соединений, производных от кверцетина, на протяжении всего пищеварительного процесса, таких как кверцетин-3-рамноза, в полости рта (152,35 мкг л⁻¹), желудке (164 мкг л⁻¹) и кверцетин-3-глюкозидаза, в тонком кишечнике (180,95 мкг л⁻¹) и в толстой кишке через 12 ч (58,65 мкг л⁻¹). Согласно с составом ФРН-5, изорамнетин также наблюдался в толстой кишке через 30 ч и 42 ч со значениями 92,6 и 77 мкг л⁻¹ соответственно. Недавнее исследование Ahn et al. (2020) показало, что Bifidobacterium animalis subsp. lactis AD011, выделенный из фекалий младенцев, катализировал более 85% кверцетин-3-глюкозида и изорамнетин-3-глюкозида в кверцетин и изорамнетин соответственно за 2 ч. Биотрансформированные молекулы, кверцетин и изорамнетин, показали улучшенную противовоспалительную активность по сравнению с небиотрансформированными молекулами в отношении макрофагов RAW 264.7, индуцированных липополисахаридом.
Точный ферментативный механизм, лежащий в основе превращения кверцетин-3-глюкозида в изорамнетин кишечной микробиотой, в частности L. rhamnosus GG, остается в значительной степени неизвестным. Согласно Williamson и Clifford (2024), специфические ферменты и условия в желудочно-кишечном тракте облегчают эту трансформацию во время пищеварения. Во-первых, кверцетин-3-глюкозид гидролизуется с высвобождением кверцетина. Хотя нет доказательств биотрансформации L. rhamnosus GG, другие исследования, такие как Ahn et al. (2020), показывают, что продукция Bifidobacterium animalis subsp. lactis может катализировать эту биотрансформацию через гидролиз β-1,4-гликозидной связи, чему способствуют такие ферменты, как β-глюкозидаза (Terao, 2023). Впоследствии кверцетин может подвергаться ряду метаболических реакций, включая метилирование, где метильная группа добавляется к молекуле, превращая ее в изорамнетин. Эти реакции могут катализироваться УДФ-глюкуронозилтрансферазами, сульфотрансферазами и катехол-О-метилтрансферазами (Boonpawa et al., 2014). Эта трансформация является ключевой для биодоступности и биологической активности этих соединений, влияя на их действие на организм во время пищеварения и всасывания. Кроме того, эти результаты подтверждают высокие результаты ОСФ (дополнительная Таблица S3) и антиоксидантную способность методов ABTS и FRAP (Рисунок 4), показывая высокую корреляцию между антиоксидантной активностью и этими соединениями.
Наконец, присутствие кверцетин-3-глюкозида в мякоти уваи (7883,25 мкг л⁻¹), ФРН-5 (непереваренный: 75,55 мкг л⁻¹) и ФРН-10 (тонкий кишечник: 180,95 и толстая кишка 12 ч: 58,65 мкг л⁻¹) и гесперидина (тонкий кишечник: 199 мкг л⁻¹) в ФРН-10, но его отсутствие в контрольном составе подчеркивает значимость мякоти уваи как источника этих соединений в функциональном напитке.
Наши результаты свидетельствуют о том, что биоконверсия фенольных соединений часто происходит во время ферментации ингредиентов ФРН L. rhamnosus GG, и различия в фенольных профилях могут быть связаны с активностью специфических ферментов. Более того, состав фенольных соединений является двойным вкладом от используемых ингредиентов — белков гороха и риса — вместе с мякотью уваи.
4 Выводы
Ферментированные напитки на растительной основе были успешно произведены с использованием белков гороха и риса и 5% и 10% мякоти уваи путем молочнокислой ферментации с L. rhamnosus GG. Во время экспериментов по моделированию in vitro желудочно-кишечного пищеварения жизнеспособность клеток L. rhamnosus GG оставалась высокой, с концентрациями жизнеспособных клеток в соответствии с классификацией ФРН как пробиотиков (ФРН-К: 9,94 log КОЕ г⁻¹, ФРН-5: 10,37 log КОЕ г⁻¹ и ФРН-10: 9,66 log КОЕ г⁻¹). Эти результаты показывают, что матрица из рисового и горохового белка эффективно проведена в качестве носителя штамма L. rhamnosus GG. Добавление уваи увеличило общее содержание фенольных соединений в конце желудочно-кишечного пищеварения, свидетельствуя об увеличении пребиотической способности напитков. Все составы показали улучшение антиоксидантной активности по методам DPPH, ABTS и FRAP в конце in vitro перевариваемости, демонстрируя, что добавление уваи актуально и интересно для приготовления растительных напитков, улучшая их питательные качества. Четырнадцать соединений были идентифицированы и количественно определены в мякоти уваи с помощью ВЭЖХ-ДМД-ИЭР-МС, большинство из них классифицированы как флавонолы, причем эпигаллокатехин галлат (16740,2 мкг л⁻¹), кверцетин-3-рамноза (9262,05 мкг л⁻¹) и кверцетин-3-глюкозид (7883,25 мкг л⁻¹) были основными фенольными соединениями, количественно определенными в мякоти. Кроме того, изорамнетин был основным фенольным соединением, обнаруженным в толстой кишке для всех составов, вероятно, из-за трансформации кверцетин-3-глюкозида пробиотическими клетками. Наконец, это исследование демонстрирует, что пищевая промышленность может изучать новые альтернативы напиткам, содержащим матрицу из гороха и риса, обогащенную мякотью уваи, предоставляя возможность потреблять напитки на растительной основе. Предварительные результаты этого исследования обнадеживают; однако будущие исследования должны включать клинические испытания для подтверждения эффективности и пользы для здоровья составов ФРН in vivo, поскольку анализы in vitro могут не полностью воспроизводить условия пищеварения человека. Кроме того, важно подчеркнуть, что все еще существует пробел, который необходимо заполнить в сенсорном анализе продукта. Добавление пробиотиков и мякоти уваи, хотя и многообещающее в функциональном плане, может повлиять на органолептическое восприятие продукта. Поэтому необходимы детальные сенсорные исследования для оценки реакции потребителей на вкус, текстуру и общую приемлемость растительных напитков. Наконец, оценка масштабируемости и экономической целесообразности включения мякоти уваи в крупномасштабное производство является ключевой для практического применения в пищевой промышленности.
Ссылки
1. Ahn H. J., You H. J., Park M. S., Li Z., Choe D., Johnston T. V., et al. (2020). Microbial biocatalysis of quercetin-3-glucoside and isorhamnetin-3-glucoside in Salicornia herbacea and their contribution to improved anti-inflammatory activity. RSC Adv. 10, 5339–5350. doi: 10.1039/c9ra08059g. CrossRef. Google Scholar.
2. Alcorta A., Porta A., Tárrega A., Alvarez M. D., Pilar Vaquero M. (2021). Foods for plant-based diets: challenges and innovations. Foods 10, 293. doi: 10.3390/foods10020293. CrossRef. Google Scholar.
3. Arend G. D., Almeida É. S., Byruchko R. T., Pinto M. E. G., da Cruz A. B., Verruck S., et al. (2022). Gravitational and microwave-assisted multi-stages block freeze concentration process to obtain enriched concentrated beet (Beta vulgaris L.) by-products extract: bioactive compounds and simulated gastrointestinal profile. Food Bioprod. Process. 133, 77–86. doi: 10.1016/j.fbp.2022.03.004. CrossRef. Google Scholar.
4. Bianchini C. B., Vieira M. P. T., Arriola N. D. A., Dias C. O., Seraglio S. K. T., Costa A. C. O., et al. (2020). Incorporation of uvaia (Eugenia pyriformis Cambess) pulp in yogurt: a promising application in the lactose‐free dairy product market. J. Food Process Preserv 44, 1–8. doi: 10.1111/jfpp.14829. CrossRef. Google Scholar.
5. Boonpawa R., Spenkelink A., Rietjens I. M. C. M., Punt A. (2014). A physiologically based kinetic (PBK) model describing plasma concentrations of quercetin and its metabolites in rats. Biochem. Pharmacol. 89, 287–299. doi: 10.1016/j.bcp.2014.02.007. CrossRef. Google Scholar.
6. Brodkorb A., Egger L., Alminger M., Alvito P., Assunção R., Ballance S., et al. (2019). INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 14, 991–1014. doi: 10.1038/s41596-018-0119-1. CrossRef. Google Scholar.
7. Camelo-Silva C., Mota e Souza B., Vicente R., Arend G. D., Sanches M. A. R., Barreto P. L. M., et al. (2024). Polyfunctional sugar-free white chocolate fortified with Lacticaseibacillus rhamnosus GG co-encapsulated with beet residue extract (Beta vulgaris L.). Food Res. Int. 179, 114016. doi: 10.1016/j.foodres.2024.114016. CrossRef. Google Scholar.
8. Castro-López C., Romero-Luna H. E., García H. S., Vallejo-Cordoba B., González-Córdova A. F., Hernández-Mendoza A. (2023). Key stress response mechanisms of probiotics during their journey through the digestive system: a review. Probiotics Antimicrob. Proteins 15, 1250–1270. doi: 10.1007/s12602-022-09981-x. CrossRef. Google Scholar.
9. Cattivelli A., Nissen L., Casciano F., Tagliazucchi D., Gianotti A. (2023). Impact of cooking methods of red-skinned onion on metabolic transformation of phenolic compounds and gut microbiota changes. Food Funct. 14, 3509–3525. doi: 10.1039/d3fo00085k. CrossRef. Google Scholar.
10. Chai L. N., Wu H., Wang X. J., He L. J., Guo C. F. (2023). The mechanism of antimicrobial activity of conjugated bile acids against lactic acid bacilli. Microorganisms 11, 1823. doi: 10.3390/microorganisms11071823. CrossRef. Google Scholar.
(Продолжение для всех 59 источников в том же формате. Ниже дан шаблон для остальных — при необходимости полный список можно предоставить аналогично номерам 1–10.)
11. Chen H. J., Dai F. J., Chen C. Y., Fan S. L., Zheng J. H., Chau C. F., et al. (2023). Effects of molecular weight fraction on antioxidation capacity of rice protein hydrolysates. Sci. Rep. 13, 3464. doi: 10.1038/s41598-022-14314-7. CrossRef. Google Scholar.
... и так до 59-го источника.
продолжай
Продолжаю оформление списка литературы с 12 по 59 источники в том же формате (CrossRef и Google Scholar в одной строке, ссылки зашиты в слова).
12. Cuvas-Limón R. B., Ferreira-Santos P., Cruz M., Teixeira J. A., Belmares R., Nobre C. (2022). Novel bio-functional Aloe vera beverages fermented by probiotic Enterococcus faecium and Lactobacillus lactis. Molecules 27, 2473. doi: 10.3390/molecules27082473. CrossRef. Google Scholar.
13. Deziderio M. A., de Souza H. F., Kamimura E. S., Petrus R. R. (2023). Plant-based fermented beverages: development and characterization. Foods 12, 4128. doi: 10.3390/foods12224128. CrossRef. Google Scholar.
14. Duda-Chodak A. (2012). The inhibitory effect of polyphenols on human gut microbiota. J. Physiology Pharmacol. 63, 497–503. CrossRef (no DOI available). Google Scholar.
15. Elbatreek M. H., Mahdi I., Ouchari W., Mahmoud M. F., Sobeh M. (2023). Current advances on the therapeutic potential of pinocembrin: an updated review. Biomed. Pharmacother. 157, 114032. doi: 10.1016/j.biopha.2022.114032. CrossRef. Google Scholar.
16. Farias D. de P., Araújo F. F., Neri-Numa I. A., Dias-Audibert F. L., Delafiori J., Catharino R. R., et al. (2021). Effect of in vitro digestion on the bioaccessibility and bioactivity of phenolic compounds in fractions of Eugenia pyriformis fruit. Food Res. Int. 150, 110767. doi: 10.1016/j.foodres.2021.110767. CrossRef. Google Scholar.
17. Fernandes P. A. R., Coimbra M. A. (2023). The antioxidant activity of polysaccharides: a structure-function relationship overview. Carbohydr. Polym. 314, 120965. doi: 10.1016/j.carbpol.2023.120965. CrossRef. Google Scholar.
18. Galanakis C. M. (2021). Functionality of Food Components and Emerging Technologies. Foods 10, 128. doi: 10.3390/foods10010128. CrossRef. Google Scholar.
19. Gentile L. (2020). Protein–polysaccharide interactions and aggregates in food formulations. Curr. Opin. Colloid Interface Sci. 48, 18–27. doi: 10.1016/j.cocis.2020.03.002. CrossRef. Google Scholar.
20. Grand View Research (2024). Dairy alternatives market size and trends. *Rep. ID GVR-1-68038-070-5*. Available at: https://www.grandviewresearch.com/industry-analysis/dairy-alternatives-market (Accessed August, 2024). CrossRef. Google Scholar.
21. Hoff R., Daguer H., Deolindo C. T. P., de Melo A. P. Z., Durigon J. (2022). Phenolic compounds profile and main nutrients parameters of two underestimated non-conventional edible plants: Pereskia aculeata Mill. (ora-pro-nóbis) and Vitex megapotamica (Spreng.) Moldenke (tarumã) fruits. Food Res. Int. 162, 112042. doi: 10.1016/j.foodres.2022.112042. CrossRef. Google Scholar.
22. Kupikowska-Stobba B., Domagała J., Kasprzak M. M. (2024). Critical review of techniques for food emulsion characterization. Appl. Sci. 14, 1069. doi: 10.3390/app14031069. CrossRef. Google Scholar.
23. Liu L., Firrman J. (2016). Genetic expression profile analysis of the temporal inhibition of quercetin and naringenin on Lactobacillus rhamnosus GG. J. Probiotics Health 04, 1000139. doi: 10.4172/2329-8901.1000139. CrossRef. Google Scholar.
24. Lopes J. M. M., Lage N. N., Guerra J. F. C., Silva M., Bonomo L. F., Paulino A. H. S., et al. (2018). A preliminary exploration of the potential of Eugenia uvalha Cambess juice intake to counter oxidative stress. Food Res. Int. 105, 563–569. doi: 10.1016/j.foodres.2017.11.067. CrossRef. Google Scholar.
25. Lu Z. X., He J. F., Zhang Y. C., Bing D. J. (2020). Composition, physicochemical properties of pea protein and its application in functional foods. Crit. Rev. Food Sci. Nutr. 60, 2593–2605. doi: 10.1080/10408398.2019.1651248. CrossRef. Google Scholar.
26. Ma J., Miao Y., Li J., Ma Y., Wu M., Wang W., et al. (2022). Incorporation of blue honeysuckle juice into fermented goat milk: physicochemical, sensory and antioxidant characteristics and in vitro gastrointestinal digestion. Foods 11, 3065. doi: 10.3390/foods11193065. CrossRef. Google Scholar.
27. Manus J., Millette M., Uscanga B. R. A., Salmieri S., Maherani B., Lacroix M. (2021). In vitro protein digestibility and physico-chemical properties of lactic acid bacteria fermented beverages enriched with plant proteins. J. Food Sci. 86, 4172–4182. doi: 10.1111/1750-3841.15859. CrossRef. Google Scholar.
28. Masiá C., Geppel A., Jensen P. E., Buldo P., Smith C. J., Lluna A. G. (2021). Effect of Lactobacillus rhamnosus on physicochemical properties of fermented plant-based raw materials. Foods 10, 573. doi: 10.3390/foods10030573. CrossRef. Google Scholar.
29. McCoubrey L. E., Favaron A., Awad A., Orlu M., Gaisford S., Basit A. W. (2023). Colonic drug delivery: formulating the next generation of colon-targeted therapeutics. J. Control. Release 353, 1107–1126. doi: 10.1016/j.jconrel.2022.12.029. CrossRef. Google Scholar.
30. Mohamadzadeh M., Fazeli A., Shojaosadati S. A. (2024). Polysaccharides and proteins-based bionanocomposites for microencapsulation of probiotics to improve stability and viability in the gastrointestinal tract: a review. Int. J. Biol. Macromol. 259, 129287. doi: 10.1016/j.ijbiomac.2024.129287. CrossRef. Google Scholar.
31. Muncey L., Hekmat S. (2021). Development of probiotic almond beverage using Lacticaseibacillus rhamnosus GR-1 fortified with short-chain and long-chain inulin fibre. Fermentation 7, 90. doi: 10.3390/fermentation7020090. CrossRef. Google Scholar.
32. Olagunju A. I., Omoba O. S., Enujiugha V. N., Alashi A. M., Aluko R. E. (2018). Pigeon pea enzymatic protein hydrolysates and ultrafiltration peptide fractions as potential sources of antioxidant peptides: an in vitro study. LWT 97, 269–278. doi: 10.1016/j.lwt.2018.07.003. CrossRef. Google Scholar.
33. Olthof M. R., Hollman P. C. H., Vree T. B., Katan M. B. (2000). Bioavailabilities of quercetin-3-glucoside and quercetin-4'-glucoside do not differ in humans. J. Nutr. 130 (5), 1200–1203. doi: 10.1093/jn/130.5.1200. CrossRef. Google Scholar.
34. Popova A., Mihaylova D., Lante A. (2023). Insights and Perspectives on Plant-Based Beverages. Plants 12, 3345. doi: 10.3390/plants12193345. CrossRef. Google Scholar.
35. Prestes A. A., Verruck S., Vargas M. O., Canella M. H. M., Silva C. C., da Silva Barros E. L., et al. (2021). Influence of guabiroba pulp (Campomanesia xanthocarpa O. Berg) added to fermented milk on probiotic survival under in vitro simulated gastrointestinal conditions. Food Res. Int. 141, 110135. doi: 10.1016/j.foodres.2021.110135. CrossRef. Google Scholar.
36. Ramirez M. R., Schnorr C. E., Feistauer L. B., Apel M., Henriques A. T., Moreira J. C. F., et al. (2012). Evaluating the polyphenolic content, anti-inflammatory, and antioxidant activities of total extract from Eugenia pyriformes (uvaia) fruit. J. Food Biochem. 36, 405–412. doi: 10.1111/j.1745-4514.2011.00558.x. CrossRef. Google Scholar.
37. Reese I., Schäfer C., Ballmer-Weber B., Beyer K., Dölle-Bierke S., van Dullemen S., et al. (2023). Vegan diets from an allergy point of view - position paper of the DGAKI working group on food allergy. Allergol. Sel. 7, 57–83. doi: 10.5414/ALX02400E. CrossRef. Google Scholar.
38. Rodrigues L. M., Romanini E. B., Silva E., Pilau E. J., Da Costa S. C., Madrona G. S. (2021). Uvaia (Eugenia pyriformis Cambess) residue as a source of antioxidants: an approach to ecofriendly extraction. LWT 138, 110785. doi: 10.1016/j.lwt.2020.110785. CrossRef. Google Scholar.
39. Rufino S. M., Alves R. E., De Brito E. S., Selene M. D. M., Sampaio C. D. G., Pérez-Jiménez J., et al. (2007). Scientific methodology: determination of total antioxidant activity in fruits by DPPH free radical capture. Brasília, DF (Brazil): EMBRAPA, 127. CrossRef (no DOI available). Google Scholar.
40. Rumpf J., Burger R., Schulze M. (2023). Statistical evaluation of DPPH, ABTS, FRAP, and Folin-Ciocalteu assays to assess the antioxidant capacity of lignins. Int. J. Biol. Macromol. 233, 123470. doi: 10.1016/j.ijbiomac.2023.123470. CrossRef. Google Scholar.
41. Sganzerla W. G., Komatsu R. A., Mangrich dos Passos J. F., Costa M. D., Dobler Stroschein M. R., Schons P. F., et al. (2021). Variability in the molecular, phenological, and physicochemical characteristics of uvaia (Eugenia pyriformis Cambess - Myrtaceae) accessions from the Brazilian Atlantic rainforest. Biocatal. Agric. Biotechnol. 35, 102082. doi: 10.1016/j.bcab.2021.102082. CrossRef. Google Scholar.
42. Sheen F., Lim A. J. Y., Forde C. G. (2023). Diversity among flexitarian consumers; stratifying meat reducers by their underlying motivations to move to a plant-based diet. Food Qual. Prefer. 112, 105022. CrossRef. Google Scholar.
43. Sigüenza-Andrés T., Gómez M., Rodríguez-Nogales J. M., Caro I. (2023). Development of a fermented plant-based beverage from discarded bread flour. LWT 182, 114795. doi: 10.1016/j.lwt.2023.114795. CrossRef. Google Scholar.
44. Silva A. P. G., Sganzerla W. G., Jacomino A. P., da Silva E. P., Xiao J., Simal-Gandara J. (2022). Chemical composition, bioactive compounds, and perspectives for the industrial formulation of health products from uvaia (Eugenia pyriformis Cambess - Myrtaceae): a comprehensive review. J. Food Compos. Analysis 109, 104500. doi: 10.1016/j.jfca.2022.104500. CrossRef. Google Scholar.
45. Singleton V. L., Rossi J. A. (1965). Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 16, 144–158. doi: 10.5344/ajev.1965.16.3.144. CrossRef. Google Scholar.
46. Sorita G. D., Leimann F. V., Ferreira S. R. S. (2022). Food Bioproc. Tech. CrossRef (no DOI available). Google Scholar.
47. Spricigo P. C., Almeida L. S., Ribeiro G. H., Correia B. S. B., Taver I. B., Jacomino A. P., et al. (2023). Quality attributes and metabolic profiles of uvaia (Eugenia pyriformis), a native Brazilian Atlantic forest fruit. Foods 12, 1881. doi: 10.3390/foods12091881. CrossRef. Google Scholar.
48. Tang Z., Zhao Z., Chen S., Lin W., Wang Q., Shen N., et al. (2023). Dragon fruit-kiwi fermented beverage: in vitro digestion, untargeted metabolome analysis and anti-aging activity in Caenorhabditis elegans. Front. Nutr. 9, 1–15. doi: 10.3389/fnut.2022.1052818. CrossRef. Google Scholar.
49. Terao J. (2023). Potential role of quercetin glycosides as anti-atherosclerotic food-derived factors for human health. Antioxidants 12, 258. doi: 10.3390/antiox12020258. CrossRef. Google Scholar.
50. Verruck S., Barretta C., Miotto M., Canella M. H. M., de Liz G. R., Maran B. M., et al. (2020). Evaluation of the interaction between microencapsulated Bifidobacterium BB-12 added in goat's milk Frozen Yogurt and Escherichia coli in the large intestine. Food Res. Int. 127, 108690. doi: 10.1016/j.foodres.2019.108690. CrossRef. Google Scholar.
51. Verruck S., Prudêncio E. S., Vieira C. R. W., Amante E. R., de Mello Castanho Amboni R. D. (2015). The buffalo Minas Frescal cheese as a protective matrix of Bifidobacterium BB-12 under in vitro simulated gastrointestinal conditions. LWT 63, 1179–1183. doi: 10.1016/j.lwt.2015.04.014. CrossRef. Google Scholar.
52. Wanderley B. R. da S. M., Haas I. C. da S., Biluca F. C., Brugnerotto P., Gomes T. M., Aquino A. C. M. de S., et al. (2022). Phenolic profiling, organic acids and sugars composition of feijoa (Acca sellowiana (O. Berg) Burret) and uvaia (Eugenia pyriformis Cambess) from the southern Brazilian highlands. Cienc. Rural. 52, 1–6. doi: 10.1590/0103-8478cr20210458. CrossRef. Google Scholar.
53. Wang H., Chen L., Yang B., Du J., Chen L., Li Y., et al. (2023). Structures, sources, identification/quantification methods, health benefits, bioaccessibility, and products of isorhamnetin glycosides as Phytonutrients. Nutrients 15, 1947. doi: 10.3390/nu15081947. CrossRef. Google Scholar.
54. Wang X., Qi Y., Zheng H. (2022). Dietary polyphenol, gut microbiota, and health benefits. Antioxidants 11, 1212. doi: 10.3390/antiox11061212. CrossRef. Google Scholar.
55. Williamson G., Clifford M. N. (2024). A critical examination of human data for the biological activity of quercetin and its phase-2 conjugates. Crit. Rev. Food Sci. Nutr., 1–37. doi: 10.1080/10408398.2023.2299329. CrossRef. Google Scholar.
56. Wu C., Wang J., Liu N., Chen X., Xu H., Lei H. (2023a). Phytochemical properties and antioxidant capacities of apple juice fermented by probiotics during refrigerated storage and simulated gastrointestinal digestion. Appl. Biochem. Biotechnol. 195, 6032–6049. doi: 10.1007/s12010-022-04255-2. CrossRef. Google Scholar.
57. Wu D. T., Li W. X., Wan J. J., Hu Y. C., Gan R. Y., Zou L., et al. (2023b). Comparison of soluble dietary fibers extracted from ten traditional legumes: physicochemical properties and biological functions. Foods 12, 2352. doi: 10.3390/foods12122352. CrossRef. Google Scholar.
58. Zhang L., Wu T., Zhang Y., Chen Y., Ge X., Sui W., et al. (2023). Release of bound polyphenols from wheat bran soluble dietary fiber during simulated gastrointestinal digestion and colonic fermentation in vitro. Food Chem. 402, 134111. doi: 10.1016/j.foodchem.2022.134111. CrossRef. Google Scholar.
59. Zhang Y., Li Y., Quan Z., Xiao P., Duan J.-A. (2024). New insights into antioxidant peptides: an overview of efficient screening, evaluation models, molecular mechanisms, and applications. Antioxidants 13, 203. doi: 10.3390/antiox13020203. CrossRef. Google Scholar.

Santana de Oliveira T, Sant’Anna R, Arend GD, Sorita GD, Camelo-Silva C, Hoff RB and Verruck S (2024) Fermented plant-based beverage supplemented with uvaia (Eugenia pyriformis) pulp: an innovative and pioneering approach to diversify plant-based diet product market. Front. Food. Sci. Technol. 4:1460402. doi: 10.3389/frfst.2024.1460402
Перевод статьи «Fermented plant-based beverage supplemented with uvaia (Eugenia pyriformis) pulp: an innovative and pioneering approach to diversify plant-based diet product market» авторов Santana de Oliveira T, Sant’Anna R, Arend GD, Sorita GD, Camelo-Silva C, Hoff RB and Verruck S., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык


















































































































Комментарии (0)