Ферментация ламинарии: секрет микробов-аборигенов
В этом исследовании изучалось использование собственной (аборигенной) микробиоты сахарной ламинарии Saccharina latissima в форме ферментированной жидкости из ламинарии (ФЖЛ), полученной в результате спонтанного анаэробного процесса брожения, в качестве стартовой культуры (СК) для ферментации свежесобранной биомассы того же вида.
Аннотация
Быстрое (<48 ч) подкисление (pH < 4,3) было достигнуто для S. latissima, инокулированной ФЖЛ, при температуре ферментации 21°C. Ламинария, инокулированная коммерческим штаммом Lactiplantibacillus plantarum (Lp, положительный контроль), достигла аналогичного уровня pH через 5 дней, тогда как ламинария без СК (отрицательный контроль) не достигла pH ниже 4,3 в течение 9 дней. Микробиота ФЖЛ-СК, а также образцов S. latissima, инокулированных ФЖЛ, была доминирована молочнокислыми бактериями (МКБ), идентифицированными как L. plantarum. СК в этих образцах успешно преобразовывала маннит в молочную кислоту как основной продукт ферментации. Напротив, в образцах отрицательного контроля было зафиксировано более высокое содержание уксусной кислоты и этанола по сравнению с другими группами; это отражает иной микробный профиль, включающий морские бактерии, которые не удалось идентифицировать с помощью MALDI-TOF биотинга. Экспериментальные испытания образцов S. latissima из экспериментальных и коммерческих процессов ферментации с Bacillus cereus не привели к росту этого пищевого патогена, даже при уровнях pH, находящихся в пределах жизнеспособного диапазона для данного вида (pH > 4,3). Эти предварительные результаты создают основу для дальнейшего выделения подходящих СК для ферментации ламинарии в промышленном производстве и для оценки безопасности ферментированной ламинарии. Эффективные и безопасные процессы ферментации повысят устойчивость производства ламинарии и позволят шире использовать ингредиенты из ламинарии в пищевых продуктах.
1 Введение
Макроводоросли рассматриваются как перспективный альтернативный ресурс для получения ингредиентов для пищи и кормов, а также как соответствующее сырье для множества промышленных применений (Philis и др., 2018; Rotter и др., 2020; Blikra и др., 2021). Коммерческое культивирование макроводорослей, в основном видов ламинарии Saccharina latissima (сахарная ламинария) и Alaria esculenta, расширяется в Европе (Stévant и др., 2017). Однако крупномасштабное культивирование ламинарии влечет за собой технические проблемы, связанные с обработкой, переработкой и хранением больших объемов биомассы. Saccharina latissima портится в течение 9 дней после сбора урожая при охлажденной температуре из-за высокого содержания воды (Wirenfeldt и др., 2022). Поэтому для стабилизации сырья необходимо применять эффективные методы. Замораживание и низкотемпературная сушка являются широко применяемыми методами, обеспечивающими получение высококачественных и универсальных продуктов с длительным сроком хранения при правильном хранении (Stévant and Rebours, 2021). Эти методы также являются энергоемкими, что снижает рентабельность и экологическую устойчивость производственно-сбытовой цепочки и ограничивает их применимость при переработке больших объемов биомассы (Philis и др., 2018; Thomas и др., 2020). Решение этих технических проблем с акцентом на устойчивость является ключевым для развития процветающей и экологически ответственной индустрии культивирования ламинарии в Европе. В этом контексте ферментация была определена как устойчивый метод консервации биомассы ламинарии (Stévant and Rebours, 2021; Thomas и др., 2020; Emblemsvåg и др., 2020; Sørensen и др., 2021).
Ферментация, индуцированная молочнокислыми бактериями (МКБ), является интересной альтернативой для стабилизации собранной ламинарии из-за низких потребностей в энергии и сложном оборудовании и набирает популярность среди производителей ламинарии (Stévant and Rebours, 2021). Методы ферментации МКБ в анаэробных условиях (также называемые "силосованием") являются методами консервации, широко применяемыми в сельском хозяйстве для продуктов питания и кормов. Успешная ферментация снижает pH (обычно ниже 4,0), чтобы ингибировать рост микроорганизмов, вызывающих порчу (например, клостридий и плесени). Низкий pH может быть достигнут непосредственно добавлением органической кислоты или косвенно путем инокуляции биомассы стартовой культурой (СК) МКБ, которые затем преобразуют сбраживаемые сахара в молочную кислоту. Ферментация макроводорослей, индуцированная МКБ, является новой стратегией, и оптимизированные протоколы, включающие адаптированные СК для стабилизации коммерчески значимых видов (например, S. latissima и A. esculenta), еще не установлены. Макроводоросли богаты полисахаридами, такими как альгинат, каррагинан и агар, которые структурно сложны и нелегко сбраживаются обычными МКБ. Это противоречит хорошо зарекомендовавшим себя методам ферментации, основанным на овощах, содержащих сбраживаемые глюкозу, сахарозу и фруктозу, в то время как молочные продукты содержат лактозу, которая может легко сбраживаться МКБ. Кроме того, овощи естественным образом содержат МКБ, а молочная среда благоприятствует росту МКБ (например, Lactobacillus spp. и Lactococcus spp.). Собственная микробиота макроводорослей может адаптироваться к морским условиям и может конкурировать с желательными ферментативными микробами, потенциально производя неприятные привкусы и токсины.
Химический состав макроводорослей, характеризующийся низким содержанием сухого вещества и высокой буферной емкостью, связанной с высоким уровнем анионов (например, хлорида, сульфата), представляется проблемой для успешной ферментации, с вариабельными результатами в разных видов (Herrmann и др., 2015; Cabrita и др., 2017). Ламинария S. latissima показала лучший потенциал ферментации в анаэробных условиях по сравнению с другими видами и производила молочную кислоту из собственных популяций МКБ (Herrmann и др., 2015). Спонтанная анаэробная ферментация S. latissima (т.е. без добавления СК) также была описана в более поздних исследованиях (Sørensen и др., 2021; Stévant, 2019; Krook и др., 2024). Однако из-за вариабельности собственной микробиоты, приводящей к ограниченному контролю над процессом ферментации и непредсказуемому качеству ферментированной биомассы (например, развитию микроорганизмов, вызывающих порчу, и несогласованным сенсорным профилям), внедрение этой техники в коммерческом масштабе остается ограниченным. Другие проблемы для прогнозируемой ферментации ламинарии включают вариабельность содержания водорастворимых полисахаридов и полиолов (ламинарана и маннита в ламинариях) в качестве субстратов для ферментации, индуцированной МКБ, низкую исходную численность МКБ и неоптимальные СК для сырья. Дополнительные этапы, такие как ферментативный гидролиз (Uchida and Miyoshi, 2013), тепловая обработка (Bruhn и др., 2019) или добавление сбраживаемых сахаров (например, патоки) (Larsen и др., 2021), могут способствовать быстрому подкислению, но также добавляют сложность, которая может ограничить применимость процесса к крупномасштабному коммерческому производству.
В настоящее время доступные СК для ферментации ламинарии полагаются на коммерческие штаммы из сельского хозяйства, и знания о морских МКБ для ферментации макроводорослей отсутствуют (Uchida and Miyoshi, 2013; Uchida и др., 2004). Благодаря их адаптируемости к специфической среде ламинарии, такой как высокая соленость и маннит в качестве основного источника углерода, собственная микробиота может предоставить превосходную альтернативу существующим коммерческим штаммам МКБ для ферментации ламинарии. Основная цель этого исследования состояла в том, чтобы выяснить, может ли собственная микробиота (предположительно МКБ) S. latissima быть использована в качестве СК для дальнейшей ферментации ламинарии. Эта собственная микробиота была отобрана из спонтанной ферментации культивируемой S. latissima в анаэробных условиях, причем жидкость, полученная в результате этого процесса, называемая "ферментированной жидкостью из ламинарии" (ФЖЛ), использовалась в качестве СК. Ферментация S. latissima с использованием этого метода была сравнена с аналогичным процессом с использованием обычной СК Lactiplantibacillus plantarum (Lp, положительный контроль) или без СК (т.е. спонтанная ферментация, отрицательный контроль). pH образцов контролировали на протяжении всего процесса, и микробиота ферментированной и свежей ламинарии, а также ФЖЛ-СК была охарактеризована. Также были рассмотрены аспекты безопасности ферментированной S. latissima.
Патогенные Bacillus spp., Vibrio spp. и Aeromonas spp. были идентифицированы как основные собственные бактерии, вызывающие особую озабоченность с точки зрения пищевой безопасности макроводорослей, тогда как несколько других видов бактерий, включая Escherichia coli, Salmonella spp., Staphylococcus aureus и Listeria monocytogenes, считаются потенциальными проблемами пищевой безопасности из-за повторного заражения во время обработки. Эти бактерии эффективно инактивируются снижением pH ниже 4,3 при хранении при охлажденных температурах или ниже pH 3,7 при хранении при комнатной температуре (Løvdal и др., 2021). Пищевые патогены из рода Bacillus представляют особый интерес, поскольку они могут продуцировать споры, которые позволяют бактериям оставаться в состоянии покоя в экстремальных условиях, и могут прорастать, развиваться и продуцировать токсины после наступления благоприятных условий. Токсинпродуцирующие Bacillus spp., включая B. cereus, B. pumilus и B. licheniformis, были идентифицированы в низких концентрациях на поверхности свежих и сушеных съедобных ламинарий (Blikra и др., 2018; Lytou и др., 2021; Martelli и др., 2021). Микробиологическая безопасность ферментированной ламинарии в отношении этих патогенов в настоящее время не установлена. Вторая цель этого исследования состояла в том, чтобы провести экспериментальные испытания с B. cereus на образцах S. latissima из экспериментальных и коммерческих процессов ферментации для оценки безопасности продуктов из ламинарии в отношении патогена.
2 Материалы и методы
2.1 Экспериментальная ферментация S. latissima с изучением потенциала ферментированной жидкости из ламинарии в качестве стартовой культуры
2.1.1 Приготовление ФЖЛ и СК L. plantarum
Saccharina latissima культивировали в Tango Seaweed AS (Херёй, Норвегия; широта 62.1903 °N, долгота 5.4473 °E) и собрали 26 апреля 2022 года. Биомасса, состоящая из слоевищ, ножек и ризоидов, была грубо измельчена, а затем перенесена в 30-литровые ферментационные ведра, оснащенные гидрозатвором. Три ведра-повторности получили 18.2 ± 1.1 кг свежей S. latissima, которую плотно спрессовали, чтобы минимизировать захваченные пузырьки воздуха и свободное пространство под крышкой. Ведра транспортировали в лабораторию и хранили при 21°C в темноте. Через 15 дней хранения в этих условиях их открыли и собрали жидкую фракцию (т.е. ФЖЛ, образовавшуюся во время этой спонтанной ферментации). Каждое ведро произвело приблизительно 7 л ФЖЛ. ФЖЛ из одной из повторностей характеризовалась неприятным кислым запахом и поэтому была отброшена. ФЖЛ из двух оставшихся повторностей гомогенизировали для использования в качестве СК для ферментации свежей биомассы ламинарии. Ведра с ФЖЛ были извлечены из хранилища при 21°C до комнатной температуры окружающей среды (прибл. 15°C) примерно за 2 часа до начала эксперимента. Температура ФЖЛ-СК при инокуляции предположительно была близка к 21°C. Оценка плотности МКБ в ФЖЛ-СК через 12 дней хранения с помощью высева на чашки (3M Petrifilm Пластины для подсчета молочнокислых бактерий) не удалась из-за переросших чашек, поэтому плотность МКБ во время инокуляции свежей ламинарии ФЖЛ-СК была неизвестна. Высев на чашки был предпринят снова в начале эксперимента и показал плотность МКБ 6 log (КОЕ мл−1). pH ФЖЛ-СК составлял 3.6.
Коммерческий штамм L. plantarum (PAL LACTIC LP 2233 VEG RM D10, Laboratoire Standa, Франция) использовали в качестве положительного контроля для ферментации S. latissima на основе предыдущих успешных результатов (Stévant, 2019). Лиофилизированные бактерии регидратировали в пептонной воде в течение 30 минут перед инокуляцией в количестве 6 log (КОЕ г−1) свежей ламинарии, как рекомендовано производителем закваски.
2.1.2 Эксперименты по ферментации
Биомассу S. latissima культивировали в Tango Seaweed AS и собирали 9 мая 2022 года. Слоевища ламинарии, все еще прикрепленные к несущим тросам, транспортировали в лабораторию в течение 2 часов, а затем перенесли в емкости с подачей морской воды при 7.5°C.
Эксперимент по ферментации включал пять обработок, предусматривающих добавление различных СК (ФЖЛ, коммерческий штамм L. plantarum и без добавления СК) и различные температуры хранения образцов во время ферментации (10°C и 21°C). Эти температуры были выбраны в соответствии с оптимальным ростом МКБ (21°C) и реалистичными условиями переработки ламинарии при комнатной температуре в Норвегии в мае (10°C). Для каждой повторности 3 кг S. latissima брали из емкости, грубо измельчали и оставляли стекать на 5 минут. Затем биомассу тщательно смешивали с СК в большом чане, предварительно продезинфицированном, и разделяли на полиэтиленовые вакуумные пакеты примерно по 500 г. Вакуумные пакеты хранили в темноте либо при 21°C в помещении с контролируемой температурой, либо при 10°C в инкубаторе. Вес каждого пакета регистрировали с точностью до одного десятичного знака. Экспериментальный протокол, использованный в этом исследовании, и обработка образцов обобщены на Рисунке 1. Обработки ферментации проводили в трех повторностях. Детали каждой обработки обобщены в Таблице 1. Образцы в отдельных вакуумных пакетах вскрывали на дни 1, 2, 5 и 9 после инокуляции СК для мониторинга показателей качества во время процесса и для дальнейшей характеристики.
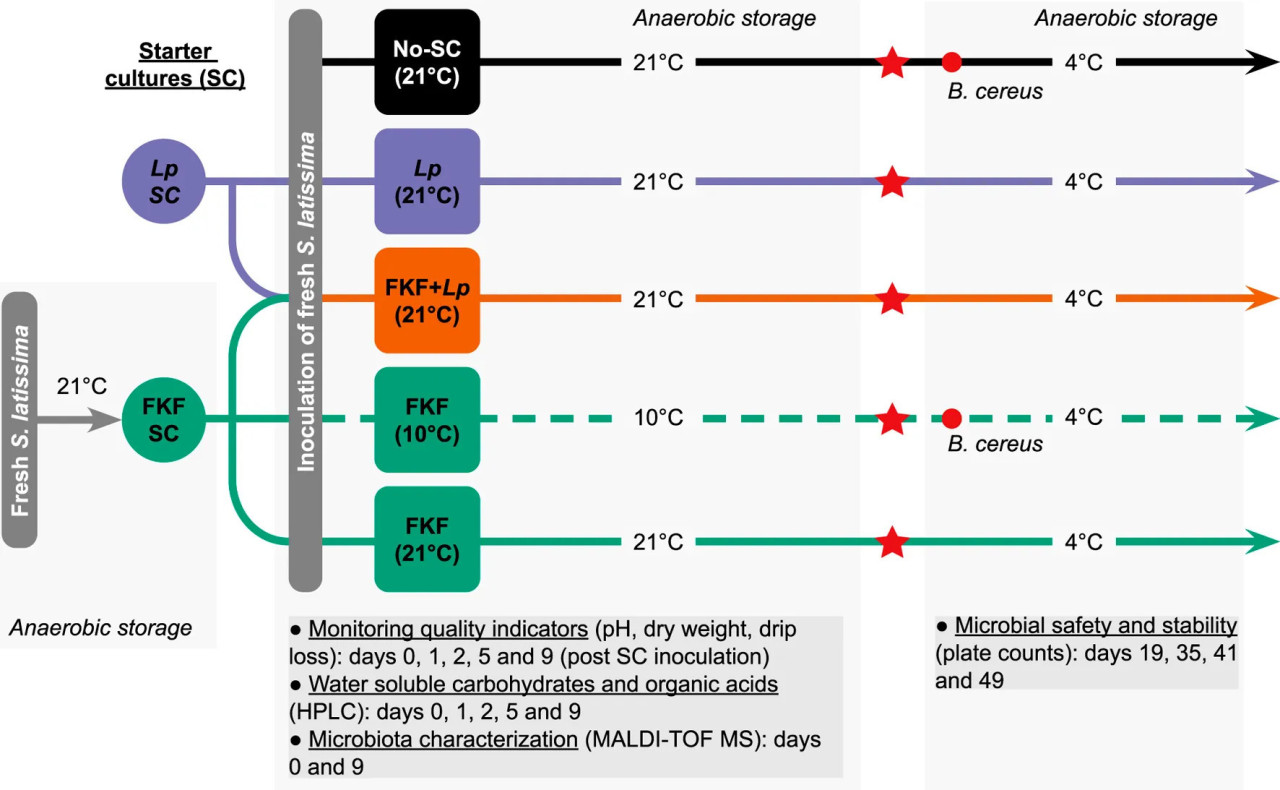
РИСУНОК 1 Экспериментальный дизайн для изучения потенциала ферментированной жидкости из ламинарии (ФЖЛ) по сравнению с Lactiplantibacillus plantarum (Lp) в качестве стартовой культуры (СК) для ферментации Saccharina latissima. Красные звездочки указывают, что образцы были вскрыты, осушены и восстановлены (вакуумированы) по прибытии в лабораторные помещения Nofima. Красные кружки указывают на заражение образцов Bacillus cereus на 19 день после инокуляции свежих образцов S. latissima.
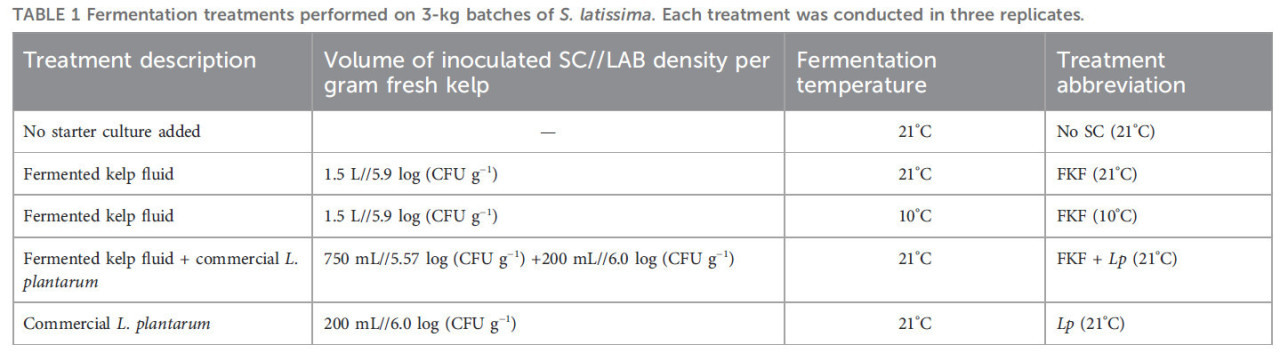
ТАБЛИЦА 1 Обработки ферментации, проведенные на 3-кг партиях S. latissima. Каждая обработка проводилась в трех повторностях.
2.1.3 Сухой вес и потери при стекании
После вскрытия содержимое каждого пакета с образцом переносили на сито, помещенное над стаканом, и давали стечь в течение 5 минут. Потери при стекании (дрип-потери) рассчитывали с использованием Уравнения 1, где M0 — масса исходного образца, а Mt — масса твердой фракции в момент времени t.
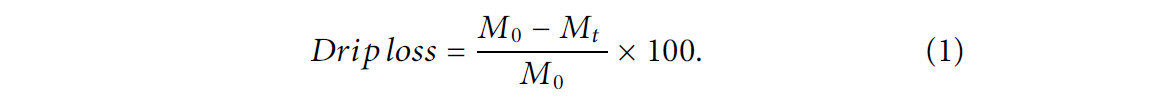
Сухой вес (СВ) твердой фракции образцов определяли гравиметрически как остаток после высушивания в лабораторной печи при 105°C в течение 24 часов. Каждое измерение СВ проводили в трех повторностях, используя случайно выбранные кусочки образца. pH жидкой фракции образцов измеряли с помощью pH-зонда (pH-метр 340 с pH-электродом InLab Expert Pro-C, Mettler Toledo, Огайо, США).
2.1.4 Анализ водорастворимых углеводов и органических кислот
Содержание водорастворимых углеводов (ВРУ) и органических кислот в исходной биомассе до ферментации и ферментированных образцах анализировали методом высокоэффективной жидкостной хроматографии (ВЭЖХ) согласно Sørensen и др. (2021). Для каждого образца несколько граммов влажного твердого материала переносили в zip-пакет и затем хранили замороженными (-20°C) до окончания эксперимента. Один грамм размороженных образцов добавляли к 5 мл серной кислоты (H2SO4, 5 мМ) и затем хранили overnight при 4°C, после чего снова добавляли еще 5 мл H2SO4 той же концентрации. Образцы перемешивали пятикратным переворачиванием пробирок и центрифугировали при 2,795 g в течение 8 минут. 1 мл супернатанта фильтровали через 0.22-мкм полиэфирсульфоновый (ПЭС) фильтр во флаконы для ВЭЖХ. ВЭЖХ анализ проводили на колонке Aminex HPX-87H (Bio-Rad, Геркулес, США) на системе Ultimate 3000 HPLC (Dionex, Thermo Fischer Scientific, США), оснащенной рефрактометрическим детектором Shodex RI-101 (Showa Denko K.K., Токио, Япония) и диодно-матричным детектором УФ (Thermo Fischer Scientific, США). H2SO4 (5 мМ) использовали в качестве подвижной фазы. Температура колоночного термостата была установлена на 60°C, и скорость потока 0.5 мл мин−1 использовалась для всех образцов. Стандарты для каждого углевода и первичных органических метаболитов класса ВЭЖХ (глюкоза, маннит, фукоза, молочная кислота, уксусная кислота и этанол) были приготовлены в концентрациях 50, 100, 500 и 1,000 мкг мл−1 в 5 мМ H2SO4 и профильтрованы через 0.22 ПЭС-фильтр. Результаты были проанализированы с использованием программного обеспечения Chromeleon 6.0 (Thermo Fischer Scientific, Массачусетс, США).
2.1.5 Характеристика микробиоты образцов с помощью MALDI-TOF MS
Микробиоту ФЖЛ-СК, свежей биомассы S. latissima до инокуляции СК и ферментированных образцов через 9 дней исследовали методом матрично-активированной лазерной десорбции/ионизации времяпролетной масс-спектрометрии (MALDI-TOF MS) биотинга в соответствии с протоколом, установленным в Техническом университете Дании (Sedó Molina и др., 2022). В 0.5 мл глицерина (50%) в криопробирке добавляли 0.5 мл жидкого образца, который затем аккуратно перемешивали и хранили при -80°C до анализа. Для свежих образцов S. latissima до ферментации стерильным тампоном проводили по измельченным слоевищам ламинарии в течение 10 секунд, а затем переносили в 0.5 мл 50% глицерина. Замороженные глицериновые запасы высевали на следующие четыре твердые питательные среды: агар de Man, Rogosa и Sharpe (MRS) (Oxoid Ltd., Великобритания) для МКБ; агар Luria–Bertani (LB) (Sigma-Aldrich, Миссури, США) в качестве неселективной питательной среды; агар на основе дрожжевого экстракта, пептона и декстрозы (YPD) (Sigma-Aldrich, Миссури, США); и морской агар (Condalab, Испания) для гетеротрофных морских бактерий. Отдельные колонии пересевали до трех раз до чистоты, а затем свежие бактериальные колонии инокулировали в соответствующие бульонные среды без агара на 18 часов при 30°C. Для экстракции белков (в соответствии с протоколом производителя) культуры overnight центрифугировали при 3,500 × g (5 минут при 4°C), дважды промывали 0.9% солевым раствором и смешивали с i) водой MilliQ и ii) 96% этанолом перед еще одним этапом центрифугирования для удаления интерферирующих веществ. После этого клеточные осадки смешивали с 70% муравьиной кислотой и чистым ацетонитрилом с последующим встряхиванием на вортексе и центрифугировали при 14,000 g в течение 3 минут. По 1 мкл супернатанта наносили на многоразовую стальную мишень MALDI-TOF (Bruker Corp., Массачусетс, США) в двух повторностях и оставляли высыхать перед добавлением насыщенного раствора матрицы, содержащего α-циано-4-гидроксикоричную кислоту (трифторуксусная кислота:вода MilliQ:ацетонитрил 1:19:20). Планшет загружали в MALDI-TOF Biotyper Sirius (Bruker Corp., Массачусетс, США), и спектры образцов сравнивали с бактериальной (BDAL) и грибковой (FFL) библиотеками спектров от Bruker Corp.
2.1.6 Микробиологическая стабильность и безопасность при охлажденном хранении ферментированных образцов
Три повторных образца из каждой обработки были отправлены с доставкой overnight в лабораторию Nofima (Ставангер, Норвегия) на 12 день после инокуляции СК свежей биомассы S. latissima. Образцы транспортировали и хранили в охлажденном состоянии (4°C). Вакуумные пакеты были вскрыты на 19 день после инокуляции СК, затем содержимое осушено, разделено на аликвоты по 25 г, повторно упаковано под вакуумом в пакеты для су-вид и хранилось при 4°C до дальнейшего анализа (Рисунок 1). Отбор проб для анализа pH и микробиологического анализа проводили на дни 19, 35, 41 и 49 после инокуляции СК.
Аликвоты по 10 г помещали в пакеты Stomacher 400 с фильтрами (Seward Ltd., Великобритания) и разводили в десять раз в стерильном буферном пептонном растворе (Merck, Германия) и гомогенизировали в течение 120 секунд в гомогенизаторе Smasher (bioMérieux, Франция). Гомогенаты далее разводили до соответствующих концентраций в стерильной воде перед высевом. Общее количество аэробных бактерий (ОКАБ) определяли высевом на триптиказо-соевый агар с 0.6% дрожжевым экстрактом (TSAYE) (Oxoid AS, Норвегия) и инкубировали при 25°C в течение 48 часов перед подсчетом колоний и расчетом концентраций, выраженных в колониеобразующих единицах (КОЕ) на грамм образца. Для подсчета МКБ гомогенаты высевали на агар MRS, дополненный амфотерицином (Oxoid AS, Норвегия), и инкубировали в анаэробных условиях при 25°C в течение 48 часов в соответствии с Северным комитетом по анализу пищевых продуктов (NMKL:140, 2023). Дрожжи и плесени подсчитывали высевом на агар с дихлораном и розой бенгальской (DRBC) (Oxoid AS, Норвегия) и инкубированием при 25°C в течение 5–7 дней в соответствии с NMKL:98 (NMKL:98, 2005). Присутствие аэробных и анаэробных спорообразователей анализировали нагреванием аликвот гомогенатов при 80°C в течение 10 минут перед высевом на кровяной агар. Параллельные чашки инкубировали аэробно и анаэробно, соответственно, в течение 24 и 48 часов при 37°C (NMKL:189, 2011). Колиформные бактерии исследовали высевом 0.1 мл и 1 мл гомогената образца на пластины Neogen Petrifilm для подсчета колиформных бактерий (Nerliens Meszansky, Норвегия) в соответствии с инструкциями производителя (Blikra и др., 2018). Listeria monocytogenes анализировали после обогащения в бульоне Фрейзера половинной и полной концентрации (Oxoid AS, Норвегия) и подтверждения высевом на агар Brilliance Listeria (Oxoid AS, Норвегия) в соответствии с NMKL:136 (2010). Патогенные виды Vibrio анализировали в соответствии с NMKL:156 (1997) высевом на агар с тиосульфатом, цитратом, желчными солями и сахарозой (TCBS) (Oxoid AS, Норвегия) и инкубированием при 37°C в течение 24 часов.
2.1.7 Экспериментальное заражение B. cereus
Экспериментальное заражение было проведено на двух образцах с самым высоким pH через 19 дней после инокуляции образцов ламинарии, т.е. контроль без добавленной СК (Без-СК; pH 4.38 ± 0.06) и образцы с добавлением ФЖЛ, хранившиеся при 10°C (pH 4.34 ± 0.04) (Рисунок 1). Bacillus cereus (штамм NVH 1230-88), холодоустойчивый штамм, выделенный из восточного рагу, вызывающий пищевые отравления (Ehling-Schulz и др., 2005); его выращивали overnight в TSBYE при 37°C и встряхивании (225 об/мин). Полученную культуру разводили в фосфатно-солевом буфере (PBS), и 1 мл разведенной культуры инокулировали в 10 г образцов S. latissima в пластиковых пакетах, что давало конечную концентрацию примерно 3 log (КОЕ г−1). Пластиковые пакеты вакуумировали и хранили при 4°C. Отбор проб для B. cereus проводили на дни 0, 7, 21 и 30 после инокуляции патогеном. Содержимое пакетов переносили в стомачерный пакет с фильтром и разводили в десять раз в буферном пептонном растворе (BPW) и гомогенизировали в течение 120 секунд, как описано выше. Соответствующие разведения гомогенатов высевали на кровяной агар (Oxoid AS, Норвегия) и инкубировали аэробно при 30°C в течение 24 часов (NMKL:67, 2021).
2.2 Экспериментальное заражение B. cereus коммерчески ферментированной S. latissima
Saccharina latissima, культивируемая Arctic Seaweed AS (Вэрландет, Норвегия; широта 61.3001 °N, долгота 4.7485 °E) и собранная в мае 2023 года, была нарезана на кусочки примерно 2 × 2 см, и по 100 г помещено в 24 пакета для су-вид. Патоку (93.8 г, Champion Melasse, Felleskjøpet, Норвегия) растворяли в 375 мл морской воды, предварительно профильтрованной через 47-мм поликарбонатный фильтр (Poretics™) с размером пор 0.45 мкм (Thermo Fisher Scientific, Массачусетс, США), и добавляли 0.94 г коммерческой лиофилизированной СК Latilactobacillus sakei (SafePro Flora Ctrl 01, Chr. Hansen, Дания). К S. latissima в пакетах для су-вид добавляли 10 мл этого раствора. Культуры B. cereus NVH 1230-88 (Ehling-Schulz и др., 2005) и B. cereus ATCC 9139 (López-Pedemonte и др., 2003), которые способны расти при температурах <10°C, были приготовлены, как описано выше. 1 мл каждого штамма осаждали центрифугированием и ресуспендировали в 1 мл PBS и объединяли. Суспензию B. cereus дополнительно разводили в автоклавированной воде MilliQ, и 1 мл инокулировали в 15 из пакетов для су-вид для достижения конечной концентрации B. cereus примерно 4 log (КОЕ г−1). В оставшиеся девять пакетов для су-вид (контроли) добавляли 1 мл автоклавированной воды MilliQ. Три инокулированных и три контрольных пакета были немедленно отобраны для анализа pH, B. cereus, МКБ и ОКАБ. Оставшиеся пакеты вакуумировали, и инокулированные пакеты хранили при 10 или 15°C соответственно. Остальные контрольные пакеты хранили при 10°C. Каждое условие обработки (т.е. инокуляция B. cereus и температура хранения) было повторено трижды. Образцы из каждой группы обработки отбирали через 48 и 72 часа хранения и анализировали. pH измеряли в жидкой фазе с использованием pH-метра FiveEasy Plus с зондом LE410 (Mettler Toledo, Огайо, США). Аликвоты по 10 г разводили в десять раз в BPW, гомогенизировали, и подсчитывали B. cereus, ОКАБ и МКБ, как описано выше.
2.3 Анализ данных
Статистический анализ результатов проводили с использованием R [версия 4.2.2 (R Development Core Team, 2022)]. Результаты анализа повторных образцов описывали как среднее ± стандартное отклонение. Различия в эволюции pH, ВРУ, органических кислот и потерь при стекании между условиями обработки (СК и температура хранения) анализировали с помощью дисперсионного анализа повторных измерений [RM ANOVA, функция R lmer() из пакета lme4 (Bates и др., 2015)], рассматривая образцы из повторностей ферментационной обработки как случайный фактор. Попарные сравнения наименьших квадратов средних [функция R lsmeans() из пакета emmeans (Lenth, 2016)] проводили после значимых результатов RM ANOVA. В случаях ненормального распределения данных и неоднородности дисперсий результаты анализировали с помощью дисперсионного анализа на основе выровненных ранговых преобразований [ART ANOVA, функция R art() из пакета ARTool (Kay и др., 2021)], который представляет собой непараметрический подход к факторному дисперсионному анализу. Попарные сравнения проводили с использованием функции art.con().
3 Результаты
3.1 Экспериментальная ферментация S. latissima с использованием собственной флоры ламинарии
Жидкая фракция, полученная в результате спонтанной ферментации предыдущей партии культивируемой S. latissima (т.е. ФЖЛ-СК), была проверена в качестве инокулята для ферментации свежесобранной биомассы. Эффективность полученного процесса ферментации при 21°C, характеризующаяся снижением pH, была сравнена с ферментацией S. latissima в тех же условиях с использованием Lp в качестве СК, без СК и комбинированной инокуляции биомассы ламинарии ФЖЛ и Lp (ФЖЛ + Lp). Также была изучена эффективность ФЖЛ при низкой температуре (10°C). Проверенные условия (т.е. СК и температура) значительно повлияли на изменения pH в ходе эксперимента (RM ANOVA, F4, 10 = 18.38, p < 0.001). pH всех групп тестирования был значительно ниже, чем в отрицательном контроле (Без-СК) на протяжении всего эксперимента, за исключением группы с добавлением ФЖЛ, хранившейся при 10°C (Рисунок 2). Образцы, инокулированные ФЖЛ и ФЖЛ + Lp, хранившиеся при 21°C, достигли pH ниже 4.3 (т.е. 4.1 ± 0.0 и 4.0 ± 0.2 соответственно) в течение 2 дней. Образцы, инокулированные только Lp, достигли аналогичного низкого pH (3.9 ± 0.1) через 5 дней, тогда как средний pH отрицательного контроля и образцов с добавлением ФЖЛ, хранившихся при 21°C, составлял 4.4 ± 0.5 и 4.3 ± 0.2 соответственно через 10 дней.
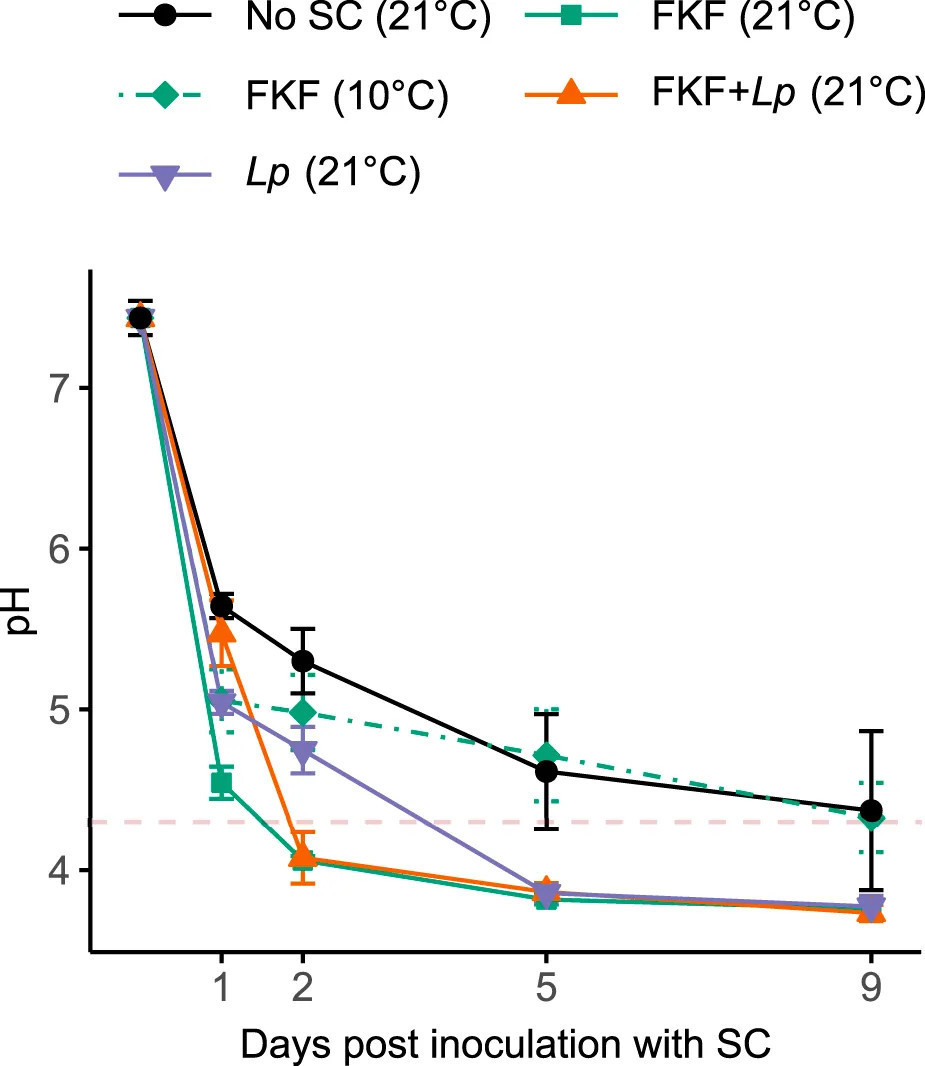
РИСУНОК 2 Изменение pH во время ферментации образцов S. latissima после добавления различных СК. Значения представлены как среднее ± ст. откл. (n = 3). Пунктирная красная линия представляет порог безопасности pH 4.3.
Результаты анализа сахаров и органических кислот в образцах в ходе эксперимента показывают более высокие уровни молочной кислоты в образцах, инокулированных ФЖЛ и/или Lp и хранившихся при 21°C, по сравнению с отрицательным контролем и образцами, инокулированными ФЖЛ и хранившимися при 10°C (LSmeans, p < 0.009) (Рисунок 3). Повышение уровней молочной кислоты в образцах соответствовало снижению маннита во всех образцах (незначимое различие между обработками, RM ANOVA F4, 40 = 2.60, p = 0.50). Исходное содержание глюкозы и фукозы было низким — 0.43 ± 0.06 и 0.30 ± 0.03 г (100 г)−1 СВ соответственно — по сравнению с маннитом 24.1 ± 7.4 г (100 г)−1 СВ. Небольшие количества уксусной кислоты и этанола были обнаружены во всех образцах. В каждой группе уровни уксусной кислоты и этанола значительно не различались между образцами, взятыми на 1, 2, 5 и 9 дни, за исключением образцов Без-СК, в которых были измерены значительно более высокие уровни этанола через 9 дней ферментации (LSmeans, p < 0.001). Относительно высокие стандартные отклонения, наблюдаемые в образцах, могут отражать переменное бактериальное равновесие между повторностями обработок во время экспериментальной ферментации. Подготовка образцов для анализа, вероятно, является дополнительным источником вариабельности.
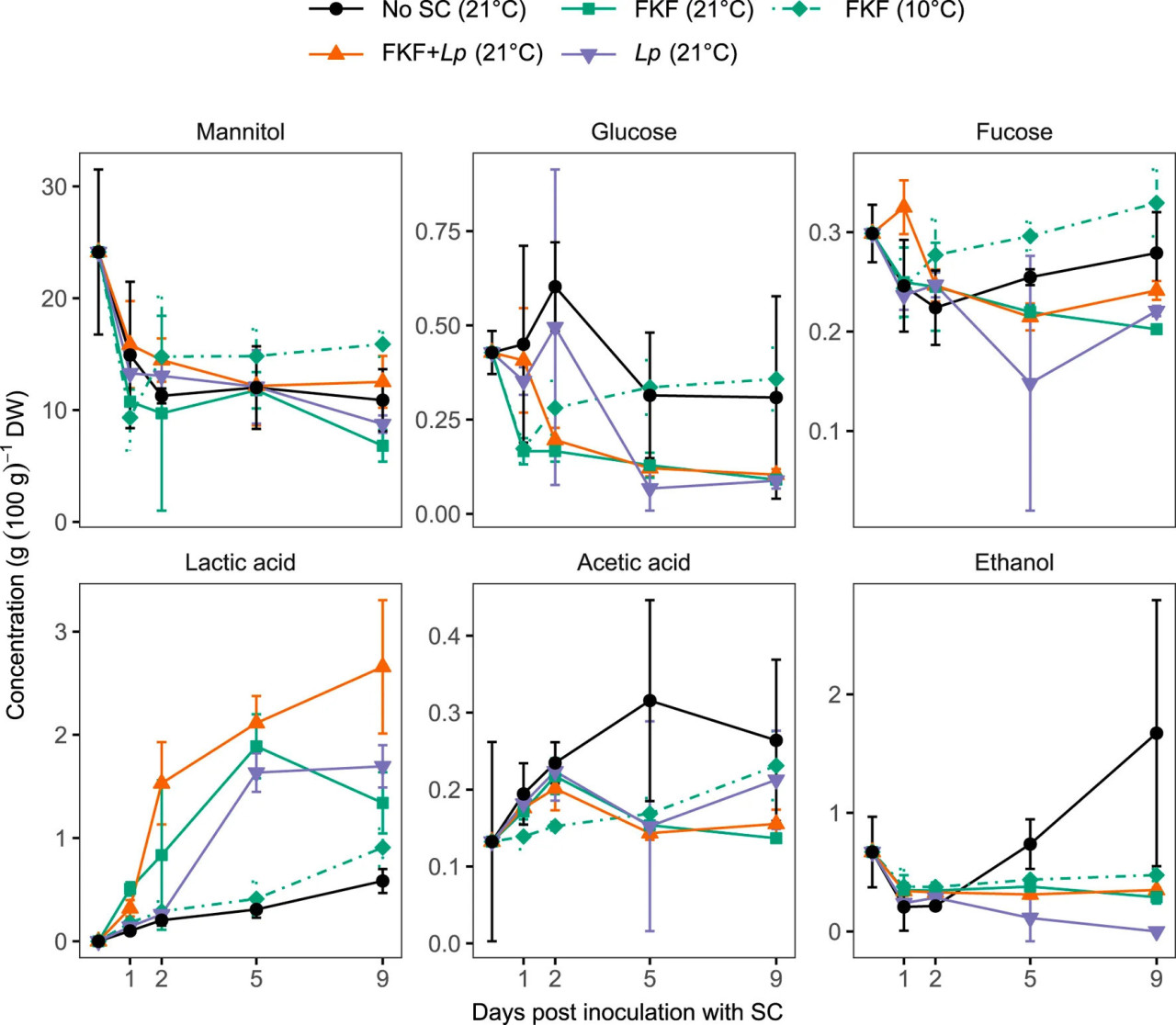
РИСУНОК 3 Изменение содержания водорастворимых углеводов, органических кислот и этанола в образцах S. latissima во время ферментации после добавления различных СК. Значения представлены как среднее ± ст. откл. (n = 3).
Потери при стекании (т.е. образование экссудата), измеренные в группах обработки через 1 день, находились в диапазоне от 18% до 31%, затем от 31% до 37% через 10 дней (Рисунок 4). Несмотря на относительно высокие вариации, измеренные в повторностях образцов, наблюдалась тенденция к более высоким потерям при стекании в образцах ФЖЛ + Lp и более низким в образцах Без-СК на протяжении эксперимента. Личные наблюдения при обращении с образцами показали более слизистую текстуру образцов, инокулированных ФЖЛ (как хранившихся при 10°C, так и при 21°C), по сравнению с другими группами образцов.
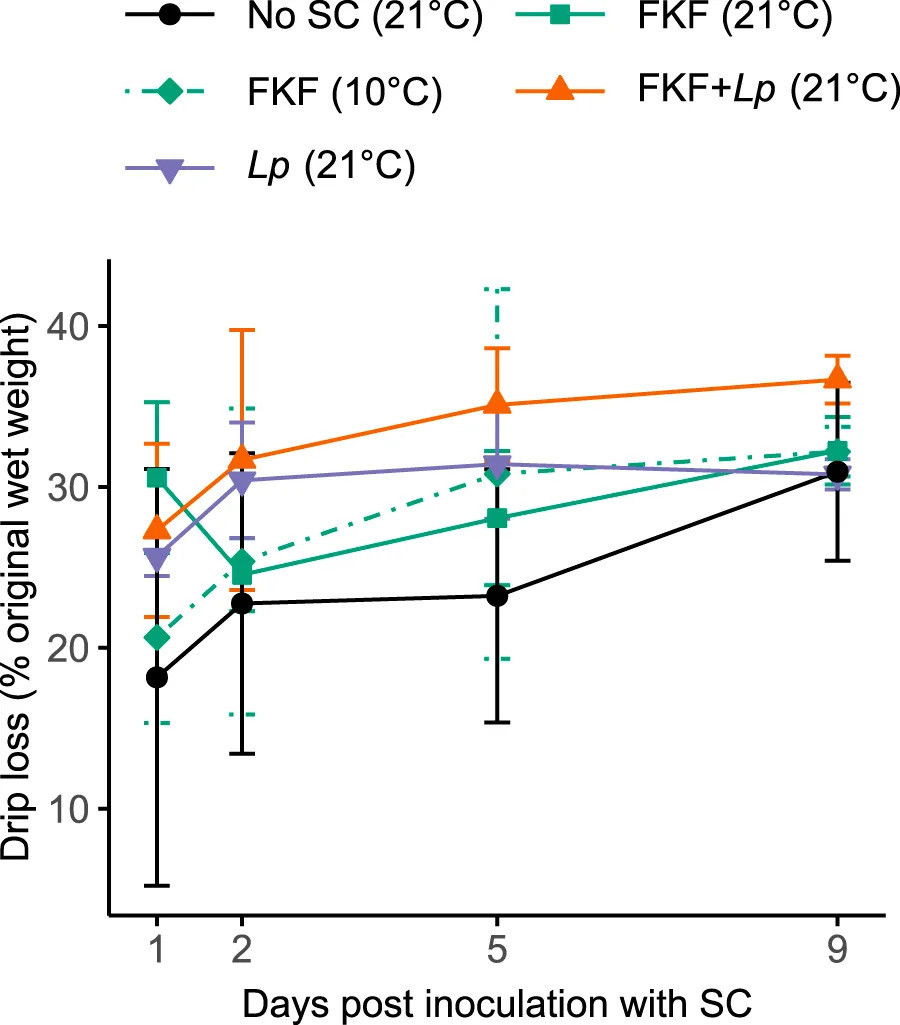
РИСУНОК 4 Потери при стекании во время ферментации S. latissima после добавления различных СК. Значения представлены как среднее ± ст. откл. (n = 3).
3.2 Микробиология ферментированной S. latissima с использованием собственной биоты ламинарии
Микробиота сырой (неферментированной) S. latissima, ФЖЛ-СК (т.е. жидкости, использованной в качестве СК) и ферментированной биомассы была охарактеризована с помощью MALDI-TOF MS. Микробиота свежей ламинарии была доминирована морскими бактериями, которые не могли быть идентифицированы с помощью MALDI-TOF Biotyper (Рисунок 5). Из этих образцов не было выделено колоний на питательной среде MRS, тогда как некоторые были выделены, но не идентифицированы в ферментированных контрольных образцах (Без-СК). Во многих случаях идентификация изолятов с морского агара не удалась (из-за низких значений оценки), за исключением Lelliotia amnigena и Pseudomonas luteola, идентифицированных в образцах Без-СК. Состав микробиоты инокулированных образцов и ФЖЛ-СК был относительно схожим между собой и доминировался лактобациллами. Морские бактерии не были выделены из этих образцов. В случае агара LB, который является неселективной средой, поддерживающей быстрый рост высокого выхода бактериальных клеток, большинство колоний не были идентифицированы с приемлемым значением оценки. Наибольшее количество колоний из ферментированной ламинарии было получено из образцов, инокулированных ФЖЛ и хранившихся при 21°C, а наименьшее — из аналогичной обработки, но хранившейся при 10°C. Изоляты из очищенных колоний на агаре MRS были успешно идентифицированы как L. plantarum как в ФЖЛ-СК, так и в образцах, инокулированных ФЖЛ и Lp. Дрожжи были выделены из всех образцов и успешно идентифицированы во многих случаях.
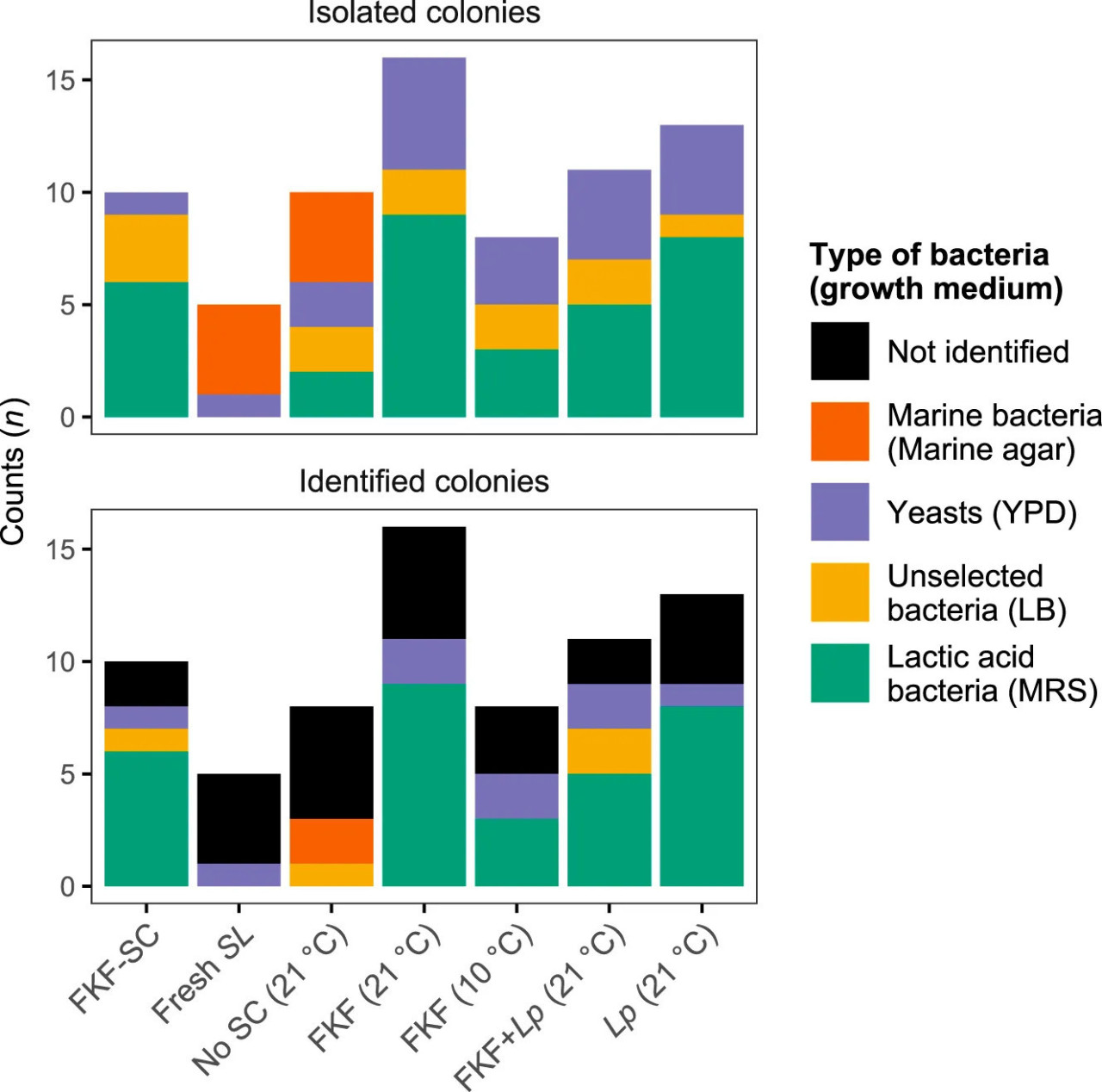
РИСУНОК 5 Количество колоний бактерий и дрожжей (средние значения для повторных образцов, n = 3), выделенных с использованием четырех различных твердых питательных сред из образцов ламинарии: свежая S. latissima (SL) перед инокуляцией, стартовая культура из ферментированной жидкости ламинарии (ФЖЛ-СК) и ферментированная S. latissima с использованием ФЖЛ и/или Lactiplantibacillus plantarum (Lp) или без стартовой культуры (Без-СК), хранившаяся при 10 или 21°C в течение 9 дней (верхний график). Эти колонии были подвергнуты идентификации на основе оценок MALDI-TOF (нижний график). Аббревиатуры: YPD, агар на основе дрожжевого экстракта, пептона и декстрозы; LB, агар Луриа–Бертани; MRS, агар de Man, Rogosa и Sharpe.
На 12 день процесса ферментации (т.е. инокуляции СК свежей ламинарии) образцы были осушены, затем повторно запечатаны под вакуумом и перенесены из соответствующих температур хранения (10 или 21°C) на охлажденную температуру (4°C, Рисунок 1) для оценки их микробиологической стабильности. Бактериальную нагрузку образцов анализировали до 49 дня эксперимента. ОКАБ отражает общую бактериальную нагрузку, включая МКБ (поскольку большинство МКБ являются факультативными анаэробами), дрожжи и плесени. Основное влияние обработки образцов на ОКАБ было значительным на протяжении периода отбора проб (ART ANOVA, F4, 10 = 3.54, p = 0.048), хотя попарные сравнения показали значительные различия только между образцами Без-СК и ФЖЛ + Lp (ART contrast, p = 0.048). ОКАБ были относительно высокими [в диапазоне от 7.3 до 7.7 log (КОЕ г−1) на 19 день] в ферментированных образцах, инокулированных СК (ФЖЛ и/или Lp), и стабильными на протяжении всего периода охлажденного хранения (Рисунок 6). Образцы Без-СК имели более низкие исходные показатели [5.9 ± 1.6 log (КОЕ г−1)] до 49 дня, когда показатели стали сравнимы с другими группами обработки. Во всех образцах микробная флора была доминирована МКБ, которые следовали той же закономерности, что и ОКАБ, на протяжении охлажденного хранения во всех группах образцов. Также наблюдалось значительное влияние обработки на количество дрожжей и плесеней (ART ANOVA, F4, 10 = 4.78, p = 0.020). Уровни дрожжей и плесеней варьировали от 2.7 ± 0.9 log (КОЕ г−1), измеренных в образцах, инокулированных Lp, до 4.4 ± 0.2 log (КОЕ г−1) в образцах ФЖЛ, ферментированных при 21°C. Стабильно более высокие показатели дрожжей и плесеней были получены из образцов, инокулированных ФЖЛ (ферментированных при 10°C и 21°C), во время хранения по сравнению с другими группами.
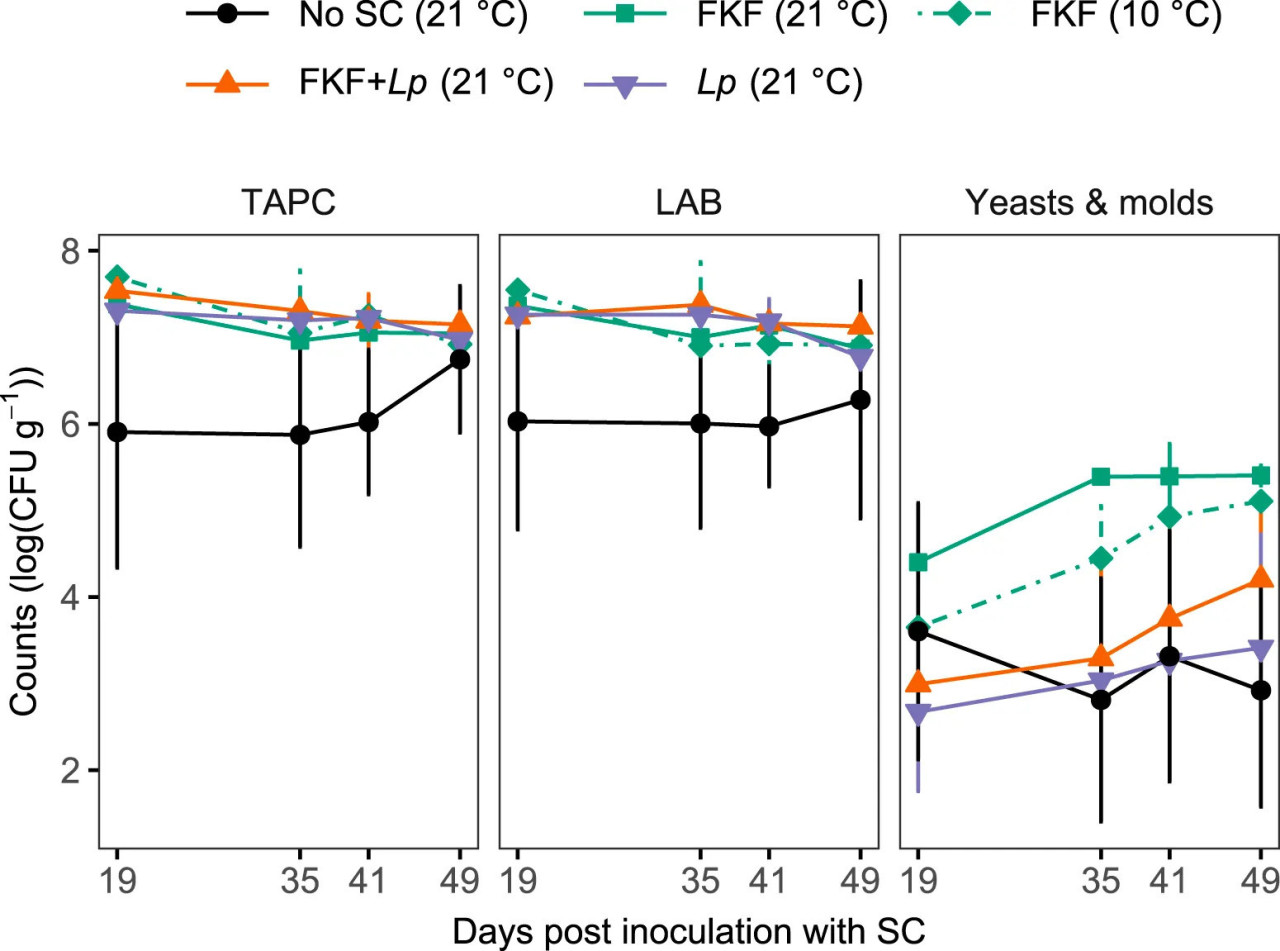
РИСУНОК 6 Количества колоний [log (КОЕ г−1)] общих аэробных бактерий (ОКАБ), молочнокислых бактерий (МКБ) и дрожжей и плесеней во время охлажденного (4°C) хранения ферментированных образцов S. latissima после добавления различных СК. Охлаждение образцов началось на 12 день после инокуляции образцов ламинарии СК. До 12 дня образцы хранились либо при 10°C, либо при 21°C. Значения представлены как среднее ± ст. откл. (n = 3).
Аэробные спорообразователи иногда были выше предела обнаружения 2 log (КОЕ г−1) в ферментированной S. latissima. На 19 день аэробные спорообразователи были обнаружены во всех трех повторностях образцов Без-СК, двух из трех повторностей образцов ФЖЛ + Lp [2.2 ± 0.4 log (КОЕ г−1)] и в 1 из 3 повторностей в остальных обработках (Рисунок 7). Аэробные спорообразователи были обнаружены на 35 день в образцах, инокулированных ФЖЛ + Lp и Lp, и на 49 день в образцах Без-СК и Lp. Анаэробные спорообразователи были обнаружены выше 2 log (КОЕ г−1) в трех случаях (данные не показаны): в одной из трех повторностей образцов ФЖЛ + Lp на 35 день [чашка переросла, оценено выше 4 log (КОЕ г−1)], в двух из трех повторностей образцов ламинарии Без-СК, из которых одна параллель была переросшей [>4 log (КОЕ г−1)], а другая составляла 2.6 log (КОЕ г−1). Listeria monocytogenes, Vibrio spp. и колиформные бактерии не были обнаружены ни в одном образце.
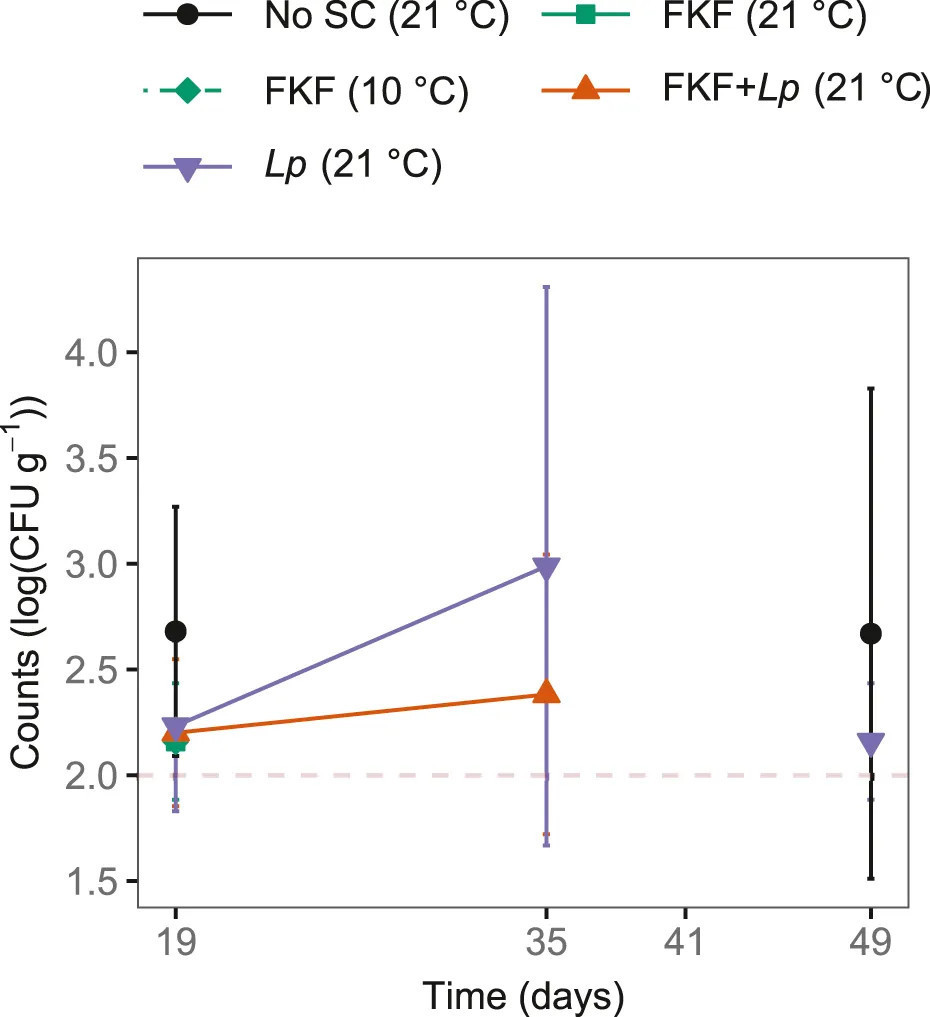
РИСУНОК 7 Количества колоний [log (КОЕ г−1)] аэробных спорообразователей во время охлажденного (4°C) хранения ферментированных образцов S. latissima после добавления различных стартовых культур (СК). Пунктирная красная линия представляет предел обнаружения 2 log (КОЕ г−1).
3.3 Экспериментальные заражения ферментированной S. latissima B. cereus
Экспериментальные заражения B. cereus были проведены на экспериментально ферментированной S. latissima с использованием ФЖЛ в качестве инокулята (хранившейся при 10°C) и контрольных образцов Без-СК (Рисунок 1) на 19 день. Измеренный pH этих образцов при инокуляции патогеном составлял 4.3 и 4.4 в двух группах образцов соответственно. Роста B. cereus не наблюдалось в обеих группах после инокуляции патогена и охлажденного хранения при 4°C в течение 30 дней. Другое экспериментальное заражение тем же и дополнительным штаммом B. cereus было проведено на коммерчески ферментированной S. latissima в момент инокуляции СК (L. sakei). Инокулированный B. cereus не рос на ферментированной ламинарии, хранившейся при 10°C и 15°C, несмотря на относительно высокие уровни pH, измеренные в этих образцах, по сравнению с пороговым значением 4.3 для роста этого пищевого патогена (Рисунок 8). Только контрольные образцы S. latissima, инокулированные L. sakei, но не B. cereus, достигли pH ниже этого порога в течение 3 дней. Во всех образцах микробная флора была доминирована МКБ. Микробная нагрузка (ОКАБ) контрольных образцов была выше, чем у ферментированной S. latissima, зараженной B. cereus, хотя значимо выше только по сравнению с зараженными образцами, хранившимися при 15°C (ART contrast, p = 0.035).
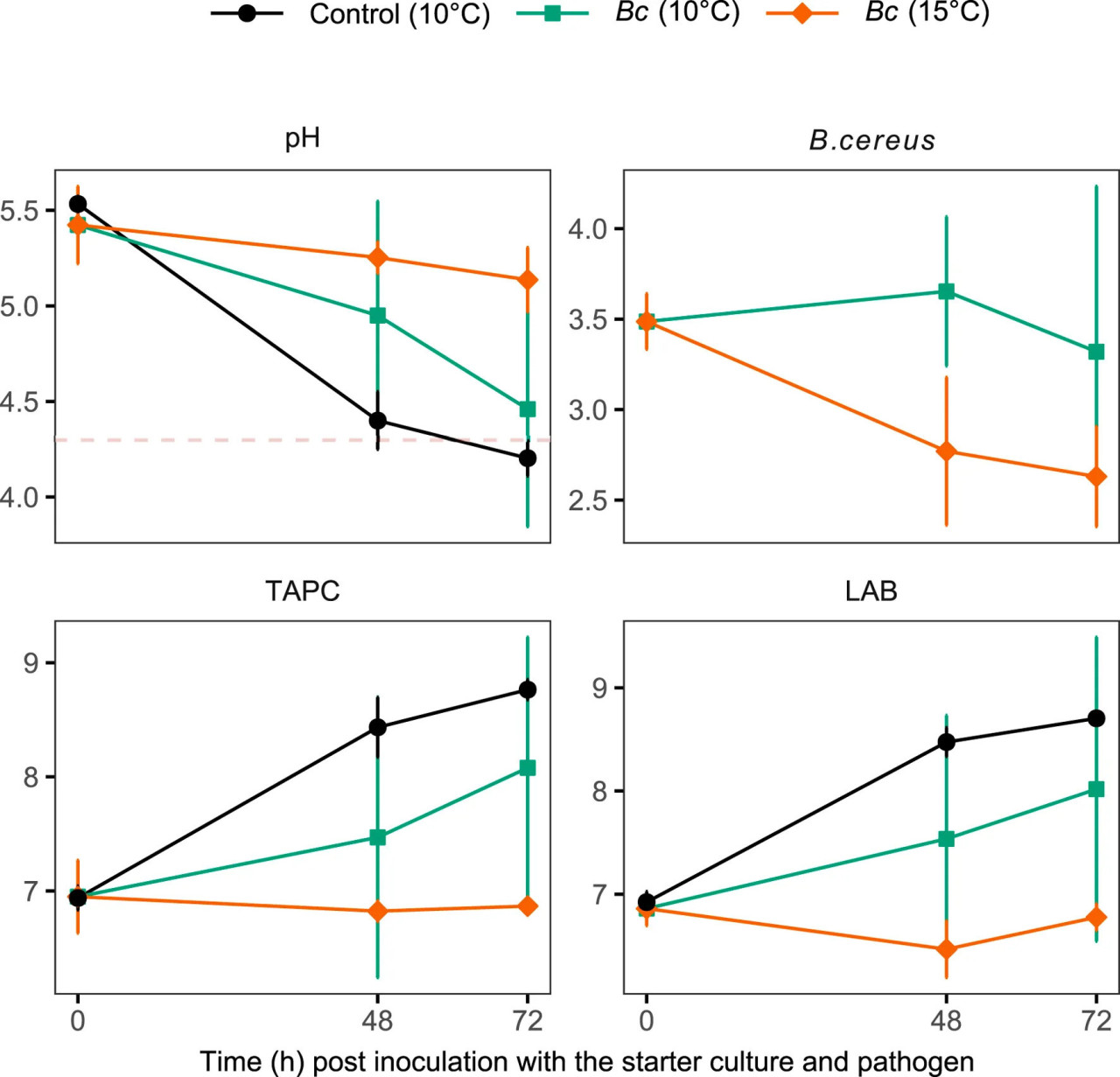
РИСУНОК 8 pH (единицы pH) и микробная нагрузка [log (КОЕ г−1)], включая Bacillus cereus, ОКАБ и МКБ, коммерчески ферментированных образцов S. latissima после добавления B. cereus (Bc) при инокуляции стартовой культурой и последующем хранении при 10°C и 15°C. Контрольные образцы не были инокулированы B. cereus. Значения представлены как среднее ± ст. откл. (n = 3). Пунктирная красная линия представляет порог безопасности pH 4.3.
4 Обсуждение
4.1 Экспериментальная ферментация S. latissima с использованием собственной биоты ламинарии
Настоящее исследование изучило использование собственной микробной флоры S. latissima в форме ФЖЛ в качестве СК для ферментации свежесобранной биомассы. pH ниже 4.3 был достигнут за 2 дня как в образцах ламинарии, инокулированных ФЖЛ, так и в образцах, получивших смесь ФЖЛ и коммерческой культуры L. plantarum, хранившихся при 21°C. Аналогичный уровень pH был достигнут через 5 дней у S. latissima, инокулированной только L. plantarum, тогда как спонтанная ферментация (Без-СК) привела к уровням pH немного выше 4.3. Кислотность ФЖЛ-СК (pH 3.6) может частично объяснить быстрое падение pH в этой группе обработки. Однако более медленное падение отчетливо наблюдалось у S. latissima, инокулированной ФЖЛ и хранившейся при 10°C, что указывает на то, что продукция кислоты после инокуляции в значительной степени отвечает за быстрое снижение pH в образцах с добавлением ФЖЛ, хранившихся при 21°C. Это было подтверждено ВЭЖХ анализом образцов, выявившим повышающиеся уровни органических кислот, в основном молочной кислоты, и самые высокие в ламинарии, инокулированной ФЖЛ и/или L. plantarum и хранившейся при 21°C. Увеличение молочной кислоты соответствовало снижению маннита в твердых образцах, предполагая, что маннит используется в качестве источника углерода ферментирующей микробиотой. Исходное содержание других ВРУ, т.е. глюкозы и фукозы, было низким и не следовало аналогичной снижающейся тенденции, предполагая, что маннит является основным источником углерода для ферментации S. latissima, что подтверждает аналогичные выводы об анаэробной ферментации ламинарии (Stévant, 2019; Krook и др., 2024; Horn and Østgaard, 2001; Hwang и др., 2011; Sandbakken и др., 2018). Незначительная продукция уксусной кислоты и этанола была обнаружена во всех образцах, хотя выше в контрольных образцах без добавленной СК. Эти продукты ферментации, вероятно, являются результатом присутствия гетероферментативных бактерий и дрожжей в этих образцах, как сообщалось ранее при спонтанной анаэробной ферментации S. latissima (Herrmann и др., 2015; Stévant, 2019; Sandbakken и др., 2018) и A. esculenta (Sørensen и др., 2021). Более медленное и менее выраженное подкисление ламинарии с добавлением ФЖЛ, хранившейся при 10°C, вероятно, связано с субоптимальной температурой для микробиоты, отобранной во время производства ФЖЛ-СК при 21°C. Было замечено подкисление S. latissima до pH 4.4 при спонтанной ферментации при 10°C с использованием того же протокола, что и в этом исследовании для приготовления ФЖЛ-СК (P. Stévant, личное наблюдение). Этот процесс может способствовать росту микробов, способных ферментировать биомассу ламинарии при низких температурах, и должен быть изучен в будущем скрининге.
Результаты этого исследования и литературные данные (Sørensen и др., 2021; Herrmann и др., 2015; Stévant, 2019; Sandbakken и др., 2018) сообщают о значительном падении pH после ферментации S. latissima без добавления СК, часто стабилизирующемся на уровнях pH между 4.0 и 4.5 после относительно длительного времени (>10 дней). Однако качество полученного ферментированного продукта в некоторой степени непредсказуемо и может позволить развитие патогенных микроорганизмов, таких как плесени (Stévant, 2019). Это было проиллюстрировано в данном исследовании появлением неприятного запаха, напоминающего рвоту, характерного для масляной кислоты, производимой Clostridium sp., в 1 из 3 повторностей во время приготовления ФЖЛ-СК (см. Раздел 2.1.1). Хотя масляная кислота не была обнаружена ни в одном из образцов (включая Без-СК) во время экспериментальной ферментации, предполагая, что рост и активность клостридий были ингибированы, спонтанная ферментация не является надежным методом для крупномасштабного сохранения биомассы ламинарии для пищевых применений. Это может быть связано с вариабельностью содержания сбраживаемых сахаров в разных партиях S. latissima, высокой буферной емкостью и низким количеством МКБ, исходно присутствующих на биомассе. Оптимальные СК будут способствовать быстрому подкислению ламинарии и конкурировать с ростом потенциально присутствующих патогенов. Быстрое подкисление ферментированных продуктов (например, кимчи и квашеной капусты) имеет ключевое значение для ингибирования роста патогенов на ранней стадии процесса и создания селективной среды, благоприятствующей кислотоустойчивым МКБ, которые будут доминировать в ферментации и предотвращать дальнейшую порчу неизвестными микроорганизмами (Jung и др., 2011; Zabat и др., 2018). Быстрое снижение pH также может предотвратить ферментативную и микробную деградацию, потенциально приводящую к неприятным привкусам и измененному nutritional content (например, витаминов и антиоксидантов).
Предварительная характеристика образцов с помощью MALDI-TOF MS показала, что ФЖЛ-СК, использованная для ферментации свежесобранной S. latissima, была доминирована МКБ, идентифицированными как L. plantarum. Это противоречит микробной флоре свежей неферментированной S. latissima (того же происхождения, что и биомасса, использованная для производства ФЖЛ-СК, но собранной на несколько недель позже), которая была доминирована морскими бактериями и некоторыми дрожжами, тогда как МКБ обнаружены не были. МКБ могут естественным образом присутствовать на свежесобранной S. latissima, но в слишком малом количестве для обнаружения. Эти бактерии могут конкурировать с другими микроорганизмами и расти после наступления анаэробных условий (как в ФЖЛ-СК). Однако L. plantarum является одним из наиболее распространенных видов МКБ, и его доминирование в образцах ФЖЛ-СК также может возникать из-за загрязнения при сборе урожая и обработке биомассы на коммерческом производственном объекте. Напротив, МКБ не были идентифицированы в образцах спонтанно ферментированного контроля (Без-СК). Среди бактериальных изолятов, идентифицированных с надежной идентификацией рода (значение оценки >2.0), в этих образцах были обнаружены оппортунистические бациллы родов Pseudomonas и Lelliotia. Это отличается от микробной флоры спонтанно ферментированной S. latissima, о которой сообщили Sørensen и др. (2021), которая состояла в основном из Psychromonas, Marinomonas и Aliivibrio.
Протокол, использованный для подготовки образцов, включал лизизис и последующий анализ лизатов, что обеспечивает более надежный и последовательный метод идентификации бактерий по сравнению с использованием целых клеток с прямым нанесением бактериальных колоний с твердых питательных сред (Böhme и др., 2010; Emami и др., 2012). Значительная часть микробиоты образцов, особенно морские бактерии, не могла быть идентифицирована с помощью MALDI-TOF MS из-за отсутствия информации о морской микробиоте в базе данных. В частности, низкие оценки, полученные для этой фракции изолированных колоний, могли дать представление только на уровне рода. Однако некоторые роды, такие как Aeromonas, демонстрируют высокое сходство спектральных паттернов, что ограничивает надежную идентификацию среди возможных видов-кандидатов. Более того, принцип MALDI-TOF/спектрального биотинга создает ограничения при использовании неподходящих эталонных баз данных, например, микроорганизмов наземного происхождения, и это описанный случай для морских ассоциированных бактерий (Emami и др., 2012). Хотя ампликонное секвенирование 16s рРНК было предпринято для характеристики микробиоты, анализ не мог быть завершен из-за непредвиденного неправильного обращения с образцами во время транспортировки. Идентификация микробиоты из ферментированной ламинарии с использованием методов высокопроизводительного секвенирования необходима в будущих исследованиях.
4.2 Микробиология и пищевая безопасность ферментированной S. latissima
Ферментированные продукты обычно содержат от пяти до семи log МКБ на мл или г, тогда как ферментированные молочные продукты могут содержать до девяти log (КОЕ г−1) (Rezac и др., 2018). Бактериальная нагрузка как экспериментально, так и коммерчески ферментированной S. latissima в настоящем исследовании, доминируемая МКБ, находилась в этом диапазоне. Суммарные показатели для дрожжей и плесеней в экспериментально ферментированных образцах были ниже (в случае образцов Без-СК, ФЖЛ + Lp и Lp) или в нижнем диапазоне (образцы, инокулированные ФЖЛ, хранившиеся при 10°C и 21°C) показателей дрожжей для типичных ферментированных продуктов [от пяти до восьми log (КОЕ г-1)] (Rezac и др., 2018). Ни L. monocytogenes, ни Vibrio spp., ни колиформные бактерии не были обнаружены ни в одном из этих образцов.
Пищевые патогены, включая Bacillus spp., L. monocytogenes, E. coli и Salmonella spp., были обнаружены на свежих или обезвоженных и готовых к употреблению съедобных макроводорослях, включая виды ламинарий (Blikra и др., 2018; Lytou и др., 2021; Martelli и др., 2021; Gupta и др., 2010; Barberi и др., 2020). Эти микроорганизмы могут присутствовать в морской среде, в которой выращиваются и собираются макроводоросли, или могут возникать из-за загрязнения во время послеуборочной обработки. Хотя в большинстве случаев эти микробные патогены обнаруживались на низких уровнях по сравнению с инфекционными дозами, необходимо принять меры для предотвращения их возникновения и роста во время сбора урожая, переработки и хранения макроводорослей, предназначенных для пищевых применений. Среди микробных патогенов токсинпродуцирующие спорообразующие бактерии (Bacillus spp.) представляют особый интерес, поскольку их споры могут выдерживать высокие температуры (>100°C) и низкий pH (<3.0) в течение коротких периодов. Споры, присутствующие в продукте, могут прорастать в жизнеспособных условиях, размножаться и в конечном итоге продуцировать токсины, что приводит к пищевым отравлениям. Концентрация B. cereus и других патогенных видов Bacillus, необходимая для производства токсинов в уровнях, способных вызвать пищевое отравление, обычно считается превышающей log 5 КОЕ г−1 (Kramer и др., 1989; Granum и др., 2000). Когда макроводоросли собирают в местах с удовлетворительным качеством воды, их концентрации патогенных Bacillus spp. предположительно низки по сравнению с порогом инфекционной дозы (Løvdal и др., 2021). Однако важно внедрить меры для ограничения их роста во время обработки и хранения. В настоящем исследовании начальная инокулированная концентрация B. cereus примерно log 3.5 КОЕ г−1 не повысилась в течение 72 часов при 10 или 15°C (Рисунок 8). Это предполагает, что эффективная ферментация и достаточное снижение pH действительно могут быть достигнуты при температурах до 10°C, и что комбинация низкого pH и конкурирующей микробиоты эффективно ингибирует рост B. cereus в этих условиях. Доступные руководства и законодательство, касающиеся съедобных макроводорослей, не предназначены специально для ферментированных продуктов, а скорее для сушеных и свежих продуктов (CEVA, 2019). Основываясь на известных пределах роста пищевых патогенов в зависимости от температуры, pH, активности воды и водно-фазового NaCl, Løvdal и др. (2021) рекомендовали pH ниже 4.3, при котором большинство патогенов, имеющих значение для макроводорослей, включая Listeria spp, Bacillus spp., Clostridium spp. и Vibrio spp., инактивируются при охлажденной температуре (4°C). Если продукт должен храниться при комнатной температуре, pH должен быть снижен ниже 3.7, чтобы предотвратить рост Salmonella spp., S. aureus и патогенной E. coli. В настоящее время не выпущено никаких руководств относительно того, насколько быстро должен быть снижен pH ферментированной ламинарии. Снижение pH ниже порога в течение 48 часов в настоящее время считается подходящей целью для успешной ферментации S. latissima производителями ламинарии. Однако, основываясь на настоящих выводах, могут быть основания для увеличения этого временного лимита до 72 часов, когда ферментация проводится при низких температурах. Однако для дальнейших рекомендаций необходимы дополнительные исследования и оценки риска.
Экспериментально ферментированные образцы S. latissima, показывающие самые высокие уровни pH — образцы, инокулированные ФЖЛ и хранившиеся при 10°C, и контрольные образцы Без-СК — были заражены B. cereus на 19 день перед хранением в анаэробных условиях при 4°C. Несмотря на уровни pH, близкие к порогу, но не ниже его в обеих группах (pH 4.3 и 4.4 соответственно), роста инокулированного патогена не наблюдалось. В этом исследовании заражения патоген инокулировали на поздней стадии процесса ферментации, когда pH был значительно ниже исходных уровней, измеренных в свежем сырье (pH 7.4), что имитирует перекрестное загрязнение ферментированной ламинарии во время переработки. Следует отметить, что как анаэробные, так и аэробные термоустойчивые спорообразователи спорадически обнаруживались во всех группах образцов экспериментально ферментированной S. latissima. Хотя обнаруженные колонии не были дополнительно идентифицированы, эти наблюдения требуют получения дополнительных данных о присутствии и поведении патогенных спорообразователей в ферментированной ламинарии. Другой тест на заражение с использованием того же штамма B. cereus и другого, ответственного за пищевые отравления, был проведен на коммерческих образцах ламинарии в начале процесса ферментации, при инокуляции СК (L. sakei). Роста патогена не наблюдалось в зараженных образцах, хранившихся как при 10°C, так и при 15°C через 72 часа, несмотря на уровни pH выше порога (pH 4.5 и 5.1 в двух температурных группах соответственно). Это может быть объяснено конкуренцией с СК, которая предотвратила рост B. cereus. Более того, заражение S. latissima B. cereus задерживало снижение pH по сравнению с контрольными образцами, которые не подвергались воздействию патогена и достигли pH 4.2 менее чем за 72 часа. Эти результаты предполагают, что подкисление при ферментации S. latissima и конкуренция с другими микроорганизмами (скорее всего, L. sakei) предотвратили рост этих штаммов B. cereus, ответственных за пищевые отравления. Тесты на заражение следует повторить, используя широкий спектр патогенных штаммов Bacillus spp. наряду с другими микробными патогенами, чтобы сделать выводы о пищевой безопасности ферментированной S. latissima.
Успешная ферментация пищевых продуктов продлевает срок годности продукта и улучшает безопасность пищевых продуктов, предотвращая микробную порчу. В случае S. latissima процесс снижает содержание йода (Krook и др., 2024; Bruhn и др., 2019), которое в противном случае может быть чрезмерным и ограничивать использование этого вида в крупномасштабных пищевых целях. Контролируемая ферментация ламинарии также может дать дополнительные преимущества, включая повышенные уровни пробиотических бактерий в конечном продукте (Gupta and Abu-Ghannam, 2012), пребиотическую функцию за счет деградации крупных неперевариваемых полисахаридов (например, альгинатов) (O’Sullivan и др., 2010) и развитие желаемых вкусовых характеристик (Bruhn и др., 2019). Согласно недавним исследованиям, ферментация ламинарии не оказывает или почти не оказывает влияния на содержание сухого вещества, содержание минералов и сырого белка (Bruhn и др., 2019; Larsen и др., 2021). Однако может происходить деградация белка (Larsen и др., 2021). Будущие исследования должны изучить влияние процессов ферментации на nutritional quality ламинарии.
5 Заключение и будущие исследования
Быстрое подкисление S. latissima ниже 4.3 после инокуляции ФЖЛ-СК подчеркивает потенциал микробиоты спонтанно ферментированной ламинарии в качестве СК для низкоэнергетического сохранения культивируемой биомассы. ФЖЛ-СК была доминирована МКБ, идентифицированными как L. plantarum, хотя из этого эксперимента было неясно, был ли этот штамм МКБ собственным или произошел от потенциального загрязнения после коммерческого сбора урожая. При хранении при 21°C микробиота образцов S. latissima с добавлением ФЖЛ использовала маннит в качестве основного источника углерода для производства молочной кислоты в качестве основного продукта ферментации, что приводило к быстрому подкислению биомассы. Скрининг и выделение релевантных СК для коммерческой ферментации ламинарии следует продолжать с использованием методов, основанных на секвенировании. Успешная и прогнозируемая ферментация биоматериала зависит от микроорганизмов с известными требованиями и характеристиками. Одной из таких характеристик является использование углеводов. Хотя некоторые штаммы могут ферментировать только простые сахара, другие могут метаболизировать более сложные углеводы. Микробиота из спонтанно ферментированной ламинарии в различных условиях в контролируемой среде, а также из других морских источников, таких как пищеварительный тракт травоядных, питающихся ламинарией, и естественно разлагающейся ламинарии, может быть изучена в качестве источников СК для ферментации ламинарии в будущих исследованиях. Будущие исследования должны быть направлены не только на поиск и улучшение ферментации с помощью природных изолятов L. plantarum, но и на использование адаптивной лабораторной эволюции (Mavrommati и др., 2022) для дальнейшего улучшения изолятов L. plantarum из ФЖЛ для ферментации ламинарии.
Предварительные результаты экспериментальных исследований ферментации S. latissima с помощью токсинпродуцирующих спорообразующих бактерий B. cereus показали отсутствие роста инокулированного патогена. Это наблюдалось даже при уровнях pH выше порога 4.3, который, как известно, ингибирует рост и развитие этого вида. Повторные испытания на заражение с использованием нескольких штаммов и микробных видов необходимы в будущих исследованиях. Адекватные СК, которые будут конкурировать с любыми потенциально присутствующими патогенными микроорганизмами в низком количестве для дальнейшего развития, в сочетании со строгими гигиеническими мерами на этапах сбора урожая и переработки, обеспечат пищевую безопасность ферментированной ламинарии. В дополнение к безопасности пищевых продуктов, будущие исследования должны изучить nutritional и сенсорный профиль, а также биоактивный (например, пребиотический) потенциал и оценить усвояемость ферментированных продуктов из ламинарии. Ферментация, индуцированная МКБ, является энергоэффективной альтернативой обычным методам стабилизации ламинарий (например, сушке на воздухе, замораживанию) (Thomas и др., 2020) в соответствии с текущими целями по улучшению устойчивости систем производства продуктов питания. Спрос на устойчивые и здоровые пищевые продукты продолжает расти, и ферментация ламинарии открывает многообещающий путь для низкоэнергетической стабилизации биомассы ламинарии.
Ссылки
1. Barberi O. N., Byron C. J., Burkholder K. M., Gelais A. T. St., Williams A. K. (2020). Assessment of bacterial pathogens on edible macroalgae in coastal waters. J. Appl. Phycol. 32 (1), 683–696. doi: 10.1007/s10811-019-01993-5. CrossRef. Google Scholar.
2. Bates D., Mächler M., Bolker B., Walker S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67 (1), 1–48. doi: 10.18637/jss.v067.i01. CrossRef. Google Scholar.
3. Blikra M. J., Altintzoglou T., Løvdal T., Rognså G., Skipnes D., Skåra T. et al. (2021). Seaweed products for the future: using current tools to develop a sustainable food industry. Trends Food Sci. Technol. 118, 765–776. doi: 10.1016/j.tifs.2021.11.002. CrossRef. Google Scholar.
4. Blikra M. J., Løvdal T., Vaka M. R., Roiha I. S., Lunestad B. T., Lindseth C. et al. (2018). Assessment of food quality and microbial safety of brown macroalgae (Alaria esculenta and Saccharina latissima). J. Sci. Food Agric. 99 (3), 1198–1206. doi: 10.1002/jsfa.9289. CrossRef. Google Scholar.
5. Böhme K., Fernández-No I. C., Barros-Velázquez J., Gallardo J. M., Cañas B., Calo-Mata P. (2010). Comparative analysis of protein extraction methods for the identification of seafood-borne pathogenic and spoilage bacteria by MALDI-TOF mass spectrometry. Anal. Methods-UK 2 (12), 1941–1947. doi: 10.1039/c0ay00457j. CrossRef. Google Scholar.
6. Bruhn A., Brynning G., Johansen A., Lindegaard M. S., Sveigaard H. H., Aarup B. et al. (2019). Fermentation of sugar kelp (Saccharina latissima) — effects on sensory properties, and content of minerals and metals. J. Appl. Phycol. 31, 3175–3187. doi: 10.1007/s10811-019-01827-4. CrossRef. Google Scholar.
7. Cabrita A. R. J., Maia M. R. G., Sousa-Pinto I., Fonseca A. J. M. (2017). Ensilage of seaweeds from an integrated multi-trophic aquaculture system. Algal Res. 24 (Part A), 290–298. doi: 10.1016/j.algal.2017.04.024. CrossRef. Google Scholar.
8. CEVA (2019). Réglementation algues alimentaires. Pleubian, France: Centre d'Étude et de Valorisation des Algues. Google Scholar.
9. Ehling-Schulz M., Svensson B., Guinebretiere M.-H., Lindbäck T., Andersson M., Schulz A. et al. (2005). Emetic toxin formation of Bacillus cereus is restricted to a single evolutionary lineage of closely related strains. Microbiology 151 (1), 183–197. doi: 10.1099/mic.0.27607-0. CrossRef. Google Scholar.
10. Emami K., Askari V., Ullrich M., Mohinudeen K., Anil A. C., Khandeparker L. et al. (2012). Characterization of bacteria in ballast water using MALDI-TOF mass spectrometry. Plos One 7 (6), e38515. doi: 10.1371/journal.pone.0038515. CrossRef. Google Scholar.
11. Emblemsvåg J., Kvadsheim N. P., Halfdanarson J., Koesling M., Nystrand B. T., Sunde J. et al. (2020). Strategic considerations for establishing a large-scale seaweed industry based on fish feed application: a Norwegian case study. J. Appl. Phycol. 32 (6), 4159–4169. doi: 10.1007/s10811-020-02234-w. CrossRef. Google Scholar.
12. Granum P. E., Braid-Parker T. (2000). "Bacillus species," in The microbiological safety and quality of food. 2. Editors Lund B., Braid-Parker T., Gould W. (Gaithersburg, MD, USA: Aspen Publishers), 1029–1039. Google Scholar.
13. Gupta S., Abu-Ghannam N. (2012). Probiotic fermentation of plant based products: possibilities and opportunities. Crit. Rev. Food Sci. Nutr. 52 (2), 183–199. doi: 10.1080/10408398.2010.499779. CrossRef. Google Scholar.
14. Gupta S., Rajauria G., Abu-Ghannam N. (2010). Study of the microbial diversity and antimicrobial properties of Irish edible brown seaweeds. Int. J. Food Sci. Tech. 45 (3), 482–489. doi: 10.1111/j.1365-2621.2009.02149.x. CrossRef. Google Scholar.
15. Herrmann C., FitzGerald J., O'Shea R., Xia A., O'Kiely P., Murphy J. D. (2015). Ensiling of seaweed for a seaweed biofuel industry. Bioresour. Technol. 196, 301–313. doi: 10.1016/j.biortech.2015.07.098. CrossRef. Google Scholar.
16. Horn S. J., Østgaard K. (2001). Alginate lyase activity and acidogenesis during fermentation of Laminaria hyperborea. J. Appl. Phycol. 13 (2), 143–152. doi: 10.1023/A:1011187526918. CrossRef. Google Scholar.
17. Hwang H., Lee S., Kim S., Lee S. (2011). Fermentation of seaweed sugars by Lactobacillus species and the potential of seaweed as a biomass feedstock. Bioprocess Eng. 16 (6), 1231–1239. doi: 10.1007/s12257-011-0278-1. CrossRef. Google Scholar.
18. Jung J. Y., Lee S. H., Kim J. M., Park M. S., Bae J. W., Hahn Y. et al. (2011). Metagenomic analysis of kimchi, a traditional Korean fermented food. Appl. Environ. Microbiol. 77 (7), 2264–2274. doi: 10.1128/AEM.02157-10. CrossRef. Google Scholar.
19. Kay M., Elkin L., Higgins J., Wobbrock J. (2021). ARTool: aligned rank transform for nonparametric factorial ANOVAs. Available at: https://github.com/mjskay/ARTool. Google Scholar.
20. Kramer J., Gilbert R. (1989). "Bacillus cereus and other Bacillus species," in Food borne bacterial pathogens. Editor Doyle M. (New York: Marcel Dekker Inc), 21–70. Google Scholar.
21. Krook J. L., Riboldi L., Birkeland I. M., Stévant P., Larsen W. E., Rhein-Knudsen N. et al. (2024). Acid preservation of the brown seaweed Saccharina latissima for food applications. Algal Res. 80, 103524. doi: 10.1016/j.algal.2024.103524. CrossRef. Google Scholar.
22. Larsen S. U., Ma N., Hou X., Bruhn A., Boderskov T., MacLeod A. et al. (2021). Ensiling of sugar kelp biomass for biorefining. Biomass Bioenergy 151, 106134. doi: 10.1016/j.biombioe.2021.106134. CrossRef. Google Scholar.
23. Lenth R. V. (2016). Least-squares means: the R package lsmeans. J. Stat. Softw. 69 (1), 1–33. doi: 10.18637/jss.v069.i01. CrossRef. Google Scholar.
24. López-Pedemonte T. J., Roig-Sagués A. X., Trujillo A. J., Capellas M., Guamis B. (2003). Inactivation of spores of Bacillus cereus in cheese by high hydrostatic pressure with the addition of nisin or lysozyme. J. Dairy Sci. 86 (10), 3075–3081. doi: 10.3168/jds.S0022-0302(03)73907-1. CrossRef. Google Scholar.
25. Løvdal T., Lunestad B. T., Myrmel M., Rosnes J. T., Skipnes D. (2021). Microbiological food safety of seaweeds. Foods 10 (11), 2719. doi: 10.3390/foods10112719. CrossRef. Google Scholar.
26. Lytou A. E., Schoina E., Liu Y., Michalek K., Stanley M. S., Panagou E. Z. et al. (2021). Quality and safety assessment of edible seaweeds Alaria esculenta and Saccharina latissima cultivated in Scotland. Foods 10 (9), 2210. doi: 10.3390/foods10092210. CrossRef. Google Scholar.
27. Martelli F., Marrella M., Lazzi C., Neviani E., Bernini V. (2021). Microbiological contamination of ready-to-eat algae and evaluation of Bacillus cereus behavior by microbiological challenge test. J. Food Prot. 84 (7), 1275–1280. doi: 10.4315/JFP-20-407. CrossRef. Google Scholar.
28. Mavrommati M., Daskalaki A., Papanikolaou S., Aggelis G. (2022). Adaptive laboratory evolution principles and applications in industrial biotechnology. Biotechnol. Adv. 54, 107795. doi: 10.1016/j.biotechadv.2021.107795. CrossRef. Google Scholar.
29. NMKL:136 (2010). Listeria monocytogenes. Detection in foods and feeding stuffs and enumeration in foods. Bergen, Norway: Nordisk Metodikkomité for Næringsmidler. Google Scholar.
30. NMKL:140 (2023). Lactic acid bacteria. Determination in food in association with food spoilage. Bergen, Norway: Nordisk Metodikkomité for Næringsmidler. Google Scholar.
31. NMKL:156 (1997). Pathogenic Vibrio species. Detection and enumeration in foods. Bergen, Norway: Nordisk Metodikkomité for Næringsmidler. Google Scholar.
32. NMKL:189 (2011). Aerobic or anaerobic microorganisms or bacterial spores. Enumeration on Blood Agar. Bergen, Norway: Nordisk Metodikkomité for Næringsmidler. Google Scholar.
33. NMKL:67 (2021). Presumptive Bacillus cereus. Determination in foods. Bergen, Norway: Nordisk Metodikkomité for Næringsmidler. Google Scholar.
34. NMKL:98 (2005). Mould and yeasts. Determination in foods and feed. Bergen, Norway: Nordisk Metodikkomité for Næringsmidler. Google Scholar.
35. O'Sullivan L., Murphy B., McLoughlin P., Duggan P., Lawlor P. G., Hughes H. et al. (2010). Prebiotics from marine macroalgae for human and animal health applications. Mar. Drugs 8 (7), 2038–2064. doi: 10.3390/md8072038. CrossRef. Google Scholar.
36. Philis G., Gracey E. O., Gansel L. C., Fet A. M., Rebours C. (2018). Comparing the primary energy and phosphorus consumption of soybean and seaweed-based aquafeed proteins – a material and substance flow analysis. J. Clean. Prod. 200, 1142–1153. doi: 10.1016/j.jclepro.2018.07.247. CrossRef. Google Scholar.
37. R Development Core Team (2022). R: a language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. Available at: https://www.R-project.org/. Google Scholar.
38. Rezac S., Kok C. R., Heermann M., Hutkins R. (2018). Fermented foods as a dietary source of live organisms. Front. Microbiol. 9, 1785. doi: 10.3389/fmicb.2018.01785. CrossRef. Google Scholar.
39. Rotter A., Bacu A., Barbier M., Bertoni F., Bones A. M., Cancela M. L. et al. (2020). A new network for the advancement of marine biotechnology in Europe and beyond. Front. Mar. Sci. 7 (278). doi: 10.3389/fmars.2020.00278. CrossRef. Google Scholar.
40. Sandbakken I. S., Sæther M., Funderud J., Aasen I. M. (2018). Acid preservation of Saccharina latissima for application as a carbon source for fermentation to biofuels and chemicals. J. Appl. Phycol. 30 (6), 3581–3588. doi: 10.1007/s10811-018-1489-z. CrossRef. Google Scholar.
41. Sedó Molina G. E., Shetty R., Xiao H., Wätjen A. P., Tovar M., Bang-Berthelsen C. H. (2022). Development of a novel lactic acid bacteria starter culture approach: from insect microbiome to plant-based fermentations. LWT-Food Sci. Technol. 167, 113797. doi: 10.1016/j.lwt.2022.113797. CrossRef. Google Scholar.
42. Sørensen J. S., Madsen S. K., Bang-Berthelsen C. H., Hansen L. T. (2021). Quality and safety aspects in fermentation of winged kelp (Alaria esculenta) and sugar kelp (Saccharina latissima) by the natural microbiota with or without addition of a Lactiplantibacillus plantarum starter culture. Food Res. Int. 150, 110800. doi: 10.1016/j.foodres.2021.110800. CrossRef. Google Scholar.
43. Stévant P. (2019). Seaweeds in food applications: effects of processing on product quality. Ph.D thesis. Trondheim, Norway: Norwegian University of Science and Technology NTNU. Available at: https://ntnuopen.ntnu.no/ntnu-xmlui/handle/11250/2606704. Google Scholar.
44. Stévant P., Rebours C. (2021). Landing facilities for processing of cultivated seaweed biomass: a Norwegian perspective with strategic considerations for the European seaweed industry. J. Appl. Phycol. 33, 3199–3214. doi: 10.1007/s10811-021-02525-w. CrossRef. Google Scholar.
45. Stévant P., Rebours C., Chapman A. (2017). Seaweed aquaculture in Norway: recent industrial developments and future perspectives. Aquacult Int. 25 (4), 1373–1390. doi: 10.1007/s10499-017-0120-7. CrossRef. Google Scholar.
46. Thomas J.-B. E., Sodré Ribeiro M., Potting J., Cervin G., Nylund G. M., Olsson J. et al. (2020). A comparative environmental life cycle assessment of hatchery, cultivation, and preservation of the kelp Saccharina latissima. ICES J. Mar. Sci. 78 (1), 451–467. doi: 10.1093/icesjms/fsaa112. CrossRef. Google Scholar.
47. Uchida M., Amakasu H., Satoh Y., Murata M. (2004). Combinations of lactic acid bacteria and yeast suitable for preparation of marine silage. Fish. Sci. 70, 507–517. doi: 10.1111/j.1444-2906.2004.00832.x. CrossRef. Google Scholar.
48. Uchida M., Miyoshi T. (2013). Algal Fermentation — The seed for a new fermentation industry of foods and related products. JARQ-Jpn Agr Res. 47 (1), 53–63. doi: 10.6090/jarq.47.53. CrossRef. Google Scholar.
49. Wirenfeldt C. B., Sørensen J. S., Kreissig K. J., Hyldig G., Holdt S. L., Hansen L. T. (2022). Post-harvest quality changes and shelf-life determination of washed and blanched sugar kelp (Saccharina latissima). Front. Food Sci. Technol. 2, 1030229. doi: 10.3389/frfst.2022.1030229. CrossRef. Google Scholar.
50. Zabat M. A., Sano W. H., Wurster J. I., Cabral D. J., Belenky P. (2018). Microbial community analysis of sauerkraut fermentation reveals a stable and rapidly established community. Foods Basel, Switz. 7 (5), 77. doi: 10.3390/foods7050077. CrossRef. Google Scholar.
Stévant P, Zioga E, Steenholdt Sørensen J, Heiner Bang-Berthelsen C, Dahl TH, Barnung T, Kleppe M and Løvdal T (2025) Fermentation of sugar kelp (Saccharina latissima): exploring the potential of the kelp’s native microbiota as starter culture and the microbiological food safety of fermented products. Front. Food. Sci. Technol. 4:1490547. doi: 10.3389/frfst.2024.1490547
Перевод статьи «Fermentation of sugar kelp (Saccharina latissima): exploring the potential of the kelp’s native microbiota as starter culture and the microbiological food safety of fermented products» авторов Fermentation of sugar kelp (Saccharina latissima): exploring the potential of the kelp’s native microbiota as starter culture and the microbiological food safety of fermented products., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык
Фото: wikipedia



















































































































Комментарии (0)