Холодная плазма: почему одни сальмонеллы выживают, а другие — нет
Основной целью данного исследования было оценить устойчивость клеток Salmonella Enteritidis ATCC 13076 в стационарной фазе (STAT) и фазе длительного выживания (LTS) к воздействию атмосферной холодной плазмы (ACP) в фосфатно-солевом растворе (PBS, pH 7,0) и на куриных яйцах в скорлупе.
Аннотация
Salmonella Enteritidis культивировали в триптиказо-соевом растворе с добавлением 0,6% (мас./об.) дрожжевого экстракта (35°C) в течение 20 часов (STAT) и 21 дня (LTS). Морфологию клеток определяли с помощью световой микроскопии. В PBS и на яйца в скорлупе вносили клетки STAT или LTS для достижения уровня ~7,0 log10 КОЕ/мл или на яйцо. Обработку ACP проводили при 45 кВ (PBS) и 60 кВ (яйца в скорлупе) в течение 1–4 минут и 1–5 минут соответственно. Выжившие патогены подсчитывали на тонкослойной агаризованной среде (TAL) и на агаре ксилоза-лизин-тергитол-4 (XLT-4) после 48 часов инкубации (35°C). Для выживших на яйцах в скорлупе были получены значения R2 и среднеквадратичной ошибки с использованием моделей «логарифмическая линейная с хвостом» (Log-linear with Tail) и Вейбулла. Клетки STAT были преимущественно палочковидной формы, тогда как клетки LTS имели кокковидную форму. В PBS снижение (log10 КОЕ/мл) клеток STAT составило 1,0, 0,95, 1,45 и 1,44 после воздействия ACP в течение 1, 2, 3 и 4 минут соответственно. Напротив, снижение количества клеток LTS было значительно ниже (p < 0,05): 0,04 (1 мин), 0,06 (2 мин), 0,01 (3 мин) и 0,11 (4 мин). Аналогичная закономерность наблюдалась на яйцах в скорлупе, где клетки LTS проявили гораздо более высокую устойчивость к ACP, чем клетки STAT (p < 0,05). Модель «логарифмическая линейная с хвостом» обеспечила лучшее соответствие данным по выживанию клеток STAT; время для достижения снижения на 4 и 5 log составило 5,29 и 5,78 минут соответственно. Сублетальное повреждение наблюдалось как у выживших клеток STAT, так и LTS; однако различия не были статистически значимыми (P > 0,05). Кроме того, не было обнаружено различий в прочности скорлупы и цвете желтка между яйцами, обработанными ACP, и контрольными яйцами. Основываясь на этих результатах, клетки S. Enteritidis в фазе LTS более устойчивы к ACP, чем клетки в фазе STAT, и их следует учитывать при разработке протоколов валидации процессов, включающих применение ACP для инактивации Salmonella на яйцах в скорлупе.
1 Введение
По оценкам Центров по контролю и профилактике заболеваний (CDC), Salmonella является основной причиной бактериальных пищевых заболеваний: ежегодно в США регистрируется примерно 1,35 миллиона случаев, 26 500 госпитализаций и 420 смертельных исходов. В этом отношении мясо птицы и птицеводческая продукция являются основными переносчиками этого патогена к потребителям (CDC, 2021). Salmonella Enteritidis обычно ассоциируется со вспышками заболеваний, связанных с яйцами, и чаще всего загрязняет внешнюю поверхность куриных яиц в скорлупе через помет птицы. В 2018 году Gravil Ridge Farms объявила отзыв яиц в скорлупе, загрязненных S. Enteritidis, что привело к 44 случаям заболевания и 12 госпитализациям (CDC, 2018). Два штамма S. Enteritidis в яйцах из Польши стали причиной крупной вспышки сальмонеллеза, произошедшей в 18 европейских странах (Pijnacker и др., 2019).
Во всем мире яйца являются высокопитательным основным продуктом питания в домохозяйствах, который обеспечивает организм необходимыми витаминами, аминокислотами, жирными кислотами, фолатом и другими микронутриентами (Keerthirathne и др., 2017). Согласно недавним оценкам, потребление яиц на душу населения в Соединенных Штатах в 2023 году составило 281,3 яйца, и прогнозируется, что в 2024 году оно достигнет 284,4 яйца (Statistica, 2024). Для повышения микробиологической безопасности яиц несколько стран, включая США, Канаду и Японию, применяют мойку с использованием хлора (Georgescu и др., 2017; Wan и др., 2017) для снижения микробной нагрузки на внешней поверхности яиц в скорлупе.
Хлор является наиболее широко используемым санитайзером для яиц в скорлупе, поскольку он относительно недорог; однако с его использованием связан ряд ограничений. К ним относятся его инактивация органическими веществами, реакция с органическими веществами с образованием вредных хлорорганических побочных продуктов (Goslan и др., 2009; Ölmez и Kretzschmar, 2009), образование точечной коррозии на оборудовании из нержавеющей стали при более низком pH (<6,5) и снижение противомикробной эффективности при более высоком pH (Nou и Luo, 2010; Tirpanalan и др., 2011). Эти ограничения подогрели интерес пищевой промышленности к поиску альтернатив использованию хлора в качестве санитайзера (Meireles и др., 2016). Более того, негативное восприятие потребителями синтетических химических санитайзеров (Buchmüller и др., 2020) также способствовало интересу пищевой промышленности к альтернативным противомикробным обработкам. Хотя тепловая пастеризация может использоваться в качестве метода деконтаминации, она может вызывать дефекты качества, такие как коагуляция яйца и денатурация белков. Чтобы уничтожить Salmonella на яйцах в скорлупе, смягчая при эти дефекты качества, исследователи изучили некоторые нетермические методы, такие как электролизованная кислая вода, диоксид углерода под высоким давлением, радиочастотная обработка (Keerthirathne и др., 2017) и холодная плазма (Gavahian и др., 2019; Illera и др., 2022; Wan и др., 2017).
Новый нетермический метод уничтожения патогенов на внешней поверхности яиц в скорлупе включает использование атмосферной холодной плазмы (ACP). Холодная плазма представляет собой неравновесный ионизированный газ, содержащий разнообразные атомы, молекулы, ионы, свободные радикалы и другие активные формы, сосуществующие как в основном, так и в возбужденном состояниях (Bourke и др., 2017; Misra и др., 2016). Инактивация микроорганизмов ACP является результатом аддитивных и/или синергетических эффектов различных активных форм кислорода (ROS) и активных форм азота (RNS) (Deng и др., 2010; Scholtz и др., 2015). Важным преимуществом ACP является то, что инактивация микроорганизмов может быть достигнута с незначительным тепловым повреждением пищи (Niemira, 2012). Совсем недавно сообщалось об отсутствии различий в качественных характеристиках обработанных и необработанных цельных куриных яиц после инактивации Salmonella с помощью ACP более чем на 5,0 log10 КОЕ/яйцо (Illera и др., 2023; Abdoli и др., 2024). Существует несколько сообщений об инактивации кишечных патогенов на поверхности яиц в скорлупе с помощью ACP (Dasan и др., 2018; Gavahian и др., 2019; Georgescu и др., 2017; Lin и др., 2020; Moritz и др., 2021; Wan и др., 2017). Однако во всех этих исследованиях яйца были искусственно инокулированы патогенами, находящимися в стационарной фазе (STAT) бактериального жизненного цикла.
Четырьмя наиболее признанными фазами бактериального жизненного цикла являются лаг-фаза, экспоненциальная фаза, стационарная фаза и фаза отмирания. Однако существует пятая, менее известная фаза, называемая длительной стационарной фазой (Finkel, 2006), в последнее время именуемая фазой длительного выживания (LTS) (Bhullar и др., 2021; Djebbi-Simmons и др., 2019; Wang и др., 2018; Wen и др., 2009). Переход бактерий в эту фазу индуцируется факторами окружающей среды, такими как ограничение питательных веществ, обезвоживание, накопление отходов, конкуренция или другими стрессорами (Finkel, 2006). Бактерии LTS представляют собой фенотипические варианты, которые трансформируются из палочковидных клеток в коккобациллы или кокковидные клетки по мере перехода через жизненный цикл (Wang, 2017; Wen и др., 2011). Интересно, что бактерии LTS проявляют повышенную устойчивость к антибактериальным обработкам, таким как высокая температура и давление (Wen и др., 2011), ультрафиолетовое излучение (Wang и др., 2018), ампициллин (Wang, 2017) и хлор (Bhullar и др., 2021; Djebbi-Simmons и др., 2019).
В исследованиях по микробиологической безопасности пищевых продуктов распространенной практикой является использование клеток STAT кишечных патогенов при оценке антибактериальных обработок и валидации микробиологической безопасности пищевых процессов. Согласно Finkel (2006), бактерии в естественной среде большую часть своей жизни проводят в виде клеток LTS. Следовательно, сырые сельскохозяйственные продукты, предназначенные для переработки, с большей вероятностью будут загрязнены патогенами LTS из источников окружающей среды. Хотя клетки STAT более устойчивы, чем клетки экспоненциальной фазы (EXP), появляется все больше доказательств, свидетельствующих о том, что клетки LTS проявляют более высокую устойчивость к физическим или химическим противомикробным обработкам (Bhullar и др., 2021; Djebbi-Simmons и др., 2019; Wang, 2017; Wang и др., 2018; Wen и др., 2009). Хотя существует растущий объем знаний об инактивации сальмонелл на яйцах в скорлупе с помощью ACP (Georgescu и др., 2017; Wan и др., 2017), нет опубликованных сообщений об инактивации LTS Salmonella на этих яйцах с помощью ACP. Соответственно, основной целью настоящего исследования было изучить устойчивость клеток STAT и LTS S. Enteritidis к ACP в PBS и на яйцах в скорлупе. Дополнительными задачами было определить степень сублетального повреждения выживших патогенов и оценить изменения выбранных параметров качества яиц после обработки ACP.
2 Материалы и методы
2.1 Бактериальная культура и условия культивирования
Salmonella enterica серовар Enteritidis (ATCC13076) была получена из коллекции культур Лаборатории микробиологической безопасности пищевых продуктов Университета штата Айова, Эймс, Айова. Маточную культуру хранили замороженной (−80°C) в бульоне на основе мозгово-сердечной инфузии (BHI) (Difco; Becton Dickinson, Спаркс, Мэриленд), содержащем 10% (об./об.) глицерина. Замороженные культуры во флаконах оттаивали под струей холодной водопроводной воды, затем активировали путем асептического переноса 0,1 мл оттаянной культуры в 10 мл триптиказо-соевого бульона с добавлением 0,6% (мас./об.) дрожжевого экстракта (TSBYE) с последующей инкубацией (35°C) в течение 24 ч. Активированную культуру хранили при 4°C до использования для приготовления рабочих культур. Рабочие культуры готовили путем выполнения двух последовательных 20-часовых пересевов активированной культуры в TSBYE (35°C) перед началом каждого эксперимента.
2.2 Приготовление клеток STAT и LTS
Аликвоту (2,0 мл) рабочей культуры S. Enteritidis асептически переносили в каждую из двух конических колб с завинчивающейся крышкой, содержащих по 200 мл TSBYE на колбу. Колбы с инокулированным TSBYE инкубировали (35°C) в течение 21 дня для получения клеток LTS (Bhullar и др., 2021). Перед началом эксперимента две дополнительные колбы, содержащие по 200 мл TSBYE, инокулировали, как описано ранее, и инкубировали (35°C) в течение 20 ч для получения клеток STAT.
2.3 Приготовление инокулюма и инокуляция PBS
40-мл порции культур STAT или LTS в TSBYE асептически переносили в соответствующие стерильные центрифужные пробирки объемом 50 мл. Эти клетки осаждали центрифугированием (10 000 × g в течение 10 мин, 23°C) с использованием центрифуги Sorvall Super T21 (American Laboratory Trading, Inc., Ист-Лайм, Коннектикут). Осажденные клетки отмывали, ресуспендируя их в PBS (pH 7,0) с помощью вортексирования. Отмытые клетки осаждали центрифугированием, как описано ранее. Осажденные клетки STAT и LTS ресуспендировали в 40 мл и 4 мл PBS соответственно для получения примерно одинаковой концентрации жизнеспособных клеток для каждого физиологического состояния патогена. Аликвоты (150 мкл) суспензии клеток STAT или LTS (∼10⁹ КОЕ/мл) использовали для инокуляции 15-мл порций PBS (в чашках Петри) для достижения концентрации клеток ∼10⁷ КОЕ/мл. Для приготовления инокулюма для яиц в скорлупе 40-мл порции рабочих культур STAT или LTS асептически переносили в соответствующие центрифужные пробирки объемом 50 мл и осаждали центрифугированием, как описано ранее. Осажденные клетки ресуспендировали в соответствующих объемах отработанного TSBYE для получения концентрации жизнеспособных клеток ∼10⁹ КОЕ/мл.
2.4 Приготовление и инокуляция яиц в скорлупе
Свежие органические крупные коричневые яйца (66 ± 2 г) были приобретены в местной ферме в Эймсе, Айова, и использованы в течение 10 дней после покупки. Яйца проверяли на целостность скорлупы с помощью просвечивания и осторожно ополаскивали дистиллированной водой для удаления загрязнений со скорлупы. Их протирали бумажным полотенцем, смоченным 70% этанолом, с последующим погружением в 70% этанол на 1 минуту (Wan и др., 2017) для снижения микробной контаминации. Яйца высушивали на воздухе в ламинарном шкафу с включенным вентилятором в течение минимум 20 минут перед инокуляцией S. Enteritidis. Аликвоту (100 мкл) суспензии клеток S. Enteritidis наносили точечно (13–18 точек) на продольную поверхность яйца в пределах области 2 × 2 см, обозначенной с помощью маркера. Инокулированные яйца выдерживали при комнатной температуре (22 ± 1°C) в течение 2,5 ч в ламинарном шкафу (с выключенным вентилятором) для обеспечения прикрепления клеток и высыхания инокулюма.
2.5 Обработка PBS холодной плазмой
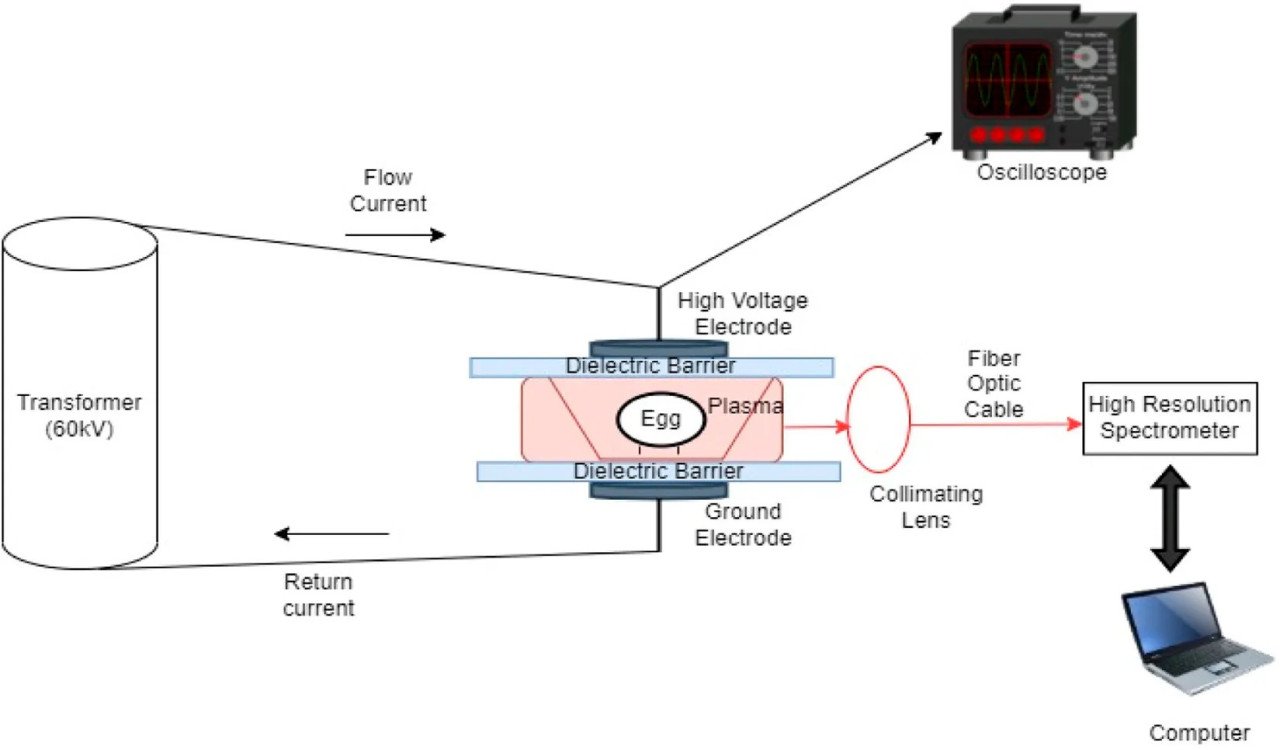
РИСУНОК 1 Схема экспериментальной установки для обработки яиц в скорлупе высоковольтной атмосферной холодной плазмой (HVACP).
Установка диэлектрического барьерного разряда (DBD) высоковольтной атмосферной холодной плазмы (HVACP), изображенная на Рисунке 1, представляет собой систему генерации плазмы при атмосферном давлении и низкой температуре, используемую для плазменной обработки внутри упаковки (Yepez и Keener, 2016). Система состоит из трансформатора (Phenix Technologies, Мэриленд, США) с входным напряжением 120 В (переменного тока) при 60 Гц, регулятора напряжения с выходным напряжением, регулируемым в пределах 0–120 кВ, двух алюминиевых электродов в форме бублика (каждый диаметром 145 мм) с межэлектродным разрядным расстоянием 55 мм. Диэлектрические барьеры состоят из слоя плексигласа и двух полипропиленовых листов Cuisinart® толщиной 1 мм, каждый размерами 355 мм × 272 мм × 2,20 мм (Cuisinart, Нью-Джерси, США). Чашку Петри (150 мм × 15 мм) без крышки, содержащую 15 мл PBS, инокулированного S. Enteritidis, помещали внутрь полипропиленовой коробки (160 × 110 × 24 мм). Крышку коробки закрывали, а затем коробку помещали внутрь полиэтиленового пакета с высоким барьером для кислорода размером 250 × 250 мм, изготовленного из пленки Cryovac® B2360 (Sealed Air North Carolina, США). Пакет продували атмосферным воздухом, затем термосваривали. Упакованную коробку с образцом располагали между двумя алюминиевыми электродами со слоем плексигласа и полипропиленовым листом непосредственно над пакетом и одним полипропиленовым листом под пакетом. Прикладывали напряжение электрического поля 45 кВ, и образцы PBS подвергали воздействию ACP в течение 0 (контроль), 1, 2, 3 и 4 минут. После обработки ACP образцы выдерживали при комнатной температуре (22°C ± 1°C) в ламинарном шкафу в течение 24 ч перед определением количества выживших патогенов. Каждый эксперимент проводили в трех повторностях.
2.6 Обработка яиц в скорлупе холодной плазмой
Ту же систему DBD HVACP, что описана ранее в Разделе 2.5, использовали для обработки искусственно инокулированных яиц в скорлупе. Каждое инокулированное яйцо помещали в стерильную полистироловую чашку для взвешивания Fisher brand (41 мм × 41 мм × 8 мм). Каждую чашку с яйцом помещали в отдельную стерильную полипропиленовую коробку Mainstays (161 мм × 85 мм × 57 мм) общим объемом 942,5 мл (Walmart Inc., Бентонвилл, Арканзас, США). Коробку (с закрытой крышкой) помещали внутрь пакета Cryovac® B2360 с высоким барьером для кислорода размером 310 × 202 мм (Sealed Air North Carolina, США), пакет продували воздухом и термосваривали. Прикладывали напряжение электрического поля 60 кВ, и яйца подвергали воздействию ACP в течение 0 (контроль), 1, 2, 3, 4 и 5 минут. После обработки ACP яйца выдерживали при комнатной температуре (22°C ± 1°C) в ламинарном шкафу в течение 24 ч перед определением количества выживших патогенов. Каждый эксперимент проводили по крайней мере три раза.
2.7 Микробиологический анализ
Аликвоты (1,0 мл) контрольного (0 мин; без обработки ACP) и обработанного ACP PBS последовательно разводили (в 10 раз) в 0,1% (мас./об.) пептоне и высевали поверхностным способом на тонкослойную агаризованную среду (TAL). Среда TAL состояла из затвердевшего агара ксилоза-лизин-тергитол-4 (XLT-4) (селективная среда для Salmonella) с верхним слоем TSAYE (∼14 мл) в качестве неселективного слоя для стимуляции реанимации сублетально поврежденных выживших клеток (Kang и Fung, 2000). Каждое яйцо переносили в отдельный соответствующим образом промаркированный стерильный мешок Seward для гомогенизатора (Seward, Уортингтон, Великобритания). В каждый мешок добавляли 50 мл 0,1% (мас./об.) пептона. Яйца вручную растирали с внешней стороны мешка, уделяя особое внимание области, где был нанесен инокулюм. Отверстие мешка закручивали, и мешок вручную встряхивали в течение 1 минуты. Из смывной жидкости готовили десятикратные последовательные разведения в 0,1% (мас./об.) пептоне, и соответствующие разведения высевали поверхностным способом на среду TAL и на агар XLT-4. Подсчет бактериальных колоний на агаровых пластинах проводили после 48 ч аэробной инкубации при 35°C.
2.8 Кинетика инактивации S. Enteritidis под действием ACP
Кинетику инактивации ACP моделировали с использованием двух из девяти моделей выживания микроорганизмов, включенных в надстройку GInaFit (версия 1.8 для Microsoft Office 365) для Excel (Geeraerd и др., 2005). Модель «логарифмическая линейная с хвостом» (Log-linear with Tail) (Geeraerd и др., 2000) использовали со следующим уравнением:
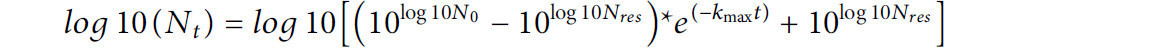
где Nt — популяция микроорганизмов в данное время обработки ACP (мин); N0 — начальная популяция микроорганизмов (log КОЕ/яйцо); Nres — остаточная популяция микроорганизмов, характеризующая «хвост» (log КОЕ/яйцо); а kmax (мин⁻¹) — скорость инактивации на логарифмически линейном участке кривой.
Модель Вейбулла (Weibull) (Mafart и др., 2002) использовала следующее уравнение:
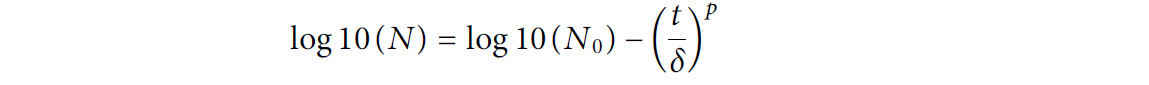
где δ — время (мин) для первой десятичной редукции популяции микроорганизмов, а p учитывает форму кривой выживания; если p > 1, кривая имеет плечо (выпуклая или вогнутая вниз), если p < 1, кривая имеет «хвост» (вогнутая), а если p = 1, кривая выживания линейна.
Для каждой модели определяли время достижения 4- и 5-логарифмической редукции. Для модели Log-linear with Tail значения D сначала рассчитывали как 2,303/kmax, умножали на x-логарифмическую редукцию (т.е. 4) с добавлением «хвоста». Для модели Вейбулла x-логарифмическую редукцию измеряли с использованием параметров модели (δ и p) в следующем уравнении (Huang и др., 2013):
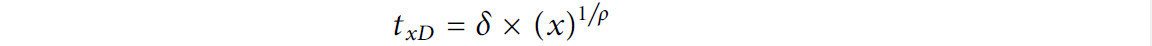
Модели оценивали с использованием четырех статистических показателей, предоставляемых GInaFit: средняя сумма квадратов ошибок (MSE), корень из средней суммы квадратов ошибок (RMSE), R² и скорректированный R² (R² adj). MSE и RMSE измеряют статистические отклонения между подобранными и наблюдаемыми экспериментальными значениями, причем меньшие значения указывают на лучшее соответствие данных. R² и R² adj оба измеряют, насколько хорошо модель соответствует данным, причем значения, близкие к единице, указывают на лучшую модель для данных.
Для оценки эффективности обеих моделей определяли фактор смещения (Bf) и фактор точности (Af) с помощью следующего уравнения (Ross, 1996):
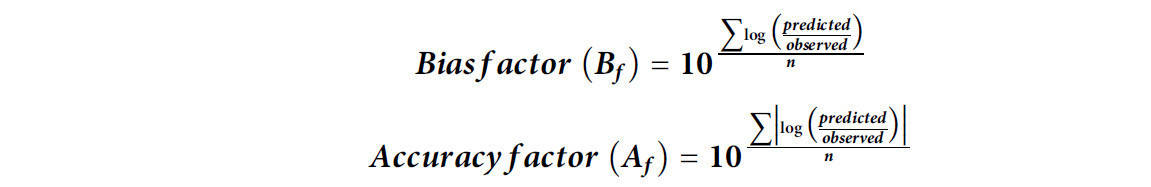
где n — количество экспериментальных наблюдений. Bf указывает, находятся ли наблюдаемые данные ниже или выше предсказанных моделью значений. Следовательно, Bf > 1 демонстрирует переоценку (завышение прогноза), Bf < 1 демонстрирует недооценку (занижение прогноза), а Bf = 1 демонстрирует точное прогнозирование. Af указывает, насколько прогнозы близки к наблюдениям.
2.9 Определение сублетального повреждения
Разницу в количестве выживших патогенов на среде TAL и селективной среде (XLT-4) использовали для расчета сублетального повреждения. Процент сублетального повреждения определяли по следующему уравнению:
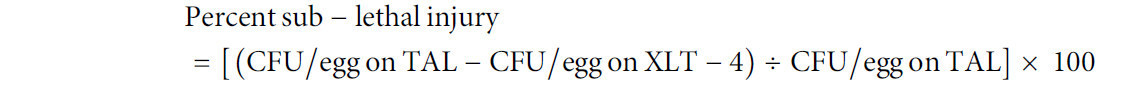
2.10 Морфология клеток EXP, STAT и LTS
Клетки EXP и STAT S. Enteritidis готовили путем штрихового посева рабочей культуры патогена на TSAYE с последующей инкубацией (35°C) в течение 8 ч (EXP) и 24 ч (STAT). Клетки из колоний EXP и STAT наносили на очищенное предметное стекло с помощью бактериологической петли, фиксировали нагреванием и окрашивали кристаллическим фиолетовым (в течение 1,0 мин) для подготовки к микроскопическому исследованию. Клетки LTS культивировали в TSBYE (35°C) в течение 21 дня (Bhullar и др., 2021), затем осаждали центрифугированием, как описано ранее. Надосадочную жидкость удаляли, а часть осажденных клеток наносили на очищенное предметное стекло и готовили для микроскопического исследования, как описано ранее. Все стекла исследовали при увеличении 1000× с использованием светового микроскопа Leica DM500, а фотографии бактериальных клеток делали с помощью микроскопной камеры Leica ICC50W (North Central Instruments, Бруклин Парк, Миннесота).
2.11 Оптическая эмиссионная спектроскопия
Спектры оптической эмиссии были собраны для анализа ROS и RNS, образующихся внутри упаковки во время обработки ACP яиц в скорлупе при 60 кВ в течение 5 минут. Спектры эмиссии во время обработки яиц регистрировали с помощью управляемого компьютером спектрометра Ocean Optics (Ocean Optics, Inc., Флорида, США). Свет, излучаемый плазмой, передавался на спектрометр через оптическое волокно Ocean Optics с диаметром сердцевины 1000 мкм. Коллимирующая линза (диаметром 5 мм), расположенная на расстоянии 15 см от края контейнера с образцом, использовалась для оптимизации попадания излучаемого плазмой света в оптическое волокно. Время интегрирования было установлено на 5 с, и усреднялось шесть спектров, в результате чего спектры эмиссии сохранялись каждые 30 секунд.
2.12 Тесты качества яиц в скорлупе
Влияние ACP на прочность скорлупы и цвет желтка оценивали через 24 ч после воздействия на яйца ACP в течение 1, 3 и 5 минут и сравнивали с теми же качественными характеристиками контрольных яиц. Прочность скорлупы как для обработанных, так и для контрольных яиц оценивали как функцию пикового положительного усилия, необходимого для 10% сжатия яйца, с использованием анализатора текстуры TA-XTplus (Stable Micro Systems, Годалминг, Англия, Великобритания). Для калибровки прибора использовали 2-килограммовый тензодатчик. Для сжатия яйца использовали зонд TA-30 диаметром 75 мм. Параметры скорости до теста, во время теста и после теста были установлены на 1,00 мм/с, 2,00 мм/с и 2,00 мм/с соответственно, с расстоянием 50 мм и пусковой силой 5 г. Цвет яичного желтка оценивали с помощью колориметра Hunter Lab ColorFlex EZ (HunterLab, Рестон, Вирджиния), который оценивал L (светлоту), a (зеленый-красный), b (синий-желтый), тон и хрому с использованием стандартного источника света CIE D65 при стандартном наблюдении 10°. Тон и хрому рассчитывали с помощью следующих уравнений (Cielab и Space, 2021):
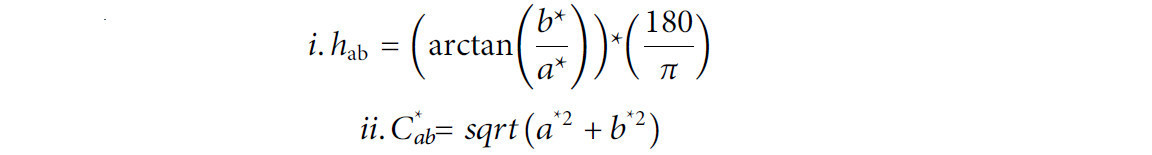
Каждый анализ проводили в четырех повторностях, и выполняли статистический анализ данных.
2.13 Статистический анализ
Эксперименты с PBS проводили в трех повторностях, тогда как испытания на яйцах в скорлупе проводили в четырех повторностях. Среднее количество выживших и стандартную ошибку среднего рассчитывали с использованием Microsoft Excel 2020 (Microsoft Inc., Редмонд, Вашингтон). Статистический анализ проводили с помощью статистического программного обеспечения JMP Pro версии 16 (SAS Institute, Inc., Кэри, Северная Каролина) с регистрацией статистической значимости при p < 0,05. Средние значения обработок оценивали на наличие статистически значимых различий (p < 0,05) с использованием двухфакторного дисперсионного анализа (ANOVA), где время рассматривалось как категориальная переменная. Для определения значимых различий между обработками использовали критерий Тьюки для Honestly Significant Difference (HSD). Оценки качества проводили в четырех повторностях с указанием среднего значения и стандартного отклонения. Средние значения обработок оценивали на статистическую значимость с использованием однофакторного ANOVA, а для оценки значимых различий между обработками использовали критерий Тьюки HSD.
3 Результаты
3.1 Инактивация S. Enteritidis в PBS
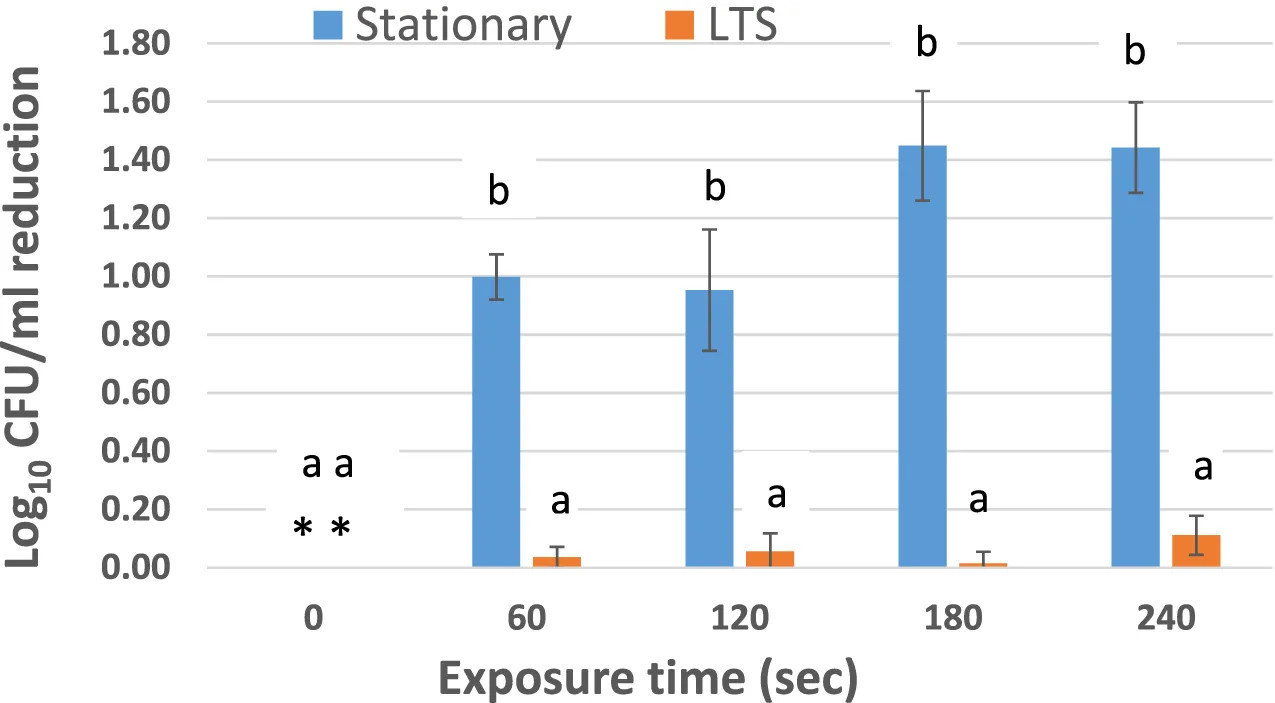
РИСУНОК 2 Логарифмическая редукция Salmonella Enteritidis ATCC 13076 после воздействия ACP (45 кВ) в PBS. Эксперимент проводили в трех повторностях. Столбики погрешностей представляют стандартную ошибку среднего. Для каждого времени обработки столбики, не имеющие одинаковых букв, значительно различаются (p < 0,05). Звездочки () означают отсутствие редукции в контрольных (не обработанных ACP) клетках STAT или LTS.
На Рисунке 2 показано снижение жизнеспособных популяций клеток STAT и LTS S. Enteritidis ATCC 13076 в PBS (pH 7,0) после прямой обработки ACP (45 кВ). В PBS исходные количества жизнеспособных клеток (log10 КОЕ/мл) STAT и LTS составляли 6,83 ± 0,06 и 7,69 ± 0,30 соответственно, на основе подсчета колоний на TSAYE. Для всех протестированных обработок ACP было значительно более высокое снижение популяций клеток STAT по сравнению с клетками LTS (p < 0,05). Среднее снижение (log10 КОЕ/мл) клеток STAT составило 1,0 ± 0,08, 0,95 ± 0,21, 1,45 ± 0,19 и 1,44 ± 0,16 после воздействия HVACP в течение 1,0, 2,0, 3,0 и 4,0 мин соответственно. Напротив, снижение log10 КОЕ/мл клеток LTS составило всего 0,04 ± 0,01 (1 мин), 0,06 ± 0,06 (2 мин), 0,01 ± 0,04 (3 мин) и 0,11 ± 0,07 (4 мин), что представляет собой <1,0 log снижения за всю продолжительность обработки.
3.2 Инактивация S. Enteritidis на яйцах в скорлупе
РИСУНОК 3 Выжившие клетки Salmonella Enteritidis ATCC 13076 на яйцах в скорлупе после воздействия ACP (60 кВ). Использованные среды для восстановления: тонкослойная агаризованная среда (TAL) (A) и агар ксилоза-лизин-тергитол 4 (XLT-4) (B). Эксперимент проводили в четырех повторностях. Столбики погрешностей представляют стандартную ошибку среднего. Для каждого времени обработки столбики, не имеющие одинаковых букв, значительно различаются (p < 0,05).
На Рисунке 3 показаны выжившие клетки STAT и LTS S. Enteritidis на внешней поверхности скорлупы яиц после прямой обработки ACP (60 кВ). Данные основаны на количестве бактериальных колоний на среде TAL (Рисунок 3A) и на агаре XLT-4 (Рисунок 3B). Средние исходные количества жизнеспособных клеток (log10 КОЕ/яйцо) S. Enteritidis на TAL составляли 6,85 ± 0,58 и 5,63 ± 0,68 для клеток STAT и LTS соответственно, тогда как на XLT-4 жизнеспособные количества составляли 6,72 ± 0,60 (STAT) и 5,55 ± 0,70 (LTS). Независимо от типа агаровой среды, после обработки ACP в течение 2–5 минут было значительно больше (p < 0,05) выживших клеток LTS по сравнению с выжившими клетками STAT (Рисунки 3A, B). Независимо от агаровой среды и времени воздействия (1–5 мин) ACP, количество выживших LTS на обработанных яйцах значимо не отличалось от такового на контрольных яйцах (p > 0,05). Напротив, все протестированные обработки ACP значительно снижали количество выживших STAT на яйцах по сравнению с контролем (p < 0,05). Выжившие STAT (независимо от агаровой среды) демонстрировали первоначальное крутое снижение до 2 минут с последующим эффектом «хвоста» с 3 по 5 минуты, который не наблюдался у выживших LTS, которые демонстрировали относительно плоский профиль кривой выживания (Рисунки 3A, B).
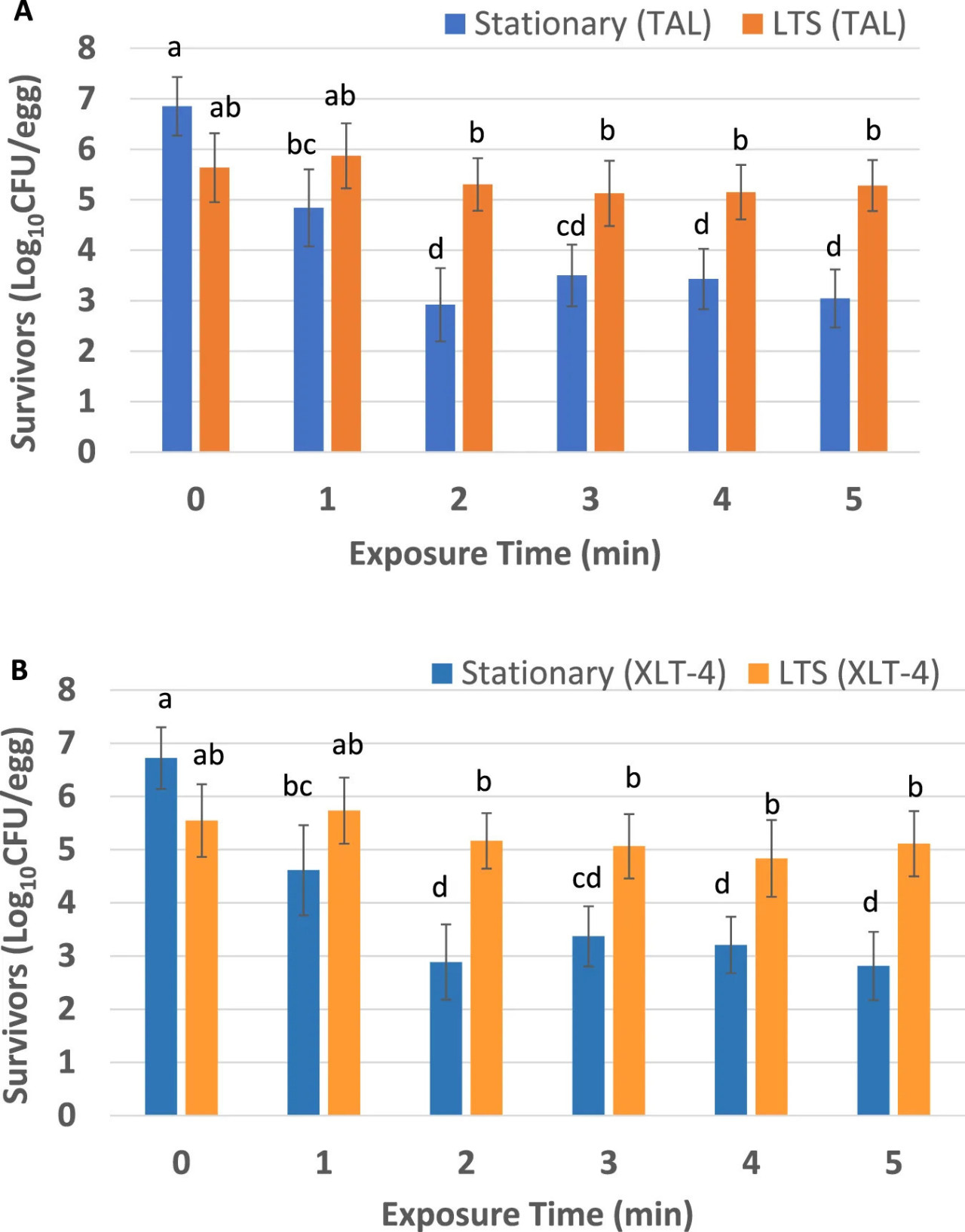
РИСУНОК 3 Выжившие клетки Salmonella Enteritidis ATCC 13076 на яйцах в скорлупе после воздействия ACP (60 кВ). Использованные среды для восстановления: тонкослойная агаризованная среда (TAL) (A) и агар ксилоза-лизин-тергитол 4 (XLT-4) (B). Эксперимент проводили в четырех повторностях. Столбики погрешностей представляют стандартную ошибку среднего. Для каждого времени обработки столбики, не имеющие одинаковых букв, значимо различаются (p < 0,05).
3.3 Кинетика инактивации S. Enteritidis под действием ACP
На Рисунке 4 показаны логарифмические графики уравнений для модели Log-linear with Tail и модели Вейбулла, подобранные к необработанным данным для выживших STAT S. Enteritidis [log10КОЕ/яйцо в зависимости от времени (мин)] на среде TAL (A) и агаре XLT-4 (B). Модель Log-linear + Tail продемонстрировала лучшее соответствие данных. Значения кинетических параметров (kmax, log10(Nres), δ- и ρ-значения) для двух моделей выживания GInaFit для обработки ACP (60 кВ) STAT S. Enteritidis на яйцах в скорлупе с использованием двух сред для восстановления показаны в Таблице 1. Статистические показатели как для модели Log-linear with Tail, так и для модели Вейбулла представлены в Таблице 2. На основе модели Log-linear with Tail были получены значения 5,29 и 5,78 минут как время обработки для достижения 4- и 5-логарифмической редукции соответственно жизнеспособных клеток STAT (Таблица 2). Значения кинетических параметров для обработки ACP клеток LTS не могли быть определены, поскольку инактивация составила менее одного (<1) log10 КОЕ/яйцо за общее время обработки 5 минут.
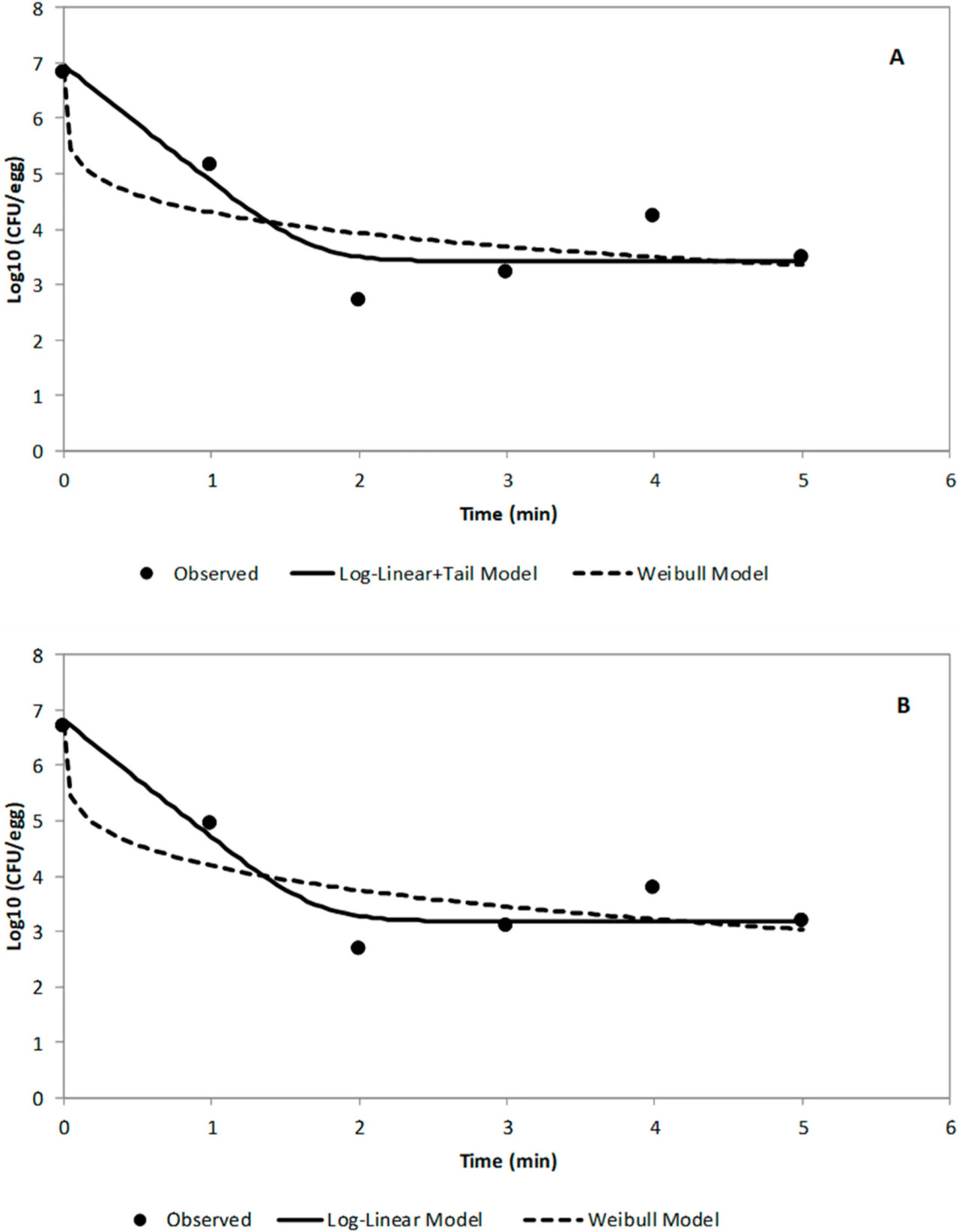
РИСУНОК 4 Логарифмические графики уравнений логарифмической линейной модели с хвостом и модели Вейбулла, подобранные к необработанным данным для выживших STAT S. Enteritidis на среде TAL (A) и агаре XLT-4 (B) как log10КОЕ/яйцо в зависимости от времени (мин).
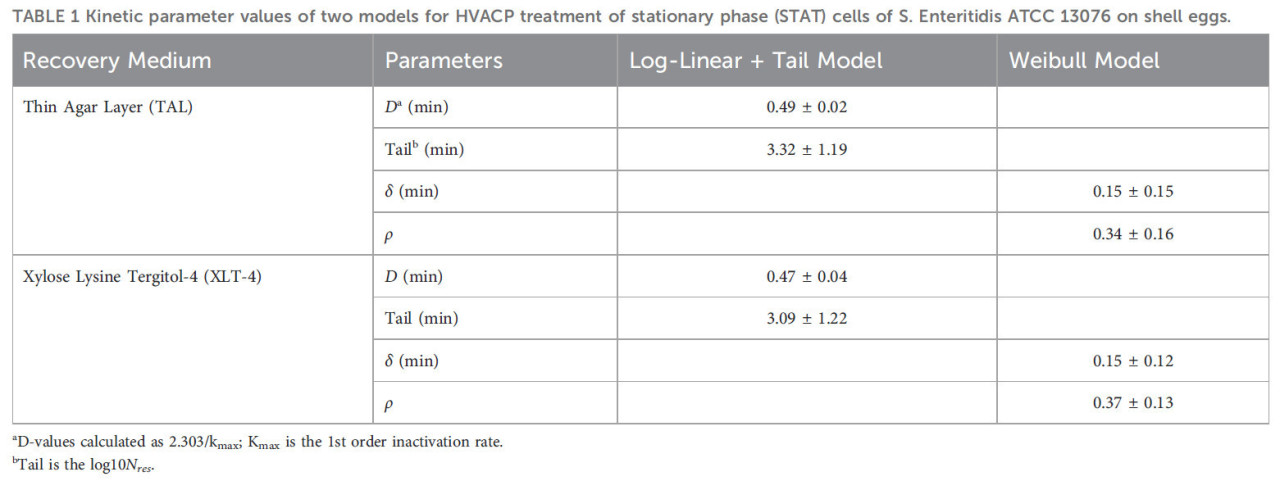
ТАБЛИЦА 1 Значения кинетических параметров двух моделей для обработки HVACP клеток стационарной фазы (STAT) S. Enteritidis ATCC 13076 на яйцах в скорлупе.
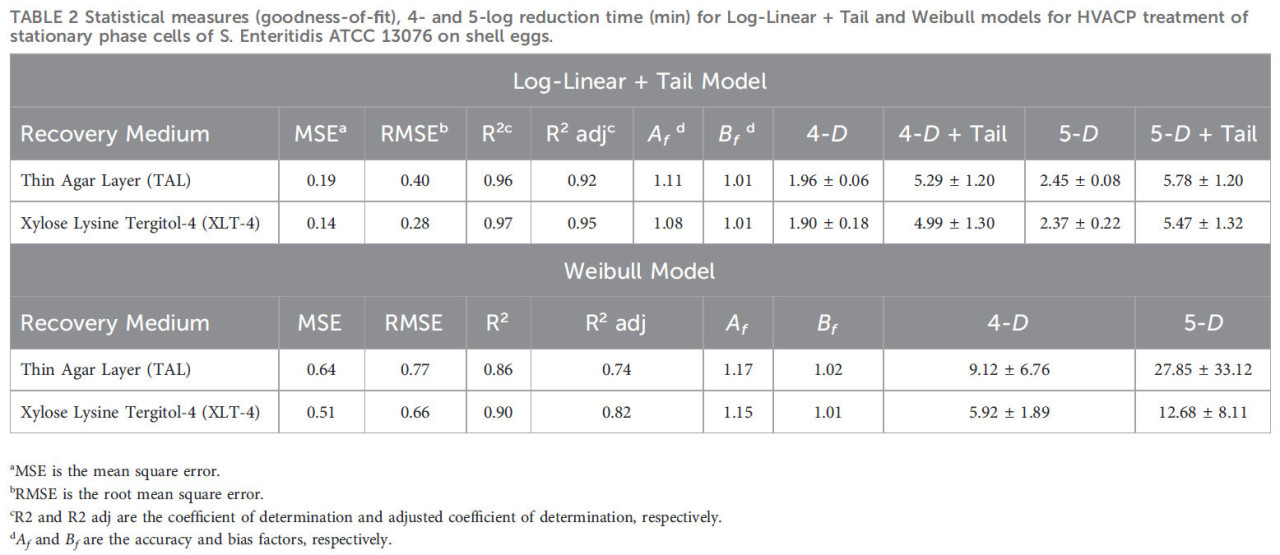
ТАБЛИЦА 2 Статистические показатели (качество соответствия), время 4- и 5-логарифмической редукции (мин) для моделей Log-linear + Tail и Вейбулла для обработки HVACP клеток стационарной фазы S. Enteritidis ATCC 13076 на яйцах в скорлупе.
a MSE — среднеквадратическая ошибка.
b RMSE — корень из среднеквадратической ошибки.
c R² и R² adj — коэффициент детерминации и скорректированный коэффициент детерминации соответственно.
d Af и Bf — факторы точности и смещения соответственно.
3.4 Сублетальное повреждение выживших S. Enteritidis на яйцах в скорлупе

ТАБЛИЦА 3 Средние значения процента сублетального повреждения ± стандартная ошибка среднего для выживших клеток стационарной фазы (STAT) и фазы длительного выживания (LTS) Salmonella Enteritidis после обработки ACP (60 кВ) в течение 1–5 минут искусственно инокулированных яиц в скорлупе.
Средние значения сублетального повреждения в выживших популяциях клеток STAT и LTS S. Enteritidis на яйцах в скорлупе после 1–5 минут воздействия ACP (60 кВ) показаны в Таблице 3. Сублетального повреждения выживших S. Enteritidis на контрольных яйцах не наблюдалось (данные не показаны). Выжившие популяции для обеих фаз жизненного цикла (STAT и LTS) патогена претерпели сублетальное повреждение из-за обработки ACP. Процент сублетального повреждения составлял от 7,22% до 37,11% и от 9,40% до 30,55% у выживших STAT и LTS соответственно. Независимо от времени воздействия ACP, не было значимых различий в сублетальном повреждении у выживших STAT по сравнению с выжившими LTS (p > 0,05).
3.5 Изображения S. Enteritidis под световым микроскопом
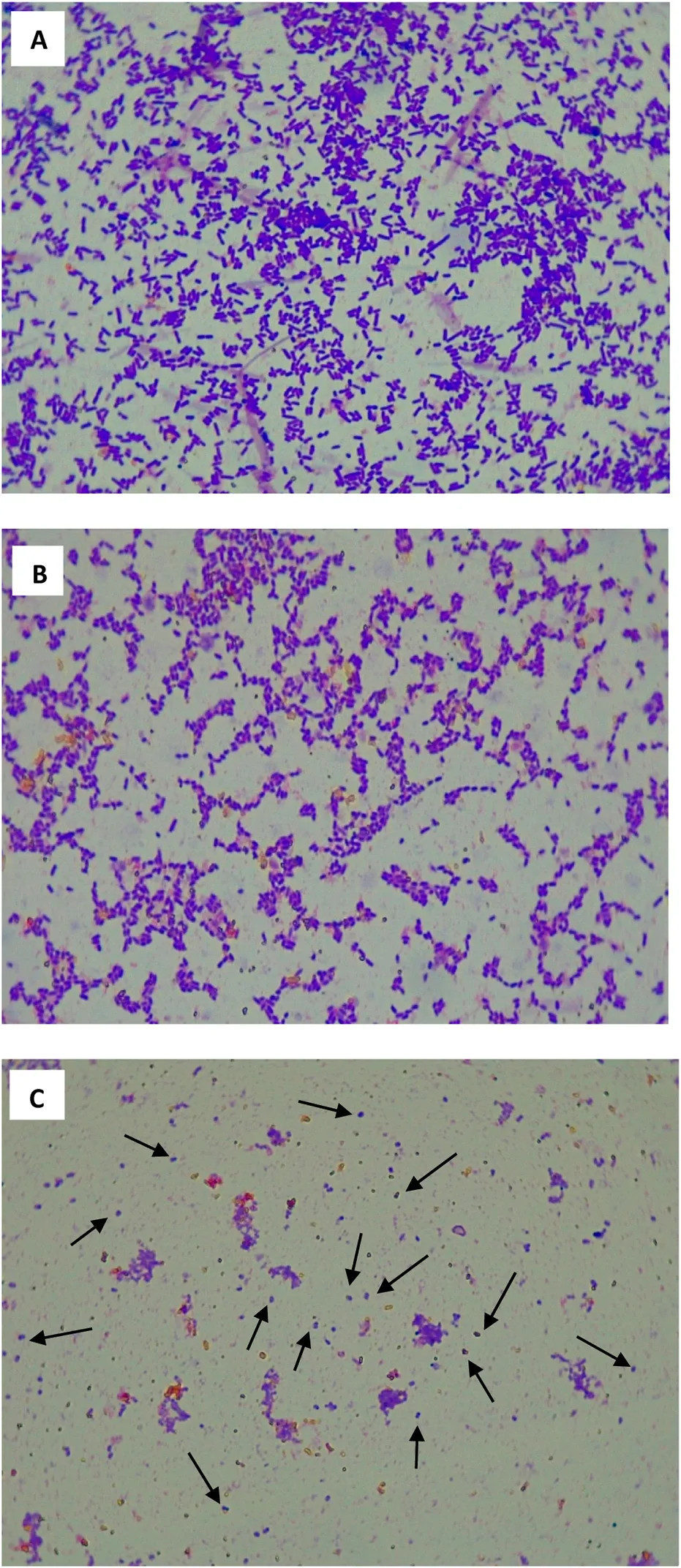
РИСУНОК 5 Изображения клеток EXP (A) , STAT (B) и LTS (C), полученных с помощью световой микроскопии (увеличение 1000 ×) . Стрелками указаны кокковидные клетки LTS.
На Рисунке 5 показаны фотографии изображений световой микроскопии клеток EXP (A), STAT (B) и LTS (C) S. Enteritidis ATCC 13076. Как популяции EXP, так и STAT (A и B соответственно) состояли преимущественно из палочковидных клеток, тогда как клетки LTS были преимущественно кокковидными (C). Клетки EXP выглядели как более длинные палочки, причем некоторые клетки находились в процессе деления. Клетки STAT представляли собой смесь более коротких палочек и кокковидных клеток. Кокковидные клетки LTS, казалось, были устойчивы к окрашиванию кристаллическим фиолетовым по сравнению с клетками EXP и STAT, которые демонстрировали интенсивный синий цвет.
3.6 Оптическая эмиссионная спектроскопия
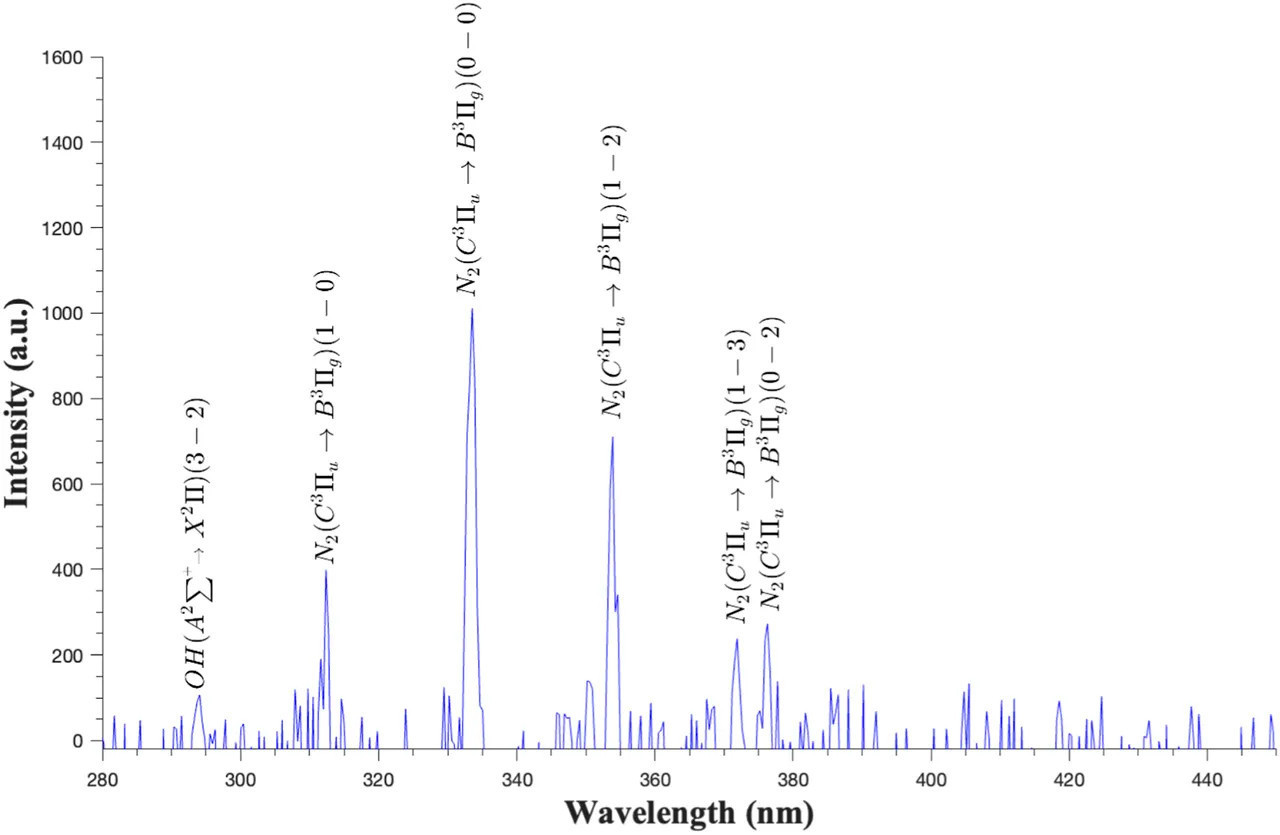
РИСУНОК 6 Спектры оптической эмиссии диэлектрического барьерного разряда ACP (60 кВ) при обработке яиц в скорлупе на воздухе при атмосферном давлении.
На Рисунке 6 показаны спектры эмиссии, полученные в результате обработки ACP (60 кВ), примененной к яйцам в скорлупе в атмосферном воздухе с относительной влажностью 60%. Преобладающие пики, наблюдаемые в воздушной плазме, были сконцентрированы в УФ-области (300–400 нм), в основном соответствуя возбужденным формам азота, таким как вторая положительная система (SPS) азота N2(C-B) и первая отрицательная система N2+(B-X) (Misra и др., 2015; Shi и др., 2017). Кроме того, пик гидроксила (OH) был обнаружен около 300 нм.
3.7 Анализ качества яиц
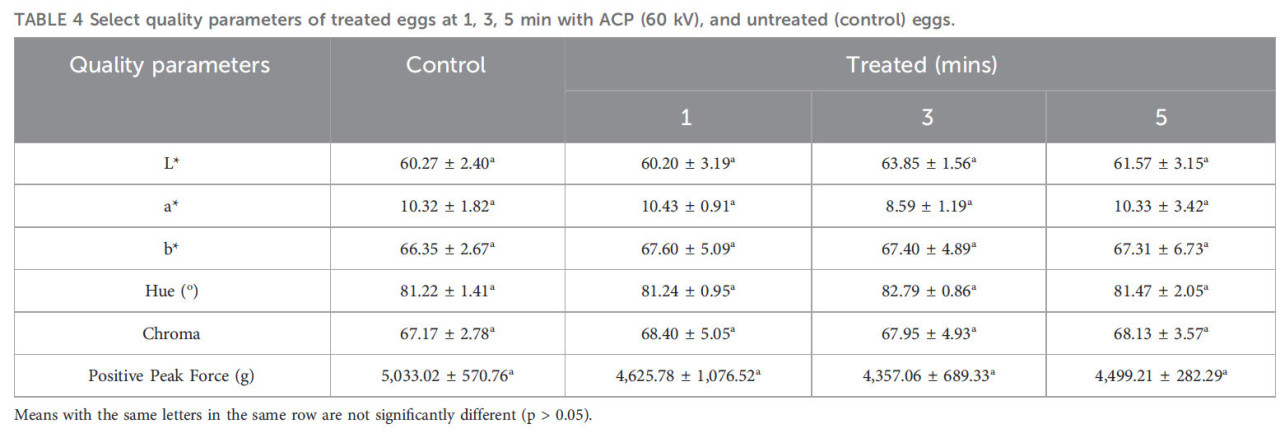
ТАБЛИЦА 4 Выбранные параметры качества обработанных яиц через 1, 3, 5 минут ACP (60 кВ) и необработанных (контрольных) яиц.
Средние значения с одинаковыми буквами в одной строке значимо не различаются (p > 0,05).
В Таблице 4 показано влияние ACP на L, a, b, тон (hab), хрому (Cab) и пиковое положительное усилие, необходимое для 10% сжатия яичной скорлупы для контрольных и обработанных яиц после 1-, 3- и 5-минутного воздействия ACP. Не было обнаружено значимых различий (p > 0,05) ни для одного из этих параметров качества. Яйца, обработанные ACP, казалось, требовали меньшего усилия: 1 мин (4625 г), 3 мин (4357 г), 5 мин (4499 г); однако эти значения оставались в пределах диапазона, наблюдаемого для контрольных яиц (4300–5900 г). Аналогично, цвет желтка, определяемый значениями L, a, b, тона и хромы для всех обработанных яиц, независимо от времени воздействия ACP, попадал в пределы наблюдаемого диапазона контрольных яиц.
4 Обсуждение
4.1 Устойчивость S. Enteritidis к HVACP в PBS и на яйцах в скорлупе
После обработки ACP PBS и яйца в скорлупе выдерживали при комнатной температуре (22°C ± 1°C) в соответствующих упаковках в ламинарном шкафу в течение 24 ч перед анализом на наличие выживших клеток. Этот шаг был предпринят для использования противомикробного эффекта относительно долгоживущих активных форм кислорода, таких как озон (Yepez и др., 2020). Значительно большая инактивация клеток STAT S. Enteritidis по сравнению с клетками LTS наблюдалась in vitro и на яйцах в скорлупе после прямого применения ACP при 45 кВ и 60 кВ соответственно. Обработка ACP при более низком напряжении (45 кВ) использовалась для PBS из-за гораздо более высокой чувствительности планктонных клеток к противомикробным обработкам по сравнению с прикрепленными клетками (Bhullar и др., 2021; Smet и др., 2019), что препятствовало адекватному восстановлению клеток для построения кривой выживания. Наши результаты о более высокой устойчивости клеток LTS к ACP по сравнению с клетками STAT согласуются с более ранними сообщениями о патогенах LTS (Bhullar и др., 2021; Djebbi-Simmons и др., 2019; Wang, 2017; Wang и др., 2018; Wen и др., 2009). Например, сообщалось о более высокой устойчивости к нагреванию и высокому гидростатическому давлению LTS L. monocytogenes по сравнению с тем же патогеном в других фазах жизненного цикла, таких как поздняя EXP, STAT, ранняя фаза отмирания и поздняя фаза отмирания (Wen и др., 2009). Для S. Typhimurium клетки LTS проявляли значительно (P < 0,05) более высокую устойчивость, чем клетки EXP и STAT, к УФ-излучению как в 0,85% (мас./об.) физиологическом растворе, так и в яблочном соке (Wang и др., 2018). Значительно более высокая устойчивость к гипохлориту натрия (0,0095%) (p < 0,05) наблюдалась для клеток LTS S. Typhimurium по сравнению с клетками STAT на пластиковых (разделочная доска, ламинат Formica) и нержавеющих стальных поверхностях (Djebbi-Simmons и др., 2019). Совсем недавно более высокие (p < 0,05) выжившие популяции LTS шига-токсин-продуцирующей E. coli по сравнению с клетками STAT наблюдались после воздействия in vitro раствора гипохлорита натрия и промывки искусственно инокулированных листьев салата растворами гипохлорита (Bhullar и др., 2021). Эти результаты указывают на то, что клетки LTS более устойчивы к противомикробным обработкам, чем клетки STAT.
Условия в высокопитательном лабораторном бульоне с оптимальным pH, активностью воды и температурой роста, а также отсутствием микробных конкурентов нетипичны для естественной среды обитания бактерий. Напротив, ограничение питательных веществ, накопление отходов, конкурирующая микрофлора и суровые условия окружающей среды, вероятно, являются нормой, а не исключением для бактерий в природе (Roszak и Colwell, 1987; Finkel, 2006). Такие условия могут вызвать переход бактерий в LTS (Finkel, 2006), таким образом входя в состояние покоя с низкой скоростью метаболизма (Kolter и др., 1993). Например, ограничение питательных веществ индуцирует покой клеток у морских бактерий (Novitsky и Morita, 1978). Более того, Pseudomonas aeruginosa проявляет фенотип покоя во время своего длительного выживания в воде (Lewenza и др., 2018). При вышеупомянутых условиях некоторые грамположительные бактерии продуцируют эндоспоры; однако большинство грамотрицательных бактерий и не образующих эндоспоры грамположительных бактерий образуют покоящиеся клетки. Покоящиеся бактериальные клетки обычно не растут, проявляя мало метаболической активности или не проявляя ее вовсе, и большинство противомикробных агентов неэффективны против бактериальных клеток, которые не растут активно и не делятся (Reygaert, 2018).
4.2 Кинетика инактивации S. Enteritidis под действием ACP на яйцах в скорлупе
Модели Log-linear with Tail и Вейбулла были подобраны к данным по выживанию, полученным из подсчетов бактерий на двух средах для восстановления: TAL и XLT-4. Выживаемость S. Enteritidis демонстрировала отклонение от линейного снижения логарифма числа выживших со временем. Наблюдалось образование «хвоста» кривых выживания или субпопуляция более устойчивых бактерий, снижающихся с более низкой скоростью (Рисунок 1; Таблица 1). Параметр формы модели Вейбулла (p < 1) указывает на то, что кривые выживания S. Enteritidis, подогнанные этой моделью, были вогнутыми. Тем не менее, модель Log-linear with Tail обеспечила лучшее соответствие обработке ACP клеток STAT на яйцах в скорлупе (на основе подсчета колоний на средах TAL и XLT-4), о чем свидетельствуют более высокие значения R² и R² adj и меньшие значения MSE и RMSE (Таблица 2). Согласно Ross и др. (2000), значения фактора точности (Af) 0,90–1,15 (увеличение или уменьшение на 0,1–0,15) считаются приемлемыми. Значения Af составляли от 1,08 до 1,11 и от 1,15 до 1,17 для моделей Log-linear with Tail и Вейбулла соответственно, что дополнительно указывает на то, что модель Log-linear with Tail давала лучшие прогнозы для обработки ACP клеток STAT на яйцах. По сравнению с моделью Вейбулла, время, необходимое для 4- и 5-логарифмической инактивации клеток STAT на яйцах в скорлупе обработкой ACP, было ниже при использовании модели Log-linear with Tail (Таблица 2).
4.3 Индуцированное ACP сублетальное повреждение
Наши результаты показывают, что часть выживших после обработки ACP S. Enteritidis на яйцах в скорлупе была сублетально повреждена независимо от фазы жизненного цикла патогена (Таблица 1). Wang и др. (2018) наблюдали, что сублетальное повреждение у LTS Salmonella Typhimurium было меньшим по сравнению с клетками STAT этого патогена после обработки ультрафиолетовым излучением (1500 мкВт/см²) в яблочном соке (p < 0,05). Выжившие LTS шига-токсин-продуцирующей Escherichia coli после обработки хлором (40 млн⁻¹) листьев салата имели значительно меньшее сублетальное повреждение по сравнению с выжившими STAT (Bhullar и др., 2021). Наши результаты отличаются от результатов вышеупомянутых исследований тем, что степень индуцированного ACP сублетального повреждения у выживших STAT и LTS S. Enteritidis значимо не различалась (p > 0,05). Это различие может быть связано с типом сублетального повреждения, вызванного ACP, которая имеет очень сложный химический состав.
Химический состав ACP включает множество различных активных форм, действующих независимо, аддитивно или синергетически для инактивации микроорганизмов (Bourke и др., 2017). Следовательно, после каждой обработки ACP, вероятно, будут возникать большие вариации в типе и степени клеточных повреждений. Как для клеток STAT, так и LTS, мы постоянно наблюдали большие вариации в степени сублетального повреждения, вызванного каждой обработкой ACP, в разных повторностях эксперимента. Это, в свою очередь, снижает статистическую значимость средних значений процента сублетального повреждения для фаз жизненного цикла (STAT и LTS). Поэтому, основываясь на результатах настоящего исследования, трудно определить, зависит ли степень индуцированного ACP сублетального повреждения выживших S. Enteritidis значимо от фазы жизненного цикла патогена. Однако наблюдаемое сублетальное повреждение как у выживших STAT, так и LTS имеет последствия для безопасности пищевых продуктов. В этом контексте определение сублетально поврежденных пищевых патогенов имеет критическое значение по двум основным причинам. Во-первых, игнорирование сублетально поврежденных патогенов может привести к переоценке противомикробной эффективности обработки, и во-вторых, пищевые процессы, наносящие сублетальные повреждения микроорганизмам, открывают возможности для их применения в сочетании с другими методами консервации пищевых продуктов для предотвращения реактивации и последующей пролиферации патогенных микроорганизмов (Thomas-Popo и др., 2019). Таким образом, мы предполагаем, что ACP имеет хороший потенциал для применения в рамках системы «барьерной технологии» для повышения микробиологической безопасности яиц в скорлупе.
4.4 Морфология клеток EXP, STAT и LTS
По мере того как палочковидные клетки S. Enteritidis переходили от EXP к STAT и LTS, их морфология менялась от вытянутых или палочковидных к кокковидным или сферическим клеткам (Рисунки 5A–C). Это наблюдение согласуется с более ранними сообщениями Nystrom (2004) и Jaishankar и Srivastava (2017), которые оба сообщали об уменьшении размера бактериальных клеток по мере их вхождения в STAT. Это уменьшение включало конденсацию нуклеоида, увеличение периплазматического пространства и снижение текучести мембраны, которые являются результатом генов, индуцируемых голоданием, что способствует повышению выживаемости клеток в стрессовых условиях. Аналогично, Mañas и Mackey (2004) сообщали о палочковидных и кокковидных формах E. coli в клетках EXP и STAT соответственно. Однако важно отметить, что клетки STAT не являются исключительно кокковидными. Поскольку некоторые клетки все еще реплицируются, культура STAT представляет собой гетерогенную смесь палочек и кокков. К моменту перехода клеток в LTS популяция становится почти полностью кокковидной (Рисунок 5C). Эти клетки уменьшаются даже больше, чем клетки STAT, и выглядят меньше. Наши результаты также согласуются с наблюдениями Wen и др. (2009), которые сообщили о значительно меньших кокковидных клетках LTS L. monocytogenes по сравнению с частично кокковидными клетками STAT и палочковидными клетками EXP. Те же авторы наблюдали очень слабое окрашивание кокковидных клеток LTS, что указывало на изменение клеточной оболочки или более конденсированную кристаллическую (стекловидную) цитоплазму. Подобно состоянию ядра бактериальных эндоспор, конденсация цитоплазмы может снижать активность воды в кокковидных клетках, таким образом повышая устойчивость клеток ко многим стрессам (Wen и др., 2009). Во время перехода L. monocytogenes в LTS в TSBYE наиболее значительно активируемыми генами были гены, ответственные за транспорт, энергетический метаболизм и структуру клеточной оболочки (Wen и др., 2011). Следовательно, наблюдаемое нами повышение устойчивости LTS S. Enteritidis к ACP может быть связано с изменением клеточной оболочки или снижением внутриклеточной активности воды, что может уменьшить внутриклеточное образование активных форм.
4.5 Оптическая эмиссионная спектроскопия
Оптическая эмиссионная спектроскопия (OES) была проведена для исследования активных газообразных форм, образующихся внутри упаковки во время обработки ACP яиц в скорлупе. Спектры эмиссии (Рисунок 6) демонстрируют сходство и сопоставимость со спектрами эмиссии, сообщавшимися в более ранних исследованиях, в которых для генерации плазмы использовался атмосферный воздух (Pankaj и др., 2017; Wan и др., 2017). Наши результаты указывают на низкую интенсивность пиков атомарного кислорода, что может быть объяснено самотушением O(³P) и O(⁵P) в воздушной плазме (Fridman, 2008; Walsh и др., 2010). Во время обработки ACP ионизация газовых молекул генерирует ROS и RNS, которые обладают бактерицидным действием и вызывают повреждения клеток, такие как повреждение ДНК и перекисное окисление липидов мембран (Han и др., 2016; Joshi и др., 2011). Наши результаты показывают, что ACP, использованная в условиях настоящего исследования, была источником как ROS, так и RNS, которые могут быть летальны для бактерий (Bourke и др., 2017).
4.6 Влияние на качество яиц
Прочность яичной скорлупы является серьезной экономической проблемой для яичной промышленности. Это вопрос первостепенной важности во время таких операций, как сбор яиц, сортировка и, что более важно, во время транспортировки, когда может происходить высокий уровень повреждения скорлупы яиц. По оценкам, поврежденные яйца составляют 8%–11% от общих потерь яиц (Dunn и др., 2009). Поэтому важно оценить влияние новых противомикробных вмешательств на прочность яичной скорлупы. В настоящем исследовании качество яиц, обработанных ACP, и необработанных (контрольных) оценивали по прочности скорлупы и цвету желтка. Наши результаты показывают, что обработки ACP, независимо от времени воздействия, значимо не влияли на прочность яичной скорлупы, хотя наблюдалось, что прочность скорлупы немного снижалась после воздействия ACP в течение 1, 3 и 5 минут (P > 0,05). Поскольку прочность скорлупы обработанных ACP и контрольных яиц оставалась в одном диапазоне, результаты для обработанных яиц, вероятно, связаны с вариабельностью между отдельными яйцами, а не с эффектом ACP. Тем не менее, наши результаты согласуются с предыдущими результатами исследований, которые продемонстрировали незначительные изменения целостности яичной скорлупы после обработки ACP яиц в скорлупе (Ragni и др., 2010). Эти результаты позволяют предположить, что ACP в условиях, протестированных в настоящем исследовании, не ухудшает прочность скорлупы.
Цвет является важным сенсорным свойством, которое влияет на восприятие потребителем и выбор определенных продуктов. Например, потребители воспринимают яйца с интенсивно окрашенным желтком как здоровую, богатую питательными веществами пищу (Williams, 1992; Saleh и др., 2021). Более того, цвет яичного желтка является одной из основных характеристик, по которым оценивается качество яиц (Beardsworth и Hernandez, 2004). Следовательно, цвет яичного желтка является важным показателем восприятия потребителем внутреннего качества яиц в скорлупе. Мы оценивали цвет яичного желтка на основе светлоты (L), цвета по осям зеленый (−), красный (+) (a), и синий (−), желтый (+) (b), тона и интенсивности цвета. Не было обнаружено визуальных различий между протестированными образцами желтка, даже после 5 минут, максимального времени обработки ACP. Эти результаты согласуются с Dasan и др. (2018), Wan и др. (2017), Georgescu и др. (2017) и Gavahian и др. (2019), которые все сообщали о незначительных изменениях цвета желтка после обработок ACP.
Каротиноиды в яичном желтке придают ему желто-оранжевый оттенок (Anton, 2007). Учитывая, что ACP может оказывать окислительное действие, окисление каротиноидов в яичном желтке требовало изучения. Окисление может вызвать выцветание цвета, что негативно влияет на восприятие потребителем; однако, поскольку ACP является поверхностной обработкой, возможно, что потенциальные окислительные эффекты не проникают за пределы оболочки скорлупы, таким образом сохраняя целостность цвета желтка неповрежденных яиц в скорлупе.
5 Заключение
Клетки LTS S. Enteritidis на яйцах в скорлупе проявляют значительно более высокую устойчивость к обработке ACP по сравнению с клетками STAT. Клетки LTS сжимаются, превращаясь преимущественно в кокковидные клетки, которые являются слегка рефрактильными. ACP вызывает большие вариации в степени сублетального повреждения S. Enteritidis независимо от фазы жизненного цикла патогена. В условиях испытаний настоящего исследования ACP оказывает незначительное влияние на прочность яичной скорлупы и цвет желтка. Основные результаты настоящего исследования показывают, что LTS обеспечивает перекрестную защиту S. Enteritidis от ACP, и это следует учитывать при разработке и валидации параметров обработки ACP для обеспечения микробиологической безопасности яиц в скорлупе.
Сокращения
ACP, атмосферная холодная плазма; BHI, бульон на основе мозгово-сердечной инфузии; DBD, диэлектрический барьерный разряд; EXP, экспоненциальная фаза; HVACP, высоковольтная атмосферная холодная плазма; LTS, фаза длительного выживания; MSE, средняя сумма квадратов ошибок; RMSE, корень из средней суммы квадратов ошибок; STAT, стационарная фаза; TAL, тонкослойная агаризованная среда; TSAYE, триптиказо-соевый агар с дрожжевым экстрактом; TSBYE, триптиказо-соевый бульон с дрожжевым экстрактом; XLT-4, ксилоза-лизин-тергитол-4.
Ссылки
1. Abdoli B., Khoshtaghaza M. H., Ghomi H., Torshizi M. A. K., Mehdizadeh S. A., Pishkar G. et al. (2024). Cold atmospheric pressure air plasma jet disinfection of table eggs: inactivation of Salmonella enterica, cuticle integrity and egg quality. Int. J. Food Microbiol. 410, 110474. doi: 10.1016/j.ijfoodmicro.2023.110474. CrossRef. Google Scholar.
2. Anton M. (2007). Composition and structure of hen egg yolk. Bioact. Egg Compd., 1–6. doi: 10.1007/978-3-540-37885-3_1. CrossRef. Google Scholar.
3. Beardsworth P. M., Hernandez J. M. (2004). Yolk colour--an important egg quality attribute. Int. Poult. Prod. 12 (5), 17–18. Google Scholar.
4. Bhullar M. S., Shaw A., Mendonca A., Monge A., Nabwire L., Thomas-Popo E. (2021). Shiga toxin--producing Escherichia coli in the long-term survival phase exhibit higher chlorine tolerance and less sublethal injury following chlorine treatment of romaine lettuce. Foodborne Pathogens Dis. 18, 276–282. doi: 10.1089/fpd.2020.2873. CrossRef. Google Scholar.
5. Bourke P., Ziuzina D., Han L., Cullen P. J., Gilmore B. F. (2017). Microbiological interactions with cold plasma. J. Appl. Microbiol. 123 (2), 308–324. doi: 10.1111/jam.13429. CrossRef. Google Scholar.
6. Buchmüller K., Bearth A., Siegrist M. (2020). Consumers' perceptions of chemical household products and the associated risks. Food Chem. Toxicol. 143, 111511. doi: 10.1016/j.fct.2020.111511. CrossRef. Google Scholar.
7. CDC (2018). Outbreak of Salmonella infections linked to gravel Ridge Farms shell eggs - final update. Atlanta, GA, United States: Centers for Disease Control and Prevention. Google Scholar.
8. CDC (2021). Salmonella and food handlers. Lancet. doi: 10.1016/s0140-6736(87)91345-6. CrossRef. Google Scholar.
9. Cielab T., Space C. (2021). The CIELAB L * a * b * system -- the method to quantify colors of coatings, 1–4. Google Scholar.
10. Dasan B. G., Yildirim T., Boyaci I. H. (2018). Surface decontamination of eggshells by using non-thermal atmospheric plasma. Int. J. Food Microbiol. 266, 267–273. doi: 10.1016/j.ijfoodmicro.2017.12.021. CrossRef. Google Scholar.
11. Deng S., Cheng C., Ni G., Meng Y., Chen H. (2010). Bacillus subtilis devitalization mechanism of atmosphere pressure plasma jet. Curr. Appl. Phys. 10 (4), 1164–1168. doi: 10.1016/j.cap.2010.02.004. CrossRef. Google Scholar.
12. Djebbi-Simmons D., Xu W., Janes M., King J. (2019). Survival and inactivation of Salmonella enterica serovar Typhimurium on food contact surfaces during log, stationary and long-term stationary phases. Food Microbiol. 84, 103272. doi: 10.1016/j.fm.2019.103272. CrossRef. Google Scholar.
13. Dunn I. C., Joseph N. T., Bain M., Edmond A., Wilson P. W., Milona P. et al. (2009). Polymorphisms in eggshell organic matrix genes are associated with eggshell quality measurements in pedigree Rhode Island Red hens. Anim. Genet. 40 (1), 110–114. doi: 10.1111/j.1365-2052.2008.01794.x. CrossRef. Google Scholar.
14. Finkel S. E. (2006). Long-term survival during stationary phase: evolution and the GASP phenotype. Nat. Rev. Microbiol. 4 (2), 113–120. doi: 10.1038/nrmicro1340. CrossRef. Google Scholar.
15. Fridman A. (2008). Plasma chemistry. New York: Cambridge University Press. Google Scholar.
16. Gavahian M., Peng H.-J., Chu Y.-H. (2019). Efficacy of cold plasma in producing Salmonella-free duck eggs: effects on physical characteristics, lipid oxidation, and fatty acid profile. J. Food Sci. Technol. 56 (12), 5271–5281. doi: 10.1007/S13197-019-03996-Z. CrossRef. Google Scholar.
17. Geeraerd A. H., Herremans C. H., Van Impe J. F. (2000). Structural model requirements to describe microbial inactivation during a mild heat treatment. Int. J. food Microbiol. 59 (3), 185–209. doi: 10.1016/s0168-1605(00)00362-7. CrossRef. Google Scholar.
18. Geeraerd A. H., Valdramidis V. P., Van Impe J. F. (2005). GInaFiT, a freeware tool to assess non--log-linear microbial survivor curves. Int. J. Food Microbiol. 102, 95–105. doi: 10.1016/j.ijfoodmicro.2004.11.038. CrossRef. Google Scholar.
19. Georgescu N., Apostol L., Gherendi F. (2017). Inactivation of Salmonella enterica serovar Typhimurium on egg surface, by direct and indirect treatments with cold atmospheric plasma. Food control. 76, 52–61. doi: 10.1016/j.foodcont.2017.01.005. CrossRef. Google Scholar.
20. Goslan E. H., Krasner S. W., Bower M., Rocks S. A., Holmes P., Levy L. S. et al. (2009). A comparison of disinfection by-products found in chlorinated and chloraminated drinking waters in Scotland. Water Res. 43 (18), 4698–4706. doi: 10.1016/j.watres.2009.07.029. CrossRef. Google Scholar.
21. Han L., Patil S., Boehm D., Milosavljević V., Cullen P. J., Bourke P. (2016). Mechanisms of inactivation by high-voltage atmospheric cold plasma differ for Escherichia coli and Staphylococcus aureus. Appl. Environ. Microbiol. 82 (2), 450–458. doi: 10.1128/AEM.02660-15. CrossRef. Google Scholar.
22. Huang K., Yu L., Liu D., Gai L., Wang J. (2013). Modeling of yeast inactivation of PEF-treated Chinese rice wine: effects of electric field intensity, treatment time and initial temperature. Food Res. Int. 54 (1), 456–467. doi: 10.1016/j.foodres.2013.07.046. CrossRef. Google Scholar.
23. Illera A. E., Souza V. R., Nikmaram N., Tang L., Keener K. M. (2022). High voltage atmospheric cold plasma decontamination of Salmonella enteritidis on chicken eggs. Innovative Food Sci. and Emerg. Technol. 82, 103210. doi: 10.1016/j.ifset.2022.103210. CrossRef. Google Scholar.
24. Illera A. E., Souza V. R., Tang L., Nikmaram N., Keener K. M. (2023). Effect of high voltage atmospheric cold plasma on chicken eggs quality during refrigerated storage. Food Biosci. 53, 102754. doi: 10.1016/j.fbio.2023.102754. CrossRef. Google Scholar.
25. Jaishankar J., Srivastava P. (2017). Molecular basis of stationary phase survival and applications. Front. Microbiol. 8, 2000. doi: 10.3389/fmicb.2017.02000. CrossRef. Google Scholar.
26. Joshi S. G., Cooper M., Yost A., Paff M., Ercan U. K., Fridman G. et al. (2011). Nonthermal dielectric-barrier discharge plasma-induced inactivation involves oxidative DNA damage and membrane lipid peroxidation in Escherichia coli. Antimicrob. agents Chemother. 55 (3), 1053–1062. doi: 10.1128/AAC.01002-10. CrossRef. Google Scholar.
27. Kang D. H., Fung D. Y. (2000). Application of thin agar layer method for recovery of injured Salmonella typhimurium. Int. J. Food Microbiol. 54 (1-2), 127–132. doi: 10.1016/s0168-1605(99)00174-9. CrossRef. Google Scholar.
28. Keerthirathne T. P., Ross K., Fallowfield H., Whiley H. (2017). Reducing risk of salmonellosis through egg decontamination processes. Int. J. Environ. Res. public health 14 (3), 335. doi: 10.3390/ijerph14030335. CrossRef. Google Scholar.
29. Kolter R., Ie D. A. S., Torma A. (1993). The stationary phase of the bacterial life cycle. Annu. Rev. Microbiol. 47, 855–874. doi: 10.1146/annurev.mi.47.100193.004231. CrossRef. Google Scholar.
30. Lewenza S., Abboud J., Poon K., Kobryn M., Humplik I., Bell J. R. et al. (2018). Pseudomonas aeruginosa displays a dormancy phenotype during long-term survival in water. PLoS ONE 13 (9), 01983844–e198419. doi: 10.1371/journal.pone.0198384. CrossRef. Google Scholar.
31. Lin L., Liao X., Li C., Abdel-Samie M. A., Cui H. (2020). Inhibitory effect of cold nitrogen plasma on Salmonella Typhimurium biofilm and its application on poultry egg preservation. LWT 126, 109340. doi: 10.1016/j.lwt.2020.109340. CrossRef. Google Scholar.
32. Mafart P., Couvert O., Gaillard S., Leguérinel I. (2002). On calculating sterility in thermal preservation methods: application of the Weibull frequency distribution model. Int. J. Food Microbiol. 72 (1–2), 107–113. doi: 10.1016/S0168-1605(01)00624-9. CrossRef. Google Scholar.
33. Mañas P., Mackey B. M. (2004). Morphological and physiological changes induced by high hydrostatic pressure in exponential- and stationary-phase cells of Escherichia coli: relationship with cell death. Appl. Environ. Microbiol. 70 (3), 1545–1554. doi: 10.1128/AEM.70.3.1545-1554.2004. CrossRef. Google Scholar.
34. Meireles A., Giaouris E., Simões M. (2016). Alternative disinfection methods to chlorine for use in the fresh-cut industry. Food Res. Int. 82, 71–85. doi: 10.1016/j.foodres.2016.01.021. CrossRef. Google Scholar.
35. Misra N., Schülter O. K., Cullen P. (2016). "Cold plasma in food and agriculture," in Cold plasma in food and agriculture. doi: 10.1016/c2014-0-00009-3. CrossRef. Google Scholar.
36. Misra N. N., Keener K. M., Bourke P., Cullen P. J. (2015). Generation of in-package cold plasma and efficacy assessment using methylene blue. Plasma Chem. Plasma Process. 35, 1043–1056. doi: 10.1007/s11090-015-9638-5. CrossRef. Google Scholar.
37. Moritz M., Wiacek C., Weihe T., Ehlbeck J., Weltmann K. D., Braun P. G. (2021). Effect of cold atmospheric pressure plasma treatment of eggshells on the total bacterial count inoculated Salmonella Enteritidis and selected quality parameters. Plasma Process. Polym. 18 (1), 2000061. doi: 10.1002/ppap.202000061. CrossRef. Google Scholar.
38. Niemira B. A. (2012). Cold plasma decontamination of foods. Annu. Rev. Food Sci. Technol. 3 (1), 125–142. doi: 10.1146/annurev-food-022811-101132. CrossRef. Google Scholar.
39. Nou X., Luo Y. (2010). Whole-leaf wash improves chlorine efficacy for microbial reduction and prevents pathogen cross-contamination during fresh-cut lettuce processing. J. Food Sci. 75 (5), M283–M290. doi: 10.1111/j.1750-3841.2010.01630.x. CrossRef. Google Scholar.
40. Novitsky J. A., Morita R. Y. (1978). Possible strategy for the survival of marine bacteria under starvation conditions. Mar. Biol. 48 (3), 289–295. doi: 10.1007/BF00397156. CrossRef. Google Scholar.
41. Nyström T. (2004). Stationary-phase physiology. Annu. Rev. Microbiol. 58, 161–181. doi: 10.1146/annurev.micro.58.030603.123818. CrossRef. Google Scholar.
42. Ölmez H., Kretzschmar U. (2009). Potential alternative disinfection methods for organic fresh-cut industry for minimizing water consumption and environmental impact. LWT - Food Sci. Technol. 42 (3), 686–693. doi: 10.1016/J.LWT.2008.08.001. CrossRef. Google Scholar.
43. Pankaj S. K., Wan Z., Colonna W., Keener K. M. (2017). Degradation kinetics of organic dyes in water by high voltage atmospheric air and modified air cold plasma. Water Sci. Technol. 76 (3), 567–574. doi: 10.2166/wst.2017.169. CrossRef. Google Scholar.
44. Pijnacker R., Dallman T. J., Tijsma A. S. L., Hawkins G., Larkin L., Kotila S. M. et al. (2019). An international outbreak of Salmonella enterica serotype Enteritidis linked to eggs from Poland: a microbiological and epidemiological study. Lancet Infect. Dis. 19 (7), 778–786. doi: 10.1016/S1473-3099(19)30047-7. CrossRef. Google Scholar.
45. Ragni L., Berardinelli A., Vannini L., Montanari C., Sirri F., Guerzoni M. E. et al. (2010). Non-thermal atmospheric gas plasma device for surface decontamination of shell eggs. J. Food Eng. 100 (1), 125–132. doi: 10.1016/j.jfoodeng.2010.03.036. CrossRef. Google Scholar.
46. Reygaert W. C. (2018). An overview of the antimicrobial resistance mechanisms of bacteria. AIMS Microbiol. 4 (3), 482–501. doi: 10.3934/microbiol.2018.3.482. CrossRef. Google Scholar.
47. Ross T. (1996). Indices for performance evaluation of predictive models in food microbiology. J. Appl. Bacteriol. 81 (5), 501–508. doi: 10.1111/j.1365-2672.1996.tb03539.x. CrossRef. Google Scholar.
48. Ross T., Dalgaard P., Tienungoon S. (2000). Predictive modelling of the growth and survival of Listeria in fishery products. Int. J. Food Microbiol. 62 (3), 231–245. doi: 10.1016/S0168-1605(00)00340-8. CrossRef. Google Scholar.
49. Roszak D. B., Colwell R. (1987). Survival strategies of bacteria in the natural environment. Microbiol. Rev. 51 (3), 365–379. doi: 10.1128/mr.51.3.365-379.1987. CrossRef. Google Scholar.
50. Saleh A. A., Gawish E., Mahmoud S. F., Amber K., Awad W., Alzawqari M. H. et al. (2021). Effect of natural and chemical colorant supplementation on performance, egg-quality characteristics, yolk fatty-acid profile, and blood constituents in laying hens. Sustainability 13 (8), 4503. doi: 10.3390/su13084503. CrossRef. Google Scholar.
51. Scholtz V., Pazlarova J., Souskova H., Khun J., Julak J. (2015). Nonthermal plasma - a tool for decontamination and disinfection. Biotechnol. Adv. 33 (6), 1108–1119. doi: 10.1016/j.biotechadv.2015.01.002. CrossRef. Google Scholar.
52. Shi H., Ileleji K., Stroshine R. L., Keener K., Jensen J. L. (2017). Reduction of aflatoxin in corn by high voltage atmospheric cold plasma. Food Bioprocess Technol. 10, 1042–1052. doi: 10.1007/s11947-017-1873-8. CrossRef. Google Scholar.
53. Smet C., Govaert M., Kyrylenko A., Easdani M., Walsh J. L., Van Impe J. F. (2019). Inactivation of single strains of Listeria monocytogenes and Salmonella typhimurium planktonic cells biofilms with plasma activated liquids. Front. Microbiol. 10 (JULY), 1539–1615. doi: 10.3389/fmicb.2019.01539. CrossRef. Google Scholar.
54. Statistica (2024). Per capita consumption of eggs in the U.S. 2000-2024. Google Scholar.
55. Thomas-Popo E., Mendonça A., Misra N. N., Little A., Wan Z., Moutiq R. et al. (2019). Inactivation of Shiga-toxin-producing Escherichia coli, Salmonella enterica and natural microflora on tempered wheat grains by atmospheric cold plasma. Food control. 104, 231–239. doi: 10.1016/j.foodcont.2019.04.025. CrossRef. Google Scholar.
56. Tirpanalan O., Zunabovic M., Domig K. J., Kneifel W. (2011). Mini review: antimicrobial strategies in the production of fresh-cut lettuce products. Sci. against Microb. Pathogens Commun. Curr. Res. Technol. Adv. 1, 176–188. Google Scholar.
57. Wan Z., Chen Y., Pankaj S. K., Keener K. M. (2017). High voltage atmospheric cold plasma treatment of refrigerated chicken eggs for control of Salmonella Enteritidis contamination on egg shell. LWT - Food Sci. Technol. 76, 124–130. doi: 10.1016/j.lwt.2016.10.051. CrossRef. Google Scholar.
58. Wang F. (2017). Influence of long-term survival on resistance of Salmonella enterica serovar Typhimurium and Escherichia coli O157:H7 to physical or chemical food processes. ProQuest dissertations and theses. Google Scholar.
59. Wang F., Mendonca A., Brehm-Stecher B. F., Dickson J., Dispirito A., Shaw A. et al. (2018). Long-term survival phase cells of Salmonella Typhimurium ATCC 14028 have significantly greater resistance to ultraviolet radiation in 0.85% saline and apple juice. Foodborne Pathogens Dis. 15 (9), 538–543. doi: 10.1089/fpd.2018.2423. CrossRef. Google Scholar.
60. Walsh J. L., Iza F., Janson N. B., Law V. J., Kong M. G. (2010). Three distinct modes in a cold atmospheric pressure plasma jet. J. Phys. D: Appl. Phys. 43 (7), 075201. doi: 10.1088/0022-3727/43/7/075201. CrossRef. Google Scholar.
61. Wen J., Anantheswaran R. C., Knabel S. J. (2009). Changes in barotolerance, thermotolerance, and cellular morphology throughout the life cycle of Listeria monocytogenes. Appl. Environ. Microbiol. 75 (6), 1581–1588. doi: 10.1128/AEM.01942-08. CrossRef. Google Scholar.
62. Wen J., Deng X., Li Z., Dudley E. G., Anantheswaran R. C., Knabel S. J. et al. (2011). Transcriptomic response of Listeria monocytogenes during the transition to the long-term-survival phase. Appl. Environ. Microbiol. 77 (17), 5966–5972. doi: 10.1128/AEM.00596-11. CrossRef. Google Scholar.
63. Williams W. D. (1992). Origin and impact of color on consumer preference for food. Poult. Sci. 71 (4), 744–746. doi: 10.3382/ps.0710744. CrossRef. Google Scholar.
64. Yepez X. V., Keener K. M. (2016). High-voltage atmospheric cold plasma (HVACP) hydrogenation of soybean oil without trans-fatty acids. Innovative Food Sci. Emerging Technol. 38, 169–174. doi: 10.1016/j.ifset.2016.09.001. CrossRef. Google Scholar.
65. Yepez X. V., Misra N. N., Keener K. M. (2020). Nonthermal plasma technology. Food Saf. Eng., 607–628. doi: 10.1007/978-3-030-42660-6_23. CrossRef. Google Scholar.

Barry K, Mendonça A, Phillips GJ, Boylston T, Fortes-Da-Silva P, Brehm-Stecher B, Juneja V and Wan Z (2024) Long-term-survival phase cells of Salmonella enteritidis ATCC 13076 exhibit significantly greater tolerance to atmospheric cold plasma treatment of shell eggs. Front. Food. Sci. Technol. 4:1442761. doi: 10.3389/frfst.2024.1442761
Перевод статьи «Long-term-survival phase cells ofSalmonellaenteritidis ATCC 13076 exhibit significantly greater tolerance to atmospheric cold plasma treatment of shell eggs» авторов Barry K, Mendonça A, Phillips GJ, Boylston T, Fortes-Da-Silva P, Brehm-Stecher B, Juneja V and Wan Z., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык



















































































































Комментарии (0)