Сельскохозяйственные культуры для углеродного земледелия
В данной перспективной статье мы предлагаем разработку идеотипов культур с двойной функциональностью: быть высокопродуктивными для производства продуктов питания, кормов и топлива, а также обеспечивать больший вклад в почвенный углерод и улучшать подземную экологию.
Аннотация
Системы возделывания сельскохозяйственных культур и пастбища занимают треть пахотных земель мира и обладают потенциалом для значительного связывания атмосферного CO₂ в виде почвенного органического углерода (ПОУ), улучшая углеродный баланс почвы. Улучшенный углеродный баланс почвы служит двойной цели: способствует здоровью почвы, что поддерживает продуктивность культур, и формирует пул, из которого углерод может преобразовываться в трудноразлагаемые формы для долгосрочного хранения в качестве меры по смягчению последствий глобального потепления. В данной перспективной статье мы предлагаем разработку идеотипов культур с двойной функциональностью: быть высокопродуктивными для производства продуктов питания, кормов и топлива, а также обеспечивать больший вклад в почвенный углерод и улучшать подземную экологию. Мы выступаем за целостный подход, объединяющий систему «растение — микроорганизмы — почва», и предполагаем, что значительных улучшений в накоплении почвенного углерода можно достичь с помощью трехстороннего подхода: (1) создание растений с повышенной прочностью корней для усиления распределения углерода под землю; (2) балансирование увеличения подземного распределения углерода с повышением силы источника (source strength) для усиления фотосинтеза и накопления биомассы; и (3) создание микробных консорциумов почвы для повышения силы стока (sink strength) ризосферы и свойств, способствующих росту растений (PGP).
Основная часть
Мы предполагаем, что значительные инвестиции времени и ресурсов следует направить на разработку однолетних культур для углеродного земледелия. Эти культуры будут распределять (allocates) возрастающее количество углерода в репродуктивные органы-стоки (sinks) и в подземные хранилища для двойной цели: смягчения последствий глобального потепления из-за роста уровня атмосферного CO₂ и улучшения здоровья почвы для повышения продуктивности сельскохозяйственных культур. С точки зрения баланса массы углерода, это сводится к увеличению поступления и хранения углерода в почве и уменьшению его потерь для достижения чистого увеличения запасов почвенного углерода. Мы утверждаем, что этого можно достичь через всестороннее понимание потоков углерода и взаимодействий источник-сток в интегрированной (integrated) системе «растение-микроорганизм-почва». Наше обоснование фокусирования на однолетних культурах включает четыре аспекта: (1) по данным Геологической службы США, в мире насчитывается 18,6 млн км² (4,6 млрд акров) пахотных земель (U.S. Geological Survey (USGS), 2019), почти 80% из которых занято однолетними сельскохозяйственными культурами, т.е. зерновыми, зернобобовыми и масличными культурами (Glover и др., 2007); (2) в отличие от многолетних культур, которые требуют временных затрат в несколько лет, однолетние культуры более удобны для внедрения и применения новых и специально сконструированных сортов; (3) многолетние растения уже инвестируют значительную часть своих фотоассимилятов в корневую биомассу, и можно ожидать, что влияние перепроектирования многолетников на улучшение углеродного статуса почвы будет менее значительным (significance) по сравнению с усилиями в отношении однолетних культур; и (4) во многих случаях системы однолетнего возделывания (cropping systems) привели к образованию углеродного долга в почве и, следовательно, могут быть особенно восприимчивы к усилиям, направленным (aimed) на улучшение углеродного бюджета почвы (Griscom и др., 2017; Houghton и Nassikas, 2017; Sanderman и др., 2017).
Пул почвенного углерода объемом 2500 гигатонн (Гт; 1 Гт = 1 млрд метрических тонн) в верхних 3 м в 3,3 раза превышает размер атмосферного пула в 760 Гт и включает 1550 Гт почвенного органического углерода (ПОУ) и 950 Гт почвенного неорганического углерода (Lal, 2004a, 2008; Jansson и др., 2010; Рисунок 1). Пул ПОУ в первых 1 м и в верхних 20 см почвенного профиля составляет 1500 и 615 Гт соответственно (Sanderman и др., 2010; Guo и др., 2016). Основной обмен углеродом между атмосферой и наземной экосистемой представляет собой включение CO₂ в размере 123 Гт в год в биомассу растений посредством фотосинтеза, из которых 3 Гт в год образуется в результате антропогенной деятельности (DOE, 2008), и выделение CO₂ из ранее фиксированного углерода в результате дыхания растений и микроорганизмов в размере 60 и 60 Гт в год соответственно (Jansson и др., 2010; Abdullahi и др., 2018). В настоящее время наземные экосистемы являются чистым стоком углерода в размере 3 Гт в год, тем самым примерно буферизуя одну треть ежегодного увеличения концентрации атмосферного CO₂ от выбросов парниковых газов (Le Quere и др., 2018). Следовательно, большая часть CO₂, который фиксируется в виде фотоассимилятов, быстро возвращается в атмосферу, и лишь незначительная часть поступает в стабильный пул почвенного углерода. Таким образом, манипулирование углеродным бюджетом почвы, даже на несколько процентов, представляет собой значительный потенциал для смягчения последствий изменения климата (Paustian и др., 2016).
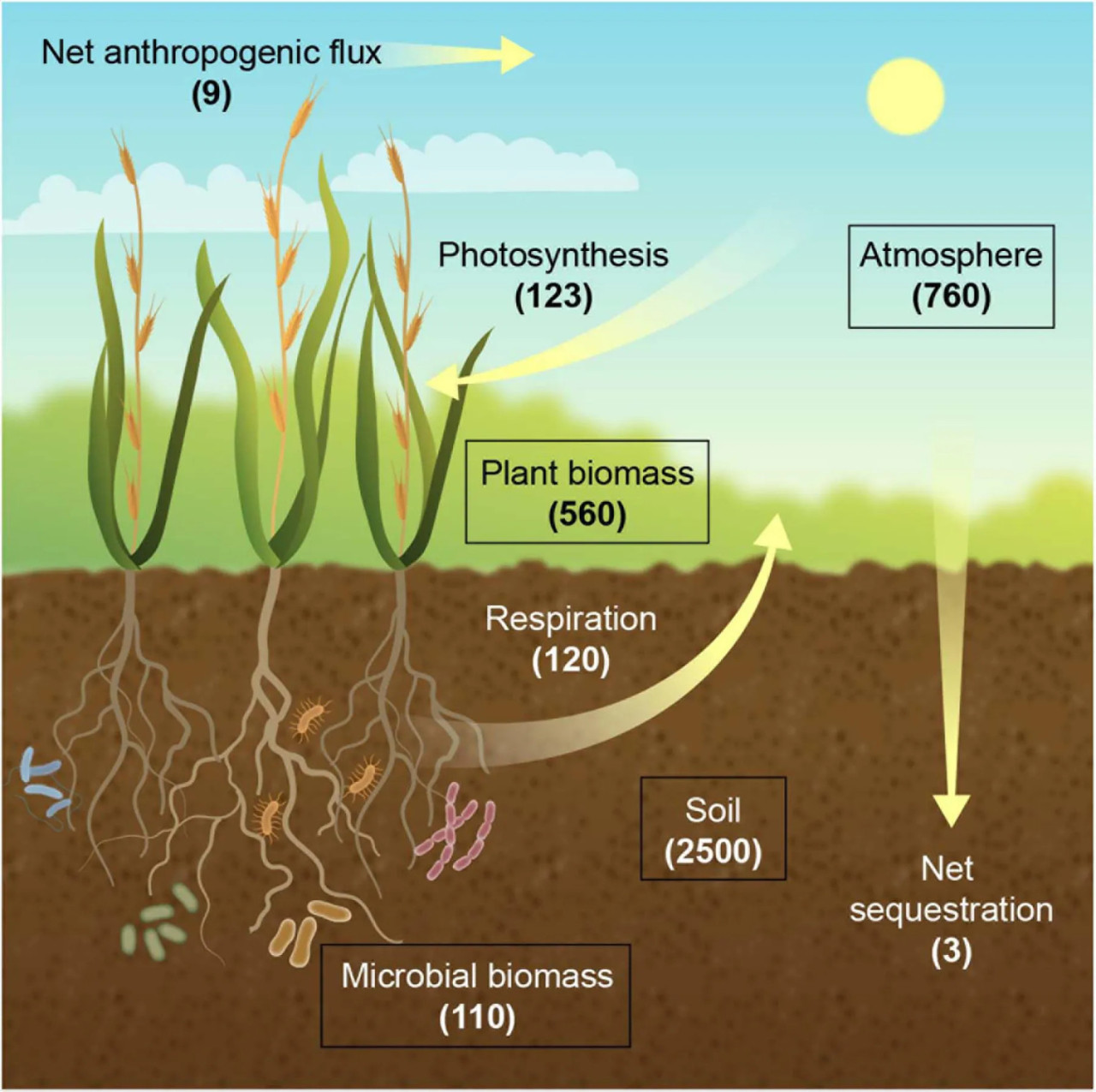
РИСУНОК 1 Наземный углеродный (C) цикл. Запасы углерода (прямоугольники) показаны в гигатоннах (Гт), а потоки (стрелки) показаны в Гт в год. Дыхание (Respiration) относится к сумме дыхания растений и микроорганизмов.
Агроэкосистемы занимают более одной трети пахотных земель мира и вносят 10–14% глобальных антропогенных выбросов парниковых газов, главным образом за счет кишечной ферментации (метан), применения синтетических удобрений (закись азота) и обработки почвы (CO₂) (Jantke и др., 2020). Однако сельскохозяйственные экосистемы также обладают потенциалом накапливать огромное количество почвенного углерода (Sanderman и Baldock, 2010; Abdullahi и др., 2018), до 1 Гт в год, что компенсировало бы около 10% годовых выбросов парниковых газов в размере 8–10 Гт в год. Как заявляет Институт углеродного цикла: «Сельское хозяйство — это ЕДИНСТВЕННЫЙ сектор, который способен трансформироваться из чистого источника CO₂ в чистый накопитель CO₂ — ни одна другая управляемая человеком сфера не обладает таким потенциалом» (The Carbon Cycle Institute, 2020). Существует множество методов управления земельными ресурсами, которые могут быть приняты для увеличения запасов почвенного углерода в агроэкосистемах, таких как изменение севооборотов, обработки почвы, управления удобрениями и органических мелиорантов (organic amendments) (Lal, 2004a). Возможно, наиболее эффективным средством для увеличения секвестрации почвенного углерода является изменение растительного покрова, например, перевод однолетних пахотных земель в леса или многолетние травы. Одним из предостережений (caveat) в отношении таких изменений в землепользовании является то, что они будут иметь негативные последствия для выхода биомассы с вытесняемых культур. Один практически неиспользованный альтернативный вариант — отбор и разработка однолетних сельскохозяйственных культур, которые распределяют (allocate) повышенное количество углерода в подземную биомассу, корневые экссудаты или ризодепозиты.
Почвенный органический углерод (как показатель почвенного органического вещества) играет две роли, когда мы решаем задачу достижения устойчивых агроэкосистем в ближайшие десятилетия: увеличивая продуктивность сельскохозяйственных культур и секвестрируя атмосферный углерод. ПОУ способствует продуктивности культур, улучшая удержание питательных веществ и влагоемкость, обеспечивая эффективный дренаж и аэрацию, минимизируя потерю верхнего слоя почвы из-за эрозии и предоставляя субстраты для почвенных микробиомов (Lal, 2004a; Sanderman и Baldock, 2010; Berazneva и др., 2019). ПОУ может секвестрироваться в устойчивых пулах, например, путем преобразования в биочар или через органо-минеральные и органо-металлические взаимодействия, со временем пребывания от десятилетий до тысяч лет и тысячелетий (Jansson и др., 2010; Sanderman и др., 2010; Abdullahi и др., 2018). ПОУ также может преобразовываться в неорганические соединения почвы, такие как карбонаты кальция и магния, для долгосрочного хранения (Guo и др., 2016). Рациональный дизайн, разработка и внедрение культур, адаптированных (tailored) для углеродного земледелия, будут частично зависеть от нашей способности моделировать метаболические потоки углерода и азота, понимать их контроль и впоследствии применять это понимание для реконфигурации взаимодействий источник-сток и путей распределения углерода в интегрированных (integrated) системах «растение-микроорганизм-почва» с помощью геномной инженерии и редактирования. В этой области существует множество текущих усилий, на которые можно опереться, таких как разработка моделей метаболических потоков (Sweetlove и Ratcliffe, 2011; Grafahrend-Belau и др., 2013; Lakshmanan и др., 2016; Töpfer и др., 2020) и метаболических сетей геномного масштаба (Matthews и Marshall-Colon, 2021).
Куда направляется углерод? – Анализ баланса массы
Схема потоков углерода в наземной экосистеме изображена на Рисунке 2. Баланс массы для углерода (и любого материала) в экосистеме может быть описан уравнением:
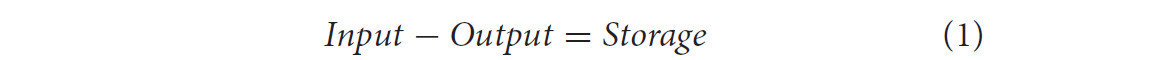
Для углерода уравнение (1) может быть обобщено следующим образом:
Входы: Фотосинтез; Почвенные мелиоранты (soil amendments)
Выходы: Дыхание растений и микроорганизмов; Выбросы летучих органических соединений (VOC)
Хранение: Биомасса растений и микроорганизмов; Почвенный углерод в виде пулов ПОУ и ПНУ (SIC)
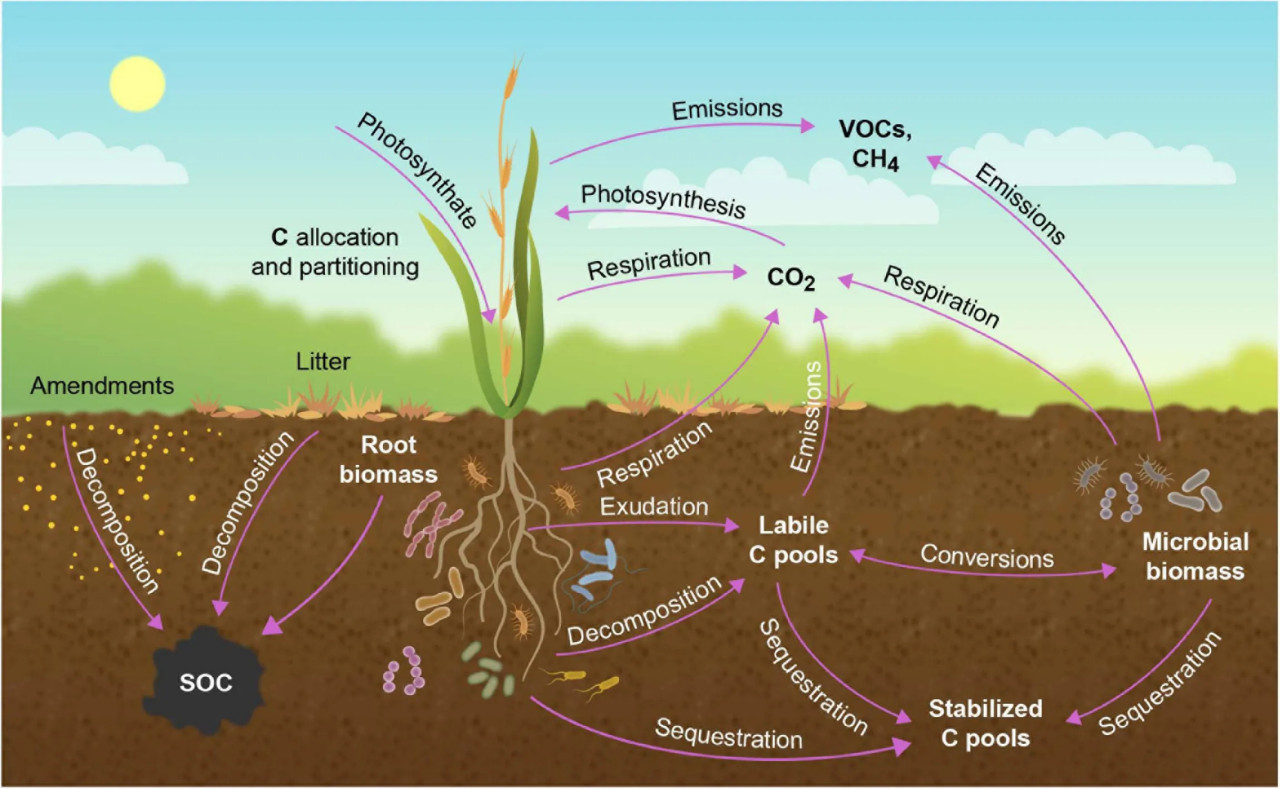
РИСУНОК 2 Перенос атмосферного CO₂ в биотические и педологические пулы углерода (C) в растительной экосистеме. Углерод поступает в почву в виде корневых экссудатов или через разложение корневой или надземной биомассы. В почве C существует в корневой или микробной биомассе, в виде биодоступного лабильного органического C или в виде более трудноразлагаемого (recalcitrant) C. Углерод покидает почву в виде прямых выбросов или через дыхание корней или микроорганизмов, при этом опосредованное микроорганизмами дыхание почвы является основным источником CO₂ из наземных экосистем. Углерод также теряется из экосистемы в виде летучих органических соединений (VOC) и метана (CH₄). Модифицировано по Jansson и др. (2018).
Содержание почвенного углерода определяется балансом между фотосинтетическими поступлениями через опад и корневую экссудацию и такими мелиорантами, как навоз и компост, и выходами через гетеротрофное дыхание корней и микроорганизмов и через выбросы VOC.
Распределение углерода является ключевым (crucial) физиологическим процессом, в ходе которого ассимилированный атмосферный CO₂ (фотоассимилят) перераспределяется между дыханием и производством биомассы, транзиторными и долгоживущими тканями, а также надземными и подземными компонентами. Функциональная, или оптимальная равновесная, теория утверждает, что растения распределяют ресурсы между органами для оптимизации приспособленности всего растения (Thornley, 1972; Bloom и др., 1985; Poorter и Nagel, 2000). Таким образом, распределение недавних фотоассимилятов между надземной и подземной биомассой для данного растения будет меняться в ответ на переменные окружающей среды, такие как влажность почвы, освещенность и доступность питательных веществ. Данные полевых исследований показывают (indicate), что растения распределяют относительно больше углерода в побеги при ограничении освещения и в корни при ограничении воды и/или питательных веществ.
Растения распределяют значительное количество (20–30%) недавних фотоассимилятов в свою подземную биомассу. Около 50% перемещенного углерода используется для роста корней, в то время как значительная часть этого углерода (до 30%) дополнительно высвобождается в ризосферу либо в виде прямого корневого отложения через экссудацию, слущивающиеся клетки корневого чехлика или через микоризу, либо теряется через дыхание (Lorenz и др., 2008; Turner и др., 2013; Kaiser и др., 2015). Как корневая экссудация, так и передача микоризным грибам происходят быстро после фотосинтеза — от нескольких часов у трав до нескольких дней у деревьев (Kaiser и др., 2015). Корневая экссудация стимулирует микробную декомпозицию SOM, что, в свою очередь, улучшает доступность питательных веществ вдоль ризосферы. Передача углерода микоризным грибам приносит пользу растению благодаря прямой передаче питательных веществ из сети грибных гиф. В обоих случаях инвестиции растения в подземное распределение углерода вознаграждаются повышенной доступностью питательных веществ, в частности азота и фосфора, а также повышенной толерантностью к абиотическим стрессам, таким как засуха, жара и засоление (Kaiser и др., 2015; Begum и др., 2019).
Значительная часть фотоассимилятов может высвобождаться в атмосферу в виде ЛОС (VOC), причем изопреноиды представляют собой доминирующий класс соединений (Guenther и др., 2012). Количество углерода, повторно выделяемого в виде изопрена (это единственное соединение составляет около половины всех выбросов изопреноидов), обычно составляет 1–3% от NPP, но может значительно (significantly) увеличиваться до 50% при неблагоприятных условиях окружающей среды, особенно когда поглощение углерода при фотосинтезе низкое (Loreto и Sharkey, 1993; Sharkey и Loreto, 1993; Harley и др., 1999). Таким образом, большая доля фиксированного углерода повторно выделяется в условиях умеренного стресса растений, в то время как общие выбросы уменьшаются при экстремальном стрессе (Holopainen и Gershenzon, 2010; Niinemets, 2010; Niinemets и др., 2013). Эти ЛОС высокореакционноспособны и легко участвуют в химии атмосферного окисления (Atkinson и Arey, 1998, 2003). Продукты окисления этих реакций подвергаются распределению между газовой и твердой фазами и способствуют образованию атмосферных твердых частиц, называемых вторичным органическим аэрозолем (SOA; Hallquist и др., 2009; Mohr и др., 2019). Атмосферные аэрозоли влияют на перенос излучения через атмосферу непосредственно, рассеивая и поглощая падающую солнечную радиацию, и косвенно, способствуя процессам облакообразования (Kerminen и др., 2005; Spracklen и др., 2008; Riipinen и др., 2012). Тип летучих веществ, выделяемых разными растениями в здоровом и стрессовом состоянии, может сильно различаться, что оказывает существенное влияние на образование SOA (Mentel и др., 2013; Joutsensaari и др., 2015; Yli-Pirilä и др., 2016; Zhao и др., 2017; Faiola и др., 2018, 2019). Это влияет на количество и характеристики света, доступного для использования растениями, включая соотношение рассеянного и прямого света, с последующим влиянием на NPP (Rap и др., 2018). Эти взаимосвязи схематично показаны на Рисунке 2 и подчеркивают (highlight) потенциальные механизмы обратной связи между системой почва-растение-атмосфера, опосредованные выбросами ЛОС и образованием аэрозолей. Значительное влияние образования вторичных аэрозолей, опосредованного ЛОС, на атмосферные процессы и здоровье человека было отмечено, например, при преобразовании сельскохозяйственных полей в растения для биотоплива, выделяющие изопрен (например, тополь) (Ashworth и др., 2013). Учет этих обратных связей при принятии решений об углеродном земледелии будет особенно важным в будущих климатических сценариях с усилением засухи, что может увеличить общую долю углерода, повторно выделяемого в виде ЛОС. Мы предостерегаем, что большинство исследований ЛОС на сегодняшний день проведено на многолетних растениях, и значимость обсуждаемых выше аспектов для однолетних культур, таких как сорго, кукуруза, пшеница и соя, необходимо оценить с помощью крупномасштабных исследований.
Усиление способности корней и ризосферы к поглощению питательных веществ.
Запасы почвенного углерода могут быть увеличены за счет повышения скорости поступления углерода в почву, увеличения удержания нового углерода, отложенного в почве, или за счет снижения скорости разложения ПОУ, уже присутствующего в почве. Потенциальный путь к увеличению запасов почвенного углерода — использование или разработка сортов сельскохозяйственных культур, которые вносят большее количество углерода в почву через свои корни. Подобно тому, как сила стока развивающихся зерновок является ключевым (key) фактором урожайности зерна, использование углерода для роста корней определяет силу стока корней и подземное накопление в виде ПОУ (Рисунок 3). Это может быть достигнуто за счет большей корневой биомассы или большей площади поверхности корней, которые активно выделяют углерод в почву. Например, сорта проса прутьевидного (switchgrass) с большей долей тонких корней и относительно большой удельной длиной корней увеличили поступление почвенного углерода в биоэнергетических системах возделывания (Adkins и др., 2016). Однако для долгосрочной секвестрации этого углерода важно, чтобы он удерживался в почве либо через ассоциации с почвенными минералами, либо через преобразование в карбонатные минералы или рекальцитрантный органический углерод, такой как древесный уголь, либо через снижение микробного дыхания, т.е. повышение эффективности использования углерода микроорганизмами. Выращивание более глубоких корневых систем представляет собой еще один путь, с помощью которого можно повысить поступление и удержание почвенного углерода. Углерод, отложенный на глубине, может иметь большее среднее время пребывания, поскольку скорость разложения в более глубоких почвенных профилях ниже по сравнению с поверхностными горизонтами, что способствует долгосрочному хранению почвенного углерода. Кроме того, более глубокие корни могут в некоторой степени смягчать последствия засух, тем самым дополнительно увеличивая поглощение углерода. Если такие растения будут разработаны, они могут быть быстро и в больших масштабах развернуты благодаря непрерывному генетическому обороту и активному управлению земельными ресурсами на сельскохозяйственных угодьях. Улучшение растений для увеличения секвестрации почвенного углерода представляет собой неиспользованный и экономичный чистый сток углерода со значительным экономическим потенциалом (Paustian и др., 2019a). Обоснование этой концепции было представлено Kell (2012). Кроме того, при сценариях повышения уровня атмосферного CO₂ из-за изменения климата, C3-культуры, которые обычно ограничены CO₂, могут вместо этого столкнуться с ограничением азота, что делает большую корневую биомассу еще более выгодной.
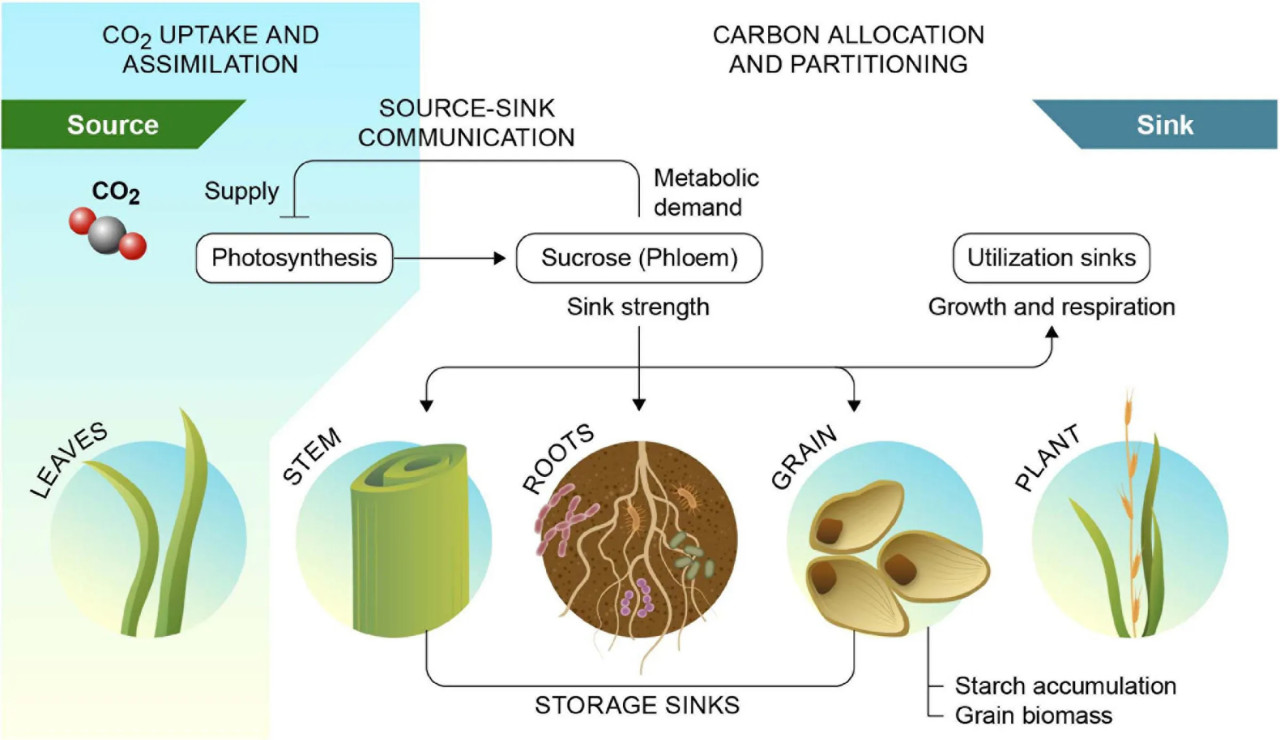
РИСУНОК 3 Взаимодействия источник-сток при производстве и использовании фотоассимилятов. Взаимодействия источник-сток связывают источники углерода, такие как зрелые листья, со стоками, такими как корни и семена, и опосредуют ингибирование фотосинтеза обратной связью через воспринимаемый спрос стока. Сила стока ризосферы складывается из корневой биомассы и связанных с ней микробных сообществ, включая арбускулярную микоризу. Перепечатано из Fan и др. (2008).
В анализе для поддержки новой программы, запущенной Министерством энергетики США, Paustian и др. (2016) выполнили «ограничивающий анализ» (bounding analysis) для оценки того, какой уровень увеличения почвенного углерода и общего смягчения выбросов парниковых газов (включая выбросы N₂O) возможен на основе достижимого увеличения общей корневой массы и изменения распределения глубины корней в сторону, характерную для многолетних трав. Они подсчитали, что широкое внедрение фенотипов однолетних культур, сконструированных для создания более глубоких и крупных корневых систем, может обеспечить увеличение запасов почвенного углерода на 0,5 Гт CO₂ га⁻¹ год⁻¹ на современных сельскохозяйственных угодьях США (Paustian и др., 2016, 2019a).
Хотя хорошо известно, что корневые системы играют важную роль в обеспечении и хранении ПОУ, какие именно характеристики корней важны для максимизации выигрыша в ПОУ и обеспечения долгосрочного хранения углерода, не очевидно. Высокое поступление углерода, поступающего из корней, является необходимым, но недостаточным условием для увеличения содержания углерода в почве. Например, большая корневая биомасса и корневая экссудация, стимулируемые более высоким уровнем CO₂, не обязательно приводят к высокому увеличению почвенного углерода — феномен, объясняемый усилением микробной активности и усилением праймирования (priming) старого почвенного органического вещества (Hungate и др., 1997; Pendall и др., 2004; Phillips и др., 2012) (однако см. Jastrow и др., 2005). Девятилетнее полевое испытание, сравнивающее две многолетние травянистые системы: монокультуру проса прутьевидного и высокобиоразнообразную сукцессионную растительность местного происхождения, показало (demonstrated), что, хотя корневая биомасса проса более чем в 10 раз превышала таковую местной растительности, улучшения уровней ПОУ под просом были заметно ниже (Kravchenko и др., 2019). Эти примеры показывают (suggest), что совсем не очевидно, что селекция на более высокую корневую биомассу является ответом на более быстрое и эффективное накопление ПОУ. Среди характеристик растений, которые следует рассматривать как потенциально ведущие к увеличению ПОУ, можно выделить: (1) физические характеристики архитектуры корневой системы — не только общая корневая биомасса, но и морфология корней, благоприятная для формирования структуры почвы (Bardgett и др., 2014; Postma и др., 2017; Kravchenko и др., 2019; Voges и др., 2019); (2) количества углерода, поступающего в почву в виде ризодепозитов во время роста корней и их активного функционирования (Kuzyakov и Domanski, 2000; Bengtson и др., 2012; Mwafulirwa и др., 2016); (3) химический состав корневых тканей и экссудатов (Keiluweit и др., 2015; Naveed и др., 2017); и (4) развитие ризосферного микробиома, способного преобразовывать корневой углеродный вход в защищенный ПОУ с большей эффективностью (Lange и др., 2015).
Однако, возможно, ни один из этих факторов сам по себе не имеет значения, но вместо этого для получения прироста ПОУ необходимо благоприятное сочетание всех (Yang и др., 2019). Как уже упоминалось выше, то, сколько углерода остается в почве, зависит от того, сколько вносится и сколько остается защищенным. Недавние исследования показали (revealed), что низкомолекулярные корневые экссудаты особенно важны для формирования и удержания ПОУ. Это связано с тем, что они способствуют образованию микробных остатков, а соединения микробного происхождения составляют большую долю стабильного ПОУ (Cotrufo и др., 2013; Kallenbach и др., 2016). Тем не менее, остается много вопросов относительно взаимодействий между корневой экссудацией, структурой микробного сообщества, его физиологией и, в конечном счете, влиянием на соотношение углерода, преобразованного в микробную биомассу, и количества углерода, выделяемого из почвы в виде CO₂. Идеальным сценарием может быть большое количество высококачественного углерода, поступающего из корней, сопровождающееся формированием порового пространства, благоприятного для защиты нового углерода от дальнейшего разложения (Kravchenko и др., 2019), т.е. большей гетерогенностью порового пространства (De Deyn и др., 2011).
Также необходимо учитывать, что даже в пределах одного вида/генотипа конкретный вклад отдельных растений может зависеть от их возраста, условий роста с точки зрения обеспечения питательными веществами и водой, наличия стрессов (Uren, 2007; Helliwell и др., 2019) и конкуренции с соседними растениями того же или других видов/генотипов (Fan и др., 2008). В многолетней травянистой растительности было показано, что дополнительность (complementarity) высокого растительного разнообразия очень полезна для хранения почвенного углерода; обработки с высоким разнообразием показали значительно (significantly) лучшие результаты, чем любые монокультуры (Yang и др., 2019), и биоразнообразие с точки зрения большего количества видов растений, по-видимому, было более важным, чем поступление биомассы растений (Steinbeiss и др., 2008). Это те аспекты, которые необходимо будет учитывать при принятии решения о том, куда направлять инженерные усилия по модификации корневых систем однолетних растений для повышения их способности защищать ПОУ.
В дополнение к разнообразию растений и морфологии корней, изменение количества и состава корневых экссудатов для увеличения ПОУ является привлекательной областью внимания по двум причинам. Во-первых, прогнозируемый контроль общего производства корневых экссудатов может напрямую влиять на взаимоотношения источник-сток между корнем и надземными органами растения. Во-вторых, состав корневых экссудатов влияет на сукцессию микробного сообщества ризосферы и, следовательно, предоставляет селекционерам способ потенциально направлять состав ризобиома культур в пользу микроорганизмов, которые являются лучшими стоками углерода, улучшают характеристики почвы или увеличивают долговечность ПОУ. Роль AM грибов как стоков углерода для растений хорошо документирована (Churland и Grayston, 2014; Gorzelak и др., 2015; Kaiser и др., 2015), и есть данные, что бактерии и другие члены ризосферного микробиома обеспечивают аналогичные функции (Trivedi и др., 2013; Kallenbach и др., 2016). Например, недавно было показано, что кумарины растительного происхождения в корневых экссудатах ограничивают рост определенных таксонов бактерий в ризобиоме (Voges и др., 2019). Таким образом, контроль общего производства корневых экссудатов как стратегия увеличения силы стока корней, а также состава экссудатов как стратегия влияния на динамику сообщества ризосферы должны быть изучены как вариант для распределения большего количества углерода в почву для долгосрочного хранения. Здесь смещение баланса состава ризосферного микробиома в пользу AMF может обеспечить необходимые преимущества как для урожайности, так и для хранения почвенного углерода (Berger, 2019; Zhang и др., 2019).
Хорошо документировано, что микробиом растений оказывает множество эффектов, способствующих росту растений (plant growth-promoting, PGP), которые приносят пользу растениям-хозяевам, таких как обеспечение повышенной толерантности к абиотическим и биотическим стрессам и улучшенное усвоение питательных веществ, включая азотфиксацию (Ahkami и др., 2017). Таким образом, адаптация (tailoring) ризосферного микробиома для сочетания увеличения силы стока с усиленными PGP-свойствами должна значительно (go a long way) способствовать достижению двойной цели: улучшению углеродного бюджета почвы и повышению продуктивности (Рисунок 4). Мы предполагаем, что, хотя разработка микробиомов с заданными функциональными свойствами будет использовать огромное существующее микробное разнообразие почвы, она также будет включать возможность использования геномной инженерии/редактирования для конструирования высокоспецифичных синтетических микробных сообществ (SynComs), которые являются облигатными симбионтами растения-хозяина, чтобы обеспечить стратегию биосдерживания. Дизайн и полезность SynComs для улучшения признаков растений недавно обсуждались в de Souza и др. (2020).
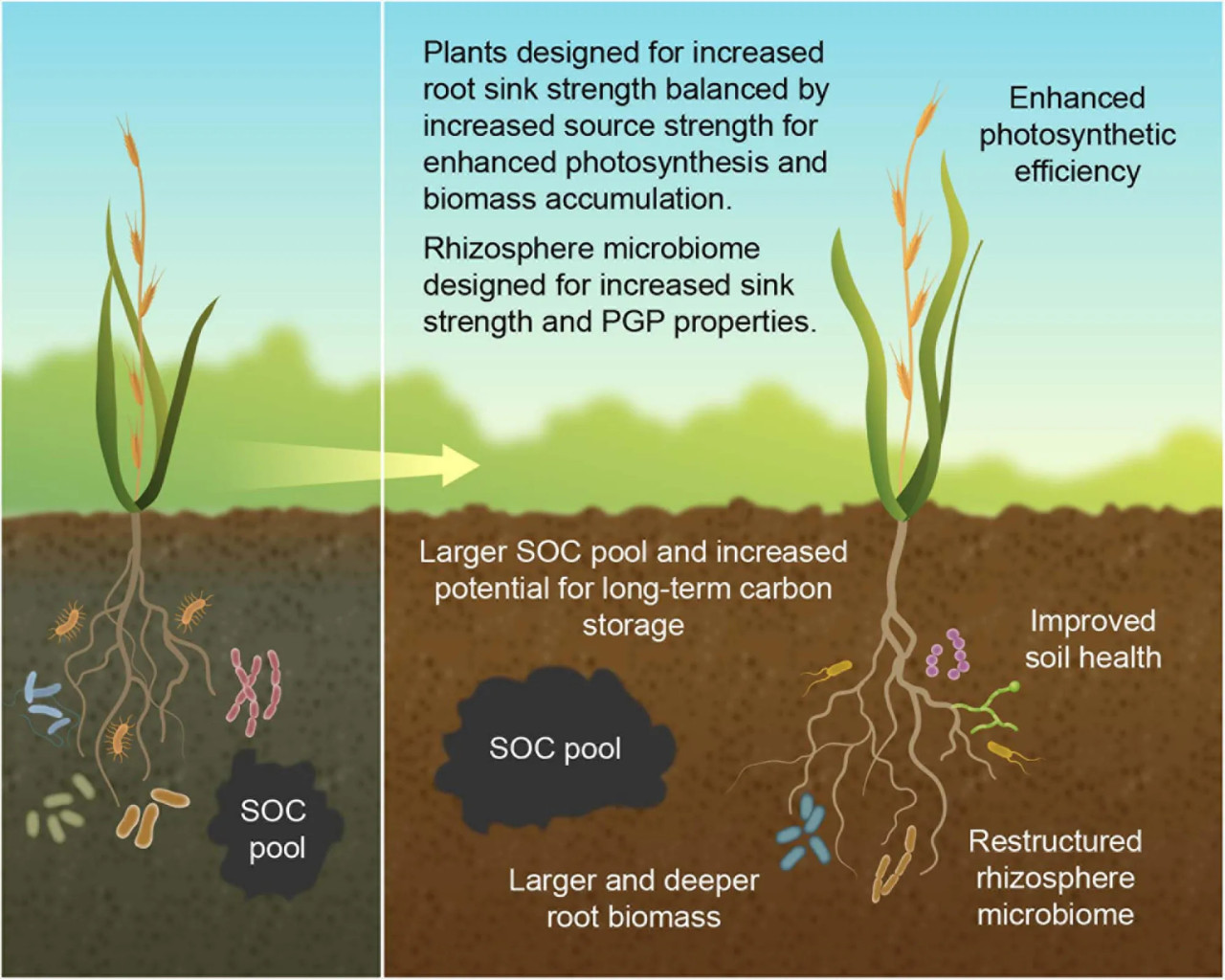
РИСУНОК 4 Обоснование разработки интегрированной системы «растение-микроорганизм-почва» с двойной целью: улучшение углеродного бюджета почвы при сохранении урожайности сельскохозяйственных культур, показывающее современную культуру (левая панель) и желаемый идеотип культуры (правая панель). Большая корневая биомасса обеспечивает повышенную силу стока, которая направляет больше углерода в почву, а более глубокие корни увеличивают вероятность долгосрочного хранения почвенного углерода. Индивидуально подобранные (custom-made) ризосферные микробиомы созданы для дальнейшего увеличения спроса на подземный углерод, тем самым увеличивая силу стока ризосферы, а также обеспечивая PGP-свойства. Чтобы дополнить стимулирование продуктивности растений за счет улучшенного здоровья почвы, PGP-микроорганизмов и усиленного фотосинтеза за счет повышенного спроса со стороны стока, растения также разработаны для увеличения силы источника (source strength) для дальнейшего улучшения фотосинтеза и накопления биомассы.
… Без ущерба для урожайности
Разработка культур для углеродного земледелия и улучшения углеродного бюджета почвы вряд ли станет жизнеспособным подходом, если фермеры не увидят финансовой отдачи. Хотя такая отдача может принимать форму налоговых льгот и программ ограничения и торговли выбросами (cap-and-trade), это не обеспечивает устойчивого долгосрочного решения. Хотя улучшение здоровья почвы за счет увеличения пула ПОУ и получение выгоды от усиленных PGP-эффектов синтетических ризосферных микробиомов в конечном итоге будет способствовать продуктивности растений, маловероятно, что это сохранит, не говоря уже об увеличении, урожайность культур у растений, созданных для направления значительной части фотоассимилятов в почву. Поэтому углеродное земледелие также должно будет включать меры по повышению эффективности преобразования фотосинтетической энергии и, следовательно, производства биомассы. Было изучено несколько таких вариантов (Ort и др., 2015; Kromdijk и др., 2016; Jansson и др., 2018; South и др., 2019). В частности, увеличение скорости восстановления от фотозащитного состояния привело к увеличению фотосинтеза и производства биомассы (Kromdijk и др., 2016). Аналогично, конструирование фотодыхательных обходных путей может привести к повышению уровня CO₂ вокруг Рубиско и снижению фотодыхательных потерь (South и др., 2019). Модификация биохимических и анатомических особенностей листьев и снижение сопротивления диффузии CO₂ к месту его фиксации вокруг Рубиско имеет потенциал для увеличения активности источника без увеличения потерь воды через устьица (Flexas и др., 2016). Увеличение скорости регуляции устьичной проводимости или активности цикла Кальвина-Бенсона при колеблющемся свете также может повысить эффективность фотосинтеза и эффективность использования воды культурами (Lawson и Blatt, 2014; Taylor и Long, 2017). В дополнение к этому, радикальные манипуляции с фотосинтетическим аппаратом, такие как конструирование механизмов концентрирования CO₂ (CCM), например, C4-фотосинтетического аппарата или цианобактериального карбоксисомного CCM, в C3-культурах также имеют большой потенциал для значительного увеличения активности источника (Long и др., 2015). Основное предположение здесь заключается в том, чтобы эти варианты были полезны для увеличения урожайности и корневой биомассы, повышенные скорости фотосинтеза не приведут к снижению распределения в корневые ткани. Действительно, когда рассматриваются растения одного функционального типа, получаются относительно консервативные соотношения корень:побег, хотя растения разных функциональных типов, или выращенные в разных условиях, или на разных стадиях роста показывают резко различные соотношения корень:побег (Mokany и др., 2006). Мы признаем, что наши знания о биохимических путях в растениях часто недостаточно полны, чтобы мы могли разумно предсказать последствия изменения экспрессии целевых генов. Здесь ненаправленные (open-ended) подходы генетики вперед (forward genetics) с помощью геномного редактирования могут быть полезны в качестве дополнения к рациональному дизайну (Zsögön и др., 2017; Andres и др., 2019; Belcher и др., 2020).
Рост и урожайность сельскохозяйственных культур ограничены не только фотосинтезом в зрелых листьях (сила источника), но также и характером распределения фотоассимилятов в другие органы, т.е. силой стока. Важно отметить, что если сила стока не поспевает за силой источника, это может привести к накоплению углеводов в листьях и ингибированию фотосинтеза обратной связью (White и др., 2016; Jansson и др., 2018; Рисунок 3). Поскольку повышенная концентрация атмосферного CO₂ увеличивает силу источника больше, чем силу стока, будет еще более важным сосредоточиться на повышении силы стока наряду с фотосинтезом при улучшении сельскохозяйственных культур (Chang и Zhu, 2017). Повышенной урожайности и улавливания углерода растениями можно было бы достичь за счет оптимизации регуляторных процессов, определяющих силу стока в гетеротрофных органах, в сочетании с преодолением ингибирования фотосинтеза обратной связью. Таким образом, вполне вероятно, что повышенная сила стока в корнях или ризосфере для направления большего количества углерода в почву может сочетаться с сохраненной или даже повышенной урожайностью за счет использования повышенных уровней атмосферного CO₂ и/или разобщения фотосинтеза от ингибирования обратной связью спросом стока. Это особенно верно для C3-культур, которые напрямую выиграют от повышенной силы источника за счет растущих концентраций CO₂.
При рассмотрении описанного выше сценария крайне важно понимать, что степень, в которой сельскохозяйственная культура ограничена источником или стоком, зависит от ее стадии развития и окружающей среды, а также варьируется между видами и генотипами. Однолетние растения обычно переходят от ограничения стоком к ограничению источником в ходе развития, когда они переключаются с вегетативного на репродуктивный рост (Arp, 1991; Burnett и др., 2016), хотя злаки, такие как пшеница и ячмень, могут оставаться ограниченными стоком во время размножения (Serrago и др., 2013). Многолетники, напротив, могут избегать ограничения стока и акклиматизации к повышенному CO₂ за счет пластичности развития (Burnett и др., 2016).
Еще одним важным аспектом является диверсификация севооборотов и годичного покрова. Долгосрочные эксперименты и недавние измерения вихревой ковариации показали противоречивые результаты относительно того, являются ли сельскохозяйственные угодья стоком или источником CO₂, которые варьируются между сортами и методами управления (Poyda и др., 2019). Поглощение углерода в течение вегетационного периода может компенсироваться более сильным гетеротрофным дыханием голой почвы после уборки урожая (Schmidt и др., 2012). Включение покровных культур (вместо того, чтобы оставлять пар в зимние месяцы) может увеличить запас ПОУ в почвах сельскохозяйственных угодий и, таким образом, быть еще одной эффективной мерой для компенсации выбросов CO₂. Таким образом, было показано, что возделывание покровных культур улучшает чистый углеродный баланс экосистемы, заменяя период чистого пара, когда углерод теряется за счет дыхания почвы, дополнительным периодом ассимиляции углерода (Lal, 2004b). Недавний мета-анализ оценил потенциальную глобальную секвестрацию ПОУ в 0,12 Пг C в год, что компенсировало бы 8% прямых годовых выбросов парниковых газов от сельского хозяйства (Poeplau и Don, 2015). Совместное выращивание (intercropping) часто применялось либо для улучшения питания почвы, например, с помощью азотфиксирующих растений, либо, наоборот, для обеспечения поглощения избыточного азота и снижения вымывания азота из почвы, а также для улучшения P и почвенных микробных сообществ. Таким образом, покровные культуры в основном исследовались на предмет их способности улучшать качество почвы (Hallama и др., 2019). Еще одна неиспользованная возможность для смягчения последствий изменения климата заключается в создании покровных культур с более глубокими корневыми системами и улучшенным распределением почвенного углерода, которую они обеспечивают.
Необходимость моделирования потоков
Желательна разработка комплексных, высокомеханистических системных моделей роста и развития сельскохозяйственных культур (Chang и Zhu, 2017) для руководства созданием новых интегрированных систем «культура-микроорганизм» или агрономических практик. Например, поскольку корни являются гетеротрофными органами, большая корневая биомасса еще больше увеличит уже значительное корневое дыхание. Способность точно прогнозировать потери углерода через дыхательные потоки становится важным инструментом при разработке культур. В более общем плане около 50% ассимилированных фотоассимилятов впоследствии теряется на дыхание (Amthor и др., 2019). Минимизация «неэссенциальной» дыхательной активности, ведущей к ненужному выделению CO₂, вряд ли подвергалась давлению отбора в процессе эволюции или учитывалась в программах селекции сельскохозяйственных культур, и сокращение этих больших потерь могло бы дополнить и усилить другие усилия по созданию культур с повышенной продуктивностью. Такие стратегии снижения дыхательных затрат могли бы включать: (1) замедление ненужного оборота белков; (2) замену, перемещение и/или изменение времени метаболической активности; (3) подавление бесполезных циклов; (4) повышение эффективности ионного транспорта (Amthor и др., 2019). Всестороннее понимание краткосрочных и долгосрочных потоков углерода (¹³C) и азота (¹⁵N) внутри модельного злака могло бы быть поучительным, показывая долю углерода, распределяемого в источники и стоки (зерно и корни), и процент, выделяемый в почву и атмосферу. Это интегрированное понимание движения углерода должно рассматриваться в связи с ростом растений, биомассой, урожайностью и продуктивностью растений. Эти данные послужат основой (blueprint) для понимания изменений в распределении углерода в зависимости от развития и окружающей среды, для начала разработки инженерных стратегий и для лучшей поддержки прогностических моделей. Например, для понимания влияния сниженного азота, повышенного CO₂, засухи и тепла, по отдельности и в различных комбинациях, на континуум растение-микроорганизм-окружающая среда-углерод. Другой пример, который необходимо выделить (highlight), — это необходимость измерения и прогнозирования потоков, связанных с выбросами N₂O (самого мощного биогенного парникового газа в расчете на единицу массы) в результате увеличения поступления азота, когда это необходимо для соответствия повышенным уровням атмосферного CO₂. Также требуются модели, которые учитывают стимуляцию фотосинтеза стоком для оценки того, в какой степени углеродные затраты, возникающие в рамках взаимодействий или симбиозов с ризосферным микробиомом, компенсируются усиленным фотосинтезом и производством биомассы (Kaschuk и др., 2009). Разработка прогностических метаболических моделей требует детальных карт метаболических потоков в различных органах и во всей интегрированной системе «растение-микроорганизм-почва-атмосфера» в различных условиях, а также того, как они изменяются при изменении внешних или внутренних условий. Это представляет собой крупную область исследований для будущей физиологии растений и молекулярной физиологии. Анализ метаболических потоков широко используется для изучения микробных сообществ, и он может быть адаптирован для получения всеобъемлющей карты переноса фотоассимилятов и соединений между различными органами сельскохозяйственных культур и других растений (Wiechert, 2001; Ma и др., 2014).
Заключительные замечания
Углеродное земледелие направлено (aims) на повышение скорости, с которой CO₂ удаляется из атмосферы и преобразуется в растительный материал и почвенное органическое вещество. Положительный результат этой амбиции двоякий: улучшенное здоровье почвы, которое, в свою очередь, способствует продуктивности сельскохозяйственных культур, и повышенный потенциал для долгосрочного хранения углерода для смягчения выбросов парниковых газов. В данной перспективной статье мы утверждаем, что культуры, созданные для углеродного земледелия, должны быть наделены следующими атрибутами: (1) повышенное подземное распределение углерода для большей и более глубокой корневой биомассы; (2) взаимодействие с адаптированным (tailored) синтетическим почвенным микробиомом для увеличения силы стока ризосферы и улучшенных PGP-свойств, которые облегчают усвоение питательных веществ и эффективность использования воды; и (3) повышенная сила источника для усиления фотосинтеза и накопления биомассы. Это представляет собой амбициозную инициативу, которая влечет за собой геномную инженерию/редактирование интегрированной системы «растение-микроорганизм-почва», поддерживаемую системным многоомиксным анализом, а также анализом и моделированием метаболических потоков. Также будет важно привлечь международные селекционные программы и использовать ресурсы из обширных коллекций зародышевой плазмы (Lenaerts и др., 2019; Voss-Fels и др., 2019). Это становится особенно актуальным в усилиях по интеграции и балансировке признаков для отложения почвенного углерода с урожайностью биомассы, устойчивостью к стрессам и способностью к восстановлению.
В заключение мы признаем, что хранение почвенного углерода не является бесконечным решением для сокращения выбросов парниковых газов, прежде всего потому, что почвы имеют верхний предел или уровень насыщения углеродом (Six и др., 2002; Paustian и др., 2019b). Мы осознаем проблемы, связанные с общественным признанием культур с отредактированным геномом и новых агрономических практик при внедрении культур для углеродного земледелия. Однако углеродное земледелие предоставляет возможность в ближайшие десятилетия использовать значительный потенциал, присущий сочетанию сельского хозяйства с ризосферным микробиомом для содействия секвестрации почвенного углерода. Таким образом, разработка культур для углеродного земледелия согласуется с консенсусом Парижского климатического соглашения, в котором говорится, что экономически оптимальные пути достижения парижской цели по ограничению глобального потепления требуют не только сокращения выбросов парниковых газов, но и должны включать технологии отрицательных выбросов, такие как стимулирование почвы к накоплению большего количества углерода (Economist, 2017).
Ссылки
1. Abdullahi A. C., Siwar C., Shaharudin M. I., Anizan I. (2018). Carbon Capture, Utilization and Sequestration, ed. Agarwal R. K. (London: IntechOpen). CrossRef. Google Scholar.
2. Adkins J., Jastrow J. D., Morris G. P., Six J., de Graaff M. A. (2016). Effects of switchgrass cultivars and intraspecific differences in root structure on soil carbon inputs and accumulation. Geoderma 262, 147–154. CrossRef. Google Scholar.
3. Ahkami A., White I., Handakumbura R. A., Jansson C. (2017). Rhizosphere engineering: enhancing sustainable plant ecosystem productivity in a challenging climate. Rhizosphere 3, 233–343. CrossRef. Google Scholar.
4. Amthor J. S., Bar-Even A., Hanson A. D., Millar A. H., Stitt M., Sweetlove L. J., et al. (2019). Engineering strategies to boost crop productivity by cutting respiratory carbon loss. Plant Cell 31, 297–314. Google Scholar.
5. Andres J., Blomeier T., Zurbriggen M. D. (2019). Synthetic switches and regulatory circuits in plants. Plant Physiol. 179, 862–884. CrossRef. Google Scholar.
6. Arp W. J. (1991). Effects of source-sink relations on photosynthetic acclimation to elevated CO₂. Plant Cell Environ. 14, 869–875. CrossRef. Google Scholar.
7. Ashworth K., Wild O., Hewitt C. N. (2013). Impacts of biofuel cultivation on mortality and crop yields. Nat. Clim. Change 3, 492–496. CrossRef. Google Scholar.
8. Atkinson R., Arey J. (1998). Atmospheric chemistry of biogenic organic compounds. Acc. Chem. Res. 31, 574–583. CrossRef. Google Scholar.
9. Atkinson R., Arey J. (2003). Atmospheric degradation of volatile organic compounds. Chem. Rev. 103, 4605–4638. CrossRef. Google Scholar.
10. Bardgett R. D., Mommer L., De Vries F. T. (2014). Going underground: root traits as drivers of ecosystem processes. Trends Ecol. Evol. 29, 692–699. CrossRef. Google Scholar.
11. Begum N., Qin C., Ahanger M. A., Raza S., Khan M. I., Ashraf M., et al. (2019). Role of arbuscular mycorrhizal fungi in plant growth regulation: implications in abiotic stress tolerance. Front. Plant Sci. 10:1068. CrossRef. Google Scholar.
12. Belcher M. S., Vuu K. M., Zhou A., Mansoori N., Ramos A. A., Thompson M. G., et al. (2020). Design of orthogonal regulatory systems for modulating gene expression in plants. Nat. Chem. Biol. 16:857. CrossRef. Google Scholar.
13. Bengtson P., Barker J., Grayston S. J. (2012). Evidence of a strong coupling between root exudation, C and N availability, and stimulated SOM decomposition caused by rhizosphere priming effects. Ecol. Evol. 2, 1843–1852. CrossRef. Google Scholar.
14. Berazneva J., Conrad J. M., Guerena D. T., Lehman J., Woolf D. (2019). Agricultural productivity and soil carbon dynamics: a bioeconomic model. Am. J. Agric. Econ. 101, 1021–1046. CrossRef. Google Scholar.
15. Berger J. J. (2019). Can soil microbes slow climate change? Sci. Am. Google Scholar.
16. Bloom A. J., Chapin F. S., Mooney H. A. (1985). Resource limitation in plants – an economic analogy. Annu. Rev. Ecol. Syst. 16, 363–392. CrossRef. Google Scholar.
17. Burnett A. C., Rogers A., Rees M., Osborne C. P. (2016). Carbon source-sink limitations differ between two species with contrasting growth strategies. Plant Cell Environ. 39, 2460–2472. CrossRef. Google Scholar.
18. Chang T.-G., Zhu X.-G. (2017). Source-sink interaction: a century old concept under the light of modern molecular systems biology. J. Exp. Bot. 68:1002. CrossRef. Google Scholar.
19. Churland C., Grayston S. J. (2014). Specificity of plant-microbe interactions in the tree mycorrhizosphere biome and consequences for soil C cycling. Front. Microbiol. 5:261. CrossRef. Google Scholar.
20. Cotrufo M. F., Wallenstein M. D., Boot C. M., Denef K., Paul E. (2013). The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: do labile plant inputs form stable soil organic matter? Glob. Change Biol. 19, 988–995. CrossRef. Google Scholar.
21. De Deyn G. B., Shiel R. S., Ostle N. J., McNamara N. P., Oakley S., Young I., et al. (2011). Additional carbon sequestration benefits of grassland diversity restoration. J. Appl. Ecol. 48, 600–608. CrossRef. Google Scholar.
22. de Souza R. S. C., Armanhi J. S. L., Arruda P. (2020). From microbiome to traits: designing synthetic microbial communities for improved crop resiliency. Front Plant Sci 11:1179. CrossRef. Google Scholar.
23. DOE (2008). Carbon Cycling and Biosequestration: Integrating Biology and Climate Through Systems Science. U.S. Department of Energy Office of Science. Google Scholar.
24. Economist T. (2017). Sucking up Carbon. Greenhouse Gases Must be Scrubbed From the Air. Google Scholar.
25. Faiola C. L., Buchholz A., Kari E., Yli-Pirilä P., Holopainen J. K., Kivimäenpää M., et al. (2018). Terpene composition complexity controls secondary organic aerosol yields from scots pine volatile emissions. Sci. Rep. 8:21045. CrossRef. Google Scholar.
26. Faiola C. L., Pullinen I., Buchholz A., Khalaj F., Ylisirnio A., Kari E., et al. (2019). Secondary organic aerosol formation from healthy and aphid-stressed scots pine emissions. ACS Earth Space Chem. 3, 1756–1772. CrossRef. Google Scholar.
27. Fan F. L., Zhang F. S., Qu Z., Lu Y. H. (2008). Plant carbon partitioning below ground in the presence of different neighboring species. Soil Biol. Biochem. 40, 2266–2272. CrossRef. Google Scholar.
28. Flexas J., Díaz-Espejo A., Conesa M. A., Coopman R. E., Douthe C., Gago J., et al. (2016). Mesophyll conductance to CO₂ and Rubisco as targets for improving intrinsic water use efficiency in C₃ plants. Plant Cell Environ. 39, 965–982. CrossRef. Google Scholar.
29. Glover J. D., Cox C. M., Reganold J. P. (2007). Future Farming: a return to roots? Sci. Am. 2007, 82–89. Google Scholar.
30. Gorzelak M. A., Assay A. K., Pickles B. J., Simard S. W. (2015). Inter-plant communication through mycorrhizal networks mediates complex adaptive behaviour in plant communities. AoB Plants 7:lv050. CrossRef. Google Scholar.
31. Grafahrend-Belau E., Junker A., Eschenroeder A., Muller J., Schreiber F., Junker B. (2013). Multiscale metabolic modeling: dynamic flux balance analysis on a whole-plant scale. Plant Physiol. 163, 637–647. CrossRef. Google Scholar.
32. Griscom B. W., Adams J., Ellis P. W., Houghton R. A., Lomax G., Miteva D. A., et al. (2017). Natural climate solutions. Proc. Natl. Acad. Sci. U.S.A. 114, 11645–11650. CrossRef. Google Scholar.
33. Guenther A. B., Jiang X., Heald C. L., Sakulyanontvittaya T., Duhl T., Emmons L. K., et al. (2012). The model of emissions of gases and aerosols from nature version 2.1 (MEGAN2.1): an extended and updated framework for modeling biogenic emissions. Geosci. Model Dev. 5, 1471–1492. CrossRef. Google Scholar.
34. Guo Y., Wang X. J., Li X. L., Wang J. P., Xu M. G., Li D. W., et al. (2016). Dynamics of soil organic and inorganic carbon in the cropland of upper Yellow River Delta, China. Sci. Rep. 6:36105. CrossRef. Google Scholar.
35. Hallama M., Pekrun C., Lambers H., Kandeler E. (2019). Hidden miners – the roles of cover crops and soil microorganisms in phosphorus cycling through agroecosystems. Plant Soil 434, 7–45. CrossRef. Google Scholar.
36. Hallquist M., Wenger J. C., Baltensperger U., Rudich Y., Simpson D., Claeys M., et al. (2009). The formation, properties and impact of secondary organic aerosol: current and emerging issues. Atmos. Chem. Phys. 9, 5155–5236. CrossRef. Google Scholar.
37. Harley P. C., Monson R. K., Lerdau M. T. (1999). Ecological and evolutionary aspects of isoprene emission from plants. Oecologia 118, 109–123. CrossRef. Google Scholar.
38. Helliwell J. R., Sturrock C. J., Miller A. J., Whalley W. R., Mooney S. J. (2019). The role of plant species and soil condition in the structural development of the rhizosphere. Plant Cell Environ. 42, 1974–1986. CrossRef. Google Scholar.
39. Holopainen J. K., Gershenzon J. (2010). Multiple stress factors and the emission of plant VOCs. Trends Plant Sci. 15, 176–184. CrossRef. Google Scholar.
40. Houghton R. A., Nassikas A. A. (2017). Global and regional fluxes of carbon from land use and land cover change 1850-2015. Glob. Biogeochem. Cycles 31, 456–472. CrossRef. Google Scholar.
41. Hungate B. A., Holland E. A., Jackson R. B., Chapin F. S., Mooney H. A., Field C. B. (1997). The fate of carbon in grasslands under carbon dioxide enrichment. Nature 388, 576–579. CrossRef. Google Scholar.
42. Jansson C., Vogel J., Hazen S., Brutnell T., Mockler T. (2018). Climate-smart crops with enhanced photosynthesis. J. Exp. Bot. 69, 3801–3809. CrossRef. Google Scholar.
43. Jansson C., Wullschleger S. D., Udaya C. K., Tuskan G. A. (2010). Phytosequestration: carbon biosequestration by plants and the prospects of genetic engineering. Bioscience 60, 685–696. CrossRef. Google Scholar.
44. Jantke K., Hartmann M. J., Rasche L., Blanz B., Schneider U. A. (2020). Agricultural greenhouse gas emissions: knowledge and positions of german farmers. Land 9:130. Google Scholar.
45. Jastrow J. D., Miller R. M., Matamala R., Norby R. J., Boutton T. W., Rice C. W., et al. (2005). Elevated atmospheric carbon dioxide increases soil carbon. Glob. Change Biol. 11, 2057–2064. CrossRef. Google Scholar.
46. Joutsensaari J., Yli-Pirilä P., Korhonen H., Arola A., Blande J. D., Heijari J., et al. (2015). Biotic stress accelerates formation of climate-relevant aerosols in boreal forests. Atmos Chem. Phys. 15, 12139–12157. CrossRef. Google Scholar.
47. Kaiser C., Kilburn M. R., Clode P. L., Fuchslueger L., Koranda M., Cliff J. B., et al. (2015). Exploring the transfer of recent plant photosynthates to soil microbes: mycorrhizal pathway vs direct root exudation. New Phytol. 205, 1537–1551. CrossRef. Google Scholar.
48. Kallenbach C. M., Frey S. D., Grandy A. S. (2016). Direct evidence for microbial-derived soil organic matter formation and its ecophysiological controls. Nat. Commun. 7:13630. Google Scholar.
49. Kaschuk G., Kuyper T. W., Leffelaar P. A., Hungria M., Giller K. E. (2009). Are the rates of photosynthesis stimulated by the carbon sink strength of rhizobial and arbuscular mycorrhizal symbioses? Soil Biol. Biochem. 41, 1233–1244. CrossRef. Google Scholar.
50. Keiluweit M., Bougoure J. J., Nico P. S., Pett-Ridge J., Weber P. K., Kleber M. (2015). Mineral protection of soil carbon counteracted by root exudates. Nat. Clim. Change 5, 588–595. Google Scholar.
51. Kell D. B. (2012). Large-scale sequestration of atmospheric carbon via plant roots in natural and agricultural ecosystems: why and how. Philos. Transact. R. Soc. B Biol. Sci. 367, 1589–1597. CrossRef. Google Scholar.
52. Kerminen V. M., Lihavainen H., Komppula M., Viisanen Y., Kulmala M. (2005). Direct observational evidence linking atmospheric aerosol formation and cloud droplet activation. Geophys. Res. Lett. 32:23130. CrossRef. Google Scholar.
53. Kravchenko A. N., Guber A. K., Razavi B. S., Koestel J., Quigley M. Y., Robertson G. P., et al. (2019). Microbial spatial footprint as a driver of soil carbon stabilization (vol 10, 3121, 2019). Nat. Commun. 10:12000. Google Scholar.
54. Kromdijk J., Głowacka K., Leonelli L., Gabilly S. T., Iwai M., Niyogi K. K., et al. (2016). Improving photosynthesis and crop productivity by accelerating recovery from photoprotection. Science 354:857. Google Scholar.
55. Kuzyakov Y., Domanski G. (2000). Carbon input by plants into the soil. Review. J. Plant Nutr. Soil Sci. 163, 421–431. CrossRef. Google Scholar.
56. Lakshmanan M., Cheung C. Y. M., Mohanty B., Lee D. Y. (2016). Modeling rice metabolism: from elucidating environmental effects on cellular phenotype to guiding crop improvement. Front. Plant Sci. 7:1795. CrossRef. Google Scholar.
57. Lal R. (2004a). Soil carbon sequestration impacts on global climate change and food security. Science 304, 1623–1627. Google Scholar.
58. Lal R. (2004b). Soil carbon sequestration to mitigate climate change. Geoderma 123, 1–22. CrossRef. Google Scholar.
59. Lal R. (2008). Sequestration of atmospheric CO₂ in global carbon pools. Energy Environ. Sci. 1, 86–100. CrossRef. Google Scholar.
60. Lange M., Eisenhauer N., Sierra C. A., Bessler H., Engels C., Griffiths R. I., et al. (2015). Plant diversity increases soil microbial activity and soil carbon storage. Nat. Commun. 6:7707. CrossRef. Google Scholar.
61. Lawson T., Blatt M. R. (2014). Stomatal size, speed, and responsiveness impact on photosynthesis and water use efficiency. Plant Physiol. 164, 1556–1570. CrossRef. Google Scholar.
62. Le Quéré C., Andrew R. M., Friedlingstein P., Sitch S., Hauck J., Pongratz J., et al. (2018). Global carbon budget 2018. Earth Syst. Sci. Data 10, 2141–2194. CrossRef. Google Scholar.
63. Lenaerts B., Collard B. C. Y., Demont M. (2019). Review: Improving global food security through accelerated plant breeding. Plant Sci. 287:110207. CrossRef. Google Scholar.
64. Long S. P., Marshall A. M., Zhu X. G. (2015). Engineering crop photosynthesis and yield potential to meet global food demand of 2050. Cell 161, 56–66. Google Scholar.
65. Lorenz K., Lal R., Shipitalo M. J. (2008). Chemical stabilization of organic carbon pools in particle size fractions in no-till and meadow soils. Biol. Fertil. Soils 44, 1043–1051. Google Scholar.
66. Loreto F., Sharkey T. D. (1993). Isoprene emission by plants is affected by transmissible wound signals. Plant Cell Environ. 16, 563–570. CrossRef. Google Scholar.
67. Ma F., Jazmin L. J., Young J. D., Allen D. K. (2014). Isotopically nonstationary 13C flux analysis of changes in Arabidopsis thaliana leaf metabolism due to high light acclimation. Proc. Natl. Acad. Sci. U.S.A. 111:16967. CrossRef. Google Scholar.
68. Matthews M. L., Marshall-Colon A. (2021). Multiscale plant modeling: from genome to phenome and beyond. Emerg. Top. Life Sci. 5:ETLS20200276. CrossRef. Google Scholar.
69. Mentel T. F., Kleist E., Andres S., Dal Maso M., Hohaus T., Kiendler-Scharr A., et al. (2013). Secondary aerosol formation from stress-induced biogenic emissions and possible climate feedbacks. Atmos Chem. Phys. 13, 8755–8770. CrossRef. Google Scholar.
70. Mohr C., Thornton J. A., Heitto A., Lopez-Hilfiker F. D., Lutz A., Riipinen I., et al. (2019). Molecular identification of organic vapors driving atmospheric nanoparticle growth. Nat. Commun. 10:12473. CrossRef. Google Scholar.
71. Mokany K., Raison R. J., Prokushkin A. S. (2006). Critical analysis of root : shoot ratios in terrestrial biomes. Glob. Change Biol. 12, 84–96. CrossRef. Google Scholar.
72. Mwafulirwa L., Baggs E. M., Russell J., George T., Morley N., Sim A., et al. (2016). Barley genotype influences stabilization of rhizodeposition-derived C and soil organic matter mineralization. Soil Biol. Biochem. 95, 60–69. CrossRef. Google Scholar.
73. Naveed M., Brown L. K., Raffan A. C., George T. S., Bengough A. G., Roose T., et al. (2017). Plant exudates may stabilize or weaken soil depending on species, origin and time. Eur. J. Soil Sci. 68, 806–816. CrossRef. Google Scholar.
74. Niinemets U. (2010). Mild versus severe stress and BVOCs: thresholds, priming and consequences. Trends Plant Sci. 15, 145–153. CrossRef. Google Scholar.
75. Niinemets U., Kannaste A., Copolovici L. (2013). Quantitative patterns between plant volatile emissions induced by biotic stresses and the degree of damage. Front. Plant Sci. 4:262. Google Scholar.
76. Ort D. R., Merchant S. S., Alric J., Barkan A., Blankenship R. E., Bock R., et al. (2015). Redesigning photosynthesis to sustainably meet global food and bioenergy demand. Proc. Natl. Acad. Sci. U.S.A. 112, 8529–8536. CrossRef. Google Scholar.
77. Paustian K., Campbell N., Dorich C., Marx E., Swan A. (2016). Assessment of Potential Greenhouse Gas Mitigation From Changes to Crop Root Mass and Architecture. Final Report for ARPA-E. Google Scholar.
78. Paustian K., Collier S., Baldock J., Burgess R., Creque J., DeLonge M., et al. (2019a). Quantifying carbon for agricultural soil management: from the current status toward a global soil information system. Carbon Manag. 10, 567–587. CrossRef. Google Scholar.
79. Paustian K., Larson E., Kent J., Marx E., Swan A. (2019b). Soil C sequestration as a biological negative emission strategy. Front. Climate 1:8. CrossRef. Google Scholar.
80. Pendall E., Mosier A. R., Morgan J. A. (2004). Rhizodeposition stimulated by elevated CO₂ in a semiarid grassland. New Phytol. 162, 447–458. CrossRef. Google Scholar.
81. Phillips R. P., Meier I. C., Bernhardt E. S., Grandy A. S., Wickings K., Finzi A. C. (2012). Roots and fungi accelerate carbon and nitrogen cycling in forests exposed to elevated CO₂. Ecol. Lett. 15, 1042–1049. CrossRef. Google Scholar.
82. Poeplau C., Don A. (2015). Carbon sequestration in agricultural soils via cultivation of cover crops – A meta-analysis. Agricult. Ecosyst. Environ. 200, 33–41. CrossRef. Google Scholar.
83. Poorter H., Nagel O. (2000). The role of biomass allocation in the growth response of plants to different levels of light, CO₂, nutrients and water: a quantitative review. Aust. J. Plant Physiol. 27, 595–607. CrossRef. Google Scholar.
84. Postma J. A., Kuppe C., Owen M. R., Mellor N., Griffiths M., Bennett M. J., et al. (2017). OPENSIMROOT: widening the scope and application of root architectural models. New Phytol. 215, 1274–1286. CrossRef. Google Scholar.
85. Poyda A., Wizemann H. D., Ingwersen J., Eshonkulov R., Hogy P., Demyan M. S., et al. (2019). Carbon fluxes and budgets of intensive crop rotations in two regional climates of southwest Germany. Agricult. Ecosyst. Environ. 276, 31–46. CrossRef. Google Scholar.
86. Rap A., Scott C. E., Reddington C. L., Mercado L., Ellis R. J., Garraway S. A., et al. (2018). Enhanced global primary production by biogenic aerosol via diffuse radiation fertilization. Nat. Geosci. 11:640. CrossRef. Google Scholar.
87. Riipinen I., Yli-Juuti T., Pierce J. R., Petaja T., Worsnop D. R., Kulmala M., et al. (2012). The contribution of organics to atmospheric nanoparticle growth. Nat. Geosci. 5, 453–458. CrossRef. Google Scholar.
88. Sanderman J., Baldock J. A. (2010). Accounting for soil carbon sequestration in national inventories: a soil scientist's perspective. Environ. Res. Lett. 5:34003. CrossRef. Google Scholar.
89. Sanderman J., Farquharson R., Baldock J. A. (2010). Soil Carbon Sequestration Potential: A Review for Australian Agriculture. CSIRO. Google Scholar.
90. Sanderman J., Hengl T., Fiske G. J. (2017). Soil carbon debt of 12,000 years of human land use. Proc. Natl. Acad. Sci. U.S.A. 114, 9575–9580. CrossRef. Google Scholar.
91. Schmidt M., Reichenau T. G., Fiener P., Schneider K. (2012). The carbon budget of a winter wheat field: an eddy covariance analysis of seasonal and inter-annual variability. Agricult. For. Meteorol. 165, 114–126. CrossRef. Google Scholar.
92. Serrago R. A., Alzueta I., Savin R., Slafer G. A. (2013). Understanding grain yield responses to source-sink ratios during grain filling in wheat and barley under contrasting environments. Field Crops Res. 150, 42–51. CrossRef. Google Scholar.
93. Sharkey T. D., Loreto F. (1993). Water-stress, temperature, and light effects on the capacity for isoprene emission and photosynthesis of kudzu leaves. Oecologia 95, 328–333. CrossRef. Google Scholar.
94. Six J., Conant R. T., Paul E. A., Paustian K. (2002). Stabilization mechanisms of soil organic matter: implications for C-saturation of soils. Plant Soil 241, 155–176. CrossRef. Google Scholar.
95. South P. F., Cavanagh A. P., Liu H. W., Ort D. R. (2019). Synthetic glycolate metabolism pathways stimulate crop growth and productivity in the field. Science 363:9077. CrossRef. Google Scholar.
96. Spracklen D. V., Carslaw K. S., Kulmala M., Kerminen V.-M., Sihto S.-L., Riipinen I., et al. (2008). Contribution of particle formation to global cloud condensation nuclei concentrations. Geophys. Res. Lett. 35:33038. CrossRef. Google Scholar.
97. Steinbeiss S., Bessler H., Engels C., Temperton V. M., Buchmann N., Roscher C., et al. (2008). Plant diversity positively affects short-term soil carbon storage in experimental grasslands. Glob. Change Biol. 14, 2937–2949. CrossRef. Google Scholar.
98. Sweetlove L. J., Ratcliffe R. G. (2011). Flux-balance modeling of plant metabolism. Front. Plant Sci. 2:38. CrossRef. Google Scholar.
99. Taylor S. H., Long S. P. (2017). Slow induction of photosynthesis on shade to sun transitions in wheat may cost at least 21% of productivity. Philos. Transact. R. Soc. B Biol. Sci. 372:20160543. CrossRef. Google Scholar.
100. The Carbon Cycle Institute (2020). Carbon Farming. Google Scholar.
101. Thornley J. H. (1972). Model to describe partitioning of photosynthate during vegetative plant-growth. Ann. Bot. 36, 419. Google Scholar.
102. Töpfer N., Braam T., Shameer S., Ratcliffe R. G., Sweetlove L. J. (2020). Alternative crassulacean acid metabolism modes provide environment-specific water-saving benefits in a leaf metabolic model. Plant Cell 32, 3689–3705. CrossRef. Google Scholar.
103. Trivedi P., Anderson I. C., Singh B. K. (2013). Microbial modulators of soil carbon storage: integrating genomic and metabolic knowledge for global prediction. Trends Microbiol. 21, 641–651. CrossRef. Google Scholar.
104. Turner T. R., James E. K., Poole P. S. (2013). The plant microbiome. Genome Biol. 14:209. Google Scholar.
105. U.S. Geological Survey (USGS) (2019). Global Food Security-Support Analysis Data at 30 m. Google Scholar.
106. Uren N. C. (2007). The Rhizosphere, eds Pinton R., Varanini Z., Nannipieri P. (Boca Raton, FL: CRC Press), 1–23. Google Scholar.
107. Voges M. J. E. E. E., Bai Y., Schulze-Lefert P., Sattely E. S. (2019). Plant-derived coumarins shape the composition of an Arabidopsis synthetic root microbiome. Proc. Natl. Acad. Sci. U.S.A. 116, 12558–12565. CrossRef. Google Scholar.
108. Voss-Fels K. P., Stahl A., Hickey L. T. (2019). Q&A: modern crop breeding for future food security. BMC Biol. 17:638. CrossRef. Google Scholar.
109. White A. C., Rogers A., Rees M., Osborne C. P. (2016). How can we make plants grow faster? A source-sink perspective on growth rate. J. Exp. Bot. 67, 31–45. CrossRef. Google Scholar.
110. Wiechert W. (2001). 13C metabolic flux analysis. Metab. Eng. 3, 195–206. CrossRef. Google Scholar.
111. Yang Y., Tilman D., Furey G., Lehman C. (2019). Soil carbon sequestration accelerated by restoration of grassland biodiversity. Nat. Commun. 10:8636. CrossRef. Google Scholar.
112. Yli-Pirilä P., Copolovici L., Kännaste A., Noe S., Blande J. D., Mikkonen S., et al. (2016). Herbivory by an outbreaking moth increases emissions of biogenic volatiles and leads to enhanced secondary organic aerosol formation capacity. Environ. Sci. Technol. 50, 11501–11510. CrossRef. Google Scholar.
113. Zhang H. Q., Liu T., Wang Y. Y., Tang M. (2019). Exogenous arbuscular mycorrhizal fungi increase soil organic carbon and change microbial community in poplar rhizosphere. Plant Soil Environ. 65, 152–158. CrossRef. Google Scholar.
114. Zhao D. F., Buchholz A., Tillmann R., Kleist E., Wu C., Rubach F., et al. (2017). Environmental conditions regulate the impact of plants on cloud formation. Nat. Commun. 8:14067. CrossRef. Google Scholar.
115. Zsögön A., Čermák T., Voytas D., Peres L. E. P. (2017). Genome editing as a tool to achieve the crop ideotype and de novo domestication of wild relatives: case study in tomato. Plant Sci. 256, 120–130. CrossRef. Google Scholar.
Jansson C, Faiola C, Wingler A, Zhu X-G, Kravchenko A, de Graaff M-A, Ogden AJ, Handakumbura PP, Werner C and Beckles DM (2021) Crops for Carbon Farming. Front. Plant Sci. 12:636709. doi: 10.3389/fpls.2021.636709
Перевод статьи «Crops for Carbon Farming» авторов ansson C, Faiola C, Wingler A, Zhu X-G, Kravchenko A, de Graaff M-A, Ogden AJ, Handakumbura PP, Werner C and Beckles DM., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык



















































































































Комментарии (0)