Разнообразие и функции микробиома ризосферы сои в условиях природного земледелия
Природное земледелие — это система возделывания культур без использования химических удобрений и пестицидов. В данном исследовании изучались бактериальные и грибковые сообщества в ризосфере сои, выращенной на почвах, обрабатываемых традиционным способом, и на почвах, обрабатываемых естественным способом, с использованием дикого типа и не образующих клубеньки мутантных сортов сои. Также был проанализирован эффект почвенного фумиганта, чтобы выяснить его влияние на микробные сообщества и последующее воздействие на рост сои.
Аннотация
В целом, дикий тип сои показал лучшие показатели роста по сравнению с мутантной соей, особенно в условиях природного земледелия. Образование клубеньков и колонизация арбускулярными микоризными грибами были выше у растений при природном земледелии, чем в почвах с традиционным управлением; однако фумигация резко повлияла на эти симбиозы, оказав большее воздействие на растения в почве природного земледелия.
Разнообразие микробиома ризосферы при природном земледелии было выше, чем при традиционном, для обоих сортов. Однако разнообразие значительно снизилось после обработки фумигантом, причем большее влияние наблюдалось именно в условиях природного земледелия. Анализ главных координат показал, что почвы при природном и традиционном земледелии содержат различные микробные сообщества, и фумигация почвы значительно изменила сообщества в почвах природного земледелия, но не в почвах традиционного.
Что интригует, некоторые полезные таксоны микробов, связанные с ростом и здоровьем растений, включая Rhizobium, Streptomyces и Burkholderia, были обнаружены как характерные микробы в почве при природном земледелии, но были избирательно уничтожены обработкой фумигантом. Сетевой анализ выявил высокосложную микробную сеть с высокой связностью таксонов в почве природного земледелия по сравнению с традиционной почвой; однако фумигация сильно разрушила эту сеть.
В целом, результаты подчеркнули, что природное земледелие способствует более высокому микробному разнообразию и обилию полезных почвенных микробов со сложной и взаимосвязанной сетевой структурой, а также продемонстрировали устойчивость микробного сообщества к воздействию внешних факторов, что имеет решающее значение в условиях природного земледелия, где химические удобрения и пестициды не применяются.
1. Введение
Сельское хозяйство имеет критическое значение для удовлетворения потребностей постоянно растущего населения Земли в продовольствии. С момента зеленой революции большинство методов управления в сельском хозяйстве полагались на использование химических удобрений и пестицидов для увеличения производства сельскохозяйственных культур. Использование этих агрохимикатов привело не только к повышению урожайности, но и к ряду экологических проблем, включая деградацию почв, выщелачивание агрохимикатов в поверхностные водоемы, появление устойчивых к пестицидам видов и потерю микробного биоразнообразия почвы, что привело к неустойчивой сельскохозяйственной практике (Singh and Mondal, 2017; Kumar et al., 2021). Следовательно, текущие методы ведения сельского хозяйства должны быть существенно изменены, чтобы сократить использование избыточных химикатов и их воздействие на окружающую среду, при этом сохраняя высокую урожайность.
Цель устойчивого сельского хозяйства — поддерживать биологическую функцию почвы и способствовать здоровью растений. «Природное земледелие», зародившееся в Японии, способствует достижению этой цели, используя такие методы, как севооборот и сидерацию, для поддержания плодородия почвы и роста культур вместо использования химических удобрений и пестицидов. Природное земледелие отличается от других систем органического земледелия тем, что оно строго запрещает использование необработанного навоза животных и городских сточных вод, поскольку неправильно компостированные сточные воды и навоз (Martin, 2005) могут быть источником загрязнения патогенами и тяжелыми металлами для сельскохозяйственной продукции, предназначенной для потребления человеком (Xu, 2001). В результате, по сравнению с традиционным земледелием, природное земледелие является более устойчивым, экономически эффективным, экологически безопасным и улучшает плодородие почвы и биоразнообразие.
В условиях систем природного земледелия здоровье и продуктивность растений в значительной степени зависят от микробиоты корней. Растения получают выгоду от ассоциированных с корнями микробов различными способами, включая усвоение питательных веществ, устойчивость к патогенам и толерантность к стрессу (Bulgarelli et al., 2013). Ризосфера — это непосредственная узкая зона, окружающая корни растений, которая заселена огромной популяцией микробов (Hartmann et al., 2008). Некоторые ассоциированные с корнями бактерии и грибы были идентифицированы как микроорганизмы, способствующие росту растений, которые играют разнообразные важные роли в регулировании роста и урожайности растений. Рикобии являются наиболее известными мутуалистическими бактериальными симбионтами, способными фиксировать атмосферный азот в корнях бобовых растений через образование клубеньков (Sugiyama, 2019). Аналогично, арбускулярные микоризные грибы (АМ грибы) являются наиболее распространенными мутуалистическими грибными симбионтами, которые устанавливают симбиоз с большинством наземных растений (Smith et al., 2011), принося пользу растениям-хозяевам разнообразными способами, включая обеспечение неорганическим фосфором и повышение толерантности растений к стрессу и болезням (Berendsen et al., 2012; Haney et al., 2015; Li et al., 2021). В последнее время некоторые исследования оценили влияние внесения удобрений на структуру микробного сообщества в условиях традиционного и природного земледелия (Liao et al., 2019; Sinong et al., 2021). Эти исследования показали более высокое обилие и разнообразие микробов в почвах природного земледелия по сравнению с почвами традиционного земледелия (Liao et al., 2019; Sinong et al., 2021). Несмотря на эти фундаментальные работы, мало известно о составе и функциях ассоциированных с корнями микробных сообществ, связанных с растениями в условиях систем природного земледелия, что требует дальнейших исследований функций микробных сообществ в почвах природного земледелия. Например, ни одно исследование не изучало микробиом ризосферы сои природного земледелия в Японии, несмотря на то, что соя является одной из наиболее возделываемых культур в системе природного земледелия.
Фумигация почвы широко используется для борьбы с почвенными патогенами и сорняками в различных системах земледелия (Strauss and Kluepfel, 2015). Хлорпикрин обычно используется в качестве почвенного фумиганта после отказа от бромистого метила во многих развитых странах (Mao et al., 2014). Хотя хлорпикрин показал перспективную биоцидную активность, такие фумиганты являются неселективными биоцидами с губительным воздействием на нецелевые организмы, включая полезные почвенные микроорганизмы. Очень мало известно о влиянии фумигантов на производство бобовых, особенно на симбиотическую ассоциацию бобовых с ризобиями, микоризными грибами и другими полезными почвенными симбионтами. Оценка воздействия почвенных фумигантов на симбиотические системы предоставит важную фундаментальную информацию для долгосрочного и эффективного использования полезных почвенных микроорганизмов в сельскохозяйственном производстве.
Настоящее исследование проверяло гипотезу о том, что в условиях природного земледелия существует отличный и разнообразный микробиом ризосферы, способствующий росту растений, с использованием сои в качестве модельной системы. Также было исследовано влияние почвенного фумиганта на микробиом ризосферы сои. С помощью высокопроизводительного секвенирования маркерных генов 16S рРНК и ITS было оценено бактериальное и грибное сообщество ризосферы сои в условиях природного земледелия и проведено его сравнение с системами традиционного земледелия.
2. Материалы и методы
2.1. Отбор почвенных образцов и фумигация хлорпикрином
Почва природного земледелия была собрана 5 июня 2021 года на экспериментальном поле Международного исследовательского центра природного земледелия (Нагано, Япония, 36°11′57.5″N 137°52′27.7″E), где непрерывно практикуется отказ от химических удобрений и пестицидов более 10 лет. Почва традиционного земледелия была собрана 6 июня 2021 года на экспериментальном поле Токийского университета сельского хозяйства и технологий (Токио, Япония, 35°41′02.3″N 139°29′03.9″E). Обе почвы классифицируются как андосоли по японской системе классификации почв (Amano, 1985). Образцы почвы отбирались с верхних 15 см. Аликвоты почвы высушивались на воздухе и пропускались через сито с ячейкой 2 мм для получения тонкозема для рутинного физико-химического анализа почв. Каждый образец полевой почвы был разделен на две части: обрабатываемую фумигантом и необрабатываемую. Хлорпикрин (Mitsui Chemicals Agro Co., Ltd., Токио, Япония) использовался в качестве фумиганта и вносился в виде водного раствора в рекомендуемой дозе 12 мл на 6 кг почвы. Почва герметизировалась в пластиковом пакете и перемешивалась каждые 2 дня в течение 10 дней, а затем открывалась для аэрации на 2 дня. Приблизительно 2,5 л почвы использовалось для заполнения сосудов Вагнера объемом 1/5000 для выращивания сои.
2.2. Химический анализ почвы
pH почвы и электрическая проводимость (ЕС) измерялись в 1 М KCl и деионизированной воде при соотношении почва:раствор 1:5. Общий углерод (TC) и общий азот (TN) измерялись методом сухого сжигания на NC-анализаторе (SUMIGRAPH NC TR22, Sumika Chemical Analysis Service Ltd., Осака, Япония). Неорганический азот в почве экстрагировали 2 М KCl, N-NH4+ в экстракте анализировали модифицированным индофенольным методом, а N-NO3- в экстракте анализировали методом проточного инжекционного анализа с использованием проточного видимого спектрофотометра, оснащенного меднокадмиевой колонкой для восстановления нитрата до нитрита в растворах образцов. Доступный фосфат (Available-P) оценивался методом Брея 2.
2.3. Выращивание сои
Два сорта сои (Glycine max L.), Enrei (дикий тип) и его немутирующий мутант En1282, были использованы в этом эксперименте. Все семена сои поверхностно стерилизовали погружением в 20 мл раствора каплата (Mitsui Chemicals Agro Co., Ltd., Токио, Япония) (10 мг/мл) на 1 час при 28°C. Затем семена несколько раз промывали стерильной дистиллированной водой, помещали на стерильную влажную бумажную салфетку и инкубировали в течение 72 часов при 25°C в темноте для проращивания. Проросшие семена высевали непосредственно в подготовленные сосуды по два семени на сосуд с 12 повторностями. Растения выращивали в фитотроне (20–25°C день/ночь) и регулярно поливали. Значения SPAD и длину стебля измеряли еженедельно с 3-й по 10-ю неделю после посадки. Медный фунгицид, бордоская смесь (Nihon Nohyaku Co., Ltd., Япония), разводили в 500 раз и опрыскивали листья сои, выращиваемой на фумигированных почвах, через 3 недели после посадки.
2.4. Сбор ризосферной почвы
Через 10 недель после посева по четыре растения из каждой обработки были собраны для анализа микробиома ризосферы, в то время как остальные растения выращивались в течение 12 недель. Растения сои осторожно удаляли из сосудов, и почву, слабо прикрепленную к корням, удаляли легким встряхиванием. Клубеньки отделяли от корней и подсчитывали, а также измеряли свежую массу корней. Ризосферную почву сои собирали согласно модифицированной процедуре McPherson и др. (2018). Кратко: 2 г корней с плотно связанной почвой помещали в 50-мл центрифужную пробирку, заполненную 30 мл автоклавированного фосфатного буфера (г/л: 6,33 г NaH2PO4.H2O, 16,5 г Na2HPO4.7H2O, 200 мкл поверхностно-активного вещества, pH 6,5). Пробирку встряхивали на максимальной скорости (об/мин) в течение 2 минут, и суспензию фильтровали через нейлоновый фильтр с ячейкой 100 мкм в новую 50-мл пробирку. Затем суспензию центрифугировали при 3000×g в течение 5 минут для осаждения почвенных частиц. Осадок ресуспендировали в 1,5 мл фосфатного буфера (без поверхностно-активного вещества) и затем переносили в 2-мл микроцентрифужную пробирку. Пробирку центрифугировали при 15871×g в течение 2 минут, после чего супернатант сливали, а осадок почвы хранили при -20°C до дальнейшей обработки.
2.5. Оценка колонизации АМ грибами
Для определения степени колонизации АМ грибами корни, использованные для сбора ризосферной почвы, помещали в 50-мл пробирки. Корни осветляли 10% KOH на водяной бане в течение 15 минут, подкисляли 5% HCl и окрашивали 0,05% трипановым синим в лактоглицерине (Phillips and Hayman, 1970). Десять фрагментов корня (длиной 1 см) на обработку монтировали на предметные стекла в трех повторностях для оценки морфологических структур АМ грибов в корнях, которые включали гифы, везикулы и арбускулы, следуя процедуре McGonigle и др. (1990).
2.6. Экстракция ДНК из почвы и ампликонный анализ
ДНК ризосферной почвы экстрагировали с использованием набора Nucleospin soil kit (MACHEREY-NAGEL, Дюрен, Германия) следуя протоколу производителя. Ампликонное секвенирование бактериальной области V3/V4 и грибной области ITS проводилось с использованием платформы Miseq в Bioengineering Lab. Co. (Сагамихара, Япония). Бактериальная область V4 амплифицировалась с использованием прямого праймера 1st_PCR_515F_MIX:(5′-ACACTCTTTCCCTACACGACGCTCTTCCGATCT-NNNNN-GTGCCAGCMGCCGCGGTAA-3′) и обратного праймера 1st_PCR_806R_MIX:(5′-GTGACTGGAGTTCAGACGTGTGCTCTTCCGATCT-NNNNN-GGACTACHVGGGTWTCTAAT-3′), и внутренний транскрибируемый спейсер (ITS) 1 область для грибов амплифицировали с использованием прямого праймера ITS1F_KYO1 (5′ACACTCTTTCCCTACACGACGCTCTTCCGATCTCTHGGTCATTTAGAGGAASTAA3′) и обратного праймера ITS2_KYO2 (5′GTGACTGGAGTTCAGACGTGTGCTCTTCCGATCTTTRCTRCGTTCTTCATC-3′) с помощью ПЦР. ПЦР-реакция, содержащая матрицу ДНК (5 нг), проводилась в конечном объеме 20 мкл с использованием набора ExTaq polymerase kit (Takara). Продукты ПЦР очищали с использованием AMPure XP (Beckman coulter) и количественно определяли с использованием Synergy H1 (Bio Tek) и системы QuantiFlour dsDNA. Контроль качества библиотек проводили с использованием Fragment Analyzer и набора реактивов sdDNA 915 Reagent Kit (Advanced Analytical Technologies). Библиотеки объединяли и загружали в прибор Illumina MiSeq в соответствии с инструкциями производителя (Illumina, Сан-Диего, Калифорния, США).
2.7. Биоинформатика
Набор инструментов Quantitative Insights into Microbial Ecology (Qiime 2.0) использовался для обработки необработанных данных высокопроизводительного секвенирования. Затем данные, полученные с ампликонов 16S и ITS, были проанализированы с использованием MicrobiomeAnalyst (Dhariwal и др., 2017), а OTU были аннотированы как метки SILVA.
2.8. Обработка данных секвенирования и идентификация вариантов ампликонной последовательности
Биоинформатический анализ микробиома и идентификация были проведены с помощью QIIME2 2020.8 (Bolyen и др., 2019). Адаптеры и праймеры были удалены с помощью Cutadapt v2.4 из непроцессированных ридов (Martin, 2011). Последовательности были демультиплексированы с использованием плагина q2-demux с последующим контролем качества, усечением длины, денизингом, удалением химер и PhiX и построением таблицы признаков с помощью DADA2 с настройками по умолчанию, за исключением «-ptrunc-len-f» и «-p-trunc-len-r», которые были установлены на 250 и 200 соответственно для данных 16S и на 160 и 200 соответственно для данных ITS (через q2-dada2; Callahan и др., 2016). Полученные ASV были выровнены с помощью mafft (Katoh и др., 2002). Таксономия была присвоена ASV с использованием qiime feature-classifier classify-sklearn (Bokulich и др., 2018) с предварительно обученной наивной байесовской классификацией SILVA v132, усеченной до области V4 гена 16S рДНК (Quast и др., 2012) для бактерий, и предварительно обученной базой данных UNITE ver8 99% (UNITE Community, 2019), обученной на полных референсных последовательностях без какой-либо экстракции для грибов. Небактериальные и негрибные риды были удалены из полученной таблицы ASV. Мы нормализовали библиотеку, используя масштабирование с ранжированной субдискретизацией с помощью функции «SRS» в «SRS» с «qiime srs SRS» (Beule and Karlovsky, 2020). Различия в разнообразии микробных сообществ оценивали как индекс разнообразия Шеннона в qiime2, и для сравнения разнообразия использовали критерий Краскела-Уоллиса. Боксплоты для отображения показателей альфа-разнообразия были созданы с использованием ggplot2 (Wickham и др., 2016), установленного в R Core Team (2020). Матрица несходства Брея-Кертиса была измерена для бактериальных и грибных сообществ и использована для ординационной диаграммы PCoA в PAST4.04 (Hammer и др., 2001). Мы сравнили различия в составе и структуре сообществ между образцами (β-разнообразие), используя анализ сходства (ANOSIM; Clarke, 1993) и пермутационный многомерный дисперсионный анализ (PERMANOVA; Anderson, 2014). Наименьший дискриминантный анализ был выполнен с использованием microbiomeAnalyst (Chong и др., 2020). Функции бактерий были предсказаны с помощью программного обеспечения PICRUSt2 (ver. 2.3.0 b) на основе функциональной базы данных KEGG (Douglas и др., 2020).
2.9. Статистический анализ
Дисперсионный анализ (ANOVA) использовался для определения того, где средние значения обработок значительно отличаются друг от друга (p < 0,05), и тест SNK использовался для сравнения наборов средних. Вычисления проводились с использованием подпрограмм, встроенных в программное обеспечение Microsoft Excel ystat2013.
3. Результаты
3.1. Химические свойства почвы
Химический анализ почвы был выполнен на образцах, отобранных с полей традиционного и природного земледелия до выращивания сельскохозяйственных культур. Обе почвы были характерно кислыми, при этом почва природного земледелия показала немного более низкий pH, чем почва традиционного земледелия (Supplementary Table 1). Содержание общего углерода и общего азота, а также уровни аммонийного азота (NH4+-N) были выше в почве традиционного земледелия, в то время как содержание доступного P и уровни нитратного азота (NO3−-N) были выше в почве природного земледелия (Supplementary Table 1).
3.2. Ростовая производительность сои
Что касается длины побега, не было обнаружено значительных различий в средних значениях между растениями дикого типа, выращенными в почвах традиционного и природного земледелия, независимо от обработки фумигантом (Table 1). Аналогично, независимо от системы земледелия или обработки фумигантом, длины побегов мутантных растений не показали значительных различий; однако все они были значительно ниже по сравнению с диким типом (Table 1), что указывает на вклад азота, обеспечиваемого азотфиксирующими клубеньками. Не было обнаружено значительных различий в свежей массе корней всех обработок, за исключением мутантов, выращенных на нефумигированной традиционной почве (Table 1). В почве традиционного земледелия обработки хлорпикрином не изменили значительно рост и урожайность сои дикого типа; однако в почве природного земледелия обработки хлорпикрином значительно снизили количество семян и семенных бобов у сои дикого типа (Table 1), что позволяет предположить, что фумигация оказала большее влияние на азотфиксирующие ризобии в почве природного земледелия по сравнению с почвой традиционного земледелия. В почве традиционного земледелия обработки хлорпикрином увеличили количество семян, особенно у немутирующего мутанта сои (Table 1), вероятно, потому что микробное разложение хлорпикрином увеличило количество доступного азота. Напротив, в почве природного земледелия обработка хлорпикрином значительно снизила урожайность немутирующего мутанта сои (Table 1), что предполагает, что микробы, отличные от ризобий, в значительной степени способствуют урожайности сои в этой системе земледелия.
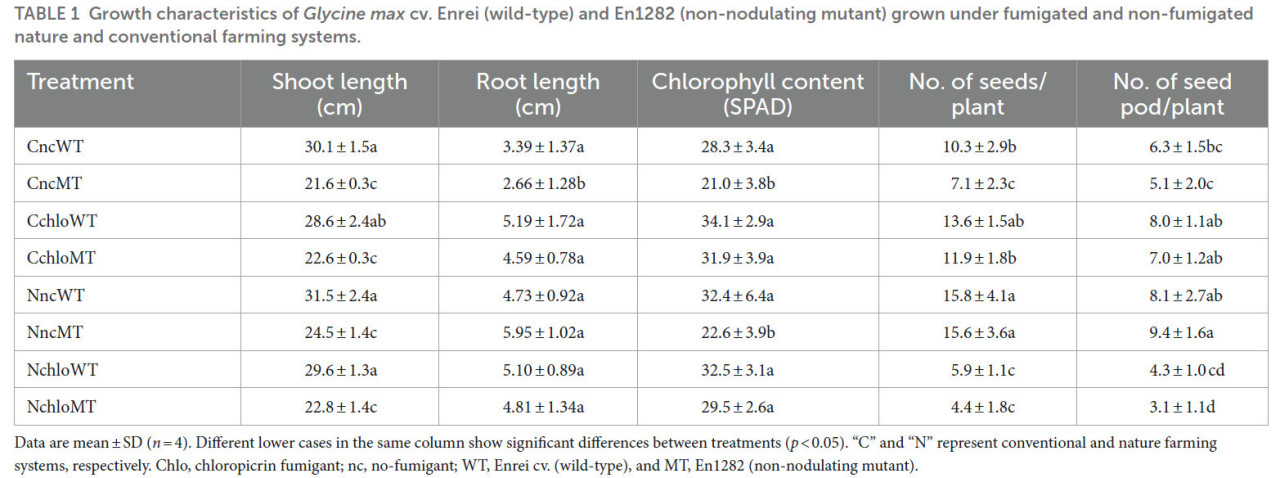
Таблица 1 Характеристики роста Glycine max cv. Enrei (дикий тип) и En1282 (немутирующий мутант), выращенных в фумигированных и нефугированных системах природного и традиционного земледелия.
Данные представлены как среднее ± стандартное отклонение (n = 4). Разные строчные буквы в одном столбце показывают значительные различия между обработками (p < 0,05). «C» и «N» представляют системы традиционного и природного земледелия соответственно. Chlo, фумигант хлорпикрин; nc, без фумиганта; WT, cv. Enrei (дикий тип), и MT, En1282 (немутирующий мутант).
3.3. Клубенькообразование и колонизация АМ грибами
Сорт сои Enrei и его немутирующий мутант En1282 выращивались в почвах традиционного и природного земледелия с обработкой фумигантом и без нее. Как и ожидалось, на корнях мутантных (немутирующих) растений клубеньков не обнаружено (Figure 1B), тогда как в среднем 17 и 59 клубеньков на растение было обнаружено на корнях растений дикого типа, выращенных в почвах традиционного и природного земледелия соответственно (Figure 1A). Однако фумигация снизила количество клубеньков в обеих системах земледелия, особенно резко в почве природного земледелия (Figure 1A). Было обнаружено высокозначимое взаимодействие (p < 0,001; Supplementary Table 2) между системой земледелия, обработкой фумигантом и сортом сои по количеству образованных клубеньков.
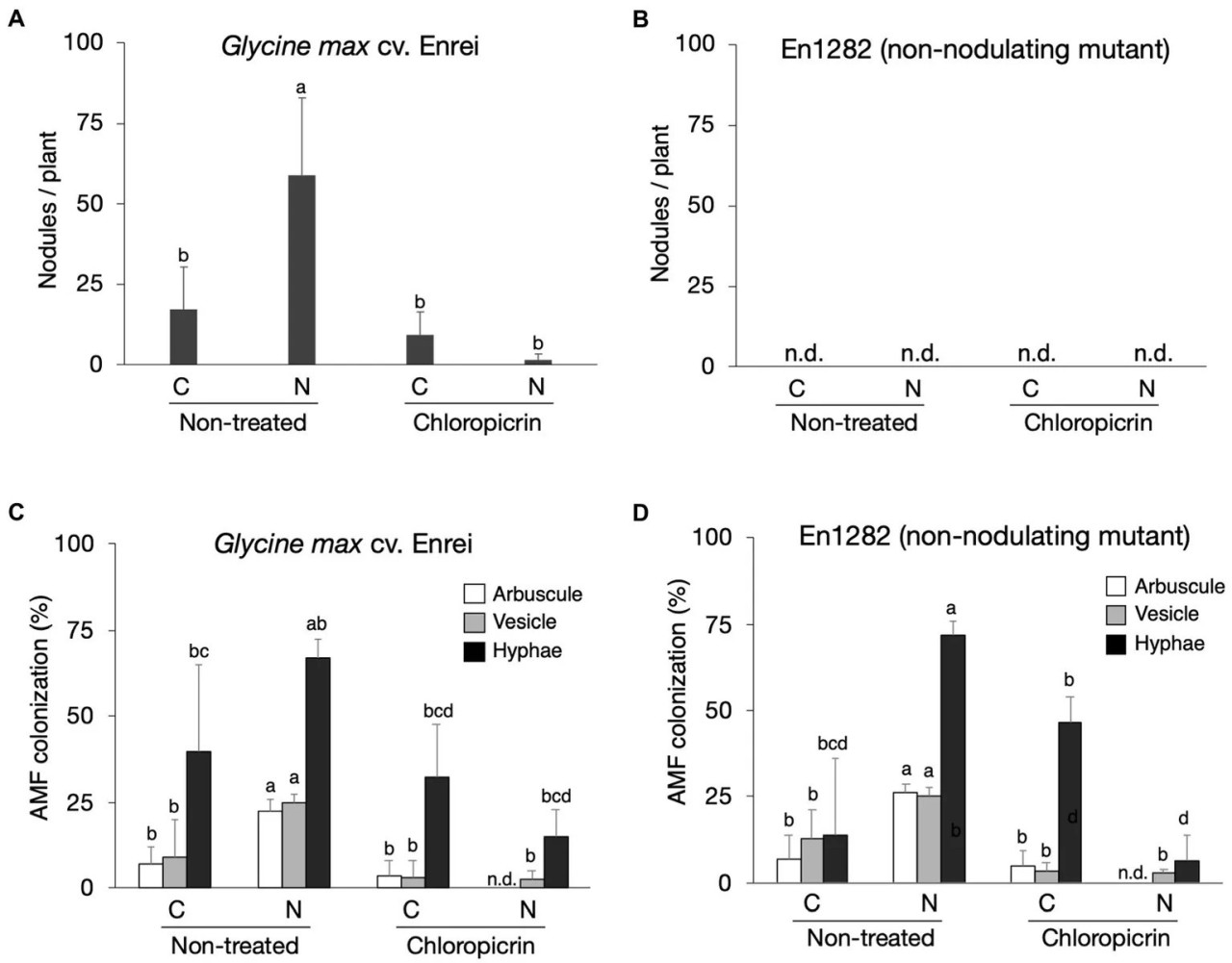
Figure 1 Влияние почв, фумигации и генотипов сои на среднее количество клубеньков (A,B) и скорость колонизации корней сои АМ грибами на основе наличия арбускул, везикул и гиф (C,D). Разные буквы указывают на значительные различия между образцами на основе теста SNK (p < 0,05). C, почва традиционного земледелия, N, почва природного земледелия, n = 4.
Микроскопические наблюдения окрашенных корней сои показали наличие морфологических структур АМ грибов, включая гифы, везикулы и арбускулы (Supplementary Figure 1). Процент колонизации арбускулами и везикулами как для растений дикого типа, так и для мутантов был заметно выше при выращивании в почве природного земледелия по сравнению с почвой традиционного земледелия, когда не применялся фумигант (Figures 1C,D). Процент колонизации гифами был выше у растений сои природного земледелия (66,8% для дикого типа; 72,0% для мутанта), чем у растений традиционного земледелия (40,0% для дикого типа; 13,8% для мутанта), хотя эти различия не были статистически значимыми, особенно для растений дикого типа (Figures 1C,D; Supplementary Table 2). Фумигация почв хлорпикрином привела к значительному снижению уровня колонизации корней сои АМ грибами как при традиционном, так и при природном земледелии. Было определено, что процент колонизации АМ грибами у растений сои не зависит от эффекта взаимодействия. Однако на него значительно влияли основные эффекты системы земледелия (p < 0,001) и обработки фумигантом (p < 0,001; Figures 1C,D; Supplementary Table 2). Средние показатели колонизации АМ грибами после обработки фумигантом были выше у растений, выращенных в условиях традиционного земледелия, чем в условиях природного земледелия (Figures 1C,D), что указывает на то, что примененный фумигант оказал серьезное воздействие на растения в условиях природного земледелия.
3.4. Результаты ампликонного секвенирования
Высокопроизводительное ампликонное секвенирование на платформе Illumina MiSeq дало 1 114 090 ридов для 16S и 895 540 ридов для ITS, распределенных по 24 образцам, принадлежащим к 4 919 и 2 369 различным вариантам ампликонной последовательности микробов для бактерий и грибов соответственно. Кривые разрежения неуклонно увеличивались по мере увеличения количества последовательностей (Supplementary Figure 1), что указывало на то, что размер выборки был адекватным для дальнейшего анализа.
3.5. Микробиом ризосферы сои в условиях природного земледелия более разнообразен и отличен, чем в условиях традиционного земледелия
Анализы альфа-разнообразия, основанные на индексах Шеннона и Chao1, выявили более разнообразное и богатое бактериальное сообщество в ризосфере растений сои, выращиваемых в условиях природного земледелия, чем у растений, выращиваемых в условиях традиционного земледелия (Figure 2A; Supplementary Table 3). Этот эффект, однако, был более выражен в ризосфере сои дикого типа, чем у мутанта, хотя сорт сои не оказывал значительного влияния на альфа-разнообразие ризосферы (Figure 2A; Supplementary Table 3). Аналогичная тенденция наблюдалась в грибном альфа-разнообразии (оба индекса Шеннона и Chao1), за исключением значительно более высокого индекса Шеннона, наблюдаемого в ризосфере мутантных растений, выращенных в почве традиционного земледелия, по сравнению с почвой природного земледелия (Figure 2B; Supplementary Table 4).
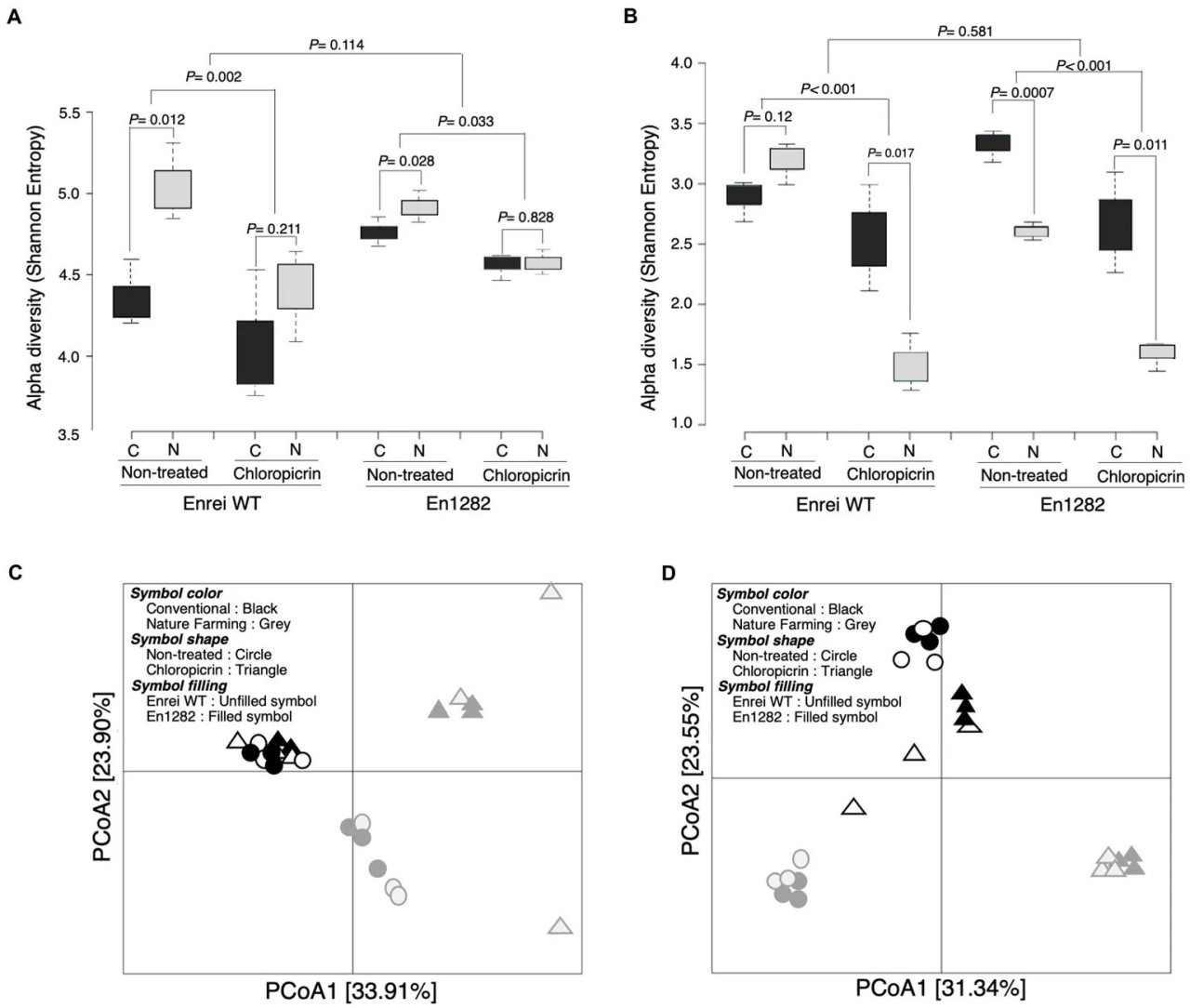
Figure 2 Бактериальное и грибное сообщество ризосферы сои под влиянием сорта, системы земледелия и обработки хлорпикрином. Альфа-разнообразие (индекс Шеннона) бактерий (A) и грибов (B). Анализ главных координат (PCoA) на основе индекса несходства Брея-Кертиса для бактерий (C) и грибов (D). C, почва традиционного земледелия, N, почва природного земледелия.
Согласно результатам различий между образцами (β-разнообразие) с использованием неограниченного анализа главных координат на основе расстояния Брея-Кертиса, наблюдалось четкое разделение по оси 1 для бактерий (Figure 2C) и по оси 2 для грибов (Figure 2D), что подтверждает, что почвы традиционного и природного земледелия содержат различные микробиомы. В то же время не наблюдалось четкого разделения растений дикого типа или мутантов, выращенных на любой из обрабатываемых почв, что позволяет предположить незначительное влияние сорта на структуру сообщества, что было дополнительно подтверждено тестами PERMANOVA и ANOSIM (Supplementary Figure 3).
3.6. Разнообразие микробиома ризосферы сои изменяется в ответ на фумигацию почвы
Фумигант оказал губительное воздействие на микробное разнообразие ризосферы сои, о чем свидетельствует значительно более низкое альфа-разнообразие в ризосферах растений, выращенных на фумигированных почвах, по сравнению с нефумигированными почвами (Figures 2A,B). После фумигации показатели разнообразия ризосферы (Шеннона и Chao1) были сходными между почвами традиционного и природного земледелия (Figure 2A; Supplementary Table 3). Не было заметных различий в альфа-разнообразии между растениями, растущими на фумигированной традиционной и фумигированной природной почве для бактерий, но наблюдалось значительное различие в грибном альфа-разнообразии, причем фумигант оказывал выраженное действие в ризосфере растений, растущих на почве природного земледелия (Figures 2A,B). Интересно, что видовое разнообразие бактерий растений, растущих на фумигированной почве природного земледелия, было выше, чем на нефумигированной почве традиционного земледелия (Figure 2A).
Анализ главных координат на основе расстояния Брея-Кертиса показал значительные различия в структуре микробного сообщества ризосферы между фумигированными и нефумигированными обработками, и это было подтверждено тестами PERMANOVA и ANOSIM (Figures 2C,D; Supplementary Figure 3). Сообщества ризосферы (бактерий и грибов) растений, растущих на фумигированной и нефумигированной почве традиционного земледелия, группировались вместе (Figures 2C,D), что указывает на сходство микробных сообществ. Однако сообщества растений, выращенных на почве природного земледелия, были относительно отличны друг от друга (Figures 2C,D), что указывает на более сильное действие фумигантов на микробные сообщества в почве природного земледелия.
3.7. Влияние системы земледелия на состав микробиома ризосферы сои
Состав бактериальных и грибных сообществ в основном различался между системами земледелия с точки зрения обилия или отсутствия определенных таксонов. Proteobacteria была наиболее обильным бактериальным типом во всех образцах, составляя 34,5–65,8% от общего числа бактериальных ридов (Supplementary Figure 4A). Firmicutes были более обильны в почве традиционного земледелия, в то время как Acidobacteria и Bacteroidetes доминировали в почве природного земледелия (Supplementary Figure 4A). Топ-10 бактериальных родов показаны на Figure 3A, большинство из которых принадлежали к Proteobacteria. Agrobacterium, Bacillus и Cupriavidus имели тенденцию к большему обилию в ризосфере растений, выращенных в почве традиционного земледелия, в то время как Rhizobium более доминировал у растений, растущих в почве природного земледелия. Неидентифицированные бактерии составляли ~30,0–48,6% относительной численности во всех образцах ризосферы.
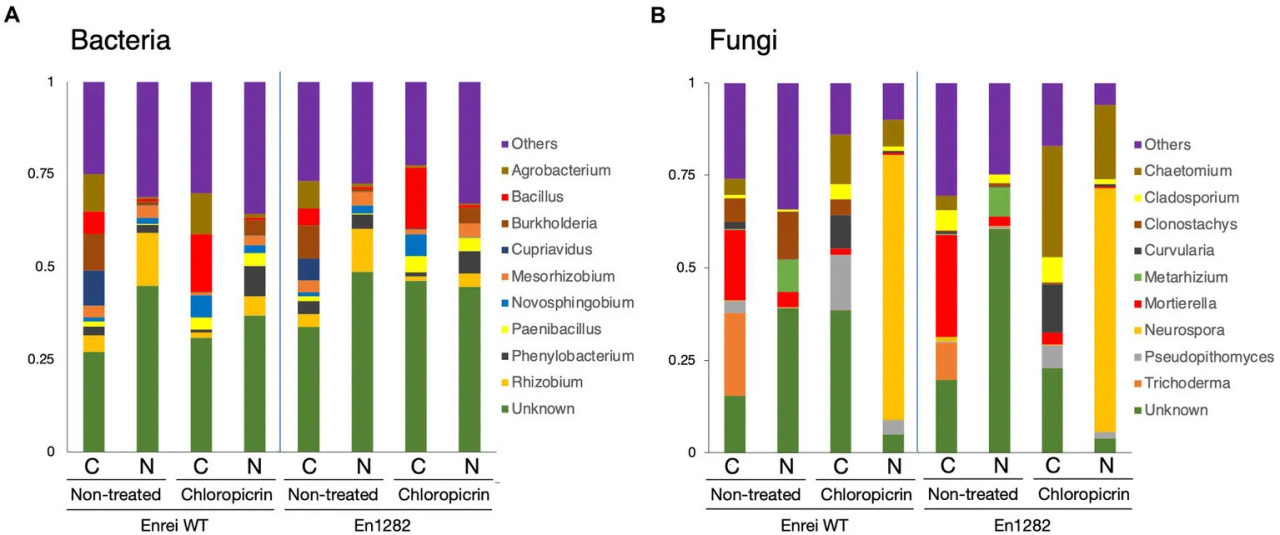
Figure 3 Таксономический состав микробных сообществ, населяющих ризосферу сои, выращенной в почвах традиционного и природного земледелия с обработкой хлорпикрином и без нее. Столбчатые графики представляют относительную численность бактериальных (A) и грибных (B) сообществ на уровне рода. Показаны только таксоны с численностью >1%. WT = cv. Enrei. C, почва традиционного земледелия, N, почва природного земледелия.
Что касается таксономического состава грибов, Ascomycota был доминирующим типом, варьируя от 37,1 до 97,2% во всех обработках (Supplementary Figure 4B). Другие часто сообщаемые типы грибов, такие как Basidiomycota, Chytridiomycota и Mortierellomycota, были обнаружены в очень низком количестве, и их распределение варьировалось в зависимости от обработки (Supplementary Figure 4B). Mortierella и Trichoderma доминировали в ризосфере растений сои в почве традиционного земледелия, в то время как Clonostachys и Metarhizium были более доминирующими в почве природного земледелия (Figure 3B).
3.8. Фумигация почвы влияет на состав бактериального и грибного сообщества ризосферы сои
Фумигация привела к изменению относительной численности определенных микробных таксонов. Количество Firmicutes и Bacteroidetes резко увеличилось, заметно в почве традиционного и природного земледелия соответственно, в то время как численность Proteobacteria относительно снизилась в обеих системах земледелия (Supplementary Figure 4A). Chloroflexi и Acidobacteria были истощены или элиминированы после фумигации, и этот эффект был более выражен в почве природного земледелия (Supplementary Figure 4A). Фумигация привела к тому, что численность Bacillus, Novosphingobium и Paenibacillus специфически увеличилась в ризосфере растений в почве традиционного земледелия, в то время как Phenylobacterium было более многочисленным в ризосфере растений в почве природного земледелия (Figure 3A). Burkholderia, Cupriavidus, Rhizobium и Mesorhizobium снизились по относительной численности в обеих системах земледелия после фумигации (Figure 3A).
Что касается грибного состава, фумигация привела к увеличению доли Ascomycota, в то время как численность Mortierellomycota, Chytridiomycota и Basidiomycota снизилась (Supplementary Figure 4B). Грибные роды были по-разному представлены среди обработок. Например, Chaetomium и Neurospora, которые не обнаруживались в ризосфере растений, выращенных в почве природного земледелия, внезапно появились и значительно увеличили свою численность после фумигации (Figure 3B). Отмечалось заметное увеличение численности Curvularia и Pseudopithomyces с более высоким распределением в почве традиционного земледелия, чем в почве природного земледелия (Figure 3B). Напротив, Trichoderma и Metarhizium, обнаруженные только в почве традиционного и природного земледелия соответственно, были элиминированы после фумигации (Figure 3B). Аналогично, численность Mortierella значительно снизилась после фумигации, и это было более выражено в ризосфере растений, выращенных в почве традиционного земледелия (Figure 3B).
3.9. Дифференциально обильные таксоны под влиянием системы земледелия и фумигации почвы
Был проведен линейный дискриминантный анализ (LefSe) для выявления дифференциально обильных таксонов, на которые значимо повлияла система земледелия или обработка фумигантом, с пороговым значением LDA-оценки, равным двум. Что касается бактериальных таксонов, ASV, классифицированные как Rhizobium, Phenylobacterium и некоторые представители семейства Comamonadaceae, среди прочих, были значительно обогащены в ризосфере растений, выращенных в почве природного земледелия (Figure 4A). Напротив, Bacillus sp., Cupriavidus, некоторые представители типа Gemmatimonadetes (Gemm 1, Ellin5301) и Paenibacillus были селективно истощены в почве природного земледелия и обогащены в почве традиционного земледелия (Figure 4A). Кроме того, некоторые полезные таксоны, такие как азотфиксирующие Rhizobium и Mesorhizobium, Streptomyces и Burkholderia, были сильно ассоциированы с нефумигированными почвами как ключевые таксоны (Figure 4C), в то время как Herbaspirillum, некоторые представители Shingobacteriaceae, Amycolatopsis и Bosea были ключевыми таксонами в фумигированных почвах (Figure 4C). Кроме того, степень обогащения таксонами варьировалась в зависимости от вовлеченного сорта. Например, ASV_002 (Bacillus flexus) стабильно обогащался в традиционной почве, но был более обильным у мутантов, выращенных на фумигированной традиционной почве (Supplementary Figures 4A, 5; Supplementary Table 5), в то время как ASV_020 и ASV_021 (Burkholderia bryophila и Sphingomonas sp., соответственно) стабильно обогащались в ризосфере дикого типа и мутантов, выращенных на нефумигированных почвах природного земледелия, соответственно (Supplementary Figures 5A, 6; Supplementary Table 5).
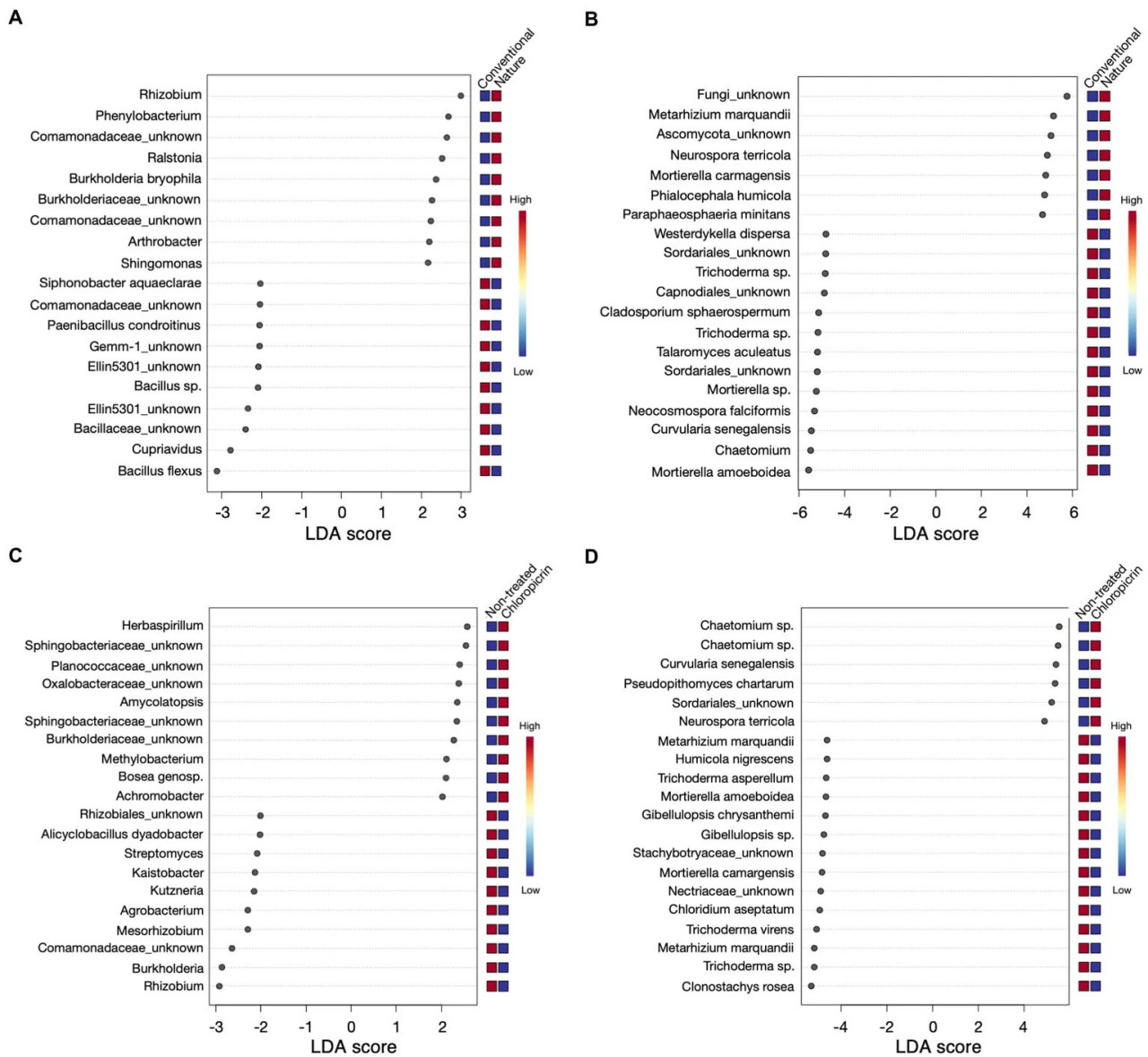
Figure 4 Результаты линейного дискриминантного анализа (LEfSe) для топ-20 бактерий (A) и грибов (B), которые были значительно обогащены либо в почве традиционного, либо природного земледелия, и бактерий (C) и грибов (D) со значимой численностью либо в необработанных, либо в обработанных хлорпикрином почвах. Красные рамки показывают высокую численность, а синие рамки — низкую численность конкретного рода.
Что касается грибного сообщества, ключевые таксоны в ризосфере растений, выращенных в почве природного земледелия, включали Metarhizium sp. (Figure 4B), который также был дифференциально обилен в нефумигированных почвах (Figure 4D). Напротив, Neurospora была значима в почвах природного земледелия и фумигированных почвах. Mortierella sp. также была среди таксонов, избирательно рекрутируемых в ризосферу как дикого типа, так и мутантов, выращенных на нефумигированной почве традиционного земледелия (Supplementary Figures 5B, 7; Supplementary Table 6). С другой стороны, грибной ASV_039 (Chaetomium grande) был дискриминационно выше у растений, ассоциированных с фумигированной традиционной почвой, но более обилен в ризосфере сои дикого типа (Supplementary Figures 5B, 7; Supplementary Table 6).
3.10. Анализ функционального предсказания бактериальных сообществ ризосферы сои
Филогенетическое исследование сообществ путем реконструкции ненаблюдаемых состояний 2 (PICRUSt2) использовалось для предсказания предполагаемых функциональных различий бактериальных сообществ, колонизирующих ризосферу сои как в условиях традиционного, так и природного земледелия, с обработкой фумигантом и без нее. Метаболические пути, связанные с биосинтезом кофакторов и витаминов, и пути, связанные с деградацией, утилизацией и ассимиляцией соединений в качестве источников питательных веществ и энергии, включая деградацию аминов и полиаминов, биосинтез жирных кислот и липидов, а также суперпуть D-глюкарата и D-галактарата, преимущественно ассоциировались с ризобактериями в почве природного земледелия (Figure 5A). Это важно, поскольку микробы используют питательные вещества и энергию для своего роста и пролиферации в ризосфере растений в почве природного земледелия. Такие пути, как биосинтез нуклеозидов и нуклеотидов, глиоксилатный цикл, деградация вторичных метаболитов и цикл трикарбоновых кислот, более доминировали в обработках, связанных с традиционным земледелием (Figure 5A). В нефумигированной почве было больше предсказанных функций по сравнению с фумигированной почвой. Например, деградация аминокислот, деградация альдегидов, биосинтез аминов и полиаминов, биосинтез жирных кислот и липидов и фотосинтез были более выражены в нефумигированных почвах (Figure 5B). Такие пути, как метаболизм неорганических питательных веществ, деградация карбоксилатов, деградация углеводов и биосинтез гликанов, были более заметны в фумигированных почвах (Figure 5B).
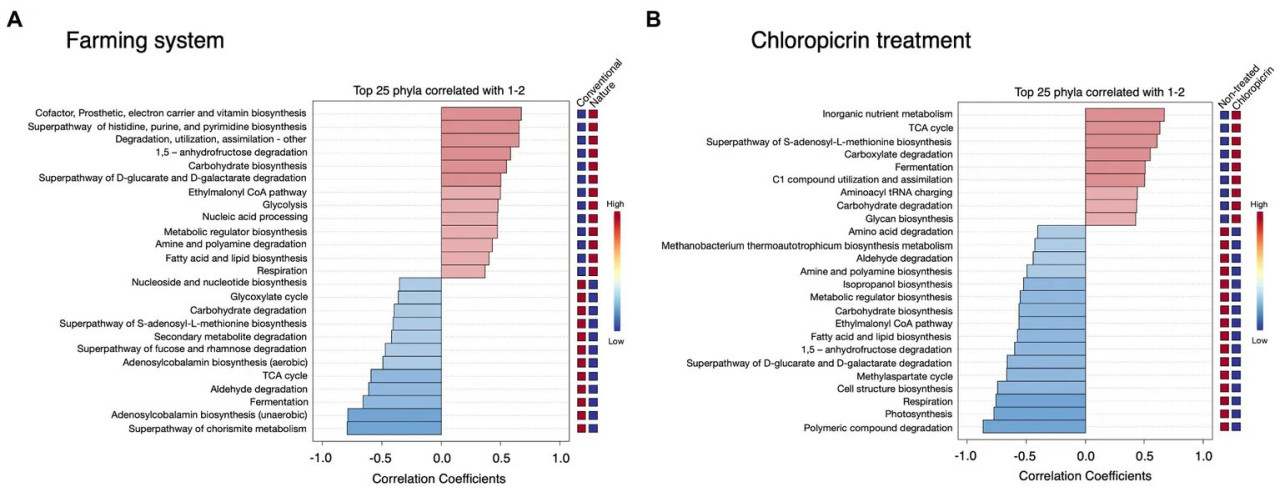
Figure 5 Предполагаемое функциональное предсказание бактериальных сообществ в ризосфере сои, выращенной в различных системах земледелия (A), и при обработке хлорпикрином (B) с использованием PICRUSt. Показаны топ-25 наиболее значимо доминирующих функциональных путей.
3.11. Анализ сетей совместной встречаемости ризосферы сои
Микробные сообщества в нефумигированной почве природного земледелия были наиболее сложными и плотно связанными по сравнению с соответствующими сетями обработок традиционной почвой, в то время как сеть ризосферы была менее связанной и менее плотной в фумигированной почве природного земледелия (Figure 6). Положительные попарные взаимодействия наблюдались в обработанной традиционной почве между Nitrospira (принадлежащей к типу Nitrospirota) и Mesorhizobium, но были отрицательно связаны с Rhizobium и Bradyrhizobium, все из которых принадлежат к типу Proteobacteria (Figure 6B). Rhizobium, Bosea, Mesorhizobium, Methylobacterium и Bradyrhizobium были среди наиболее тесно связанных таксонов, совместно встречающихся в нефумигированной почве природного земледелия (Figure 6C), и все они участвовали в положительных взаимодействиях между собой и с другими таксонами. Положительные взаимодействия с этими таксонами подразумевают их синергетическую активность в усилении N2-фиксации.
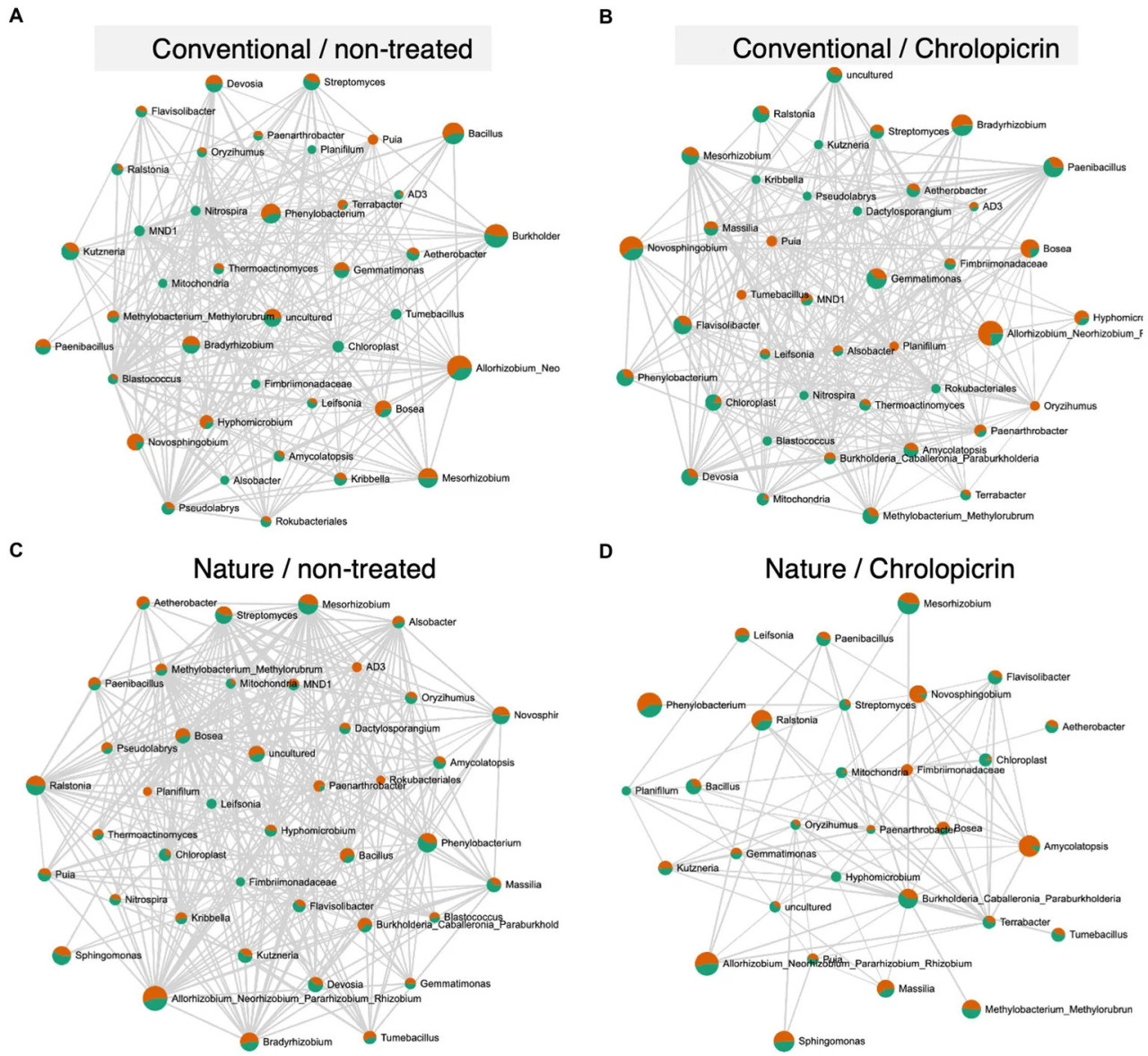
Figure 6 Анализ сетей совместной встречаемости бактериальных сообществ ризосферы сои, выращенной в необработанной традиционной почве (A), обработанной хлорпикрином традиционной почве (B), необработанной почве природного земледелия (C) и обработанной хлорпикрином почве природного земледелия (D). Корреляционная сеть, сгенерированная с использованием алгоритма SparCC, с узлами, представляющими таксоны на уровне рода, и ребрами, представляющими корреляции между бактериальными таксонами. Сети совместной встречаемости основаны на корреляции Пирсона с порогом r > 0,5.
4. Обсуждение
Хотя взаимодействие «растение-микроб» и «микроб-микроб» в ризосфере хорошо изучено на нескольких культурах (Edwards и др., 2015; Durán и др., 2018; Thiergart и др., 2019), взаимодействие микроорганизмов в ризосфере сельскохозяйственных культур в условиях систем природного земледелия изучено слабо. В настоящем исследовании микробиом ризосферы сои и его влияние на рост сои в почве природного земледелия сравнивались с методами традиционного земледелия в условиях обработки почвы фумигантом. В настоящем исследовании был изучен микробиом ризосферы, чтобы понять влияние агрохимических добавок на полезные почвенные микроорганизмы и рост растений, а также то, как альтернативные методы ведения сельского хозяйства, такие как природное земледелие, где не используются внешние удобрения и пестициды, могут влиять на обилие полезных почвенных микробов и рост растений.
4.1. Влияние системы земледелия и фумигации на симбиотические ассоциации сои
В настоящем исследовании значительно большее количество клубеньков наблюдалось на корнях сои дикого типа в почве природного земледелия, чем в почве традиционного земледелия, когда не применялся фумигант (Figure 1A). Это может быть связано с более низкой местной популяцией ризобий в почве традиционного земледелия по сравнению с почвой природного земледелия или с отсутствием в почве традиционного земледелия совместимых с соей штаммов ризобий для симбиоза. Было обнаружено высокозначимое взаимодействие между системой земледелия, обработкой фумигантом и сортом сои по количеству образованных клубеньков. Это может означать, что эффект системы земледелия или применяемого фумиганта на образование клубеньков зависит от используемого сорта сои, что очевидно из клубеньков, образовавшихся у растений дикого типа, но не у мутантных растений.
Арбускулярные микоризные грибы, наиболее распространенные и очень полезные ассоциированные с растениями грибы (Smith и др., 2011), представляют особый интерес в системах природного земледелия, где плодородие почвы поддерживается без использования агрохимикатов. Интересно, что значительно более высокий процент колонизации корней с точки зрения гиф, везикул и арбускул наблюдался в необработанных фумигантом обработках природного земледелия, чем в необработанных обработках традиционного земледелия (Figures 1C,D). Это согласуется с другими сообщениями, утверждающими, что растения, выращенные в почвах с органическим управлением, имели более высокую скорость колонизации АМ грибами по сравнению с растениями в интенсивных традиционных почвах (Entz и др., 2004; Gosling и др., 2010; Gazdag и др., 2019). Это может быть частично объяснено тем фактом, что АМ грибы процветают в условиях с низким поступлением питательных веществ, как при природном земледелии, где питательные вещества медленно рециркулируются почвенными микроорганизмами. Это контрастирует с методами традиционного земледелия, где большие количества химикатов часто вносятся, тем самым влияя на симбиотические взаимодействия сои с грибами в отношении взаимного обмена питательными веществами и углеродом между ними (Oehl и др., 2004). Однако следует отметить, что содержание доступного P было выше в почве природного земледелия, чем в почве традиционного земледелия в нашем исследовании (Supplementary Table 1). Таким образом, другие факторы, кроме P, такие как популяция жизнеспособных спор АМ грибов, могли повлиять на колонизацию АМ грибами в этом исследовании. Поэтому необходимы дальнейшие исследования для характеристики видового разнообразия АМ грибов и обилия спор в традиционном и природном земледелии, прежде чем можно будет сделать более обоснованные выводы.
С другой стороны, фумигация хлорпикрином значительно снизила колонизацию АМ грибами и количество корневых клубеньков, с более сильным эффектом на растения, выращенные в почве природного земледелия (Figures 1A–C). Это предполагает, что в почве природного земледелия существовала активная местная популяция ризобий и АМ грибов, которая была сильно затронута фумигацией. Из этих результатов можно выдвинуть гипотезу, что фумигант отрицательно повлиял на клубенькообразование и, следовательно, на биологическую фиксацию азота у сои, нарушая сигнальную активность Rhizobium, необходимую для инфицирования корней, или воздействуя на корневые волоски сои, где происходит инфицирование и образование клубеньков (Meena и др., 2020). Предыдущие исследования показали, что почвенные фумиганты, такие как хлорпикрин и формальдегид, ингибируют инфицирование корней АМ грибами и развитие спор (Dangi и др., 2017), что объясняется токсическим действием фумигантов на споры грибов и развитие гиф. Хотя не было значимого взаимодействия между всеми тремя переменными, участвовавшими в этом исследовании, наблюдалось значимое взаимодействие (p < 0,001; Supplementary Table 2) между системой земледелия и обработкой фумигантом по скорости колонизации АМ грибами. Таким образом, эффект фумиганта на колонизацию АМ грибами больше зависел от типа системы земледелия, что могло бы объяснить, почему фумигант оказал более серьезное влияние на колонизацию АМ грибами в природном земледелии независимо от сорта сои. Это означает, что в одной из двух обрабатываемых почв могут присутствовать разные виды АМ грибов, которые либо более восприимчивы, либо более толерантны к фумиганту из-за методов управления земледелием, применяемых в традиционном или природном земледелии.
4.2. Зависимая от системы земледелия и фумигации ростовая производительность сои
В этом исследовании высота растений сои дикого типа, растущей в почве традиционного или природного земледелия, не различалась значимо независимо от обработки фумигантом, но все они были значительно выше, чем обработки мутантных растений (Table 1). Поскольку все почвенные системы были одинаковыми для обоих сортов, этот эффект можно объяснить генетической структурой мутанта, которая привела к ограниченному образованию клубеньков и симбиозу с ризобиями. В этом исследовании было показано, что масса корней сои относительно не зависела ни от системы земледелия, ни от применения фумиганта, за исключением мутантных растений на нефумигированной традиционной почве (Table 1). Это может быть связано с уничтожением корневых патогенов фумигантом вокруг корней сои или с наличием микробов биоконтроля в почве природного земледелия, что отражает то, как культура должна выглядеть в здоровой почвенной системе.
Содержание хлорофилла в листьях мутанта увеличилось в фумигированных почвах (традиционной и природной) по сравнению с таковыми в нефумигированной почве и было сходно с растениями дикого типа (Table 1). У мутантов, выращенных на нефумигированных почвах, более низкое содержание хлорофилла в листьях не удивительно из-за отсутствия азотфиксации в результате отсутствия образования клубеньков (Figures 1A,B). Однако предыдущие исследования показали, что фумигация увеличивает содержание азота в почвах из-за деградации и высвобождения питательных веществ из мертвых микробных клеток, что может объяснить более высокое содержание хлорофилла у мутантных растений, ассоциированных с фумигированными почвами (Yan и др., 2022). Кроме того, конечными продуктами деградации хлорпикрина, который был использован в качестве фумиганта в этом исследовании, являются диоксид углерода, хлор и азот. Следовательно, высвобождение азота после деградации хлорпикрина также может объяснить повышенное содержание хлорофилла в листьях мутантных растений, выращенных на фумигированных почвах, по сравнению с мутантами на нефумигированных почвах. Однако текущее исследование было ограничено анализом питательных веществ почвы в фумигированных почвах и поглощением питательных веществ растениями после сбора урожая, что следует учитывать в будущих исследованиях. Интересно, что мы наблюдали более выраженное и значительно более высокое среднее количество бобов и семян на растениях сои, выращенных в почве природного земледелия у мутанта, но количество значительно снизилось при фумигированной обработке (Table 1). Поэтому можно постулировать, что этот эффект может быть результатом присутствия некоторых полезных микробных симбионтов, таких как АМ грибы, в ризосфере мутанта, которые помогали способствовать образованию бобов и семян по сравнению с растениями дикого типа, потому что у мутанта не было симбиоза с ризобиями для образования клубеньков.
4.3. Практики управления земледелием формируют сообщество ризосферы сои
В то время как сорт сельскохозяйственной культуры оказал меньшее влияние на разнообразие микробиома ризосферы сои, практика ведения сельского хозяйства оказала большое влияние на него. Поэтому большинство обсуждений ниже было сосредоточено на системе земледелия. Текущее исследование показало, что ризосфера растений сои, выращиваемых в условиях природного земледелия, имела большее бактериальное и грибное разнообразие (индексы Шеннона и Chao1), чем соя, выращиваемая в условиях традиционного земледелия (Figures 2A,B; Supplementary Tables 3, 4). Аналогично этому исследованию, другие обнаружили более высокое микробное разнообразие в системах органического/природного земледелия по сравнению с традиционными системами (Chaudhry и др., 2012; Hartmann и др., 2015; Lupatini и др., 2017; Ares и др., 2021; Sinong и др., 2021). Более высокое альфа-разнообразие в ризосферной почве природного земледелия также обнаружено в почве, где ограничено использование агрохимикатов. Lupatini и др. (2017) сообщили, что непрерывное использование агрохимикатов в традиционной практике земледелия может прямо или косвенно уничтожать и ингибировать рост определенных микробных групп, что приводит к более низкому микробному разнообразию в традиционных почвах по сравнению с почвами с органическим управлением.
Интересно, что анализы альфа-разнообразия показали, что грибное разнообразие мутанта было значительно выше в традиционной, нефугимированной почве, чем в почве природного земледелия (Figure 2B; Supplementary Table 4). Возможно, существует избирательное действие корневых экссудатов, выделяемых мутантными растениями, на специфические таксоны, которые могут быть полезны для их роста и здоровья, как это наблюдалось ранее (Mendes и др., 2013; Philippot и др., 2013). Наблюдалось четкое разделение бактериального и грибного сообщества ризосферы между почвой традиционного и природного земледелия, в то время как влияние сорта сои было незначительным (Figures 2C,D; Supplementary Figure 3). Хотя сорт растения не оказал сильного влияния на структуру сообществ ризосферы в текущем исследовании, исследования на других системах выявили такой эффект (Ares и др., 2021). Результаты этого исследования показали, что, независимо от сорта растения, практика управления хозяйством может оказывать заметное влияние на структуру микробиома ризосферы растений.
4.4. Фумигация почвы губительно влияет на микробное разнообразие и изменяет структуру сообщества ризосферы сои
Некоторые исследования сообщают, что почвенные микробиомы значительно изменяются химическими фумигантами (Ibekwe и др., 2001; Rokunuzzaman и др., 2016; Li и др., 2017). В этом исследовании наблюдалось, что применяемый почвенный фумигант значительно снизил микробное разнообразие ризосферы сои по сравнению с обработками без фумигации в обеих системах земледелия (Figures 2A,B; Supplementary Tables 3, 4), возможно, потому что токсичность фумиганта привела к увеличению смертности клеток, таким образом предотвращая восстановление некоторых микробных таксонов и приводя к значительному снижению микробного богатства и разнообразия (Xiong и др., 2022). Некоторые другие исследования сообщили о более низком альфа-разнообразии микробов из-за фумигации почвы и объяснили это прямой токсичностью фумигантов, которые обладают широким спектром биоцидной активности против некоторых местных почвенных микроорганизмов (Ibekwe и др., 2001; Rokunuzzaman и др., 2016; Li и др., 2017).
Это исследование показало, что сообщества ризосферы в почве традиционного земледелия, обработанной фумигантом, имели сходную структуру бактериального и грибного сообщества с таковыми в фумигированной почве природного земледелия. Однако в почве природного земледелия сообщества ризосферы в фумигированных образцах различались по сравнению с нефугированными образцами как для бактерий, так и для грибов (Figures 2C,D). Это, скорее всего, потому, что сообщество ризосферы в традиционной почве было более устойчиво к стрессу от фумиганта, возможно, из-за механизма адаптации, развившегося в результате непрерывного использования химикатов в традиционной практике земледелия. Напротив, поскольку различные микробы имеют разную степень толерантности к внешним стрессорам (Bouskill и др., 2013), воздействие фумигации на почву природного земледелия вызвало сдвиг микробных профилей ризосферы в сторону более устойчивых таксонов, которые могли выдержать стресс фумиганта и занять пустующие ниши, что привело к возникновению двух различных сообществ (Figure 2C,D; Supplementary Figure 3).
4.5. Влияние практики управления земледелием на таксономический состав ризосферы сои
Proteobacteria была наиболее преобладающим бактериальным типом (от 34,5 до 65,8%), обнаруженным во всех образцах ризосферной почвы в нашем исследовании (Supplementary Figure 4A). Представители Proteobacteria обычно считаются копиотрофами, которые адаптированы к богатой углеродом среде для своего быстрого роста и размножения (Ling и др., 2022). Свойством ризосферы, сходным у разных видов растений, является секреция корневых экссудатов, которые обеспечивают производный от корней углеродный субстрат для ризосферных микроорганизмов. Поэтому неудивительно, что тип Proteobacteria стабильно обогащался даже в фумигированных образцах ризосферы. Firmicutes, хотя и присутствовали в ризосфере растений как традиционных, так и природных почв, были значительно обогащены в традиционной почве по сравнению с природной (Supplementary Figure 4A). Известно, что представители этого типа, включая Bacillus, производят антибиотики, которые контролируют широкий спектр почвенных патогенов растений. Acidobacteria, которые были более обогащены в ризосфере растений природного земледелия, обычно считаются олиготрофами, которые доминируют в почвах с низкой доступностью ресурсов (Fierer и др., 2012). Это может объяснять их обогащение в ризосферной почве растений сои природного земледелия.
Rhizobium, Phenylobacterium и Metarhizium в почве природного земледелия и Bacillus, Cupriavidus, Mortierella, Chaetomium и Curvularia в почве традиционного земледелия были среди бактериальных и грибных таксонов, которые способствовали дифференциации сообществ ризосферы сои, ассоциированных с двумя системами земледелия, согласно анализу LEfSe (Figures 4A,B). Помимо их вклада в клубенькообразование у бобовых и фиксацию азота, сообщалось, что некоторые виды ризобий способствуют росту растений, помогая в растворении P (Ahmad и др., 2008) и производстве индолил-3-уксусной кислоты (IAA), которая способствует удлинению корней для поглощения питательных веществ (Igiehon и др., 2019). Совсем недавно Baron и др. (2020) сообщили о способности вида Metarhizium Metarhizium marquandii производить IAA и растворять нерастворимый фосфат, а также способствовать росту сои.
4.6. Влияние фумигации почвы на таксономический состав ризосферы сои
Результаты показали, что фумигация привела к изменениям в составе микробиома ризосферы сои, увеличивая или уменьшая относительную численность определенных таксонов по сравнению с нефумигированными образцами. В бактериальных сообществах относительная численность Proteobacteria немного снизилась по всем образцам ризосферы, в то время как Acidobacteria была ингибирована в образцах ризосферы природного земледелия после фумигации (Supplementary Figure 4A). Предыдущие исследования сообщали, что представители типа Acidobacteria часто чувствительны к экологическим стрессорам (Xiong и др., 2022), и эта чувствительность может быть возможной причиной их ингибирования в ризосферных почвах после фумигации. Относительная численность Firmicutes увеличилась после фумигации с более высокими долями в образцах ризосферы традиционного земледелия, чем в образцах природного земледелия (Supplementary Figure 4A). Liu и др. (2015) также обнаружили увеличение доли грамположительных бактерий в почве после фумигации и сообщили, что эти микробные группы более толерантны к химикатам, включая фумиганты, из-за состава их клеточной стенки и способности образовывать споры, что делает их устойчивыми к внешним стрессорам и позволяет им быстрее адаптироваться к парам фумигантов.
Бактериальные роды Rhizobium, Burkholderia и Streptomyces были специфическими таксонами, которые больше всего способствовали значительной дифференциации микробных сообществ ризосферы после фумигации, а грибные роды Trichoderma, Clonostachys, Mortierella и Metarhizium — к нефумигированным образцам (Figures 4C,D). Burkholderia bryophila, которая была дискриминационно обогащена в почве природного земледелия, была описана как проявляющая антагонистическую активность против нескольких патогенов растений, включая Xanthomonas campestris и Rhizoctonia solani, и способствующая выработке гормонов роста растений, таких как ACC-деаминаза и сидерофоры (Vandamme и др., 2007). Streptomyces известен биоконтролем почвенных грибных патогенов благодаря своей антагонистической активности через синтез различных противогрибных метаболитов (Viaene и др., 2016; Vurukonda и др., 2018) и своей роли в растворении неорганического фосфата (de Jesus Sousa and Olivares, 2016). Trichoderma — это гриб, способствующий росту растений, который улучшает поглощение питательных веществ растениями, производит гормоны роста и защищает растения от заражения патогенами (Bononi и др., 2020; Chen и др., 2021). Род Mortierella известен производством антибиотиков и действует как потенциальный антагонист против различных патогенов растений (Xiong и др., 2017). Однако фумигация способствовала значительному снижению численности этих полезных микроорганизмов, что дополнительно подтверждает воздействие химических фумигантов на уничтожение или ингибирование нецелевых полезных микроорганизмов в почве.
Анализ LEfSe показал, что бактериальные роды Herbaspirillum и Bacillus, а также грибные роды Neurospora и Chaetomium положительно реагировали на примененный фумигант и были значительно обогащены в образцах ризосферы сои (Figures 4C,D). Повышенная численность этих микробных таксонов после фумигации может быть связана с их способностью разлагать и использовать фумигант в качестве источника питательных веществ для своего роста и колонизации пустующих ниш (Castellano-Hinojosa и др., 2021). Вид Neurospora со способностью растворять фосфат был выделен из ризосферы Sorghum bicolor L. (Sharma и др., 2016). Определение точных функций Neurospora sp. в микробиоме сои потребует дальнейших экспериментов и функциональной характеристики. Chaetomium, другой гриб, который был значительно обогащен в фумигированных обработках, широко используется в качестве агента биоконтроля (Castellano-Hinojosa и др., 2021), что позволяет предположить, что растения сои рекрутируют эти толерантные к фумиганту организмы для своего роста.
4.7. Предполагаемые функциональные признаки микробиома сои, предсказанные PICRUST2
Анализ функционального предсказания в этом исследовании показал, что микробиом ризосферы растений в почве природного земледелия может быть связан с большим количеством предполагаемых функций, чем в почве традиционного земледелия (Figure 5A), что может быть связано с более высоким микробным разнообразием, наблюдаемым в ризосфере растений в почве природного земледелия по сравнению с почвой традиционного земледелия. Кроме того, более разнообразные метаболические функции были ассоциированы с микробиомом ризосферы растений в нефумигированных почвах, чем в фумигированных почвах (Figure 5B). Это понятно, поскольку мы наблюдали более высокое микробное разнообразие ризосферы у растений, ассоциированных с нефумигированными почвами, по сравнению с растениями на фумигированных почвах, таким образом, более разнообразные микробные сообщества могут отвечать за выполнение более разнообразных функций. Однако дальнейший анализ, такой как транскриптомика, обеспечил бы лучшее понимание микробных сообществ и их функциональной активности.
4.8. Структуры сетей совместной встречаемости микробов в ризосфере сои
Сетевой анализ был проведен для более глубокого понимания сложности микробных взаимодействий, действующих в различных образцах ризосферной почвы. Микробные сети в ризосферной почве природного земледелия без фумиганта были более плотными и имели более сложную сеть с большим количеством узлов и ребер (Figure 6C), чем в обработках традиционного земледелия или фумигированной почве природного земледелия. Это, несомненно, связано с более высоким микробным разнообразием в ризосфере растений сои, выращиваемых в природных условиях. Результаты ясно показывают, что практика ведения сельского хозяйства влияет на сложность микробиома ризосферы и что природное земледелие способствует более сложным взаимодействиям между различными таксонами, обеспечивая тем самым более высокую стабильность сообщества. Наши результаты сходны с результатами Banerjee и др. (2019), которые наблюдали значительно более связанные микробные сообщества в почве органического земледелия, чем в традиционной и почве с нулевой обработкой. Сложные сети с высокой связностью более устойчивы к экологическим стрессорам, чем простые сети с ограниченными взаимодействиями (Banerjee и др., 2019). Таким образом, более высокая сложность структуры микробной сети ризосферы в почве природного земледелия может указывать на то, что различные таксоны в ризосфере растений в почве природного земледелия могут дополнять друг друга, чтобы противостоять воздействиям окружающей среды, таким как абиотический стресс или атака патогенов. Результаты далее показали, что природное земледелие содержит уникальный микробиом, на который фумигант оказал значительное влияние по сравнению с микробиомом почвы традиционного земледелия. Однако этот вывод нуждается в дальнейшей проверке с помощью анализа более широкого спектра почв природного земледелия из разных местоположений ферм и функционального анализа полученной микробиоты (чтобы понять, как она связана с поглощением питательных веществ и устойчивостью к болезням) для обоснованного заключения по этой гипотезе.
5. Заключение
В настоящем исследовании изучались рост сои и микробиом ризосферы в почвах традиционного и природного земледелия, а также влияние почвенного фумиганта на микробные сообщества ризосферы. Это первое исследование, в котором сообщается о влиянии микробных сообществ ризосферы сои на рост растений в системе природного земледелия. Результаты показали, что по сравнению с почвой традиционного земледелия в ризосфере растений, выращенных в условиях природного земледелия, наблюдались высокоразнообразные бактериальные и грибные сообщества, способствующие сильному симбиозу ризобий и АМ грибов. Это наиболее важные бактериальные и грибные микроссимбионты, которые помогают в усвоении питательных веществ и устойчивости растений даже без использования химических удобрений и пестицидов. Результаты этого исследования подтвердили более высокую сложность сети и связность узлов в почве природного земледелия без обработки хлорпикрином, что может указывать на более устойчивую структуру сообщества, устойчивую к воздействию окружающей среды. Кроме того, мы показали, что почвенный фумигант изменил состав микробиома ризосферы, сформировавшийся вокруг корней сои, путем элиминации или снижения численности некоторых полезных ассоциированных с растениями микробов, включая Rhizobium, Streptomyces, Burkholderia, по сравнению с нефумигированной почвой. Настоящее исследование расширяет наши знания об уникальности природного земледелия и предоставляет новое понимание положительных микробных взаимодействий в ризосфере растений, возделываемых в условиях природного земледелия.
Дополнительный материал
Дополнительный материал к этой статье можно найти онлайн по адресу: https://www.frontiersin.org/articles/10.3389/fmicb.2023.1130969/full#supplementary-material
Ссылки
1. Ahmad F., Ahmad I., Khan M. S. (2008). Screening of free-living rhizospheric bacteria for their multiple plant growth promoting activities. Microbiol. Res. 163, 173–181. doi: 10.1016/j.micres.2006.04.001. CrossRef. Google Scholar.
2. Amano Y. U. J. I. (1985). Classification of cultivated soils in Japan. Jpn. Agric. Res. Q. 18, 275–283. Google Scholar.
3. Anderson M. J. (2014). Permutational multivariate analysis of variance (PERMANOVA). Wiley Stat. Ref. Online, 1–15. doi: 10.1002/9781118445112.stat07841. CrossRef. Google Scholar.
4. Ares A., Costa J., Joaquim C., Pintado D., Santos D., Messmer M. M. et al. (2021). Effect of low-input organic and conventional farming systems on maize rhizosphere in two Portuguese open-pollinated varieties (OPV), "Pigarro" (improved landrace) and "SinPre" (a composite cross population). Front. Microbiol. 12:636009. doi: 10.3389/fmicb.2021.636009. CrossRef. Google Scholar.
5. Banerjee S., Walder F., Büchi L., Meyer M., Held A. Y., Gattinger A. et al. (2019). Agricultural intensification reduces microbial network complexity and the abundance of keystone taxa in roots. ISME J. 13, 1722–1736. doi: 10.1038/s41396-019-0383-2. CrossRef. Google Scholar.
6. Baron N. C., de Souza Pollo A., Rigobelo E. C. (2020). Purpureocillium lilacinum and Metarhizium marquandii as plant growth-promoting fungi. PeerJ 8:e9005. doi: 10.7717/peerj.9005. CrossRef. Google Scholar.
7. Berendsen R. L., Pieterse C. M., Bakker P. A. (2012). The rhizosphere microbiome and plant health. Trends Plant Sci. 17, 478–486. doi: 10.1016/j.tplants.2012.04.001. CrossRef. Google Scholar.
8. Beule L., Karlovsky P. (2020). Improved normalization of species count data in ecology by scaling with ranked subsampling (SRS): application to microbial communities. PeerJ 8:e9593. doi: 10.7717/peerj.9593. CrossRef. Google Scholar.
9. Bokulich N. A., Kaehler B. D., Rideout J. R., Dillon M., Bolyen E., Knight R. et al. (2018). Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2's q2-feature-classifier plugin. Microbiome 6, 90–17. doi: 10.1186/s40168-018-0470-z. CrossRef. Google Scholar.
10. Bolyen E., Rideout J. R., Dillon M. R., Bokulich N. A., Abnet C. C., Al-Ghalith G. A. et al. (2019). Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 37, 852–857. doi: 10.1038/s41587-019-0209-9. CrossRef. Google Scholar.
11. Bononi L., Chiaramonte J. B., Pansa C. C., Moitinho M. A., Melo I. S. (2020). Phosphorus-solubilizing Trichoderma spp. from Amazon soils improve soybean plant growth. Sci. Rep. 10, 1–13. doi: 10.1038/s41598-020-59793-8. CrossRef. Google Scholar.
12. Bouskill N. J., Lim H. C., Borglin S., Salve R., Wood T. E., Silver W. L. et al. (2013). Pre-exposure to drought increases the resistance of tropical forest soil bacterial communities to extended drought. ISME J. 7, 384–394. doi: 10.1038/ismej.2012.113. CrossRef. Google Scholar.
13. Bulgarelli D., Schlaeppi K., Spaepen S., Van Themaat E. V. L., Schulze-Lefert P. (2013). Structure and functions of the bacterial microbiota of plants. Annu. Rev. Plant Biol. 64, 807–838. doi: 10.1146/annurev-arplant-050312-120106. CrossRef. Google Scholar.
14. Callahan B. J., McMurdie P. J., Rosen M. J., Han A. W., Johnson A. J. A., Holmes S. P. (2016). DADA2: high-resolution sample inference from illumina amplicon data. Nat. Methods 13, 581–583. doi: 10.1038/nmeth.3869. CrossRef. Google Scholar.
15. Castellano-Hinojosa A., Boyd N. S., Strauss S. L. (2021). Impact of fumigants on non-target soil microorganisms: a review. J. Hazard. Mater. 427:128149. doi: 10.1016/j.jhazmat.2021.128149. CrossRef. Google Scholar.
16. Chaudhry V., Rehman A., Mishra A., Chauhan P. S., Nautiyal C. S. (2012). Changes in bacterial community structure of agricultural land due to long-term organic and chemical amendments. Microb. Ecol. 64, 450–460. doi: 10.1007/s00248-012-0025-y. CrossRef. Google Scholar.
17. Chen J., Zhou L., Din I. U., Arafat Y., Li Q., Wang J. et al. (2021). Antagonistic activity of Trichoderma spp. against fusarium oxysporum in rhizosphere of radix pseudostellariae triggers the expression of host defense genes and improves its growth under long-term monoculture system. Front. Microbiol. 12:579920. doi: 10.3389/fmicb.2021.579920. CrossRef. Google Scholar.
18. Chong J., Liu P., Zhou G., Xia J. (2020). Using microbiome analyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nat. Protoc. 15, 799–821. doi: 10.1038/s41596-019-0264-1. CrossRef. Google Scholar.
19. Clarke K. R. (1993). Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 18, 117–143. doi: 10.1111/j.1442-9993.1993.tb00438.x. CrossRef. Google Scholar.
20. Dangi S. R., Tirado-Corbalá R., Gerik J., Hanson B. D. (2017). Effect of long-term continuous fumigation on soil microbial communities. Agronomy 7:37. doi: 10.3390/agronomy7020037. CrossRef. Google Scholar.
21. de Jesus Sousa J. A., Olivares F. L. (2016). Plant growth promotion by streptomycetes: ecophysiology, mechanisms and applications. Chem. Biol. Technol. Agric. 3, 1–12. doi: 10.1186/s40538-016-0073-5. CrossRef. Google Scholar.
22. Dhariwal A., Chong J., Habib S., King I. L., Agellon L. B., Xia J. (2017). Microbiome analyst: a web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 45, W180–W188. doi: 10.1093/nar/gkx295. CrossRef. Google Scholar.
23. Douglas G. M., Maffei V. J., Zaneveld J. R., Yurgel S. N., Brown J. R., Taylor C. M. et al. (2020). PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 38, 685–688. doi: 10.1038/s41587-020-0548-6. CrossRef. Google Scholar.
24. Durán P., Thiergart T., Garrido-Oter R., Agler M., Kemen E., Schulze-Lefert P. et al. (2018). Microbial interkingdom interactions in roots promote Arabidopsis survival. Cells 175, 973–983.e14. doi: 10.1016/j.cell.2018.10.020. CrossRef. Google Scholar.
25. Edwards J., Johnson C., Santos-Medellín C., Lurie E., Podishetty N. K., Bhatnagar S. et al. (2015). Structure, variation, and assembly of the root-associated microbiomes of rice. Proc. Natl. Acad. Sci. 112, E911–E920. doi: 10.1073/pnas.1414592112. CrossRef. Google Scholar.
26. Entz M. H., Penner K. R., Vessey J. K., Zelmer C. D., Thiessen Martens J. R. (2004). Mycorrhizal colonization of flax under long-term organic and conventional management. Can. J. Plant Sci. 84, 1097–1099. doi: 10.4141/p04-055. CrossRef. Google Scholar.
27. Fierer N., Lauber C. L., Ramirez K. S., Zaneveld J., Bradford M. A., Knight R. (2012). Comparative metagenomic, phylogenetic and physiological analyses of soil microbial communities across nitrogen gradients. ISME J. 6, 1007–1017. doi: 10.1038/ismej.2011.159. CrossRef. Google Scholar.
28. Gazdag O., Kovács R., Parádi I., Füzy A., Ködöböcz L., Mucsi M. et al. (2019). Density and diversity of microbial symbionts under organic and conventional agricultural management. Microbes Environ. 34, 234–243. doi: 10.1264/jsme2.me18138. CrossRef. Google Scholar.
29. Gosling P., Ozaki A., Jones J., Turner M., Rayns F., Bending G. D. (2010). Organic management of tilled agricultural soils results in a rapid increase in colonisation potential and spore populations of arbuscular mycorrhizal fungi. Agric. Ecosyst. Environ. 139, 273–279. doi: 10.1016/j.agee.2010.08.013. CrossRef. Google Scholar.
30. Hammer Ø., Harper D. A., Ryan P. D. (2001). PAST: paleontological statistics software package for education and data analysis. Palaeontol. Electron. 4:9. Google Scholar.
31. Haney C. H., Samuel B. S., Bush J., Ausubel F. M. (2015). Associations with rhizosphere bacteria can confer an adaptive advantage to plants. Nat. Plants 1, 1–9. doi: 10.1038/nplants.2015.51. CrossRef. Google Scholar.
32. Hartmann M., Frey B., Mayer J., Mäder P., Widmer F. (2015). Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 9, 1177–1194. doi: 10.1038/ismej.2014.210. CrossRef. Google Scholar.
33. Hartmann A., Rothballer M., Schmid M. (2008). Lorenz Hiltner, a pioneer in rhizosphere microbial ecology and soil bacteriology research. Plant Soil 312, 7–14. doi: 10.1007/s11104-007-9514-z. CrossRef. Google Scholar.
34. Ibekwe A. M., Papiernik S. K., Gan J., Yates S. R., Yang C. H., Crowley D. E. (2001). Impact of fumigants on soil microbial communities. Appl. Environ. Microbiol. 67, 3245–3257. doi: 10.1128/aem.67.7.3245-3257.2001. CrossRef. Google Scholar.
35. Igiehon N. O., Babalola O. O., Aremu B. R. (2019). Genomic insights into plant growth promoting rhizobia capable of enhancing soybean germination under drought stress. BMC Microbiol 19, 1–22. doi: 10.1186/s12866-019-1536-1. CrossRef. Google Scholar.
36. Katoh K., Misawa K., Kuma K. I., Miyata T. (2002). MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 30, 3059–3066. doi: 10.1093/nar/gkf436. CrossRef. Google Scholar.
37. Kumar S., Sindhu S. S., Kumar R. (2021). Biofertilizers: an ecofriendly technology for nutrient recycling and environmental sustainability. Curr. Res. Microb. Sci. 3:100094. doi: 10.1016/j.crmicr.2021.100094. CrossRef. Google Scholar.
38. Li J., Huang B., Wang Q., Li Y., Fang W., Yan D. et al. (2017). Effect of fumigation with chloropicrin on soil bacterial communities and genes encoding key enzymes involved in nitrogen cycling. Environ. Pollut. 227, 534–542. doi: 10.1016/j.envpol.2017.03.076. CrossRef. Google Scholar.
39. Li J., Wang C., Liang W., Liu S. (2021). Rhizosphere microbiome: the emerging barrier in plant-pathogen interactions. Front. Microbiol. 12:772420. doi: 10.3389/fmicb.2021.772420. CrossRef. Google Scholar.
40. Liao J., Xu Q., Xu H., Huang D. (2019). Natural farming improves soil quality and alters microbial diversity in a cabbage field in Japan. Sustain. For. 11:3131. doi: 10.3390/su11113131. CrossRef. Google Scholar.
41. Ling N., Wang T., Kuzyakov Y. (2022). Rhizosphere bacteriome structure and functions. Nat. Commun. 13, 836–813. doi: 10.1038/s41467-022-28448-9. CrossRef. Google Scholar.
42. Liu X., Cheng X., Wang H., Wang K., Qiao K. (2015). Effect of fumigation with 1,3-dichloropropene on soil bacterial communities. Chemosphere 139, 379–385. doi: 10.1016/j.chemosphere.2015.07.034. CrossRef. Google Scholar.
43. Lupatini M., Korthals G. W., De Hollander M., Janssens T. K., Kuramae E. E. (2017). Soil microbiome is more heterogeneous in organic than in conventional farming system. Front. Microbiol. 7:2064. doi: 10.3389/fmicb.2016.02064. CrossRef. Google Scholar.
44. Mao L., Wang Q., Yan D., Ma T., Liu P., Shen J. et al. (2014). Evaluation of chloropicrin as a soil fumigant against Ralstonia solanacarum in ginger (Zingiber officinale Rosc.) production in China. PLoS One 9:e91767. doi: 10.1371/journal.pone.0091767. CrossRef. Google Scholar.
45. Martin H. (2005). Manure Composting as a Pathogen Reduction Strategy. Ontario, Canada: Ministry of Agriculture & Food. Google Scholar.
46. Martin M. (2011). Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 17, 10–12. doi: 10.14806/ej.17.1.200. CrossRef. Google Scholar.
47. McGonigle T. P., Miller M. H., Evans D. G., Fairchild G. L., Swan J. A. (1990). A new method which gives an objective measure of colonization of roots by vesicular---arbuscular mycorrhizal fungi. New Phytol. 115, 495–501. doi: 10.1111/j.1469-8137.1990.tb00476.x. CrossRef. Google Scholar.
48. McPherson M. R., Wang P., Marsh E. L., Mitchell R. B., Schachtman D. P. (2018). Isolation and analysis of microbial communities in soil, rhizosphere, and roots in perennial grass experiments. JoVE 137:e57932. doi: 10.3791/57932. CrossRef. Google Scholar.
49. Meena R. S., Kumar S., Datta R., Lal R., Vijayakumar V., Brtnicky M. et al. (2020). Impact of agrochemicals on soil microbiota and management: a review. Landscape 9:34. doi: 10.3390/land9020034. CrossRef. Google Scholar.
50. Mendes R., Garbeva P., Raaijmakers J. M. (2013). The rhizosphere microbiome: significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 37, 634–663. doi: 10.1111/1574-6976.12028. CrossRef. Google Scholar.
51. Oehl F., Sieverding E., Mäder P., Dubois D., Ineichen K., Boller T. et al. (2004). Impact of long-term conventional and organic farming on the diversity of arbuscular mycorrhizal fungi. Oecologia 138, 574–583. doi: 10.1007/s00442-003-1458-2. CrossRef. Google Scholar.
52. Philippot L., Raaijmakers J. M., Lemanceau P., Van Der Putten W. H. (2013). Going back to the roots: the microbial ecology of the rhizosphere. Nat. Rev. Microbiol. 11, 789–799. doi: 10.1038/nrmicro3109. CrossRef. Google Scholar.
53. Phillips J. M., Hayman D. S. (1970). Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 55:158. doi: 10.1016/s0007-1536(70)80110-3. CrossRef. Google Scholar.
54. Quast C., Pruesse E., Yilmaz P., Gerken J., Schweer T., Yarza P. et al. (2012). The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 41, D590–D596. doi: 10.1093/nar/gks1219. CrossRef. Google Scholar.
55. R Core Team. (2020). R: A Language and Environment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing. Google Scholar.
56. Rokunuzzaman M., Hayakawa A., Yamane S., Tanaka S., Ohnishi K. (2016). Effect of soil disinfection with chemical and biological methods on bacterial communities. Egypt. J. Basic Appl. Sci. 3, 141–148. doi: 10.1016/j.ejbas.2016.01.003. CrossRef. Google Scholar.
57. Sharma S. B., Sayyed R. Z., Sonawane M., Trivedi M. H., Thivakaran G. A. (2016). Neurospora sp. SR8, a novel phosphate solubiliser from rhizosphere soil of sorghum in Kachchh, Gujarat, India. Indian J. Exp. Biol. 54, 644–649. Google Scholar.
58. Singh R. L., Mondal S. (Eds.) (2017). Biotechnology For Sustainable Agriculture: Emerging Approaches and Strategies. Woodhead publishing. Google Scholar.
59. Sinong G. F., Yasuda M., Nara Y., Lee C. G., Dastogeer K. M. G., Tabuchi H. et al. (2021). Distinct root microbial communities in nature farming rice harbor bacterial strains with plant growth-promoting traits. Front. Sustain. Food Systems 4:629942. doi: 10.3389/fsufs.2020.629942. CrossRef. Google Scholar.
60. Smith S. E., Jakobsen I., Grønlund M., Smith F. A. (2011). Roles of arbuscular mycorrhizas in plant phosphorus nutrition: interactions between pathways of phosphorus uptake in arbuscular mycorrhizal roots have important implications for understanding and manipulating plant phosphorus acquisition. Plant Physiol. 156, 1050–1057. doi: 10.1104/pp.111.174581. CrossRef. Google Scholar.
61. Strauss S. L., Kluepfel D. A. (2015). Anaerobic soil disinfestation: a chemical-independent approach to pre-plant control of plant pathogens. J. Integr. Agric. 14, 2309–2318. doi: 10.1016/s2095-3119(15)61118-2. CrossRef. Google Scholar.
62. Sugiyama A. (2019). The soybean rhizosphere: Metabolites, microbes, and beyond—A review. J. Adv. Res. 19, 67–73. doi: 10.1016/j.jare.2019.03.005. CrossRef. Google Scholar.
63. Thiergart T., Zgadzaj R., Bozsóki Z., Garrido-Oter R., Radutoiu S., Schulze-Lefert P. (2019). Lotus japonicus symbiosis genes impact microbial interactions between symbionts and multikingdom commensal communities. MBio 10, e01833–e01819. doi: 10.1128/mbio.01833-19. CrossRef. Google Scholar.
64. UNITE Community (2019). UNITE general FASTA release for fungi. UNITE community. Version 18.11.2018. Google Scholar.
65. Vandamme P., Opelt K., Knöchel N., Berg C., Schönmann S., De Brandt E. et al. (2007). Burkholderia bryophila sp. nov. and Burkholderia megapolitana sp. nov., moss-associated species with antifungal and plant-growth-promoting properties. Int. J. Syst. Evol. Microbiol. 57, 2228–2235. doi: 10.1099/ijs.0.65142-0. CrossRef. Google Scholar.
66. Viaene T., Langendries S., Beirinckx S., Maes M., Goormachtig S. (2016). Streptomyces as a plant's best friend? FEMS Microbiol. Ecol. 92. doi: 10.1093/femsec/fiw119. CrossRef. Google Scholar.
67. Vurukonda S. S. K. P., Giovanardi D., Stefani E. (2018). Plant growth promoting and biocontrol activity of Streptomyces spp. as endophytes. Int. J. Mol. Sci. 19:952. doi: 10.3390/ijms19040952. CrossRef. Google Scholar.
68. Wickham H., Chang W., Wickham M. H. (2016). Package 'ggplot2'. Create elegant data visualisations using the grammar of graphics. Version 2, 1–189. Google Scholar.
69. Xiong W., Li R., Ren Y., Liu C., Zhao Q., Wu H. et al. (2017). Distinct roles for soil fungal and bacterial communities associated with the suppression of vanilla fusarium wilt disease. Soil Biol. Biochem. 107, 198–207. doi: 10.1016/j.soilbio.2017.01.010. CrossRef. Google Scholar.
70. Xiong J., Peng S., Liu Y., Yin H., Zhou L., Zhou Z. et al. (2022). Soil properties, rhizosphere bacterial community, and plant performance respond differently to fumigation and bioagent treatment in continuous cropping fields. Front. Microbiol. 13:2656. doi: 10.3389/fmicb.2022.923405. CrossRef. Google Scholar.
71. Xu H. L. (2001). Nature farming: history, principles and perspectives. J. Crop. Prod. 3, 1–10. doi: 10.1300/j144v03n01_01. CrossRef. Google Scholar.
72. Yan D., Wang Q., Song Z., Fang W., Wang Q., Li Y. et al. (2022). Activation effect of soil available nitrogen, manganese and cobalt after addition of different fumigants. Environ. Res. Commun. 4:041002. doi: 10.1088/2515-7620/ac64ed. CrossRef. Google Scholar.

Agyekum DVA, Kobayashi T, Dastogeer KMG, Yasuda M, Sarkodee-Addo E, Ratu STN, Xu Q, Miki T, Matsuura E and Okazaki S (2023) Diversity and function of soybean rhizosphere microbiome under nature farming. Front. Microbiol. 14:1130969. doi: 10.3389/fmicb.2023.1130969
Перевод статьи «Diversity and function of soybean rhizosphere microbiome under nature farming» авторов Agyekum DVA, Kobayashi T, Dastogeer KMG, Yasuda M, Sarkodee-Addo E, Ratu STN, Xu Q, Miki T, Matsuura E and Okazaki S., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык. Лицензия: CC BY. Изменения: переведено на русский язык
Фото: wikipedia




















































































































Комментарии (0)