Сравнение почвенных бактерий в органических и обычных теплицах: кто отвечает за азот и разложение органики
Органическое тепличное земледелие — это инновационная система, которая может обеспечивать высокую урожайность и здоровье агроэкосистемы. До сих пор не проводилось масштабных строгих исследований, сравнивающих общее сообщество бактерий и сообщество бактерий, участвующих в круговороте азота, в овощных почвах при органическом и традиционном управлении хозяйством. Было проведено исследование бактериального сообщества и циклов азота в почвах под органическими и традиционными теплицами на 30 участках, охватывающих семь типов почв с историей органического земледелия от 4 до 18 лет.
Аннотация
Сообщества общих бактерий, азотфиксаторов и аммоний-окисляющих бактерий изучали с помощью высокопроизводительного секвенирования генов 16S рРНК, nifH и amoA соответственно. Органическое тепличное земледелие не влияло на альфа-разнообразие. Бета-разнообразие среди общих (26 из 30 участков) и азотфиксирующих (17 из 19) бактерий различалось в зависимости от системы земледелия, однако различия в составе аммоний-окисляющих бактерий между двумя системами были обнаружены только на 6 участках. Несмотря на то, что влияние системы земледелия на большинство бактериальных родов варьировалось в зависимости от участка, органическое тепличное земледелие устойчиво отбирало несколько родов, возможно, участвующих в biodegradation (разложении) органического углерода с высокой молекулярной массой (Hyphomicrobium, Rubinisphaera, Aciditerrimonas, Planifilum, Phaselicystis и Ohtaekwangia), но подавляло предполагаемые аммоний-окисляющие (Nitrosospira, Nitrosopumilus) и азотфиксирующие (Bradyrhizobium) роды бактерий, согласно анализу 16S рРНК. Азотфиксирующие бактерии, относящиеся к кластеру nifH 1J, были предпочтительно связаны с органическим тепличным земледелием, в отличие от Paenibacillus borealis. Резюмируя, это исследование дает представление о сложных эффектах органического тепличного земледелия на сообщества общих, азотфиксирующих и аммоний-окисляющих бактерий в различных экологических условиях.
Введение
В Китае производство овощей в теплицах быстро росло, достигнув урожайности около 260 миллионов тонн в 2014 году, что составило 35% от общего производства овощей (Национальное бюро статистики Китайской Народной Республики [NBSC], 2014; Институт городского и сельского планирования и дизайна Чжунцзин Хуэйчэн [ZIURPD], 2014). Интенсивное сельскохозяйственное управление, такое как непрерывное выращивание и чрезмерное использование химических удобрений, пестицидов, орошения или обработки почвы, может поддерживать высокую, но краткосрочную продуктивность. Однако деградация почвы (Hannachi и др., 2015), значительные выбросы парниковых газов (Alam и др., 2019), загрязнение грунтовых вод (Yang и др., 2018), потеря органического вещества (Scotti и др., 2015) и накопление пестицидов (Chen и др., 2015) часто приводят к невозможности устойчивого производства, особенно в тепличных условиях. Адаптация органического сельскохозяйственного управления к тепличному производству овощей может смягчить эти неблагоприятные экологические проблемы. В целом, органическое тепличное земледелие обеспечивает большие экологические услуги, но меньшую (от 5 до 34%) продуктивность, чем традиционное земледелие (Bengtsson и др., 2005; Seufert и др., 2012). Благодаря ценовым премиям на органические продукты, органические фермы могут достигать уровней дохода, аналогичных или более высоких, чем у сопоставимых традиционных ферм (Qiao и др., 2018). В Китае общая площадь под органическое производство овощей достигла 148 500 гектаров к 2017 году, что составляет 6% от площади коммерческого выращивания овощей. Хотя является спорным вопрос о том, может ли органическое тепличное земледелие смягчить потепление климата (Searchinger и др., 2018; Smith и др., 2019), положительные эффекты органического тепличного земледелия на функции экосистемы, такие как плодородие почвы (Gomiero и др., 2011) и здоровье растений (Van Bruggen и др., 2016), являются очень перспективными.
Почвенные бактерии, обеспечивающие большую часть биоразнообразия в почвенных экосистемах, участвуют практически в каждом взаимодействии между культурой и почвой, включая круговорот углерода и азота в почве (Thorpe и Callaway, 2011; Che и др., 2018), рост и здоровье растений (Janvier и др., 2007; van Bruggen и др., 2015) и поддержание структуры почвы (Bonanomi и др., 2016). Например, аэробное окисление аммиака, опосредованное β- и γ-протеобактериями, является лимитирующей стадией нитрификации, которая связана с доступностью азота для растений, выщелачиванием нитратов в грунтовые воды или выбросами N2O (Di и Cameron, 2016). Азотфиксирующие бактерии могут преобразовывать атмосферный N2 в биологически полезную форму, и ген nifH, кодирующий субъединицу азотазоредуктазы (Kuypers и др., 2018), часто используется в качестве генетического маркера для молекулярного анализа азотфиксирующих бактерий (Levy-Booth и др., 2014). Функциональные популяции, участвующие в азотфиксации или нитрификации, связаны с плодородием почвы и высокой эффективностью использования азота (Levy-Booth и др., 2014). Свободноживущие или симбиотические азотфиксирующие бактерии способствуют получению биологически полезного азота, в то время как аммоний-окисляющие бактерии (AOB) или аммоний-окисляющие археи (AOA) могут окислять аммиак до нитрита и далее до нитрата, который может легко теряться при выщелачивании. Здесь мы изучали тепличное сельское хозяйство, где избыточный азот часто вносится для поддержания высокой продуктивности овощей, AOB была выбрана в качестве показательной микробной популяции, поскольку она более чувствительна к более высоким дозам азота (Hink и др., 2018).
Эффекты органического земледелия на почвенный микробиом сложны. Ранее его влияние на таксономический состав почвенного микробиома было исследовано в полевых (Dennert и др., 2018) и тепличных (Hartmann и др., 2015; Ding и др., 2019) условиях. Органическое земледелие повышает микробное обилие и активность почвы (Lori и др., 2017; Ouyang и др., 2018b) и изменяет состав микробных сообществ (Moeskops и др., 2010; Bonanomi и др., 2016). Однако влияние на микробное альфа-разнообразие (Lupatini и др., 2017; Chen и др., 2018) и специфические таксономические группы (Tuck и др., 2014) варьировало в разных исследованиях (Postma и др., 2008; Bonanomi и др., 2016) и во времени (Wu и др., 2019). Влияние органических удобрений на разнообразие и обилие аммоний-окисляющих бактерий было широко оценено (Chu и др., 2007; Ouyang и др., 2018a). Обилие или относительное обилие аммоний-окисляющих бактерий, как правило, ниже в органических системах, чем в традиционных или интегрированных системах земледелия (Wessen и др., 2011; Ding и др., 2019). В полевом исследовании на фермерском уровне состав сообщества аммоний-окисляющих бактерий и архей не показал различий между органической и интегрированной системами земледелия (Wessen и др., 2011). В тепличном эксперименте органическое земледелие изменило сообщество аммоний-окисляющих бактерий (Ding и др., 2019). Сельскохозяйственное управление может обогащать обилие и разнообразие азотфиксирующих бактерий (Köberl и др., 2016), но прямое сравнение между органической и традиционной системами земледелия встречается редко (Ding и др., 2019). Эффекты органического тепличного земледелия на микробное разнообразие почвы, вероятно, зависят от контекста. Здесь мы предположили, что органическое тепличное земледелие может все же вызывать устойчивые, показательные изменения в сообществах общих, азотфиксирующих и аммоний-окисляющих бактерий в почвенном микробиоме в различных экологических условиях. Это знание важно для углубления нашего понимания механизмов, связанных с экологическими услугами, предоставляемыми органическим тепличным земледелием.
В настоящем исследовании в июле 2017 года было проведено обследование, охватывающее тридцать участков в Китае, для изучения взаимодействия между органическим тепличным земледелием и почвенным микробиомом. Разнообразие общих, азотфиксирующих и аммоний-окисляющих бактерий было изучено с помощью высокопроизводительного секвенирования ПЦР-амплифицированных фрагментов генов 16S рРНК, nifH и amoA. Цели настоящего исследования состояли в том, чтобы (1) изучить взаимодействие между органическим тепличным земледелием и общими бактериями, азотфиксаторами и аммоний-окисляющими бактериями на разных участках; (2) выявить ключевой фактор, лежащий в основе этого взаимодействия; и (3) выявить устойчивые, предсказуемые изменения в этих микробных популяциях.
Материалы и методы
Отбор почвенных образцов
Было выбрано в общей сложности 30 участков, расположенных в регионах с наибольшим количеством органических ферм в Китае (РИСУНОК 1). Эти участки представляют от 4 до 18 лет органического тепличного управления и семь типов почв. Восемь и шесть участков находились в Пекине и Шанхае соответственно (РИСУНОК 1). Все изученные органические фермы были сертифицированы. Органическое тепличное земледелие осуществлялось в соответствии с основными стандартами Международной федерации движений за органическое сельское хозяйство (IFOAM) с использованием биологических и физических методов защиты растений и органических удобрений (навоз и коммерческие органические удобрения) (ТАБЛИЦА 1). На традиционных фермах часто использовались как органические, так и минеральные удобрения, а также химические средства для борьбы с сорняками (глифосат и оксаламин), болезнями (азоксистробин и тиофанат-метил) и насекомыми (хлорфенапир и люфенурон), а также синтетические регуляторы роста растений (ТАБЛИЦА 1). Для каждого участка путем опроса местных фермеров были выбраны три соседние традиционные и органические теплицы с одинаковой культурой (томат, огурец, баклажан или зеленый перец), одинаковыми типами почв и схожими севооборотами. Для каждой повторности было собрано десять проб верхнего слоя почвы (0–20 см) (приблизительно 1 кг). Образцы были транспортированы в лабораторию в охлаждаемом контейнере и хранились при температуре −20°C до экстракции ДНК, которая выполнялась после просеивания образцов через сито с размером ячеек 2 мм. Физико-химические свойства почвы: общий азот (TN), общий углерод (TC), pH, электропроводность (EC), плотность, доступный азот (AN), фосфор по Ольсену (Olsen P), доступный калий (K) и катионообменная способность (CEC) были определены в соответствии со стандартными протоколами (Bao, 2000).
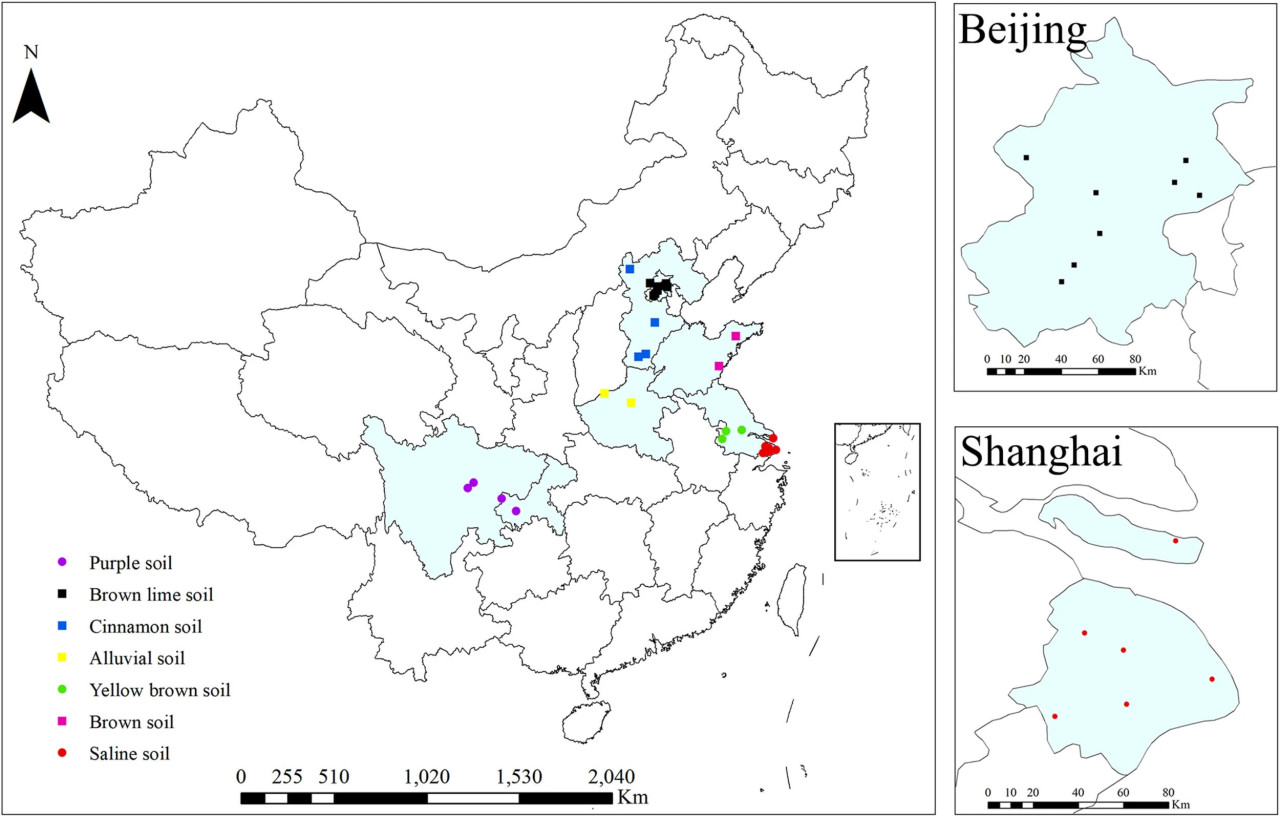
РИСУНОК 1 Расположение органических и традиционных ферм для тепличного производства овощей. Цвета на точках указывают типы почв: черный (Пекин, бурая известковая почва), синий (Хэбэй, циннамонная почва), желтый (Хэнань, аллювиальная почва), пурпурный (Шаньдун, бурая почва), зеленый (Цзянсу, желто-бурая почва), красный (Шанхай, солончаковая почва) и фиолетовый (Чэнду/Чунцин, пурпурная почва).
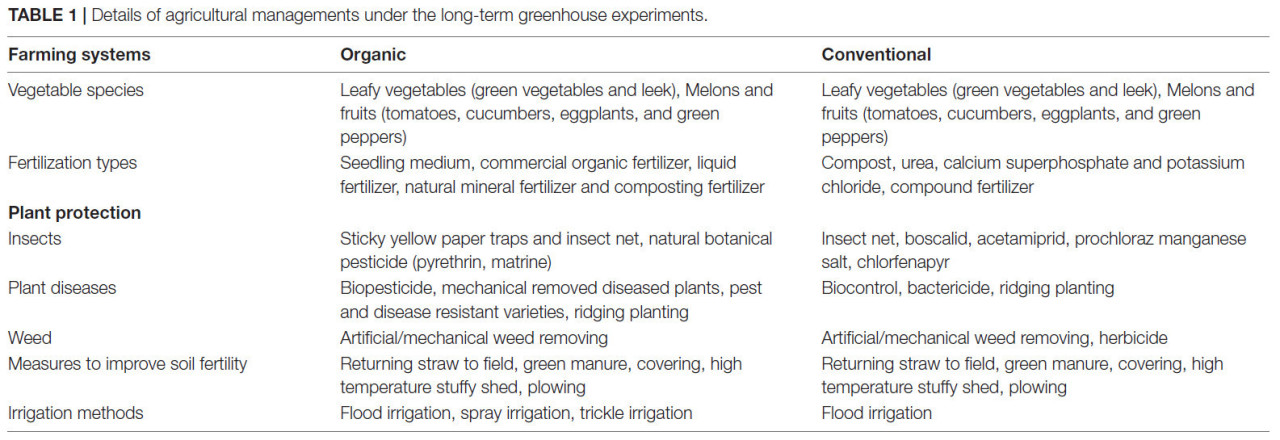
ТАБЛИЦА 1 Детали сельскохозяйственного управления в условиях длительных тепличных экспериментов.
ПЦР-амплификация генов 16S рРНК, amoA и nifH
Общая ДНК сообщества была экстрагирована с использованием набора FastDNA Spin Kit for Soil (MP Biomedicals, Санта-Ана, Карлсбад, Калифорния, США) в соответствии с инструкциями производителя. Праймеры, использованные для амплификации генов 16S рРНК, amoA и nifH, приведены во Вспомогательной таблице S1 (Rotthauwe и др., 1997; Poly и др., 2001; Tamaki и др., 2011). Продукты ПЦР были очищены из геля, количественно определены и объединены в эквимолярных количествах для каждого образца для высокопроизводительного секвенирования на платформе Illumina Hi-Seq с использованием набора реагентов Reagent Kit v2 2 × 250 bp (платформа Hi-Seq 2500).
Биоинформатический и статистический анализы
Последовательности высокого качества (длина >300 п.н., без неоднозначного основания «N» и средний показатель качества оснований >30) были использованы для последующего анализа. Последовательности генов 16S рРНК и nifH были собраны с использованием программного пакета mothur v1.39 (Schloss и др., 2009) и в дальнейшем распределены по каждому образцу на основе штрих-кодов и праймеров. Двунаправленные последовательности генов amoA были распределены по образцам отдельно из-за недостаточной длины прочтений. Денизинг, назначение OTU и классификация гена 16S рРНК были выполнены, как описано ранее (Schloss и др., 2009; Caporaso и др., 2010; Cole и др., 2013; Ding и др., 2012, 2019). Для генов nifH и amoA был выполнен автономный анализ BLASTX против соответствующих последовательностей функциональных генов, загруженных из базы данных RDP, для выявления правильных рамок трансляции. Для дальнейшего анализа были включены только те последовательности, которые не имели стоп-кодонов в пределах выведенных аминокислотных последовательностей. Выведенные аминокислотные последовательности были дополнительно подвергнуты анализу hmmscan для идентификации последовательностей генов nifH или amoA. Hmm-профиль был получен с веб-сайта RDP FunGene. Аминокислотные последовательности nifH были дополнительно распределены по OTU (>95% идентичности последовательностей) с использованием программного обеспечения vsearch. Репрезентативные OTU nifH были распределены по различным подгруппам с использованием курируемой базы данных, как описано Zehr и др. (2017). Для генов amoA выбранная последовательность была сгруппирована в OTU с 80% идентичностью путем анализа репрезентативных последовательностей различных подгрупп, как описано ранее (Li и др., 2011; Guo и др., 2017). Дискриминационные таксоны, OTU и различия в составе сообщества и сети совместной встречаемости были в основном исследованы, как описано ранее (Hothorn и др., 2008; Li и др., 2019). Кратко: значительные различия в составе микробного сообщества были сравнены с помощью теста перестановок (Kropf и др., 2004) с использованием рассчитанного попарного расстояния Брея-Кертиса. На основе пакета «Vegan» индексы альфа-разнообразия (Chao1, Simpson и выравненность Пилу) были рассчитаны путем 1000 повторных выборок равного количества последовательностей из каждого образца для уменьшения смещения, вызванного количеством прочтений или индивидуальным отбором проб. Сравнение состава сообщества, идентификация таксонов со значительно различающейся относительной численностью и сетевой анализ были выполнены, как описано ранее (Li и др., 2019). Анализ классификации случайного леса также был выполнен с помощью R-дополнения «randomForest» (Liaw и Wiener, 2002) для выявления ключевых таксонов для систем земледелия. Эта сеть была проанализирована с использованием программного обеспечения Gephi (версия 0.91) (Bastian и др., 2009). Все статистические анализы и построение графиков были выполнены с использованием программного обеспечения R 3.1.2, и эти инструменты были реализованы в экземпляре Galaxy.
Результаты
Влияние органического тепличного земледелия на альфа- и бета-разнообразие почвенных бактерий
Бактериальное разнообразие почв с органических и традиционных ферм на 30 участках было изучено с помощью секвенирования гена 16S рРНК. Proteobacteria, Acidobacteria, Actinobacteria, Firmicutes, Bacteroidetes, Chloroflexi, Planctomycetes, Gemmatimonadetes и Cyanobacteria доминировали во всех почвенных образцах (Вспомогательный рисунок S1). Альфа-разнообразие (богатство Chao1 и индекс выравненности Пилу) не подвергалось влиянию органического тепличного земледелия и было значительно связано с pH почвы (Chao1: R2 = 0.23, P = 0; выравненность Пилу: R2 = 0.12, P = 0) (РИСУНОК 2A). Бета-разнообразие явно различалось между органической и традиционной системами земледелия на 26 из 30 изученных участков (РИСУНОК 2B и Вспомогательный рисунок S2). Пермутационный анализ также подтвердил, что бактериальное сообщество было значительно (p = 0; d = 20.2%) различным между органической и традиционной системами земледелия. Однако среднее несходство между органической и традиционной системами земледелия не увеличивалось с периодом органического земледелия (Вспомогательный рисунок S3). Гетерогенность состава сообщества была немного выше в органической системе, чем в традиционной (РИСУНОК 2C).
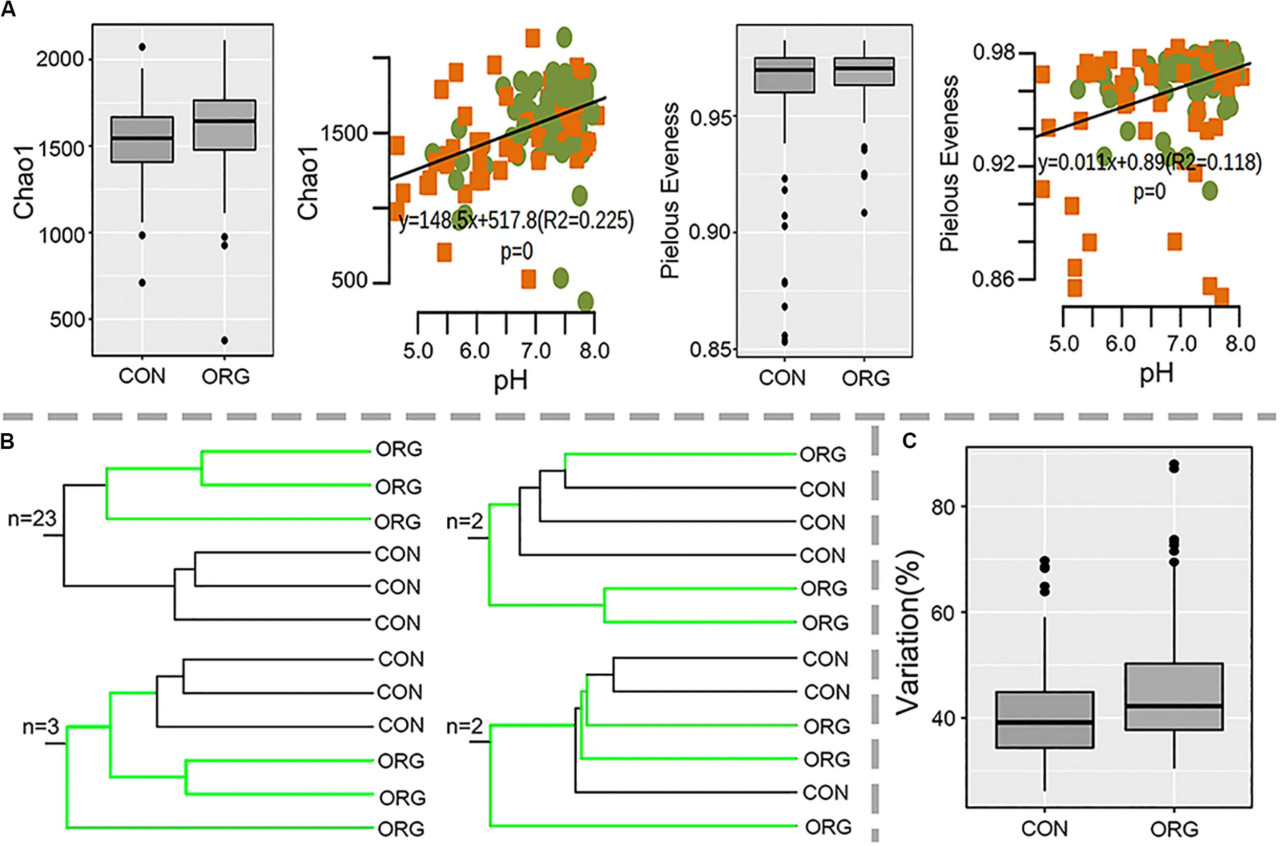
РИСУНОК 2 Альфа- и бета-разнообразие микробных сообществ бактерий в почвах при органической (ORG) и традиционной (CON) системах земледелия на 30 участках. (A) Индексы Chao1 и выравненности Пилу и их корреляции с pH; зеленые точки и оранжевые квадраты указывают на органическое и традиционное земледелие соответственно. (B) Количество участков со схематичной UPGMA кластеризацией. (C) Вариация бета-разнообразия в пределах органической (ORG) или традиционной (CON) систем земледелия.
Общие и специфичные для участка таксоны, связанные с органическим тепличным земледелием
Роды со значительно (P < 0.05) различающейся относительной численностью между системами земледелия были идентифицированы для тех 26 участков, где состав сообщества различался между системами земледелия. Эффекты органического тепличного земледелия на большинство родов были непостоянными, о чем свидетельствует низкая частота или зависимая от участка реакция на системы земледелия (РИСУНОК 3A). Девяносто четыре рода, которые часто (>12 участков) различались по относительной численности между системами земледелия, были дополнительно проанализированы на предмет их моделей реагирования (РИСУНОК 3B). Несмотря на широкое географическое распределение изученных участков, роды, которые были предпочтительно (Группа 1) или отрицательно (Группа 5 и 6) связаны с органическим тепличным земледелием, были все же идентифицированы (РИСУНОК 3B). Hyphomicrobium, Rubinisphaera, Aciditerrimonas, Planifilum, Phaselicystis и Ohtaekwangia, известные как хемоорганотрофные микроорганизмы, были предпочтительно связаны с органическим тепличным земледелием (РИСУНОК 3B). Bradyrhizobium, Nitrosospira, Nitrosopumilus, Gaiella, Arthrobacter, Flavisolibacter, Acidobacter Gp1, GP3, Gp4 и Gp25 были отрицательно связаны с органическим тепличным земледелием (РИСУНОК 3B). Интересно, что 23 из этих родов вошли в топ-50 наиболее влиятельных родов между системами земледелия, как показал анализ случайного леса (РИСУНОК 3B). Примечательно, что семнадцать из них продемонстрировали устойчивые модели реагирования на органическое земледелие (РИСУНОК 3B). Эти результаты показали, что органическое тепличное земледелие, как правило, отбирает бактерии, разлагающие органическое вещество, но подавляет азотфиксирующие, аммоний-окисляющие и олиготрофные бактерии.
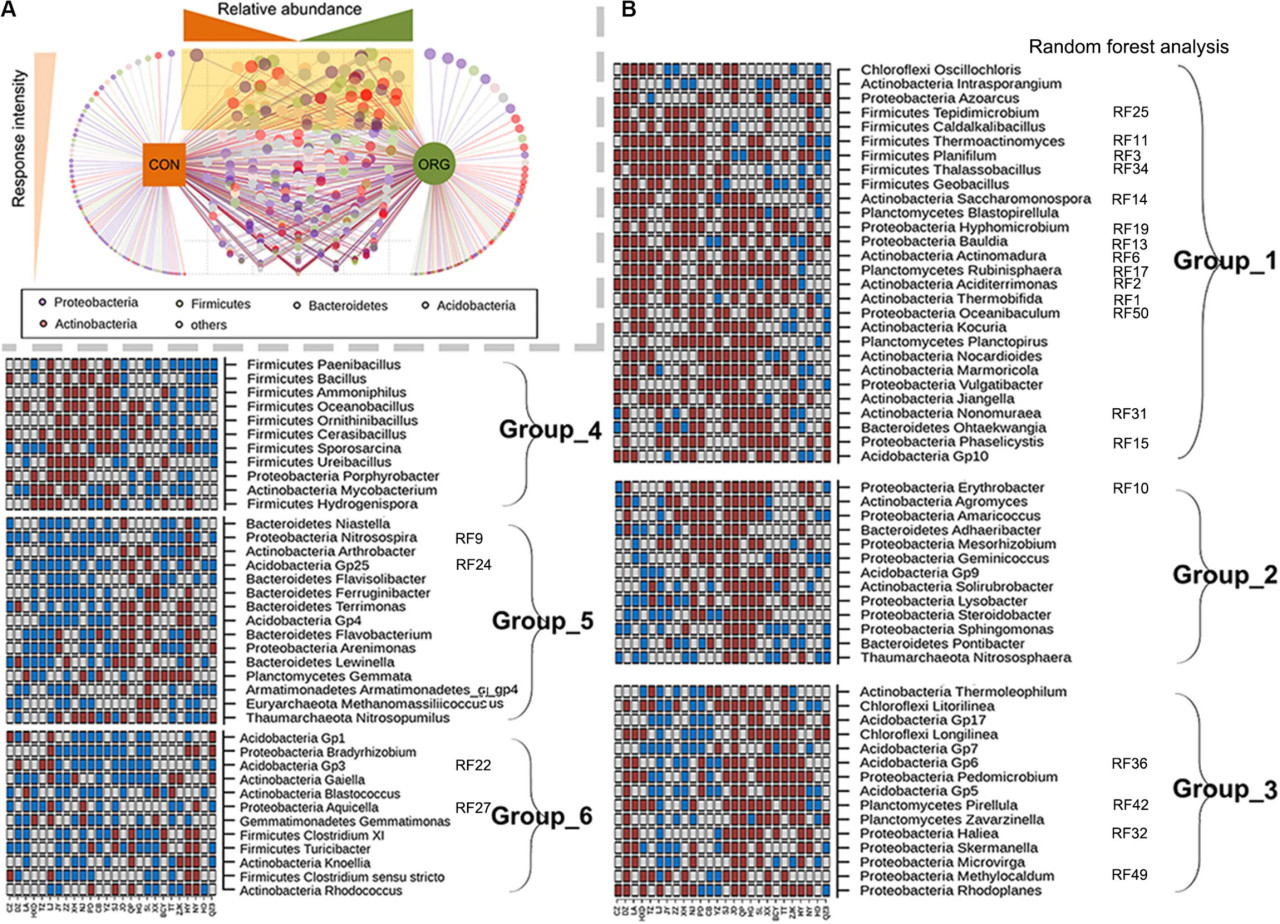
РИСУНОК 3 Бактериальные роды со значительно (p < 0.05) различающейся относительной численностью между системами земледелия и их связь с различными физико-химическими свойствами почвы. (A) Частота, с которой род является дискриминационным между системами земледелия. Светло-желтый треугольник указывает на повышенную частоту дискриминации рода между системами земледелия. (B) Группы родов со сходными моделями реагирования на системы земледелия, которые вошли в 50 наиболее влиятельных родов на всех участках; ORG, органическая система земледелия; CON, традиционная система земледелия. Коричневые и голубые квадраты указывают на значительно (p < 0.05) более высокую или более низкую относительную численность в ORG по сравнению с CON соответственно. Серые квадраты указывают на отсутствие значимой разницы между системами земледелия.
Разнообразие и состав сообщества азотфиксирующих популяций
Разнообразие азотфиксирующих бактерий было изучено с помощью высокопроизводительного секвенирования гена nifH. Эффекты органического тепличного земледелия были успешно изучены только на 19 участках, где было получено достаточное количество последовательностей по крайней мере для двух образцов на систему земледелия. Большинство последовательностей попадали в кластеры nifH 1 и 3 (РИСУНОК 4A), которые были в основном связаны с Proteobacteria, Cyanobacteria и Firmicutes. Альфа-разнообразие (индексы богатства Chao1 и выравненности Пилу) не подвергалось влиянию органического тепличного земледелия (РИСУНОК 4B), но было значительно связано с CEC (Chao1: R2 = 0.09, P = 0; выравненность Пилу: R2 = 0.08, P = 0.001) (РИСУНОК 4B). Состав сообщества азотфиксирующих популяций различался между органическим и традиционным земледелием на 17 из 19 участков (РИСУНОК 4C и Вспомогательный рисунок S4). Относительная численность подгруппы J кластера nifH 1 была значительно выше в почве, управляемой органически, на девяти участках (РИСУНОК 4D), где pH почвы, K и плотность были значительно ниже, чем на других участках (РИСУНОК 4E). Относительная численность кластера 3 и подгрупп G и C кластера 1, как правило, была отрицательно связана с органическим тепличным земледелием (РИСУНОК 4D). Было обнаружено расходящееся соотношение подгруппы A кластера 1 с органическим тепличным земледелием (РИСУНОК 4D). Доминирующие (топ-5) OTU в каждом образце были выбраны для выявления ключевых азотфиксирующих популяций, реагирующих на органическое тепличное земледелие. Семьдесят из 81 OTU варьировали между двумя системами земледелия на одном или нескольких участках, и большинство из этих OTU были идентифицированы как кластер nifH 1 или 3 (РИСУНОК 4F). Интересно, что относительная численность гена nifH, связанного с Paenibacillus borealis, была отрицательно связана с органическим тепличным земледелием (РИСУНОК 4G). Некоторые другие азотфиксирующие популяции были расходящимся образом связаны с органическим тепличным земледелием (РИСУНОК 4G). Предполагаемые образы жизни этих популяций разнообразны, включая симбиотический (Bradyrhizobium), свободноживущий (Azoarcus, Paenibacillus и Bacteroidales) и анаэробный (Desulfofustis, Desulfovibrio) образы жизни (РИСУНОК 4G). Анализ избыточности (RDA) показал, что pH почвы, TC и CEC могут объяснить большую часть вариаций этих OTU в большом масштабе (РИСУНОК 4H).
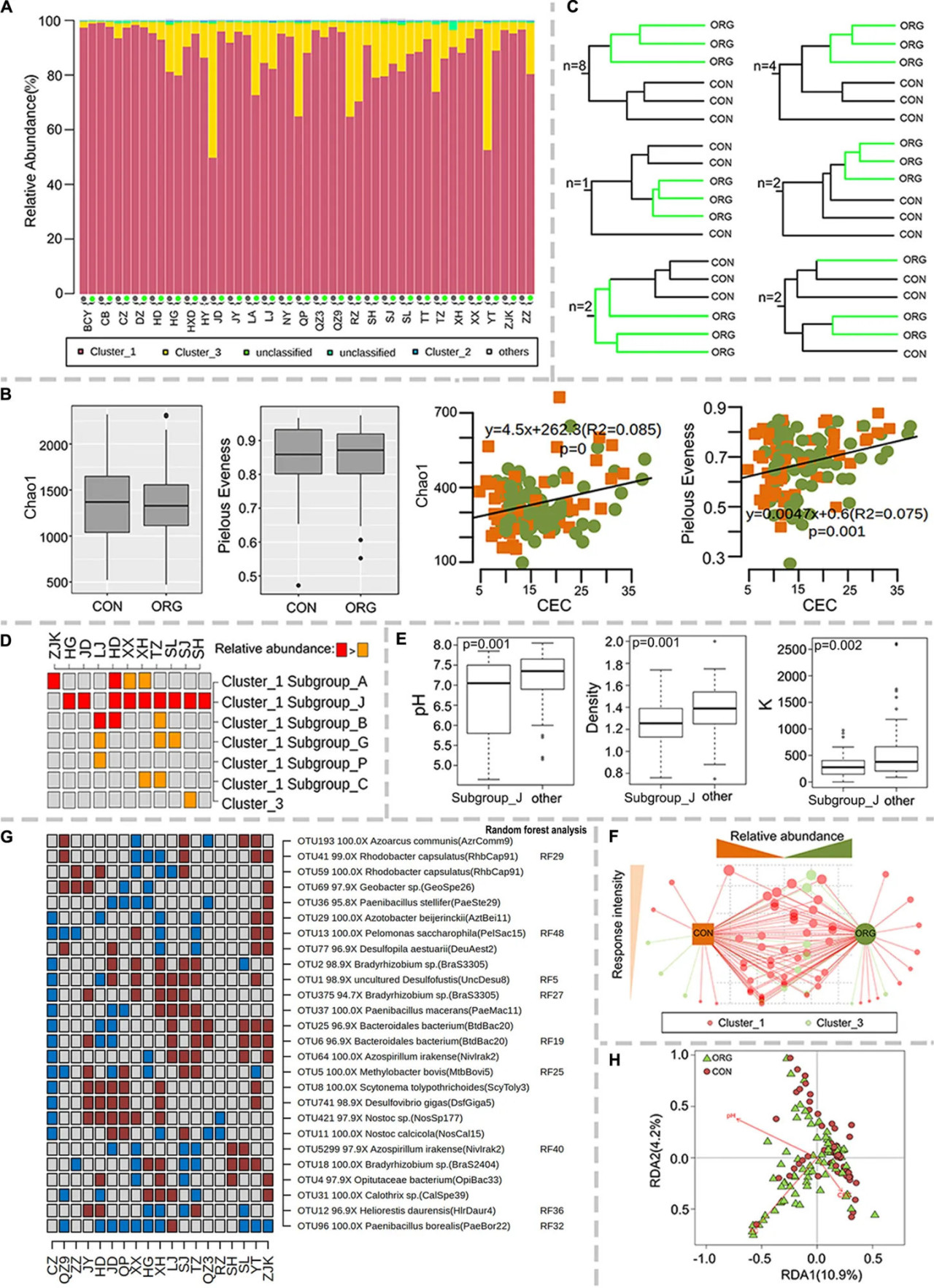
РИСУНОК 4 Связь разнообразия азотфиксирующих популяций с системой земледелия. (A) Относительная численность основных кластеров гена nifH, (B) индексы Chao1 и выравненности Пилу и их корреляции с CEC; зеленые точки и оранжевые квадраты указывают на органическое и традиционное земледелие соответственно. (C) Количество участков со схематичной UPGMA кластеризацией. (D) Дискриминационные подгруппы генов nifH. (E) Ящичковая диаграмма различий в уровнях pH, плотности и K между участками с повышенной и неповышенной относительной численностью подгруппы J кластера nifH 1. (F) Связь каждого азотфиксирующего OTU с системой земледелия. Размер точек указывает на частоту, с которой каждый OTU является дискриминационным между системами земледелия. Тенденция каждого OTU к органическим или традиционным системам земледелия была указана их расстоянием до систем земледелия. (G) Дискриминационные азотфиксирующие OTU между системами земледелия, ранжированные в 50 наиболее влиятельных OTU. Коричневые и голубые квадраты указывают на значительно (p < 0.05) более высокую или более низкую относительную численность в ORG по сравнению с CON соответственно. Серые квадраты указывают на отсутствие значимой разницы между системами земледелия. (H) Анализ избыточности (RDA) почвенных характеристик и азотфиксирующих микробных сообществ. ORG, органическая система земледелия; CON, традиционная система земледелия.
Разнообразие и состав сообщества аммоний-окисляющих популяций
Здесь эффекты органического тепличного земледелия на аммоний-окисляющие бактерии были проанализированы на 16 участках. Разнообразие последовательностей гена amoA также было высоким, достигая 6 769 OTU (>80% идентичности), большинство из которых были связаны с бактериями, подобными Nitrosospira, Nitrosovibrio и Nitrosomonas. Опять же, альфа-разнообразие amoA не подвергалось влиянию органического тепличного земледелия, и Chao1 был значительно связан с TC (прямой: R2 = 0.097 P = 0.001; обратный: R2 = 0.036, P = 0.035) (РИСУНОК 5A). Состав сообщества аммоний-окисляющих бактерий различался между органической и традиционной системами земледелия только на шести участках (РИСУНОК 5B и Вспомогательные рисунки S5, S6). OTU amoA, связанные с бактериями, подобными Nitrosovibrio, как правило, были отрицательно связаны с органическим тепличным земледелием, в отличие от бактерий, подобных Nitrosospira (Вспомогательный рисунок S7). Эти результаты показали, что эффекты органического тепличного земледелия на состав аммоний-окисляющих бактерий были фрагментарными.
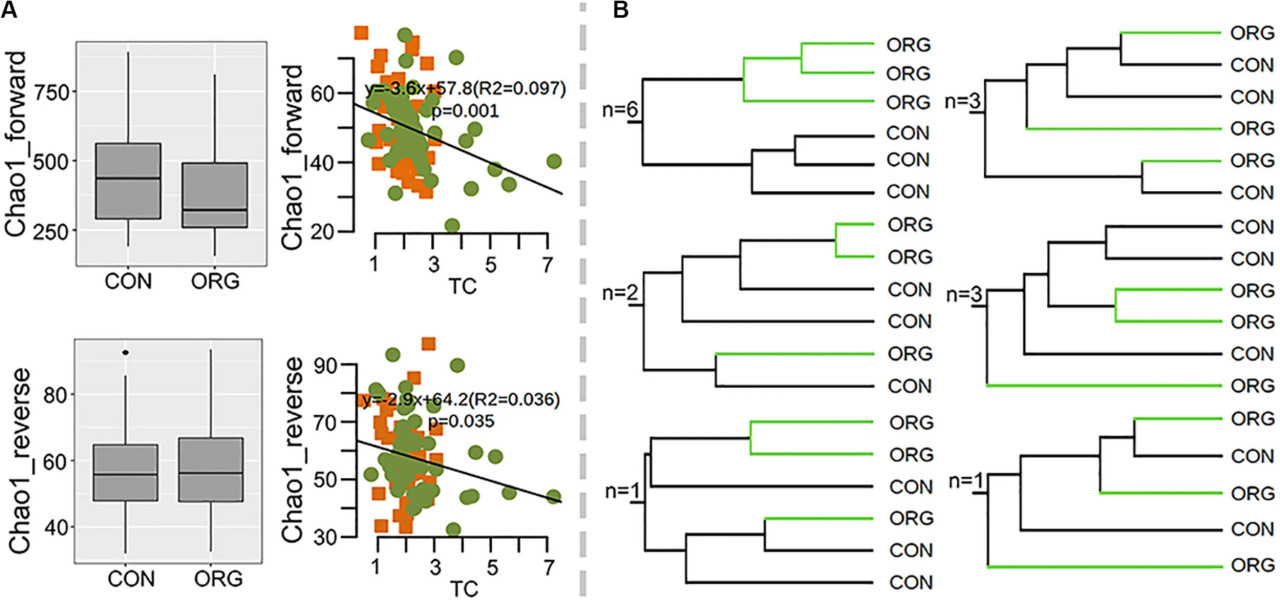
РИСУНОК 5 Связь разнообразия аммоний-окисляющих бактерий с системой земледелия. (A) Индексы Chao1 аммоний-окисляющих бактерий, выявленные с помощью прямого и обратного наборов данных, и их корреляции с TC; зеленые точки и оранжевые квадраты указывают на органическое и традиционное земледелие соответственно. (B) Количество участков со схематичной UPGMA кластеризацией. Коричневые и голубые квадраты указывают на значительно (p < 0.05) более высокую или более низкую относительную численность в ORG по сравнению с CON соответственно. Серые квадраты указывают на отсутствие значимой разницы между системами земледелия. ORG, органическая система земледелия; CON, традиционная система земледелия.
Связи между общими, азотфиксирующими и аммоний-окисляющими популяциями и физико-химическими свойствами исследуемых участков
Известно, что почвенные микробные сообщества чувствительны к изменениям окружающей среды, таким как физико-химические характеристики и изменение климата, что приводит к различиям в микробных сообществах исследуемых участков. Доминирующие (>1% относительной численности) общие (112), азотфиксирующие (177) и аммоний-окисляющие (97) бактериальные OTU были подвергнуты анализу сетей совместной встречаемости (Вспомогательный рисунок S8). В среднем 37.3% выбранных таксонов демонстрировали значительную совместную встречаемость друг с другом, и эти роды были в основном связаны с Proteobacteria (16), Acidobacteria (13), Bacteroidetes (6), Firmicutes (5), Actinobacteria (4) и Chloroflexi (3) (Вспомогательный рисунок S8). Среди этих типов самая высокая доля совместно встречающихся родов была связана с Acidobacteria — типом, который часто называют k-стратегами (Вспомогательный рисунок S8). Эти совместно встречающиеся микробные популяции образовали 11 основных микробных узлов (более пяти узлов) по данным сетевого анализа (РИСУНОК 6A). Интересно, что общие таксоны, а также азотфиксирующие и аммоний-окисляющие бактерии в основном образовывали отдельные узлы, что свидетельствует о низкой корреляции доминирующих бактерий с обеими функциональными популяциями (РИСУНОК 6A). Относительная численность различных узлов сильно варьировала между участками, но демонстрировала небольшую связь с системами земледелия (Вспомогательные рисунки S9A–C). pH почвы и CEC были двумя основными факторами, влияющими на относительную численность трех основных узлов общих бактерий (зеленый, голубой и синий) (РИСУНОК 6B). Значения pH почвы, CEC, TC, TN и EC были тесно связаны с составом азотфиксирующих бактерий, причем пять основных узлов были значительно связаны с pH, TC или CEC (РИСУНОК 6C). Интересно, что относительная численность оранжевого узла имела тенденцию быть высокой в условиях нейтрального pH, в отличие от пурпурного узла (РИСУНОК 6C). В дополнение к pH, два узла (красный или коричневый) были положительно связаны с CEC, а желтый узел был положительно связан с TC (РИСУНОК 6C). Доминирующие OTU аммоний-окисляющих бактерий образовали два основных узла (РИСУНОК 6D). Относительная численность зеленого узла имела тенденцию к снижению, когда pH был ниже 6.5 (РИСУНОК 6D), в то время как противоположная тенденция наблюдалась для черного узла (РИСУНОК 6D). RDA дополнительно показал, что pH почвы объяснял значительную вариацию внутри совместно встречающихся популяций для всех трех популяций (РИСУНКИ 6B–D). Эти результаты показывают, что pH почвы был ключевым фактором, формирующим встречаемость микробов в сообществах общих, азотфиксирующих и аммоний-окисляющих бактерий.
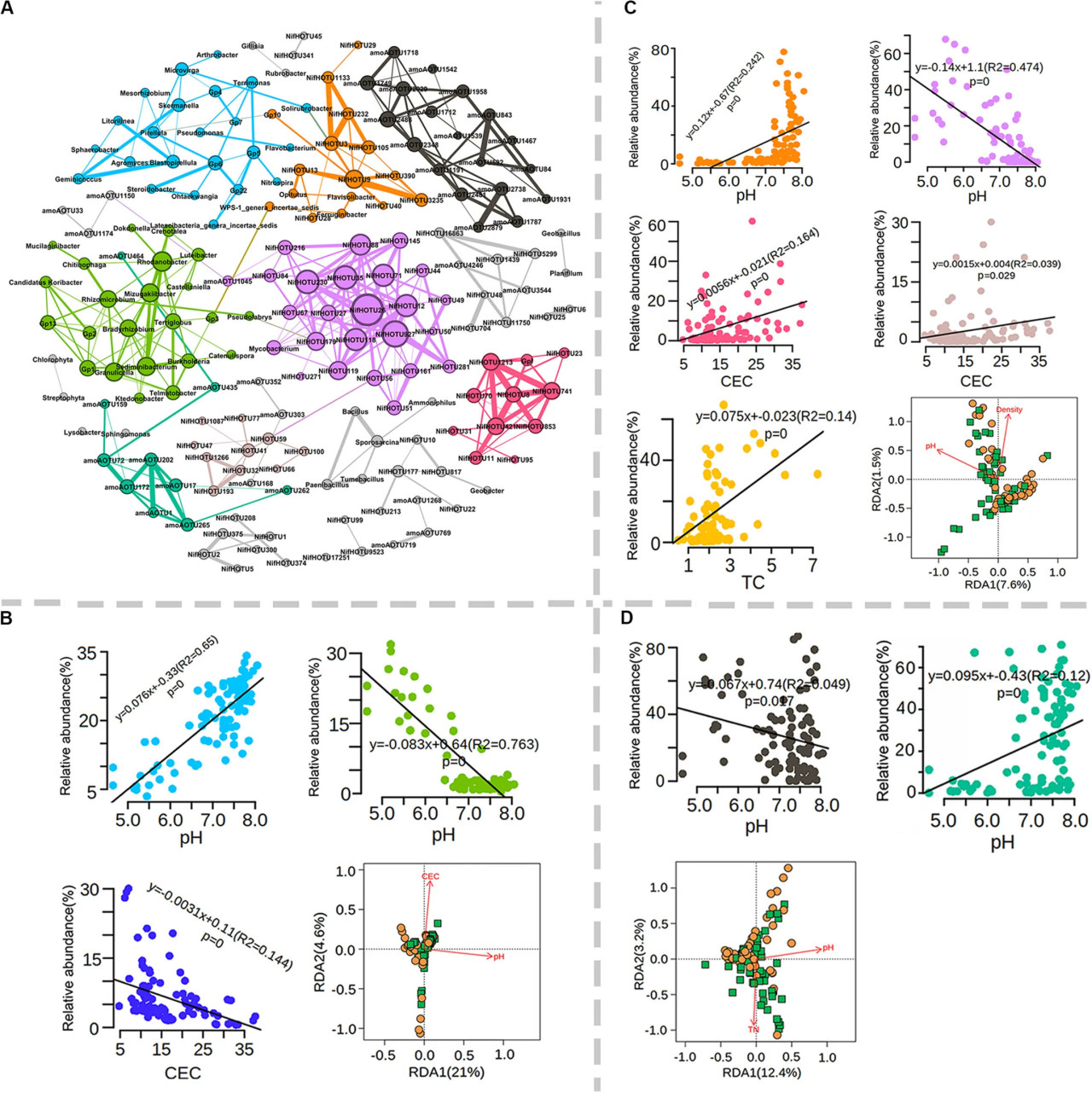
РИСУНОК 6 Анализ сетей совместной встречаемости доминирующих общих бактерий, азотфиксаторов и аммоний-окисляющих бактерий. (A) Сеть совместной встречаемости и основные микробные узлы, обозначенные разными цветами. Количество и доля совместно встречающихся микробных популяций; связь микробных узлов с физико-химическими свойствами почвы для общих бактерий (B), азотфиксаторов (C) и аммоний-окисляющих бактерий (D).
Обсуждение
Понимание взаимодействия между органическим земледелием и почвенным микробиомом важно для повышения экологических выгод органического земледелия для устойчивого производства продуктов питания. Здесь было проведено масштабное обследование почв с широким диапазоном физико-химических свойств, географических регионов и периодов органического земледелия для изучения влияния на разнообразие общих, азотфиксирующих и аммоний-окисляющих бактерий, что предоставляет возможность изучить влияние органического земледелия на почвенный микробиом. Кроме того, органическое тепличное земледелие может вызывать устойчивые, предсказуемые изменения в сообществах общих, азотфиксирующих и аммоний-окисляющих бактерий в разных почвах. Это знание важно для углубления нашего понимания влияния органического земледелия на почвенный микробиом в тепличных условиях в разных пространствах.
Органическое земледелие изменило состав бактериального сообщества, но не повлияло на альфа-разнообразие и не увеличило гетерогенность сообщества в тепличных условиях
Ранее эффекты органического земледелия на почвенные микробные сообщества были обнаружены как в тепличных, так и в полевых условиях на нескольких изученных участках (Hartmann и др., 2015; Bonanomi и др., 2016; Ding и др., 2019). Однако пространственная и временная изменчивость обычны для почвенных микробных сообществ (Uksa и др., 2014; Hill и др., 2016); таким образом, все еще неясно, может ли органическое тепличное земледелие вызывать изменения в микробном разнообразии почвы. Здесь были обнаружены различные бактериальные составы между органической и традиционной системами тепличного земледелия на 26 из 30 участков. Эти данные показали, что органическое тепличное земледелие было в значительной степени важным драйвером бактериальных сообществ на разных участках. Альфа-разнообразие почвенных бактерий не различалось в зависимости от системы земледелия, что согласуется с другими исследованиями (Prober и др., 2015; Banerjee и др., 2019). Оно было значительно связано с pH почвы. Решающая роль pH в альфа-разнообразии почвенных бактерий была продемонстрирована ранее (Rousk и др., 2010; Wang и др., 2017). Наши результаты показали, что влияние pH на бактериальное альфа-разнообразие может также распространяться на тепличные условия. Гетерогенность состава сообщества была сопоставима между системами земледелия. Другие исследования показали, что почвенный микробиом был более гетерогенным в органических системах, чем в традиционных (Tuck и др., 2014; Lupatini и др., 2017). По сравнению с полем или пастбищем, сельскохозяйственное управление в тепличных условиях является высокоинтенсивным, и эти методы управления могут снижать гетерогенность почвы.
Органическое тепличное земледелие вызывало устойчивые изменения в нескольких бактериальных таксонах на разных участках
Эффекты органического тепличного земледелия на большинство бактериальных родов (включая группы 2, 3 и 4 на РИСУНКЕ 2B) варьировали на разных участках, поскольку некоторые другие факторы, такие как физико-химические свойства почвы, типы и количество органических удобрений, также могут влиять на почвенные микробные сообщества (Chu и др., 2007; Yao и др., 2016; Ouyang и др., 2018a). Тем не менее, мы обнаружили роды, которые стабильно реагировали на органическое тепличное земледелие. Некоторые роды (группа 1 на РИСУНКЕ 3), предпочтительно связанные с органическим тепличным земледелием, часто были распространены в средах с обильным органическим материалом (Dennert и др., 2018; Zhang и др., 2019). Напротив, другие роды, такие как Nitrosospira и Bradyrhizobium, были отрицательно связаны с органическим тепличным земледелием. Представители Nitrosospira способны осуществлять окисление аммиака, и изменения предполагаемых аммоний-окисляющих бактерий согласуются с другими исследованиями органического земледелия (Wessen и др., 2011; Ding и др., 2019). Представители Bradyrhizobium способны образовывать симбиоз с бобовыми растениями для биологической фиксации азота (Liu и др., 2015). Кроме того, группы родов со сходными моделями реагирования на системы земледелия, которые вошли в 50 наиболее влиятельных родов на всех участках по данным анализа случайного леса, дают представление о реакции важных таксонов на органическое тепличное земледелие.
Органическое тепличное земледелие изменило сообщества азотфиксирующих бактерий, и его влияние на различные азотфиксирующие популяции в значительной степени зависело от участка
Свободноживущие или симбиотические азотфиксирующие бактерии могут фиксировать атмосферный N2 и добавлять биологически активный азот в агроэкосистемы (Orr и др., 2011). Альфа-разнообразие азотфиксирующих бактерий не различалось в зависимости от системы земледелия, но было связано с CEC. В недавнем исследовании Wang (Wang и др., 2019) наибольшее разнообразие азотфиксирующих бактерий было обнаружено в почве с самой высокой CEC. Состав сообщества азотфиксирующих бактерий имеет значение для скорости азотфиксации в сельскохозяйственных системах (Hsu и Buckley, 2009; Feng и др., 2018). Здесь азотфиксирующее сообщество различалось в зависимости от системы земледелия на 17 из 19 участков (значительное различие в составе сообщества между ORG и CON было подтверждено 1000-кратным двусторонним пермутационным анализом с p-значением 0.000 и несходством сообщества 21.8%). Хотя прямые сравнения азотфиксирующих сообществ между органическими и другими системами земледелия редки (Orr и др., 2011; Ding и др., 2019), было высказано предположение, что методы управления, связанные с органическим земледелием, такие как органическое удобрение, являются фактором, влияющим на популяции азотфиксирующих бактерий, включая альфа- и бета-разнообразие (Tang и др., 2017; Wang и др., 2017; Lin и др., 2018).
Кластеры nifH 1 и 3 доминировали в тепличных почвах независимо от системы земледелия, и их распространенность в наземных экосистемах была сообщена ранее (Zehr и др., 2017). Влияние органического земледелия на различные группы азотфиксаторов в значительной степени неизвестно (Grossman и др., 2005; Ramos и др., 2011). Здесь мы обнаружили, что кластер nifH 1J, часто встречающийся у Proteobacteria и Cyanobacteria (Krausfeldt и др., 2017; Zehr и др., 2017), был предпочтительно связан с органическим земледелием на участках, где pH почвы, K и плотность были значительно низкими. Азотфиксирующий P. borealis был отрицательно связан с органическим земледелием. Изменения P. borealis, по-видимому, не согласуются с результатами анализа гена 16S рРНК, в котором связь Paenibacillus с органическим земледелием была расходящейся. Возможно, что только часть Paenibacillus несет ген nifH (Grube и др., 2009). Влияние органического тепличного земледелия на некоторые другие азотфиксирующие бактерии, такие как Bradyrhizobium, Azoarcus, Paenibacillus, Bacteroidales, Desulfofustis и Desulfovibrio, с предполагаемым симбиотическим, свободноживущим и анаэробным образом жизни, было неодинаковым на разных участках (Wakelin и др., 2010). RDA дополнительно показал, что pH почвы может объяснить большую часть вариации, что согласуется с другими исследованиями (Reardon и др., 2014; Zarraonaindia и др., 2015; Wang и др., 2017). Органическое тепличное земледелие, возможно, оказывает давление отбора на азотфиксирующие бактерии, адаптированные к разным нишам, путем изменения ключевых физико-химических свойств почвы.
Эффекты органического тепличного земледелия на аммоний-окисляющие бактерии были в значительной степени фрагментарными
Ранее эффекты органического тепличного земледелия на альфа-разнообразие аммоний-окисляющих микроорганизмов были изучены лишь в немногих исследованиях (Wessen и др., 2011; Ding и др., 2019) и были в значительной степени неясны. Здесь мы обнаружили, что альфа-разнообразие аммоний-окисляющих бактерий не различалось в зависимости от системы земледелия, но было отрицательно связано с содержанием органического вещества. Аммоний-окисляющие бактерии в значительной степени являются литоавтотрофами, и внесение аммиака, вероятно, является наиболее важным драйвером их состава сообщества (Li и др., 2018). Зависящие от азотных удобрений изменения численности, разнообразия и активности этих бактерий часто обнаруживались в нескольких агроэкосистемах (Shen и др., 2012; Guo и др., 2017). Реакции AOB на добавление аммиака наблюдались как в микрокосмных, так и в полевых экспериментах (Daebeler и др., 2015; Ying и др., 2017), что подчеркивает, что добавление аммиака, которое обычно встречается в традиционной системе земледелия, может играть роль в изменении сообщества AOB. Однако по сравнению с общими и азотфиксирующими бактериальными сообществами, эффекты органического тепличного земледелия на аммоний-окисляющие бактерии были неожиданно непостоянными, поскольку состав сообщества различался в зависимости от системы земледелия только на шести из семнадцати участков. Эти результаты показали, что органическое тепличное земледелие не обязательно является основным драйвером аммоний-окисляющих бактерий. В полевом исследовании на фермерском уровне состав сообщества демонстрировал фрагментарную картину (Wessen и др., 2011). Некоторые другие факторы, такие как EC и плотность, которые были менее связаны с органическим земледелием, могут синергетически формировать сообщества аммоний-окисляющих бактерий.
pH почвы на исследуемом участке был связан с микробными узлами общих, азотфиксирующих и аммоний-окисляющих бактерий
Органическое земледелие часто приводит к увеличению разнообразия макроорганизмов (Wanjiku Kamau и др., 2019); однако влияние органического тепличного земледелия на микробное разнообразие варьирует в разных исследованиях. В целом, физико-химические свойства почвы, такие как pH, тип почвы, а не системы земледелия, были основными факторами, формирующими альфа-разнообразие общих, азотфиксирующих и аммоний-окисляющих бактерий в почве в большом масштабе, по данным анализа дисперсии (Вспомогательный рисунок S10). Анализ сетей совместной встречаемости показал, что доминирующие общие, азотфиксирующие и аммоний-окисляющие бактерии в основном образовывали отдельные узлы, что свидетельствует о том, что азотфиксирующие или аммоний-окисляющие бактерии не были тесно связаны с доминирующими общими бактериями. Предыдущие исследования показали, что численность как аммоний-окисляющих, так и азотфиксирующих бактерий часто была низкой в массе почвы, поскольку количество энергии, получаемой при окислении аммиака, очень низкое, а фиксация N2 требует высоких энергетических затрат (González-Cabaleiro и др., 2019; Luo и др., 2019). Связь между почвенным микробиомом и физико-химическими свойствами почвы была в значительной степени исследована на основе таксономии (Hansel и др., 2008; Rousk и др., 2010; Zhao и др., 2019), и было обнаружено несколько устойчивых корреляций. Возможно, существует расхождение между таксономией и физиологическими свойствами почвенного микробиома или что несколько таксонов сходным образом реагируют на определенные изменения. Здесь мы обнаружили, что относительная численность различных микробных узлов была в значительной степени связана с pH почвы, CEC и TC. Эти данные дополнительно показывают, что анализ сетей совместной встречаемости может также помочь выявить связь микробных популяций как узлов с ключевыми физико-химическими свойствами почвы или подчеркнуть, что органическое земледелие может изменять микробные взаимодействия путем изменения физико-химических свойств почвы.
Заключение
Резюмируя, альфа-разнообразие сообществ общих, азотфиксирующих и аммоний-окисляющих бактерий не различалось в зависимости от системы земледелия. Бета-разнообразие общих и азотфиксирующих, но не аммоний-окисляющих бактериальных сообществ, в значительной степени варьировало между системами земледелия на разных участках. Органическое тепличное земледелие стабильно отбирало бактерии, разлагающие органический углерод (Hyphomicrobium, Rubinisphaera, Aciditerrimonas, Planifilum, Phaselicystis и Ohtaekwangia), и азотфиксаторов кластера 1J, но не аммоний-окисляющие (Nitrosospira, Nitrosopumilus) и предполагаемые симбиотические азотфиксаторы (Bradyrhizobium), возможно, путем изменения ключевых физико-химических свойств почвы, таких как pH или CEC. Эти результаты подчеркивают, что органическое тепличное земледелие управляет перестройкой почвенного микробиома, связанного с круговоротом почвенного углерода, биологической фиксацией азота и окислением аммиака, что играет ключевую роль в поддержании плодородия почвы.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти онлайн по адресу: https://www.frontiersin.org/articles/10.3389/fmicb.2020.01861/full#supplementary-material
ДОПОЛНИТЕЛЬНЫЙ РИСУНОК S1
Относительная численность доминирующих бактерий между системами земледелия. ORG, органическая система земледелия; CON, традиционная система земледелия.
ДОПОЛНИТЕЛЬНЫЙ РИСУНОК S2
Бета-разнообразие бактериальных микробных сообществ в почвах при органической (ORG) и традиционной (CON) системах земледелия по данным UPGMA кластерного анализа.
ДОПОЛНИТЕЛЬНЫЙ РИСУНОК S3
Линейная взаимосвязь между средним несходством органической и традиционной систем земледелия и периодом органического земледелия.
ДОПОЛНИТЕЛЬНЫЙ РИСУНОК S4
Бета-разнообразие азотфиксирующих микробных сообществ в почвах при органической (ORG) и традиционной (CON) системах земледелия по данным UPGMA кластерного анализа.
ДОПОЛНИТЕЛЬНЫЙ РИСУНОК S5
Бета-разнообразие микробных сообществ аммоний-окисляющих бактерий (прямая область) в почвах при органической (ORG) и традиционной (CON) системах земледелия по данным UPGMA кластерного анализа.
ДОПОЛНИТЕЛЬНЫЙ РИСУНОК S6
Бета-разнообразие микробных сообществ аммоний-окисляющих бактерий (обратная область) в почвах при органической (ORG) и традиционной (CON) системах земледелия по данным UPGMA кластерного анализа.
ДОПОЛНИТЕЛЬНЫЙ РИСУНОК S7
Дискриминационные OTU аммоний-окисляющих бактерий (обратная область) между системами земледелия, которые вошли в 50 наиболее влиятельных OTU. Коричневые и голубые квадраты указывают на обогащение или истощение в системе органического тепличного земледелия на изученных участках. ORG, органическая система земледелия; CON, традиционная система земледелия.
ДОПОЛНИТЕЛЬНЫЙ РИСУНОК S8
Доминирующие (>1% относительной численности) общие, азотфиксирующие и аммоний-окисляющие бактериальные OTU и доля родов по данным анализа сетей совместной встречаемости.
ДОПОЛНИТЕЛЬНЫЙ РИСУНОК S9
Относительная численность микробных узлов для общих бактерий (A), азотфиксаторов (B) и аммоний-окисляющих бактерий (C) по данным анализа сетей совместной встречаемости.
ДОПОЛНИТЕЛЬНЫЙ РИСУНОК S10
Величина вариации в сообществах общих, азотфиксирующих и аммоний-окисляющих бактерий, объясняемая pH, типами почв и системами земледелия.
ДОПОЛНИТЕЛЬНАЯ ТАБЛИЦА S1
Детали сельскохозяйственного управления в условиях длительных тепличных экспериментов.
Сноски
Ссылки
1. Alam M. K., Bell R. W., Biswas W. K. (2019). Increases in soil sequestered carbon under conservation agriculture cropping decrease the estimated greenhouse gas emissions of wetland rice using life cycle assessment. J. Clean. Product. 224, 72–87. doi: 10.1016/j.jclepro.2019.03.215. CrossRef. Google Scholar.
2. Banerjee S., Walder F., Büchi L., Meyer M., Held A. Y., Gattinger A., et al. (2019). Agricultural intensification reduces microbial network complexity and the abundance of keystone taxa in roots. ISME J. 13, 1722–1736. doi: 10.1038/s41396-019-0383-2. CrossRef. Google Scholar.
3. Bao S. D. (2000). Analysis of Soil Agrochemicals. Beijing: China Agriculture Press. Google Scholar.
4. Bastian M., Heymann S., Jacomy M. (2009). "Gephi: an open source software for exploring and manipulating networks," in Proceedings of the International AAAI Conference on Weblogs and Social Media, San Jose, CA. Google Scholar.
5. Bengtsson J., Ahnström J., Weibull A. (2005). The effects of organic agriculture on biodiversity and abundance: a meta-analysis. J. Appl. Ecol. 42, 261–269. doi: 10.1111/j.1365-2664.2005.01005.x. CrossRef. Google Scholar.
6. Bonanomi G., De Filippis F., Cesarano G., La Storia A., Ercolini D., Scala F. (2016). Organic farming induces changes in soil microbiota that affect agro-ecosystem functions. Soil Biol. Biochem. 103, 327–336. doi: 10.1016/j.soilbio.2016.09.005. CrossRef. Google Scholar.
7. Caporaso J. G., Kuczynski J., Stombaugh J., Bittinger K., Bushman F. D., Costello E. K., et al. (2010). QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 7, 335–336. doi: 10.1038/nmeth.f.303. CrossRef. Google Scholar.
8. Che R., Qin J., Tahmasbian I., Wang F., Zhou S., Xu Z., et al. (2018). Litter amendment rather than phosphorus can dramatically change inorganic nitrogen pools in a degraded grassland soil by affecting nitrogen-cycling microbes. Soil Biol. Biochem. 120, 145–152. doi: 10.1016/j.soilbio.2018.02.006. CrossRef. Google Scholar.
9. Chen B., Ma X., Liu G., Xu X., Pan F., Zhang J., et al. (2015). An endophytic bacterium Acinetobacter calcoaceticus Sasm3-enhanced phytoremediation of nitrate-cadmium compound polluted soil by intercropping Sedum alfredii with oilseed rape. Environ. Sci. Pollut. Res. 22, 17625–17635. doi: 10.1007/s11356-015-4933-5. CrossRef. Google Scholar.
10. Chen H., Xia Q., Yang T., Shi W. (2018). Eighteen-year farming management moderately shapes the soil microbial community structure but promotes habitat-specific taxa. Front. Microbiol. 9:1776. doi: 10.3389/fmicb.2018.01776. CrossRef. Google Scholar.
11. Chu H., Fujii T., Morimoto S., Lin X., Yagi K., Hu J., et al. (2007). Community structure of ammonia-oxidizing bacteria under long-term application of mineral fertilizer and organic manure in a sandy loam soil. Appl. Environ. Microbiol. 73, 485–491. doi: 10.1128/AEM.01536-06. CrossRef. Google Scholar.
12. Cole J. R., Wang Q., Fish J. A., Chai B., McGarrell D. M., Sun Y., et al. (2013). Ribosomal database project: data and tools for high throughput rRNA analysis. Nucleic Acids Res. 42, 633–642. doi: 10.1093/nar/gkt1244. CrossRef. Google Scholar.
13. Daebeler A., Bodelier P. L. E., Hefting M. M., Laanbroek H. J. (2015). Ammonia-limited conditions cause of thaumarchaeal dominance in volcanic grassland soil. FEMS Microbiol. Ecol. 91:fiv014. Google Scholar.
14. Dennert F., Imperiali N., Staub C., Schneider J., Laessle T., Zhang T., et al. (2018). Conservation tillage and organic farming induce minor variations in Pseudomonas abundance, their antimicrobial function and soil disease resistance. FEMS Microbiol. Ecol. 94:fiy075. Google Scholar.
15. Di H. J., Cameron K. C. (2016). Inhibition of nitrification to mitigate nitrate leaching and nitrous oxide emissions in grazed grassland: a review. J. Soils Sediment. 16, 1401–1420. doi: 10.1007/s11368-016-1403-8. CrossRef. Google Scholar.
16. Ding G., Bai M., Han H., Li H., Ding X., Yang H., et al. (2019). Microbial taxonomic, nitrogen cycling and phosphorus recycling community composition during long-term organic greenhouse farming. FEMS Microbiol. Ecol. 95:fiz042. doi: 10.1093/femsec/fiz042. CrossRef. Google Scholar.
17. Ding G., Heuer H., Smalla K. (2012). Dynamics of bacterial communities in two unpolluted soils after spiking with phenanthrene: soil type specific and common responders. Front. Microbiol. 3:290. doi: 10.3389/fmicb.2012.00290. CrossRef. Google Scholar.
18. Feng M., Adams J. M., Fan K., Shi Y., Sun R., Wang D., et al. (2018). Long-term fertilization influences community assembly processes of soil diazotrophs. Soil Biol. Biochem. 126, 151–158. doi: 10.1016/j.soilbio.2018.08.021. CrossRef. Google Scholar.
19. Gomiero T., Pimentel D., Paoletti M. G. (2011). Environmental impact of different agricultural management practices: Conventional vs. Organic agriculture. Critical Rev. Plant Sci. 30, 95–124. doi: 10.1080/07352689.2011.554355. CrossRef. Google Scholar.
20. González-Cabaleiro R., Curtis T. P., Ofiteru I. D. (2019). Bioenergetics analysis of ammonia-oxidizing bacteria and the estimation of their maximum growth yield. Water Res. 154, 238–245. doi: 10.1016/j.watres.2019.01.054. CrossRef. Google Scholar.
21. Grossman J. M., Sheaffer C., Wyse D., Graham P. H. (2005). Characterization of slow-growing root nodule bacteria from Inga oerstediana in organic coffee agroecosystems in Chiapas, Mexico. Appl. Soil Ecol. 29, 236–251. doi: 10.1016/j.apsoil.2004.12.008. CrossRef. Google Scholar.
22. Grube M., Cardinale M., de Castro J. J. V., Müller H., Berg G. (2009). Species-specific structural and functional diversity of bacterial communities in lichen symbioses. ISME J. 3, 1105–1115. doi: 10.1038/ismej.2009.63. CrossRef. Google Scholar.
23. Guo J., Ling N., Chen H., Zhu C., Kong Y., Wang M., et al. (2017). Distinct drivers of activity, abundance, diversity and composition of ammonia-oxidizers: evidence from a long-term field experiment. Soil Biol. Biochem. 115, 403–414. doi: 10.1016/j.soilbio.2017.09.007. CrossRef. Google Scholar.
24. Hannachi N., Cocco S., Fornasier F., Agnelli A., Brecciaroli G., Massaccesi L., et al. (2015). Effects of cultivation on chemical and biochemical properties of dryland soils from southern Tunisia. Agric. Ecosyst. Environ. 199, 249–260. doi: 10.1016/j.agee.2014.09.009. CrossRef. Google Scholar.
25. Hansel C. M., Fendorf S., Jardine P. M., Francis C. A. (2008). Changes in bacterial and archaeal community structure and functional diversity along a geochemically variable soil profile. Appl. Environ. Microbiol. 74, 1620–1633. doi: 10.1128/AEM.01787-07. CrossRef. Google Scholar.
26. Hartmann M., Frey B., Mayer J., Mader P., Widmer F. (2015). Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 9, 1177–1194. doi: 10.1038/ismej.2014.210. CrossRef. Google Scholar.
27. Hill R., Saetnan E. R., Scullion J., Gwynn-Jones D., Ostle N., Edwards A. (2016). Temporal and spatial influences incur reconfiguration of Arctic heathland soil bacterial community structure. Environ. Microbiol. 18, 1942–1953. doi: 10.1111/1462-2920.13017. CrossRef. Google Scholar.
28. Hink L., Gubry-Rangin C., Nicol G. W., Prosser J. I. (2018). The consequences of niche and physiological differentiation of archaeal and bacterial ammonia oxidisers for nitrous oxide emissions. ISME J. 12, 1084–1093. doi: 10.1038/s41396-017-0025-5. CrossRef. Google Scholar.
29. Hothorn T., Bretz F., Westfall P. (2008). Simultaneous inference in general parametric models. Biom. J. 50, 346–363. doi: 10.1002/bimj.200810425. CrossRef. Google Scholar.
30. Hsu S., Buckley D. H. (2009). Evidence for the functional significance of diazotroph community structure in soil. ISME J. 3, 124–136. doi: 10.1038/ismej.2008.82. CrossRef. Google Scholar.
31. Janvier C., Villeneuve F., Alabouvette C., Edel-Hermann V., Mateille T., Steinberg C. (2007). Soil health through soil disease suppression: which strategy from descriptors to indicators? Soil Biol. Biochem. 39, 1–23. doi: 10.1016/j.soilbio.2006.07.001. CrossRef. Google Scholar.
32. Köberl M., Erlacher A., Ramadan E. M., El-Arabi T. F., Müller H., Bragina A., et al. (2016). Comparisons of diazotrophic communities in native and agricultural desert ecosystems reveal plants as important drivers in diversity. FEMS Microbiol. Ecol. 92:v166. doi: 10.1093/femsec/fiv166. CrossRef. Google Scholar.
33. Krausfeldt L. E., Tang X., van de Kamp J., Gao G., Bodrossy L. (2017). Spatial and temporal variability in the nitrogen cyclers of hypereutrophic Lake Taihu. FEMS Microbiol. Ecol. 93:24. doi: 10.1093/femsec/fix024. CrossRef. Google Scholar.
34. Kropf S., Läuter J., Eszlinger M., Krohn K., Paschke R. (2004). Nonparametric multiple test procedures with data-driven order of hypotheses and with weighted hypotheses. J. Statist. Plan. Inferen. 125, 31–47. doi: 10.1016/j.jspi.2003.07.021. CrossRef. Google Scholar.
35. Kuypers M. M. M., Marchant H. K., Kartal B. (2018). The microbial nitrogen-cycling network. Nat. Rev. Microbiol. 16, 263–276. doi: 10.1038/nrmicro.2018.9. CrossRef. Google Scholar.
36. Levy-Booth D. J., Prescott C. E., Grayston S. J. (2014). Microbial functional genes involved in nitrogen fixation, nitrification and denitrification in forest ecosystems. Soil Biol. Biochem. 75, 11–25. doi: 10.1016/j.soilbio.2014.03.021. CrossRef. Google Scholar.
37. Li H., Cai X., Gong J., Xu T., Ding G., Li J. (2019). Long-Term organic farming manipulated rhizospheric microbiome and bacillus antagonism against pepper blight (Phytophthora capsici). Front. Microbiol. 10:342. doi: 10.3389/fmicb.2019.00342. CrossRef. Google Scholar.
38. Li H., Mu B., Jiang Y., Gu J. (2011). Production processes affected prokaryotic amoA gene abundance and distribution in high-temperature petroleum reservoirs. Geomicrobiol. J. 28, 692–704. doi: 10.1080/01490451.2010.514026. CrossRef. Google Scholar.
39. Li Y., Chapman S. J., Nicol G. W., Yao H. (2018). Nitrification and nitrifiers in acidic soils. Soil Biol. Biochem. 116, 290–301. doi: 10.1016/j.soilbio.2017.10.023. CrossRef. Google Scholar.
40. Liaw A., Wiener M. (2002). Classification and regression by randomForest. R News 2:23. Google Scholar.
41. Lin Y., Ye G., Liu D., Ledgard S., Luo J., Fan J., et al. (2018). Long-term application of lime or pig manure rather than plant residues suppressed diazotroph abundance and diversity and altered community structure in an acidic Ultisol. Soil Biol. Biochem. 123, 218–228. doi: 10.1016/j.soilbio.2018.05.018. CrossRef. Google Scholar.
42. Liu L., He X., Wang K., Xie Y., Xie Q., O'Donnell A. G., et al. (2015). The bradyrhizobium-legume symbiosis is dominant in the shrubby ecosystem of the Karst region, Southwest China. Eur. J. Soil Biol. 68, 1–8. doi: 10.1016/j.ejsobi.2015.02.004. CrossRef. Google Scholar.
43. Lori M., Symnaczik S., Mäder P., De Deyn G., Gattinger A. (2017). Organic farming enhances soil microbial abundance and activity- a meta-analysis and meta-regression. PLoS One 12:e180442. doi: 10.1371/journal.pone.0180442. CrossRef. Google Scholar.
44. Luo Y. W., Shi D., Kranz S. A., Hopkinson B. M., Hong H., Shen R., et al. (2019). Reduced nitrogenase efficiency dominates response of the globally important nitrogen fixer Trichodesmium to ocean acidification. Nat. Commun. 10:1521. doi: 10.1038/s41467-019-09554-7. CrossRef. Google Scholar.
45. Lupatini M., Korthals G. W., de Hollander M., Janssens T. K. S., Kuramae E. E. (2017). Soil microbiome is more heterogeneous in organic than in conventional farming system. Front. Microbiol. 7:2064. doi: 10.3389/fmicb.2016.02064. CrossRef. Google Scholar.
46. Moeskops B., Sukristiyonubowo O., Buchan D., Sleutel S., Herawaty L., Husenb E., et al. (2010). Soil microbial communities and activities under intensive organic and conventional vegetable farming in West Java, Indonesia. Appl. Soil Ecol. 2, 112–120. doi: 10.1016/j.apsoil.2010.03.005. CrossRef. Google Scholar.
47. National Bureau of Statistics of the People's Republic of China [NBSC] (2014). China Rural Statistical Yearbook. Beijing: China Statistics Press. Google Scholar.
48. Orr C. H., James A., Leifert C., Cooper J. M., Cummings S. P. (2011). Diversity and activity of free-living nitrogen-fixing bacteria and total bacteria in organic and conventionally managed soils. Appl. Environ. Microbiol. 77, 911–919. doi: 10.1128/AEM.01250-10. CrossRef. Google Scholar.
49. Ouyang Y., Evans S. E., Friesen M. L., Tiemann L. K. (2018a). Effect of nitrogen fertilization on the abundance of nitrogen cycling genes in agricultural soils: a meta-analysis of field studies. Soil Biol. Biochem. 127, 71–78. doi: 10.1016/j.soilbio.2018.08.024. CrossRef. Google Scholar.
50. Ouyang Y., Reeve J. R., Norton J. M. (2018b). Soil enzyme activities and abundance of microbial functional genes involved in nitrogen transformations in an organic farming system. Biol. Ferti. Soils 54, 437–450. doi: 10.1007/s00374-018-1272-y. CrossRef. Google Scholar.
51. Poly F., Monrozier L. J., Bally R. (2001). Improvement in the RFLP procedure for studying the diversity of nifH genes in communities of nitrogen fixers in soil. Res. Microbiol. 152, 95–103. doi: 10.1016/s0923-2508(00)01172-4. CrossRef. Google Scholar.
52. Postma J., Schilder M. T., Bloem J., van Leeuwen-Haagsma W. K. (2008). Soil suppressiveness and functional diversity of the soil microflora in organic farming systems. Soil Biol. Biochem. 40, 2394–2406. doi: 10.1016/j.soilbio.2008.05.023. CrossRef. Google Scholar.
53. Prober S. M., Leff J. W., Bates S. T., Borer E. T., Firn J., Harpole W. S., et al. (2015). Plant diversity predicts beta but not alpha diversity of soil microbes across grasslands worldwide. Ecol. Lett. 18, 85–95. doi: 10.1111/ele.12381. CrossRef. Google Scholar.
54. Qiao Y., Martin F., Cook S., He X., Halberg N., Scott S., et al. (2018). Certified organic agriculture as an alternative livelihood strategy for small-scale farmers in China: a case study in Wanzai county, Jiangxi province. Ecol. Econ. 145, 301–307. doi: 10.1016/j.ecolecon.2017.10.025. CrossRef. Google Scholar.
55. Ramos P. L., Van Trappen S., Thompson F. L., Rocha R. C. S., Barbosa H. R., De Vos P., et al. (2011). Screening for endophytic nitrogen-fixing bacteria in Brazilian sugar cane varieties used in organic farming and description of Stenotrophomonas pavanii sp. Nov. Intern. J. Syst. Evol. Microbiol. 61, 926–931. doi: 10.1099/ijs.0.019372-0. CrossRef. Google Scholar.
56. Reardon C. L., Gollany H. T., Wuest S. B. (2014). Diazotroph community structure and abundance in wheat-fallow and wheat-pea crop rotations. Soil Biol. Biochem. 69, 406–412. doi: 10.1016/j.soilbio.2013.10.038. CrossRef. Google Scholar.
57. Rotthauwe J. H., Witzel K. P., Liesack W. (1997). The ammonia monooxygenase structural gene amoA as a functional marker: molecular fine-scale analysis of natural ammonia-oxidizing populations. Appl. Environ. Microbiol. 63, 4704–4712. doi: 10.1128/AEM.63.12.4704-4712.1997. CrossRef. Google Scholar.
58. Rousk J., Bååth E., Brookes P. C., Lauber C. L., Lozupone C. (2010). Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 4, 1340–1351. doi: 10.1038/ismej.2010.58. CrossRef. Google Scholar.
59. Schloss P. D., Westcott S. L., Ryabin T., Hall J. R., Hartmann M., Hollister E. B., et al. (2009). introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 75, 7537–7541. doi: 10.1128/AEM.01541-09. CrossRef. Google Scholar.
60. Scotti R., Bonanomi G., Scelza R., Zoina A., Rao M. A. (2015). Organic amendments as sustainable tool to recovery fertility in intensive agricultural systems. J. Soil Sci. Plant Nutr. 15, 333–352. Google Scholar.
61. Searchinger T. D., Wirsenius S., Beringer T., Dumas P. (2018). Assessing the efficiency of changes in land use for mitigating climate change. Nature 564, 249–253. doi: 10.1038/s41586-018-0757-z. CrossRef. Google Scholar.
62. Seufert V., Ramankutty N., Foley J. A. (2012). Comparing the yields of organic and conventional agriculture. Nature 485, 229–232. doi: 10.1038/nature11069. CrossRef. Google Scholar.
63. Shen J., Zhang L., Di H. J., He J. (2012). A review of ammonia-oxidizing bacteria and archaea in Chinese soils. Front. Microbiol. 3:296. doi: 10.3389/fmicb.2012.00296. CrossRef. Google Scholar.
64. Smith L. G., Kirk G. J. D., Jones P. J., Williams A. G. (2019). The greenhouse gas impacts of converting food production in England and Wales to organic methods. Nat. Commun. 10:622. doi: 10.1038/s41467-019-12622-7. CrossRef. Google Scholar.
65. Tamaki H., Wright C. L., Li X., Lin Q., Hwang C., Wang S., et al. (2011). Analysis of 16S rRNA amplicon sequencing options on the Roche/454 next-generation titanium sequencing platform. PLoS One 6:e25263. doi: 10.1371/journal.pone.0025263. CrossRef. Google Scholar.
66. Tang Y., Zhang M., Chen A., Zhang W., Wei W., Sheng R. (2017). Impact of fertilization regimes on diazotroph community compositions and N2-fixation activity in paddy soil. Agric. Ecosyst. Environ. 247, 1–8. doi: 10.1016/j.agee.2017.06.009. CrossRef. Google Scholar.
67. Thorpe A. S., Callaway R. M. (2011). Biogeographic differences in the effects of Centaurea stoebe on the soil nitrogen cycle: novel weapons and soil microbes. Biol. Invas. 13, 1435–1445. doi: 10.1007/s10530-010-9902-9. CrossRef. Google Scholar.
68. Tuck S. L., Winqvist C., Mota F., Ahnström J., Turnbull L. A., Bengtsson J. (2014). Land-use intensity and the effects of organic farming on biodiversity: a hierarchical meta-analysis. J. Appl. Ecol. 51, 746–755. doi: 10.1111/1365-2664.12219. CrossRef. Google Scholar.
69. Uksa M., Fischer D., Welzl G., Kautz T., Köpke U., Schloter M. (2014). Community structure of prokaryotes and their functional potential in subsoils is more affected by spatial heterogeneity than by temporal variations. Soil Biol. Biochem. 75, 197–201. doi: 10.1016/j.soilbio.2014.04.018. CrossRef. Google Scholar.
70. Van Bruggen A. H., Gamliel A., Finckh M. R. (2016). Plant disease management in organic farming systems. Pest. Manag. Sci. 72, 30–44. doi: 10.1002/ps.4145. CrossRef. Google Scholar.
71. van Bruggen A. H. C., Sharma K., Kaku E., Karfopoulos S., Zelenev V. V., Blok W. J. (2015). Soil health indicators and Fusarium wilt suppression in organically and conventionally managed greenhouse soils. Appl. Soil Ecol. 86, 192–201. doi: 10.1016/j.apsoil.2014.10.014. CrossRef. Google Scholar.
72. Wakelin S. A., Gupta V. V. S. R., Forrester S. T. (2010). Regional and local factors affecting diversity, abundance and activity of free-living, N2-fixing bacteria in Australian agricultural soils. Pedobiologia 53, 391–399. doi: 10.1016/j.pedobi.2010.08.001. CrossRef. Google Scholar.
73. Wang C., Zheng M., Song W., Wen S., Wang B., Zhu C., et al. (2017). Impact of 25 years of inorganic fertilization on diazotrophic abundance and community structure in an acidic soil in southern China. Soil Biol. Biochem. 113, 240–249. doi: 10.1016/j.soilbio.2017.06.019. CrossRef. Google Scholar.
74. Wang X., Liu B., Ma J., Zhang Y., Hu T., Zhang H., et al. (2019). Soil aluminum oxides determine biological nitrogen fixation and diazotrophic communities across major types of paddy soils in China. Soil Biol. Biochem. 131, 81–89. doi: 10.1016/j.soilbio.2018.12.028. CrossRef. Google Scholar.
75. Wanjiku Kamau J., Biber-Freudenberger L., Lamers J. P. A., Stellmacher T., Borgemeister C. (2019). Soil fertility and biodiversity on organic and conventional smallholder farms in Kenya. Appl. Soil Ecol. 134, 85–97. doi: 10.1016/j.apsoil.2018.10.020. CrossRef. Google Scholar.
76. Wessen E., Soderstrom M., Stenberg M., Bru D., Hellman M., Welsh A., et al. (2011). Spatial distribution of ammonia-oxidizing bacteria and archaea across a 44-hectare farm related to ecosystem functioning. ISME J. 5, 1213–1225. doi: 10.1038/ismej.2010.206. CrossRef. Google Scholar.
77. Wu M., Han H., Zheng X., Bai M., Xu T., Ding G., et al. (2019). Dynamics of oxytetracycline and resistance genes in soil under long-term intensive compost fertilization in Northern China. Environ. Sci. Pollut. Res. 26, 21381–21393. doi: 10.1007/s11356-019-05173-3. CrossRef. Google Scholar.
78. Yang S., Wang Y., Liu R., Xing L., Yang Z. (2018). Improved crop yield and reduced nitrate nitrogen leaching with straw return in a rice-wheat rotation of Ningxia irrigation district. Sci. Rep. 8:9458. doi: 10.1038/s41598-018-27776-5. CrossRef. Google Scholar.
79. Yao H., Huang S., Qiu Q., Li Y., Wu L., Mi W., et al. (2016). Effects of different fertilizers on the abundance and community structure of ammonia oxidizers in a yellow clay soil. Appl. Microbiol. Biotechnol. 100, 6815–6826. doi: 10.1007/s00253-016-7502-z. CrossRef. Google Scholar.
80. Ying J., Li X., Wang N., Lan Z., He J., Bai Y. (2017). Contrasting effects of nitrogen forms and soil pH on ammonia oxidizing microorganisms and their responses to long-term nitrogen fertilization in a typical steppe ecosystem. Soil Biol. Biochem. 107, 10–18. doi: 10.1016/j.soilbio.2016.12.023. CrossRef. Google Scholar.
81. Zarraonaindia I., Owens S. M., Weisenhorn P., West K., Hampton-Marcell J., Lax S., et al. (2015). The soil microbiome influences grapevine-associated microbiota. mBio 6:e002527-14. doi: 10.1128/mBio.02527-14. CrossRef. Google Scholar.
82. Zehr J. P., Shilova I. N., Farnelid H. M., Muñoz-Marín M. D. C., Turk-Kubo K. A. (2017). Unusual marine unicellular symbiosis with the nitrogen-fixing cyanobacterium UCYN-A. Nat. Microbiol. 2:214. doi: 10.1038/nmicrobiol.2016.214. CrossRef. Google Scholar.
83. Zhang W., Yu C., Wang X., Hai L. (2019). Increased abundance of nitrogen transforming bacteria by higher C/N ratio reduces the total losses of N and C in chicken manure and corn stover mix composting. Bioresour. Technol. 297:122410. doi: 10.1016/j.biortech.2019.122410. CrossRef. Google Scholar.
84. Zhao Z., He J., Geisen S., Han L., Wang J., Shen J., et al. (2019). Protist communities are more sensitive to nitrogen fertilization than other microorganisms in diverse agricultural soils. Microbiome 7:33. doi: 10.1186/s40168-019-0647-0. CrossRef. Google Scholar.
85. Zhong Jing Hui Cheng Institute of Urban-Rural Planning and Design [ZIURPD] (2014). Greenhouse Agricultural Industry Analysis. Industrial Planning Net of China. Available online at: http://www.chanyeguihua.com/1262.html (accessed August, 2014). Google Scholar.

Chen C, Han H, Xu T, Lv Y, Hu K, Li XX, Qiao Y, Ding G-C and Li J (2020) Comparison of the Total, Diazotrophic and Ammonia-Oxidizing Bacterial Communities Between Under Organic and Conventional Greenhouse Farming. Front. Microbiol. 11:1861. doi: 10.3389/fmicb.2020.01861
Перевод статьи «Comparison of the Total, Diazotrophic and Ammonia-Oxidizing Bacterial Communities Between Under Organic and Conventional Greenhouse Farming» авторов Chen C, Han H, Xu T, Lv Y, Hu K, Li XX, Qiao Y, Ding G-C and Li J., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык
Фото: Designed by Freepik




















































































































Комментарии (0)