Органическое земледелие усиливает защитные механизмы растений в полевых условиях
Растения используют разнообразные химические и физические средства защиты от травоядных и патогенов. Органическое земледелие стремится усилить эти реакции за счёт улучшения качества почвы, что в конечном итоге изменяет низовую регуляцию («bottom-up» regulation) защитных механизмов растений. Хотя лабораторные исследования подтверждают эффективность такого подхода, остаётся неясным, стимулирует ли органическое сельское хозяйство более активную защиту растений в реальных условиях.
Аннотация
Работая на полях сотрудничающих с нами фермеров, мы изучили экспрессию генов в листьях двух сортов картофеля (Solanum tuberosum), выращенных в органических и традиционных (конвенциональных) хозяйствах. Для одного сорта — Норкота — мы обнаружили значительное повышение активности генов, связанных с защитными путями растений, в образцах с органических полей по сравнению с конвенциональными. На органических полях с Норкотой наблюдались более низкие уровни нитратов в почве и азота в листве растений, а также различия в сообществах почвенных бактерий, что указывает на возможные связи между методами обработки почвы и наблюдаемыми различиями в защитных реакциях растений. Кроме того, численность хищных насекомых и насекомых, питающихся флоэмой, была выше на органических полях, чем на конвенциональных.
Второй сорт картофеля — Алтурас, который обычно выращивается с меньшим количеством агрохимикатов и на менее плодородных почвах, показал в целом более низкую численность травоядных и хищников, незначительные различия в почвенной экологии и отсутствие различий в активности генов между органической и традиционной системами земледелия.
В совокупности наши результаты позволяют предположить, что органическое земледелие способно повышать устойчивость растений к травоядным, что, возможно, снижает потребность в применении инсектицидов. Однако эти преимущества, по-видимому, зависят от сорта растения и/или конкретных условий хозяйства.
Введение
Традиционные методы ведения сельского хозяйства часто в значительной степени полагаются на химические инсектициды для борьбы с насекомыми-фитофагами (например, Soffe, 2002; Stockdale et al., 2002; Klonsky, 2012). Такой подход может непреднамеренно нарушить биологический контроль и усугубить вспышки численности вредителей, когда химические вещества широкого спектра действия уничтожают естественных врагов (Penman and Chapman, 1988; Prischmann et al., 2005; Woods et al., 2012). Однако, возможно, менее известны способы, которыми традиционные методы ведения сельского хозяйства могут нарушать «низовое» подавление травоядных растений. Растения используют широкий спектр химических и физических средств защиты от травоядных (Feeny, 1976; Price, 1991; Howe and Jander, 2008; Mithöfer and Boland, 2012), и на индукцию этих защитных механизмов могут влиять методы ведения сельского хозяйства (например, Davis et al., 2001; Bonanomi et al., 2010). Например, синтетические химические удобрения обеспечивают внезапные импульсы питательных веществ, которые могут быть физиологически стрессовыми для растений, включая нарушение путей, связанных с защитой растений от травоядных (Altieri and Nicholls, 2003; Bhardwaj et al., 2014). Аналогичным образом, известно, что некоторые инсектициды изменяют физиологию растений к непреднамеренной выгоде для насекомых, питающихся растениями (Karthikeyan et al., 2009; Ford et al., 2010; Szczepaniec et al., 2013). Например, инсектициды-неоникотиноиды могут подавлять экспрессию важных защитных генов растений, изменять уровни фитогормонов, участвующих в защите растений, и снижать устойчивость растений к таким травоядным, как паутинные клещи (например, James and Price, 2002; Szczepaniec et al., 2013). Роль травоядных и их значение в вызывании крупномасштабных изменений экспрессии генов широко изучены (Cheong et al., 2002; Delessert et al., 2004; Reymond et al., 2004; Smith et al., 2004; Voelckel and Baldwin, 2004; Zhu-Salzman et al., 2004; De Vos et al., 2005; Schmidt et al., 2005; Ralph et al., 2006; Thompson and Goggin, 2006; Broekgaarden et al., 2007). Следовательно, традиционное современное сельское хозяйство может влиять как на подавление травоядных «сверху вниз», нанося вред естественным врагам, так и на подавление травоядных «снизу вверх», нарушая защитные механизмы растений.
В качестве альтернативного подхода органическое земледелие стремится заменить синтетические химические инсектициды естественными методами борьбы с вредителями (Rigby and Cáceres, 2001; Pimentel et al., 2005). Действительно, сокращение количества опрыскиваний инсектицидами в этих системах, вероятно, способствует более обильным и биоразнообразным сообществам хищных насекомых (Crowder et al., 2010), пауков (Letourneau and Goldstein, 2001) и певчих птиц (Beecher et al., 2002; Bengtsson et al., 2005; Hole et al., 2005; Bouvier et al., 2011) на органических фермах по сравнению с традиционными, что улучшает биологический контроль насекомых-вредителей (Bengtsson et al., 2005; Birkhofer et al., 2008; Crowder et al., 2010). Также возможно, что органическое земледелие может улучшить низовую регуляцию вредителей (Altieri and Nicholls, 2003). Например, за счет отказа от синтетических химических инсектицидов органические методы должны позволить избежать любых негативных последствий, которые эти химические вещества оказывают на защитные механизмы растений (обсуждалось выше). Кроме того, органическое земледелие заменяет синтетические химические удобрения компостированным навозом животных, сидератами и другими биологическими источниками плодородия (Stockdale et al., 2002; Watson et al., 2002; Pimentel et al., 2005). Эти добавки, как правило, высвобождают азот и другие питательные вещества относительно медленно по сравнению с синтетическими удобрениями (Eigenbrode and Pimentel, 1988; Phelan et al., 1995, 1996; Hsu et al., 2009), и стабильное высвобождение питательных веществ, вероятно, оказывает широкомасштабное влияние на физиологию растений, включая защиту (McGuiness, 1993; Phelan et al., 1995). Эти подходы к управлением плодородием также увеличивают содержание органического вещества в почве, что является явной целью органического земледелия (например, Jenny, 1980; Reganold et al., 1987; McGuiness, 1993; Altieri and Nicholls, 2003), что может увеличить биомассу и биоразнообразие почвенных организмов (например, Gunapala and Scow, 1998; Swezey et al., 1998; Altieri, 1999; Mäder et al., 2002). Известно, что почвенные микробы индуцируют или праймируют защитные механизмы растений, которые обеспечивают косвенную защиту от насекомых-фитофагов (Davis et al., 2001; Bonanomi et al., 2010). Хотя исследования в контролируемых условиях действительно показали усиление защитных механизмов у растений, выращенных с использованием органических почвенных добавок по сравнению с традиционными (например, Bloemberg and Lugtenberg, 2001; Lugtenberg et al., 2002; Bais et al., 2004; Morrissey et al., 2004; Goh et al., 2013), остается неясным, будет ли это также наблюдаться в более сложных условиях реальных ферм, различающихся по широкому разнообразию методов ведения сельского хозяйства.
Чтобы изучить многочисленные физиологические механизмы, с помощью которых органическое земледелие может улучшить устойчивость к вредителям, мы использовали секвенирование РНК (RNA-seq) и сравнили экспрессию защитных генов в листве коммерчески выращиваемого картофеля (Solanum tuberosum), собранного с пяти органических и семи традиционных полей. Эти фермы были расположены в регионе выращивания картофеля в центральной и южной части штата Вашингтон (Supplementary Table 1), и мы рассмотрели два наиболее распространенных сорта картофеля, выращиваемых в этом регионе, Norkotah и Alturas. Наши образцы были отобраны в середине вегетационного периода, когда растения на всех полях цвели и когда повреждения от вредителей, а следовательно, и риски для урожайности, наиболее велики (O. Johnson, коммерческий производитель картофеля, личное сообщение). Чтобы оценить экологические факторы, помимо системы земледелия, которые могли способствовать экспрессии защитных генов, мы также провели интенсивный отбор проб сообществ членистоногих и собрали почву для документирования химических и биологических свойств почвы (включая оценку микробного разнообразия почвы с помощью секвенирования 16S рРНК) одновременно со сбором листвы картофеля. Наша цель состояла не только в том, чтобы обнаружить стабильные различия в экспрессии защитных генов между двумя системами земледелия, но и в том, чтобы инициировать поиск конкретных факторов, различающихся между двумя системами земледелия, которые могли бы в конечном итоге лежать в основе коррелятивных различий в характере экспрессии генов.
Материалы и методы
Сбор и подготовка образцов
С полей сортов картофеля Norkotah и Alturas сотрудничающих с нами производителей по всему центральному Вашингтону (5 органических и 7 традиционных, Supplementary Table 1) мы собрали поврежденную и неповрежденную листву картофеля для анализа RNA-seq; провели интенсивный отбор проб сообществ хищных членистоногих и фитофагов; а также отобрали почву для химической и микробиологической характеристики. Ниже мы предоставляем подробные методы для каждого из этих компонентов.
В штате Вашингтон на традиционных картофельных полях для борьбы с насекомыми-вредителями обычно применяют инсектициды широкого спектра действия — неоникотиноиды и пиретроиды, в то время как с возбудителями болезней растений борются с помощью предпосадочного фумигирования почвы с последующей обработкой семян фунгицидами, профилактических средств против фитофтороза, таких как манкоцеб, и различных продуктов, направленных против патогенов, которые появляются на культуре в течение вегетационного периода (Murray et al., 2019). Напротив, с насекомыми-вредителями на органических картофельных полях обычно борются с помощью экстракта нима и Bacillus thuringiensis, в то время как с возбудителями болезней растений борются с помощью препаратов меди (Murray et al., 2019). Дополнительные различия включают использование гербицидов для подавления сорняков и синтетических химических удобрений для повышения плодородия на традиционных полях по сравнению с частой обработкой почвы и внесением навоза и мульчи в органической системе (Koss et al., 2005; Murray et al., 2019).
Все растения картофеля, обоих сортов и в каждой из двух систем земледелия, были отобраны в фазе цветения, которое происходит в течение нескольких недель в середине вегетационного периода. Это время отбора совпадает с периодом особенно быстрого роста листвы и клубнеобразования, когда повреждения от вредителей могут быть особенно вредны для последующей урожайности (Stark et al., 2020). На площади 400 м² каждой фермы мы случайным образом выбрали 10 листьев, поврежденных насекомыми (с видимыми повреждениями от грызущих или сосущих насекомых), и 10 неповрежденных листьев с 20 отдельных растений, расположенных на среднем уровне растения (верхние листья моложе и склонны к избыточной защите; Zangerl and Bazzaz, 1992; Hartley and Jones, 1997). В нитриловых перчатках мы использовали стерильное лезвие бритвы, чтобы быстро срезать каждый лист у стебля, помещая каждый лист в 50-мл коническую пробирку Falcon и немедленно помещая ее в контейнер с жидким азотом. Хотя все растения имели некоторые повреждения, выбранные неповрежденные листья были визуально свободны от повреждений. Затем пробирки хранились в кулерах с сухим льдом во время транспортировки обратно в лабораторию, где они немедленно помещались на хранение при температуре −80°C.
Экстракция РНК из листвы растений
Тотальная РНК из каждого листа, собранного, как описано выше, была экстрагирована с использованием набора RNeasy Plant Mini Kit (Qiagen Inc., Германия). Затем РНК была объединена с равными концентрациями РНК в общей сложности 5 уникальных образцов на комбинацию обработок (тип управления и тип повреждения) в пределах сорта (см. Supplementary Figure 1). Переваривание ДНКазой I, свободной от РНКазы, было проведено после экстракции с использованием набора TURBO DNA-free Kit (Invitrogen Thermo Scientific, Литва). Начальная целостность и качество общей РНК были проверены с помощью спектрофотометра NanoDrop 1000 и формальдегид-агарозного гель-электрофореза. Окончательные образцы РНК были оценены с использованием Fragment Analyzer (Advanced Analytical Technologies, Ankeny, IA) с набором High Sensitivity RNA Analysis Kit, и для приготовления библиотек РНК использовались только образцы с RQN от 6 до 10 в соответствии со стандартными протоколами (TruSeq v2 Stranded mRNA Library Prep Kit; Illumina, San Diego, CA). Концентрации библиотек РНК были измерены с помощью системы StepOnePlus Real-Time PCR System (ThermoFisher Scientific, San Jose, CA) с набором KAPA Library Quantification Kit (Kapabiosystems, Wilmington, MA). Библиотеки были разбавлены до 2 нМ с помощью RSB (10 мМ Tris-HCl, pH 8,5) и денатурированы 0,1 М NaOH. Библиотеки концентрацией 18 пМ были кластеризованы в проточной ячейке высокой производительности с использованием набора HiSeq Cluster Kit v4 на приборе cBot (Illumina) и секвенированы с использованием парных концов (100 п.н.) на приборе HiSeq 2500 с использованием набора HiSeq SBS kit v4 (Illumina). Сырые bcl-файлы были преобразованы в fastq-файлы с помощью программного обеспечения bcl2fastq2.17.1.14.
РНК-seq риды были очищены от адаптеров, а затем выровнены по геному S. tuberosum с помощью программы выравнивания Tophat и Bowtie (Langmead et al., 2009; Trapnell et al., 2009; Bolger et al., 2014; Version12x; Phytozome Version 9, Joint Genome Institute) с параметрами по умолчанию, которые допускают до двух несоответствий и сообщают о до 40 выравниваний для ридов, картированных на множественных позициях. Подсчеты количества генов были сделаны с использованием Python Package HTSeq (Anders et al., 2015). Сырые данные последовательностей были депонированы в Sequence Read Archive (http://www.ncbi.nlm.nih.gov/sra) под номерами BioProject PRJNA417905 и PRJNA417851.
Отбор проб членистоногих
Нашей следующей задачей было оценить плотность хищников и фитофагов на каждом из картофельных полей, где были собраны образцы листвы. Все хищники были собраны в той же области, где собирали листовую ткань, с использованием устройства для отбора проб D-vac и ранее описанных методов (например, Koss et al., 2005). Кратко, мы произвольно идентифицировали 10 растений картофеля на поле для отбора проб, двигаясь зигзагообразно от края поля к центру. Мы держали собирательный конус над каждым растением, осторожно встряхивая листву в течение 20 секунд, и меняли собирательные мешки между каждой группой из 10 растений (Koss et al., 2005). Мы также визуально осмотрели дополнительный набор из 10 растений, выбранных произвольно, подсчитывая любых присутствующих членистоногих в течение 30 секунд. Членистоногие в мешках D-vac были немедленно помещены в кулер с сухим льдом и транспортированы обратно в лабораторию для дальнейшей идентификации до уровня семейства с использованием дихотомических ключей в (Triplehorn et al., 2005).
Анализы почвы и экстракция ДНК
На каждом фермерском участке пять образцов почвы были отобраны случайным образом из каждого угла и из центра в пределах той же области 400 м² на поле, как указано выше. Каждый из пяти образцов состоял из почвы глубиной 20 см, отобранной с помощью почвенного бура. Повторности почвы в пределах каждого поля были перемешаны и упакованы в стерильные пакеты whirlpak, каждый пакет был запечатан и транспортирован в лабораторию при 4°C. 100-г фракция смешанного образца была сохранена при −20°C для молекулярного анализа, в то время как ~200 г почвы были немедленно отправлены в лабораторию SoilTest Labs (Moses Lake, WA). Там образцы почвы были просеяны через сито с ячейками 2 мм и проанализированы на следующие свойства в соответствии с рекомендуемыми методами анализа почвы Gavlak et al. (2003): гранулометрический состав (% песка, ила и глины); нитратный азот и аммонийный азот; экстрагируемый фосфор, калий, кальций, магний, натрий, сера, бор, цинк, марганец, медь и железо; катионообменная способность; сумма оснований; % насыщенности основаниями; обменный процент натрия (ESP); pH; электропроводность; минерализация азота; и процент органического вещества (см. Supplementary Table 2). Кроме того, микробная биомасса была оценена с использованием теста Solvita (Haney et al., 2008) (Supplementary Table 2).
ДНК была экстрагирована из 0,25 г образца с использованием набора PowerSoil DNA Isolation Kit (MOBIO Laboratories, Inc., Carlsbad, CA, USA). Экстракции ДНК из почвы были отправлены в Центр геномных исследований и биоинформатики Университета штата Орегон для секвенирования 16S. Этот центр следует протоколу подготовки библиотеки для метагеномного секвенирования 16S Illumina для секвенирования на MiSeq. Последовательности, специфичные для генов, используемые в этом протоколе, нацелены на область V3 и V4 бактериального гена 16S рРНК. Пары праймеров для бактерий и анализ последовательностей были выполнены с использованием ранее описанных методов в работе Krey et al. (2019). Данные микробного секвенирования доступны в GenBank под номером проекта PRJNA553015.
Анализ метаболитов и питательных веществ листвы
Дериватизация первичных метаболитов и газовая хроматография были проведены с использованием небольшой модификации установленной процедуры (Lee and Fiehn, 2008). Определенное количество порошкообразных лиофилизированных листьев картофеля (ок. 20 мг) было суспендировано в 500 мкл экстракционного растворителя, содержащего метанол, 2-пропанол и воду в соотношении 5:2:2. После добавления 1,5 мкг внутреннего стандарта рибитола материал экстрагировали встряхиванием при комнатной температуре в течение 10 минут (Vortex) и ультразвуковой обработкой при комнатной температуре в течение 10 минут (ультразвуковая баня Branson 5510). Затем экстракты центрифугировали в течение 10 минут при 21 000 g, и супернатанты переносили в новые флаконы. Экстракты высушивали в вакууме. Сухие остатки суспендировали в 500 мкл 50% водного ацетонитрила и реэкстрагировали, как описано выше, путем последовательного встряхивания на вортексе и ультразвуковой обработки. Осадок снова удаляли центрифугированием, и супернатанты высушивали в вакууме. Сухие остатки суспендировали в 10 мкл O-метоксиламина гидрохлорида (30 мг/мл в пиридине, оба от Sigma) и инкубировали в течение 90 минут при 30°C и 1000 об/мин (Eppendorf Thermomixer).
Впоследствии образцы дериватизировали с помощью 90 мкл MSTFA с 1% TMCS (Thermo-Pierce кат. № TS-48915) в течение 30 минут при 37°C и 1000 об/мин (Eppendorf Thermomixer). Анализ методом газовой хроматографии-масс-спектрометрии был выполнен с использованием времяпролетного масс-спектрометра Pegasus 4D (LECO), оснащенного автосамплером Gerstel MPS2 и печью Agilent 7890A. Продукты дериватизации разделяли на колонке Rxi-5Sil® длиной 30 м, внутренним диаметром 0,25 мм, толщиной пленки 0,25 мкм (Restek) с предколонкой IntegraGuard® с использованием сверхчистого He при постоянном потоке 1 мл/мин в качестве газа-носителя. Линейный температурный градиент начинался с выдержки 1 минуты при 50°C, с последующим повышением до 330°C со скоростью 20°C/мин. Конечная температура поддерживалась в течение 5 минут перед возвращением к исходным условиям. Масс-спектры собирали со скоростью 17 спектров/с. Температура инжекционного порта поддерживалась на уровне 250°C, и 1 мкл образца вводили при соответствующем коэффициенте деления потока.
Процентный содержания азота и углерода в листве был количественно определен в листовой ткани с использованием элементного анализатора (ECS4010, Costech Analytical, Valencia, California, USA), следуя методам, описанным Krey et al. (2019).
Статистический анализ
Мы оценили различия в экспрессии генов растений, почвенных микробов и насекомых между системами производства (органическая и традиционная) и сортами картофеля (Norkotah и Alturas). Статистика была проведена в R (версия 3.2.1; R Core Team, 2015), если не указано иное.
Дифференциальная экспрессия
Пакеты EdgeR (Robinson et al., 2010) и limma (Ritchie et al., 2015) были использованы в R studio (v.0.99.903) для расчета дифференциальной экспрессии генов для генов с количеством прочтений > 2. В нашем исследовании мы изучили, какие гены экспрессируются на разных уровнях при сравнении листьев, собранных с растений, выращенных в двух системах управления (органическая и традиционная), а также когда листья имели физические повреждения или выглядели неповрежденными. Для проведения этих сравнений мы создали матрицу дизайна с управлением и насекомыми в качестве «обработки» (где поле является единицей репликации) и сделали контрасты для попарных сравнений отдельно для каждого сорта (см. Supplementary Table 3). Мы добавили поле как случайный эффект, используя функцию duplicateCorrelation, поскольку образцы листьев происходили из одного поля и, следовательно, не были независимы относительно единицы репликации. Количества с соответствующими весами были преобразованы в значения logCPM с использованием функции voom перед тестами на дифференциальную экспрессию в соответствии со стандартными практиками (Chen et al., 2014; Smyth et al., 2020). Значимость была определена с использованием скорректированного значения p FDR на уровне отсечения 10% (adj P < 0,1).
Аннотация дифференциально экспрессируемых генов
Используя Blast2GO (v.4.1.3, Conesa et al., 2005) с базой данных Blast, установленной на «nr», и E-значением 1,0E-3, а также инструментарий MapMan (Thimm et al., 2004), мы смогли присвоить классы генной онтологии нашим дифференциально экспрессируемым генам с совпадениями BLAST с известными последовательностями. Blast2GO также присваивает биологические функции на основе гомологий последовательностей BLAST и аннотаций GO (в отношении биологических процессов, молекулярных функций и клеточных компонентов); каждому контрасту, имеющему дифференциально экспрессируемые гены, была присвоена биологическая функция и соответствующая категоризация (Conesa et al., 2005). Анализ обогащения GO был использован для вычисления обогащения терминов GO, обнаруженных в MapMan, в каждой из контрастных групп. P-значения были скорректированы с использованием FDR Бенджамини и Хохберга с отсечением < 0,1. Анализ обогащения был выполнен с использованием точного теста Фишера, доступного в пакете R «stats».
Обилие насекомых-вредителей и питательные вещества почвы
Предположения модели были проверены с использованием гистограмм, тестов Шапиро-Уилка и графиков остатков по мере необходимости. Для изучения различий в плотности насекомых между режимами производства мы выполнили шесть обобщенных линейных моделей, предполагающих распределение Пуассона: по одной для фитофагов, питающихся флоэмой, трипсов и естественных врагов в пределах каждого из двух исследованных сортов картофеля. Мы рассматривали два сорта картофеля отдельно из-за значительных различий в среде их выращивания. Для изучения различий в физических и питательных профилях почвы между системами земледелия и сортами картофеля мы использовали неметрическое многомерное шкалирование (NMDS) для сжатия многомерных почвенных данных (Supplementary Table 2) до двух осей. Мы использовали функцию metaMDS в пакете vegan языка R для создания матрицы различий с использованием расстояний Брея-Кертиса и использовали функции ordihull и ordiellipse для визуализации расстояний между участками (Oksanen et al., 2007). Значения стресса были <0,2 (0,14), что указывает на то, что ординация была адекватным представлением данных в уменьшенном количестве измерений. Для проверки различий в управлении почвой между режимами производства и сортами картофеля мы выполнили анализ сходства (функция anosim в vegan), сравнивая ранжированные расстояния для каждой группы с нулевыми распределениями, используя 2000 перестановок. Кроме того, концентрации питательных веществ в почве были сравнены с использованием линейной модели, функции lm в lmer4, с режимом производства (традиционный и органический), сортом (Norkotah и Alturas) и их членом взаимодействия в качестве предикторных переменных.
Микробные сообщества почвы
Последовательности почвенных бактерий были обработаны с использованием ранее описанных методов в работе Krey et al. (2019). Phyloseq v1.18.1 (McMurdie and Holmes, 2013) был использован для визуализации данных. Дифференциальная распространенность бактерий между органическими и традиционными почвами была оценена с использованием Deseq2 (Love et al., 2014) внутри пакета phyloseq (test = «Wald», fitType = «local») со значением FDR < 0,1 и log2FoldChange > 4. Композиционное разнообразие было оценено путем применения индекса разнообразия Шеннона, учитывающего количество и обилие видов, с использованием функции estimate_richness в пакете «phyloseq». С каждого поля мы рассчитали выравненность бактериального сообщества на уровне семейства, используя Shannon (H)/ln(богатство). Показатели разнообразия и выравненности были сравнены с помощью ANOVA с режимом производства (традиционный и органический) и сортом (Norkotah и Alturas) в качестве предикторных переменных. Обилие 10 наиболее распространенных семейств бактерий было ln(x+1) трансформировано и сравнено с использованием ANOVA, с режимом производства (традиционный и органический), сортом (Norkotah и Alturas) и их членом взаимодействия в качестве предикторных переменных.
Первичные метаболиты
Идентификация пиков метаболитов листвы была проведена с использованием библиотеки первичных метаболитов Fiehn (Kind et al., 2009). Для идентификации использовался порог идентичности 700. Выравнивание пиков и сравнение спектров были выполнены с использованием функции Statistical Compare программного обеспечения ChromaTOF® (LECO). Внутренний стандарт рибитол и начальный вес ткани были использованы для нормализации. Статистический анализ выбранных метаболитов был проведен с использованием Metaboanalyst 3.0 (Xia and Wishart, 2016). Нормализованные данные были загружены и автоматически масштабированы для дальнейшего одно- и многомерного анализа. Конкретные метаболиты (см. Supplementary Table 4), связанные с защитой и здоровьем растений, были отобраны, и их концентрации были сравнены с использованием перестановочного многомерного дисперсионного анализа с использованием матриц расстояний (перестановки = 999) с использованием функций adonis в пакете vegan языка R, с режимом производства (традиционный и органический) и сортом (Norkotah и Alturas) в качестве предикторных переменных.
Питательные вещества листвы
Процентное содержание углерода, азота и соотношение C:N были измерены в листовой ткани из 120 образцов листьев, поврежденных и неповрежденных фитофагами, с каждого участка отбора проб. Повреждение фитофагами не изменило питательный состав листвы (см. Supplementary Table 5), и это позволило нам объединить повторы (поврежденные и неповрежденные листья), чтобы увеличить нашу способность обнаруживать почвенно-опосредованные различия в качестве растений на разных участках. Мы сравнили C, N и соотношения C:N с помощью 3 отдельных линейных моделей со смешанными эффектами, используя функцию lme в пакете nlme языка R, с режимом производства (традиционный и органический), сортом (Norkotah и Alturas) и их членом взаимодействия в качестве предикторных переменных и полем в качестве случайного эффекта.
Результаты
Транскриптомика растений и дифференциально экспрессируемые гены
Сорта Norkotah и Alturas значительно различались по количеству дифференциально экспрессируемых генов во всех обработках (DEG) (Supplementary Table 3). Исследование транскриптома выявило 6,985 дифференциально экспрессируемых генов у Norkotah в поврежденной ткани (объединяя органическую и традиционную) по сравнению с неповрежденной листовой тканью, в то время как у Alturas было 1,585 DE генов. Для Norkotah питание насекомыми привело к более высокому обогащению защитными генами в транскриптах растений из органических систем по сравнению с традиционными (гены, связанные со стрессом, P = 0,03; гены, связанные с вторичными метаболитами, P < 0,001; Figure 1, stress perception; Table 1; Supplementary Table 6), и имело больше активированных генов, связанных со стрессом растений (P = 0,01; Figure 1, stress perception; Table 1; Supplementary Table 6). У Norkotah мы наблюдали сильный индуцированный повреждением ответ в сигнальном пути этилена (ET) в органических системах (Figure 1, stress perception; Table 1), в то время как у Alturas в целом было меньше дифференциально экспрессируемых генов, и большинство из них были подавлены во всех обработках (Table 1; Supplementary Table 3). В органических поврежденных насекомыми листьях Norkotah гены, связанные со светособиранием и фотосинтетической ассимиляцией углерода, значительно снизились (Figure 1, growth and development; Table 1; Supplementary Table 6), гены, связанные с фотосинтетической ассимиляцией (PS) и транспортом питательных веществ, были более обогащены в транскриптах (PS, P < 0,001; Table 1; Supplementary Table 6, transport, P = 0,01; Table 1; Supplementary Table 6), и больше генов PS были подавлены (P < 0,001; Table 1; Supplementary Table 6).
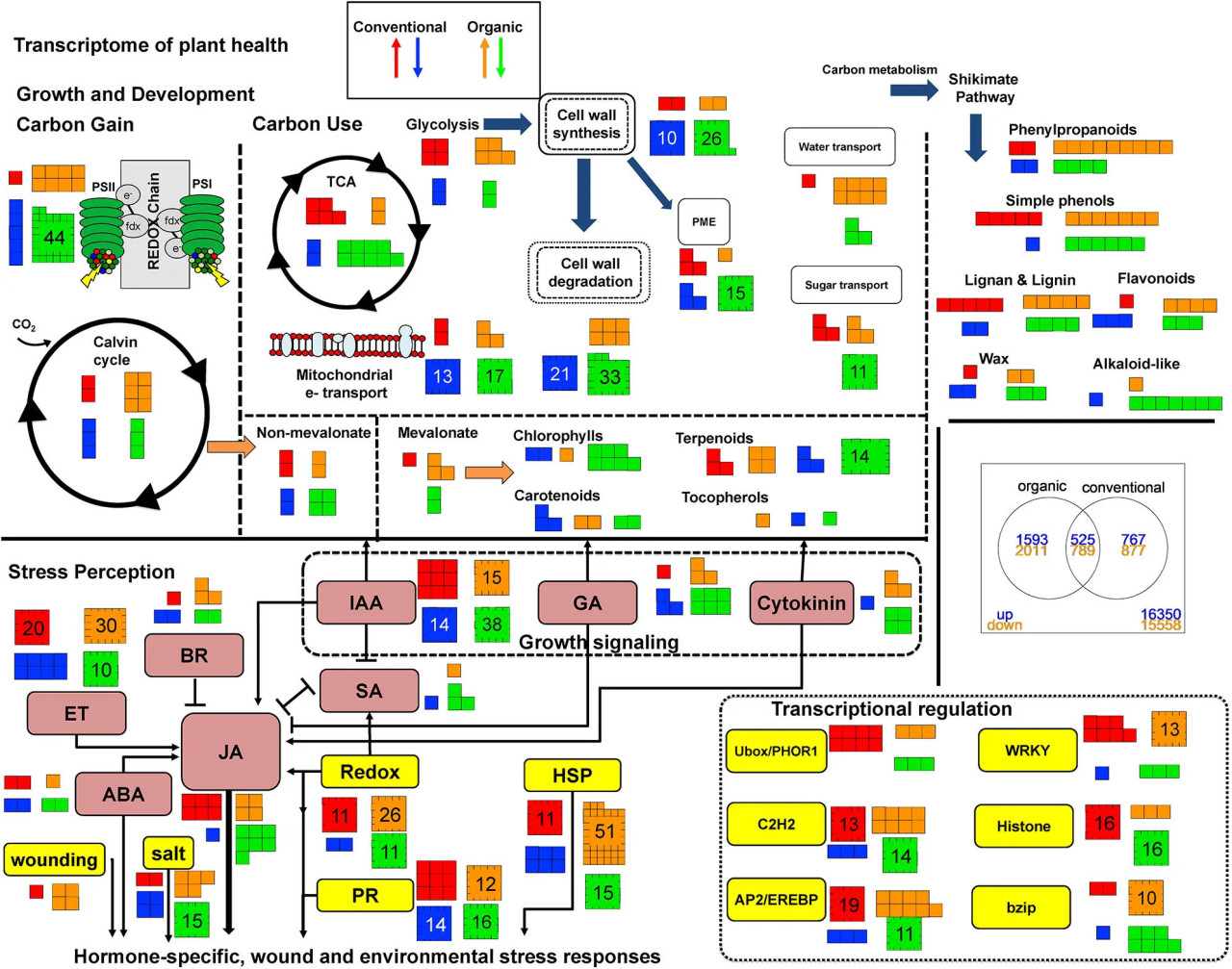
Фото 1 Это графическое представление транскриптома для процессов, связанных со здоровьем и защитой растений, сравнивает растения картофеля Norkotah, выращенные с использованием органических (оранжевые и зеленые стрелки) и традиционных (красные и синие стрелки) методов ведения сельского хозяйства, между неповрежденными и поврежденными насекомыми листьями. Повышение или понижение активности генов указано относительно того, как растения реагируют на повреждения при разных режимах управления. Цифры показывают, сколько генов активируется или подавляется. Текстовые поля с рамками имеют цветовую кодировку для метаболитов, специфически участвующих в гормональной сигнализации/восприятии: розовый; и негормональных процессах, опосредующих стрессовый ответ/восприятие: желтый. Стрелки связывают поток от первичного ко вторичному метаболизму (вверху) и синергическую или антагонистическую сигнализацию во время восприятия стресса (внизу).
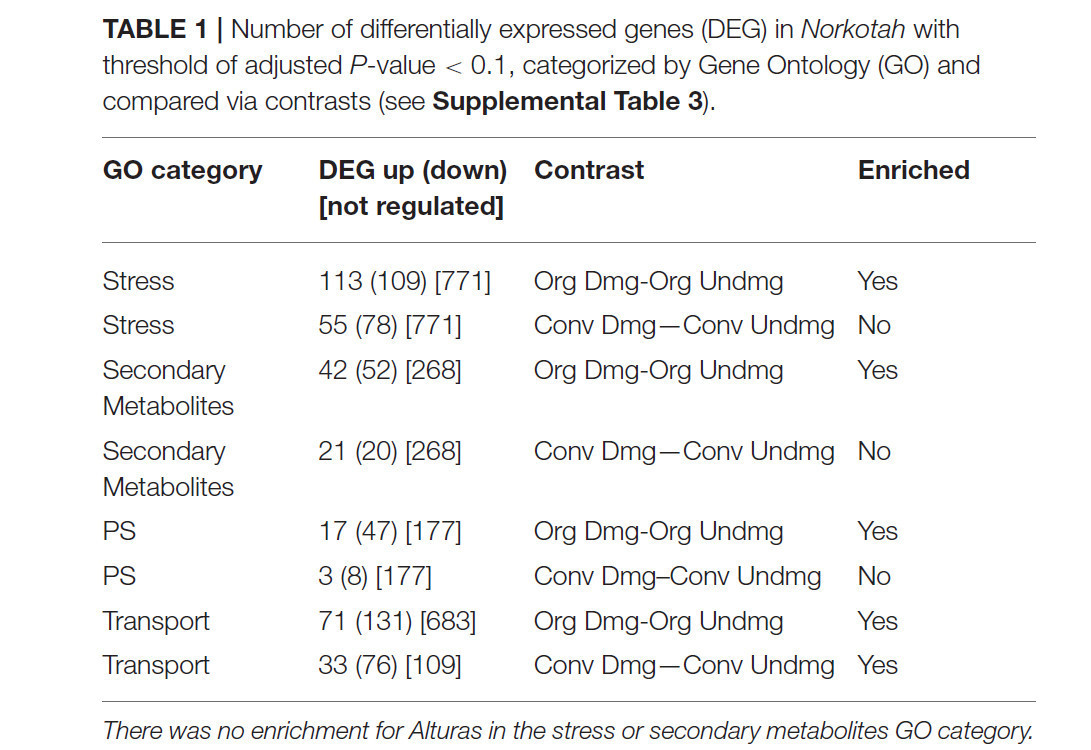
Таблица 1 Количество дифференциально экспрессируемых генов (DEG) у Norkotah с порогом скорректированного P-значения < 0,1, категоризированных по генной онтологии (GO) и сравненных с помощью контрастов (см. Supplemental Table 3).
Экспрессия генов, связанных со стрессом, была обогащена в транскриптоме для обоих сортов поврежденных листьев (Norkotah, P = 0,005; Alturas, P = 0,001), в то время как больше генов, связанных со стрессом, были активированы у Norkotah (P < 0,001; Table 1) и подавлены (P < 0,001) у Alturas. В поврежденных листьях Norkotah наблюдалось увеличение количества активированных генов в сигнальных путях салициловой кислоты (SA) и ET по сравнению с неповрежденными листьями (Figure 1), в то время как в поврежденных листьях Alturas наблюдалось больше подавленных генов (Supplementary Table 3, contrast 2 and 3). Когда мы объединили образцы из органических и традиционных систем, чтобы повысить нашу способность изучать воздействие фитофагов, поврежденные листья привели к большему количеству подавленных генов, связанных с фотосинтетической ассимиляцией (PS), у Norkotah (P < 0,001; Table 1), в то время как у Alturas этого не произошло (P = 0,9445).
Плотность вредителей на картофельных полях
Органический картофель Norkotah имел более высокую численность фитофагов, питающихся флоэмой, по сравнению с традиционными растениями (Коэффициент = 0,606, SE = 0,191, Z = 3,16, P = 0,0016, Figure 2A), схожую численность трипсов (Коэффициент = 0,051, SE = 0,073, Z = 0,694, P = 0,488, Figure 2B) и больше естественных врагов (Коэффициент = 1,17, SE = 0,155, Z = 7,511, P = < 0,0001, Figure 2C). Органический картофель Alturas имел меньше трипсов, чем традиционные растения (Коэффициент = 0,49, SE = 0,135, Z = −3,62, P = 0,0002, Figure 2D), схожую численность фитофагов, питающихся флоэмой (Коэффициент = −0,265, SE = 0,222, Z = −1,193, P = 0,233, Figure 2E), и схожую численность естественных врагов (Коэффициент = 0,361, SE = 0,228, Z = 1,583, P = 0,114, Figure 2F).
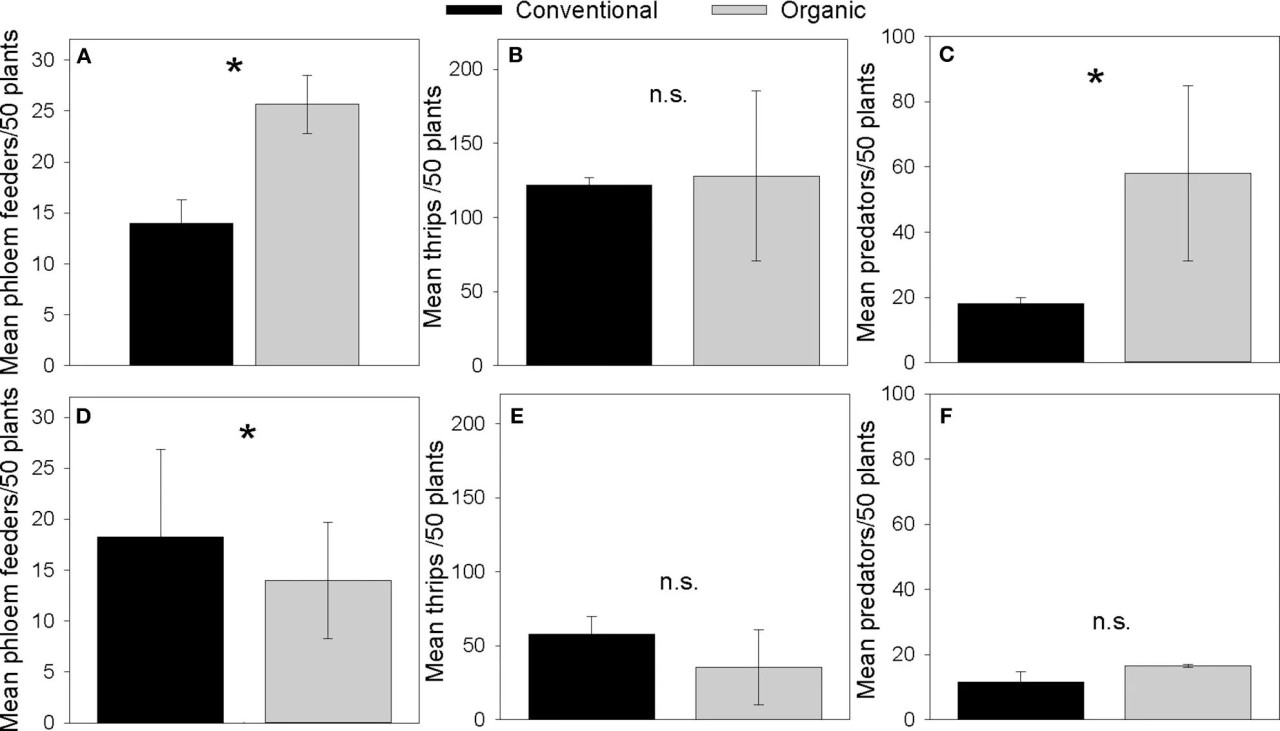
Фото 2 Средняя численность фитофагов, питающихся флоэмой (A,D), трипсов (B,E) и естественных врагов (C,F), собранных в вакуумных пробах с картофельных полей Norkotah (A–C) и Alturas (D–F) в 2015 году. указывает на P < 0,01.
Свойства почвы
Физические и химические характеристики почвы на органически управляемых участках, где выращивался картофель Norkotah, были значительно непохожи на традиционные участки и участки, где выращивался картофель Alturas (ANOSIM R = 0,2946, P = 0,049, Figure 3). Почти в 4 раза больше нитратов было доступно на традиционных полях Norkotah по сравнению с органическими, тогда как на полях Alturas уровень нитратов был в 4 раза ниже на традиционных полях по сравнению с органическими (взаимодействие управлениесорт: F1,7 = 15,338, P = 0,005; Figure 4A). Концентрации фосфора примерно удвоились на органических и традиционных полях Alturas и незначительно различались между сортами [F(1, 7) = 4,162, P = 0,06; Figure 4B]. Микробная биомасса была выше на органических полях Norkotah, чем на Alturas [взаимодействие управлениесорт: F(1, 7) = 6,458, P = 0,038; Figure 4E].
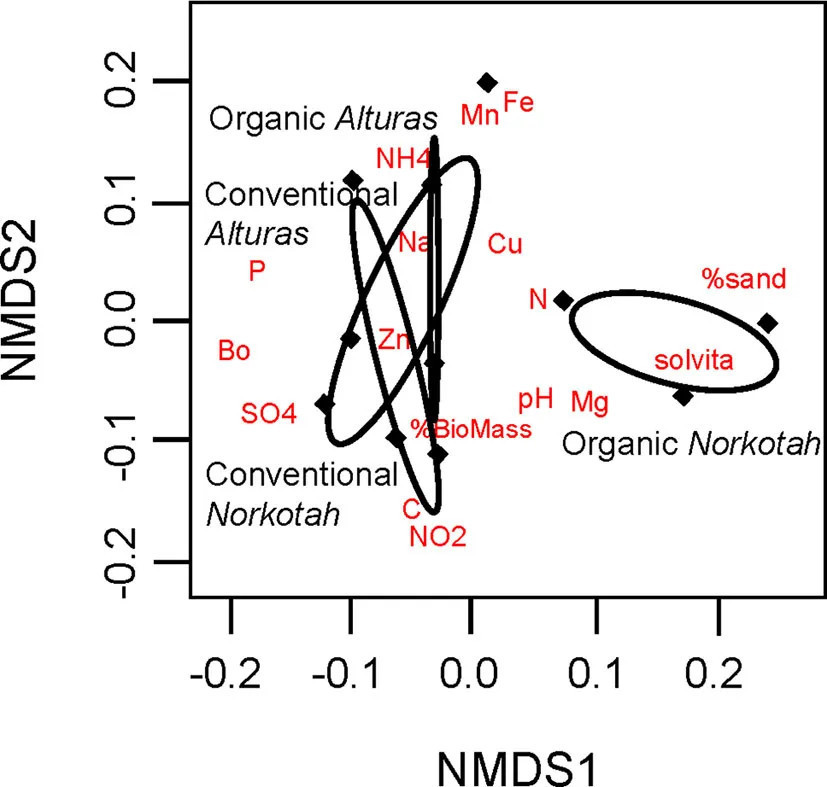
Фото 3 Физические и химические профили почвы, отображенные на двух осях неметрического многомерного шкалирования. Эллипсы показывают кластеризацию наших четырех групп обработки, которые состояли из двух систем земледелия (традиционная и органическая) и двух сортов картофеля (Norkotah и Alturas). Красные метки указывают на расположение почвенных характеристик, которые различают кластеры.
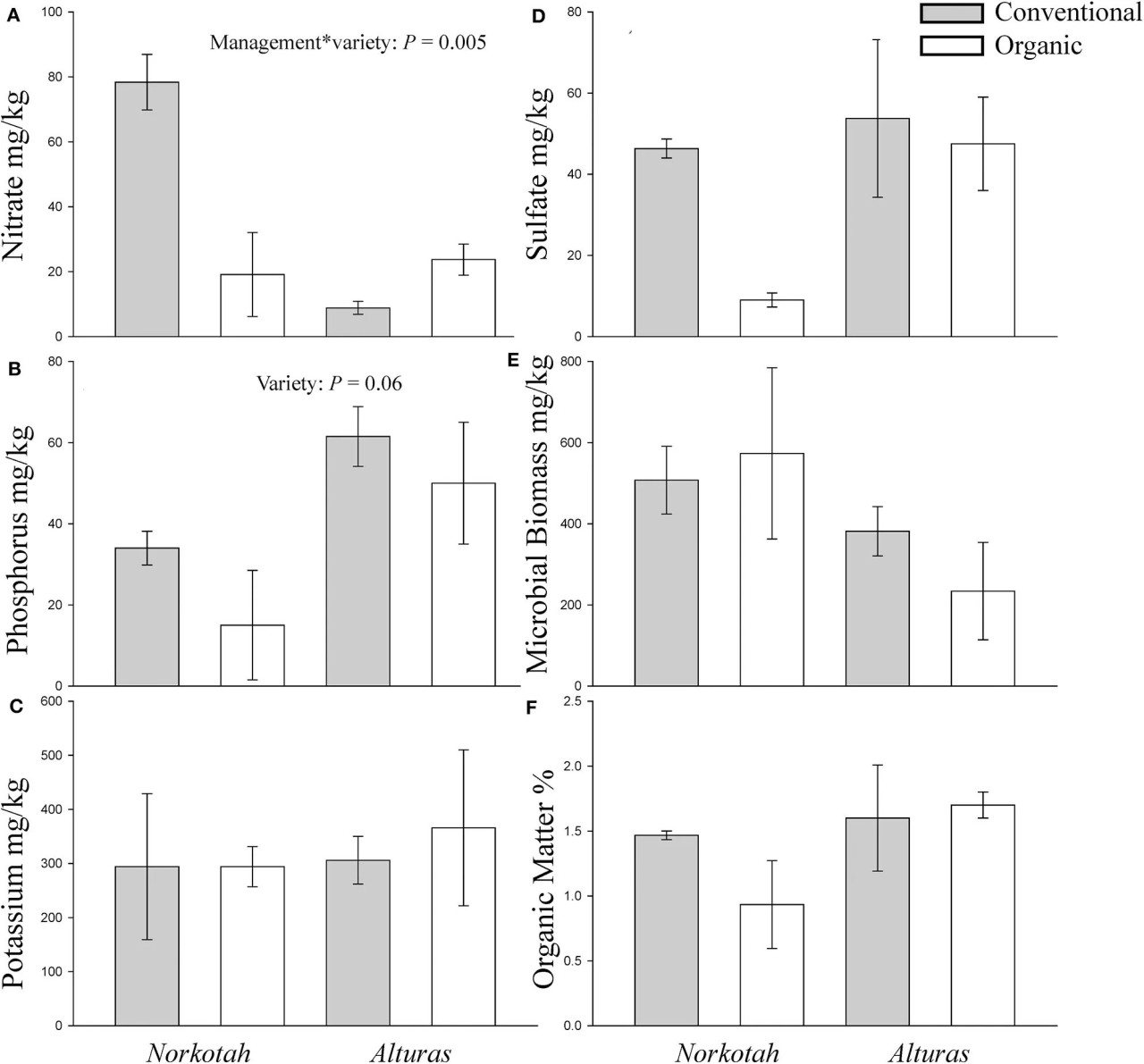
Фото 4 Концентрации питательных веществ в почве, отобранной на органических и традиционных картофельных полях в середине сезона. (A) Нитрат, (B) Фосфор, (C) Калий, (D) Сульфат, (E) Микробная биомасса и (F) Органическое вещество. Серые столбцы указывают на традиционную почву, белые столбцы — на органическую.
Микробные сообщества почвы
Не было различий в бактериальном разнообразии почвы между системами управления [F(1, 7) = 0,025, P = 0,879; Supplementary Table 7] или сортами [F(1, 7) = −0,05, P = 0,830; Supplementary Table 7], и выравненность сообществ также не различалась в зависимости от системы управления [F(1, 7) = 0,009, P = 0,926; Supplementary Table 7] или сорта [F(1, 7) = 0,005, P = 0,944; Supplementary Table 7]. Десять бактериальных таксонов были дифференциально обильны при органическом управлении по сравнению с традиционным (Padj < 0,1; Supplementary Table 8). Бактерии из семейств Bacilliaceae, Chitinophagaceae и Planctomycetaceae были более обильны на органических полях Norkotah по сравнению с традиционными, но менее обильны на органических полях Alturas по сравнению с традиционными (Bacilliaceae; взаимодействие управлениесорт: F = 5,768, P = 0,0173, Chitinophagaceae; взаимодействие управлениесорт: F = 3,921, P = 0,0478, Planctomycetaceae; взаимодействие управлениесорт: F = 25,550, P < 0,001), в то время как другие таксоны не различались значимо по сортам.
Анализ питательных веществ листвы и первичные метаболиты
Процентное содержание углерода в листовой ткани не различалось между обработками (t = −0,321, P = 0,7491; Supplementary Table 5) или сортами (t = 0,436, P = 0,6637; Supplementary Table 5). Процентное содержание азота было выше у Norkotah по сравнению с Alturas (t = 2,667, P = 0,009), и соотношение углерод:азот в традиционных системах незначительно снижалось в листовой ткани, но только на полях Norkotah, в то время как традиционное управление увеличивало соотношение углерод:азот на полях Alturas (t = 0,1,963, P = 0,052). Набор первичных метаболитов (Supplementary Table 4) в листовой ткани не различался в зависимости от сорта, управления или повреждения фитофагами (R² = 0,126, df = 1, 11, P = 0,22).
Обсуждение
Нашей главной целью было сравнить характер экспрессии генов растений в традиционных и органических системах земледелия. Для этого мы оценили экспрессию защитных генов от фитофагов в растительной ткани, собранной в открытом поле на действующих фермах, и количественно определили несколько потенциально связанных с этим переменных окружающей среды (например, питательные вещества почвы, микробные сообщества, прессинг фитофагов и т.д.). Наиболее выраженным эффектом органического земледелия, который мы наблюдали, было сильное увеличение активности нескольких генов, связанных с защитой от фитофагов, в поврежденной насекомыми листовой ткани, тогда как эти гены не экспрессировались дифференциально у растений, подвергшихся нападению фитофагов на традиционных полях. Важно отметить, что эти реакции были специфичны для сорта: мы наблюдали эту картину повышенной экспрессии защитных генов у Norkotah, который обычно продается как свежий продукт и более восприимчив к стрессу, но не у Alturas, который в основном перерабатывается на обезвоженные продукты и выведен для большей толерантности к стрессам (Novy et al., 2003; Stark et al., 2020).
Различия в экспрессии генов растений между неповрежденными традиционными и органическими листьями картофеля
Boorse (1977) определяет нездоровые или болезненные состояния как возникающие, когда нормальная функция нарушена или ограничена. Таким образом, здоровье — это отсутствие нарушений или ограничений. Учитывая, что нормальная функция зависит от множества параметров, наше определение здоровья растений для этой статьи использует в качестве базового уровня экспрессию генов у традиционно выращиваемого картофеля. Любое изменение обилия транскриптов, которое представляет собой усиление или ограничение процессов, связанных с метаболическим гомеостазом или восприятием стрессоров окружающей среды (абиотических или биотических), считается изменением здоровья растений. Учитывая количество взаимодействий и попарных сравнений, которые можно было бы сделать, логичной базовой линией является оценка вариаций между неповрежденными традиционно выращенными растениями и неповрежденными органически выращенными растениями, в дальнейшем называемыми традиционными и органическими растениями соответственно. На этом базовом уровне органические растения имеют более высокое обилие транскриптов роста и транскриптов, связанных с патогенным стрессом, и более низкое обилие транскриптов, связанных с абиотическим стрессом, по сравнению с традиционными растениями (Figure 1). Более конкретно, органические растения показывают более высокую экспрессию генов фотосинтеза и метаболизма клеточной стенки (P < 0,01) и тенденции к более высокой экспрессии, связанной с развитием, и сниженной экспрессии, связанной со стрессом (P < 0,1). Напротив, Pacifico et al. (2017) обнаружили больше транскриптов, связанных со стрессом, с акцентом на метаболизм фенилпропаноидов, в один год и обогащение вторичным метаболизмом (флавоноидами) во второй год на посевах картофеля. Энергетический метаболизм, метаболизм клеточной стенки, сигнализация и регуляция РНК были стабильно дифференциально экспрессированы в течение обоих лет по сравнению с традиционными растениями, хотя величины и направления генов были вариабельны (Pacifico et al., 2017). Кроме того, van Dijk et al. (2012) обнаружили, что обработка органическими удобрениями на посевах картофеля увеличивала экспрессию защитных генов, опосредованных липоксигеназой, но органическая защита растений подавляла этот тип экспрессии генов. Эта направленная экспрессия будет взаимодействовать в полевых условиях, где одновременно применяются как стратегии удобрения, так и защиты растений, и может объяснить, почему в наших данных было обнаружено, что только два защитных транскрипта, связанных с lox, были активированы у органических растений по сравнению с традиционными (Figure 1; Supplementary Table 6).
Некоторые специфические процессы, которые показали повышенное обилие транскриптов, включали процессы, участвующие в светособирании, ген снижения деградации крахмала (B-амилаза) и увеличение мобилизации сахаров для формирования клеточной стенки (Supplementary Table 6). Интерес представляет активация нескольких генов, связанных с метилэстеразами пектина (PME), ключевыми ферментами, которые деэтерифицируют пектины для укрепления клеточных стенок. PME также увеличиваются во время распознавания патогенов и зависят от сигнализации JA, смягченной факторами ответа на этилен (ERF), когда атакуют некротрофы, и от сигнализации как ERF, так и MYC, когда атакуют биотрофы (Bethke et al., 2014). Конститутивная активность JA у дикого типа картофеля увеличивает PME для снижения метилэтерификации пектина и изменения матрицы пектина в пользу повышенной устойчивости к некоторым патогенам (Taurino et al., 2014).
GO анализ показал, что стрессовые реакции имели тенденцию к обогащению (P < 0,1; Table 1), но несколько отдельных транскриптов накапливались больше в органических растениях. В частности, транскрипты защитного вторичного метаболизма (терпеноиды, алкалоиды) и гормональной сигнализации (синтез JA, синтез/сигнализация ET) были выше, а реакция на абиотический стресс, связанный с температурой и ранением, была снижена по сравнению с традиционными растениями. Lox1 и HPL показывают более высокую экспрессию в органических растениях, что указывает на то, что биосинтез жирных кислот (C-6 альдегиды и w-кето FA) или вышестоящие защитные метаболиты могут быть повышены. Это предполагало бы активную систему защиты в случае атаки или более высокий конститутивный уровень защиты из-за лучшего здоровья растений, праймирования почвенными микробами или того и другого. Гены биотического стресса, связанные с реакцией на патогены, показали повышенную экспрессию, как и различные p450 и UGT, гены, участвующие в изменении растворимости метаболитов для вторичного метаболизма и детоксикации. Также следует подчеркнуть, что, хотя активация ряда генов, участвующих в защитной реакции, указывает на сильный ответ, это не обязательно указывает на эффективный ответ. Отмечаем, что требуются дальнейшие исследования, чтобы продемонстрировать, что любые различия в экспрессии генов между системами земледелия действительно повышают способность растения подавлять повреждения от вредителей.
В целом, неповрежденные органические растения показали лучшее здоровье, определяемое накоплением транскриптов в пользу роста, развития и защитной сигнализации. Учитывая, что компромиссы между ростом и защитой в значительной степени опосредованы гормональными взаимодействиями, включая антагонизмы, положительная реакция роста наряду с защитной сигнализацией, вероятно, означает, что растения находятся в более высоком конститутивном или праймированном состоянии здоровья, а не в защитном состоянии, вызванном индукцией вредителями. Праймирование наиболее выгодно, когда атака неизбежна, что делает органическое картофельное поле в середине сезона оптимальным кандидатом для инвестиций в защиту от атаки в конце сезона, когда клубнеобразование наиболее интенсивно. Таким образом, будущая гипотеза для проверки касается временной природы праймирования: всегда ли органическое управление вызывает более высокие конститутивные процессы роста и защиты, или праймирование специфично с фенологической или временной точки зрения?
Различия в экспрессии генов растений, индуцированные грызущими фитофагами
Когда поврежденные и неповрежденные растения (объединяя системы производства), 6985 генов были дифференциально экспрессированы. Из них 1402 становятся значимыми только при объединении традиционных и органических образцов, что указывает на взаимодействие между стратегией управления и повреждением. Действительно, график MDS иллюстрирует то же взаимодействие (Supplementary Figure 1). См. Supplemental Table 4 для данных по отдельным генам (базовый уровень O против C неповрежденных). В условиях повреждения органические растения показывают больше дифференциально экспрессируемых генов, чем традиционные (3604 против 1644), но имеют 1314 общих генов, экспрессируемых в одном направлении при повреждении (Supplementary Figures 2, 3). Было обнаружено, что 30 генов экспрессируются в противоположных направлениях в зависимости от управления, а остальные гены были либо активированы, либо подавлены в одной системе производства, но не были значимо экспрессированы в другой. Количество генов, дифференциально экспрессируемых между традиционным и органическим производством во всех образцах, невелико (2), потому что неповрежденные растения имеют картины экспрессии генов в противоположных направлениях (Supplementary Figure 2). В неповрежденных органических растениях активировано больше генов, чем подавлено, по сравнению с неповрежденными традиционными растениями (Supplementary Figure 2). Однако эти различия в экспрессии генов исчезают, когда на растения нападают грызущие фитофаги. Независимо от стратегии управления, у поврежденных растений Norkotah было меньше транскриптов, связанных с ростом, и больше защитных транскриптов. Например, гены, связанные с фотосинтезом, мобилизацией углерода, клеточным циклом, ростом и развитием, показали больше подавлений, чем активаций (Figure 1). Гены, связанные с метаболизмом аминокислот, вторичным метаболизмом, редокс-регуляцией и деградацией ксенобиотиков, показали больше активаций, чем подавлений (Figure 1). Эти картины позволяют предположить, что защитный метаболизм активен для защиты от биотического стресса, вызванного дефолиацией или другой атакой вредителей на поврежденных листьях, в то время как рост подавлен.
При раздельном сравнении, на 1960 транскриптов больше дифференциально экспрессируется в органических растениях, чем в традиционных, испытывающих биотическую атаку. Генные онтологии, которые чрезмерно представлены, включают PS, липидный метаболизм, метаболизм клеточной стенки, вторичный метаболизм, стресс и развитие, тогда как традиционные растения показали чрезмерную представленность в генах фотосинтеза, метаболизма белков и сигнализации (P < 0,05). Разница в реакции интригует, потому что профили обогащения экспрессии генов показывают, что органические растения реагируют с большим компромиссом между ростом и защитой, что можно охарактеризовать как типичную или здоровую реакцию. Традиционные растения показывают обогащение в меньшем количестве путей, которые включают ожидаемое подавление фотосинтеза при повреждении, но показывают дифференциальную экспрессию в сигнализации и обороте белков. Таким образом, компромисс между ростом и защитой активен при атаке, но вторичный метаболизм и опосредованные гормонами защитные пути не обогащены, даже если отдельные гены поддерживают этот компромисс.
Различия в экспрессии защитных генов между двумя сортами картофеля в органическом и традиционном земледелии показывают, что Alturas в целом менее чувствителен к повреждениям фитофагами, чем Norkotah. Это может быть связано с тем, что Alturas в основном выращивается для промышленности по производству обезвоженных продуктов (где форма клубня не контролируется) и был выведен на толерантность к низким дозам азота, устойчивость к Verticillium dahliae (возбудителю раннего увядания) и другим стрессам (Novy et al., 2003). Эти признаки могут вступать в компромисс с устойчивостью к фитофагам (например, Fineblum and Rausher, 1995; Rudgers et al., 2004). Кроме того, из-за их толерантности к неоптимальным почвенным условиям, картофель Alturas может успешно выращиваться в менее благоприятных обстоятельствах (например, на полях с низким плодородием почвы, маргинальных землях с камнями, в интенсивных картофельных севооборотах без сидератов), чем некоторые другие сорта (Novy et al., 2003). По этой причине растения Alturas могут быть хозяевами более низкого качества для фитофагов и поэтому подвергаться меньшему повреждению (например, Awmack and Leather, 2002). С другой стороны, растения Norkotah обычно производятся для свежего рынка в Тихоокеанском Северо-Западе и относительно восприимчивы к фитофагам, но методы производства будут существенно различаться в зависимости от того, будет ли собранный урожай храниться или продаваться непосредственно с поля (Zvomuya et al., 2002; Stark et al., 2020). Хотя Norkotah устойчив к обыкновенной парше и толерантен к некоторым вирусам, исследование показало, что у персиковой тли скорость роста на растениях Norkotah была ниже, чем на растениях Alturas (Draper et al., 2002; Davis et al., 2007; Stark et al., 2020). Это также может частично объяснить, почему Norkotah имел более сильную защитную реакцию на атаку фитофагов, чем Alturas (Davis et al., 2007). Как правило, лучшие поля и более высокие дозы удобрений используются для поддержания растений Norkotah как можно дольше в конце сезона, и они могут получать меньше удобрений, чем другие сорта (Zvomuya et al., 2002; Stark et al., 2020). Действительно, процент азота в листовой ткани был значительно выше у Norkotah, чем у Alturas, что делает его более качественным источником пищи для фитофагов (Mattson, 1980; Awmack and Leather, 2002). Таким образом, индукция химической защиты может быть более важна для Norkotah из-за признаков растения и относительно более высокого прессинга со стороны фитофагов (например, Wink, 1988).
Мы измерили несколько факторов окружающей среды, чтобы выявить те, которые могут объяснить более сильную активацию защитных генов растений в органических системах по сравнению с традиционными. Во-первых, мы количественно определили питательные вещества почвы и провели секвенирование микробных сообществ почвы на каждом участке, где мы брали образцы растений. На традиционных полях мы наблюдали большую концентрацию нитратов там, где выращивался Norkotah, по сравнению с Alturas (Figure 4). Азотное удобрение может улучшить качество растения-хозяина для фитофагов и связано с увеличением численности насекомых, питающихся флоэмой (Strauss, 1987; Lou and Baldwin, 2004; Staley et al., 2010). Дополнение азотом может заставить растения сокращать инвестиции в химическую защиту (Glynn et al., 2003; Donaldson et al., 2006), особенно быстро высвобождаемые азотные добавки, вносимые на традиционных полях (Staley et al., 2010). Следовательно, избыточная доступность N на традиционных полях, где рос Norkotah (Figure 4), могла снизить экспрессию защитных генов, как это наблюдалось в других обработках.
Использование органических почвенных добавок также может быть связано с желательными свойствами почвы, такими как повышенная плотность полезных почвенных микроорганизмов (Drinkwater et al., 1995; Doran, 2002). Мы наблюдали увеличение микробной биомассы почвы на органических полях Norkotah по сравнению с Alturas (Figure 4E). Несколько исследований показали, что полезные подземные микроорганизмы могут повышать устойчивость растений к стрессу и усвоение питательных веществ (Bloemberg and Lugtenberg, 2001; Lugtenberg et al., 2002; Bais et al., 2004; Morrissey et al., 2004; Goh et al., 2013; Blundell et al., 2020). Многие исследования обнаруживают увеличение микробной биомассы (Gunapala and Scow, 1998; Swezey et al., 1998; Altieri, 1999; Mäder et al., 2002; Reganold et al., 2010) и разнообразия (Ge et al., 2008; Reilly et al., 2013; Lupatini et al., 2017) в ответ на органическое управление почвой, но это не стабильно во всех системах (Liu et al., 2007) и может быть специфичным для участка (Blundell et al., 2020). Картофель Alturas, который выживает в условиях дефицита питательных веществ, может представлять собой еще одно исключение. Важно также признать, что наши поля не были попарно сопоставлены с соответствующими почвообразующими факторами, которые, как было показано, влияют на различия между органическим и традиционным земледелием (Reganold et al., 1993, 2010; Reganold, 2013). Когда мы сравнили широкие сообщества бактерий в почве, в целом мы обнаружили мало различий между органическими и традиционными полями (Supplementary Table 7).
В целом, Norkotah испытывал более высокое давление со стороны фитофагов, чем Alturas, и имел соответственно более сильную реакцию в активности защитных генов (Figures 1, 2). Эта картина давления фитофагов является одной из потенциальных причин, по которой индукция защитных генов была специфична для сорта. Предыдущее повреждение фитофагами может праймировать индуцированные защиты для более сильной экспрессии позже в сезоне (Conrath et al., 2006; Frost et al., 2008) и также может отвечать за различные картины индукции в нашей системе. Использование инсектицидов является еще одним потенциальным механизмом, объясняющим различия в защитной активности, которые мы наблюдали. В традиционных посевах картофеля обычно применяются неоникотиноиды, но их использование запрещено на органических полях (Koss et al., 2005). Предыдущие работы показывают, что неоникотиноиды могут ослаблять защитную активность растений (например, James and Price, 2002; Szczepaniec et al., 2013) и, возможно, подавляли экспрессию защитных генов на наших традиционно управляемых полях. Кроме того, наше исследование уникально тем, что оно оценивает регуляцию защитных генов в реальных системах ведения сельского хозяйства, характеризующихся многочисленными взаимодействующими факторами окружающей среды. Важно признать, что этот подход не позволил нам полностью исключить фитофагов с наших «неповрежденных» растений, особенно фитофагов, питающихся флоэмой, чьи повреждения часто не видны визуально. Поэтому мы не можем однозначно различить изменения в экспрессии генов стресса растений, вызванные фитофагией и характеристиками почвы. Тем не менее, низовые силы в органическом производстве должны изучаться в полевых условиях, если мы хотим понять их истинное значение в обеспечении устойчивости к фитофагам.
Секвенирование РНК (RNA-seq) является мощным инструментом для транскрипционного профилирования для выявления генов, связанных с растениями, раскрывая сети генов, которые функционируют скоординированно для управления метаболическими путями или клеточными процессами. Например, было показано, что системы земледелия влияют на экспрессию генов, при этом органическое управление влияет на биологические процессы и пути, связанные с качеством растений (Lu et al., 2005; Pacifico and Paris, 2016; Pacifico et al., 2017). Однако почти все исследования, изучающие экспрессию генов и системы земледелия, проводятся в контролируемых условиях и могут быть ограничены в понимании реакций растений на множественные одновременные стрессы, как это происходит в реальных условиях. Когда стрессы возникают последовательно или вместе, активируются определенные гены, которые могут не индуцироваться независимо (Rizhsky et al., 2002, 2004; Mittler, 2006). Будущие исследования должны учитывать все факторы, с которыми растения на коммерческих фермах сталкиваются в окружающей среде. Наше исследование позволило нам показать, что специфические картины экспрессии генов могут иметь полезные приложения для определения различий между органически и традиционно выращенными растениями. Связывание экспрессии генов с конкретными защитными признаками растений является ключевым первым шагом к использованию этих защитных механизмов для улучшения борьбы с травоядными сельскохозяйственными вредителями (Mitchell et al., 2016). Для сельскохозяйственных культур этот подход может предложить способы использования систем земледелия для усиления защитных механизмов растений и, следовательно, естественной борьбы с вредителями.
Дополнительный материал
Дополнительный материал к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fsufs.2020.00097/full#supplementary-material
Ссылки
1. Altieri M. A. (1999). The ecological role of biodiversity in agroecosystems. Agric. Ecosyst. Environ. 74, 19–31. doi: 10.1016/S0167-8809(99)00028-6. CrossRef Google Scholar.
2. Altieri M. A., Nicholls C. I. (2003). Soil fertility management and insect pests: harmonizing soil and plant health in agroecosystems. Soil Tillage Res. 72, 203–211. doi: 10.1016/S0167-1987(03)00089-8. CrossRef Google Scholar.
3. Anders S., Pyl P. T., Huber W. (2015). HTSeq—a Python framework to work with high-throughput sequencing data. Bioinformatics 31, 166–169. doi: 10.1093/bioinformatics/btu638. CrossRef Google Scholar.
4. Awmack C. S., Leather S. R. (2002). Host plant quality and fecundity in herbivorous insects. Ann. Rev. Entomol. 47, 817–844. doi: 10.1146/annurev.ento.47.091201.145300. CrossRef Google Scholar.
5. Bais H. P., Park S.-W., Weir T. L., Callaway R. M., Vivanco J. M. (2004). How plants communicate using the underground information superhighway. Trends Plant Sci. 9, 26–32. doi: 10.1016/j.tplants.2003.11.008. CrossRef Google Scholar.
6. Beecher N. A., Johnson R. J., Brandle J. R., Case R. M., Young L. J. (2002). Agroecology of birds in organic and nonorganic farmland. Conserv. Biol. 16, 1620–1631. doi: 10.1046/j.1523-1739.2002.01228.x. CrossRef Google Scholar.
7. Bengtsson J., Ahnström J., Weibull A. C. (2005). The effects of organic agriculture on biodiversity and abundance: a meta-analysis. J. Appl. Ecol. 42, 261–269. doi: 10.1111/j.1365-2664.2005.01005.x. CrossRef Google Scholar.
8. Bethke G., Grundman R. E., Sreekanta S., Truman W., Katagiri F., Glazebrook J. (2014). Arabidopsis pectin methylesterases contribute to immunity against Pseudomonas syringae. Plant Physiol. 164, 1093–1107. doi: 10.1104/pp.113.227637. CrossRef Google Scholar.
9. Bhardwaj D., Ansari M. W., Sahoo R. K., Tuteja N. (2014). Biofertilizers function as key player in sustainable agriculture by improving soil fertility, plant tolerance and crop productivity. Microb. Cell Factor. 13:66. doi: 10.1186/1475-2859-13-66. CrossRef Google Scholar.
10. Birkhofer K., Bezemer T. M., Bloem J., Bonkowski M., Christensen S., Dubois D., et al. (2008). Long-term organic farming fosters below and aboveground biota: Implications for soil quality, biological control and productivity. Soil Biol. Biochem. 40, 2297–2308. doi: 10.1016/j.soilbio.2008.05.007. CrossRef Google Scholar.
11. Bloemberg G. V., Lugtenberg B. J. J. (2001). Molecular basis of plant growth promotion and biocontrol by rhizobacteria. Curr. Opin. Plant Biol. 4, 343–350. doi: 10.1016/S1369-5266(00)00183-7. CrossRef Google Scholar.
12. Blundell R., Schmidt J. E., Igwe A., Cheung A. L., Vannette R. L., Gaudin A. C., et al. (2020). Organic management promotes natural pest control through enhanced plant resistance to insects. Nat. Plants 6, 483–491. doi: 10.1038/s41477-020-0656-9. CrossRef Google Scholar.
13. Bolger A. M., Lohse M., Usadel B. (2014). Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120. doi: 10.1093/bioinformatics/btu170. CrossRef Google Scholar.
14. Bonanomi G., Antignani V., Capodilupo M., Scala F. (2010). Identifying the characteristics of organic soil amendments that suppress soilborne plant diseases. Soil Biol. Biochem. 42, 136–144. doi: 10.1016/j.soilbio.2009.10.012. CrossRef Google Scholar.
15. Boorse C. (1977). Health as a theoretical concept. Philos. Sci. 44, 542–573. doi: 10.1086/288768. CrossRef Google Scholar.
16. Bouvier J.-C., Ricci B., Agerberg J., Lavigne C. (2011). Apple orchard pest control strategies affect bird communities in southeastern France. Environ. Toxicol. Chem. 30, 212–219. doi: 10.1002/etc.377. CrossRef Google Scholar.
17. Broekgaarden C., Poelman E. H., Steenhuis G., Voorrips R. E., Dicke M., Vosman B. (2007). Genotypic variation in genome-wide transcription profiles induced by insect feeding: Brassica oleracea – Pieris rapae interactions. BMC Genomics 8:239. doi: 10.1186/1471-2164-8-239. CrossRef Google Scholar.
18. Chen Y., McCarthy D., Robinson M., Smyth G. K. (2014). EdgeR: Differential Expression Analysis of Digital Gene Expression Data User's Guide. Bioconductor User's Guide. Available online at: http://www.bioconductor.org/packages/release/bioc/vignettes/edgeR/inst/doc/edgeRUsersGuide.pdf (accessed May 16, 2020). Google Scholar.
19. Cheong Y. H., Chang H.-S., Gupta R., Wang X., Zhu T., Luan S. (2002). Transcriptional profiling reveals novel interactions between wounding, pathogen, abiotic stress, and hormonal responses in Arabidopsis. Plant Physiol. 129, 661–677. doi: 10.1104/pp.002857. CrossRef Google Scholar.
20. Conesa A., Götz S., García-Gómez J. M., Terol J., Talón M., Robles M. (2005). Blast2GO: a universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 21, 3674–3676. doi: 10.1093/bioinformatics/bti610. CrossRef Google Scholar.
21. Conrath U., Beckers G. J. M., Flors V., García-Agustín P., Jakab G., Mauch F., et al. (2006). Priming: getting ready for battle. Mol. Plant Microbe Interact. 19, 1062–1071. doi: 10.1094/MPMI-19-1062. CrossRef Google Scholar.
22. Crowder D. W., Northfield T. D., Strand M. R., Snyder W. E. (2010). Organic agriculture promotes evenness and natural pest control. Nature 466, 109–112. doi: 10.1038/nature09183. CrossRef Google Scholar.
23. Davis J. A., Radcliffe E. B., Ragsdale D. W. (2007). Resistance to green peach aphid, Myzus persicae (Sulzer), and potato aphid, Macrosiphum euphorbiae (Thomas), in potato cultivars. Am. J. Potato Res. 84, 259–269. doi: 10.1007/BF02986276. CrossRef Google Scholar.
24. Davis J. R., Huisman O. C., Everson D. O., Schneider A. T. (2001). Verticillium wilt of potato: a model of key factors related to disease severity and tuber yield in Southeastern Idaho. Am. J. Potato Res. 78:291. doi: 10.1007/BF02875694. CrossRef Google Scholar.
25. De Vos M., van Oosten V. R., van Poecke R. M. P., van Pelt J. A., Pozo M. J., Mueller M. J., et al. (2005). Signal signature and transcriptome changes of Arabidopsis during pathogen and insect attack. Mol. Plant Microbe Interact. 18, 923–937. doi: 10.1094/MPMI-18-0923. CrossRef Google Scholar.
26. Delessert C., Wilson I., Straeten D. V. D., Dennis E., Dolferus R. (2004). Spatial and temporal analysis of the local response to wounding. Plant Mol. Biol. 55, 165–181. doi: 10.1007/s11103-004-0112-7. CrossRef Google Scholar.
27. Donaldson J. R., Kruger E. L., Lindroth R. L. (2006). Competition- and resource-mediated tradeoffs between growth and defensive chemistry in trembling aspen (Populus tremuloides). New Phytol. 169, 561–570. doi: 10.1111/j.1469-8137.2005.01613.x. CrossRef Google Scholar.
28. Doran J. W. (2002). Soil health and global sustainability: translating science into practice. Agric. Ecosyst. Environ. 88, 119–127. doi: 10.1016/S0167-8809(01)00246-8. CrossRef Google Scholar.
29. Draper M. D., Pasche J. S., Gudmestad N. C. (2002). Factors influencing PVY development and disease expression in three potato cultivars. Am. J. Potato Res. 79, 155–165. doi: 10.1007/BF02871931. CrossRef Google Scholar.
30. Drinkwater L. E., Letourneau D. K., Workneh F., van Bruggen A. H. C., Shennan C. (1995). Fundamental differences between conventional and organic tomato in California. Ecol. Appl. 5, 1098–1112. doi: 10.2307/2269357. CrossRef Google Scholar.
31. Eigenbrode S. D., Pimentel D. (1988). Effects of manure and chemical fertilizers on insect pest populations on collards. Agric. Ecosyst. Environ. 20, 109–125. doi: 10.1016/0167-8809(88)90151-X. CrossRef Google Scholar.
32. Feeny P. (1976). Plant apparency and chemical defense, in Biochemical Interaction Between Plants and Insects, eds Wallace J. W., Mansell R. L. (Boston, MA: Springer), 1–40. doi: 10.1007/978-1-4684-2646-5_1. CrossRef Google Scholar.
33. Fineblum W. L., Rausher M. D. (1995). Tradeoff between resistance and tolerance to herbivore damage in a morning glory. Nature 377:517. doi: 10.1038/377517a0. CrossRef Google Scholar.
34. Ford K. A., Casida J. E., Chandran D., Gulevich A. G., Okrent R. A., Durkin K. A., et al. (2010). Neonicotinoid insecticides induce salicylate-associated plant defense responses. Proc. Natl. Acad. Sci. U.S.A. 107, 17527–17532. doi: 10.1073/pnas.1013020107. CrossRef Google Scholar.
35. Frost C. J., Mescher M. C., Carlson J. E., Moraes C. M. D. (2008). Plant defense priming against herbivores: getting ready for a different battle. Plant Physiol. 146, 818–824. doi: 10.1104/pp.107.113027. CrossRef Google Scholar.
36. Gavlak R., Horneck D., Miller R. O. (2003). Soil, Plant and Water Reference Methods for the Western Region, 3rd Edn. Fort Collins, CO: WCC-103 Publication. Available online at: https://www.naptprogram.org/files/napt/western-states-method-manual-2005.pdf (accessed December 21, 2019). Google Scholar.
37. Ge Y., Zhang J., Zhang L., Yang M., He J. (2008). Long-term fertilization regimes affect bacterial community structure and diversity of an agricultural soil in northern China. J. Soils Sediment. 8, 43–50. doi: 10.1065/jss2008.01.270. CrossRef Google Scholar.
38. Glynn C., Herms D. A., Egawa M., Hansen R., Mattson W. J. (2003). Effects of nutrient availability on biomass allocation as well as constitutive and rapid induced herbivore resistance in poplar. Oikos 101, 385–397. doi: 10.1034/j.1600-0706.2003.12089.x. CrossRef Google Scholar.
39. Goh C.-H., Veliz Vallejos D. F., Nicotra A. B., Mathesius U. (2013). The impact of beneficial plant-associated microbes on plant phenotypic plasticity. J. Chem. Ecol. 39, 826–839. doi: 10.1007/s10886-013-0326-8. CrossRef Google Scholar.
40. Gunapala N., Scow K. M. (1998). Dynamics of soil microbial biomass and activity in conventional and organic farming systems. Soil Biol. Biochem. 30, 805–816. doi: 10.1016/S0038-0717(97)00162-4. CrossRef Google Scholar.
41. Haney R. L., Brinton W. F., Evans E. (2008). Soil CO2 respiration: comparison of chemical titration, CO2 IRGA analysis and the solvita gel system. Renew. Agric. Food Syst. 23, 171–176. doi: 10.1017/S174217050800224X. CrossRef Google Scholar.
42. Hartley S. E., Jones C. G. (1997). Plant chemistry and herbivory, or why the World is green, in Plant Ecology, 2nd Edn, ed. Crawley M. J. (Oxford, UK: Blackwell Publishing Ltd), 284–324. Google Scholar.
43. Hole D. G., Perkins A. J., Wilson J. D., Alexander I. H., Grice P. V., Evans A. D. (2005). Does organic farming benefit biodiversity? Biol. Conserv. 122, 113–130. doi: 10.1016/j.biocon.2004.07.018. CrossRef Google Scholar.
44. Howe G. A., Jander G. (2008). Plant immunity to insect herbivores. Ann. Rev. Plant Biol. 59, 41–66. doi: 10.1146/annurev.arplant.59.032607.092825. CrossRef Google Scholar.
45. Hsu Y.-T., Shen T.-C., Hwang S.-Y. (2009). Soil fertility management and pest responses: a comparison of organic and synthetic fertilization. J. Econ. Entomol. 102, 160–169. doi: 10.1603/029.102.0123. CrossRef Google Scholar.
46. James D. G., Price T. S. (2002). Fecundity in twospotted spider mite (Acari: Tetranychidae) is increased by direct and systemic exposure to imidacloprid. J. Econ. Entomol. 95, 729–732. doi: 10.1603/0022-0493-95.4.729. CrossRef Google Scholar.
47. Jenny H. (1980). The Soil Resource: Origin and Behavior. New York, NY: Springer Verlag. Google Scholar.
48. Karthikeyan G., Doraisamy S., Rabindran R. (2009). Induction of systemic resistance in blackgram (Vigna mungo) against urdbean leaf crinkle virus by chemicals. Arch. Phytopathol. Plant Protect. 42, 1–15. doi: 10.1080/03235400600914314. CrossRef Google Scholar.
49. Kind T., Wohlgemuth G., Lee D. Y., Lu Y., Palazoglu M., Shahbaz S., et al. (2009). FiehnLib: mass spectral and retention index libraries for metabolomics based on quadrupole and time-of-flight gas chromatography/mass spectrometry. Anal. Chem. 81, 10038–10048. doi: 10.1021/ac9019522. CrossRef Google Scholar.
50. Klonsky K. (2012). Comparison of production costs and resource use for organic and conventional production systems. Am. J. Agric. Econ. 94, 314–321. doi: 10.1093/ajae/aar102. CrossRef Google Scholar.
51. Koss A. M., Jensen A. S., Schreiber A., Pike K. S., Snyder W. E. (2005). Comparison of predator and pest communities in Washington potato fields treated with broad-spectrum, selective, or organic insecticides. Environ. Entomol. 34, 87–95. doi: 10.1603/0046-225X-34.1.87. CrossRef Google Scholar.
52. Krey K. L., Blubaugh C. K., Van Leuven J. T., Snyder W. E. (2019). Organic soils control beetle survival while competitors limit aphid population growth. Environ. Entomol. 48:nvz100. doi: 10.1093/ee/nvz100. CrossRef Google Scholar.
53. Langmead B., Trapnell C., Pop M., Salzberg S. L. (2009). Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 10:R25. doi: 10.1186/gb-2009-10-3-r25. CrossRef Google Scholar.
54. Lee D. Y., Fiehn O. (2008). High quality metabolomic data for Chlamydomonas reinhardtii. Plant Methods 4:7. doi: 10.1186/1746-4811-4-7. CrossRef Google Scholar.
55. Letourneau D. K., Goldstein B. (2001). Pest damage and arthropod community structure in organic vs. conventional tomato production in California. J. Appl. Ecol. 38, 557–570. doi: 10.1046/j.1365-2664.2001.00611.x. CrossRef Google Scholar.
56. Liu B., Tu C., Hu S., Gumpertz M., Ristaino J. B. (2007). Effect of organic, sustainable, and conventional management strategies in grower fields on soil physical, chemical, and biological factors and the incidence of Southern blight. Appl. Soil Ecol. 37, 202–214. doi: 10.1016/j.apsoil.2007.06.007. CrossRef Google Scholar.
57. Lou Y., Baldwin I. T. (2004). Nitrogen supply influences herbivore-induced direct and indirect defenses and transcriptional responses in Nicotiana attenuata. Plant Physiol. 135, 496–506. doi: 10.1104/pp.104.040360. CrossRef Google Scholar.
58. Love M. I., Huber W., Anders S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15:550. doi: 10.1186/s13059-014-0550-8. CrossRef Google Scholar.
59. Lu C., Hawkesford M. J., Barraclough P. B., Poulton P. R., Wilson I. D., Barker G. L., et al. (2005). Markedly different gene expression in wheat grown with organic or inorganic fertilizer. Proc. Biol. Sci. 272, 1901–1908. doi: 10.1098/rspb.2005.3161. CrossRef Google Scholar.
60. Lugtenberg B. J. J., Chin-A-Woeng T. F. C., Bloemberg G. V. (2002). Microbe–plant interactions: principles and mechanisms. Antonie Van Leeuwenhoek 81, 373–383. doi: 10.1023/A:1020596903142. CrossRef Google Scholar.
61. Lupatini M., Korthals G. W., de Hollander M., Janssens T. K. S., Kuramae E. E. (2017). Soil microbiome is more heterogeneous in organic than in conventional farming system. Front. Microbiol. 7:2064. doi: 10.3389/fmicb.2016.02064. CrossRef Google Scholar.
62. Mäder P., Fliessbach A., Dubois D., Gunst L., Fried P., Niggli U. (2002). Soil fertility and biodiversity in organic farming. Science 296, 1694–1697. doi: 10.1126/science.1071148. CrossRef Google Scholar.
63. Mattson W. J. (1980). Herbivory in relation to plant nitrogen content. Ann. Rev. Ecol. Syst. 11, 119–161. doi: 10.1146/annurev.es.11.110180.001003. CrossRef Google Scholar.
64. McGuiness H. (1993). Living Soils: Sustainable Alternatives to Chemical Fertilizers for Developing Countries. New York, NY: Consumers Policy Institute. Google Scholar.
65. McMurdie P. J., Holmes S. (2013). phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 8:e61217. doi: 10.1371/journal.pone.0061217. CrossRef Google Scholar.
66. Mitchell C., Brennan R. M., Graham J., Karley A. J. (2016). Plant defense against herbivorous pests: exploiting resistance and tolerance traits for sustainable crop protection. Front. Plant Sci. 7:1132. doi: 10.3389/fpls.2016.01132. CrossRef Google Scholar.
67. Mithöfer A., Boland W. (2012). Plant defense against herbivores: chemical aspects. Ann. Rev. Plant Biol. 63, 431–450. doi: 10.1146/annurev-arplant-042110-103854. CrossRef Google Scholar.
68. Mittler R. (2006). Abiotic stress, the field environment and stress combination. Trends Plant Sci. 11, 15–19. doi: 10.1016/j.tplants.2005.11.002. CrossRef Google Scholar.
69. Morrissey J. P., Dow J. M., Mark G. L., O'Gara F. (2004). Are microbes at the root of a solution to world food production? EMBO Rep. 5, 922–926. doi: 10.1038/sj.embor.7400263. CrossRef Google Scholar.
70. Murray K., Jepson P., Sandlin I., Jensen A. (2019). Integrated pest management Strategic Plan for Potatoes in Oregon, Washington and Idaho. NW Potato Research Consortium Oregon State University extension catalog. EM 9275. Available online at: https://catalog.extension.oregonstate.edu/em9275 (accessed May 11, 2020). Google Scholar.
71. Novy R. G., Corsini D. L., Love S. L., Pavek J. J., Mosley A. R., James S. R., et al. (2003). Alturas: a multi-purpose, Russet potato cultivar with high yield and tuber specific gravity. Am. J. Potato Res. 80:295. doi: 10.1007/BF02854312. CrossRef Google Scholar.
72. Oksanen J., Kindt R., Legendre P., O'Hara B., Stevens M. H. H., Oksanen M. J., et al. (2007). The vegan package. Commun. Ecol. Package 10, 631–637. Available online at: www.sortie-nd.org/lme/R%20Packages/vegan.pdf. Google Scholar.
73. Pacifico D., Onofri C., Parisi B., Ostano P., Mandolino G. (2017). Influence of organic farming on the potato transcriptome. Sustainability 9:779. doi: 10.3390/su9050779. CrossRef Google Scholar.
74. Pacifico D., Paris R. (2016). Effect of organic potato farming on human and environmental health and benefits from new plant breeding techniques. Is it only a matter of public acceptance? Sustainability 8:1054. doi: 10.3390/su8101054. CrossRef Google Scholar.
75. Penman D. R., Chapman R. B. (1988). Pesticide-induced mite outbreaks: pyrethroids and spider mites. Exp. Appl. Acarol. 4, 265–276. doi: 10.1007/BF01196190. CrossRef Google Scholar.
76. Phelan P. L., Mason J. F., Stinner B. R. (1995). Soil-fertility management and host preference by European corn borer, Ostrinia nubilalis (Hübner), on Zea mays L.: a comparison of organic and conventional chemical farming. Agric. Ecosyst. Environ. 56, 1–8. doi: 10.1016/0167-8809(95)00640-0. CrossRef Google Scholar.
77. Phelan P. L., Norris K. H., Mason J. F. (1996). Soil-management history and host preference by Ostrinia nubilalis: evidence for plant mineral balance mediating insect–plant interactions. Environ. Entomol. 25, 1329–1336. doi: 10.1093/ee/25.6.1329. CrossRef Google Scholar.
78. Pimentel D., Hepperly P., Hanson J., Douds D., Seidel R. (2005). Environmental, energetic, and economic comparisons of organic and conventional farming systems. BioScience 55, 573–582. doi: 10.1641/0006-3568(2005)055[0573:EEAECO]2.0.CO;2. CrossRef Google Scholar.
79. Price P. W. (1991). The plant vigor hypothesis and herbivore attack. Oikos 62, 244–251. doi: 10.2307/3545270. CrossRef Google Scholar.
80. Prischmann D. A., James D. G., Snyder W. E. (2005). Impact of management intensity on mites (Acari: Tetranychidae, Phytoseiidae) in Southcentral Washington wine grapes. Int. J. Acarol. 31, 277–288. doi: 10.1080/01647950508684432. CrossRef Google Scholar.
81. R Core Team (2015). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. Available online at: http://www.R-project.org/ (accessed May 6, 2017). Google Scholar.
82. Ralph S., Oddy C., Cooper D., Yueh H., Jancsik S., Kolosova N., et al. (2006). Genomics of hybrid poplar (Populus trichocarpa × deltoides) interacting with forest tent caterpillars (Malacosoma disstria): normalized and full-length cDNA libraries, expressed sequence tags, and a cDNA microarray for the study of insect-induced defences in poplar. Mol. Ecol. 15, 1275–1297. doi: 10.1111/j.1365-294X.2006.02824.x. CrossRef Google Scholar.
83. Reganold J. P. (2013). Comparing organic and conventional farming systems: metrics and research approaches. Crop Manag. 12, 1–6. doi: 10.1094/CM-2013-0429-01-RS. CrossRef Google Scholar.
84. Reganold J. P., Andrews P. K., Reeve J. R., Carpenter-Boggs L., Schadt C. W., Alldredge J. R., et al. (2010). Fruit and soil quality of organic and conventional strawberry agroecosystems. PLoS ONE 5:e12346. doi: 10.1371/annotation/1eefd0a4-77af-4f48-98c3-2c5696ca9e7a. CrossRef Google Scholar.
85. Reganold J. P., Elliott L. F., Unger Y. L. (1987). Long-term effects of organic and conventional farming on soil erosion. Nature 330, 370–372. doi: 10.1038/330370a0. CrossRef Google Scholar.
86. Reganold J. P., Palmer A. S., Lockhart J. C., Macgregor A. N. (1993). Soil quality and financial performance of biodynamic and conventional farms in New Zealand. Science 260, 344–349. doi: 10.1126/science.260.5106.344. CrossRef Google Scholar.
87. Reilly K., Cullen E., Lola-Luz T., Stone D., Valverde J., Gaffney M., et al. (2013). Effect of organic, conventional and mixed cultivation practices on soil microbial community structure and nematode abundance in a cultivated onion crop. J. Sci. Food Agric. 93, 3700–3709. doi: 10.1002/jsfa.6206. CrossRef Google Scholar.
88. Reymond P., Bodenhausen N., Poecke R. M. P. V., Krishnamurthy V., Dicke M., Farmer E. E. (2004). A conserved transcript pattern in response to a specialist and a generalist herbivore. Plant Cell Online 16, 3132–3147. doi: 10.1105/tpc.104.026120. CrossRef Google Scholar.
89. Rigby D., Cáceres D. (2001). Organic farming and the sustainability of agricultural systems. Agric. Syst. 68, 21–40. doi: 10.1016/S0308-521X(00)00060-3. CrossRef Google Scholar.
90. Ritchie M. E., Phipson B., Wu D., Hu Y., Law C. W., Shi W., et al. (2015). limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 43:e47. doi: 10.1093/nar/gkv007. CrossRef Google Scholar.
91. Rizhsky L., Liang H., Mittler R. (2002). The combined effect of drought stress and heat shock on gene expression in tobacco. Plant Physiol. 130, 1143–1151. doi: 10.1104/pp.006858. CrossRef Google Scholar.
92. Rizhsky L., Liang H., Shuman J., Shulaev V., Davletova S., Mittler R. (2004). When defense pathways collide. The response of Arabidopsis to a combination of drought and heat stress. Plant Physiol. 134, 1683–1696. doi: 10.1104/pp.103.033431. CrossRef Google Scholar.
93. Robinson M. D., McCarthy D. J., Smyth G. K. (2010). edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140. doi: 10.1093/bioinformatics/btp616. CrossRef Google Scholar.
94. Rudgers J. A., Strauss S. Y., Wendel J. F. (2004). Trade-offs among anti-herbivore resistance traits: insights from Gossypieae (Malvaceae). Am. J. Botany 91, 871–880. doi: 10.3732/ajb.91.6.871. CrossRef Google Scholar.
95. Schmidt D. D., Voelckel C., Hartl M., Schmidt S., Baldwin I. T. (2005). Specificity in ecological interactions. Attack from the same lepidopteran herbivore results in species-specific transcriptional responses in two solanaceous host plants. Plant Physiol. 138, 1763–1773. doi: 10.1104/pp.105.061192. CrossRef Google Scholar.
96. Smith C. M., Rodriguez-Buey M., Karlsson J., Campbell M. M. (2004). The response of the poplar transcriptome to wounding and subsequent infection by a viral pathogen. New Phytol. 164, 123–136. doi: 10.1111/j.1469-8137.2004.01151.x. CrossRef Google Scholar.
97. Smyth G. K., Ritchie M., Thorne N., Wettenhall J., Shi W., Hu Y. (2020). Limma: Linear Models for Microarray and RNA-Seq Data User's Guide. Melbourne, VIC: Bioinformatics Division, The Walter and Eliza Hall Institute of Medical Research. Google Scholar.
98. Soffe R. J. (2002). Primrose McConnell's the Agricultural Notebook, 20th Edn. Oxford, UK: Wiley-Blackwell Science. Google Scholar.
99. Staley J. T., Stewart-Jones A., Pope T. W., Wright D. J., Leather S. R., Hadley P., et al. (2010). Varying responses of insect herbivores to altered plant chemistry under organic and conventional treatments. Proc. Biol. Sci. 277, 779–786. doi: 10.1098/rspb.2009.1631. CrossRef Google Scholar.
100. Stark J. C., Thornton M., Nolte P. (eds.) (2020). Potato Production Systems. Cham: Springer Nature. doi: 10.1007/978-3-030-39157-7. CrossRef Google Scholar.
101. Stockdale E. A., Shepherd M. A., Fortune S., Cuttle S. P. (2002). Soil fertility in organic farming systems – fundamentally different? Soil Use Manage. 18, 301–308. doi: 10.1079/SUM2002143. CrossRef Google Scholar.
102. Strauss S. Y. (1987). Direct and indirect effects of host-plant fertilization on an insect community. Ecology 68, 1670–1678. doi: 10.2307/1939859. CrossRef Google Scholar.
103. Swezey S. L., Werner M. R., Buchanan M., Allison J. (1998). Comparison of conventional and organic apple production systems during three years of conversion to organic management in coastal California. Am. J. Alternat. Agric. 13, 162–180. doi: 10.1017/S0889189300007876. CrossRef Google Scholar.
104. Szczepaniec A., Raupp M. J., Parker R. D., Kerns D., Eubanks M. D. (2013). Neonicotinoid insecticides alter induced defenses and increase susceptibility to spider mites in distantly related crop plants. PLoS ONE 8:e62620. doi: 10.1371/journal.pone.0062620. CrossRef Google Scholar.
105. Taurino M., Abelenda J. A., Río-Alvarez I., Navarro C., Vicedo B., Farmaki T., et al. (2014). Jasmonate-dependent modifications of the pectin matrix during potato development function as a defense mechanism targeted by Dickeya dadantii virulence factors. Plant J. 77, 418–429. doi: 10.1111/tpj.12393. CrossRef Google Scholar.
106. Thimm O., Bläsing O., Gibon Y., Nagel A., Meyer S., Krüger P., et al. (2004). mapman: a user-driven tool to display genomics data sets onto diagrams of metabolic pathways and other biological processes. Plant J. 37, 914–939. doi: 10.1111/j.1365-313X.2004.02016.x. CrossRef Google Scholar.
107. Thompson G. A., Goggin F. L. (2006). Transcriptomics and functional genomics of plant defence induction by phloem-feeding insects. J. Exp. Botany 57, 755–766. doi: 10.1093/jxb/erj135. CrossRef Google Scholar.
108. Trapnell C., Pachter L., Salzberg S. L. (2009). TopHat: discovering splice junctions with RNA-Seq. Bioinformatics 25, 1105–1111. doi: 10.1093/bioinformatics/btp120. CrossRef Google Scholar.
109. Triplehorn C. A. J., Borror N. F., Triplehorn D. J. C. A., Johnson N. F. (2005). Borror and DeLong's Introduction to the Study of Insects. Belmont, CA: Thomson Learning Inc. Google Scholar.
110. van Dijk J. P., Cankar K., Hendriksen P. J., Beenen H. G., Zhu M., Scheffer S., et al. (2012). The identification and interpretation of differences in the transcriptomes of organically and conventionally grown potato tubers. J. Agric. Food Chem. 60, 2090–2101. doi: 10.1021/jf204696w. CrossRef Google Scholar.
111. Voelckel C., Baldwin I. T. (2004). Herbivore-induced plant vaccination. Part II. Array-studies reveal the transience of herbivore-specific transcriptional imprints and a distinct imprint from stress combinations. Plant J. 38, 650–663. doi: 10.1111/j.1365-313X.2004.02077.x. CrossRef Google Scholar.
112. Watson C. A., Atkinson D., Gosling P., Jackson L. R., Rayns F. W. (2002). Managing soil fertility in organic farming systems. Soil Use Manag. 18, 239–247. doi: 10.1079/SUM2002131. CrossRef Google Scholar.
113. Wink M. (1988). Plant breeding: importance of plant secondary metabolites for protection against pathogens and herbivores. Theor. Appl. Genet. 75, 225–233. doi: 10.1007/BF00303957. CrossRef Google Scholar.
114. Woods J. L., Dreves A. J., Fisher G. C., James D. G., Wright L. C., Gent D. H. (2012). Population density and phenology of Tetranychus urticae (Acari: Tetranychidae) in hop is linked to the timing of sulfur applications. Environ. Entomol. 41, 621–635. doi: 10.1603/EN11279. CrossRef Google Scholar.
115. Xia J., Wishart D. S. (2016). Using metaboanalyst 3.0 for comprehensive metabolomics data analysis. Curr. Protocols Bioinformatics 55, 14–10. doi: 10.1002/cpbi.11. CrossRef Google Scholar.
116. Zangerl A. R., Bazzaz F. A. (1992). Theory and pattern in plant defense allocation, in Plant Resistance to Herbivores and Pathogens, Ecology, Evolution and Genetics, eds Fritz R. S., Simms E. L. (Chicago, IL: University of Chicago Press), 363–391. Google Scholar.
117. Zhu-Salzman K., Salzman R. A., Ahn J.-E., Koiwa H. (2004). Transcriptional regulation of sorghum defense determinants against a phloem-feeding aphid. Plant Physiol. 134, 420–431. doi: 10.1104/pp.103.028324. CrossRef Google Scholar.
118. Zvomuya F., Rosen C. J., Miller J. C. (2002). Response of Russet Norkotah clonal selections to nitrogen fertilization. Am. J. Potato Res. 79:231. doi: 10.1007/BF02986355. CrossRef Google Scholar.
Krey KL, Nabity PD, Blubaugh CK, Fu Z, Van Leuven JT, Reganold JP, Berim A, Gang DR, Jensen AS and Snyder WE (2020) Organic Farming Sharpens Plant Defenses in the Field. Front. Sustain. Food Syst. 4:97. doi: 10.3389/fsufs.2020.00097
Перевод статьи «Organic Farming Sharpens Plant Defenses in the Field» авторов Krey KL, Nabity PD, Blubaugh CK, Fu Z, Van Leuven JT, Reganold JP, Berim A, Gang DR, Jensen AS and Snyder WE., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык
Фото: Designed by Freepik





















































































































Комментарии (0)