Вопреки ожиданиям: традиционное земледелие усиливает микоризу на малоплодородных почвах
Арбускулярные микоризные (АМ) грибы имеют перспективные области применения на малоплодородных сельскохозяйственных угодьях по всему миру, однако исследования их реакции на системы традиционного и органического земледелия в условиях низко плодородных почв ограничены. Мы выдвинули гипотезу о том, что колонизационная активность и видовое разнообразие сообществ АМ грибов в системах традиционного земледелия могут быть не ниже, чем в органическом земледелии, на малоплодородных угодьях, где необходимы полезные симбиотические взаимодействия.
Аннотация
Мы собрали образцы почвы и корней кукурузы со средне- и низкоплодородных полей, использующих системы традиционного или органического земледелия, в западной части провинции Цзилинь, Китай. В обеих системах земледелия определили и сравнили процент колонизации и её интенсивность, таксономическое и филогенетическое разнообразие, состав сообществ почвенных АМ грибов, а также почвенные факторы. Интенсивность колонизации и таксономическое разнообразие операционных таксономических единиц (ОТЕ) на полях традиционного земледелия были выше, чем на органических полях. Glomus был наиболее распространённым родом на полях традиционного земледелия, тогда как Paraglomus и Glomus преобладали на органических полях. Мы также обнаружили более простую структуру сети АМ грибов с более низким филогенетическим разнообразием ОТЕ на полях традиционного земледелия. Наши результаты позволяют предположить, что, хотя система традиционного земледелия приводит к иному составу и более простой структуре сообщества почвенных АМ грибов, на традиционных полях в настоящее время присутствуют потенциально разнообразные ресурсы ОТЕ. Эта работа имеет значение для понимания влияния различных систем земледелия на почвенные АМ грибы на малоплодородных угодьях и для разработки эффективных методов производства микоризных инокулянтов.
Введение
Биология почвы является ключевым фактором, определяющим эффективность почвенных питательных ресурсов, и играет важную роль в устойчивом сельском хозяйстве (Bender и van der Heijden, 2015). Арбускулярные микоризные (АМ) грибы представляют интерес благодаря их вкладу в поглощение питательных веществ растениями (Govindarajulu и др., 2005; Davison и др., 2015), особенно в условиях дефицита питательных веществ или экологических стрессов (Akiyama и др., 2005; Begum и др., 2019). По сравнению с другими типами экосистем, сельскохозяйственные угодья обычно имеют более высокое содержание питательных веществ в почве, но всё же требуют ежегодного внесения большого количества удобрений для получения высоких урожаев. Повышенное содержание питательных веществ в почве может ослаблять взаимовыгодный симбиоз, образуемый АМ грибами с корнями растений (Scullion и др., 1998; Liu и др., 2019), и снижать интенсивность колонизации и разнообразие АМ грибов (van Geel и др., 2017; Zeng и др., 2021). Поэтому в ряде предыдущих исследований было показано, что АМ грибы приносят ограниченную пользу для сельскохозяйственного производства, особенно на высокопродуктивных угодьях (Ryan и Graham, 2002, 2018). Однако АМ грибы всё еще потенциально важны для обширных площадей низкокачественных сельскохозяйственных земель, широко распространенных по всему миру. Неблагоприятные экологические условия, такие как засуха или засоление, часто встречаются на низкокачественных землях (Bi и др., 2018), тогда как мутуалистический симбиоз может усиливать рост растений за счет улучшения использования почвенных питательных веществ и воды (Begum и др., 2019). Поскольку АМ грибы имеют перспективные применения на малоплодородных угодьях, исследования в условиях низкоплодородных почв, например, влияния систем традиционного и органического земледелия на сообщества АМ грибов, должны привлекать больше внимания (Sale и др., 2015; Roy и др., 2017).
Традиционное сельское хозяйство — это система земледелия, широко применяемая во всем мире, которая использует большие количества химических удобрений, биоцидов, высокоурожайные сорта сельскохозяйственных культур, обработку почвы и управляемые оросительные системы для максимизации урожайности. На практике норма внесения химических удобрений достигла, а иногда и превысила потребности роста культур (Tang и др., 2008). Длительное применение химических удобрений может снижать плотность спор (Wu и др., 2011; Lin и др., 2020), видовое богатство (Sheng и др., 2013), разнообразие (Oehl и др., 2004) и изменять структуру сообщества АМ грибов в почвах сельскохозяйственных угодий (He и др., 2016). Напротив, органическое земледелие с использованием органических мелиорантов, таких как навоз животных, быстро развивалось в последние несколько десятилетий из-за растущих проблем, связанных с повреждением химическими удобрениями почвенных организмов, нарушениями питательного режима и загрязнением окружающей среды (Yue и др., 2016). Высокое содержание органического углерода в органических мелиорантах может стимулировать пролиферацию мицелия АМ грибов (Hart и Reader, 2002), тем самым увеличивая инокуляционную способность АМ грибов на корнях растений. Поэтому в некоторых исследованиях было обнаружено, что колонизационная активность (Qin и др., 2015; Jiang Y. J. и др., 2020) и разнообразие АМ грибов (Bhadalung и др., 2005; Lin и др., 2012) были выше на органических фермах, чем на традиционных. Однако мы предполагаем, что эти эффекты систем традиционного и органического земледелия на почвенные АМ грибы могут быть иными на малоплодородных угодьях по следующим причинам.
Во-первых, концентрации питательных веществ и скорость их высвобождения у химических удобрений, вносимых в системах традиционного земледелия, отличаются от таковых у органических мелиорантов, применяемых в органическом земледелии (Niedziński и др., 2021). При внесении химических удобрений в почву АМ грибы могут подавляться, поскольку содержание питательных веществ в окружающей среде повышается за счет быстрого высвобождения высокой концентрации питательных веществ (Xiang и др., 2016). Повышенное содержание питательных веществ в почве в ответ на удобрение также снижает микоризную зависимость сельскохозяйственных культур. Однако когда содержание почвенных питательных веществ на малоплодородных угодьях возвращается к исходному уровню из-за поглощения растениями или потерь вследствие выщелачивания, симбиоз растения и АМ грибов может вновь широко устанавливаться на традиционных фермах. В отличие от химических удобрений, относительно низкие концентрации питательных веществ и медленная скорость высвобождения органических мелиорантов, применяемых на органических фермах, по-видимому, не ингибируют инокуляцию корней растений АМ грибами. Однако не следует упускать из виду негативные последствия продолжительного высвобождения питательных веществ для симбиоза.
Во-вторых, поскольку химические удобрения содержат практически только одну форму питательного вещества, например, одинаковую химическую форму N, P или K, независимо от марки и производителя, они создают стабильный экологический фильтр для почвенных организмов из года в год. Поэтому широко распространено мнение, что разнообразие АМ грибов на традиционных фермах впоследствии снижается по мере отфильтровывания некоторых видов, родов или семейств (Lin и др., 2012). Однако благодаря полиморфизму рибосомальной ДНК (гетерокариозис) в отдельных спорах АМ грибов (Pawlowska и Taylor, 2004) было обнаружено высокое генетическое разнообразие в спорах и экстраматрикальных гифах (Bever и Wang, 2005; Tisserant и др., 2013). Это свидетельствует о том, что даже если разнообразие семейств, родов или видов снижается, разнообразие более мелких таксономических единиц, таких как операционные таксономические единицы (ОТЕ), может и не снижаться (Rosendahl, 2008). Недавние исследования с использованием методов высокопроизводительного секвенирования также подтвердили высокое разнообразие ОТЕ в сообществах АМ грибов в почве традиционных ферм (Chen и др., 2014).
Основываясь на этих двух причинах, мы выдвинули гипотезу о том, что колонизационная активность и разнообразие ОТЕ сообщества АМ грибов в системах традиционного земледелия не обязательно будут ниже, чем в органическом земледелии, особенно на малоплодородных угодьях, где необходим полезный симбиоз. Чтобы проверить нашу гипотезу, мы собрали образцы почвы и корней кукурузы на малоплодородных (средне- и низкоплодородные земли Китая, подробности см. ниже) кукурузных полях в западной части провинции Цзилинь, Китай, при традиционном и органическом управлении соответственно. Во-первых, мы оценили, была ли активность почвенных АМ грибов (процент и интенсивность колонизации корней и плотность спор в почве) ниже на традиционных фермах, чем на органических, как принято считать. Затем были сравнены родовой состав, таксономические и филогенетические индексы разнообразия ОТЕ сообществ АМ грибов, чтобы определить, снижают ли системы традиционного земледелия разнообразие АМ грибов на малоплодородных угодьях. Состав сообщества также является основным фактором, влияющим на функцию сообщества. Поэтому мы создали сети ОТЕ сообществ АМ грибов и проанализировали их с помощью структурных параметров, чтобы определить состав сообществ АМ грибов на традиционных и органических фермах и наличие значительных различий в их композиционных характеристиках. Наконец, мы проанализировали, связаны ли активность, разнообразие и состав сообщества АМ грибов с почвенными факторами и относительное влияние каждого фактора. Это исследование способствует пониманию влияния систем традиционного и органического земледелия на почвенные АМ грибы на малоплодородных угодьях и стимулирует рациональное применение микоризных удобрений в агроэкосистемах для восстановления и поддержания экологических функций почв сельскохозяйственных угодий.
Материалы и методы
Район отбора проб и история возделывания
Провинция Цзилинь является крупным производителем кукурузы в Китае и относится к умеренному кукурузному поясу мира. Это исследование было проведено в полузасушливой равнине западной части провинции Цзилинь (Рисунок 1). Этот район имеет умеренный континентальный муссонный климат с сухими веснами, среднегодовой температурой 4–6°C и среднегодовым количеством осадков 350–500 мм, при этом осадки выпадают преимущественно в июле и августе (Li и др., 2005). Почва представляет собой супесчаный суглинок (Typic Hapludolls) с высокой засоленностью и низким содержанием питательных веществ (Zhu, 1994; Yang и др., 2014). Согласно критериям градации питательности почв второй всекитайской почвенной переписи (Shi и др., 2004; Yang и др., 2014), классы плодородия почв на наших участках обследования были средне- и низкоплодородными. После 2 лет полевых выездов с 2019 по 2020 год пятнадцать участков в шести уездах были окончательно определены как районы отбора проб для этого исследования (Дополнительная таблица 1). На каждом участке были как традиционные, так и органические фермы, засаженные кукурузой, то есть большие площади традиционных полей и небольшие участки органических кукурузных полей. Пара участков отбора проб — одно традиционное (Con) кукурузное поле и одно органическое (Org) кукурузное поле — были случайным образом выбраны на каждом участке, вдали от дороги. Таким образом, в это исследование было включено в общей сложности 15 участков с обеими системами земледелия, всего 30 районов отбора проб.
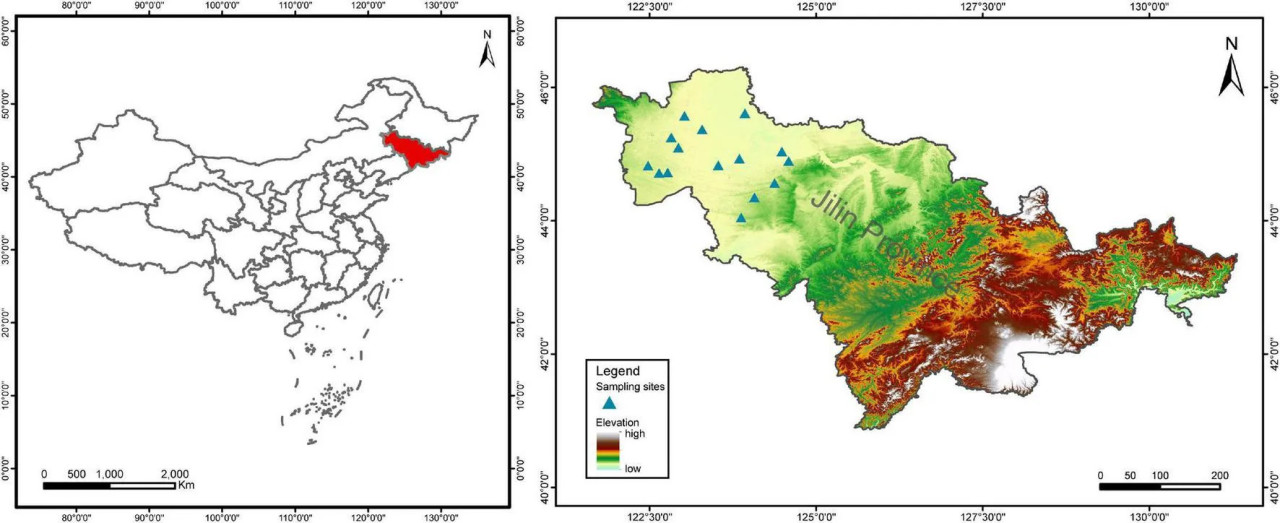
РИСУНОК 1 Распределение исследованных участков в западной части провинции Цзилинь, Китай.
Все исследуемые нами традиционные фермы находятся в системе непрерывного монокультурного возделывания кукурузы. За последние 4 года традиционная обработка почвы (ротационная вспашка и удаление пожнивных остатков) в районе исследования постепенно заменялась ресурсосберегающей обработкой (гребневая вспашка и сохранение остатков) по рекомендации правительства (Министерство сельского хозяйства и сельских дел Китая, 2020). На традиционных фермах ежегодно применялись химические комплексные удобрения и гербициды. Удобрения, используемые в органическом земледелии, как правило, основаны на навозе животных, и гербициды не применялись. Сорняки на органических фермах контролировались вручную в начале вегетационного сезона, а затем подавлялись пологом кукурузы. Органические фермы находились в системе органического земледелия не менее 5 лет, а некоторые из них — более 30 лет (Дополнительная таблица 1). Согласно нашему полевому обследованию, сорта кукурузы, высаживаемые на традиционных и органических фермах, различались не только от места к месту, но и от года к году. В основном, тип кукурузы на органических фермах — это восковидная кукуруза, в то время как зубовидная кукуруза распространена на традиционных фермах в нашем районе отбора проб.
Сбор образцов
Все образцы почвы и корней с кукурузных полей были собраны в конце августа 2020 года (репродуктивная стадия кукурузы). На каждом поле отбора проб было случайным образом выбрано три точки отбора проб с расстоянием около 5–10 м между каждой точкой. После удаления примерно 2 см верхнего слоя почвы на горизонтальном расстоянии около 15 см от стебля кукурузы выкапывался объем почвы 20 см × 20 см × 30 см, и извлекались мелкие корни кукурузы. Почва из трех точек отбора проб тщательно перемешивалась, затем случайным образом отбиралось около 2 кг свежей почвы и помещалось в пластиковые герметичные пакеты (образец почвы I). Около 10 г свежей почвы случайным образом отбиралось в пластиковые герметичные пакеты и помещалось в холодильник с льдом (образец почвы II). Мелкие корни кукурузы из трех точек отбора проб смешивались и помещались в пластиковые бутылки с фиксатором формалин-уксусная кислота-спирт (FAA). Затем все образцы почвы и корней были доставлены обратно в лабораторию. Примеси, такие как растительные и животные остатки, были удалены из образца почвы I, и он был высушен на воздухе в помещении для измерения плотности спор АМ грибов и почвенных параметров. Образец почвы II хранился в холодильнике при −80°C в течение 3 дней, а затем был отправлен в Majorbio Co., Ltd. для выделения ДНК и секвенирования АМ грибов. Корни, фиксированные в FAA, использовались для измерения процента и интенсивности колонизации АМ грибов.
Измерение почвенных параметров
Образец почвы I был высушен на воздухе и просеян через сито с размером ячеек 40 меш (<0,425 мм). pH почвы измеряли в почвенно-водной суспензии (1:5, вес/об) с использованием pH-метра (Mettler-Toledo pH reader, Швейцария). Для определения содержания органического углерода в почве использовался метод дихромата калия (Lu, 1999). Общее содержание азота в почве определялось методом Кьельдаля (Lu, 1999). Гидролизуемый азот почвы измерялся методом щелочного гидролиза-диффузии (Lu, 1999). Содержание общего и доступного фосфора определялось спектрофотометрически с помощью проточного анализатора (SAN + +, Skalar, Бреда, Голландия).
Определение колонизации арбускулярными микоризными грибами и плотности спор
Корни, фиксированные в FAA, промывали 10% раствором KOH при 90°C в течение 60 мин, подкисляли 1% HCl в течение 30 мин, а затем нарезали на сегменты корней длиной примерно 1 см. Сегменты корней выдерживали в 0,05% (об.) растворе трипанового синего, а затем помещали на горячую плиту при 90°C на 30 мин (Koske и Gemma, 1989). Окрашенные корни помещали на предметные стекла для оценки колонизации АМ грибами в соответствии с методом Trouvelot и др. (1986). Затем были рассчитаны процент колонизации (частота микоризы в корневой системе) и интенсивность колонизации (интенсивность микоризной колонизации в корневой системе) (Trouvelot и др., 1986).
Споры АМ грибов выделяли из 20 г случайно разделенного высушенного образца почвы I методом мокрого просеивания и градиентного центрифугирования (Gerdemann и Nicolson, 1963). Плотность спор (общее количество спор в 20 г сухой почвы) определяли по количеству спор, подсчитанных под стереомикроскопом (Daniels и Skipper, 1982). Для подсчета отбирались только споры с полной структурой формы.
Секвенирование ДНК
Выделение ДНК и ампликонное секвенирование
Общая ДНК из 0,5 г образцов почвы была выделена в соответствии с инструкциями производителя с использованием набора E.Z.N.A.® soil (Omega Bio-Tek, Норкросс, Джорджия, США). Праймеры AML1F/AML2R использовались для амплификации гипервариабельных областей V4-V5 гена 18S рРНК грибов в системе ПЦР-термоциклера (Axygen Biosciences, Юнион-Сити, Калифорния, США). Вторая амплификация проводилась с использованием специфичного для АМ грибов набора праймеров AMV4.5NF/AMDGR. ПЦР-реакции проводились по следующей программе: 3 мин денатурации при 95°C, 27 циклов по 30 с при 95°C, 30 с для отжига при 55°C и 45 с для элонгации при 72°C, и финальная элонгация при 72°C в течение 10 мин. ПЦР-реакции выполнялись в трех повторностях в 20 мкл смеси, содержащей 4 мкл 5 × FastPfu Buffer, 2 мкл 2,5 мM dNTPs, 0,8 мкл каждого праймера (5 мкM), 0,4 мкл FastPfu Polymerase и 10 нг матричной ДНК.
Биоинформатика
Сырые FASTQ-файлы были демультиплексированы, прошли фильтрацию качества с помощью Trimmomatic и объединены с помощью FLASH со следующими критериями: (i) чтения усекались в любом месте, где средний балл качества за 50 п.н. скользящее окно был < 20. (ii) Праймеры точно сопоставлялись с допуском 2 несовпадающих нуклеотидов, и чтения, содержащие неоднозначные основания, удалялись. (iii) Последовательности, перекрытие которых превышало 10 п.н., были объединены в соответствии с их последовательностью перекрытия. Операционные таксономические единицы (ОТЕ) были кластеризованы с порогом сходства 97% с использованием UPARSE (версия 7.11), а химерные последовательности были идентифицированы и удалены с использованием UCHIME. Таксономическая идентичность репрезентативных последовательностей была проверена по онлайн-базе данных АМ грибов Maarjam081 (2 Accession для цитирования этих данных SRA: PRJNA819886).
Анализ данных
Были рассчитаны четыре индекса разнообразия ОТЕ сообществ АМ грибов, включая индекс Chao1, индекс разнообразия Шеннона, индекс выравненности Симпсона и индекс филогенетического разнообразия (Caporaso и др., 2010). Традиционные и органические фермы являются сопряженными парами образцов, поэтому для анализа различий в почвенных параметрах, микоризной колонизации, плотности спор и индексах разнообразия АМ грибов между системами традиционного и органического земледелия были проведены парные t-тесты с использованием программного обеспечения SPSS 25.0 (IBM). Нормальность и однородность распределения остатков были проверены, при необходимости проводились логарифмические или арксинусные преобразования.
Для выявления различий в структуре сообщества почвенных АМ грибов между традиционными и органическими кукурузными фермами было проведено неметрическое многомерное шкалирование (NMDS) с использованием пакета R "vegan" на основе расстояний Брея-Кертиса матрицы композиционных данных. Различия в расстоянии Брея-Кертиса между двумя системами земледелия также были проверены на значимость с помощью теста перестановок Монте-Карло.
С использованием пакета "SpiecEasi" в программном обеспечении R были отдельно построены карты сетей ОТЕ почвенных АМ грибов с традиционных и органических кукурузных полей. Было рассчитано несколько параметров, характеризующих сети. ОТЕ с относительной численностью > 0,05% были отобраны для корреляционного анализа Спарка, и корреляционные сети были построены путем отбора значимых (P < 0,01) и сильных (| r| > 0,3) корреляций (Kim и др., 2020). Визуализация сетей была выполнена на платформе Gephi (v.0.9.2)3 и раскрашена по классификации семейств.
Для изучения того, связаны ли колонизация АМ грибов, плотность спор и индексы разнообразия с почвенными факторами, в двух системах земледелия были построены корреляционные тепловые карты на основе коэффициентов корреляции Спирмена между пятью почвенными факторами (pH, содержание органического вещества, общий и доступный азот, общий и доступный фосфор) и вышеуказанными показателями. С использованием пакета "gbmplus" (R v.3.6.3) с 500 деревьями для бустинга (De'ath, 2007) был проведен анализ агрегированных бустинг-деревьев (ABT) для количественной оценки влияния пяти почвенных факторов на состав ОТЕ сообществ АМ грибов.
Результаты
Инокуляция корней и плотность спор
Процент колонизации корней растений варьировал незначительно (86,67–100%), но интенсивность колонизации (1,86–44,5%) и плотность спор (25–103 спор/20 г почвы) сильно варьировали по всем районам отбора проб, включая традиционные и органические фермы (Рисунок 2). Процент колонизации (t = 1,733, P = 0,105) и плотность спор (t = −0,822, P = 0,425) не различались между двумя системами земледелия (Рисунки 2A,C), но интенсивность колонизации различалась (t = 2,159, P = 0,049, Рисунок 2B).
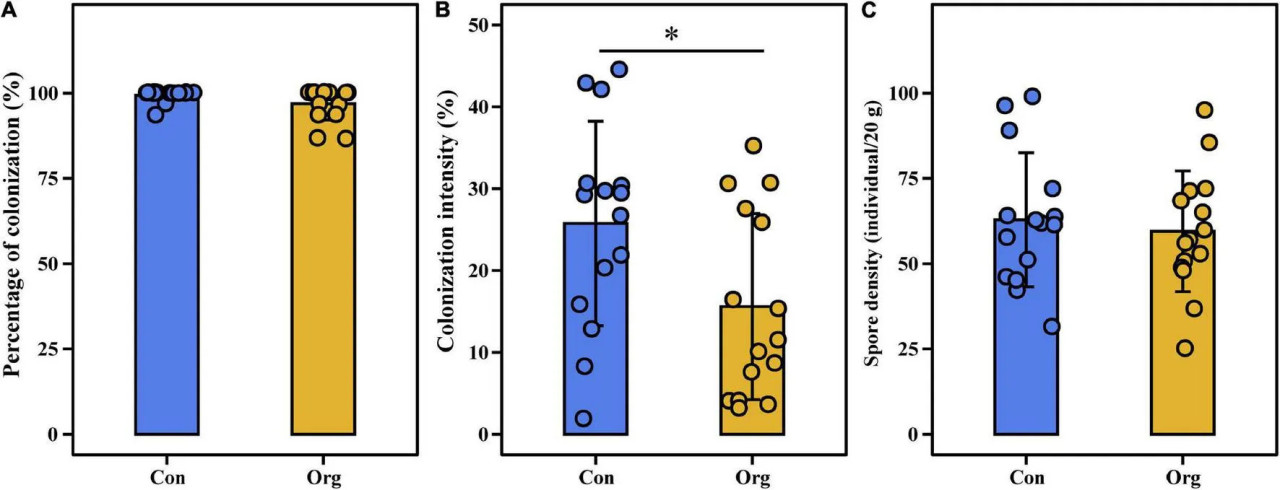
РИСУНОК 2 Процент колонизации (A), интенсивность колонизации (B) корней и плотность спор в почве (C) на традиционных (Con) и органических (Org) кукурузных фермах. означает, что различие между двумя системами земледелия значимо на уровне P = 0,05.
Разнообразие сообщества арбускулярных микоризных грибов
Всего во всех образцах было обнаружено 171 ОТЕ АМ грибов, из которых 107 и 125 ОТЕ были обнаружены на традиционных и органических фермах соответственно. Разнообразие сообщества почвенных АМ грибов различалось между двумя системами земледелия. Индекс разнообразия Шеннона и индекс Chao1 ОТЕ сообществ АМ грибов были выше на традиционных фермах, чем на органических (Рисунки 3A,C), в то время как различие в индексе выравненности Симпсона не было значимым (Рисунок 3B). Индекс филогенетического разнообразия (PD) ОТЕ был ниже для сообществ АМ грибов на традиционных фермах, чем на органических (Рисунок 3D).
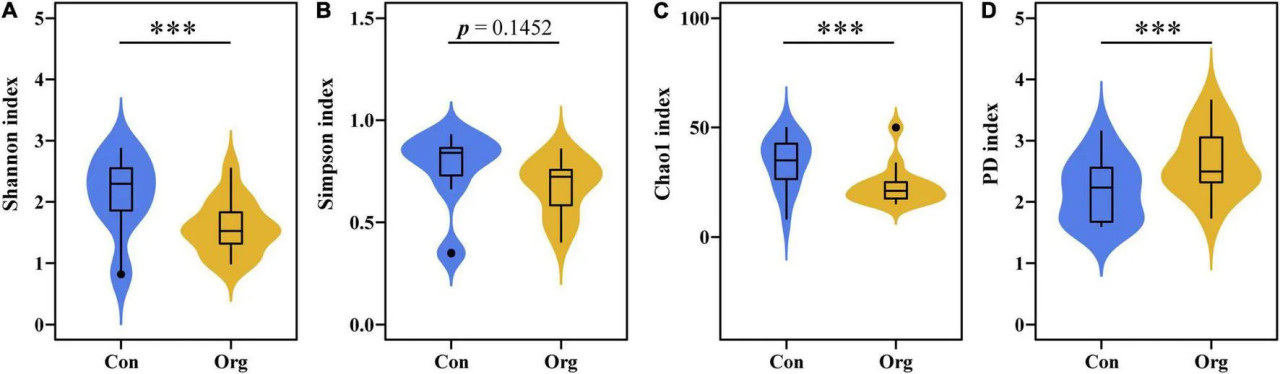
РИСУНОК 3 Индексы разнообразия ОТЕ сообществ АМ грибов на традиционных (Con) и органических (Org) кукурузных фермах, включая индекс разнообразия Шеннона (A), индекс выравненности Симпсона (B), индекс Chao 1 (C) и индекс филогенетического разнообразия (D). означает, что различие между двумя системами земледелия значимо на уровне P = 0,001.
Состав сообщества арбускулярных микоризных грибов и сетевой анализ
Для кукурузных полей в системе традиционного земледелия 64,79% ОТЕ АМ грибов принадлежали к семейству Glomeraceae (включая 41,8% Glomus, 14,08% Septoglomus, 8,68% Funneliformis и 0,23% Kamienskia), 13,88% — к Paraglomeraceae (Paraglomus), 9,7% — к Claroideoglomeraceae (Claroideoglomus) и 5,45% — к Diversisporaceae (Diversispora). Другие семейства, такие как Archaeosporaceae (Archaeospora), Geosiphonaceae (Geosiphon), Gigasporaceae (Scutellospora), Ambisporaceae (Ambispora) и Diversisporaceae (Redeckera), составляли менее 10% от общего числа ОТЕ (Рисунок 4).
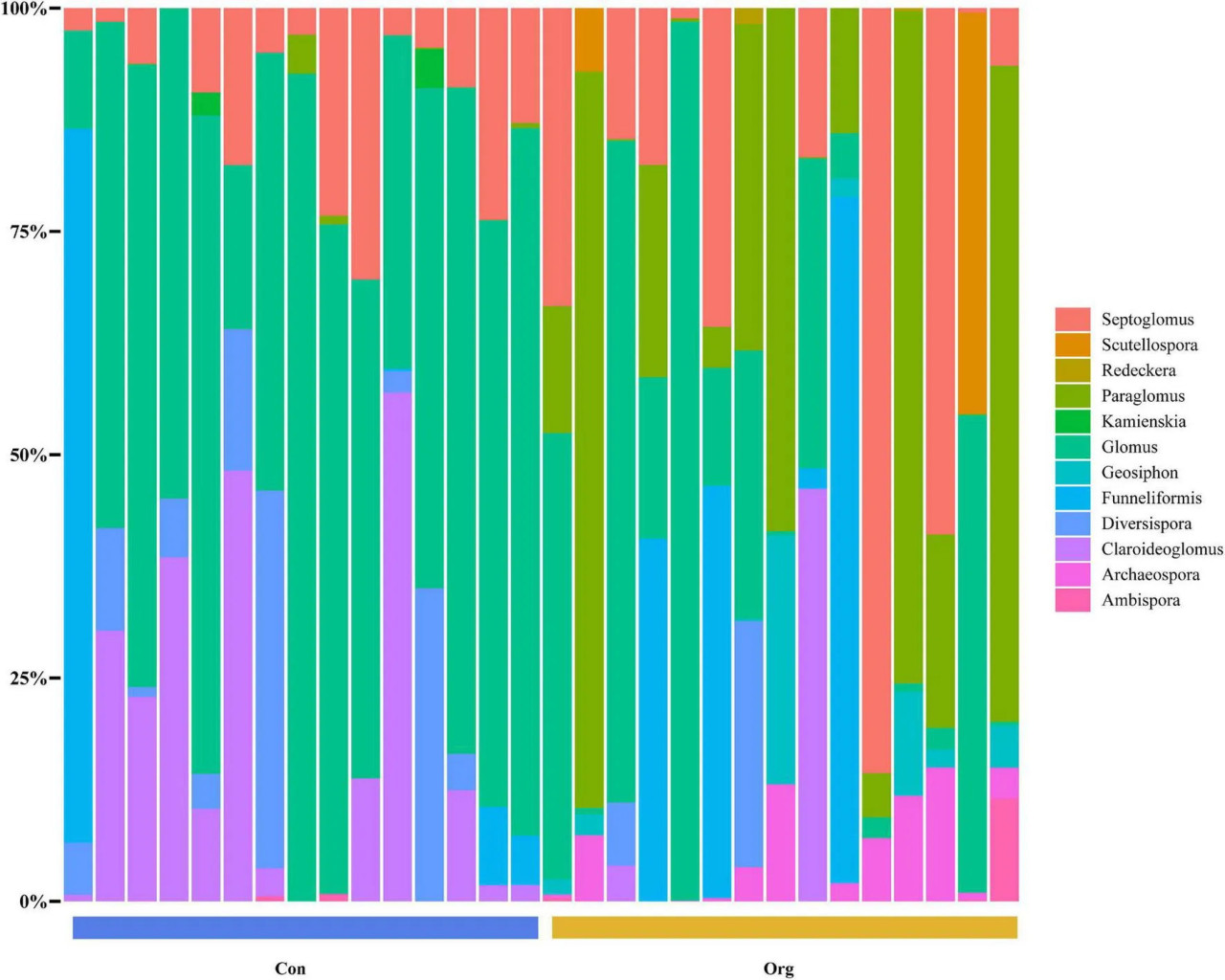
РИСУНОК 4 Родовой состав сообществ почвенных АМ грибов на традиционных (Con) и органических (Org) кукурузных фермах.
Для кукурузных полей в системе органического земледелия 54,73% ОТЕ АМ грибов принадлежали к семейству Glomeraceae (включая 25,63% Glomus, 18,04% Septoglomus, 11,06% Funneliformis), 27,34% — к Paraglomeraceae (Paraglomus). Другие семейства (каждое менее 5%), включая Ambisporaceae (Ambispora), Archaeosporaceae (Archaeospora), Claroideoglomeraceae (Claroideoglomus), Diversisporaceae (Diversispora, Redeckera), Geosiphonaceae (Geosiphon) и Gigasporaceae (Scutellospora), составляли в сумме около 18% (Рисунок 4).
На основе результатов анализа неметрического многомерного шкалирования расстояние Брея-Кертиса различалось между двумя системами земледелия, что указывает на то, что состав ОТЕ сообщества почвенных АМ грибов различался между традиционными и органическими фермами (Рисунок 5).
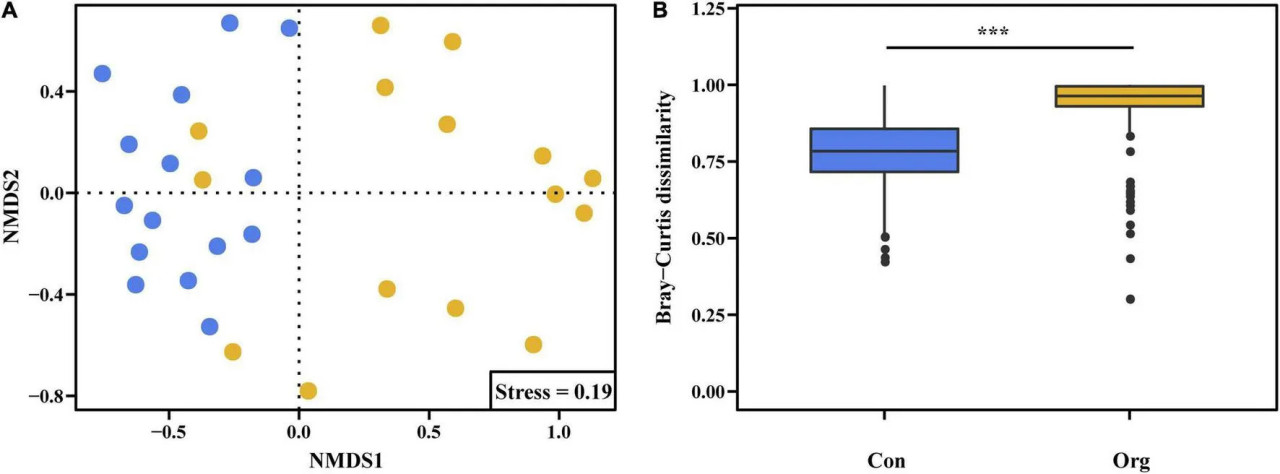
РИСУНОК 5 Ординации неметрического многомерного шкалирования (A) сообществ АМ грибов на традиционных (синий) и органических (желтый) фермах на основе их состава ОТЕ, полученного из расстояний Брея-Кертиса (B). означает, что различие между двумя системами земледелия значимо на уровне P = 0,001.
Мы обнаружили, что сеть почвенных АМ грибов как на традиционных (модульность 0,814), так и на органических фермах (модульность 0,724) имела модульную структуру (>0,4) (Таблица 1). Узловые ОТЕ с высокой связностью принадлежали преимущественно к семействам Glomeraceae и Diversisporaceae в системе традиционного земледелия, тогда как в системе органического земледелия — к семействам Glomeraceae и Paraglomeraceae (Рисунок 6). Более того, ключевой таксон семейства Glomeraceae включал полностью разные компоненты ОТЕ на традиционных и органических фермах соответственно.
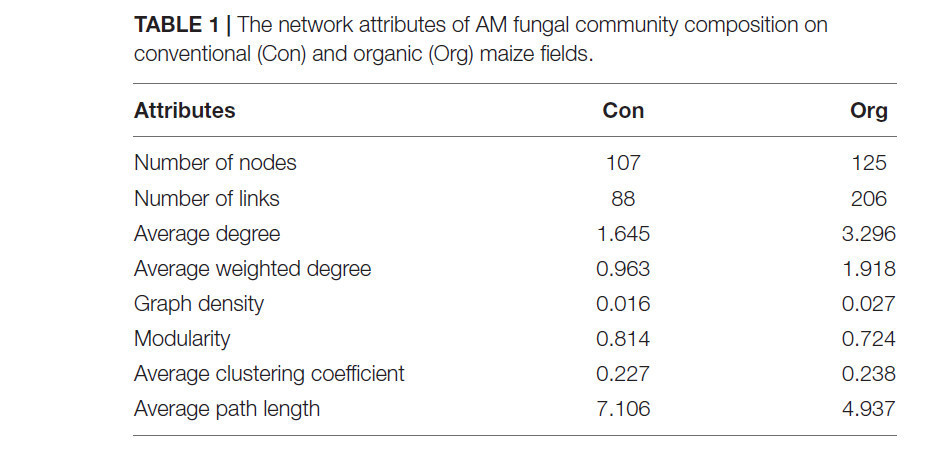
ТАБЛИЦА 1 Атрибуты сети состава сообщества АМ грибов на традиционных (Con) и органических (Org) кукурузных полях.
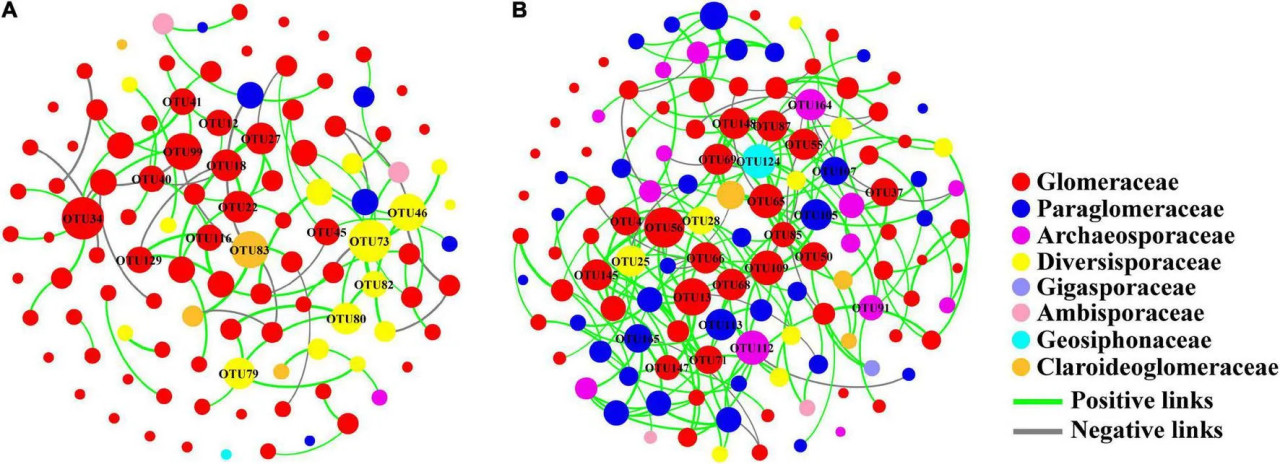
РИСУНОК 6 Схема сети сообщества АМ грибов на традиционных (A) и органических (B) кукурузных фермах. Размер каждого узла пропорционален количеству связей (степени).
Сеть почвенных АМ грибов органических ферм была более сложной, поскольку имела больше узлов и связей, более высокую среднюю связанность, плотность графа и средние коэффициенты кластеризации (Таблица 1 и Рисунок 6). Некоторые отрицательные связи между узлами появлялись в сети традиционных ферм (21,59% от общего числа связей) и органических ферм (12,14% от общего числа связей).
Почвенные факторы
За исключением pH почвы, содержание органического вещества, общего азота, общего фосфора, доступного азота и фосфора в почве было выше на органических фермах (Дополнительная таблица 2). В частности, разница в содержании доступного фосфора в почве между двумя системами земледелия была примерно восьмикратной.
Корреляции между почвенными факторами и почвенными АМ грибами (колонизация, плотность спор и разнообразие) различались в двух системах земледелия (Рисунок 7). В целом, почвенные факторы коррелировали с разнообразием АМ грибов на традиционных фермах, тогда как на органических фермах они едва ли были значимы. На традиционных фермах все четыре индекса разнообразия почвенных АМ грибов увеличивались с повышением содержания питательных веществ в почве (особенно фосфора) и снижением pH почвы (Рисунок 7A). Только индекс филогенетического разнообразия отрицательно коррелировал с pH почвы на органических фермах (Рисунок 7B). Во всех системах земледелия ни процент, ни интенсивность колонизации корней не коррелировали с почвенными факторами. На органических фермах плотность спор отрицательно коррелировала с общим и доступным содержанием фосфора в почве (Рисунок 7B).
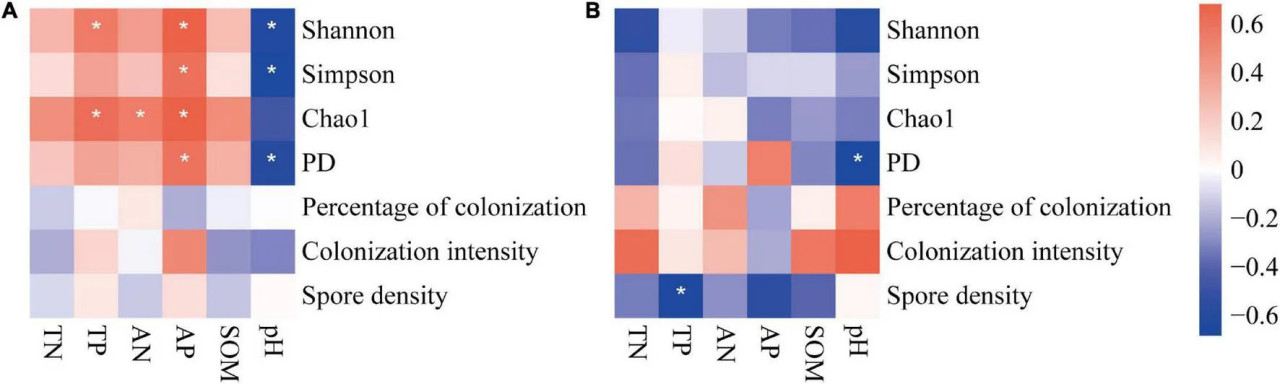
РИСУНОК 7 Корреляция между почвенными факторами и индексами сообществ АМ грибов на традиционных (A) и органических (B) кукурузных фермах. , и означают, что корреляция значима на уровне P = 0,05, 0,01 и 0,001 соответственно.
Как на традиционных, так и на органических фермах, общее содержание фосфора в почве и содержание доступного фосфора были наиболее важными факторами, влияющими на состав сообщества почвенных АМ грибов (Рисунок 8). Помимо фосфора, общее содержание азота в почве на традиционных фермах и pH почвы на органических фермах также влияли на состав сообщества АМ грибов на кукурузных полях (Рисунок 8).
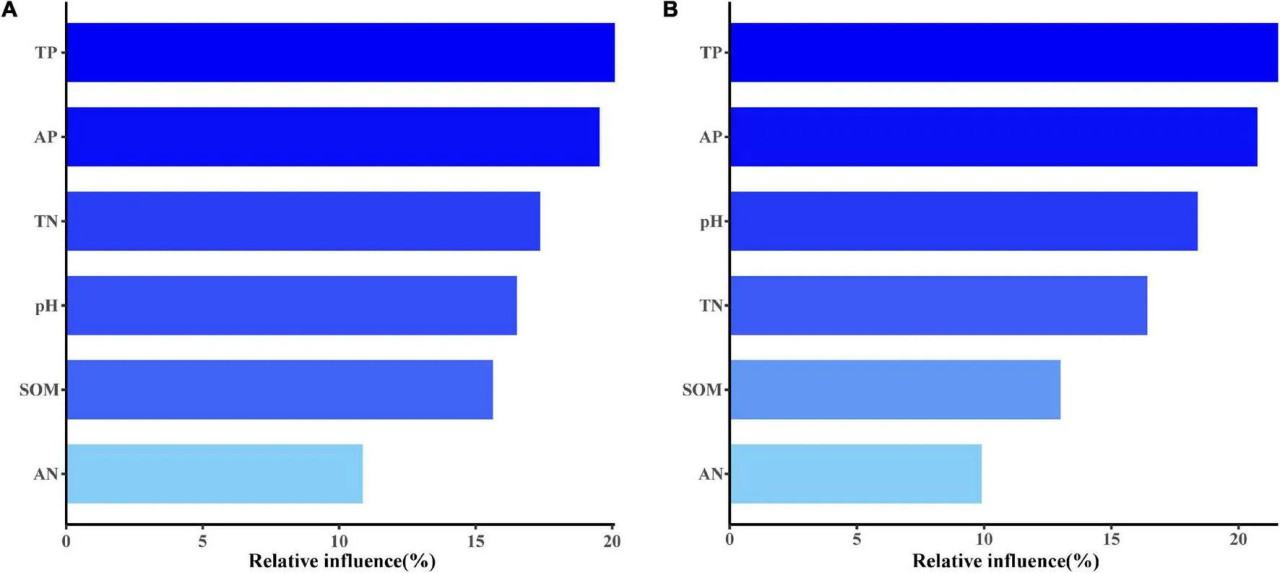
РИСУНОК 8 Анализ агрегированных бустинг-деревьев (ABT), показывающий относительное влияние каждого почвенного фактора на состав сообщества АМ грибов на традиционных (A) и органических (B) кукурузных фермах.
Обсуждение
Более высокая колонизационная активность арбускулярных микоризных грибов на традиционных фермах
Существует общепринятое мнение, что системы традиционного земледелия ингибируют колонизацию АМ грибами, тогда как системы органического земледелия усиливают колонизацию корней растений АМ грибами (Cely и др., 2016; Jiang Y. J. и др., 2020). Однако в настоящем исследовании мы обнаружили противоположный результат: интенсивность колонизации была выше на традиционных фермах (Рисунок 2B). Более того, во многих предыдущих исследованиях было подтверждено, что почвенные питательные вещества являются основным фактором, влияющим на колонизацию и размножение АМ грибов (Johnson, 2010; Begum и др., 2019). Но наше исследование показало, что, хотя содержание питательных веществ в почве было выше на органических фермах, чем на традиционных (Дополнительная таблица 2), процент и интенсивность колонизации и плотность спор не зависели почти от всех измеренных нами почвенных факторов (Рисунок 7). Различные результаты, полученные в этом исследовании, могут быть обусловлены адаптацией таксонов АМ грибов к уровню питательных веществ в низкокачественной почве данного региона.
Согласно критериям градации питательности почв второй всекитайской почвенной переписи (Zhu, 1994; Shi и др., 2004), все классы плодородия почв в этом районе были средне- и низкоплодородными. Независимо от того, являются ли вносимые питательные вещества химическими или органическими удобрениями, содержание питательных веществ в почве может увеличиваться только в течение определенного периода и в ограниченном почвенном пространстве (Cavagnaro и др., 2015; Yang и др., 2017). В этом исследовании почти все корни, отобранные на традиционных и органических фермах, были колонизированы АМ грибами (Рисунок 2A), что предполагает, что сельскохозяйственные растения могут полагаться на сотрудничество с АМ грибами для получения дополнительных ресурсов. Это согласуется с представлением о том, что симбиоз растений и АМ грибов легко устанавливается в условиях низкого содержания питательных веществ (Graham и Eissenstat, 1994; Lekberg и др., 2021). Кроме того, содержание питательных веществ в почвах традиционных полей с большей вероятностью возвращается к исходному низкокачественному уровню из-за поглощения культурами питательных веществ, быстро высвобождающихся из химических удобрений, или потерь питательных веществ вследствие выщелачивания (Bender и van der Heijden, 2015). Содержание питательных веществ в почвах традиционных полей в конце вегетационного сезона (конец августа) было ниже, чем в почвах органических полей, что могло способствовать повышению интенсивности колонизации на традиционных фермах в этом исследовании.
Таким образом, фоновый уровень плодородия почвы, а также другие экологические факторы (например, засоление, засуха и т.д.) должны учитываться при оценке влияния систем традиционного и органического земледелия на почвенные АМ грибы (Franke-Snyder и др., 2001; Boomsma и Vyn, 2008). Поскольку их сильное воздействие на АМ грибы может помешать нашему правильному пониманию полезных и вредных эффектов различных систем земледелия на экологические функции АМ грибов и/или других почвенных организмов.
Более низкое филогенетическое, но более высокое таксономическое разнообразие операционных таксономических единиц арбускулярных микоризных грибов на традиционных фермах
Системы традиционного и органического земледелия не только по-разному влияют на установление симбиоза растения и АМ грибов, но и приводят к различному разнообразию почвенных грибных сообществ (Sale и др., 2015; Roy и др., 2017). Общепризнано, что разнообразие сообществ почвенных АМ грибов на уровне семейств или родов часто снижается, когда сельскохозяйственные угодья подвергаются воздействию высоких уровней химических удобрений, гербицидов и механизированной обработки почвы при традиционном управлении (Zeng и др., 2021). Более того, наиболее распространенным таксоном сообщества АМ грибов в таких системах земледелия, как правило, является Glomus — более устойчивый к нарушениям и широко распространенный род (Daniell и др., 2001; Hart и Reader, 2002). Наше исследование согласуется с этим (Рисунок 4). Однако, хотя на традиционных фермах было обнаружено меньше семейств и родов, их разнообразие ОТЕ было выше, чем на органических фермах (Рисунки 3A,C).
Плодородие почвы было низким и средним как на традиционных, так и на органических фермах (Zhu, 1994; Shi и др., 2004), но уровни питательных веществ в почве были выше на органических фермах, чем на традиционных, особенно в отношении содержания доступного фосфора (Дополнительная таблица 2). Навоз животных является основным компонентом органического удобрения в этом регионе и содержит высокое содержание фосфора (Liu и др., 2019). Содержание фосфора является важным фактором в установлении симбиоза и разнообразия АМ грибов (Xiao и др., 2019). Обычно увеличение разнообразия АМ грибов с повышением содержания фосфора в почве происходит в малоплодородных почвах (Johnson и др., 2003), до достижения порогового уровня, за пределами которого фосфор отрицательно влияет на АМ грибы (Kiers и др., 2011; He и др., 2016). Относительно адекватная доступность фосфора в бедных питательными веществами почвах может снижать зависимость растений от АМ грибов, что, в свою очередь, может снижать поступление углеводов к грибам из корневой системы (Jakobsen и др., 1992; Kiers и др., 2011). Таким образом, более высокий уровень питательных веществ в почве, особенно содержание фосфора, на органических фермах может быть одной из причин их более низкого разнообразия ОТЕ.
С другой стороны, поскольку разные нуклеотипы, несущие разную генетическую информацию, могут быстро расходиться (Angelard и др., 2014), нуклеотипы, адаптированные к почвенным условиям традиционных ферм (стабильное нарушение почвы, засуха и засоление), затем накапливаются и занимают экологические ниши, оставленные теми, которые были отфильтрованы (для подтверждения этого необходимы долгосрочные исследования). Это позволяет АМ грибам претерпевать быстрые генотипические изменения путем изменения относительных частот различных нуклеотипов в ответ на изменение окружающей среды и приводит к высокому функциональному разнообразию (Munkvold и др., 2004; Kruger и др., 2012). Поэтому, хотя на традиционных фермах было обнаружено относительно мало семейств, разнообразие ОТЕ АМ грибов было высоким (Рисунки 3, 4). Это имеет важное значение для производства и применения микоризных инокулянтов, поскольку ОТЕ АМ грибов, присутствующие в настоящее время на малоплодородных или подверженных стрессу сельскохозяйственных полях, могут быть потенциальными и ценными ресурсами. По сравнению с обычными коммерческими микоризными агентами, местные штаммы, отобранные из местных условий, более эффективны для симбиоза растений и грибов (Chenchouni и др., 2020; Wu и др., 2021). Кроме того, грибные инокулянты с использованием местных штаммов могут быть более полезны для поддержания или восстановления стабильных почвенных микропищевых сетей.
Простая и отличающаяся структура сетей арбускулярных микоризных грибов на традиционных фермах
Поскольку некоторые микробные группы более чувствительны к экологическим нарушениям, чем другие (Bhadalung и др., 2005; Qin и др., 2015), не только разнообразие почвенных АМ грибов, но и структура сообщества может быть изменена сельскохозяйственным управлением на традиционных фермах (Bhadalung и др., 2005; Chen и др., 2014). Glomus часто считается наиболее распространенным родом грибов в почвах сельскохозяйственных угодий (Berthrong и др., 2013). Наши данные показали, что Glomus составлял 41,8% сообщества АМ грибов на традиционных фермах, что выше, чем 25,63% на органических фермах (Рисунок 4). Традиционные коммерческие фермы подвергаются более механизированной обработке почвы, чем органические фермы, и некоторые репродуктивные характеристики Glomus, такие как способность размножаться не только через споры, но и через фрагменты мицелия (Biermann и Linderman, 1983), в некоторой степени смягчают негативное влияние частой обработки почвы на выживание АМ грибов. Эти репродуктивные свойства и широко известная устойчивость к стрессам дают Glomus преимущество в сообществах АМ грибов, подвергающихся различным сельскохозяйственным нарушениям (Hijri и др., 2006; Brito и др., 2012).
В отличие от уменьшающейся доли Glomus в сообществе почвенных АМ грибов, Paraglomus увеличивался на органических фермах и даже становился наиболее распространенным родом на некоторых участках отбора проб (Рисунок 4). Несколько исследований показали, что этот род встречается в основном на органических, а не на традиционных сельскохозяйственных почвах (Gosling и др., 2014). Менее нарушающее сельскохозяйственное управление и более высокий уровень фосфора в почве могли частично способствовать увеличению Paraglomus на органических фермах (Jiang S. и др., 2020). Доминирующее семейство в системах традиционного и органического земледелия было одинаковым, а именно Glomeraceae.
Сетевой анализ дополнительно показал, что доминирующий таксон Glomeraceae также был ключевым таксоном в структуре сети почвенных грибов, независимо от системы земледелия. Ключевой таксон занимает уникальное положение в микробных сообществах, и его отсутствие или изменение оказывает значительное влияние на структуру и функцию сообщества (Banerjee и др., 2018). Хотя Glomeraceae является ключевым таксоном грибной сети в обеих системах земледелия, это семейство включало полностью разные компоненты ОТЕ на традиционных и органических фермах соответственно (Рисунок 6). Это означало, что генетический состав доминирующих Glomeraceae был полностью разным, что может приводить к разным функциям сообщества почвенных АМ грибов в этих двух системах земледелия. Более того, традиционное управление фермой привело к более простой структуре сети сообществ почвенных АМ грибов и более слабым взаимодействиям между узлами ОТЕ. Более высокий процент отрицательных связей на традиционных фермах может подразумевать более конкурентные взаимодействия или дифференциацию ниш между ОТЕ (Deng и др., 2016). Несмотря на высокое разнообразие ОТЕ сообществ почвенных АМ грибов на традиционных фермах, более простая структура сети может снижать их функции и устойчивость к изменениям окружающей среды по сравнению с органическими фермами.
Заключение
Сравнение различий в составе сообщества почвенных АМ грибов между традиционными и органическими фермами важно как для сельскохозяйственного производства, так и для восстановления деградированных пахотных почв. Мы обнаружили более высокую интенсивность колонизации корней кукурузы АМ грибами и более высокое таксономическое разнообразие ОТЕ на традиционных фермах по сравнению с органическими, что противоречит ранее обнаруженному. Это свидетельствует о том, что фоновый уровень плодородия почвы должен учитываться при оценке влияния систем традиционного и органического земледелия на почвенные АМ грибы. Хотя системы традиционного земледелия приводят к иному составу и более простой структуре сообщества почвенных АМ грибов, на традиционных фермах в настоящее время присутствуют потенциально разнообразные ресурсы ОТЕ, которые могут быть ценны для эффективного производства микоризных инокулянтов. Штаммы АМ грибов, отобранные на традиционных фермах, могут быть потенциальными штаммами для создания более эффективных биоудобрений для промышленного применения в сельском хозяйстве, в то время как штаммы с органических ферм могут быть потенциальным ресурсом для восстановления фауны деградированных пахотных земель или устойчивого сельского хозяйства.
Дополнительный материал
Дополнительный материал к этой статье можно найти онлайн по адресу: https://www.frontiersin.org/articles/10.3389/fmicb.2022.914627/full#supplementary-material
Сноски
3. ^ https://gephi.org/
Ссылки
1. Akiyama K., Matsuzaki K., Hayashi H. (2005). Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi. Nature 435, 824–827. doi: 10.1038/nature03608. CrossRef. Google Scholar.
2. Angelard C., Tanner C. J., Fontanillas P., Niculita-Hirzel H., Masclaux F., Sanders I. R. (2014). Rapid genotypic change and plasticity in arbuscular mycorrhizal fungi is caused by a host shift and enhanced by segregation. ISME J. 8, 284–294. doi: 10.1038/ismej.2013.154. CrossRef. Google Scholar.
3. Banerjee S., Schlaeppi K., van der Heijden M. G. A. (2018). Keystone taxa as drivers of microbiome structure and functioning. Nat. Rev. Microbiol. 16, 567–576. doi: 10.1038/s41579-018-0024-1. CrossRef. Google Scholar.
4. Begum N., Qin C., Ahanger M. A., Raza S., Khan M. I., Ashraf M., et al. (2019). Role of arbuscular mycorrhizal fungi in plant growth regulation: implications in abiotic stress tolerance. Front. Plant Sci. 10:1068. doi: 10.3389/fpls.2019.01068. CrossRef. Google Scholar.
5. Bender S. F., van der Heijden M. G. A. (2015). Soil biota enhance agricultural sustainability by improving crop yield, nutrient uptake and reducing nitrogen leaching losses. J. Appl. Ecol. 52, 228–239. doi: 10.1111/1365-2664.12351. CrossRef. Google Scholar.
6. Berthrong S. T., Buckley D. H., Drinkwater L. E. (2013). Agricultural management and labile carbon additions affect soil microbial community structure and interact with carbon and nitrogen cycling. Microb. Ecol. 66, 158–170. doi: 10.1007/s00248-013-0225-0. CrossRef. Google Scholar.
7. Bever J., Wang M. (2005). Hyphal fusion and multigenomic structure. Nature 433, E3–E4. doi: 10.1038/nature03294. CrossRef. Google Scholar.
8. Bhadalung N. N., Suwanarit A., Dell B., Nopamornbodi O., Thamchaipenet A., Rungchuang J. (2005). Effects of long-term NP-fertilization on abundance and diversity of arbuscular mycorrhizal fungi under a maize cropping system. Plant Soil 270, 371–382. doi: 10.1007/s11104-004-1829-4. CrossRef. Google Scholar.
9. Bi Y., Qiu L., Zhakypbek Y., Jiang B., Cai Y., Sun H. (2018). Combination of plastic film mulching and AMF inoculation promotes maize growth, yield and water use efficiency in the semiarid region of Northwest China. Agric. Water Manag. 201, 278–286. doi: 10.1016/j.agwat.2017.12.008. CrossRef. Google Scholar.
10. Biermann B., Linderman R. (1983). Use of vesicular-arbuscular mycorrhizal roots, intraradical vesicles and extraradical vesicles as inoculum. New Phytologist 95, 97–105. doi: 10.1111/j.1469-8137.1983.tb03472.x. CrossRef. Google Scholar.
11. Boomsma C. R., Vyn T. J. (2008). Maize drought tolerance: potential improvements through arbuscular mycorrhizal symbiosis? Field Crops Res. 108, 14–31. doi: 10.1016/j.fcr.2008.03.002. CrossRef. Google Scholar.
12. Brito I., Goss M. J., de Carvalho M., Chatagnier O., van Tuinen D. (2012). Impact of tillage system on arbuscular mycorrhiza fungal communities in the soil under Mediterranean conditions. Soil Tillage Res. 121, 63–67. doi: 10.1016/j.still.2012.01.012. CrossRef. Google Scholar.
13. Caporaso J. G., Kuczynski J., Stombaugh J., Bittinger K., Bushman F. D., Costello E. K., et al. (2010). QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 7, 335–336. doi: 10.1038/nmeth.f.303. CrossRef. Google Scholar.
14. Cavagnaro T. R., Bender S. F., Asghari H. R., van der Heijden M. G. A. (2015). The role of arbuscular mycorrhizas in reducing soil nutrient loss. Trends Plant Sci. 20, 283–290. doi: 10.1016/j.tplants.2015.03.004. CrossRef. Google Scholar.
15. Cely M. V. T., de Oliveira A. G., de Freitas V. F., de Luca M. B., Barazetti A. R., dos Santos I. M. O., et al. (2016). Inoculant of arbuscular mycorrhizal fungi (Rhizophagus clarus) increase yield of soybean and cotton under field conditions. Front. Microbiol. 7:720. doi: 10.3389/fmicb.2016.00720. CrossRef. Google Scholar.
16. Chen Y. L., Zhang X., Ye J. S., Han H. Y., Wan S. Q., Chen B. D. (2014). Six-year fertilization modifies the biodiversity of arbuscular mycorrhizal fungi in a temperate steppe in Inner Mongolia. Soil Biol. Biochem. 69, 371–381. doi: 10.1016/j.soilbio.2013.11.020. CrossRef. Google Scholar.
17. Chenchouni H., Mekahlia M. N., Beddiar A. (2020). Effect of inoculation with native and commercial arbuscular mycorrhizal fungi on growth and mycorrhizal colonization of olive (Olea europaea L.). Sci. Hortic. 261:108969. doi: 10.1016/j.scienta.2019.108969. CrossRef. Google Scholar.
18. Daniell T. J., Husband R., Fitter A. H., Young J. P. W. (2001). Molecular diversity of arbuscular mycorrhizal fungi colonising arable crops. FEMS Microbiol. Ecol. 36, 203–209. doi: 10.1016/s0168-6496(01)00134-9. CrossRef. Google Scholar.
19. Daniels B. A., Skipper H. D. (1982). Methods for the Recovery and Quantitative Estimation of Propagules From Soil. St. Paul: American Phytopathological Society. Google Scholar.
20. Davison J., Moora M., Opik M., Adholeya A., Ainsaar L., Ba A., et al. (2015). Global assessment of arbuscular mycorrhizal fungus diversity reveals very low endemism. Science 349, 970–973. doi: 10.1126/science.aab1161. CrossRef. Google Scholar.
21. De'ath G. (2007). Boosted trees for ecological modeling and prediction. Ecology 88, 243–251. doi: 10.1890/0012-9658(2007)88[243:btfema]2.0.co;2. CrossRef. Google Scholar.
22. Deng Y., Zhang P., Qin Y., Tu Q., Yang Y., He Z., et al. (2016). Network succession reveals the importance of competition in response to emulsified vegetable oil amendment for uranium bioremediation. Environ. Microbiol. 18, 205–218. doi: 10.1111/1462-2920.12981. CrossRef. Google Scholar.
23. Franke-Snyder M., Douds D. D., Galvez L., Phillips J. G., Wagoner P., Drinkwater L., et al. (2001). Diversity of communities of arbuscular mycorrhizal (AM) fungi present in conventional versus low-input agricultural sites in eastern Pennsylvania, USA. Appl. Soil Ecol. 16, 35–48. doi: 10.1016/s0929-1393(00)00100-1. CrossRef. Google Scholar.
24. Gerdemann J., Nicolson T. H. (1963). Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Trans. Br. Mycol. Soc. 46, 235–244. doi: 10.1016/S0007-1536(63)80079-0. CrossRef. Google Scholar.
25. Gosling P., Proctor M., Jones J., Bending G. D. (2014). Distribution and diversity of Paraglomus spp. in tilled agricultural soils. Mycorrhiza 24, 1–11. doi: 10.1007/s00572-013-0505-z. CrossRef. Google Scholar.
26. Govindarajulu M., Pfeffer P. E., Jin H. R., Abubaker J., Douds D. D., Allen J. W., et al. (2005). Nitrogen transfer in the arbuscular mycorrhizal symbiosis. Nature 435, 819–823. doi: 10.1038/nature03610. CrossRef. Google Scholar.
27. Graham J. H., Eissenstat D. M. (1994). Host genotype and the formation and function of VA mycorrhizae. Plant Soil 159, 179–185. doi: 10.1007/bf00000107. CrossRef. Google Scholar.
28. Hart M. M., Reader R. J. (2002). Taxonomic basis for variation in the colonization strategy of arbuscular mycorrhizal fungi. New Phytol. 153, 335–344. doi: 10.1046/j.0028-646X.2001.00312.x. CrossRef. Google Scholar.
29. He D., Xiang X., He J. S., Wang C., Chu H. (2016). Composition of soil fungal community is more sensitive to phosphorus than nitrogen addition in alpine meadow on the Qinghai-Tibetan Plateau. Biol. Fertil. Soils 52, 1059–1072. doi: 10.1007/s00374-016-1142-4. CrossRef. Google Scholar.
30. Hijri I., Sýkorová Z., Oehl F., Ineichen K., Mäder P., Wiemken A., et al. (2006). Communities of arbuscular mycorrhizal fungi in arable soils are not necessarily low in diversity. Mol. Ecol. 15, 2277–2289. doi: 10.1111/j.1365-294X.2006.02921.x. CrossRef. Google Scholar.
31. Jakobsen J. I., Abbott L., Robson A. (1992). External hyphae of vesicular-arbuscular mycorrhizal fungi associated with Trifolium subterraneum L. 1. Spread of hyphae and phosphorus inflow into roots. New Phytol. 120, 371–380. doi: 10.1111/j.1469-8137.1992.tb01077.x. CrossRef. Google Scholar.
32. Jiang S., Hu X., Kang Y., Xie C., An X., Dong C., et al. (2020). Arbuscular mycorrhizal fungal communities in the rhizospheric soil of litchi and mango orchards as affected by geographic distance, soil properties and manure input. App. Soil Ecol. 152:103593. doi: 10.1016/j.apsoil.2020.103593. CrossRef. Google Scholar.
33. Jiang Y. J., Luan L., Hu K. J., Liu M. Q., Chen Z. Y., Geisen S., et al. (2020). Trophic interactions as determinants of the arbuscular mycorrhizal fungal community updates with cascading plant-promoting consequences. Microbiome 8:142. doi: 10.1186/s40168-020-00918-6. CrossRef. Google Scholar.
34. Johnson N. C. (2010). Resource stoichiometry elucidates the structure and function of arbuscular mycorrhizas across scales. New Phytol. 185, 631–647. doi: 10.1111/j.1469-8137.2009.03110.x. CrossRef. Google Scholar.
35. Johnson N. C., Rowland D. L., Corkidi L., Egerton-Warburton L. M., Allen E. B. (2003). Nitrogen enrichment alters mycorrhizal allocation at five mesic to semiarid grasslands. Ecology 84, 1895–1908. Google Scholar.
36. Kiers E. T., Duhamel M., Beesetty Y., Mensah J. A., Franken O., Verbruggen E., et al. (2011). Reciprocal rewards stabilize cooperation in the mycorrhizal symbiosis. Science 333, 880–882. doi: 10.1126/science.1208473. CrossRef. Google Scholar.
37. Kim H., Lee K. K., Jeon J., Harris W. A., Lee Y. H. (2020). Domestication of Oryza species eco-evolutionarily shapes bacterial and fungal communities in rice seed. Microbiome 8, 1–17. doi: 10.1186/s40168-020-00805-0. CrossRef. Google Scholar.
38. Koske R., Gemma J. (1989). A modified procedure for staining roots to detect VA mycorrhizas. Mycol. Res. 92, 486–488. doi: 10.1016/S0953-7562(89)80195-9. CrossRef. Google Scholar.
39. Kruger M., Kruger C., Walker C., Stockinger H., Schussler A. (2012). Phylogenetic reference data for systematics and phylotaxonomy of arbuscular mycorrhizal fungi from phylum to species level. New Phytol. 193, 970–984. doi: 10.1111/j.1469-8137.2011.03962.x. CrossRef. Google Scholar.
40. Lekberg Y., Arnillas C. A., Borer E. T., Bullington L. S., Fierer N., Kennedy P. G., et al. (2021). Nitrogen and phosphorus fertilization consistently favor pathogenic over mutualistic fungi in grassland soils. Nat. Commun. 12, 1–8. doi: 10.1038/s41467-021-23605-y. CrossRef. Google Scholar.
41. Li Q. S., Willardson L. S., Deng W., Li X. J., Liu C. J. (2005). Crop water deficit estimation and irrigation scheduling in western Jilin province, Northeast China. Agric. Water Manag. 71, 47–60. doi: 10.1016/j.agwat.2004.07.003. CrossRef. Google Scholar.
42. Lin C., Wang Y., Liu M., Li Q., Xiao W., Song X. (2020). Effects of nitrogen deposition and phosphorus addition on arbuscular mycorrhizal fungi of Chinese fir (Cunninghamia lanceolata). Sci. Rep. 10:12260. doi: 10.1038/s41598-020-69213-6. CrossRef. Google Scholar.
43. Lin X. G., Feng Y. Z., Zhang H. Y., Chen R. R., Wang J. H., Zhang J. B., et al. (2012). Long-term balanced fertilization decreases arbuscular mycorrhizal fungal diversity in an arable soil in north China revealed by 454 pyrosequencing. Environ. Sci. Technol. 46, 5764–5771. doi: 10.1021/es3001695. CrossRef. Google Scholar.
44. Liu W., Zhang Y. L., Jiang S. S., Murray P. J., Liao L. Q., Li X. L., et al. (2019). Spatiotemporal differences in the arbuscular mycorrhizal fungi communities in soil and roots in response to long-term organic compost inputs in an intensive agricultural cropping system on the North China Plain. J. Soils Sediments 19, 2520–2533. doi: 10.1007/s11368-019-02244-3. CrossRef. Google Scholar.
45. Lu R. K. (1999). Soil and Agro-Chemical Analytical Methods. Beijing: China Agricultural Science and Technology Press, 146–195. Google Scholar.
46. Munkvold L., Kjoller R., Vestberg M., Rosendahl S., Jakobsen I. (2004). High functional diversity within species of arbuscular mycorrhizal fungi. New Phytol. 164, 357–364. doi: 10.1111/j.1469-8137.2004.01169.x. CrossRef. Google Scholar.
47. Niedziński T., Sierra M. J., Łabętowicz J., Noras K., Cabrales C., Millán R. (2021). Release of nitrogen from granulate mineral and organic fertilizers and its effect on selected chemical parameters of soil. Agronom. Basel 11:1981. doi: 10.3390/agronomy11101981. CrossRef. Google Scholar.
48. Oehl F., Sieverding E., Mäder P., Dubois D., Ineichen K., Boller T., et al. (2004). Impact of long-term conventional and organic farming on the diversity of arbuscular mycorrhizal fungi. Oecologia 138, 574–583. doi: 10.1007/s00442-003-1458-2. CrossRef. Google Scholar.
49. Pawlowska T. E., Taylor J. W. (2004). Organization of genetic variation in individuals of arbuscular mycorrhizal fungi. Nature 427, 733–737. doi: 10.1038/nature02290. CrossRef. Google Scholar.
50. Qin H., Lu K., Strong P. J., Xu Q., Wu Q., Xu Z., et al. (2015). Long-term fertilizer application effects on the soil, root arbuscular mycorrhizal fungi and community composition in rotation agriculture. Appl. Soil Ecol. 89, 35–43. doi: 10.1016/j.apsoil.2015.01.008. CrossRef. Google Scholar.
51. Rosendahl S. (2008). Communities, populations and individuals of arbuscular mycorrhizal fungi. New Phytol. 178, 253–266. doi: 10.1111/j.1469-8137.2008.02378.x. CrossRef. Google Scholar.
52. Roy J., Reichel R., Brueggemann N., Hempel S., Rillig M. C. (2017). Succession of arbuscular mycorrhizal fungi along a 52-year agricultural recultivation chronosequence. FEMS Microbiol. Ecol. 93:fix102. doi: 10.1093/femsec/fix102. CrossRef. Google Scholar.
53. Ryan M. H., Graham J. H. (2002). Is there a role for arbuscular mycorrhizal fungi in production agriculture? Plant Soil 244, 263–271. doi: 10.1023/a:1020207631893. CrossRef. Google Scholar.
54. Ryan M. H., Graham J. H. (2018). Little evidence that farmers should consider abundance or diversity of arbuscular mycorrhizal fungi when managing crops. New Phytol. 220, 1092–1107. doi: 10.1111/nph.15308. CrossRef. Google Scholar.
55. Sale V., Aguilera P., Laczko E., Mader P., Berner A., Zihlmann U., et al. (2015). Impact of conservation tillage and organic farming on the diversity of arbuscular mycorrhizal fungi. Soil Biol. Biochem. 84, 38–52. doi: 10.1016/j.soilbio.2015.02.005. CrossRef. Google Scholar.
56. Scullion J., Eason W. R., Scott E. P. (1998). The effectivity of arbuscular mycorrhizal fungi from high input conventional and organic grassland and grass-arable rotations. Plant Soil 204, 243–254. doi: 10.1023/a:1004319325290. CrossRef. Google Scholar.
57. Sheng M., Lalande R., Hamel C., Ziadi N. (2013). Effect of long-term tillage and mineral phosphorus fertilization on arbuscular mycorrhizal fungi in a humid continental zone of Eastern Canada. Plant Soil 369, 599–613. doi: 10.1007/s11104-013-1585-4. CrossRef. Google Scholar.
58. Shi X., Yu D., Warner E., Pan X., Petersen G., Gong Z., et al. (2004). Soil database of 1:1,000,000 digital soil survey and reference system of the Chinese genetic soil classification system. Soil Surv. Horiz. 45, 129–136. doi: 10.2136/sh2004.4.0129. CrossRef. Google Scholar.
59. Tang X., Li J., Ma Y., Hao X., Li X. (2008). Phosphorus efficiency in long-term (15 years) wheat-maize cropping systems with various soil and climate conditions. Field Crop. Res. 108, 231–237. doi: 10.1016/j.fcr.2008.05.007. CrossRef. Google Scholar.
60. The Ministry of Agriculture and Rural Affairs of China (2020). Action Plan For Conservation Tillage Of Black Soil In The Northeast China (2020–2025). Available online at: http://www.moa.gov.cn/gk/tzgg_1/tz/202003/t20200318_6339304.htm (accessed March 18, 2020). Google Scholar.
61. Tisserant E., Malbreil M., Kuo A., Kohler A., Symeonidi A., Balestrini R., et al. (2013). Genome of an arbuscular mycorrhizal fungus provides insight into the oldest plant symbiosis. PNAS 110, 20117–20122. doi: 10.1073/pnas.1313452110. CrossRef. Google Scholar.
62. Trouvelot A., Kough J. L., Gianinazzi-Pearson V. (1986). "Mesure du taux de mycorhization VA d'un système radiculaire. Recherche de méthodes d'estimation ayant une signification fonctionnelle," in Physiological and Genetic Aspects of Mycorrhizae, eds V. Gianinazzi-Pearson and S. Gianinazzi (Paris: INRA), 217–221. Google Scholar.
63. van Geel M., Verbruggen E., De Beenhouwer M., van Rennes G., Lievens B., Honnay O. (2017). High soil phosphorus levels overrule the potential benefits of organic farming on arbuscular mycorrhizal diversity in northern vineyards. Agric. Ecosyst. Environ. 248, 144–152. doi: 10.1016/j.agee.2017.07.017. CrossRef. Google Scholar.
64. Wu F., Dong M., Liu Y., Ma X., An L., Young J. P. W., et al. (2011). Effects of long-term fertilization on AM fungal community structure and Glomalin-related soil protein in the Loess Plateau of China. Plant Soil 342, 233–247. doi: 10.1007/s11104-010-0688-4. CrossRef. Google Scholar.
65. Wu Y. H., Wang H., Liu M., Li B., Chen X., Ma Y. T., et al. (2021). Effects of native arbuscular mycorrhizae isolated on root biomass and secondary metabolites of Salvia miltiorrhiza Bge. Front. Plant Sci. 12:617892. doi: 10.3389/fpls.2021.617892. CrossRef. Google Scholar.
66. Xiang X., Gibbons S. M., He J. S., Wang C., He D., Li Q., et al. (2016). Rapid response of arbuscular mycorrhizal fungal communities to short-term fertilization in an alpine grassland on the Qinghai-Tibet Plateau. PeerJ 4:e2226. doi: 10.7717/peerj.2226. CrossRef. Google Scholar.
67. Xiao D., Che R., Liu X., Tan Y., Yang R., Zhang W., et al. (2019). Arbuscular mycorrhizal fungi abundance was sensitive to nitrogen addition but diversity was sensitive to phosphorus addition in karst ecosystems. Biol. Fertil. Soils 55, 457–469. doi: 10.1007/s00374-019-01362-x. CrossRef. Google Scholar.
68. Yang W., Guo Y., Wang X., Chen C., Hu Y., Cheng L., et al. (2017). Temporal variations of soil microbial community under compost addition in black soil of Northeast China. Appl. Soil Ecol. 121, 214–222. doi: 10.1016/j.apsoil.2017.10.005. CrossRef. Google Scholar.
69. Yang Z., Yu T., Hou Q., Xia X., Feng H., Huang C., et al. (2014). Geochemical evaluation of land quality in China and its applications. J. Geochem. Explor. 139, 122–135. doi: 10.1016/j.gexplo.2013.07.014. CrossRef. Google Scholar.
70. Yue X., Zhang J., Shi A., Yao S., Zhang B. (2016). Manure substitution of mineral fertilizers increased functional stability through changing structure and physiology of microbial communities. Eur. J. Soil Biol. 77, 34–43. doi: 10.1016/j.ejsobi.2016.10.002. CrossRef. Google Scholar.
71. Zeng H. L., Yu L. L., Liu P., Wang Z. G., Chen Y., Wang J. W. (2021). Nitrogen fertilization has a stronger influence than cropping pattern on AMF community in maize/soybean strip intercropping systems. Appl. Soil Ecol. 167:104034. doi: 10.1016/j.apsoil.2021.104034. CrossRef. Google Scholar.
72. Zhu K. G. (1994). Soil Type of China, Vol. 2. Beijing: China Agriculture Press. Google Scholar.

Chen J, Li J, Yang Y, Wang Y, Zhang Y and Wang P (2022) Effects of Conventional and Organic Agriculture on Soil Arbuscular Mycorrhizal Fungal Community in Low-Quality Farmland. Front. Microbiol. 13:914627. doi: 10.3389/fmicb.2022.914627
Перевод статьи «Effects of Conventional and Organic Agriculture on Soil Arbuscular Mycorrhizal Fungal Community in Low-Quality Farmland» авторов Chen J, Li J, Yang Y, Wang Y, Zhang Y and Wang P., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык
Фото: Designed by Freepik





















































































































Комментарии (0)