Что происходит с почвой и бактериями при разном земледелии? Результаты многолетнего тепличного опыта
Рост сельского хозяйства привел к нерациональному использованию синтетических пестицидов и удобрений, а также к неустойчивым методам управления посевными площадями, что может усугубить негативное воздействие на микробную популяцию, а также на физические и химические характеристики почвенной экосистемы.
Аннотация
Исходя из этого, настоящее исследование было спланировано для оценки влияния длительных агротехнических практик на различные физико-химические параметры почвы, а также на численность и разнообразие почвенных микроорганизмов на разных глубинах почвы (0–20 см и 20–40 см) на экспериментальной станции Цюйчжоу Китайского сельскохозяйственного университета (провинция Хэбэй, Китай) в октябре и декабре 2016 года. Влияние агротехнических практик на численность и разнообразие почвенных микроорганизмов изучалось с помощью методов фосфолипидного анализа жирных кислот (PLFA) и высокопроизводительного секвенирования ДНК. Полученные результаты показали, что почва имеет нейтральную или слабощелочную реакцию, с наибольшим содержанием влаги при органическом земледелии (ORF) на глубине 0–20 см и наименьшим — при традиционном земледелии на глубине 20–40 см. Было установлено, что органическое земледелие (ORF) значительно увеличивало содержание общего органического углерода (TOC), общего углерода (TC), аммонийного азота, доступного азота (AN), общего азота (TN), общего фосфора (TP) и доступного фосфора (AP) по сравнению с режимами низкозатратного и традиционного земледелия в образцах почвы как за октябрь, так и за декабрь. Корреляционный анализ показал значительную (при p ≤ 0,05 и 0,01) сильную положительную связь между различными физическими и химическими свойствами исследуемой почвы. Дисперсионный анализ (ANOVA) и многомерный дисперсионный анализ (MANOVA) показали значительное влияние взаимодействия между глубиной почвы и режимами земледелия на почвенные параметры. Анализ главных компонент (PCA) показал наиболее значимую корреляцию между большинством типов бактерий (G+ бактерии, G− бактерии, актиномицеты) и содержанием доступного фосфора (AP), общего доступного азота, общего органического углерода (TOC) и влажности почвы (WC). Корреляционный анализ Пирсона выявил значительную корреляцию между филумами микроорганизмов (Proteobacteria, Bacteroidetes и Latescibacteria) и классами микроорганизмов (Alphaproteobacteria, Sphingobacteriia, Flavobacteriia) с большинством физико-химических свойств почвы.
Введение
Почва выступает ключевым элементом для производства продуктов питания, от которого зависит жизнь на Земле. Почвенная экосистема предоставляет различные функциональные услуги, такие как поддержание почвенного плодородия, обеспечение стабильности экосистемы и регулирование изменения климата (Medan et al., 2011; Jing et al., 2015). Хорошо известно, что антропогенные нарушения, такие как нерациональное использование химических удобрений и режимы ведения сельского хозяйства, в значительной степени влияют на физические и химические свойства почвы, а также на биохимическую активность. Агротехнические практики могут влиять на эффективность использования воды и влажность почвы из-за нерационального внесения удобрений, выбора типа сельскохозяйственных культур, управления орошением и т. д. (Niyogi et al., 2010). Постоянная обработка почвы может нарушать образование макроагрегатов, что приводит к потере органического углерода (OC) (Nandan et al., 2019), и, как следствие, к снижению концентрации CO₂ в атмосфере (Xu and Zhang, 2021). Общий органический углерод (TOC) почвы может служить лучшим показателем качества почвы и запасов углерода (Venkatesh et al., 2013). Азот является еще одним важнейшим питательным веществом, необходимым для лучшего производства сельскохозяйственных культур. Он улучшает адаптацию почвы к стрессовым факторам окружающей среды, таким как ограниченная доступность воды и засоленность (Hou et al., 2007). Содержание общего азота (TN) и органического углерода в почве может варьировать по почвенному профилю и, как правило, уменьшается с увеличением глубины почвы. Другое важное питательное вещество — фосфор (P) — его удержание в приповерхностной зоне почвы колеблется в зависимости от различных методов обработки почвы (Carvalho et al., 2014), и только органическая форма фосфора остается доступной для поглощения растениями (Zhu et al., 2016). Доступность фосфора (AP) в почве может ограничиваться как доступностью, так и извлечением фосфора из-за архитектуры корневой системы растений и микоризной ассоциации (Xomphoutheb et al., 2020).
В сельскохозяйственной практике обычно применяются такие системы земледелия, как традиционное (COF), низкозатратное (LOF), органическое (ORF) и многие другие. Традиционный тип земледелия может приводить к снижению плодородия почвы, производству продуктов питания с остаточными количествами химических веществ и увеличению рыночной стоимости (Riedo et al., 2021). COF ухудшает физическое состояние почвы, снижает эффективность использования воды и удобрений, что приводит к задержке роста и снижению урожайности зерна (Zhou et al., 2014). Однако было установлено, что COF увеличивает накопление азота и фосфора в растениях, которые обычно остаются для них недоступными (Shannon et al., 2002). Было обнаружено, что система LOF увеличивает накопление избыточного азота в почвенной среде (Clark et al., 1998). ORF является одной из наиболее широко используемых систем земледелия во всем мире. Она предполагает использование промежуточных культур, навоза животных, органических удобрений и рециклинг растительных остатков, а также избегает вспашки полей (Gajda et al., 2020). ORF строго избегает использования химических средств защиты растений (синтетических минеральных удобрений), что приводит к улучшению биофизической среды почвы и получению качественной сельскохозяйственной продукции (Kwiatkowski et al., 2020). Было обнаружено, что ORF повышает содержание органического вещества и микроэлементов в почве, что может улучшить ее качество, структуру и повысить биологическую активность почвы, водоудерживающую способность и скорость инфильтрации (Morshedi et al., 2017; Sankaramoorthy and Rangasamy, 2019; Kwiatkowski and Harasim, 2020; Hietala et al., 2021). Несмотря на множество преимуществ, органический тип земледелия может приводить к дефициту азота в почве и нежелательному росту сорняков, что приводит к снижению урожайности (Ponisio et al., 2015). ORF производит более питательные продукты с меньшим содержанием остатков пестицидов по сравнению с другими системами земледелия (Seufert and Ramankutty, 2017). На содержание питательных веществ в почве могут влиять тип сельскохозяйственных угодий и используемый метод ведения хозяйства, что может изменить биогеохимические циклы углерода и азота и привести к снижению запасов углерода в почве (Wang et al., 2011). Многие предыдущие исследования сообщали о влиянии агротехнических практик на почвенные микроорганизмы и их активность (Zhu et al., 2018). Бессменное возделывание культур является одним из факторов, которые могут привести к деградации и нарушению баланса в составе и структуре почвенного микробного сообщества (Lu et al., 2013; Li Y. et al., 2014).
Качество почвы — комплексная способность поддерживать лучшее качество окружающей среды, а также высокую биологическую продуктивность, что приводит к улучшению здоровья растений. Это можно оценить с помощью почвенных индикаторов, т. е. физико-химических свойств и биохимических процессов (Bunemann et al., 2018; Ukalska-Jaruga et al., 2019). В почве присутствие различных микро- и макроэлементов (Yang et al., 2020), содержание азота и некоторых элементов (Fe, K, Mg, Mn, Zn); влажность, соотношение C:N, минерализуемый и окисляемый перманганатом углерод, микробный состав почвы и активность почвенных ферментов (Gannett et al., 2019; Ozlu et al., 2019; Thomas et al., 2019) могут служить важными индикаторами здоровья/плодородия почвы. Почвенные микроорганизмы содержат большое количество жирных кислот, которые различаются у каждого отдельного организма, обитающего в почве (Willers et al., 2015). Оценка этих профилей жирных кислот с помощью анализа фосфолипидных жирных кислот (PLFA) позволяет определить наличие или отсутствие и относительную численность любого типа микроорганизмов (грамотрицательные и грамположительные бактерии, актинобактерии, актиномицеты, грибы и т. д.) в почве (Balser et al., 2019; Brinton, 2020). Ранее исследования влияния агротехнических практик на почвенные микробные сообщества включали использование библиотек генов, ПЦР-амплификацию и гель-электрофорез, которые позволяют обнаружить наличие или отсутствие некоторых специфических микробных групп с высокой численностью (Chen et al., 2012). Использование методов секвенирования следующего поколения (NGS), таких как методы на основе Illumina и пиросеквенирование, предоставляет больше информации и позволяет идентифицировать таксоны микроорганизмов с низкой численностью в почве. Это позволяет детально проанализировать огромное количество таксономических профилей почвенных микроорганизмов (Uroz et al., 2013; Zhu et al., 2018).
Информация о влиянии методов ведения сельского хозяйства или изменений, связанных с внесением удобрений, на сельскохозяйственные почвы все еще недостаточно изучена. В прошлом большинство исследований были посвящены влиянию различных систем земледелия на физико-химические и микробиологические свойства почвы, однако влияние на численность и разнообразие почвенных микроорганизмов рассматривалось редко. Поэтому комплексная оценка влияния стратегий ведения хозяйства на важные почвенные параметры (C, N, P), а также на численность почвенных микроорганизмов в различных слоях почвы необходима как с агрономической, так и с экологической точек зрения. Исходя из этих фактов, настоящее исследование было направлено на: (i) проверку влияния трех агротехнических практик (традиционной, низкозатратной и органической систем) на физико-химические параметры почвы, включая pH, влажность (WC) (%), общий углерод (TC) (%), общий органический углерод (TOC) (%), аммонийный азот (NH₄N) (%), нитратный азот (NO₃N) (%), доступный азот (AN) (%), общий азот (TN) (%), общий фосфор (TP) (%) и доступный фосфор (AP) в разных слоях почвы (0–20 и 20–40 см) и в разные месяцы отбора проб (октябрь и декабрь); и (ii) исследовать изменения в численности и разнообразии микроорганизмов в ответ на различные системы земледелия. Мы предположили, что три исследуемые системы земледелия будут по-разному влиять на физико-химические параметры почвы и численность почвенных микроорганизмов на разных глубинах.
Материалы и методы
Район исследования
Настоящее исследование было проведено в Сельскохозяйственном университете Китая, Цюйчжоу (36°87′ с.ш., 115°02′ в.д.), провинция Хэбэй, Китай. На экспериментальной станции Цюйчжоу долгосрочный тепличный эксперимент был заложен в марте 2002 года и с тех пор поддерживается производство овощей. Температура и влажность, поддерживаемые во всех трех теплицах, составляли 14,8–24,6°C (октябрь), 8,2–17,5°C (декабрь) и 60%, а интенсивность освещения соответствовала 12-часовому световому/темновому циклу. Параметры почвы, такие как органическое вещество (COF — 1,893%; LOF — 1,525%; ORF — 1,663%), доступный азот (AN) (COF — 0,136%; LOF — 0,119%; ORF — 0,117%) и доступный фосфор (AP) (COF — 0,0188%; LOF — 0,00816%; ORF — 0,01391%), были измерены до начала эксперимента в 2002 году. Станция характеризуется теплым полувлажным климатом с сухой холодной зимой и дождливым летом. Среднегодовая температура составляет 13,2°C, а основной тип почвы — илисто-флюво-аквический (Li X. et al., 2014).
В настоящем эксперименте рассматривались три различные тепличные системы (длиной 52 м и шириной 7 м) при трех режимах ведения хозяйства: COF, LOF и ORF (Wang et al., 2021). Все три теплицы организованы интегрированно, как смежные участки в одном месте с одинаковым типом почвы (илисто-флюво-аквической). Выращиваемые сорта овощей, тип орошения и методы обработки почвы одинаковы для всех теплиц в течение одного вегетационного сезона. Овощи, такие как огурец, сельдерей, цветная капуста, баклажан, фенхель и томат, высаживаются в севообороте в разные сезоны. Сельдерей и цветная капуста выращиваются во всех теплицах в осенний сезон одновременно, а баклажан высаживается весной. В традиционном типе земледелия использовались химические удобрения, пестициды и куриный + коровий навоз, образующийся в местных теплицах, в то время как в LOF использовалось 50% химических удобрений и 50% компостированных удобрений, а также малотоксичные биологические химические пестициды для защиты растений. ORF-практика осуществлялась в соответствии с Международной федерацией органического сельского хозяйства (IFOAM) и использовала компост из куриного и коровьего навоза, а также биологические и физические методы защиты культурных растений.
Физико-химический анализ
Образцы почвы были отобраны в октябре (после колонизации) и декабре (перед уборкой урожая/созреванием) 2016 года из всех трех теплиц под посевами сельдерея. Образцы отбирались на глубине 0–20 и 20–40 см по схеме «конверт» в рамках 5-точечного метода отбора проб. Всего из каждой теплицы для каждой системы земледелия было отобрано по четыре образца в октябре и декабре. Свежие образцы почвы, доставленные в лабораторию, были немедленно использованы для анализа численности и разнообразия почвенных микроорганизмов, в то время как для физико-химического анализа образцы почвы высушивали на воздухе, помещали в асептические пакеты, маркировали и хранили в холодильнике при 4°C для дальнейшего анализа.
Были определены различные физико-химические параметры: pH, WC, TC, TOC, NH₄N, NO₃N, AN, TN, TP и AP, а также численность и разнообразие почвенных микроорганизмов в образцах почвы, отобранных на полях с сельдереем. pH (1:5 почва:вода) определяли с помощью pH-метра; влажность измеряли путем высушивания образцов почвы в течение 24 часов при 105°C, в то время как общее содержание C и N измеряли методом сухого сжигания (950°C) с использованием анализатора Vario Max CN (Elementar; Ханау, Германия). Содержание TOC в почве определяли с помощью анализатора TOC (Elementar, жидкостной TOC II, Германия) (Wang et al., 2021). Содержание NH₄N в почве измеряли путем добавления 5 г образца почвы в 25 мл 2 моль/л KCl в колбу объемом 250 мл (Robertson et al., 1999). Колбу помещали на шейкер на 1 час, и раствор фильтровали. В мерную колбу на 50 мл к 10 мл фильтрата добавляли 5 мл 0,1 моль/л раствора фенола и 5 мл 0,1 моль/л NaClO, и раствор оставляли в покое на 1 час. Через 1 час содержание аммонийного азота в растворе определяли путем измерения оптической плотности при 625 нм с использованием спектрофотометра (Shimazu UV-3600, Япония). Далее NO₃N определяли, помещая 1 мл фильтрованного раствора в колориметрическую пробирку, добавляя 25 мл дистиллированной воды и измеряя оптическую плотность при 210 нм с помощью спектрофотометра. AN определяли методом диффузионного поглощения, при котором 2 г воздушно-сухой почвы пропускали через сито 1 мм и гидролизовали 1 мл/л NaOH для высвобождения NH₃, а затем NH₃ поглощался добавлением 20 г/л H₃BO₃ (Griesheim and Mulvaney, 2019). Раствор титровали 0,005 мл/л H₂SO₄. AP и TP определяли методами Olsen et al. (1954) и Bowman (1988) соответственно.
Численность и разнообразие почвенных микроорганизмов
Численность и разнообразие почвенных микроорганизмов определяли методами анализа фосфолипидных жирных кислот (PLFA) и высокопроизводительного секвенирования ДНК.
Технология фосфолипидных жирных кислот
PLFA в образцах почвы разделяли и детектировали с помощью газовой хроматографии по стандартному методу Bligh and Dyer (1959) с небольшими модификациями. Для экстракции фосфолипидов в образцах почвы использовали цитратный буфер. К 8 г лиофилизированного образца почвы в белых толстостенных центрифужных пробирках добавляли однофазный экстрагент — раствор цитратного буфера [хлороформ:метанол:цитратный буфер в соотношении 1:2:0,8 (об/об)], почву многократно пропитывали экстрагирующим раствором и тщательно встряхивали. Экстракт переносили в делительную воронку, добавляли 12 мл хлороформа и фосфатного буфера, тщательно встряхивали в течение 2 минут и оставляли в покое на ночь. Для очистки жирных кислот использовали SPE-колонку (активированную 3 мл хлороформа). К экстракту добавляли 5 мл хлороформа, затем 10 мл ацетона и, наконец, промывали 5 мл метанола. Метанольную фазу собирали в пробирки, помещали на водяную баню при 32°C и концентрировали под N₂.
Для этерификации в пробирки добавляли 1 мл смеси толуол:метанол (1:1) и 0,2 моль/л раствора KOH, тщательно встряхивали, инкубировали при 37°C в течение 1 часа. После инкубации добавляли 0,3 мл 1 моль/л раствора уксусной кислоты, 2 мл н-гексана и 2 мл сверхчистой воды, встряхивали на низкой скорости в течение 10 минут (повторяли встряхивание и выщелачивание один раз), верхний слой н-гексана переносили во флаконы. Образец обезвоживали и высушивали на водяной бане под N₂ и получали образцы метилированных жирных кислот. Образцы метилированных жирных кислот растворяли в 200 мкл н-гексана, а PLFA в образцах идентифицировали и анализировали на газовом хроматографе (Agilent 6890N, США) с использованием программного обеспечения MIDI (MINDI, Newark, Delaware, USA) в сочетании с системой SherlockMIS4.5 (Sherlock Microbial Identification System). PLFA были идентифицированы с различными биомаркерами для различных микробных групп в соответствии с методом Kandler and Konig (1998).
Технология высокопроизводительного секвенирования ДНК
Метод высокопроизводительного секвенирования использовался для изучения влияния агротехнических практик на относительную численность и разнообразие почвенных микроорганизмов. Для экстракции микробной ДНК из образцов почвы использовали набор FastDNA SPIN Kit (MP Biomedicals, LLC, США; Протокол: #116560200-201908/#116560000-201908) в соответствии с протоколом производителя, ранее использованным во многих исследованиях (Song et al., 2017; Hemkemeyer et al., 2018; Paul et al., 2018). Набор SPIN позволяет эффективно выделять готовую к ПЦР геномную ДНК непосредственно из образцов почвы менее чем за 30 минут. ДНК экстрагировали из 0,5 г свежей почвы с использованием MT- и фосфатных буферов и повторного центрифугирования с использованием фильтров SPIN™. Пробирка для сбора с элюированной ДНК хранилась при -20 или -80°C для дальнейшей обработки (Niu et al., 2017). Для выполнения парно-концевого секвенирования продуктов ПЦР-амплификации использовали платформу секвенирования Illumina MiSeq 2 × 300. Концентрацию образцов ДНК оценивали с помощью флуориметра Qubit 2.0 (Invitrogen, Carlsbad, CA), а библиотеки секвенирования создавали с использованием набора MetaVx™ Library Construction Kit (GENEWIZ, Inc., South Plainfield, NJ, USA). Все реакции выполнялись в двух повторах. Для каждой индивидуальной библиотеки было взято по два прочтения. ДНК амплифицировали методом ПЦР с использованием прокариотических праймеров к гену 16S рРНК. Праймеры 515F (GTG CCA GCM GCC GCG G) и 907R (CCG TCA ATT CMT TTR AGT TT) были разработаны для амплификации гипервариабельных областей (V3 и V4) гена 16S рРНК бактерий (Roesch et al., 2007). Используя 30–50 нг ДНК в качестве матрицы, серию ПЦР-праймеров, разработанных GENEWIZ, использовали для амплификации обеих гипервариабельных областей прокариотической 16S рРНК. Также на конец продуктов ПЦР 16S рДНК добавляли линкер с индексом для NGS-секвенирования. Секвенирование было заказано в компании GENEWIZ (Сучжоу, Китай).
Статистический анализ
Результаты анализировали и обрабатывали в Microsoft Excel 2007 с использованием программного обеспечения SPSS (версия 20.0). Сначала был проведен многомерный дисперсионный анализ (MANOVA) для проверки наличия значимого влияния независимых переменных (время отбора проб, глубина почвы и режимы земледелия) по крайней мере на один из физико-химических параметров почвы. Затем был выполнен однофакторный дисперсионный анализ (ANOVA) с использованием критериев Тьюки и Дункана для различных почвенных параметров для оценки значимого влияния времени отбора проб, глубины почвы и режимов земледелия, при этом различия в средних значениях при p ≤ 0,05 считались значимыми. Корреляцию между различными физико-химическими параметрами почвы и численностью микроорганизмов на основе содержания PLFA проводили с использованием программного обеспечения Canoco 4.5. Также был проведен корреляционный анализ Пирсона (при p ≤ 0,01 и p ≤ 0,05) между различными физико-химическими параметрами почвы; между филумами микроорганизмов и физико-химическими свойствами почвы; а также между классами микроорганизмов и физико-химическими свойствами почвы. Кроме того, на основе результатов кластеризации OTU были рассчитаны индексы альфа-разнообразия, включая индекс Chao, индекс ACE, индекс Симпсона и индекс Шеннона.
MANOVA-анализ проводился для проверки дисперсионного анализа различных почвенных параметров, который может отражать уровни значимости различий различных почвенных параметров в разное время отбора проб, на разных глубинах и при разных режимах земледелия. Были выбраны значения четырех тестов MANOVA (лямбда Уилкса, Лоули-Хотеллинга, след Пиллая и наибольший корень Роя), при этом уровни различий между группами соответствуют значениям F (Matheron, 1963).
Результаты
Физико-химические параметры
Было обнаружено, что 14-летние (2002–2016) долгосрочные агротехнические практики в некоторой степени изменили физико-химические свойства почвы (Таблица 1). Было установлено, что почва при всех трех режимах имеет нейтральную или слабощелочную реакцию (pH = 7,3–8,1). Различия оказались незначительными (при p ≥ 0,05). Образцы почвы, отобранные на глубине 0–20 см в октябре и декабре, показали более низкие значения pH, чем образцы, отобранные на глубине 20–40 см. На глубине 0–20 см ORF показал увеличение pH почвы, тогда как на глубине 20–40 см традиционное земледелие повышало pH почвы, за ним следовали органический и LOF-режимы. В целом результаты показали увеличение pH почвы с увеличением глубины: низкий pH наблюдался в образцах почвы, отобранных с глубины 0–20 см, в то время как высокий pH был зарегистрирован в образцах, взятых с глубины 20–40 см.
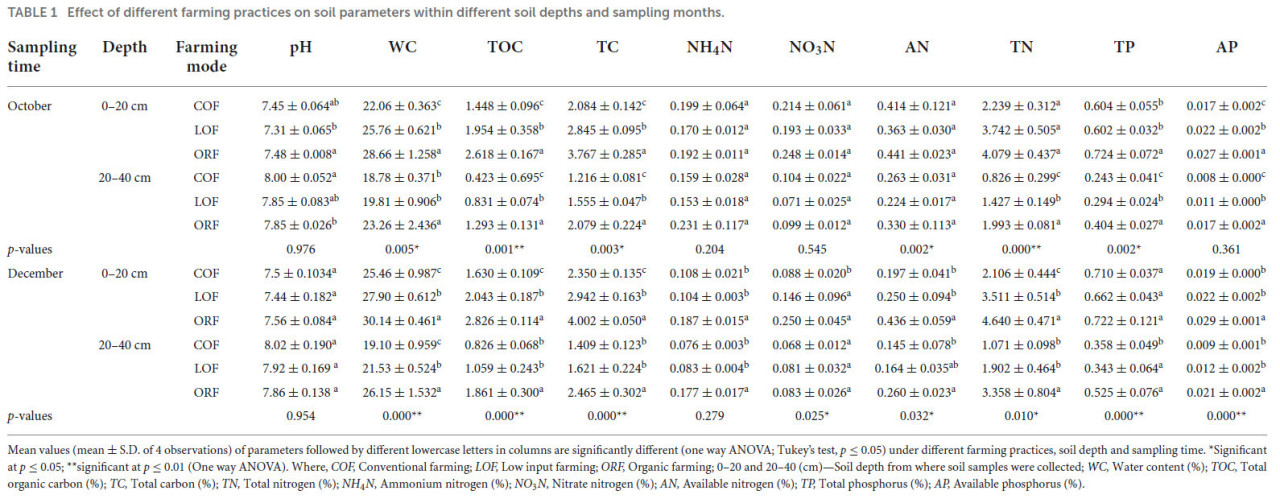
ТАБЛИЦА 1 Влияние различных агротехнических практик на параметры почвы на разных глубинах и в разные месяцы отбора проб.
Средние значения (среднее ± стандартное отклонение 4 наблюдений) параметров, за которыми следуют разные строчные буквы в столбцах, значительно различаются (однофакторный дисперсионный анализ; критерий Тьюки, p ≤ 0,05) при различных агротехнических практиках, глубине почвы и времени отбора проб. Значимо при p ≤ 0,05; значимо при p ≤ 0,01 (однофакторный дисперсионный анализ). Где: COF — традиционное земледелие; LOF — низкозатратное земледелие; ORF — органическое земледелие; 0–20 и 20–40 (см) — глубина почвы, с которой были взяты образцы; WC — влажность (%); TOC — общий органический углерод (%); TC — общий углерод (%); TN — общий азот (%); NH₄N — аммонийный азот (%); NO₃N — нитратный азот (%); AN — доступный азот (%); TP — общий фосфор (%); AP — доступный фосфор (%).
Различия по влажности почвы были значительными как в декабрьских, так и в октябрьских образцах. Влажность при органическом, низкозатратном и традиционном режимах составляла 30–43, 24–38 и 23–34% соответственно. Влажность при традиционном земледелии была значительно ниже по сравнению с другими системами. Изменение влажности на глубине 0–20 см было значительно выше, чем на глубине 20–40 см при том же режиме в тот же месяц отбора проб с диапазоном увеличения от 21 до 28%. Содержание TOC и TC в почве при низкозатратном и органическом режимах значительно увеличилось после длительного применения биокомпоста. Содержание TOC и TC в образцах почвы было значительно выше при органическом земледелии, за которым следовали низкозатратный и традиционный режимы. Содержание обеих форм углерода было выше в декабрьских образцах, отобранных с глубины 0–20 см, в то время как октябрьские образцы с глубины 0–20 см показали более низкое содержание.
Содержание аммонийного азота (NH₄N) в почве было незначительно выше при органическом земледелии и самым низким при традиционном и низкозатратном режимах, а содержание на глубине 0–20 см было выше, чем в слое 20–40 см. Было обнаружено, что содержание NH₄N при традиционном и низкозатратном режимах было выше в октябрьских образцах по сравнению с декабрьскими. При всех трех режимах изменение содержания NO₃N на глубине 0–20 см было выше, чем на глубине 20–40 см. Содержание NO₃N в образцах почвы, отобранных в декабре с глубины 0–20 см, следовало значимому тренду: ORF > низкозатратное земледелие > традиционное земледелие, тогда как на глубине 20–40 см тренд наблюдался следующим: традиционное земледелие > ORF > низкозатратное земледелие (значимо при p ≤ 0,05). Содержание AN в образцах почвы, отобранных с глубины 0–20 см, не показало значимых различий между органическим и низкозатратным режимами; однако оно было значительно выше, чем при традиционном режиме. Октябрьские образцы почвы с глубины 20–40 см показали изменение тренда содержания AN при ORF, которое было значительно выше, чем при традиционном и низкозатратном режимах. В целом результаты показали, что изменение содержания AN на глубине 0–20 см было значительно выше, чем на глубине 20–40 см, и в октябрьских образцах содержание AN было выше по сравнению с декабрьскими. Содержание TN в декабрьских образцах было выше, чем в октябрьских. На глубине 0–20 см значимых различий в содержании TN между органическим и низкозатратным режимами не наблюдалось, однако содержание было выше, чем при традиционном типе земледелия. Изменение содержания TN при всех трех режимах следовало тренду: ORF > низкозатратное земледелие > традиционный режим. Изменение содержания TN на глубине 0–20 см было выше, чем на глубине 20–40 см, и содержание TN было значительно выше в декабрьских образцах по сравнению с октябрьскими. Изменение содержания общего фосфора (TP) в почве было значительно выше на глубине 0–20 см по сравнению с глубиной 20–40 см. Содержание TP в декабрьских образцах было выше, чем в октябрьских. Образцы почвы, отобранные с глубины 0–20 см, показали содержание TP в тренде: ORF > традиционное земледелие > низкозатратное земледелие, в то время как образцы с глубины 20–40 см показали тренд: ORF > низкозатратное земледелие > традиционный режим. Изменение содержания AP в почве, отобранной на полях при трех режимах, следовало тренду: ORF > низкозатратное земледелие > традиционный режим. Содержание AP было значительно выше в октябрьских образцах, чем в декабрьских, и на глубине 0–20 см AP был выше, чем на глубине 20–40 см. Можно заметить, что применение органических удобрений значительно увеличило общее содержание фосфора и доступного фосфора в почве при органическом режиме.
MANOVA-анализ показал эффект взаимодействия между независимыми переменными (время отбора проб глубина почвы; время отбора проб режимы земледелия; глубина почвы режимы земледелия; и время отбора проб глубина почвы режимы земледелия) на зависимые переменные (pH, WC, TC, TOC, NH₄N, NO₃N, AN, TN, TP и AP). Описательные результаты MANOVA приведены в Таблице 2. Значения следа Пиллая, следа Хотеллинга и наибольшего корня Роя были значимы при p ≤ 0,01, где значение следа Хотеллинга было почти равно значению следа Пиллая, что указывает на значимое взаимодействие независимых переменных с некоторыми из зависимых переменных. Значения следа Хотеллинга и наибольшего корня Роя также были схожи, что также указывает на значимый эффект взаимодействия независимых переменных с некоторыми зависимыми переменными. Сгенерированный результат показывает, что существует значимый эффект взаимодействия между глубиной почвы и режимами земледелия на WC (F = 8,800; p = 0,001), TC (F = 10,721; p = 0,000), TN (F = 7,303; p = 0,002), TP (F = 5,942; p = 0,006) и AP (F = 5,396; p = 0,009), тогда как pH (F = 1,389; p = 0,262), TOC (F = 1,337; p = 0,275), NH₄N (F = 1,280; p = 0,290), NO₃N (F = 0,834; p = 0,443) и AN (F = 1,562; p = 0,223) были незначительно затронуты взаимодействием. Эффект взаимодействия глубина почвы время отбора проб показал значимое влияние на содержание NO₃N (F = 4,390; p = 0,043), тогда как все остальные параметры почвы показали незначимый эффект. Кроме того, не было обнаружено значимого эффекта взаимодействия различных режимов земледелия время отбора проб и режимов земледелия время отбора проб глубина почвы ни на один из параметров почвы.
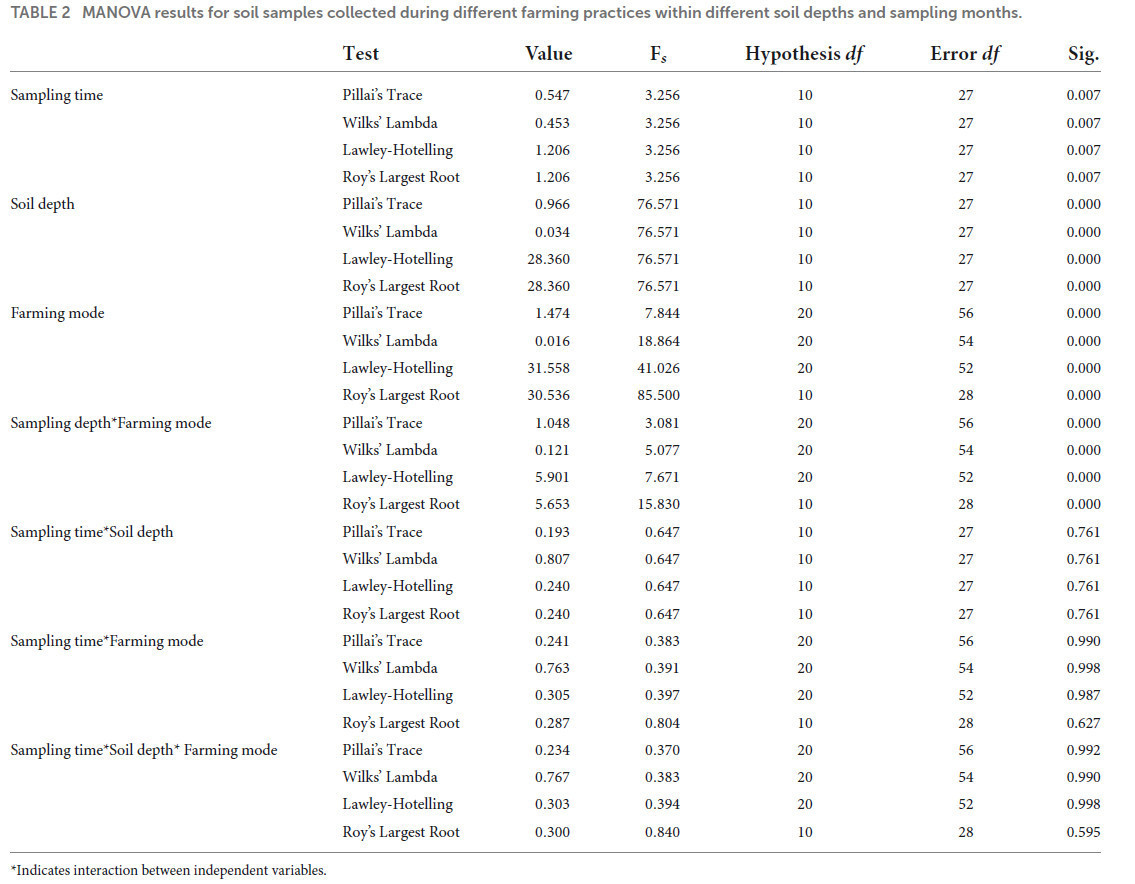
ТАБЛИЦА 2 Результаты MANOVA для образцов почвы, отобранных при различных агротехнических практиках, на разных глубинах и в разные месяцы.
Указывает на взаимодействие между независимыми переменными.
Численность и разнообразие почвенных микроорганизмов
Фосфолипидные жирные кислоты
PLFA-анализ выявил биомаркеры для 33 микроорганизмов в протестированных образцах почвы, из которых для дальнейшего исследования были отобраны только 22 биомаркера с содержанием более 0,1% (дополнительные таблицы 1, 2). Было обнаружено, что маркеры PLFA почвенных микроорганизмов при различных режимах земледелия значительно различались и были полностью распределены в пределах глубин 0–20 и 20–40 см (14:0, 16:0, 18:0, 15:0 anteiso, 15:0 iso, 16:0 10-methyl, 16:0 iso, 16:1 2OH, 17:0 anteiso, 17:0 cyclo, 17:0 iso, 17:1 iso w9c и 19:0 cyclo w8c) и т. д., в то время как некоторые биомаркеры, такие как 15:0, 17:0, 20:0, 16:1 w7c, 17:0 10-methyl, 18:0 10-methyl TBSA, 18:1 w9c, 18:3 w6c (6, 9, 12) и 20:4w6, 9c, не были распределены. Содержание биомаркеров PLFA в образцах с глубины 0–20 см было значительно выше, чем в образцах с глубины 20–40 см. Образцы почвы, отобранные в декабре, показали больше микроорганизмов с биомаркером 16:1 w7c, в то время как октябрьские образцы показали микроорганизмы с биомаркерами 15:0, 17:0, 20:0, 18:0 10-methyl TBSA, 18:3 w6c (6,9,12) и 20:4w6.
Наиболее распространенными биомаркерами PLFA, содержание которых превышало 5%, оказались 16:0 (грамположительные бактерии), за которыми следовали 19:0 cyclo w8c (грамотрицательные бактерии), 16:0 10-methyl (актиномицеты), 15:0 iso (грамположительные бактерии) и 17:1 iso w9c (грамотрицательные бактерии) (Рисунок 1). Однако содержание PLFA у пяти зарегистрированных типов микроорганизмов варьировало при различных режимах земледелия и следовало тренду: ORF > LOF > COF. В сообществе грибов на глубине 20–40 см не было зарегистрировано, в то время как в октябрьских образцах почвы содержание PLFA у грамположительных бактерий и актиномицетов было самым высоким при LOF и ORF и самым низким при COF. Содержание PLFA различных микроорганизмов было выше при ORF, за которым следовали COF и LOF в образцах почвы, отобранных в декабре. В целом результаты показали, что численность грамположительных и грамотрицательных бактерий была значительно выше при органическом режиме, за которым следовали низкозатратный и традиционный режимы. Общее содержание PLFA в почве может косвенно отражать общее количество почвенных микроорганизмов. Видно, что характер изменения общего содержания PLFA в почве при различных режимах земледелия следовал тренду: органический режим > LOF > традиционный режим (Рисунок 2). Изменение общего содержания PLFA в почве было наибольшим на глубине 0–20 см по сравнению с глубиной 20–40 см. ORF-практика показала значительное увеличение общего содержания PLFA в октябре по сравнению с декабрем. При традиционном режиме общее содержание PLFA было выше в декабре по сравнению с октябрем, в то время как LOF показал незначительное различие в общем содержании PLFA в оба периода отбора проб.
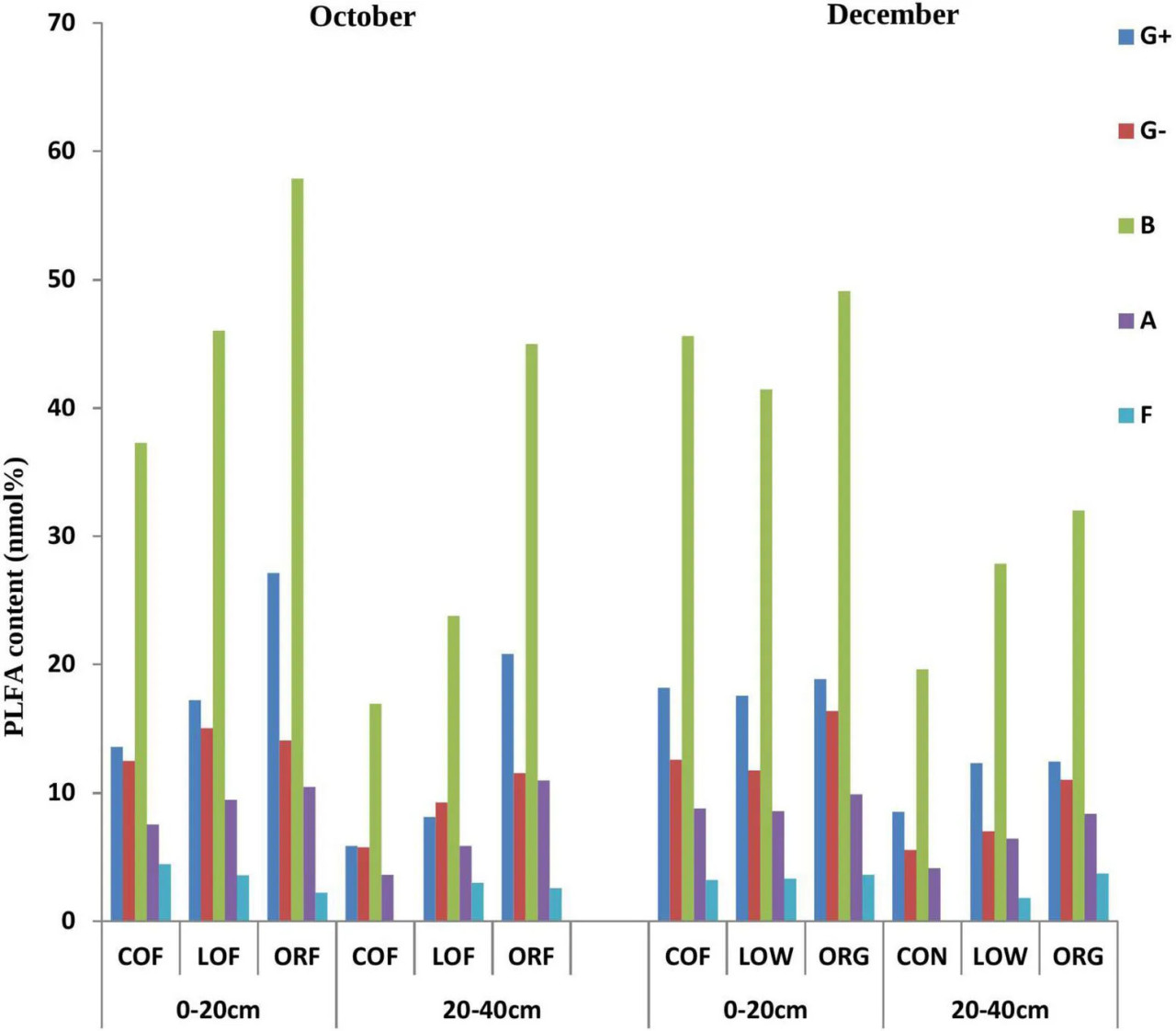
РИСУНОК 1 Влияние различных агротехнических практик на содержание PLFA различных типов почвенных микроорганизмов на разных глубинах и в разные месяцы отбора проб. Где: G+ — грамположительные бактерии; G– — грамотрицательные бактерии; B — бактерии; A — актиномицеты; F — грибы; October и December — месяцы отбора проб почвы; PLFA — фосфолипидные жирные кислоты; COF — традиционное земледелие; LOF — низкозатратное земледелие; ORF — органическое земледелие; 0–20 и 20–40 (см) — глубина почвы, с которой были взяты образцы.
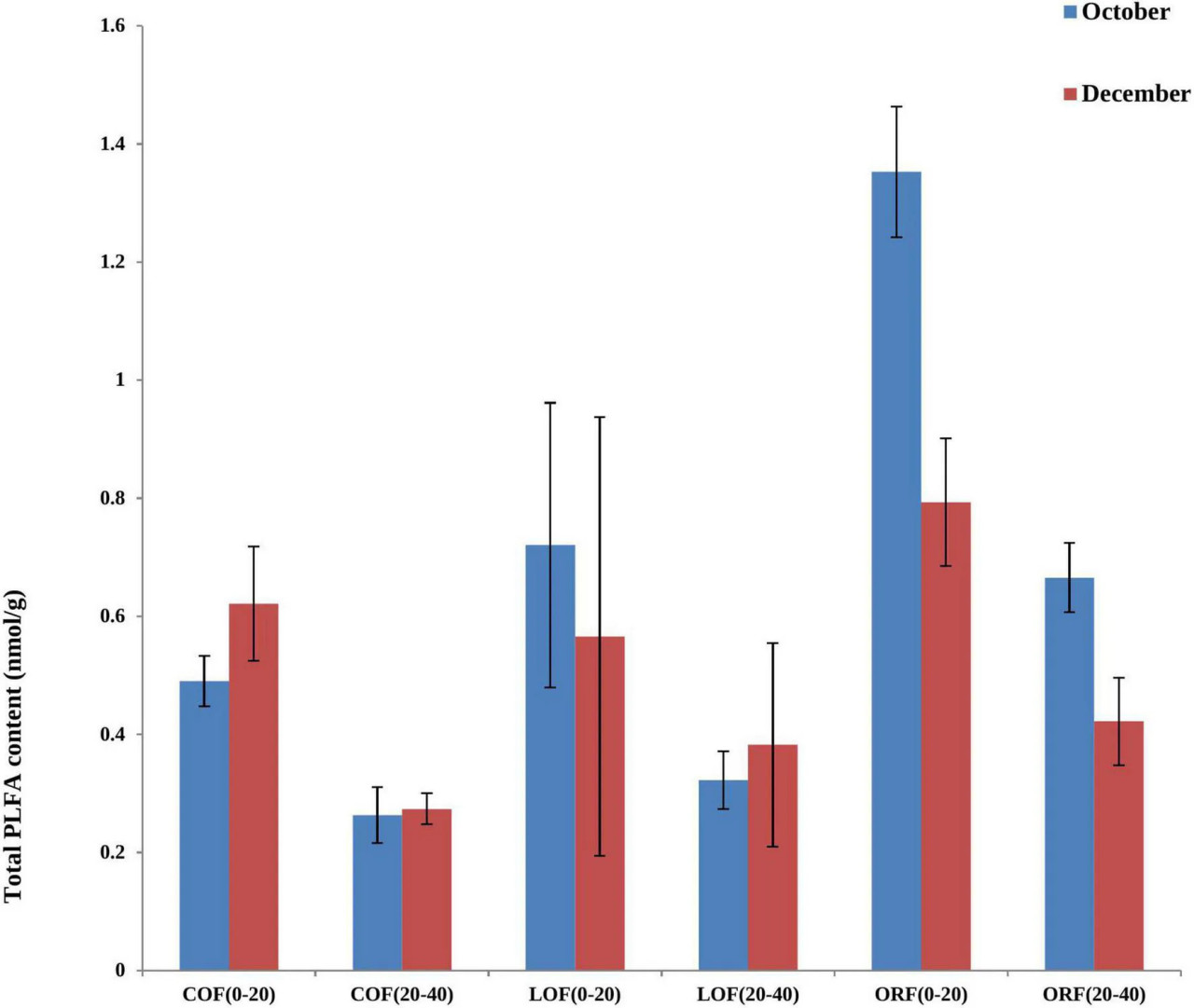
РИСУНОК 2 Влияние различных агротехнических практик на общее содержание PLFA в почве на разных глубинах и в разные месяцы отбора проб. Где: October и December — месяцы отбора проб почвы; PLFA — фосфолипидные жирные кислоты; COF — традиционное земледелие; LOF — низкозатратное земледелие; ORF — органическое земледелие; 0–20 и 20–40 (см) — глубина почвы, с которой были взяты образцы.
Технология высокопроизводительного секвенирования ДНК
С помощью высокопроизводительного секвенирования ДНК было получено 29 689 сортируемых операционных таксономических единиц (OTU) со степенью сходства последовательностей 97%. OTU выявили микроорганизмы из 2 царств, 27–30 филумов, 62–67 классов, 72–81 порядков, 129–143 семейств, 154–186 родов и 15–20 видов (дополнительная таблица 3). Было обнаружено, что при различных режимах земледелия, глубинах почвы и времени отбора проб наблюдалось небольшое влияние на численность и разнообразие почвенных микроорганизмов на различных таксономических уровнях, но сообщалось о различиях в структурном распределении микроорганизмов. В настоящем исследовании относительная численность и разнообразие микроорганизмов изучались на уровне филума и класса. На уровне филума в метагеномных образцах было зарегистрировано 30 филумов, причем на неклассифицированные филумы приходилось 0,01–0,08%. Была отобрана относительная численность более 0,1% (Рисунок 3). Рисунок 3 построен для 11 филумов, которые показали, что Proteobacteria являются доминирующим филумом, за которым следуют Acidobacteria, Bacteroidetes, Actinobacteria, Chloroflexi, Gemmatimonadetes, Firmicutes и т. д., на которые приходилось 24–44%, 19–30%, 6–23%, 3,8–6,2%, 3,8–8,3%, 0,9–4,3%, 0,7–2,2% от общего содержания микроорганизмов соответственно. Среди них Proteobacteria, Acidobacteria и Bacteroidetes показали более высокую относительную численность — 95%, за которыми следовали Actinobacteria, Chloroflexbacteria и т. д. Было обнаружено, что относительная численность различных филумов незначительно различалась на разных глубинах (0–20 и 20–40 см), но значительные различия в численности микроорганизмов наблюдались в ответ на различные режимы земледелия и месяцы отбора проб.
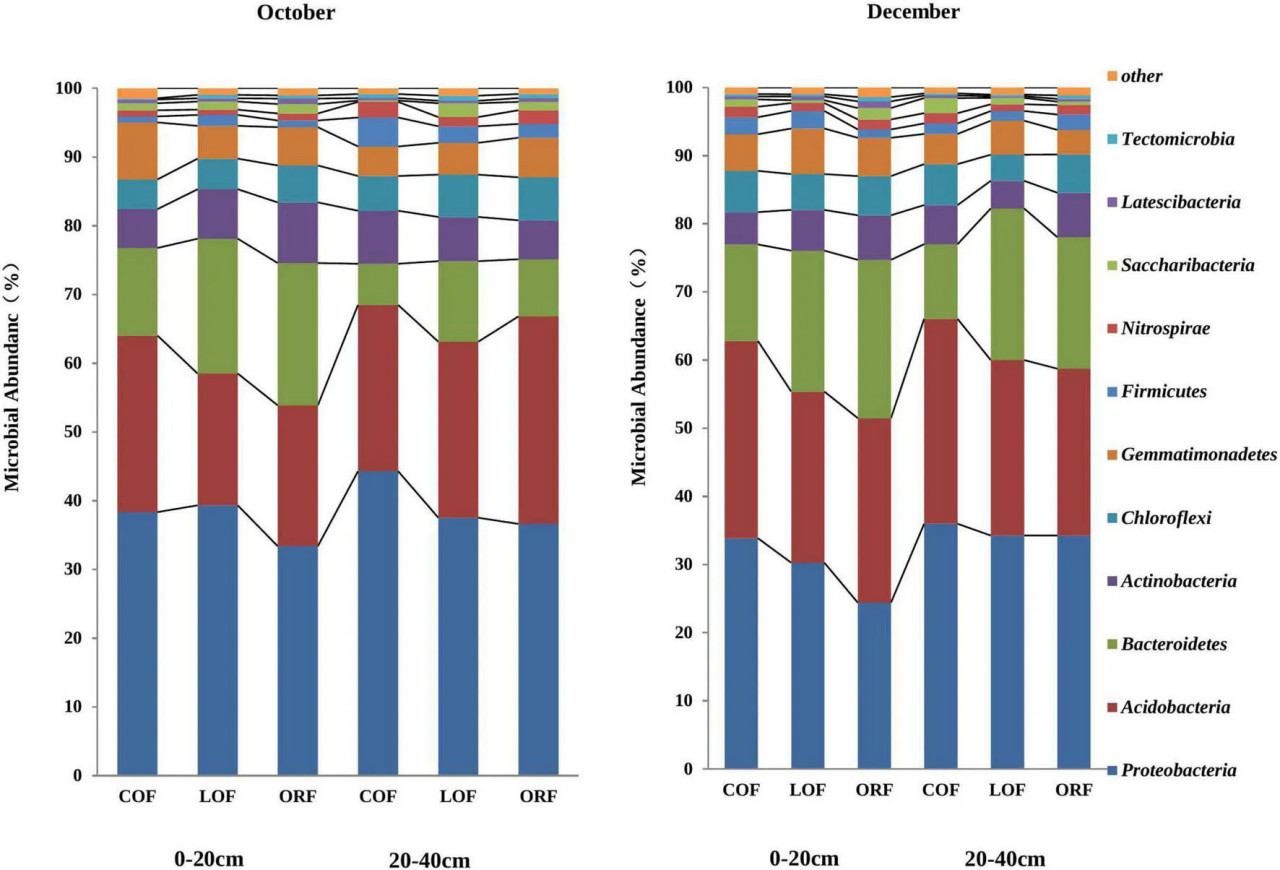
РИСУНОК 3 Влияние различных агротехнических практик на численность почвенных микроорганизмов на уровне классов на разных глубинах и в разные месяцы отбора проб. Где: October и December — месяцы отбора проб почвы; COF — традиционное земледелие; LOF — низкозатратное земледелие; ORF — органическое земледелие; 0–20 и 20–40 (см) — глубина почвы, с которой были взяты образцы.
Относительная численность Proteobacteria в октябрьских образцах почвы на глубине 0–20 см была значительно выше при LOF и немного ниже при традиционном и органическом режимах, тогда как численность Acidobacteria была значительно выше при традиционном режиме и ниже при LOF и органическом. Численность Bacteroidetes была значительно выше на глубине 0–20 см, чем на глубине 20–40 см. Относительная численность Actinobacteria в образцах почвы, отобранных с глубины 0–20 см, следовала тренду: органический режим > LOF > традиционный режим, тогда как на глубине 20–40 см тренд наблюдался: традиционный режим > LOF > органический режим. Относительная численность Chloroflexbacteria на глубине 20–40 см была значительно выше, чем на глубине 0–20 см, наибольшая при традиционном земледелии, за которым следовали органический и LOF-режимы. В декабре относительная численность Proteobacteria и Acidobacteria была наибольшей при традиционном режиме, за которым следовали LOF и ORF. Численность Bacteroidetes на глубине 0–20 см была значительно выше при органическом режиме, в то время как на глубине 20–40 см относительная численность была наибольшей при LOF по сравнению с органическим и традиционным режимами. Численность Actinobacteria и Chloroflexbacteria на глубине 20–40 см не показала значимых различий между режимами, в то время как на глубине 0–20 см численность актиномицетов была наибольшей при органическом режиме, за которым следовали LOF и традиционный режимы. Численность Chloroflexi не была затронута ни одним из режимов, в то время как филум Bacillus показал значительно более высокую численность при LOF по сравнению с двумя другими режимами.
На уровне класса в метагеномных образцах насчитывалось 67 классов бактерий, а относительная численность неклассифицированных бактериальных классов составляла 11,91–22,59% (Рисунок 4). Рисунок 4 построен для 11 классов микроорганизмов с численностью более 0,1%, и основными доминирующими классами были Alphaproteobacteria, Gammaproteobacteria, Sphingobacteriia, Blastocatellia, Betaproteobacter, Actinobacteria, Deltaproteobacteria, Gemmatimonadetes, Acidimicrobiia и Nitrospira. Среди них численность Alphaproteobacteria, Gammaproteobacteria, Sphingobacteria и Blastocatellia была относительно высокой, составляя 40–60% от общего микробного сообщества. Кроме того, относительная численность классов грибов показала явные различия при различных режимах земледелия на разных глубинах и в разные месяцы отбора проб. Относительная численность Alphaproteobacteria и Blastocatellia была выше при традиционном режиме, в то время как присутствие Sphingobacteria на глубине 0–20 см было выше при LOF, а на глубине 20–40 см численность была выше при традиционном земледелии. Дополнительная таблица 4 показала, что при уровне сходства 97% результаты секвенирования, охват созданной микробной библиотеки во всех образцах почвы составлял от 99,4 до 99,6%, что может достоверно отражать OTU-состав микробного сообщества образцов, и информация имеет высокую степень достоверности, что представляет высокую исследовательскую значимость и практическую ценность.
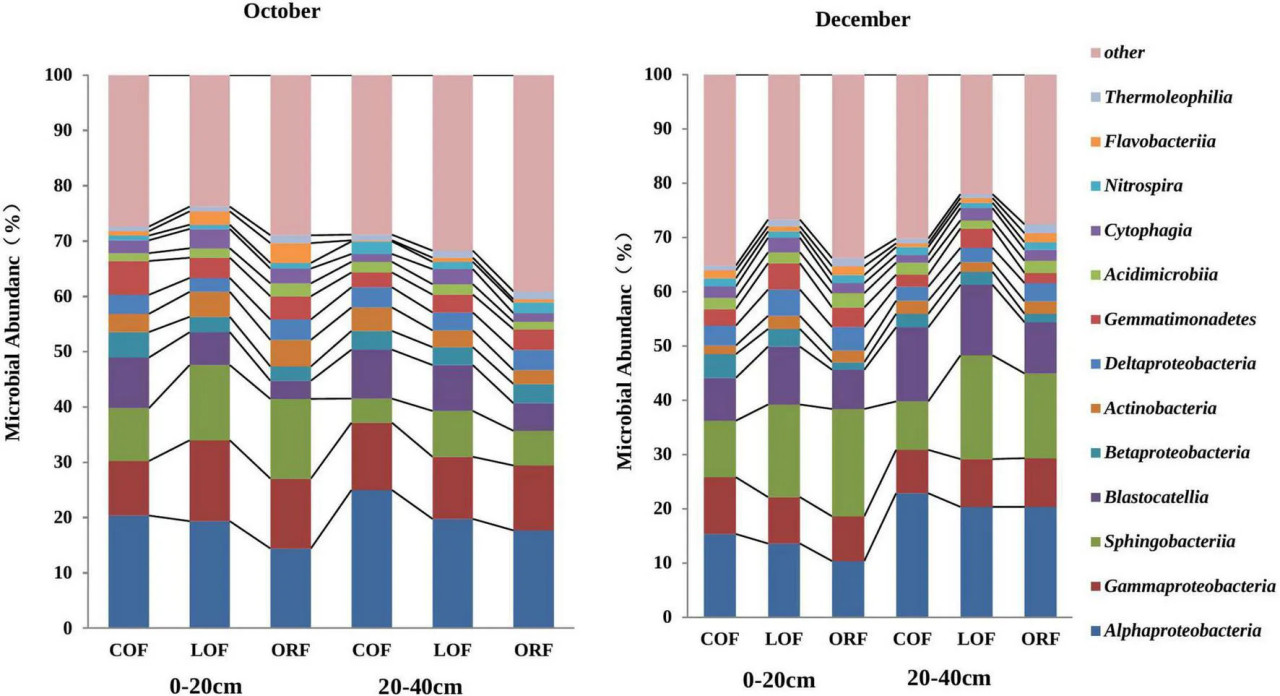
РИСУНОК 4 Влияние различных агротехнических практик на численность почвенных микроорганизмов на уровне филумов на разных глубинах и в разные месяцы отбора проб. Где: October и December — месяцы отбора проб почвы; COF — традиционное земледелие; LOF — низкозатратное земледелие; ORF — органическое земледелие; 0–20 и 20–40 (см) — глубина почвы, с которой были взяты образцы.
Было обнаружено, что индексы богатства ACE, индекс Чао и индекс разнообразия Шеннона микробного сообщества зависели от различных режимов земледелия в разные периоды отбора проб, в то время как индекс Симпсона не изменился (дополнительная таблица 4). В октябре все три индекса были выше при LOF, в то время как низкий индекс богатства Чао и ACE наблюдался при органическом и традиционном режимах. Индекс разнообразия Шеннона был выше при ORF по сравнению с традиционным режимом. На поздней стадии производства сельдерея (декабрь) индекс богатства Чао и ACE и индекс разнообразия Шеннона микробного сообщества не показали влияния режимов земледелия, что может быть связано с культурой сельдерея и ее влиянием на численность почвенных микроорганизмов.
Корреляционный анализ Пирсона
Корреляционный анализ Пирсона выявил значимую корреляцию почти между всеми физико-химическими параметрами образцов почвы, за исключением pH, NH₄N, NO₃N и TP, которые показали незначимую корреляцию с другими параметрами почвы. Были обнаружены значительные сильные положительные корреляции между AP и TOC (r = 0,969) и TC (r = 0,962) при p ≤ 0,01, в то время как содержание TN показало сильную положительную значимую корреляцию с органическим углеродом (r = 0,913, p ≤ 0,01). Другие параметры, такие как TOC-WC (r = 0,941); TC-WC (r = 0,936); TC-TOC (r = 0,989); TN-TOC (r = 0,931); TN-TC (r = 0,905); AP-WC (r = 0,944); NO₃N-AN (r = 0,919), также показали сильную значимую положительную корреляцию друг с другом при уровне значимости p ≤ 0,01.
Также была проведена корреляция между различными физико-химическими параметрами почвы и группами почвенных микроорганизмов на основе содержания PLFA с использованием программного обеспечения Canoco 4.5, RDA-анализ (Рисунок 5). Общая объясненная доля двух главных компонент в два периода достигла 99,9%. Среди них первая главная компонента (PC1) объяснила 93,4 и 86,1% переменных, а вторая главная компонента (PC2) объяснила 6,5 и 13,8% переменных. В октябре наиболее значимая корреляция наблюдалась между большинством типов бактерий и AP почвы, общим доступным азотом, TOC и влажностью почвы. Численность микробной популяции отрицательно коррелировала с pH. Наблюдалась значительная положительная корреляция между G+ бактериями, G– бактериями, грибами, актиномицетами и общим количеством бактерий, и все они находились на отрицательном конце второй главной компоненты, а корреляции между различными микробными популяциями достигали значительного и высокозначимого уровня. В декабре наблюдалась значительная положительная корреляция между G+ и G– бактериями, грибами, актиномицетами и общим количеством бактерий, и они также находились на отрицательном конце первой и второй главных компонент, а корреляции между различными микробными популяциями достигали значительного и высокозначимого уровня. Бактериальная популяция, G+ бактерии, G– бактерии, актиномицеты и грибы положительно коррелировали с TOC, TC, TN, нитратным азотом, общим доступным азотом, TP, AP и влажностью, среди которых корреляция с TN, TP и TOC была наиболее значимой. Соотношение G+ к G– бактериям было высокозначимо и отрицательно коррелировало с аммонийным азотом почвы и положительно коррелировало с pH почвы.
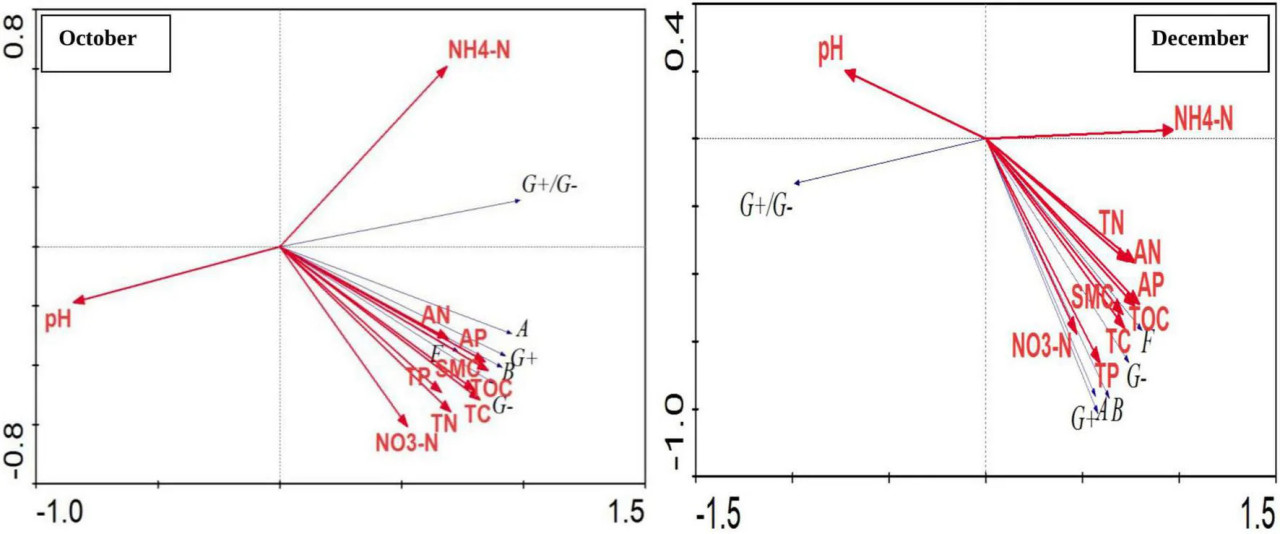
РИСУНОК 5 Корреляция между различными физико-химическими параметрами почвы и группами почвенных микроорганизмов на основе содержания PLFA. Где: October и December — месяцы отбора проб почвы; G+ — грамположительные бактерии; G– — грамотрицательные бактерии; B — бактерии; A — актиномицеты; F — грибы; WC — влажность (%); TOC — общий органический углерод (%); TC — общий углерод (%); TN — общий азот (%); NH₄N — аммонийный азот (%); NO₃N — нитратный азот (%); AN — доступный азот (%); TP — общий фосфор (%); AP — доступный фосфор (%).
Таблица 3 показывает корреляционный анализ между различными физико-химическими свойствами почвы и филумами микроорганизмов. Присутствие Proteobacteria, Bacteroidetes и Latescibacteria показало значимые корреляции с большинством физико-химических свойств почвы, где Proteobacteria отрицательно коррелировали с WC, TOC, TC и AP при уровне значимости p ≤ 0,01, а с TN и TP при p ≤ 0,05. Сообщество Bacteroidetes положительно коррелировало с WC, TOC, TC, TN и AP при уровне значимости p ≤ 0,01 и с TP при p ≤ 0,05. Популяция Actinobacteria положительно коррелировала с AN при уровне значимости p ≤ 0,01, в то время как присутствие Gemmatimonadetes отрицательно коррелировало с pH почвы при p ≤ 0,05. Присутствие Latescibacteria положительно коррелировало с WC, TOC, TC, TN, NO₃N, AP и AN при p ≤ 0,01, тогда как корреляция с TP достигала уровня значимости p ≤ 0,05. Индекс Чао отрицательно коррелировал с присутствием Latescibacteria. Таблица 4 представляет корреляционный анализ между различными физико-химическими свойствами почвы и классами микроорганизмов. Присутствие Alphaproteobacteria показало значительную отрицательную корреляцию с WC, TOC, TC, TN, TP и AP при уровне значимости p ≤ 0,01 и с NO₃N при p ≤ 0,05, в то время как pH почвы показал значительную положительную корреляцию при p ≤ 0,05. Популяция Sphingobacteriia положительно коррелировала с TN при уровне значимости p ≤ 0,01 и с WC, TOC, TC и AP при уровне p ≤ 0,05. Присутствие Blastocatellia отрицательно коррелировало с NO₄N и AN при уровне значимости p ≤ 0,01 и с AP и индексом Шеннона при уровне p ≤ 0,05. Deltaproteobacteria отрицательно коррелировали с индексом богатства Чао, в то время как присутствие Gemmatimonadetes отрицательно коррелировало с pH почвы при уровне значимости p ≤ 0,05. Присутствие Cytophagia отрицательно коррелировало с pH почвы и индексом Шеннона при p ≤ 0,01, тогда как Nitrospira положительно коррелировала с pH почвы при уровне значимости p ≤ 0,05. Популяция Flavobacteriia положительно коррелировала с TOC, TC, TN, AP (при p ≤ 0,01), WC, NO₃N, TP и индексом Шеннона (при p ≤ 0,05) и отрицательно коррелировала с pH почвы при p ≤ 0,05.
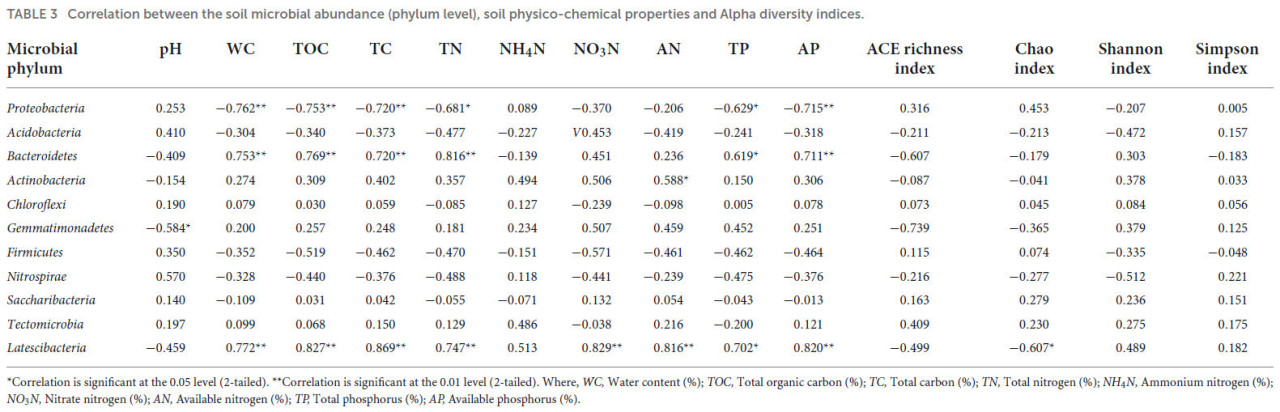
ТАБЛИЦА 3 Корреляция между численностью почвенных микроорганизмов (уровень филума), физико-химическими свойствами почвы и индексами альфа-разнообразия.
Корреляция значима на уровне 0,05 (двухсторонняя). Корреляция значима на уровне 0,01 (двухсторонняя). Где: WC — влажность (%); TOC — общий органический углерод (%); TC — общий углерод (%); TN — общий азот (%); NH₄N — аммонийный азот (%); NO₃N — нитратный азот (%); AN — доступный азот (%); TP — общий фосфор (%); AP — доступный фосфор (%).
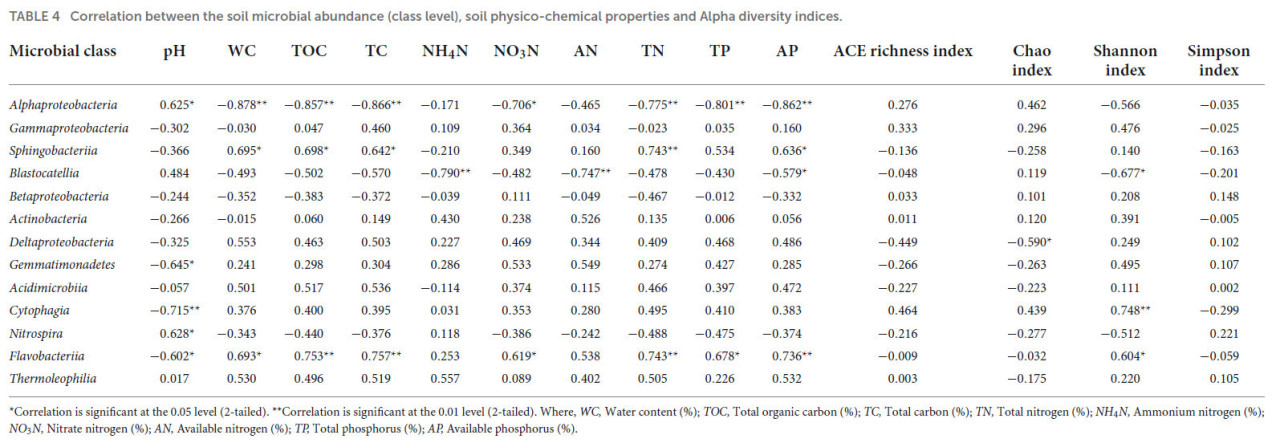
ТАБЛИЦА 4 Корреляция между численностью почвенных микроорганизмов (уровень класса), физико-химическими свойствами почвы и индексами альфа-разнообразия.
Корреляция значима на уровне 0,05 (двухсторонняя). Корреляция значима на уровне 0,01 (двухсторонняя). Где: WC — влажность (%); TOC — общий органический углерод (%); TC — общий углерод (%); TN — общий азот (%); NH₄N — аммонийный азот (%); NO₃N — нитратный азот (%); AN — доступный азот (%); TP — общий фосфор (%); AP — доступный фосфор (%).
Обсуждение
В настоящем исследовании pH почвы был слабощелочным, при этом щелочность почвы увеличивалась с глубиной при органическом земледелии. Аналогично, Kwiatkowski and Harasim (2020) сообщили о высоком pH в органически управляемой почве по сравнению с другими традиционными режимами. Органический материал, вносимый в почву, может повышать буферную способность почвы и хелатирование Al³⁺, что приводит к увеличению pH почвы (Baolan et al., 2014). Изменчивость pH на разных глубинах может быть связана с усилением нисходящего движения солей почвенных оснований в пределах почвенного профиля из-за выщелачивания (Assen et al., 2005) и повышенным содержанием карбонат-ионов и меньшей скоростью выветривания в почве (Malo et al., 2005). Влажность почвы может влиять на pH, изменяя химическое равновесие почвы и изменяя формы и эффективность питательных веществ (Morales et al., 2011). pH почвы может увеличиваться из-за внесения органических остатков (Manna et al., 2005; Huang et al., 2016) или из-за сохранения растительных остатков (Liu et al., 2010; Butterly et al., 2011). В органически обрабатываемой почве влажность была выше в верхнем слое почвы по сравнению с более глубоким. Pimentel et al. (2005) также наблюдали, что применение органических удобрений может увеличить водоудерживающую способность почвы. Наличие высокого содержания органического вещества в почве может создавать наземный покров, который помогает снизить температуру почвы, что приводит к уменьшению испарения воды (Calvino and Monzon, 2009). Hathaway-Jenkins et al. (2011) в своем исследовании сообщили о высокой скорости инфильтрации и снижении поверхностного стока в сельскохозяйственных почвах Англии, управляемых в режиме ORF. В одном из недавних исследований Durrer et al. (2021) обнаружили, что органическая обработка почвы может поддерживать высокий запас воды в почве благодаря наличию высокого содержания органического вещества.
Было обнаружено, что внесение органических удобрений быстро увеличивало содержание углерода в почве на глубине 0–20 см, что может быть связано с применением биокомпоста. Для оценки продуктивности любой сельскохозяйственной почвы органический углерод может использоваться как один из наиболее важных показателей (Shukla et al., 2006). Аналогично настоящему исследованию, относительно более высокое содержание OC в верхнем слое почвы (0–20 см) было обнаружено Selassie and Ayanna (2013) в их исследовании, и они объяснили это увеличение относительно более высоким возвратом биомассы почвы на поверхности из-за микробного разложения. Кроме того, Wakene (2001) сообщил о высоком содержании OC и TOC в поверхностной почве, которая является наиболее биологически активным слоем во всем почвенном профиле. Высокое накопление углерода в сельскохозяйственной почве может быть связано с опадом листьев на поверхности, а также с корневыми отложениями в более глубоких слоях почвы (Zhou and Wang, 2017). Настоящие результаты также подтверждаются выводами Mandal et al. (2018) и Kwiatkowski and Harasim (2020). В отличие от настоящего исследования, Sharma et al. (2020) сообщили, что интенсивное возделывание, неправильная обработка почвы и другие методы управления могут увеличить потерю органического углерода почвы из-за высокой микробной активности и процессов окисления. Другая форма углерода, т. е. неорганический углерод почвы (IC), содержит такие соединения, как карбонат-ионы, отложения карбоната кальция и магния в почвенном растворе и CO₂ в почвенном воздухе. В настоящем исследовании неорганический углерод почвы можно рассчитать, вычитая TOC из TC. Было обнаружено, что IC был относительно высоким при ORF по сравнению с LOF и COF. В основном почвы аридного и семиаридного климата имеют высокое содержание IC по сравнению с влажными почвами, которые имеют очень низкое содержание IC из-за длительного периода оборота и химических процессов, которые оказывают незначительное влияние на плодородие почвы (Kowalska et al., 2020).
Долгосрочное органическое земледелие и возделывание овощей увеличили содержание аммонийного азота в верхнем слое (0–20 см) и было самым низким при традиционном и низкозатратном режимах (глубина 20–40 см). Запасы органического азота в почве увеличились из-за снижения выщелачивания азота (Kwiatkowski and Harasim, 2020). Содержание NO₃N, AN и TN в почве было наибольшим в верхнем слое при ORF. Применение органических удобрений может не только улучшить запасы TN в почве, но и оказать определенное влияние на аммонийный азот и компоненты AN. Более высокое поступление органических веществ (опад листьев) может усилить минерализацию азота в почве (Zhou and Wang, 2017). Аналогичные результаты наблюдали Drinkwater et al. (1995) и Wang et al. (2011), которые зарегистрировали более высокое содержание общего N и более высокую скорость минерализации в органически управляемых почвах по сравнению с традиционными. Кроме того, Kwiatkowski and Harasim (2020) в своем исследовании сообщили о высоком содержании TN и NH₄N в верхнем слое сельскохозяйственной почвы и объяснили это типом удобрений, используемых при ORF, которые сильно минерализуются и могут стимулировать высокую ферментативную активность почвы, что приводит к большему высвобождению азота в почве. Они также сообщили о низком содержании NO₃N на органически управляемом поле по сравнению с традиционным, которое показало значительное увеличение с глубиной. В настоящем исследовании было обнаружено, что доступность TN стабильно снижается с увеличением глубины от 0–20 см до 20–40 см при всех режимах, как ранее сообщалось Gong et al. (2005), Geissen and Guzman (2006) и Selassie and Ayanna (2013). Напротив, Emiru and Gebrekidan (2013) сообщили, что непрерывное органическое возделывание может увеличить скорость окисления органического вещества, что может привести к снижению доступности азота для сельскохозяйственных культур.
Содержание TP и AP было наибольшим при органическом режиме на глубине 0–20 см по сравнению с глубиной 20–40 см. В настоящем исследовании количество AP уменьшалось с увеличением глубины. Аналогичные результаты наблюдали Selassie and Ayanna (2013), которые объяснили снижение доступных форм P с увеличением глубины снижением содержания органического вещества в более низких слоях почвы. Как правило, доступность P увеличивается в органически управляемых почвах из-за увеличения органических соединений, которые приводят к образованию органофосфатных комплексов, легко усваиваемых растениями, а также органическое вещество в почве может способствовать максимальному растворению неорганического фосфора, уменьшая его фиксацию и улучшая его доступность в почве (Selassie and Ayanna, 2013). В настоящем исследовании можно заметить, что AP в поверхностной почве (0–20 см) был выше, чем в нижних горизонтах (20–40 см), аналогичные результаты были получены ранее во многих исследованиях (Eshetu et al., 2004; Tuma, 2007). Долгосрочное применение фосфорных удобрений и органических удобрений в почвах показало увеличение доступности P для сельскохозяйственных культур через разложение органического вещества и выделение определенных кислот, способствующих растворению фосфатов кальция (Mohammadi et al., 2009).
Численность и разнообразие почвенных микроорганизмов изучались путем получения биомаркеров для различных микроорганизмов с помощью метода PLFA. В настоящем исследовании наиболее распространенными биомаркерами PLFA были 16:0, 19:0 cyclo w8c и т. д., что указывало на присутствие грамположительных бактерий, грамотрицательных бактерий, актиномицетов и т. д. Общее содержание PLFA в почве показало наибольшее микробное разнообразие при ORF. Durrer et al. (2021) сообщили, что наличие N-содержащих удобрений в виде компоста из птичьего помета и зеленых удобрений может привести к изменениям в структуре бактериального сообщества, что может улучшить круговорот фосфора и углерода. Аналогично, Neha et al. (2022) сообщили, что ORF является эффективным агротехническим методом, который может увеличить общее содержание PLFA в почве и улучшить микробную биомассу почвы. Аналогичные результаты наблюдались в предыдущих исследованиях Zhong et al. (2010), которые изучали долгосрочные эффекты органических и неорганических удобрений на профили PLFA и функциональное разнообразие микроорганизмов и сообщили о значительном увеличении общего количества PLFA почвенных микроорганизмов, таких как бактерии и актиномицеты, при применении как органических, так и неорганических удобрений, в то время как Jingwei (2016) показал, что применение органических и биологических удобрений может улучшить общее содержание PLFA бактерий и грибов, присутствующих в почвенной экосистеме. В отличие от настоящего исследования, Zhang et al. (2010) сообщили о снижении относительного содержания PLFA у актиномицетов и грибов при ORF.
Мы использовали высокопроизводительное секвенирование ДНК гена 16S рРНК для оценки сдвигов в разнообразии и идентификации специфических бактериальных групп, ассоциированных с почвой при органической, традиционной и низкозатратной системах земледелия. Результаты показали 97% сходство последовательностей и некоторое влияние на структурное распределение микроорганизмов при различных режимах земледелия, на разных глубинах и времени отбора проб. Использование областей V3 и V4 для секвенирования гена 16S рРНК рекомендуется, так как они предоставляют точную и адекватную информацию для таксономической классификации бактериальных сообществ (Castelino et al., 2017). Многие ранние исследования показали влияние различных методов ведения сельского хозяйства на почвенное микробное сообщество, которое может увеличиваться, уменьшаться или оставаться стабильным (Reilly et al., 2013; Lupatini et al., 2017). В нашем исследовании мы наблюдали увеличение содержания C и N при ORF, что может привести к увеличению индексов микробного разнообразия, как ранее сообщалось Hartmann et al. (2015). Из 30 зарегистрированных филумов, Proteobacteria были наиболее распространенными, за ними следовали Acidobacteria, Bacteroidetes и т. д., а из 67 классов Alphaproteobacteria, Gammaproteobacteria, Sphingobacteriia и т. д. были доминирующими в районе исследования. Было обнаружено, что на глубине 0–20 см численность микроорганизмов была значительно выше при LOF, за которым следовали органический и традиционный режимы. Shen et al. (2010) также сообщили о значительном влиянии различных режимов удобрения или земледелия на численность и разнообразие почвенных микроорганизмов. Аналогичные результаты наблюдались во многих предыдущих исследованиях Niu et al. (2017), где технология высокопроизводительного секвенирования Illumina MiSeq использовалась для изучения структуры микробного сообщества плато и было обнаружено, что численность Proteobacteria (доминирующий бактериальный филум) была наибольшей, за которой следовали Actinobacteria, Bacteroidetes и Acidobacteria. Аналогично, Megyes et al. (2021) сообщили о значительных изменениях в различных бактериальных и архейных сообществах (в основном ацидотолерантные таксоны, включая Acidobacteria, Solibacter, Bryobacter и т. д.) при долгосрочных агротехнических практиках на полях кукурузы с использованием технологии Illumina MiSeq. Кроме того, Hao et al. (2008) использовали пиросеквенирование 16S рРНК или Illumina MiSeq для изучения влияния различных режимов земледелия на доминирующее бактериальное сообщество и показали высокую относительную численность Acidobacteria, за которой следовали Proteobacteria, Actinobacteria, Bacteroidetes и Gemmatimonadetes. Среди них Proteobacteria были более стабильным филумом в почве по сравнению с Acidobacteria и не подвергались влиянию различных обработок почвы (Yin et al., 2010), что согласуется с результатами настоящего исследования. Fierer et al. (2007) показали высокую численность Acidobacteria в почвах с низким содержанием органического углерода. Lukow et al. (2000) и Nacke et al. (2011) сообщили о доминировании Actinobacteria, Gemmatimonadetes и Bacteroidetes в сельскохозяйственных почвах, в то время как Ivanova et al. (2015) обнаружили большее количество Actinobacteria и Bacillus в почвах, обработанных органическим веществом, что указывает на положительный эффект органического вещества. Wang et al. (2017) использовали технологию высокопроизводительного секвенирования ДНК для обнаружения структуры бактериального сообщества в почве при различных режимах и сообщили об увеличении численности олиготрофных бактерий, т. е. Bacteroidetes, при применении неорганических удобрений, в то время как применение органических удобрений увеличило большое количество эвтрофных бактерий (Proteus).
Все индексы разнообразия, такие как Chao1, ACE и индекс разнообразия Шеннона, были значительно затронуты различными агротехническими практиками в районе исследования. Можно заметить, что применение органических удобрений увеличило относительную численность и разнообразие почвенных микроорганизмов, в то время как комбинация органических и неорганических удобрений оказала более значительное влияние на улучшение численности и разнообразия почвенных микроорганизмов. Результаты Chao1 и ACE показали, что обработка почвы неорганическими удобрениями может значительно снизить бактериальное богатство почвы по сравнению с отсутствием обработки. Аналогично, Liu et al. (2022) сообщили об увеличении индекса разнообразия Шеннона и индекса богатства микробного сообщества почвы при применении биологических комплексных удобрений в течение 25 лет. Kirchmann et al. (2013) обнаружили, что применение неорганических и органических удобрений не оказывало значительного влияния на индекс разнообразия Шеннона бактерий и грибов по сравнению с контрольной почвой, как сообщалось в настоящем исследовании, что может быть связано с адаптацией почвенных микроорганизмов к различным режимам земледелия.
Корреляционный анализ Пирсона выявил значительную положительную связь между всеми изученными параметрами почвы, и аналогичная значимая положительная связь между физико-химическими параметрами почвы наблюдалась Gebreselassie (2002) в почвах Исследовательского центра Адет в Северо-Западной Эфиопии. Различные типы микроорганизмов и AP, TOC, общий доступный азот и влажность почвы показали наиболее значительную корреляцию между собой. Почвы с более высоким содержанием C и N могут благоприятствовать разнообразной микробной популяции, которая в конечном итоге регулирует биогеохимические циклы (Chen Q. et al., 2020; Han et al., 2021). Аналогично настоящему исследованию, Ma et al. (2010) показали, что органическое вещество почвы, азот и AP являются основными источниками питательных веществ для роста микроорганизмов, в то время как Zhong et al. (2010) выявили роль TN, AP и pH почвы, которые могут значительно влиять на профили PLFA и функциональное разнообразие микроорганизмов. На уровне филума корреляция между различными микробными группами и физико-химическими свойствами почвы была значительно различной, где Proteobacteria, Bacteroidetes и Latescibacteria показали значимые корреляции с большинством физико-химических свойств почвы. Proteobacteria и Bacteroidetes включают многие бактерии, способные фиксировать азот (Farag et al., 2017), а Latescibacteria имеют небольшую численность в почве, но могут способствовать поглощению и превращению сахаров, жирных кислот и аминокислот в качестве источников углерода. Микроорганизмы при ORF участвуют в окислительно-восстановительных реакциях и способствуют разложению органического вещества (He et al., 2017). Корреляция между этими филумами и углеродом, азотом и фосфором в почве была значительной (Jones et al., 2009). Аналогично настоящему исследованию, Sun et al. (2015) показали, что численность бактерий положительно коррелировала с питательным статусом почвы, таким как нитрат, TN, TC и AP, однако разнообразие бактерий в основном определялось pH почвы. Кроме того, Niu et al. (2017) показали, что влажность почвы, pH, AN, TN и органический углерод оказывают большое влияние на состав микробных сообществ. Большинство параметров почвы сильно коррелируют с изменениями в микробной популяции района, аналогично отчету Chen X. et al. (2020). Напротив, исследование, основанное на крупномасштабном анализе, показало, что разнообразие почвенных бактерий и грибов в основном зависит от pH почвы или пространственного расстояния (Green et al., 2004), в то время как Acidobacteria показали отрицательную корреляцию с pH почвы (Dong et al., 2014). Наше исследование положительно подтвердило, что ORF может приводить к высокой доступности питательных веществ для почвенной фауны и флоры, способствовать изменениям в бактериальном сообществе и их функциональных аспектах.
Заключение
Результаты настоящего исследования показали, что глубина почвы и режимы земледелия являются ключевыми факторами, влияющими на различные параметры почвы и микробную популяцию. Применение навоза и органических удобрений способствовало значительному повышению плодородия почвы и улучшению питательного статуса почвы за счет увеличения доступности таких питательных веществ, как углерод, азот и фосфор для растений.
PCA-анализ показал высокозначимую корреляцию между большинством типов бактерий (G+ бактерии, G– бактерии, актиномицеты) и AP, общим доступным азотом, TOC и WC. Большинство свойств почвы показали значительную взаимосвязь между филумами и классами микроорганизмов.
При долгосрочном возделывании органических овощей на исследуемой территории наблюдалось увеличение органического азота в почве, что может служить жизнеспособной стратегией для лучшего производства овощей в Северном Китае (район исследования). В заключение, использование экологически чистых методов ведения сельского хозяйства и применение большего количества органических удобрений фермерами должно поощряться правительством. В будущем следует изучить больше технологий на основе NGS для характеристики микробных сообществ и использовать подходящую и экологически безопасную практику управления сельским хозяйством.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти онлайн по адресу: https://www.frontiersin.org/articles/10.3389/fevo.2022.1026771/full#supplementary-material
Ссылки
Assen M., le Roux A. L., Barker C. H., Heluf G. (2005). Soils of jelo micro-catchment in the chercher highlands of eastern ethiopia: I. Morphological and physiochemical properties. Ethopian J. Nat. Res. 7, 55–81. CrossRef. Google Scholar.
Balser T. C., Liang C., Gutknecht J. L. M. (2019). Linking microbial community analysis and ecosystem studies: A rapid lipid analysis protocol for high throughput. Soil Ecol. Lett. 1, 22–32. doi: 10.1007/s42832-019-0003-0. CrossRef. Google Scholar.
Baolan H., Shuai L., Wei W., Lidong S., Liping L., Weiping L. et al. (2014). pH dominated niche segregation of ammonia-oxidising microorganisms in Chinese agricultural soils. FEMS Microbiol. Ecol. 90, 290–299. doi: 10.1111/1574-6941.12391. CrossRef. Google Scholar.
Bligh E. G., Dyer W. J. (1959). A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 37, 911–917. doi: 10.1139/o59-099. CrossRef. Google Scholar.
Bowman R. A. (1988). The rapid method to determine total phosphorus in soils. Soil Sci. Soc. Am. J. 52, 1301–1304. doi: 10.2136/sssaj1988.03615995005200050016x. CrossRef. Google Scholar.
Brinton W. F. (2020). Phospholipid fatty acid (PLFA) analysis: A robust indicator for soil health? Agri. Res. Tech. 24, 118–119. doi: 10.19080/ARTOAJ.2020.24.556281. CrossRef. Google Scholar.
Bunemann E. K., Bongiorno G. Z. G., Creamerb R. E., Deynb G. D., Goedeb R. D., Fleskensd L. et al. (2018). Soil quality -- a critical review. Soil Biol. Biochem. 120, 105–125. CrossRef. Google Scholar.
Butterly C. R., Kaudal B., Baldock J. A., Tang C. (2011). Contribution of soluble and insoluble fractions of agricultural residues to short-term pH changes. Eur. J. Soil Sci. 62, 718–727. doi: 10.1111/j.1365-2389-2011-01387.x. CrossRef. Google Scholar.
Calvino P., Monzon J. (2009). "Farming systems of Argentina: Yield constraints and risk management," in Crop physiology, applications for genetic improvement and agronomy, eds. Sadras V. O., Calderini D. (Cambridge, MA: Academic Press), 55–70. CrossRef. Google Scholar.
Carvalho A. M. D., Bustamante M. M. D. C., Almondes Z. A. D. P., Figueiredo C. C. D. (2014). Forms of phosphorus in an oxisol under different soil tillage systems and cover plants in rotation with maize. Rev. Bras. Ciênc. Solo 38, 972–979. doi: 10.1590/S0100-06832014000300029. CrossRef. Google Scholar.
Castelino M., Eyre S., Moat J., Fox G., Martin P., Hp P. et al. (2017). Optimization of methods for bacterial skin microbiome investigation: Primer selection and comparison of the 454 versus MiSeq platform. BMC Microbiol. 17:23. doi: 10.1186/s12866-017-0927-4. CrossRef. Google Scholar.
Chen M., Li X., Yang Q., Chi X., Pan L., Chen N. et al. (2012). Soil eukaryotic microorganism succession as affected by continuous cropping of peanut pathogenic and beneficial fungi were selected. PLoS One 7:e40659. doi: 10.1371/journal.pone.0040659. CrossRef. Google Scholar.
Chen Q., Ding J., Zhu D., Hu H. W., Delgado-Baquerizo M., Ma Y. B. et al. (2020). Rare microbial taxa as the major drivers of ecosystem multifunctionality in long-term fertilized soils. Soil Biol. Biochem. 141:107686. CrossRef. Google Scholar.
Chen X., Henriksen T. M., Svensson K., Korsaeth A. (2020). Long-term effects of agricultural production systems on structure and function of the soil microbial community. Appl. Soil Ecol. 147:103387. CrossRef. Google Scholar.
Clark M. S., Horwath W. R., Shennan C., Scow K. M. (1998). Changes in soil chemical properties resulting from organic and low-input farming practices. Agron. J. 90, 662–671. doi: 10.2134/agronj1998.00021962009000050016x. CrossRef. Google Scholar.
Dong W. Y., Zhang X. Y., Dai X. Q., Fu X. L., Yang F. T., Liu X. Y. et al. (2014). Changes in soil microbial community composition in response to fertilization of paddy soils in subtropical China. Appl. Soil Ecol. 84, 140–147. doi: 10.1016/j.apsoil.2014.06.007. CrossRef. Google Scholar.
Drinkwater L. E., Letourneau D. K., Workneh F., van Bruggen A. H. C., Shennan C. (1995). Fundamental differences between conventional and organic tomato agroecosystems in California. Ecol. Appl. 5, 1098–1112. doi: 10.2307/2269357. CrossRef. Google Scholar.
Durrer A., Gumiere T., Rumenos M., Zagatto G., Feiler H. P., Silva A. M. M. et al. (2021). Organic farming practices change the soil bacteria community, improving soil quality and maize crop yield. PeerJ 9:e11985. doi: 10.7717/peerj.11985. CrossRef. Google Scholar.
Emiru N., Gebrekidan H. (2013). Effect of land use changes and soil depth on soil organic matter, total nitrogen and available phosphorus contents of soils in Senbat Watershed, Western Ethiopia. Am. J. Agric. Biol. Sci. 8, 206–212. CrossRef. Google Scholar.
Eshetu Z., Giesler R., Högberg P. (2004). Historical land use pattern affects the chemistry of forest soils in the Ethiopian highlands. Geoderma 118, 149–165. CrossRef. Google Scholar.
Farag I. F., Youssef N. H., Elshahed M. S. (2017). Global distribution patterns and pangenomic diversity of the candidate phylum "Latescibacteria" (WS3). Appl. Environ. Microbiol. 83:e00521-17. doi: 10.1128/AEM.00521-17. CrossRef. Google Scholar.
Fierer N., Bradford M. A., Jackson R. B. (2007). Toward an ecological classification of soil bacteria. Ecology 88, 1354–1364. doi: 10.1890/05-1839. CrossRef. Google Scholar.
Gajda A. M., Czyz E. A., Ukalska-Jaruga A. (2020). Comparison of the effects of different crop production systems on soil physico-chemical properties and microbial activity under winter wheat. Agronomy 10:1130. doi: 10.3390/agronomy10081130. CrossRef. Google Scholar.
Gannett M., Pritts M. P., Lehmann J. (2019). Soil amendments affect soil health indicators and crop yield in perennial strawberry. HortTechnology 29, 179–188. doi: 10.21273/HORTTECH03959-18. CrossRef. Google Scholar.
Gebreselassie Y. (2002). Selected chemical and physical characteristics of soils of Adet Research Center and its testing sites in north-western Ethiopia. Ethiopian J. Nat. Res. 4, 199–215. CrossRef. Google Scholar.
Geissen V., Guzman G. M. (2006). Fertility of tropical soils under different land use systems-a case study of soils in Tabasco, Mexico. Appl. Soil Ecol. 31, 169–178. doi: 10.1016/j.apsoil.2005.02.012. CrossRef. Google Scholar.
Gong J. L., Chen N. F., Huang Y., Huang Z., Peng H. (2005). Effect of land use on soil nutrients in the Loess hilly area of the Loess Plateau, China. Land Degrad. Dev. 17, 453–465. doi: 10.1002/ldr.701. CrossRef. Google Scholar.
Green J. L., Holmes A. J., Westoby M., Oliver I., Briscoe D., Dangerfield M. et al. (2004). Spatial scaling of microbial eukaryote diversity. Nature 432, 747–750. doi: 10.1038/nature03034. CrossRef. Google Scholar.
Griesheim K. L., Mulvaney R. L. (2019). Improving the accuracy of diffusion for inorganic 15n analyses of soil extracts and water. Commun. Soil Sci. Plant Anal. 50, 1161–1169. doi: 10.1080/00103624.2019.1604734. CrossRef. Google Scholar.
Han S., Delgado-Baquerizo M., Luo X. S., Liu Y. R., Nostrand J. D. V., Chen W. L. et al. (2021). Soil aggregate size-dependent relationships between microbial functional diversity and multifunctionality. Soil Biol. Biochem. 154:108143. CrossRef. Google Scholar.
Hao D. C., Ge G. B., Yang L. (2008). Bacterial diversity of Taxus rhizosphere: Culture-independent and culture-dependent approaches. FEMS Microbiol. Lett. 284, 204–212. doi: 10.1111/j.1574-6968.2008.01201.x. CrossRef. Google Scholar.
Hartmann M., Frey B., Mayer J., Mader P., Widmer F. (2015). Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 9, 1177–1194. CrossRef. Google Scholar.
Hathaway-Jenkins L. J., Sakrabani R., Pearce B., Whitmore A. P., Godwin R. J. (2011). A comparison of soil and water properties in organic and conventional farming systems in England. Soil Use Manag. 27, 133–142. doi: 10.1111/j.1475-2743.2011.00335.x. CrossRef. Google Scholar.
He X. L., Song C., Zhang L. P., Wang N., Cao L. H., Wei D. S. (2017). Effects of landfill leachate on microbial diversity in contaminated soil. J. Tianjin Polytechnic Univ. 36, 36–40. doi: 10.3969/j.issn.1671-024x.2017.01.007. CrossRef. Google Scholar.
Hemkemeyer M., Dohrmann A. B., Christensen B. T., Tebbe C. C. (2018). Bacterial preferences for specific soil [article size fractions revealed by community analyses. Front. Microbiol. 9:149. doi: 10.3389/fmicb.2018.00149. CrossRef. Google Scholar.
Hietala R., Liu P., Qi S. (2021). Does small-scale organic farming contributes to the local environment---A case study in suburban Shanghai, China. Agronomy 11:1601. doi: 10.3390/agronomy11081601. CrossRef. Google Scholar.
Hou Z., Li P., Li B., Gong J., Wang Y. (2007). Effects of fertilization scheme on N uptake and N use efficiency in cotton. Plant Soil 290, 115–126. doi: 10.1007/s11104-006-9140-1. CrossRef. Google Scholar.
Huang M., Zhou X., Cao F., Zou Y. (2016). Long-term effect of no-tillage on soil organic carbon and nitrogen in an irrigated rice-based cropping system. Paddy Water Environ. 14, 367–371. doi: 10.1007/s10333-015-0506-y. CrossRef. Google Scholar.
Ivanova E. A., Kutovaya O. V., Tkhakakhova A. K., Chernov T. I., Pershina E. V., Markina L. G. et al. (2015). The structure of microbial community in aggregates of a typical Chernozem aggregates under contrasting variants of its agricultural use. Eurasian Soil Sci. 48, 1242–1256. doi: 10.1134/S1064229315110083. CrossRef. Google Scholar.
Jing X., Sanders N. J., Shi Y., Chu H. Y., Classen A. T., Zhao K. et al. (2015). The links between ecosystem multifunctionality and above- and belowground biodiversity are mediated by climate. Nat. Commun. 6:8159. doi: 10.1038/ncomms9159. CrossRef. Google Scholar.
Jingwei W. (2016). Different effects of fertilization treatment on microbial phospholipid fatty acids in reclaimed soil. Ph.D. thesis. Jinzhong: Shanxi Agricultural University. CrossRef. Google Scholar.
Jones R. T., Robeson M. S., Lauber C. L., Hamady M., Knight R., Fierer N. et al. (2009). A comprehensive survey of soil acidobacterial diversity using pyrosequencing and clone library analyses. ISME J. 3, 442–453. doi: 10.1038/ismej.2008.127. CrossRef. Google Scholar.
Kandler O., Konig H. (1998). Cell wall polymers in Archaea (Archaebacteria). Cell. Mol. Life Sci. 54, 305–308. doi: 10.1007/s000180050156. CrossRef. Google Scholar.
Kirchmann H., Schan M., Barjesson G., Hamnér K., Kätterer T. (2013). Properties of soils in the Swedish long-term fertility experiments: VII. Changes in topsoil and upper subsoil at Orja and Fors after 50 years of nitrogen fertilization and manure application. Acta Agric. Scand. B Soil Plant Sci. 63, 25–36. doi: 10.1080/09064710.2012.711352. CrossRef. Google Scholar.
Kowalska A., Pawlewicz A., Dusza M., Jaskulak M., Grobelak A. (2020). "Plant--soil interactions in soil organic carbon sequestration as a restoration tool," in Climate change and soil interactions, eds. Majeti Prasad M. N. V., Pietrzykowski M. (Amsterdam: Elsevier), 663–688. doi: 10.1016/B978-0-12-818032-7-00023-0. CrossRef. Google Scholar.
Kwiatkowski C. A., Haliniarz M., Harasim E. (2020). Weed infestation and health of organically grown Chamomile (Chamomilla recutita (L.) Rausch.) depending on selected foliar sprays and row spacing. Agriculture 10:168. doi: 10.3390/agriculture10050168. CrossRef. Google Scholar.
Kwiatkowski C. A., Harasim E. (2020). Chemical properties of soil in four-field crop rotations under organic and conventional farming systems. Agronomy 10:1045. doi: 10.3390/agronomy10071045. CrossRef. Google Scholar.
Li X., Ding C., Zhang T., Wang X. (2014). Fungal pathogen accumulation at the expense of plant-beneficial fungi as a consequence of consecutive peanut monoculturing. Soil Biol. Biochem. 72, 11–18. doi: 10.1016/j.soilbio.2014.01.019. CrossRef. Google Scholar.
Li Y., Cao Z., Hu C., Li J., Yang H. (2014). Response of nematodes to agricultural input levels in various reclaimed and unreclaimed habitats. Eur. J. Soil Biol. 60, 120–129. doi: 10.1016/j.ejsobi.2013.12.001. CrossRef. Google Scholar.
Liu E., Yan C., Mei X., He W., Bing S. H., Ding L. et al. (2010). Long-term effect of chemical fertilizer, straw, and manure on soil chemical and biological properties in northwest China. Geoderma 158, 173–180. doi: 10.1016/j.geoderma.2010.04.029. CrossRef. Google Scholar.
Liu X., Shi Y., Kong L., Tong L., Cao H., Zhou H. et al. (2022). Long-term application of bio-compost increased soil microbial community diversity and altered its composition and network. Microorganisms 10:462. doi: 10.3390/microorganisms10020462. CrossRef. Google Scholar.
Lu L., Yin S., Liu X., Zhang W., Gu T., Shen Q. et al. (2013). Fungal networks in yield-invigorating and-debilitating soils induced by prolonged potato monoculture. Soil Biol. Biochem. 65, 186–194. doi: 10.1016/j.soilbio.2013.05.025. CrossRef. Google Scholar.
Lukow T., Dunfield P. F., Liesack W. (2000). Use of the T-RFLP technique to assess spatial and temporal changes in the bacterial community structure within an agricultural soil planted with transgenic and non-transgenic potato plants. FEMS Microbiol. Ecol. 32, 241–247. doi: 10.1111/j.1574-6941.2000.tb00717.x. CrossRef. Google Scholar.
Lupatini M., Korthals G. W., de Hollander M., Janssens T. K. S., Kuramae E. E. (2017). Soil microbiome is more heterogeneous in organic than in conventional farming system. Front. Microbiol. 7:2064. doi: 10.3389/fmicb.2016.02064. CrossRef. Google Scholar.
Ma N.-N., Li T.-L., Wu C.-C., Zhang E.-P. (2010). Effects of long-term fertilization on soil enzyme activities and soil physicochemical properties of facility vegetable field. Chin. J. Appl. Ecol. 21, 1766–1771. CrossRef. Google Scholar.
Malo D. D., Schumacher T. E., Doolittle J. J. (2005). Long-term cultivation impacts on selected soil properties in the Northern Great Plains. Soil Tillage Res. 81, 277–291. doi: 10.1016/j.still.2004.09.015. CrossRef. Google Scholar.
Mandal A., Toor A. S., Dhaliwal S. S. (2018). Effect of land-uses on physico-chemical properties and nutrient status of surface (0-15 cm) and sub-surface (15-30 cm) layers in soils of South-Western Punjab, India. Int. J. Curr. Microbiol. Appl. Sci. 7, 2659–2671. doi: 10.20546/ijcmas.2018.706.315. CrossRef. Google Scholar.
Manna M. C., Swarup A., Wanjari R. H., Ravankar H. N., Mishra B., Saha M. N. et al. (2005). Long-term effect of fertilizer and manure application on soil organic carbon storage, soil quality and yield sustainability under sub-humid and semi-arid tropical India. Field Crop Res. 93, 264–280. CrossRef. Google Scholar.
Matheron G. (1963). Principles of geostatistics. Econ. Geol. 58, 1246–1266. doi: 10.2113/gsecongeo.58.8.1246. CrossRef. Google Scholar.
Medan D., Torretta J. P., Hodara K., dela Fuente E. B., Montaldo N. H. (2011). Effects of agriculture expansion and intensification on the vertebrate and invertebrate diversity in the Pampas of Argentina. Biodivers. Conserv. 20, 3077–3100. doi: 10.1007/s10531-011-0118-9. CrossRef. Google Scholar.
Megyes M., Borsodi A. K., Arendas T. M., Marialigeti K. (2021). Variations in the diversity of soil bacterial and archaeal communities in response to different long-term fertilization regimes in maize fields. Appl. Soil Ecol. 168:104120. doi: 10.1016/j.apsoil.2021.104120. CrossRef. Google Scholar.
Mohammadi S., Kalbasi M., Shariatmadri H. (2009). Cumulative and residual effects of organic fertilizer application on selected soil properties, water soluble P and phosphorus sorption index. J. Agric. Sci. Technol. 11, 487–497. CrossRef. Google Scholar.
Morales L. A., Vazquez E. V., Paz-Ferreiro J. (2011). Spatial and temporal variability of Mehlich-1 extractable Fe, Mn and Zn over a rice field as a function of lime amendment. Stoch. Environ. Res. Risk Assess. 25, 1039–1048. doi: 10.1007/s00477-011-0490-8. CrossRef. Google Scholar.
Morshedi L., Lashgarara F., Hosseini S. J. F., Najafabadi M. O. (2017). The role of organic farming for improving food security from the perspective of Fars farmers. Sustainability 9:2086. doi: 10.3390/su9112086. CrossRef. Google Scholar.
Nacke H., Thürmer A., Wollherr A., Will C., Hodac L., Herold N. et al. (2011). Pyrosequencing-based assessment of bacterial community structure along different management types in German forest and grassland soils. PLoS One 6:e17000. doi: 10.1371/journal.pone.0017000. CrossRef. Google Scholar.
Nandan R., Singh V., Singh S. S., Kumar V., Hazra K. K., Nath C. P. et al. (2019). Impact of conservation tillage in rice--based cropping systems on soil aggregation, carbon pools and nutrients. Geoderma 340, 104–114. doi: 10.1016/j.geoderma.2019.01.001. CrossRef. Google Scholar.
Neha, Bhardwaj Y., Sharma M. P., Pandey J., Dubey S. K. (2022). Response of crop types and farming practices on soil microbial biomass and community structure in tropical agroecosystem by lipid biomarkers. J. Soil Sci. Plant Nutr. 22. doi: 10.1007/s42729-022-00758-3. CrossRef. Google Scholar.
Niu S. Q., Long Y., Li H. Y., Da W. Y., Hu S., Li W. J. et al. (2017). Microbial diversity in saline alkali soil from Hexi Corridor analyzed by Illumina MiSeq high-throughput sequencing system. Microbiol. China 44, 2067–2078. doi: 10.13344/j.microbiol.china.160824. CrossRef. Google Scholar.
Niyogi D., Kishtawal C., Tripathi S., Govindaraju R. S. (2010). Observational evidence that agricultural intensification and land use change may be reducing the Indian summer monsoon rainfall. Water Resour. Res. 46:W03533. doi: 10.1029/2008WR007082. CrossRef. Google Scholar.
Olsen S. R., Cole C. V., Watanable F. S., Dean L. A. (1954). Estimation of available phosphorus in soils by extraction with sodium bicarbonate. Circular, Vol. 939. Washington, DC: US Department of Agriculture. CrossRef. Google Scholar.
Ozlu E., Sandhu S. S., Kumar S., Arriaga F. J. (2019). Soil health indicators impacted by long-term cattle manure and inorganic fertilizer application in a corn soybean rotation of South Dakota. Sci. Rep. 9:11776. doi: 10.1038/s41598-019.48207-z. CrossRef. Google Scholar.
Paul C., Bayrychenko Z., Junier T., Filippidou S., Beck K., Bueche M. et al. (2018). Dissemination of antibiotic resistance genes associated with the sporobiota in sediments impacted by wastewater. PeerJ 6:e4989. doi: 10.7717/peerj.4989. CrossRef. Google Scholar.
Pimentel D., Hepperly P., Hanson J., Douds D., Seide R. (2005). Environmental, energetic, and economic comparisons of organic and conventional farming systems. BioScience 55, 573–582. CrossRef. Google Scholar.
Ponisio L. C., M'Gonigle L. K., Mace K. C., Palomino J., de Valpine P., Kremen C. (2015). Diversification practices reduce organic to conventional yield gap. Proc. R. Soc. B Biol. Sci. 282:20141396. doi: 10.1098/rspb.2014.1396. CrossRef. Google Scholar.
Reilly K., Cullen E., Lola-luz T., Stone D., Valverde J., Gaffney M. et al. (2013). Effect of organic, conventional and mixed cultivation practices on soil microbial community structure and nematode abundance in a cultivated onion crop. J. Sci. Food Agric. 93, 3700–3709. doi: 10.1002/jsfa.6206. CrossRef. Google Scholar.
Riedo J., Wettstein F. E., Rösch A., Herzog C., Banerjee S., Büchi L. et al. (2021). Widespread occurrence of pesticides in organically managed agricultural soils---the ghost of a conventional agricultural past? Environ. Sci. Technol. 55, 2919–2928. doi: 10.1021/acs.est.0c06405. CrossRef. Google Scholar.
Robertson G. P., Sollins P., Ellis B. G., Laijtha K. (1999). "Exchangeable ions, pH, and cation exchange capacity," in Standard soil methods for long term ecological research, eds. Robertson G. P., Coleman D. C., Bledsoe C. S., Sollins P. (New York, NY: Oxford University Press). CrossRef. Google Scholar.
Roesch L. F. W., Fulthorpe R. R., Riva A., Casella G., Hadwin A. K. M., Kent A. D. et al. (2007). Pyro-sequencing enumerates and contrasts soil microbial diversity. ISME J. 1, 283–290. CrossRef. Google Scholar.
Sankaramoorthy A., Rangasamy V. (2019). Impacts of different sources of organic manures on soil physico-chemical properties, nutrient balance and yield of rice-green gram cropping sequence under organic farming. SSRG Int. J. Agric. Environ. Sci. 6, 65–85. doi: 10.14445/23942568/IJAES-V6I4P112. CrossRef. Google Scholar.
Selassie Y. G., Ayanna G. (2013). Effects of different land use systems on selected physico-chemical properties of soils in Northwestern Ethiopia. J. Agric. Sci. 5, 112–120. doi: 10.5539/jas.v5n4p112. CrossRef. Google Scholar.
Seufert V., Ramankutty N. (2017). Many shades of Gray---the context dependent performance of organic agriculture. Sci. Adv. 3:e1602638. doi: 10.1126/sciadv.1602638. CrossRef. Google Scholar.
Shannon D., Sen A. M., Johnson D. B. (2002). A comparative study of the microbiology of soils managed under organic and conventional regimes. Soil Use Manag. 18(Suppl. S1), 274–283. doi: 10.1111/j.1475-2743.2002.tb00269.x. CrossRef. Google Scholar.
Sharma S., Singh P., Sodhi G. P. S. (2020). Soil organic carbon and biological indicators of uncultivated vis-a-vis intensively cultivated soils under rice-wheat and cotton --wheat cropping systems in South-Western Punjab. Carbon Manage. 11, 681–695. doi: 10.1080/17583004.2020.1840891. CrossRef. Google Scholar.
Shen J. P., Zhang L. M., Guo J. F., Ray J. L., He J. Z. (2010). Impact of long-term fertilization practices on the abundance and composition of soil bacterial communities in Northeast China. Appl. Soil Ecol. 46, 119–124. doi: 10.1016/j.apsoil.2010.06.015. CrossRef. Google Scholar.
Shukla M. K., Lal R., Ebinger M. (2006). Determining soil quality indicators by factor analysis. Soil Tillage Res. 87, 194–204. doi: 10.1016/j.still.2005.03.011. CrossRef. Google Scholar.
Song Z.-Q., Cheng J.-E., Cheng F.-X., Zhang D.-Y., Liu Y. (2017). Development and evaluation of loop-mediated isothermal amplification assay for rapid detection of Tylenchulus semipenetrans using DNA extracted from soil. Plant Pathol. J. 33, 184–192. doi: 10.5423/PPJ.OA.10.2016.0224. CrossRef. Google Scholar.
Sun R., Zhang X. X., Guo X., Wang D., Chu H. (2015). Bacterial diversity in soils subjected to long-term chemical fertilization can be more stably maintained with the addition of livestock manure than wheat straw. Soil Biol. Biochem. 88, 9–18. doi: 10.1016/j.soilbio.2015.05.007. CrossRef. Google Scholar.
Thomas B. W., Hunt D., Bittman S., Hannam K. D., Messiga A. J., Haak D. et al. (2019). Soil health indicators after 21 yr of no-tillage in South Coastal British Columbia. Can. J. Soil Sci. 99, 222–225. doi: 10.1139/cjss-2018-0146. CrossRef. Google Scholar.
Tuma A. (2007). Effects of fruit based land use systems on soil physicochemical properties: The case of smallholders farming systems in Gamo Gofa, Southern Ethiopia. M. Sc. thesis. Hawassa: Hawassa University. CrossRef. Google Scholar.
Ukalska-Jaruga A., Klimkowicz-Pawlas A., Smreczak B. (2019). Characterization of organic matter fractions in the top layer of soils under different land uses in Central-Eastern Europe. Soil Use Manag. 35, 595–606. doi: 10.1111/sum.12514. CrossRef. Google Scholar.
Uroz S., Ioannidis P., Lengelle J., CeÂbron A., Morin E., BueÂe M. et al. (2013). Functional assays and metagenomic analyses reveals differences between the microbial communities inhabiting the soil horizons of a Norway spruce plantation. PLoS One 8:e55929. doi: 10.1371/journal.pone.0055929. CrossRef. Google Scholar.
Venkatesh M. S., Hazra K. K., Ghosh P. K., Praharaj C. S., Kumar N. (2013). Long--term effect of pulses and nutrient management on soil carbon sequestration in Indo--Gangetic plains of India. Can. J. Soil Sci. 93, 127–136. doi: 10.4141/CJSS2012-072. CrossRef. Google Scholar.
Wakene N. (2001). Assessment of important physicochemical properties of Nitosols under different management systems in Bako Area, Western Ethiopia. M. Sc. thesis. Alemaya: Alemaya University. CrossRef. Google Scholar.
Wang J., Song Y., Ma T., Raza W., Li J., Howland J. G. et al. (2017). Impacts of inorganic and organic fertilization treatments on bacterial and fungal communities in a paddy soil. Appl. Soil Ecol. 112, 42–50. doi: 10.1016/j.apsoil.2017.01.005. CrossRef. Google Scholar.
Wang L., Kaur M., Zhang P., Li J., Xu M. (2021). Effect of different agricultural farming practices on microbial biomass and enzyme activities of celery growing field soil. Int. J. Environ. Res. Public Health 18:12862. doi: 10.3390/ijerph182312862. CrossRef. Google Scholar.
Wang Y., Tu C., Cheng L., Li C., Gentry L. F., Hoyt G. D. et al. (2011). Long-term impact of farming practices on soil organic carbon and nitrogen pools and microbial biomass and activity. Soil Tillage Res. 117, 8–16. doi: 10.1016/j.still.2011.08.002. CrossRef. Google Scholar.
Willers C., van Rensburg P. J., Claassens S. (2015). Phospholipid fatty acid profiling of microbial communities--a review of interpretations and recent applications. J. Appl. Microbiol. 119, 1207–1218. doi: 10.1111/jam.12902. CrossRef. Google Scholar.
Xomphoutheb T., Jiao S., Guo X., Mabagala F. S., Sui B., Wang H. et al. (2020). The effect of tillage systems on phosphorus distribution and forms in rhizosphere and non-rhizosphere soil under maize (Zea mays L.) in Northeast China. Sci. Rep. 10:6574. doi: 10.1038/s41598-020-63567-7. CrossRef. Google Scholar.
Xu H., Zhang C. (2021). Investigating spatially varying relationships between total organic carbon contents and pH values in European agricultural soil using geographically weighted regression. Sci. Total Environ. 752:141977. doi: 10.1016/j.scitotenv.2020.141977. CrossRef. Google Scholar.
Yang T., Kadambot H. M. S., Liu K. (2020). Cropping systems in agriculture and their impact on soil health-A review. Glob. Ecol. Conserv. 23:e01118. doi: 10.1016/j.gecco.2020.e01118. CrossRef. Google Scholar.
Yin C., Jones K. L., Peterson D. E., Garrett K. A., Hulbert S. H., Paulitz T. C. et al. (2010). Members of soil bacterial communities sensitive to tillage and crop rotation. Soil Biol. Biochem. 42, 2111–2118. doi: 10.1016/j.soilbio.2010.08.006. CrossRef. Google Scholar.
Zhang Q. C., Wang X. Q., Shi Y. N., Wang G. H. (2010). Effects of different fertilizer treatments on ecological characteristics of microorganism in chemical fertilizer omission paddy soil. J. Plant Nutr. Fertil. 16, 118–123. CrossRef. Google Scholar.
Zhong W., Gu T., Wang W., Zhang B., Lin X., Huang Q. et al. (2010). The effects of mineral fertilizer and organic manure on soil microbial community and diversity. Plant Soil 326, 511–522. doi: 10.1007/s11104-009-9988-y. CrossRef. Google Scholar.
Zhou W., Lv T.-F., Chen Y., Westby A. P., Ren W.-J. (2014). Soil physicochemical and biological properties of paddy-upland rotation: A review. Sci. World J. 2014:856352. doi: 10.1155/2014/856352. CrossRef. Google Scholar.
Zhou Z., Wang C. (2017). Soil-microbe-mineralization carbon and nitrogen stoichiometry under different land-uses in the Maoershan region. Acta Ecol. Sin. 37, 2428–2436. CrossRef. Google Scholar.
Zhu S., Wang Y., Xu X., Liu T., Wu D., Zheng X. et al. (2018). Potential use of high-throughput sequencing of soil microbial communities for estimating the adverse effects of continuous cropping on ramie (Boehmeria nivea L. Gaud). PLoS One 13:e0197095. doi: 10.1371/journal.pone.0197095. CrossRef. Google Scholar.
Zhu Y., Wu F., Feng W., Liu S., Giesy J. P. (2016). Interaction of alkaline phosphatase with minerals and sediments: Activities, kinetics, and hydrolysis of organic phosphorus. Colloids Surf. A Physicochem. Eng. Asp. 495, 46–53. doi: 10.1016/j.colsurfa.2016.01.05. CrossRef. Google Scholar.

Kaur M, Li J, Zhang P, Yang H, Wang L and Xu M (2022) Agricultural soil physico-chemical parameters and microbial abundance and diversity under long-run farming practices: A greenhouse study. Front. Ecol. Evol. 10:1026771. doi: 10.3389/fevo.2022.1026771
Перевод статьи «Agricultural soil physico-chemical parameters and microbial abundance and diversity under long-run farming practices: A greenhouse study» авторов Kaur M, Li J, Zhang P, Yang H, Wang L and Xu M., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык
Фото: Designed by Freepik





















































































































Комментарии (0)