Подводные перемены: влияние лососёвых ферм на микробные сообщества донных осадков
В последние годы индустрия выращивания лосося значительно выросла во всём мире, а на Фарерских островах она стала одной из ключевых отраслей экономики с ежегодным объёмом производства более 94 000 тонн, что составляет 24% ВВП. Согласно экологическим нормативам, в течение каждого производственного цикла на всех фермерских участках проводится мониторинг состояния морского дна, включающий макрофаунальный, сенсорный и химический анализы. Однако влияние рыбоводной деятельности на микроорганизмы на Фарерских островах до сих пор остаётся неизученным.
Аннотация
Целью данного исследования было оценить воздействие выращивания атлантического лосося на бентосные микробные сообщества, чтобы глубже понять последствия для основы бентосной пищевой сети, а также определить, являются ли эти сообщества более чувствительными к антропогенному воздействию по сравнению с традиционным макрофаунальным биомониторингом.
Керны донных отложений отбирались по трансекте — от точки непосредственно под садками с лососем до фонового референтного участка. Отбор проб проводился до выпуска молоди лосося в садки (этап «до зарыбления») и непосредственно перед выловом рыбы (этап «пик биомассы»). В образцах осадка с поверхности, а также с глубины 3 см и 10 см было проведено секвенирование гена 16S рРНК (участки V4–V5) на платформе Illumina MiSeq. Были обнаружены значительные изменения в составе микробных сообществ между этапами «до зарыбления» и «пик биомассы», а также между разными слоями осадка. Микробное разнообразие увеличивалось по мере удаления от садков и было максимальным на этапе «до зарыбления», что указывает на существенное влияние лососеводства на структуру микробного сообщества. В отличие от результатов регулярно проводимого экологического мониторинга, данные этого исследования показали наличие воздействия рыбоводства на донные отложения, что подчёркивает эффективность ДНК-метабаркодинга как мощного альтернативного метода биомониторинга аквакультурных зон.
Введение
Аквакультура стала областью, имеющей крупное мировое промышленное значение (Kawahara и др., 2009). За последнее десятилетие она росла интенсивными темпами, чтобы соответствовать постоянно растущему мировому спросу на морепродукты (Stoeck и др., 2018; Leontidou и др., 2023). В глобальном масштабе продукция морской аквакультуры в 2021 году составила 57,5 миллиона тонн (FAO, 2022), при этом большинство приходится на костистых рыб (FAO, 2018). Эта крупная и растущая отрасль вызывает обеспокоенность по поводу воздействия аквакультуры на морскую среду (Kawahara и др., 2009; Verhoeven и др., 2018; Ape и др., 2019). Большинство выращиваемых костистых рыб, таких как лосось, являются хищниками с рационом, богатым фосфатами и белками (Stoeck и др., 2018). Хотя большая часть углерода, фосфора и азота, поступающих с кормом, извлекается из выловленной рыбы, значительные количества также попадают в окружающую среду в виде растворённых стоков или частиц корма и фекалий, которые оседают на морском дне (Gowen и Bradbury, 1987; Folke и Kautsky, 1989; Price и др., 2015). Таким образом, в период активного рыбоводства морское дно обогащается органическими частицами в виде несъеденного корма и фекальных материалов (Carroll и др., 2003; Holmer и др., 2005; Buschmann и др., 2006). Изменения в органическом обогащении донных отложений влияют на биогеохимические процессы и бентосные микробные сообщества (McCaig и др., 1999; Norði и др., 2011; Bannister и др., 2014; Sweetman и др., 2014). Эффекты органического обогащения зависят от нескольких факторов, таких как водные течения, турбулентность и глубина, а также от продолжительности, качества и количества отходов.
Когда количество отложений превышает естественную скорость разложения органического вещества (ОВ), на поверхности осадка оседает слой мелкозернистого материала с высоким содержанием органических частиц. Со временем океанское дно закисляется, а кислород истощается из-за процессов микробной деградации. При этом могут образовываться токсичные газы, такие как сероводород и метан, которые, в свою очередь, могут влиять на здоровье рыбы и других животных (Gowen и Bradbury, 1987; Black и др., 1996; Forrest и др., 2007; Norði и др., 2011). Эти геохимические изменения в структуре морского дна обычно приводят к изменениям в сообществах инфауны и эпифауны. Это приводит к тому, что менее устойчивые бентосные животные заменяются меньшим числом более оппортунистических и толерантных видов. В таких экстремальных условиях на поверхности осадка могут формироваться слои хемоавтотрофных бактерий. Со временем осадки вокруг рыбоводных садков могут становиться азойными (Brown и др., 1987; Holmer и др., 2005; Keeley и др., 2013b), что приводит к ухудшению здоровья морского дна (Hargrave и др., 1993; Hyland и др., 2005; Forrest и др., 2007).
Наиболее серьёзные экологические последствия от морских лососёвых ферм наблюдаются непосредственно под садками. Эти эффекты демонстрируют градиент снижения воздействия с увеличением расстояния, что согласуется с градиентами органического обогащения (Schendel и др., 2004; Hyland и др., 2005; Forrest и др., 2007). Осадочный «след» типичных лососёвых ферм простирается на сотни метров от точки сброса, которая обычно находится в центре садков и часто имеет овальную форму, смещённую в направлении преобладающих течений (Brown и др., 1987; Forrest и др., 2007; Keeley и др., 2013a; Bannister и др., 2016). Поверхностный аэробный слой осадка имеет толщину от нескольких миллиметров до сантиметров и зависит от баланса между проникновением и потреблением кислорода. Однако если вышележащие воды становятся бескислородными, поверхность осадка становится анаэробной (Kristensen, 2000; Hargrave и др., 2008; Norði и др., 2011). Тип осадка и его склонность к органическому обогащению в основном определяются гидродинамикой и глубиной участка, что также контролирует форму, интенсивность и размер следа (Keeley и др., 2013b; Keeley и др., 2019).
Поскольку эвтрофикация влияет на биогеохимические процессы в донных отложениях, бентосные сообщества, особенно микробные, сильно страдают (Asami и др., 2005). Высокий уровень обогащения ОВ непосредственно под рыбоводными садками обычно проявляется через группу различных «индикаторов», таких как анаэробные микроорганизмы или высокое содержание видов, толерантных к обогащению, например, полихеты Capitella capitata (Brown и др., 1987; Karakassis и др., 2000; Neofitou и др., 2010; Gaard и др., 2011). Предыдущие исследования микробных сообществ под фермами аквакультуры показали, что они могут быть высокочувствительными к локальным условиям окружающей среды (Quero и др., 2020; Frühe и др., 2021a; Keeley и др., 2021; Pawlowski и др., 2022). Однако существует мало исследований, если они вообще есть, которые изучают микробные сообщества на разных глубинных слоях осадка. Из-за их относительно быстрой сменяемости изменения в микробных популяциях в осадках могут служить более чувствительным и точным индикатором для оценки воздействия на морские осадки в более короткие временные масштабы и, следовательно, могут быть более мощным экологическим индикатором по сравнению с мейо- и мегафауной (Kawahara и др., 2009; Stoeck и др., 2018; Frühe и др., 2021b).
Насколько нам известно, мы представляем первое исследование с Фарерских островов, которое фокусируется на структурах микробных сообществ в колонках осадка на расстоянии от садков аквакультуры в условиях режимных сдвигов органического вещества от лососёвого хозяйства.
Материалы и методы
Отбор и подготовка проб
Пробы осадка были отобраны 25 сентября 2019 г. и 15 июля 2020 г. в районе рыбоводства A-83 в Сёрвогуре, Фарерские острова (Рисунок 1). Первый отбор проб проводился после трёхмесячного периода залежи, а второй отбор проб проводился, когда биомасса лосося была на пике.
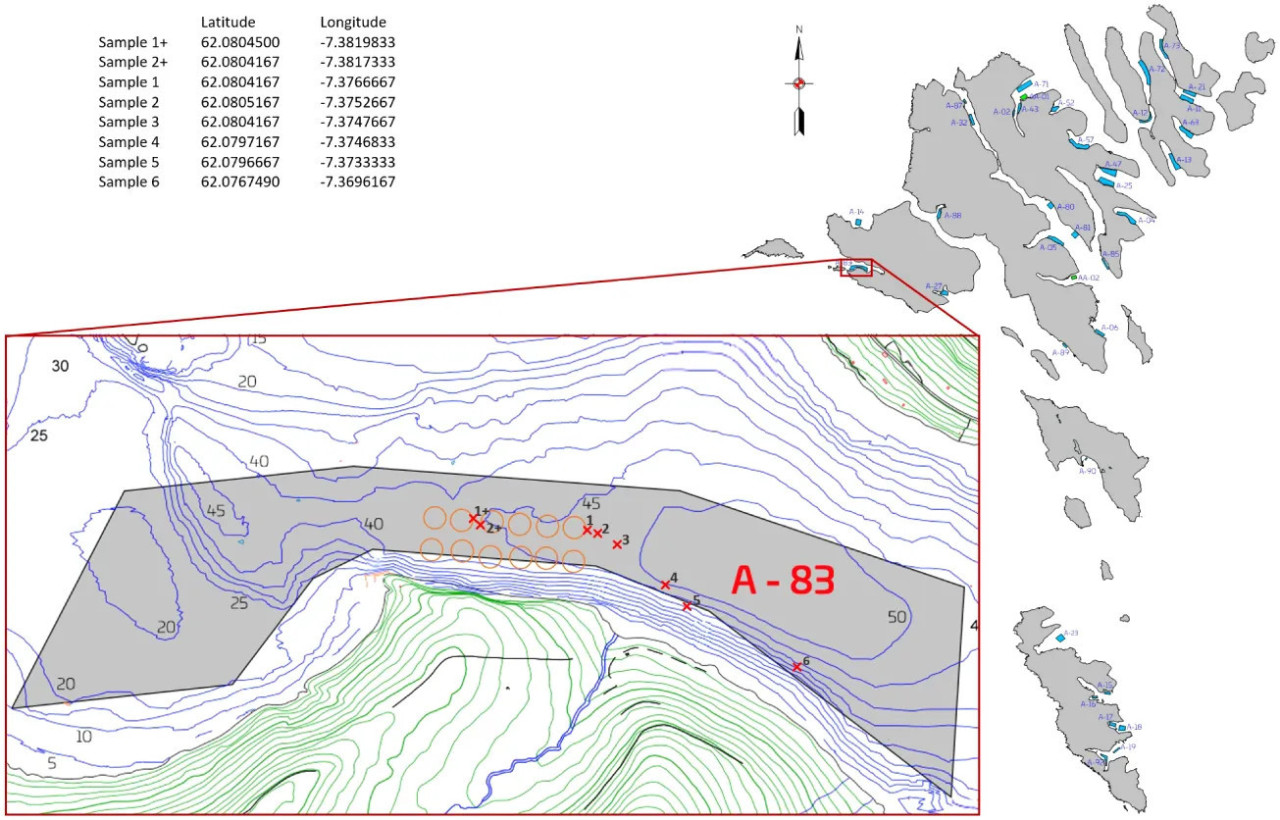
Рисунок 1 – Расположение фермерского участка A-83 на Фарерских островах. Красные крестики – станции отбора проб. Оранжевые круги – рыбоводные садки.
Начиная со станции у садков RS20, вдоль трансекты было отобрано шесть станций (Рисунок 1). Первая станция отбиралась непосредственно на периферии садка (0 м), а затем приблизительно на 12 м, 40 м, 150 м, 190 м и 600 м от RS20. Трансекта вглубь фьорда следует за направлением преобладающего течения, измеренного приблизительно в 225 м к западу от фермы (Norði и др., 2023) (Рисунок 1). Станция отбора на 600 м была определена как фоновая проба на основе результатов исследования, указывающих на то, что максимальная зона воздействия органического обогащения от рыбоводных ферм в фарерских фьордах составляет 200 м (Mortensen и др., 2021). Наш фоновый участок, находящийся в 600 м, был выбран на основе порога, установленного Советом по управлению аквакультурой (ASC), – расстояние более 500 м между рыбоводной фермой и референтной станцией (Mortensen и др., 2021). Однако станция отбора на 600 м находится на несколько меньшей глубине (≈ 35 м) по сравнению с другими пробами (≈ 45 м). Из-за небольших размеров фьорда мы выбрали наиболее подходящую референтную точку в радиусе 500 м. Во время отбора проб «пик биомассы» были взяты две дополнительные пробы (1+ и 2+, показаны на Рисунке 1) из-за перемещения садков (RS20, RS22, RS24 и RS26) в течение производственных циклов. Для обеспечения сопоставимости с отбором проб «до зарыбления» дополнительные пробы были отобраны из RS28 (представляющего расстояние 0 м и 12 м), который был самой внешней станцией с садками на той же трансекте. Таким образом, пробы отбирались на станциях 1, 2, 3, 4, 5 и 6 в оба периода отбора, но только на станциях 1+ и 2+ в период «пик биомассы». В общей сложности это составило 14 станций. Пробы отбирались с помощью трубчатой грунтовой трубки, состоящей из плексигласовых трубок длиной 47 см и диаметром 44 мм (Дополнительный файл 1). Каждый из 14 кернов осадка был нарезан на интервалы по 1 см до глубины 10 см. Анализировались три слоя осадка: поверхностный (соскоб с поверхности), глубина 3 см и глубина 10 см (Дополнительный файл 1). С каждого керна и слоя отбиралось по три повторности, что в сумме составило 126 проб. Пробы хранились при температуре -18°C в течение 2-5 часов после сбора и сохранялись до выделения ДНК, которое проводилось через 2-7 недель. Одновременно со сбором наших проб проводилась оценка экологического состояния района. Эта оценка является обязательным требованием перед запуском смолтов в садки и при достижении максимальной биомассы (т.е. перед забоем).
Пробы для законодательно установленной национальной оценки экологического состояния, как описано в ICES (2023), отбирались независимой компанией. Оценка экологического состояния проводилась в соответствии с Норвежским стандартом 9410:2016, который также используется Фарерским агентством по охране окружающей среды (Agency, 2018). Статус воздействия классифицируется следующим образом: «нет воздействия», «некоторое воздействие», «высокое воздействие» и «очень высокое воздействие» (Agency, 2018).
Выделение ДНК
Пробы размораживали при комнатной температуре, а затем гомогенизировали для обеспечения однородной консистенции. Каждая проба была разделена на три лабораторные повторности, и ДНК выделяли с использованием набора Qiagen DNeasy PowerSoil согласно инструкциям производителя. Выход ДНК после экстракции измеряли с помощью нанофотометра (3117, Implen). Средний выход ДНК для проб составил 41 ± 62 нг/мкл со средним соотношением A260/A280 1,863 ± 0,304. Схема протокола отбора проб и лабораторных исследований показана в Дополнительном файле 2.
ПЦР и секвенирование
Секвенирование проводилось для региона V4-V5 гена 16S рРНК с использованием праймеров 515F-Y и 926R согласно Parada и др. (2015). ПЦР-реакции проводились в трёх повторностях для каждой лабораторной повторности с разведением 1:10 для всех экстрактов ДНК для снижения ингибиторов ПЦР (Apprill и др., 2015; Djurhuus и др., 2017). Вкратце, 1 мкл ДНК-матрицы добавляли к мастер-миксу, состоящему из 12,5 мкл 2X Phusion Master Mix (Thermo Fisher Scientific, США), 10,50 мкл воды без нуклеаз и 0,5 мкл каждого праймера (0,2 мкМ) (Parada и др., 2015), в результате получалась реакция объёмом 25 мкл. ПЦР-реакция проходила при 98°C в течение 30 с, затем 27 циклов: 98°C – 10 с, 50°C – 30 с и 72°C – 30 с, с финальной элонгацией при 72°C в течение 10 мин. После ПЦР три повторности объединяли. Для подтверждения наличия целевых полос и проверки чистоты отрицательных контролей продукты амплификации объединённых проб разделяли на 1,5% агарозных гелях с использованием UView (Bio-Rad) для визуализации. Все отрицательные контроли без матрицы (NTC) и чистые контроли дали отрицательный результат. Очистку объединённых ампликонов проводили с использованием 1x магнитных частиц Agencourt AMPure XP (Beckman Coulter, США) согласно протоколу производителя. Концентрацию всех очищенных ПЦР-продуктов измеряли с помощью нанофотометра. Вторая ПЦР-реакция проводилась для присоединения двойных баркод-индексов и адаптеров для секвенирования Illumina с использованием набора Nextera XT Index Kit v2. Вкратце, 5 мкл каждого ампликона ДНК добавляли к мастер-миксу, состоящему из 25 мкл 2X KAPA HiFi (Kapa Biosystems Inc., Woburn, США) HotStart ReadyMix, 10 мкл воды без нуклеаз и 5 мкл праймеров Nextera XT Index 1 и Nextera XT Index 2, в результате чего общий объём составлял 50 мкл. ПЦР проводилась со следующими условиями: 95°C – 3 мин, 8 циклов: 95°C – 30 с, 55°C – 30 с и 72°C – 30 с, с финальной элонгацией при 72°C в течение 5 мин. Затем продукты очищали с использованием AMPure XP и измеряли их концентрацию с помощью Qubit 2.0 Fluorometer и набора Qubit dsDNA HS Assay Kit (Invitrogen, США). После количественного определения проводилась нормализация библиотек для обеспечения равномерной глубины секвенирования на пробу. Библиотеки секвенирования проверяли с помощью Agilent Bioanalyzer 2100 (Agilent Technologies, США) для оценки размеров ПЦР-библиотек. Затем их секвенировали на платформе Illumina MiSeq с получением спаренных концов длиной 2 x 250 п.н.
Биоинформатика и статистика
Все пробы были демоноплексированы в файлы спаренных ридов в формате fastq, и все баркоды были удалены. Все последующие анализы проводились с использованием программного обеспечения R (R Core Team, 2020). Пакет dada2 (Callahan и др., 2016) использовался для фильтрации качества и обрезки (включая удаление праймеров), оценки ошибок, выявления вариантов последовательностей, слияния ридов и удаления химер. Таксономическая классификация проводилась с использованием референсной базы данных SILVA132. Результатом стали таблица вариантов ампликонных последовательностей (ASV) и соответствующая таблица таксономии. Качество данных секвенирования визуально оценивалось с помощью кривых разрежения для каждой пробы, созданных с использованием пакета vegan (Oksanen и др., 2013). Эти кривые разрежения использовались для определения того, была ли глубина секвенирования достаточной для охвата микробного богатства в пробах (Дополнительный файл 3).
Поскольку наши пробы имели широкий диапазон количества прочтений на пробу, мы решили не разрежать данные, чтобы не терять важную информацию путём удаления данных последовательностей (Love и др., 2014; Callahan и др., 2016). Для сохранения разрешения данных мы применили поправку на множественное тестирование Бенджамини-Хохберга к нашим данным с использованием пакета R DESeq2 (Love и др., 2014). Этот метод учитывает коррекцию неравномерного количества прочтений на пробу и подходит для нормализации наборов данных с высокой дисперсией, полученных при высокопроизводительном секвенировании (McMurdie и Holmes, 2014; Djurhuus и др., 2017).
Для получения общего представления о среднем разнообразии и богатстве альфа-разнообразие оценивалось через наблюдаемое богатство (количество ASV), визуализированное на графике богатства, созданном с использованием пакета phyloseq (McMurdie и Holmes, 2013) (Рисунок 2). В следующих анализах данные были разрежены до 19 000 прочтений/проба для равномерного сравнения богатства между пробами. Данные были проверены на нормальность с помощью теста Шапиро-Уилка, который показал ненормальное распределение. Для оценки различий между микробными сообществами в два периода отбора («до зарыбления» и «пик биомассы») и различий между всеми станциями отбора применялся ранговый тест Краскела-Уоллиса. В случаях обнаружения статистических различий проводились попарные сравнения с использованием рангового теста Уилкоксона для последующего анализа.
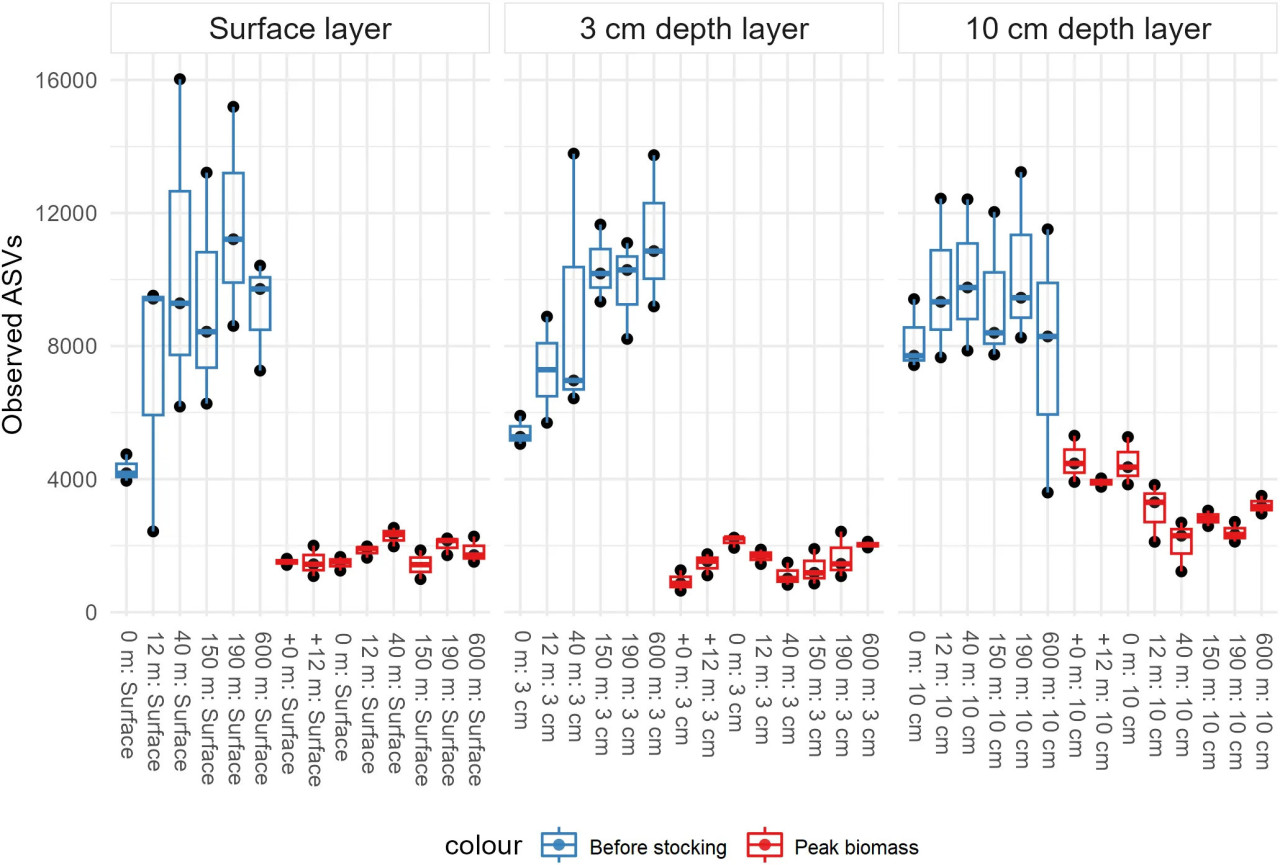
Рисунок 2 – Показатель альфа-разнообразия наблюдаемого богатства, сгруппированный по слоям осадка. Цвета соответствуют периодам отбора (до зарыбления = синий, пик биомассы = красный). Оси X – пробы, оси Y – количество наблюдаемых ASV. Для каждой пробы показаны повторности и их погрешности.
Для точного отображения порядковых расстояний между пробами в двух измерениях данные были логарифмически преобразованы, и построена диаграмма неметрического многомерного шкалирования (NMDS) с использованием индексов несходства Брея-Кёртиса в качестве меры расстояния (Рисунок 3A). Дополнительно был создан точечный график для исследования возможных корреляций между расстоянием от садков и соотношением гамма- и дельтапротеобактерий (Рисунок 3B).
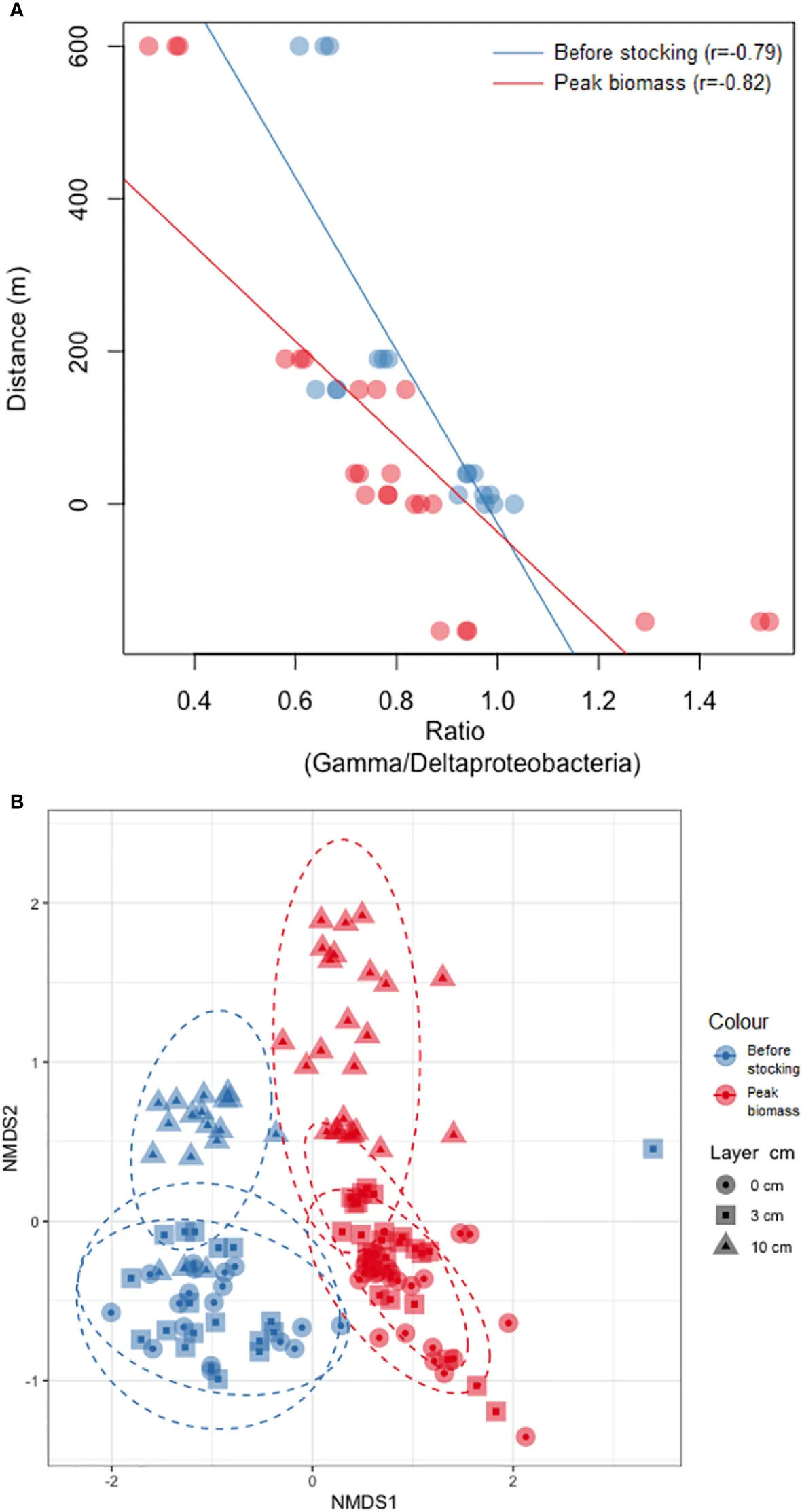
Рисунок 3 – (A) Ординация NMDS матрицы расстояний несходства Брея-Кёртиса между микробными сообществами. Синий = до зарыбления, красный = пик биомассы. Круг = 0 см, квадрат = 3 см, треугольник = 10 см. Овальные точки – эллипсы. (B) График соотношения гамма/дельтапротеобактерий. Синий = до зарыбления, красный = пик биомассы. Оси X – соотношение, оси Y – расстояние в метрах.
Для определения значимых различий в микробных сообществах в зависимости от биомассы лосося, проб и слоёв проб мы провели анализ сходства (ANOSIM) с использованием пакета R vegan (Oksanen и др., 2013). Количество перестановок было установлено на 9999. Кроме того, мы провели анализ индикаторных таксонов с помощью пакета R «indicspecies» (Cáceres и Legendre, 2009) с 9999 перестановками. Анализ проводился по расстоянию от садков и слою в керне осадка. Этот анализ проводился на данных, агрегированных на уровне семейств, из-за большого количества видов в наших пробах. Полученные данные использовались для интерпретации наших результатов и выявления некоторых тенденций в наборе данных (Рисунок 4; Дополнительные файлы 6, 8-10).
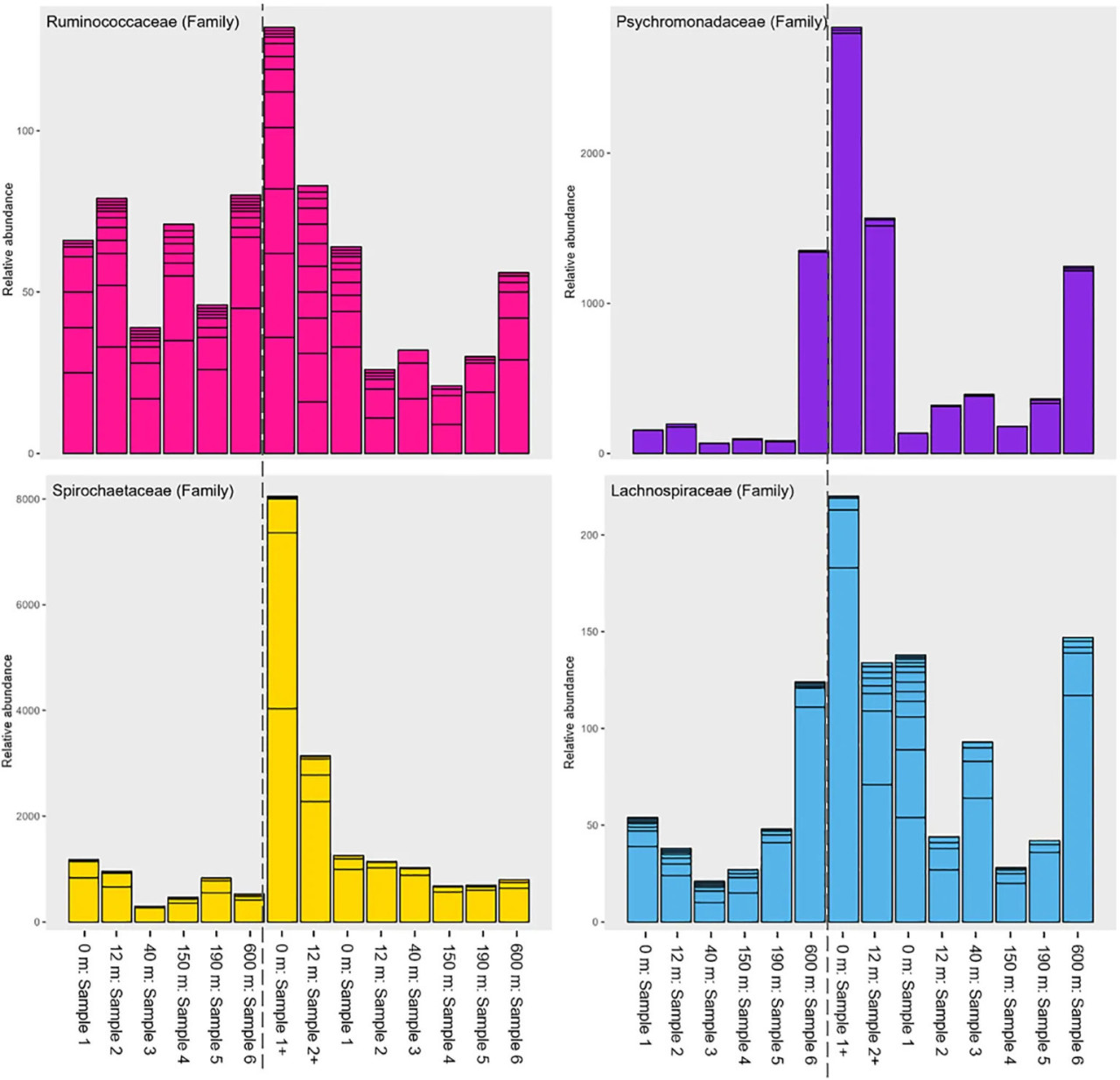
Рисунок 4 – График относительной численности семейств Ruminococcaceae, Psychromonadaceae, Spirochaetaceae и Lachnospiraceae. Слева направо – участки отбора «до зарыбления», за которыми следуют участки «пик биомассы». Оси X – расстояние от садков, оси Y – количественное отображение относительной численности.
Результаты
Национальная оценка экологического состояния
Во время отбора проб «до зарыбления» все станции у садков были в целом отнесены к категории «некоторое воздействие», за исключением одной (RS21), которая была отнесена к категории «нет воздействия» (агентство по охране окружающей среды, неопубликованные данные). Оценка также классифицировала измерения pH осадка и значений окислительно-восстановительного потенциала как «некоторое воздействие». Осадок имел тёмный цвет с сильным запахом и полутвёрдую текстуру. На станции RS20 были обнаружены виды, толерантные к органическому обогащению, – Capitella capitata (галерейный червь) и моллюски, идентифицированные на месте при визуальном осмотре после перколяции осадка из пробоотборника через сито. Кроме того, C. capitata была также обнаружена на RS28. Во время «пика биомассы» общее состояние снова было классифицировано как «некоторое воздействие» – один садок был отнесён к категории «очень высокое воздействие» (RS30), два – к «высокому воздействию» (RS29 и RS31), шесть – к «некоторому воздействию» (RS24-RS28) и три – к «нет воздействия» (RS20, RS22, RS23). На станции RS20 значения pH и окислительно-восстановительного потенциала были отнесены к категории «нет воздействия», тогда как значения для RS28 указывали на «высокое воздействие». RS20 имел бледный цвет осадка, тогда как осадок в RS28 был тёмным с сильным запахом. Тип осадка на всех участках был песчаным (неопубликованные данные).
Микробное разнообразие и структура сообщества
В целом после фильтрации качества было получено 14 407 470 высококачественных прочтений. Эти прочтения использовались для всех последующих анализов. Чистые контроли показали минимальное количество прочтений или их отсутствие (медиана = 5 прочтений), в то время как отрицательные контроли выделения ДНК имели более высокое количество прочтений (медиана = 1050 прочтений), хотя значительно более низкое по сравнению с успешными пробами. Всего в чистых контролях было обнаружено 30 ASV (Дополнительный файл 4). Из исходных 138 проб после фильтрации качества осталось 122, исключая чистые контроли, отрицательные контроли ДНК и три пробы с менее чем 19 000 прочтений. Последовательности были сгруппированы в 201 345 ASV, из которых 9440 (4,69%) не удалось отнести к типу, и они были обозначены как «NA». Остальные ASV (191 928) были отнесены к типам или более низким таксономическим классам, и 48 930 ASV (24,28%) были классифицированы на уровне рода. Только 207 (0,11%) ASV были классифицированы до уровня вида. Для отфильтрованных данных (все пробы с > 19 000 прочтений) среднее количество прочтений/проба было значительно выше в период «до зарыбления» – 173 693 по сравнению с 75 387 прочтений/проба в период «пик биомассы». Большинство проб содержали от 50 000 до 200 000 прочтений. Хотя в пробах «до зарыбления» было значительно больше прочтений/проба, ни одна из них не достигла асимптоты на кривой разрежения (Дополнительный файл 3). Для сравнения, все пробы в «пик биомассы» с меньшим числом прочтений достигли асимптоты. Это означает, что, несмотря на исключительно высокое количество прочтений в этом раунде секвенирования, разнообразие ещё больше, чем указано в данном исследовании (Рисунок 2). Наблюдаемое богатство было значительно выше в период «до зарыбления» (Рисунок 2). Все участки отбора «до зарыбления» имели значительно более высокие показатели, чем все участки «пик биомассы». Кроме того, пробы «до зарыбления» на 0 м и 12 м значительно отличались от всех остальных проб (Дополнительный файл 5).
Неметрическое многомерное шкалирование на основе несходства Брея-Кёртиса показало, что структуры бентосных бактериальных сообществ значительно изменились между двумя периодами отбора, при этом визуального перекрытия между «до зарыбления» и «пик биомассы» не наблюдалось (ANOSIM: p<0,001, Рисунок 3A). Структуры бактериальных сообществ были схожи для обоих периодов отбора: слои 0 и 3 см и слой 10 см чётко отличались друг от друга (p<0,001). Наблюдается сильная отрицательная корреляция между расстоянием от лососёвых садков и соотношением гамма/дельтапротеобактерий как в период «до зарыбления» (r = -0,79), так и в «пик биомассы» (r = -0,82) (Рисунок 3B).
Следующие семейства: Spirochaetaceae, Marinifilaceae, Ruminococcaceae, Psychromonadaceae и Lachnospiraceae – были среди тех, у которых наблюдались наибольшие изменения относительной численности с расстоянием от садков до референтного участка на основе теста индикаторных видов и графиков (Рисунок 4; Дополнительные файлы 6, 8, 9). Та же тенденция наблюдалась для дельтапротеобактерий, в то время как гаммапротеобактерии показали противоположную картину (Дополнительные файлы 7, 9). На участке 600 м в период «пик биомассы» некоторые из этих бактериальных семейств снова увеличили свою численность.
Поверхностный слой и слой 3 см показали сходные тенденции. Оба слоя демонстрировали большую вариабельность микробного биоразнообразия и численности (Дополнительный файл 9) как внутри, так и между периодами отбора и участками. Слой 10 см показал более равномерное распределение по всем участкам и периодам отбора. Анализ индикаторных таксонов по слоям осадка показал, что поверхностный слой и слой 3 см имели более сходные индикаторные таксоны по сравнению с комбинацией других двух слоёв (слой 3+10 см, поверхностный слой+слой 10 см). Анализ «indicspecies» выявил множество индикаторных таксонов как по расстоянию, так и по слоям, последовательно выделяя вышеупомянутые бактериальные семейства (например, Spirochaetaceae, Marinifilaceae, Psychromonadaceae).
Обсуждение
Микробное разнообразие
Богатство таксонов было значительно выше во всех трёх слоях осадка в период «до зарыбления» (Рисунок 2), а структура микробного сообщества значительно различалась между периодами «до зарыбления» и «пик биомассы» (Рисунок 3). Это согласуется с недавними исследованиями, в которых обнаружена общая тенденция увеличения богатства от края садков к референтным участкам (Verhoeven и др., 2018; Dully и др., 2021; Frühe и др., 2021b). Однако другие исследования сообщали о противоположных тенденциях в структуре богатства по сравнению с нашими результатами (Bissett и др., 2007; Kawahara и др., 2009; Frühe и др., 2021b) или об отсутствии изменений (Zhang и др., 2008; Kawahara и др., 2009). Эти противоречия, вероятно, возникают из-за сложной взаимосвязи экологических параметров, влияющих на микробное разнообразие и состав сообществ (Hornick и Buschmann, 2018; Dully и др., 2021), включая такие факторы, как сезонные колебания, гидрология, биогеографические регионы, текстура осадка и условия, специфичные для конкретной фермы (Frühe и др., 2021b).
Микробное богатство также демонстрировало устойчивую картину увеличения с удалением от садков (Рисунок 2), особенно в слоях 0 см и 3 см в период «до зарыбления». Эти закономерности являются чёткими признаками того, что микробные сообщества в осадках подвержены влиянию аквакультуры, скорее всего, из-за уменьшения воздействия аквакультуры с расстоянием от источника органического материала. Поскольку более высокое богатство с расстоянием наиболее выражено в период «до зарыбления», мы полагаем, что, хотя это и период наименьшего воздействия в цикле лососёвого хозяйства, микробные сообщества не успевают полностью восстановиться до повторного зарыбления садков.
Пробы «до зарыбления» также показали высокое стандартное отклонение среди повторностей (Рисунок 2), что может объясняться некоторой гетерогенностью осадка, которая становится более заметной по мере увеличения альфа-разнообразия, т.е. микросреды могут быть распределены неравномерно в трёх повторностях с каждой глубины осадка. Однако самое высокое значение богатства по крайней мере на порядок выше самого низкого, что делает наши результаты надёжными. Хотя Сёрвогсфьорд является очень динамичным фьордом, и считается, что воздействие органического поступления от аквакультуры на фарерские фьорды не превышает 200 м (Mortensen и др., 2021), эти результаты указывают на обратное, по крайней мере в отношении микробных сообществ. Исследования, проведённые в других местах, показывают, что большая дисперсия на дисперсионных участках приводит к более крупным бентосным следам, простирающимся до 300-500 м от фермы (Keeley и др., 2013b; Keeley и др., 2013a; Keeley и др., 2019), по сравнению с недисперсионными участками, где воздействие обычно находится в пределах 150 м (Brooks и др., 2002; Forrest и др., 2007; Keeley и др., 2019). Значительно различающаяся структура микробных сообществ между периодами отбора, включая слой 10 см, указывает на значительное воздействие на микробные сообщества, простирающееся как минимум на 10 см ниже поверхности осадка (Рисунок 3A). Хотя среда обитания в прибрежных илистых осадках естественно претерпевает значительные вертикальные изменения, переходя от аэробных условий к анаэробным сульфидным (Jørgensen и др., 2019), не предполагается, что осадки перемешиваются до глубины 10 см. Однако эти результаты, скорее всего, обусловлены органическим обогащением на поверхности, которое каким-то образом влияет на биогеохимические процессы в глубине осадков.
Микроорганизмы, связанные с лососёвым хозяйством
Органическое обогащение влияет на биогеохимические процессы и микробное сообщество в осадке под рыбоводными садками, что согласуется с предыдущими исследованиями (Holmer и др., 2005; Bissett и др., 2007; Dully и др., 2021). Были выявлены значительные различия между микробными сообществами в периоды отбора (Рисунок 3A), что подтверждает предыдущие исследования (Bissett и др., 2007 и Hornick и Buschmann, 2018); однако несколько групп показали некоторые особенности. Гаммапротеобактерии доминировали в более удалённых районах (190-600 м), в то время как дельтапротеобактерии были более многочисленны ближе к садкам (0-12 м), что согласуется с выводами Kawahara и др. (2009), Stoeck и др. (2018) и Dully и др. (2021). Эта закономерность наблюдалась преимущественно в поверхностном слое осадка, что предполагает, что поверхностные осадки могут быть более чувствительны к изменениям, исходящим от садков сверху.
Бактериальные семейства Spirochaetaceae, Clostridia (Lachnospiraceae, Ruminococcaceae и Defluviitaleaceae) и Psychromonadaceae показали более высокую относительную численность на участках 0-12 м как «до зарыбления», так и в период «пик биомассы» (Рисунок 4; Дополнительный файл 8), что согласуется с предыдущими исследованиями (Verhoeven и др., 2018; Frühe и др., 2021b; Miranda и др., 2021). Frühe и др. (2021a) идентифицировали Psychromonadaceae, Ruminococcaceae и Spirochaetaceae как индикаторы «высокого воздействия», что согласуется с нашими данными вблизи лососёвых садков (Рисунок 4; Дополнительный файл 8). Некоторые бактериальные семейства, которые Frühe и др. (2021a) обнаружили наиболее обильными вдали от садков, в нашем исследовании были более распространены на умеренных расстояниях (40-150 м), например Pirellulaceae (Дополнительный файл 8). Референтный участок, расположенный в 600 м от садков, находился ближе к берегу на меньших глубинах (≈35 м против ≈45 м на других участках), что потенциально могло повлиять на результаты. Таким образом, по сравнению с выводами Frühe (2021), референтный участок может не точно отражать такие бактерии, как Pirellulaceae, но всё же, по-видимому, подвержен некоторому влиянию аквакультурной деятельности. Тем не менее, соотношение гамма/дельтапротеобактерий (Рисунок 3B) показало устойчивую картину на всех участках отбора, при этом гаммапротеобактерии достигали пика в референтной точке, где дельтапротеобактерии были наименее многочисленны (Дополнительный файл 7). Однородность соотношения для обоих периодов отбора подтверждает пригодность референтного участка. Более того, выраженное различие между двумя периодами отбора подчёркивает влияние на референтную пробу. Референтный участок соответствует требованиям ASC (Совет по управлению аквакультурой) (>500 м) (Mortensen и др., 2021). Однако перенос таких мер на аквакультурные участки по всему миру может быть сложным, поскольку фарерские фьорды являются высокодинамичными. Кроме того, фарерские фьорды невелики, что затрудняет поиск мест на расстоянии более 500 м, которые не были бы слишком близко к берегу или в открытом океане для сбора проб.
Verhoeven и др. (2018) сообщили о крупномасштабных сдвигах на уровне типов, включая увеличение Bacteroidetes, Firmicutes и Spirochaetes, а также уменьшение Proteobacteria и Epsilonbacteraeota вблизи лососёвых садков, что в целом согласуется с нашими выводами (Рисунок 4; Дополнительные файлы 6, 8). Эти сдвиги происходили заметно от «до зарыбления» к «пик биомассы», особенно в поверхностном слое (пробы 1+ и 2+). Однако в этом исследовании все участки отбора показали увеличение Epsilonbacteraeota (особенно семейства Sulfurovaceae) от «до зарыбления» к «пик биомассы», что отличается от Verhoeven и др. (2018), но согласуется с Keeley и др. (2021) (Дополнительный файл 6). Sulfurovaceae – это серопродуцирующие бактерии, играющие ключевую роль в циклах серы, азота и водорода в морских осадках с высоким органическим обогащением (Miranda и др., 2021), что указывает на значительное поступление органического материала на многие из исследованных участков.
Примечательно, что Bacteroidia увеличивались по мере роста биомассы, особенно вблизи садков в период «пик биомассы», при этом несколько семейств Bacteroidia были заметно обильны в пробах на 0-12 м, включая Bacteroidetes-BD2-2, Lentimicrobiaceae, Prolixibacteraceae, Marinifilaceae, в то время как Flavobacteriaceae показали более низкую относительную численность. Этот результат согласуется с Miranda и др. (2021), но противоречит Frühe (2021). Противоречивые результаты можно объяснить метаболической универсальностью Flavobacteriaceae – таксономической группы, широко распространённой в различных средах (Dully и др., 2021; Frühe и др., 2021b; Miranda и др., 2021). Эта универсальность Flavobacteriaceae подтверждается на уровне родов, где различные виды Flavobacteriaceae доминировали в разных местах (Дополнительный файл 10). Lutibacter демонстрирует более высокую численность ближе к садкам, в то время как Actibacter преобладает в более отдалённых точках. Lutimonas оказался преобладающим родом Flavobacteriaceae на всех участках отбора, сохраняя стабильную численность, за исключением пробы 1+, где она была заметно ниже. Leontidou и др. (2023) выделили Lutimonas как потенциальный биоиндикатор ухудшения качества окружающей среды из-за органического обогащения. Для получения конкретных выводов о закономерностях распределения родов Flavobacteriaceae необходимы дальнейшие исследования.
Использование микроорганизмов в качестве экологического индикатора
Наше исследование подчёркивает изменения в структуре микробного сообщества в течение одного цикла выращивания лосося, особенно вблизи садков (0-12 м). Эти структурные изменения могут быть связаны с размером лосося, так как более крупный лосось выделяет больше фекального материала и требует больше корма для роста, что приводит к увеличению накопления органического вещества (ОВ) на морском дне (Quero и др., 2020). Хотя различные неисследованные факторы, такие как накопление ОВ, ресуспензия осадка, сезонность и биотические взаимодействия, могут вносить вклад в вариации прокариотических ассамбляжей, данное исследование демонстрирует изменения микробных сообществ за один цикл лососёвого хозяйства («до зарыбления» и «пик биомассы»). Для более глубокого понимания необходимо дальнейшее исследование этих факторов. Однако корреляция соотношения гамма/дельтапротеобактерий (Рисунок 3B) с расстоянием предполагает, что участок отбора «до зарыбления» всё ещё может находиться под влиянием предыдущего производственного цикла, что указывает на потенциально недостаточный 3-месячный период залежи. Одновременно с этим исследованием оценка экологического состояния показала «некоторое воздействие» в оба периода отбора. Однако экологическая оценка, проведённая при половине от ожидаемой максимальной биомассы (март 2020 г., между двумя нашими отборами), показала «отсутствие воздействия» на осадок (неопубликованные данные). Это подтверждается тем, что последующий анализ показал, что проба на 0 м в «до зарыбления» значительно отличалась от всех остальных проб, за исключением пробы на 12 м «до зарыбления», что позволяет предположить, что участок на 0 м мог не полностью восстановиться после предыдущего(их) цикла(ов) (Дополнительный файл 5). Одним из возможных объяснений являются открытые условия выращивания на Фарерских островах (ICES, 2023) и сезонные колебания в течение производственного цикла (сентябрь 2019 г. – июль 2020 г.).
В зимний период этот район подвержен воздействию океанской зыби, при этом высота значительных волн достигает 5-6 метров. Исследования показывают, что океанская зыбь может вызывать ресуспензию осадков, подверженных влиянию рыбоводства, значительно улучшая условия осадка, особенно когда высота волн превышает 1,5 м, а глубина составляет менее 50 м (Norði и Patursson, 2012). Снижение биомассы и кормовой активности в зимний период приводит к меньшему накоплению ОВ на морском дне. Учёт сезонных колебаний и местных факторов (например, глубины, температуры, течений, волн, ветра, приливов, условий дна) необходим при определении периода залежи. Это подтверждает полезность микробных сообществ для оценки этих динамичных и открытых участков выращивания, учитывая их быстрое время реакции, что делает их чувствительными к изменениям окружающей среды и, следовательно, эффективным индикатором здоровья осадков. Для будущих исследований приоритетное использование множества референтных участков может иметь решающее значение для эффективного мониторинга данных микробных сообществ как индикаторов здоровья экосистем. Необходимы надёжные базовые данные, отражающие воздействие на окружающую среду без антропогенного влияния. В этом исследовании расположение референтного участка ближе к берегу и на меньшей глубине могло повлиять на результаты. Общая тенденция, иллюстрируемая, например, соотношением гамма/дельтапротеобактерий, предполагает некоторый уровень воздействия. Однако определить, обусловлено ли это воздействие аквакультурной деятельностью или близостью референтного участка к берегу и связанными с этим влияниями, остаётся сложной задачей, которая должна стать приоритетом для будущих исследований.
Приоритетное использование открытых участков для выращивания может помочь снизить воздействие на окружающую среду, поскольку ОВ становится более рассредоточенным по большей площади (Keeley и др., 2013b; Keeley и др., 2019). Переход от защищённых участков к местам с сильными течениями и волновым воздействием может улучшить качество воды, что приведёт к повышению насыщения кислородом и усилению водообмена. Это, в свою очередь, снизит инфекционную нагрузку благодаря более быстрому рассеиванию морских вшей в районах с сильными течениями (Kragesteen и др., 2018; Johannesen и др., 2020). Однако важно признать, что открытые условия предъявляют более высокие требования к садкам и могут повлечь непредвиденные затраты для рыбоводных ферм (Johannesen и др., 2020).
Мониторинг окружающей среды на Фарерских островах включает различные измерения, включая pH, окислительно-восстановительный потенциал, химию ОВ, концентрации меди и цинка, сенсорные показатели цвета и запаха, а также исследования макробеспозвоночных. Хотя некоторые из этих стандартов являются субъективными, они хорошо интегрированы в местные процедуры и циклы аквакультуры. Для оптимизации методологий экологической оценки анализ микробных сообществ представляет собой перспективную альтернативу для будущих исследований. Микроорганизмы быстро реагируют на изменения окружающей среды, опережая макробеспозвоночных в чувствительности к таким колебаниям. Эти анализы предоставляют больше информации с использованием меньшего количества материала (примерно 3 г осадка по сравнению с 500-1000 г) и, при расширении на другие генетические маркеры (например, 18S рРНК, 12S рРНК, COI), потенциально могут дать оценки разнообразия макроорганизмов, присутствующих в той же пробе. Кроме того, это оказывается экономически эффективным, поскольку предоставляет более полные данные по более низкой стоимости по сравнению с традиционными методами (Dully и др., 2021).
Одним из текущих недостатков является время, необходимое для получения результатов секвенирования, которое часто занимает от нескольких недель до месяцев. Однако при интеграции в стандартные процедуры обследования и выделении соответствующих ресурсов (включая персонал, оборудование и биоинформатические конвейеры) можно сократить это время до нескольких дней или до режима реального времени с использованием таких методов, как секвенирование нанопор. Для полной замены текущих методов ДНК-метабаркодингом необходимы национальные и международные руководства для сравнения результатов между странами и аквакультурными фермами. Эти стандартизированные методы должны включать базы данных индикаторных ASV, ориентированных на микробные ASV как универсальные биоиндикаторы, чувствительные к воздействию лососёвых ферм, независимо от геопространственных эффектов или естественной сезонности (Keeley и др., 2018; Dully и др., 2021; Frühe и др., 2021a). ДНК-метабаркодинг обладает потенциалом стать мощным инструментом для корректировки практики аквакультуры в сторону большей устойчивости, рентабельности и экологичности.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fmars.2024.1266410/full#supplementary-material
Ссылки
1. Agency. (2018). Environmental Surveillance of Aquaculture at Sea. Available online at: https://www.us.fo/vernd/a-sjogvi/. CrossRef. Google Scholar.
2. Ape F., Manini E., Quero G. M., Luna G. M., Sarà G., Vecchio P., et al. (2019). Biostimulation of in situ microbial degradation processes in organically-enriched sediments mitigates the impact of aquaculture. Chemosphere 226, 715–725. doi: 10.1016/j.chemosphere.2019.03.178. CrossRef. Google Scholar.
3. Apprill A., McNally S., Parsons R., Weber L. (2015). Minor revision to V4 region SSU rRNA 806R gene primer greatly increases detection of SAR11 bacterioplankton. Aquat. Microbial Ecol. 75, 129–137. doi: 10.3354/ame01753. CrossRef. Google Scholar.
4. Asami H., Aida M., Watanabe K. (2005). Accelerated sulfur cycle in coastal marine sediment beneath areas of intensive shellfish aquaculture. Appl. Environ. Microbiol. 71, 2925–2933. doi: 10.1128/AEM.71.6.2925-2933.2005. CrossRef. Google Scholar.
5. Bannister R. J., Johnsen I. A., Hansen P. K., Kutti T., Asplin L. (2016). Near- and far-field dispersal modelling of organic waste from Atlantic salmon aquaculture in fjord systems. ICES J. Mar. Sci. 73, 2408–2419. doi: 10.1093/icesjms/fsw027. CrossRef. Google Scholar.
6. Bannister R., Valdemarsen T., Hansen P., Holmer M., Ervik A. (2014). Changes in benthic sediment conditions under an Atlantic salmon farm at a deep, well-flushed coastal site. Aquaculture Environ. Interact. 5, 29–47. doi: 10.3354/aei00092. CrossRef. Google Scholar.
7. Bissett A., Burke C., Cook P. L. M., Bowman J. P. (2007). Bacterial community shifts in organically perturbed sediments. Environ. Microbiol. 9, 46–60. doi: 10.1111/j.1462-2920.2006.01110.x. CrossRef. Google Scholar.
8. Black K. D., Kiemer M. C. B., Ezzi I. A. (1996). The relationships between hydrodynamics, the concentration of hydrogen sulphide produced by polluted sediments and fish health at several marine cage farms in Scotland and Ireland. J. Appl. Ichthyology 12, 15–20. doi: 10.1111/j.1439-0426.1996.tb00053.x. CrossRef. Google Scholar.
9. Brooks K. M., Mahnken C., Nash C. (2002). Environmental effects associated with marine netpen waste with emphasis on salmon farming in the Pacific northwest. Responsible Marine Aquaculture. CABI Publishing, pp. 159–203. doi: 10.1079/9780851996042.0000. CrossRef. Google Scholar.
10. Brown J. R., Gowen R. J., McLusky D. S. (1987). The effect of salmon farming on the benthos of a Scottish sea loch. J. Exp. Mar. Biol. Ecol. 109, 39–51. doi: 10.1016/0022-0981(87)90184-5. CrossRef. Google Scholar.
11. Buschmann A. H., Riquelme V. A., Hernández-González M. C., Varela D., Jiménez J. E., Henríquez L. A., et al. (2006). A review of the impacts of salmonid farming on marine coastal ecosystems in the southeast Pacific. ICES J. Mar. Sci. 63, 1338–1345. doi: 10.1016/j.icesjms.2006.04.021. CrossRef. Google Scholar.
12. Cáceres M. D., Legendre P. (2009). Associations between species and groups of sites: indices and statistical inference. Ecology 90, 3566–3574. doi: 10.1890/08-1823.1. CrossRef. Google Scholar.
13. Callahan B. J., McMurdie P. J., Rosen M. J., Han A. W., Johnson A. J. A., Holmes S. P. (2016). DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 13, 581–583. doi: 10.1038/nmeth.3869. CrossRef. Google Scholar.
14. Carroll M. L., Cochrane S., Fieler R., Velvin R., White P. (2003). Organic enrichment of sediments from salmon farming in Norway: environmental factors, management practices, and monitoring techniques. Aquaculture 226, 165–180. doi: 10.1016/S0044-8486(03)00475-7. CrossRef. Google Scholar.
15. Djurhuus A., Port J., Closek C. J., Yamahara K. M., Romero-Maraccini O., Walz K. R., et al. (2017). Evaluation of filtration and DNA extraction methods for environmental DNA biodiversity assessments across multiple trophic levels. Front. Mar. Sci. 4. doi: 10.3389/fmars.2017.00314. CrossRef. Google Scholar.
16. Dully V., Balliet H., Frühe L., Däumer M., Thielen A., Gallie S., et al. (2021). Robustness, sensitivity and reproducibility of eDNA metabarcoding as an environmental biomonitoring tool in coastal salmon aquaculture – An inter-laboratory study. Ecol. Indic. 121, 107049. doi: 10.1016/j.ecolind.2020.107049. CrossRef. Google Scholar.
17. FAO. (2018). World Review. In The State of World Fisheries and Aquaculture. 1–83. doi: 10.18356/eeca78e4-en. CrossRef. Google Scholar.
18. FAO. (2022). The State of World Fisheries and Aquaculture 2022: Towards Blue Transformation. (Rome: FAO) 2022266. doi: 10.4060/cc0461en. CrossRef. Google Scholar.
19. Folke C., Kautsky N. (1989). The role of ecosystems for sustainable development of aquaculture. AMBIO 18, 234–243. Google Scholar.
20. Forrest B., Keeley N., Gillespie P., Hopkins G., Knight B., Govier D. (2007). Review of the Ecological Effects of Marine Finfish Aquaculture: Final Report. Available online at: https://www.cawthron.org.nz/media_new/publications/pdf/2014_07/CR1285-Review_of_the_ecological_effects_of_marine_finfish_aquaculture_Final_report.pdf. Google Scholar.
21. Frühe L., Cordier T., Dully V., Breiner H., Lentendu G., Pawlowski J., et al. (2021a). Supervised machine learning is superior to indicator value inference in monitoring the environmental impacts of salmon aquaculture using eDNA metabarcodes. Mol. Ecol. 30, 2988–3006. doi: 10.1111/mec.15434. CrossRef. Google Scholar.
22. Frühe L., Dully V., Forster D., Keeley N. B., Laroche O., Pochon X., et al. (2021b). Global trends of benthic bacterial diversity and community composition along organic enrich. Front. Microbiol. 12. doi: 10.3389/fmicb.2021.637811. CrossRef. Google Scholar.
23. Gaard E., Norði G. Á., Simonsen K. (2011). Environmental effects on phytoplankton production in a Northeast Atlantic fjord, Faroe Islands. J. Plankton Res. 33, 947–959. doi: 10.1093/plankt/fbq156. CrossRef. Google Scholar.
24. Gowen R. J., Bradbury N. B. (1987). The ecological impact of salmonid farming in coastal waters: a review. Oceanogr Mar. Biol. 25, 563–575. Google Scholar.
25. Hargrave B., Duplisea D., Pfeiffer E., Wildish D. (1993). Seasonal changes in benthic fluxes of dissolved oxygen and ammonium associated with marine cultured Atlantic salmon. Mar. Ecol. Prog. Ser. 96, 249–257. doi: 10.3354/meps096249. CrossRef. Google Scholar.
26. Hargrave B. T., Holmer M., Newcombe C. P. (2008). Towards a classification of organic enrichment in marine sediments based on biogeochemical indicators. Mar. Pollut. Bull. 56, 810–824. doi: 10.1016/j.marpolbul.2008.02.006. CrossRef. Google Scholar.
27. Holmer M., Wildish D., Hargrave B. (2005). Organic enrichment from marine finfish aquaculture and effects on sediment biogeochemical processes. In Hargrave B. T. Environmental Effects of Marine Finfish Aquaculture 181–206. The Handbook of Environmental Chemistry, vol 5M. Berlin, Heidelberg: Springer. doi: 10.1007/b136010. CrossRef. Google Scholar.
28. Hornick K. M., Buschmann A. H. (2018). Insights into the diversity and metabolic function of bacterial communities in sediments from Chilean salmon aquaculture sites. Ann. Microbiol. 68, 63–77. doi: 10.1007/s13213-017-1317-8. CrossRef. Google Scholar.
29. Hyland J., Balthis L., Karakassis I., Magni P., Petrov A., Shine J., et al. (2005). Organic carbon content of sediments as an indicator of stress in the marine benthos. Mar. Ecol. Prog. Ser. 295, 91–103. doi: 10.3354/meps295091. CrossRef. Google Scholar.
30. ICES. (2023). Workshop on the Faroes Ecoregion Aquaculture Overview (WKFaroesAO) (Copenhagen: ICES Scientific Reports), 87. doi: 10.17895/ices.pub.21551541. CrossRef. Google Scholar.
31. Johannesen Á., Patursson Ø., Kristmundsson J., Dam S. P., Klebert P. (2020). How caged salmon respond to waves depends on time of day and currents. PeerJ 8, e9313. doi: 10.7717/peerj.9313. CrossRef. Google Scholar.
32. Jørgensen B. B., Findlay A. J., Pellerin A. (2019). The biogeochemical sulfur cycle of marine sediments. Front. Microbiol. 10. doi: 10.3389/fmicb.2019.00849. CrossRef. Google Scholar.
33. Karakassis I., Tsapakis M., Hatziyanni E., Papadopoulou K.-N., Plaiti W. (2000). Impact of cage farming of fish on the seabed in three Mediterranean coastal areas. ICES J. Mar. Sci. 57, 1462–1471. doi: 10.1006/jmsc.2000.0925. CrossRef. Google Scholar.
34. Kawahara N., Shigematsu K., Miyadai T., Kondo R. (2009). Comparison of bacterial communities in fish farm sediments along an organic enrichment gradient. Aquaculture 287, 107–113. doi: 10.1016/j.aquaculture.2008.10.003. CrossRef. Google Scholar.
35. Keeley N., Cromey C., Goodwin E., Gibbs M., Macleod C. (2013a). Predictive depositional modelling (DEPOMOD) of the interactive effect of current flow and resuspension on ecological impacts beneath salmon farms. Aquaculture Environ. Interact. 3, 275–291. doi: 10.3354/aei00068. CrossRef. Google Scholar.
36. Keeley N., Forrest B. M., Macleod C. K. (2013b). Novel observations of benthic enrichment in contrasting flow regimes with implications for marine farm monitoring and management. Mar. Pollut. Bull. 66, 105–116. doi: 10.1016/j.marpolbul.2012.10.024. CrossRef. Google Scholar.
37. Keeley N., Laroche O., Birch M., Pochon X. (2021). A substrate-independent benthic sampler (SIBS) for hard and mixed-bottom marine habitats: A proof-of-concept study. Frontiers 8. doi: 10.3389/fmars.2021.627687. CrossRef. Google Scholar.
38. Keeley N., Valdemarsen T., Woodcock S., Holmer M., Husa V., Bannister R. (2019). Resilience of dynamic coastal benthic ecosystems in response to large-scale finfish farming. Aquaculture Environ. Interact. 11, 161–179. doi: 10.3354/aei00301. CrossRef. Google Scholar.
39. Keeley N., Wood S. A., Pochon X. (2018). Development and preliminary validation of a multi-trophic metabarcoding biotic index for monitoring benthic organic enrichment. Ecol. Indic. 85, 1044–1057. doi: 10.1016/j.ecolind.2017.11.014. CrossRef. Google Scholar.
40. Kragesteen T., Simonsen K., Visser A., Andersen K. (2018). Identifying salmon lice transmission characteristics between Faroese salmon farms. Aquaculture Environ. Interact. 10, 49–60. doi: 10.3354/aei00252. CrossRef. Google Scholar.
41. Kristensen E. (2000). Organic matter diagenesis at the oxic:anoxic interface in coastal marine sediments, with emphasis on the role of burrowing animals. Hydrobiologia 426, 1–24. doi: 10.1023/A:1003980226194. CrossRef. Google Scholar.
42. Leontidou K., Rubel V., Stoeck T. (2023). Comparing quantile regression spline analyses and supervised machine learning for environmental quality assessment at coastal marine aquaculture installations. PeerJ 11, e15425. doi: 10.7717/peerj.15425. CrossRef. Google Scholar.
43. Love M. I., Huber W., Anders S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550. doi: 10.1186/s13059-014-0550-8. CrossRef. Google Scholar.
44. McCaig A. E., Phillips C. J., Stephen J. R., Kowalchuk G. A., Harvey S. M., Herbert R. A., et al. (1999). Nitrogen cycling and community structure of proteobacterial beta-subgroup ammonia-oxidizing bacteria within polluted marine fish farm sediments. Appl. Environ. Microbiol. 65, 213–220. doi: 10.1128/AEM.65.1.213-220.1999. CrossRef. Google Scholar.
45. McMurdie P. J., Holmes S. (2013). phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PLoS One 8, e61217. doi: 10.1371/journal.pone.0061217. CrossRef. Google Scholar.
46. McMurdie P. J., Holmes S. (2014). Waste not, want not: why rarefying microbiome data is inadmissible. PLoS Comput. Biol. 10, e1003531. doi: 10.1371/journal.pcbi.1003531. CrossRef. Google Scholar.
47. Miranda R. M., Aguila-Torres P., Aranda C. P., Maldonado J., Casado A. (2021). Taxonomy and diversity of bacterial communities associated with marine sediments from Chilean salmonid farms. Aquaculture Res. 52, 1605–1620. doi: 10.1111/are.15014. CrossRef. Google Scholar.
48. Mortensen H., Carstensen J., Andreasen B., Johannesen T. T., Fjallstein B. V. T., Norði G. á. (2021). Benthic macrofauna classification system for Faroese fjords Vol. Fiskaaling Rit 2021-10 (Hvalvík: Fiskaaling), 54. Google Scholar.
49. Neofitou N., Vafidis D., Klaoudatos S. (2010). Spatial and temporal effects of fish farming on benthic community structure in a semi-enclosed gulf of the Eastern Mediterranean. AQUACULTURE Environ. Interact 1, 95-105. doi: 10.3354/aei00010. CrossRef. Google Scholar.
50. Norði G. á., Glud R. N., Gaard E., Simonsen K. (2011). Environmental impacts of coastal fish farming: carbon and nitrogen budgets for trout farming in Kaldbaksfjør 7 ur (Faroe Islands). Mar. Ecol. Prog. Ser. 431, 223–241. doi: 10.3354/meps09113. CrossRef. Google Scholar.
51. Norði G., Lund I., Andreasen B., Taylor D., Johannesen T. T., Jacobsen B., et al. (2023). Modeling particulate waste assimilation by blue mussels within the spatial constraints of a commercial fish farm: implications for multitrophic aquaculture. Front. Mar. Sci. 10. doi: 10.3389/fmars.2023.1236294. CrossRef. Google Scholar.
52. Norði G., Patursson Ø. (2012). Influence of waves and current speed on resuspension of fish farm waste: Case study in Funningsfjørður (Faroe Islands). ICES CM 2012/Q:13. Google Scholar.
53. Oksanen J., Blanchet F. G., Kindt R., Legendre P. (2013). Vegan: Community Ecology. R. Package Version 2.5-7. Google Scholar.
54. Parada A. E., Needham D. M., Fuhrman J. A. (2015). Every base matters: assessing small subunit rRNA primers for marine microbiomes with mock communities, time series and global field samples. Environ. Microbiol. 18, 1403–1414. doi: 10.1111/1462-2920.13023. CrossRef. Google Scholar.
55. Pawlowski J., Bruce K., Panksep K., Aguirre F. I., Amalfitano S., Apothéloz-Perret-Gentil L., et al. (2022). Environmental DNA metabarcoding for benthic monitoring: A review of sediment sampling and DNA extraction methods. Sci. Total Environ. 818, 151783. doi: 10.1016/j.scitotenv.2021.151783. CrossRef. Google Scholar.
56. Price C., Black K., Hargrave B., Morris J. (2015). Marine cage culture and the environment: effects on water quality and primary production. Aquaculture Environ. Interact. 6, 151–174. doi: 10.3354/aei00122. CrossRef. Google Scholar.
57. Quero G. M., Ape F., Manini E., Mirto S., Luna G. M. (2020). Temporal changes in microbial communities beneath fish farm sediments are related to organic enrichment and fish biomass over a production cycle. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.00524. CrossRef. Google Scholar.
58. R Core Team. (2020). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available at: https://www.R-project.org/. Google Scholar.
59. Schendel E., Nordstrom S., Lavkulich L. (2004). Floc and sediment properties and their environmental distribution from a marine fish farm. Aquaculture Res. 35, 483–493. doi: 10.1111/j.1365-2109.2004.01042.x. CrossRef. Google Scholar.
60. Stoeck T., Frühe L., Forster D., Cordier T., Martins C. I. M., Pawlowski J. (2018). Environmental DNA metabarcoding of benthic bacterial communities indicates the benthic footprint of salmon aquaculture. Mar. Pollut. Bull. 127, 139–149. doi: 10.1016/j.marpolbul.2017.11.065. CrossRef. Google Scholar.
61. Sweetman A. K., Norling K., Gunderstad C., Haugland B. T., Dale T. (2014). Benthic ecosystem functioning beneath fish farms in different hydrodynamic environments. Limnology Oceanography 59, 1139–1151. doi: 10.4319/lo.2014.59.4.1139. CrossRef. Google Scholar.
62. Verhoeven J. T. P., Salvo F., Knight R., Hamoutene D., Dufour S. C. (2018). Temporal bacterial surveillance of salmon aquaculture sites indicates a long lasting benthic impact with minimal recovery. Front. Microbiol. 9. doi: 10.3389/fmicb.2018.03054. CrossRef. Google Scholar.
63. Zhang W., Song L., Ki J.-S., Lau C.-K., Li X.-D., Qian P.-Y. (2008). Microbial diversity in polluted harbor sediments II: Sulfate-reducing bacterial community assessment using terminal restriction fragment length polymorphism and clone library of dsrAB gene. Estuarine Coast. Shelf Sci. 76, 682–691. doi: 10.1016/j.ecss.2007.07.039. CrossRef. Google Scholar.

Johansen BO, Mikalsen S-O, Magnussen E, Patursson EJ, á Norði G and Djurhuus A (2024) Impacts on microbial communities in sediments by aquaculture farming during one salmon cycle. Front. Mar. Sci. 11:1266410. doi: 10.3389/fmars.2024.1266410
Перевод статьи «Impacts on microbial communities in sediments by aquaculture farming during one salmon cycle» авторов Johansen BO, Mikalsen S-O, Magnussen E, Patursson EJ, á Norði G and Djurhuus A., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык
Фото: Designed by Freepik























































































































Комментарии (0)