Почему компост с биоуглём дают супер-урожай
Органическое земледелие исключает использование синтетических удобрений и обещает производство продуктов питания с минимальным воздействием на окружающую среду, однако эта агропрактика зачастую не обеспечивает такую же продуктивность, как традиционное сельское хозяйство. В последние годы биоуголь привлекает всё больше внимания в качестве почвенной добавки, а его покрытие минералами с образованием биоугле-минерального комплекса (БМК) позволяет улучшить удержание углерода и доступность питательных веществ. Тем не менее, мало известно о потенциале БМК в улучшении органического земледелия. Поэтому в данном исследовании мы изучили, как реагируют почвенные, бактериальные и растительные показатели на совместное внесение БМК и органического удобрения — компоста на основе куриного помёта.
Аннотация
В вегетационном опыте с пекинской капустой БМК и компост показали синергетический эффект на свойства почвы, в частности, за счёт повышения содержания нитратов. Ранее уже было показано, что почвенные нитраты увеличивают размер листьев, и в нашем исследовании мы действительно наблюдали увеличение площади листьев пекинской капусты при совместном внесении БМК и компостированного куриного помёта. Повышение содержания нитратов в почве также коррелировало с обогащением бактериальными нитрификаторами, обусловленным действием БМК. Кроме того, мы отметили, что бактерии, присутствовавшие при внесении компоста, характеризовались высокой скоростью оборота, что, вероятно, способствовало разложению органического вещества и снижению численности потенциальных патогенов, происходящих из помёта. В целом, наши результаты показывают, что сочетание БМК и компоста может стимулировать микробные процессы в органическом земледелии, что приводит к повышению урожайности овощей и улучшению почвенных свойств для устойчивого ведения сельского хозяйства.
Введение
Для сохранения природных ресурсов и обеспечения производства продуктов питания современное сельское хозяйство и horticulture в значительной степени полагаются на почвенные мелиоранты для поддержания и улучшения качества почвы и урожайности растений (Ramankutty and Rhemtulla, 2012). Одним из таких мелиорантов является биоуголь — богатый углеродом твёрдый материал, получаемый путём термической переработки органического сырья в среде с ограниченным доступом воздуха (Chan и др., 2008). Традиционное использование биоугля связывают с высоким плодородием амазонских тёмных земель (ADE; Glaser и др., 2000), а недавние работы показали, что биоуголь из ADE может взаимодействовать с окружающим органическим веществом, золой, глиной и фрагментами костей в процессе выветривания с образованием минеральных агрегатов (Chia и др., 2012). Основываясь на этих данных, в последнее время биоуголь подвергают торрефакции (< 240°C) совместно с измельчёнными горными породами, глинами и другими минералами для формирования биоугле-минеральных комплексов (БМК), которые имитируют минеральные агрегаты биоугля, обнаруженные в ADE (Chia и др., 2010, 2014). Кроме того, процесс производства БМК можно контролировать таким образом, чтобы сохранять до 73% первоначального общего азота, присутствующего в органическом сырье, поскольку азот может включаться в гетероциклические структуры, которые обеспечивают относительно стабильную и медленно высвобождаемую форму азота (Lin и др., 2013). БМК также могут иметь высокие концентрации обменных катионов и среднее время пребывания около 150 лет (Chia и др., 2014), поскольку минеральные добавки значительно повышают стабильность органических компонентов в биоугле (Li и др., 2014).
Хотя БМК обладает потенциалом повышать доступность питательных веществ и связывать углерод в течение длительных периодов времени, его влияние на круговорот питательных веществ в почве, а также его преимущества для роста растений изучены недостаточно. Интересно, что богатые углеродом ADE, как было показано, содержат отличные от соседних почв микробные сообщества, а также более высокое микробное разнообразие и биомассу (O'Neill и др., 2009; Grossman и др., 2010). Также недавно было показано, что БМК при низких нормах внесения (1–5 т га⁻¹) изменяет бактериальные сообщества в современных почвах по сравнению с традиционным удобрением (Nielsen и др., 2014). Учитывая центральную роль бактерий в круговороте питательных веществ в почве и их многочисленные положительные и отрицательные взаимодействия с растениями, эффект БМК в сельскохозяйственных условиях может частично или опосредованно реализовываться через его влияние на саму почвенную микробиоту.
Другой распространённой органической почвенной добавкой является компост, который в основном получают из легкоразлагаемого навоза животных или зелёных отходов и часто используют в органическом земледелии (Rigby and Cáceres, 2001). Компост, как правило, не только богат питательными веществами, но и содержит огромное количество микроорганизмов, состав которых во многом зависит от исходного материала (Sun и др., 2015). Например, компост на основе навоза животных содержит бактерии, происходящие из кишечника животных (Unc and Goss, 2004). Однако вопрос о том, влияет ли внесение этих микроорганизмов из компоста на процессы питания в почве, изучен слабо (Hartmann и др., 2015). Хотя органические удобрения, такие как компост, оказывают менее негативное воздействие на почвенную экосистему, чем синтетические удобрения, сами по себе они не вполне конкурентоспособны с точки зрения урожайности в большинстве агрономических и садоводческих условий (Seufert и др., 2012). Это может быть связано с тем, что питательные вещества компоста либо высвобождаются слишком медленно, либо имеют неоптимальный состав для удовлетворения потребностей растений в питании (Berry и др., 2002).
Учитывая способность БМК влиять на микробные сообщества и процессы в почве (см. выше), мы выдвинули гипотезу о том, что БМК может быть использован для изменения превращения и доступности питательных веществ из компоста, и что это может привести к улучшению роста растений. Для проверки этой гипотезы мы проанализировали урожайность растений, свойства почвы и бактериальные сообщества при обработках либо только БМК, либо только компостом, либо их комбинацией, а также в сравнении с контрольной обработкой без добавок. В качестве модельной системы мы выбрали пекинскую капусту — основную овощную культуру, выращиваемую в Китае в интенсивных садоводческих условиях.
Материалы и методы
Экспериментальный материал
Почва, использованная в этом эксперименте, была отобрана на опытной станции Шанхайского университета Цзяотун, расположенной на острове Чунмин, Шанхай, Китай (31°48′ с.ш., 121°74′ в.д.). На этом участке с 2008 по 2011 год велось овощеводство. В 2011 году почва была обработана компостом, после чего никаких мероприятий не проводилось до момента отбора проб для эксперимента. Образцы почвы из пахотного слоя (0–20 см) были собраны в июле 2014 года после удаления верхней растительности, в основном представленной травами. Почва характеризуется как лёгкий суглинок. Почву просеяли через сито с ячейкой 5 мм и гомогенизировали в лабораторных условиях.
Свежий куриный помёт, полученный от пригородной птицефабрики, компостировали в течение 30 дней и использовали в качестве компоста в данном исследовании. Биоуголь был получен из древесины я oppa (Simcoa Pty. Ltd., Банбери, Западная Австралия) в вертикальном ретортном аппарате с временем пребывания около 12 ч и максимальной температурой 600°C. Этот биоуголь активировали фосфорной кислотой и смешивали с высокожелезистой каолинитовой глиной (30%, масс./масс.), куриным помётом (30%, масс./масс.), фосфоритной мукой, базальтовой пылью, ильменитом и доломитом. Эту смесь подвергали торрефакции при 220°C в течение 3 часов для получения БМК. После пиролиза БМК хранили в стерильных условиях в zip-пакетах. Основные физико-химические свойства почвы, компоста и БМК приведены в Таблице 1.
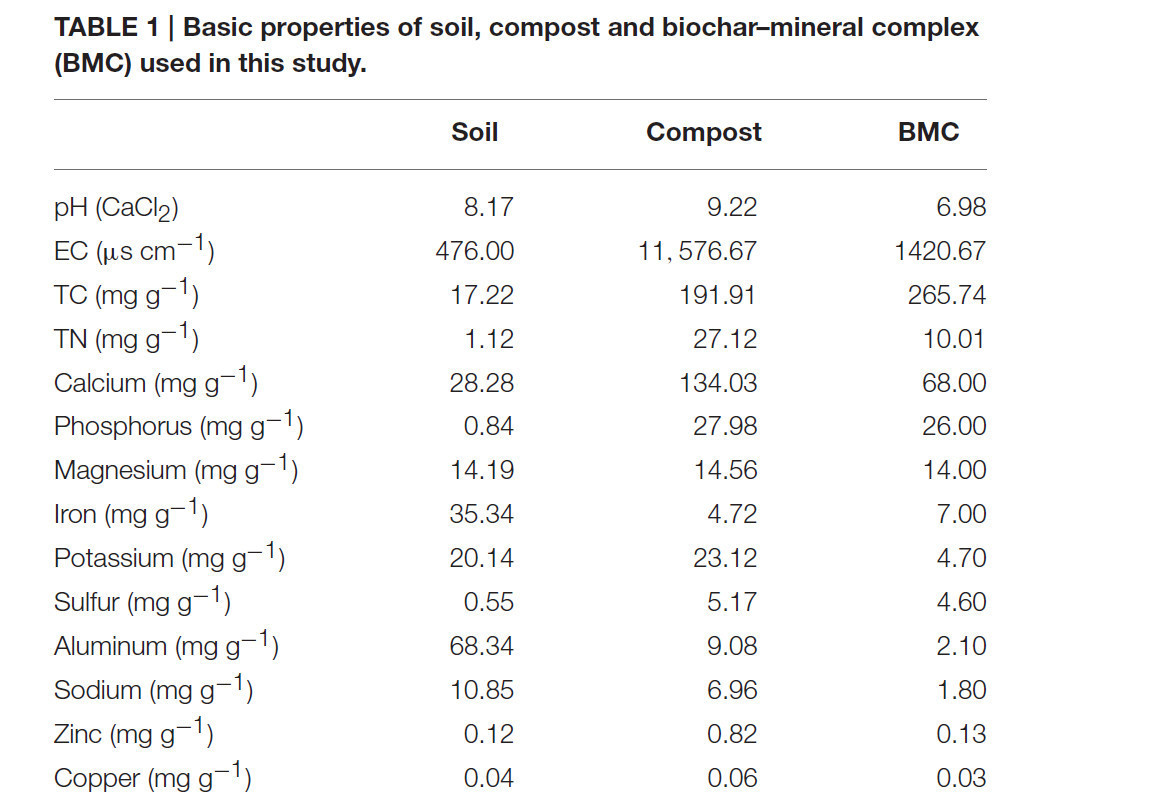
Таблица 1. Основные свойства почвы, компоста и биоугле-минерального комплекса (БМК), использованных в данном исследовании.
Вегетационный опыт
Был проведён вегетационный опыт по полной факторной схеме, включающей следующие четыре обработки: (1) без добавок (CK), (2) 0,1% (масс./масс.) БМК (эквивалентно 1,5 т га⁻¹), (3) 1,9% (масс./масс.) компоста (обработка CO, эквивалентно 28,5 т га⁻¹) и (4) 0,1% (масс./масс.) БМК и 1,9% (масс./масс.) компоста (обработка BMCO). Почву и добавки для каждой обработки гомогенизировали на стерильном лабораторном столе, и из каждой смеси было отобрано по три случайных образца для анализа микробного сообщества на 1-й день. Затем смеси для обработок разделили на 15 повторных горшков (верхний диаметр 10 см, нижний диаметр 9 см, высота 10 см, 500 г почвы на горшок).
В каждый горшок высеяли по двадцать стерильных семян коммерческого сорта пекинской капусты (Brassica rapa L. ssp. chinensis) под названием «Huawang». Горшки были случайным образом размещены в виде матрицы (с интервалом 5 см) внутри климатической камеры. В камере поддерживали дневную температуру 20°C, ночную — 18°C и фотопериод 14 ч с интенсивностью света 300 мкмоль м⁻² с⁻¹ фотосинтетически активной радиации. Полив проводили два раза в неделю. Для достижения реалистичной плотности посадки после достижения всходами стадии одного листа и одной почки (примерно через 6 дней) в каждом горшке оставляли по шесть равномерно развитых сеянцев. В течение периода выращивания дополнительные удобрения не вносили.
Через 40 дней после посадки пекинскую капусту собрали, а всю почву из каждого горшка отобрали. Почву и растения из трёх случайных горшков на обработку объединили, получив пять повторных объединённых проб для дальнейшего анализа. Почву гомогенизировали и пропускали через сито с ячейкой 2 мм для удаления корней и растительных остатков. Образцы растений и почвы разделяли на две аликвоты. Образцы растений использовали либо для физического, либо для химического анализа, тогда как образцы почвы — для химического анализа или анализа микробного сообщества. Все образцы помещали в стерильные пакеты на сухой лёд в момент отбора, а затем хранили при 4 или –80°C в зависимости от требований последующего анализа.
Анализ почвенных и растительных свойств
Влажность почвы определяли путём высушивания свежей почвы при 60°C в течение 4 дней. pH почвы измеряли в суспензии высушенной почвы в 0,01 М растворе хлорида кальция (2:1, объём/масса; Jones, 2001). Электропроводность (EC) измеряли для почвы, суспендированной в деионизированной воде (5:1, объём/масса; Corwin and Lesch, 2005). Общий углерод почвы (TC) и общий азот почвы (TN) измеряли с использованием элементного анализатора vario EL III (Elementar, Германия). Органический углерод почвы (OC) измеряли стандартным методом титрования хромовой кислотой (Walkley and Black, 1934). Доступный фосфор почвы (AP) анализировали в вытяжках 0,5 М бикарбонатом натрия (1:20, объём/масса) спектрофотометрическим методом (Olsen, 1954). Доступный калий почвы (AK) анализировали в вытяжках 1 М ацетатом аммония (1:10, объём/масса) с использованием пламенной атомно-эмиссионной спектроскопии (Brown, 1998). Автоматический дискретный анализатор (Smartchem, Франция) использовали для анализа общего растворимого азота (TSN), нитратов (NO⁻₃) и аммония (NH⁺₄) в почвенных вытяжках 2 М KCl (1:10, объём/масса).
Свежую массу, сухую массу и содержание воды в пекинской капусте определяли путём сравнения массы до и после высушивания при 105°C в течение 30 мин и при 60°C до постоянной массы. Для измерения площади и длины листьев выбирали самый крупный лист каждого растения пекинской капусты. Листья сканировали с помощью сканера Perfection V700 (Epson, США), а полученное изображение обрабатывали с использованием программного обеспечения WinRhizo (Regent Instruments Inc., Канада) для определения длины и площади поверхности листа. Свежую пекинскую капусту использовали для измерения содержания хлорофилла, растворимого белка, растворимых сахаров и нитратов стандартными методами (Li и др., 2000).
Бактериальный анализ
Образцы почвы с начала (День 1) и конца (День 40) периода выращивания подвергали экстракции тотальной ДНК и секвенированию гена 16S рРНК. Тотальную ДНК экстрагировали из 0,5 г просеянной почвы с использованием набора PowerSoil DNA Isolation Kit (MO BIO Laboratories Inc., США) в соответствии с инструкциями производителя. Качество и концентрацию ДНК определяли с помощью Nanodrop 1000 (Thermo Fisher Scientific Inc., США). Участки V1–V3 гена 16S рРНК амплифицировали с использованием штрих-кодированных праймеров 27F (5′-AGAGTTTGATCMTGGCTCAG-3′) и 519R (5′-GWATTACCGCGGCKGCTG-3′), нацеленных на консервативные последовательности, характерные для бактерий. Ампликоны из каждой ПЦР-пробы нормализовали до эквимолярных концентраций и секвенировали на платформе MiSeq (Illumina Inc., США) в Ramaciotti Centre for Genomics (Университет Нового Южного Уэльса, Австралия). Данные секвенирования депонированы в Национальном центре биотехнологической информации (NCBI) под номером BioProject PRJNA297134.
Данные секвенирования гена 16S рРНК обрабатывали с использованием конвейера Mothur (Kozich и др., 2013). Последовательности отбраковывали на уровне контигов, если они содержали основание N или имели более восьми гомополимеров. Очищенные последовательности выравнивали с использованием эталонного выравнивания гена 16S рРНК Silva (Pruesse и др., 2007), отбирали для включения только перекрывающихся участков на основе позиций выравнивания (начало = 1045, конец = 13126), подвергали предварительной кластеризации (diffs = 1) и проверяли на наличие химер с использованием UCHIME (Edgar и др., 2011). Синглетоны были удалены из набора данных для снижения шума, связанного с ошибками секвенирования, возникающими в процессе секвенирования (Reeder and Knight, 2009). Наконец, количество последовательностей было нормализовано путём случайной выборки из каждого образца до наименьшего числа последовательностей, присутствующих во всех сравниваемых образцах (т.е. 5793 отсчёта при сравнении всех образцов или 11076 при сравнении только образцов 40-го дня). Таксономическую принадлежность последовательностей определяли с использованием базы данных Greengenes (версия от мая 2013 г.) с порогом достоверности 60% и кластеризовали в операционные таксономические единицы (OTU) при 97% идентичности с консенсусной таксономией. Любые последовательности, классифицированные как «Mitochondria», «Eukaryotic» или «Chloroplast», а также любые последовательности неизвестного происхождения были удалены из набора данных. Полученный набор данных, прошедший контроль качества, использовали в последующем анализе данных.
Статистический анализ
Экспериментальные обработки были факторизованы на время культивирования (CT), компост (CO) и БМК в следующем анализе. Поскольку растения выращивали в небольших горшках, любые временные изменения в почве могут быть обусловлены как временем, так и внедрением корневой системы. Следовательно, фактор CT включает в себя эффект как времени, так и корневого воздействия. Все факторы имели два уровня: день 1 и день 40 для CT, с добавками и без добавок для CO и БМК.
Для анализа общих эффектов CO и БМК мы сравнили почвенные и растительные переменные с использованием как многомерных, так и одномерных методов. Для многомерного анализа применяли неметрическое главное координатное анализирование (PCoA) к матрице евклидовых расстояний, рассчитанной по нормализованным данным почвы и растений. Двухфакторный пермутационный дисперсионный анализ (PERMANOVA) использовали для проверки эффекта каждого фактора. Для одномерного анализа почвенные переменные и свойства растений подвергали двухфакторному дисперсионному анализу (ANOVA). Кроме того, чистые приросты TC и TN в каждой обработке рассчитывали как среднюю разницу между обработками и контролем с учётом собственного углерода и азота, внесённых с обработками (см. Таблицу 1).
Индексы Chao1 и выравненности Симпсона рассчитывали как показатели α-разнообразия для структуры бактериального сообщества, тогда как β-разнообразие сравнивали с использованием коэффициента сходства Брея–Кёртиса, рассчитанного на основе относительных численностей OTU, преобразованных по квадратному корню. Различия между индексами Chao1 и выравненности Симпсона анализировали с использованием трёхфакторного ANOVA (CT × CO × БМК). Матрицу коэффициентов сходства Брея–Кёртиса визуализировали с использованием неметрического PCoA. Трёхфакторный PERMANOVA применяли к матрице сходства Брея–Кёртиса для проверки уровней значимости различий между экспериментальными факторами и для объяснения вклада каждого фактора в общую дисперсию. Поскольку различительная способность пермутационных анализов для попарных сравнений трёх и пяти повторностей была ограничена 10 и 126 уникальными перестановками соответственно, мы получили попарные сравнения внутригрупповых и межгрупповых сходств Брея–Кёртиса с использованием непараметрического рангового критерия Уилкоксона (Wilcoxon, 1945). Учитывая, что значимые различия, обнаруженные с помощью PERMANOVA, могут быть обусловлены как сходством, так и дисперсией или тем и другим вместе (Clarke and Warwick, 2001), был выполнен тест на однородность дисперсии (PERMDISP) для проверки различий в многомерной дисперсии между группами.
Дистанционные линейные модели (DISTLM) использовали для изучения взаимосвязи между почвенными переменными и бактериальным β-разнообразием (Mcardle and Anderson, 2001). Сначала применяли маргинальный тест, который проверяет каждую переменную отдельно, и последовательный тест, который проверяет каждую переменную после включения всех переменных в одну модель, чтобы изучить прогностический потенциал переменных. В последовательных тестах использовали процедуру прямого отбора и критерий выбора скорректированного R². Далее был реализован пермутационный тест для получения уровня значимости P для этих взаимосвязей. Модель, объясняющая наибольшую долю дисперсии в бактериальном сообществе почвы, была рассчитана с использованием процедуры BEST и критерия выбора скорректированного R². Дистанционный анализ избыточности (dbRDA) с множественными частными корреляциями был выполнен для визуализации модели, полученной процедурой BEST.
OTU, проявившие значимые различия между обработками в каждый момент времени культивирования, были выявлены с использованием двухфакторного ANOVA (CO × БМК) в программе STAMP (Parks и др., 2014). Силу ассоциации каждой OTU с целевыми обработками определяли с использованием индикаторного видового анализа на основе точечно-биссериальных коэффициентов корреляции (De Cáceres and Legendre, 2009). Поскольку OTU могут быть общими для нескольких обработок, были проверены все возможные комбинации априорных групп обработок (De Cáceres и др., 2010). Анализ выполняли с использованием функции multipatt в пакете indicspecies для R. OTU, имеющие < 10 отсчётов последовательностей, были исключены, так как их индикаторный потенциал ограничен. Распределение и силу ассоциации OTU, имеющих относительную численность более 1% хотя бы в одном образце, визуализировали с использованием пакета pheatmap в R.
PERMANOVA, PERMDISP, DISTLM, BEST и dbRDA выполняли в пакете PRIMER v6 (Anderson и др., 2008). Все пермутационные тесты проводились с 10⁵ перестановками. Корректировку P-значений для множественных сравнений проводили с использованием процедуры Бенджамини–Хохберга (FDR) в R с помощью функции p.adjust. Рисунки создавали в R с использованием пакета ggplot2 (Wickham, 2009).
Результаты
Влияние компоста и биоугле-минерального комплекса на почвенные и растительные переменные
Почвенные параметры для всех четырёх обработок (CK, БМК, CO, BMCO) чётко группировались через 40 дней выращивания пекинской капусты (Рисунок 1A), тогда как растительные параметры показывали чёткое разделение только между обработками с компостом и без него (т.е. CK и БМК против CO и BMCO, Рисунок 1B). Хотя наблюдались заметные эффекты на почвенные и растительные переменные, связанные с CO и БМК, значимые взаимодействия были отмечены как для почвенных (P = 0,001), так и для растительных (P < 0,001) параметров (Таблица 2). Однако эффекты взаимодействия были невелики по сравнению с компонентом дисперсии CO, который был как минимум в три раза больше, чем компоненты для БМК или взаимодействия CO × БМК. Более того, попарные сравнения в целом показали, что комбинация БМК и CO (т.е. обработка BMCO) оказывала более выраженные эффекты по сравнению с обработками только БМК или только CO. В совокупности этот статистический анализ показывает, что CO был основным фактором, изменяющим почвенные и растительные параметры, и что БМК играет иную роль при комбинировании с компостом, чем при использовании самостоятельно.
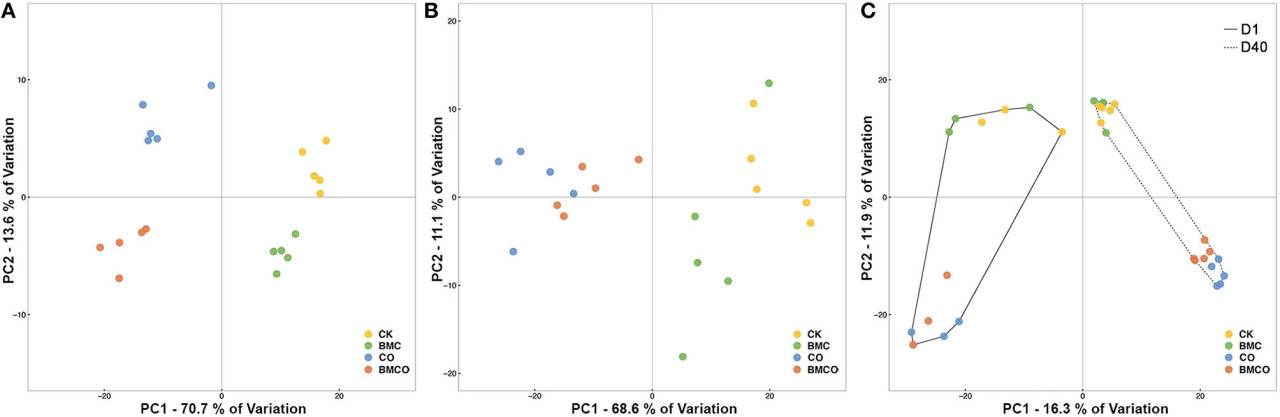
Рисунок 1. Графики неметрического главного координатного анализа (PCoA), иллюстрирующие сходство отдельных образцов на основе: (A) евклидовых расстояний почвенных переменных, (B) евклидова расстояния растительных переменных, (C) расстояния Брея–Кёртиса бактериальных сообществ. Почвенные и растительные переменные были нормализованы перед расчётом евклидова расстояния. Относительная численность бактериальных сообществ была преобразована по квадратному корню перед расчётом расстояния Брея–Кёртиса.
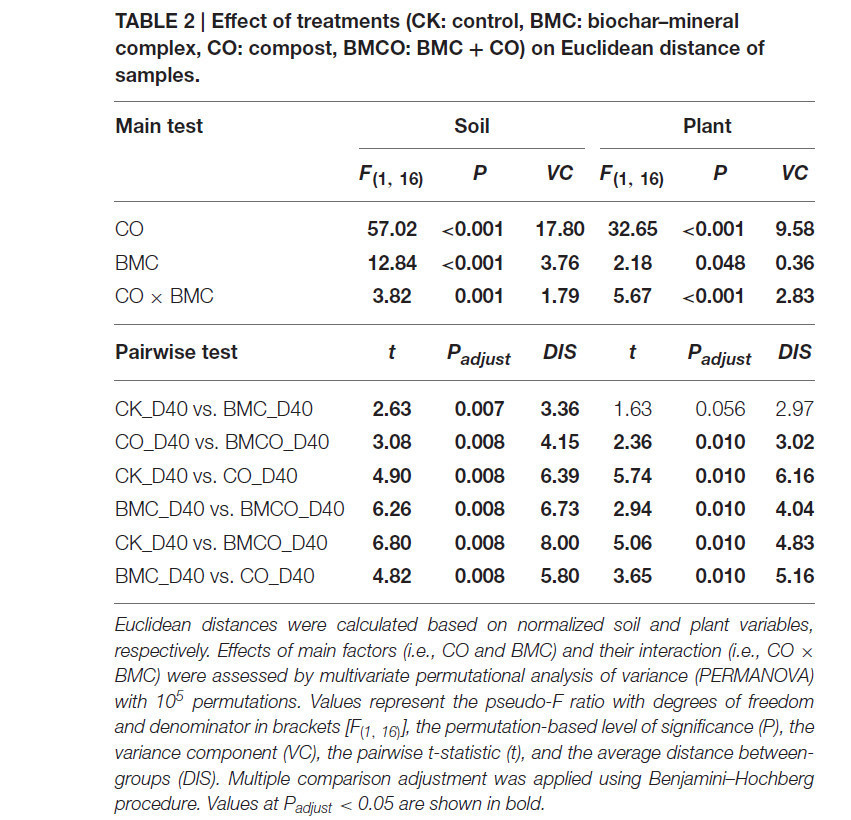
Таблица 2. Влияние обработок (CK: контроль, БМК: биоугле-минеральный комплекс, CO: компост, BMCO: БМК + CO) на евклидово расстояние образцов.
Евклидовы расстояния были рассчитаны на основе нормализованных почвенных и растительных переменных соответственно. Эффекты основных факторов (т.е. CO и БМК) и их взаимодействия (т.е. CO × БМК) оценивались с помощью многомерного пермутационного дисперсионного анализа (PERMANOVA) с 10⁵ перестановками. Значения представляют псевдо-F отношение со степенями свободы и знаменателем в скобках [F(1, 16)], пермутационный уровень значимости (P), компонент дисперсии (VC), попарную t-статистику (t) и среднее расстояние между группами (DIS). Корректировка для множественных сравнений выполнялась с использованием процедуры Бенджамини–Хохберга. Значения при Padjust < 0,05 выделены жирным шрифтом.
Одномерный анализ показал, что CO оказывал значимое влияние на все переменные, за исключением pH почвы, тогда как БМК проявлял эффекты в отношении влажности, TN, TSN, AK и площади листьев (Таблица S1). Попарные сравнения на уровне обработок дополнительно показали, что влажность почвы значительно снизилась с 16,61 ± 3,19 до 12,19 ± 1,09% при сравнении CK с CO (P = 0,025, Таблица 3). Однако добавление БМК увеличивало влажность почвы как самостоятельно (19,32 ± 2,66%), так и в комбинации с компостом (т.е. обработка BMCO, 14,71 ± 0,59%) по сравнению с обработками CK и CO соответственно. БМК и CO также увеличивали EC почвы на 6 и 26% соответственно. Не было обнаружено заметных изменений pH почвы, несмотря на то, что сами материалы компоста (pH = 9,22) и БМК (pH = 6,98) имели значения pH существенно выше и ниже, чем у почвы (pH = 8,17, Таблица 1). Как и следовало ожидать, все показатели питательных веществ значительно увеличились в почве при добавлении компоста. Дополнительное увеличение некоторых показателей питательных веществ наблюдалось при комбинировании компоста с БМК (т.е. обработка BMCO). В частности, БМК взаимодействовал с компостом синергетически, дополнительно увеличивая TC, TN, OC, NO⁻₃ и AK, тогда как БМК сам по себе вызывал лишь небольшие изменения по сравнению с обработкой CK. БМК также вызывал значительное увеличение TSN как самостоятельно, так и аддитивно с компостом. Напротив, БМК снижал содержание AP в почве, и этот эффект был ещё более выраженным в обработке BMCO.
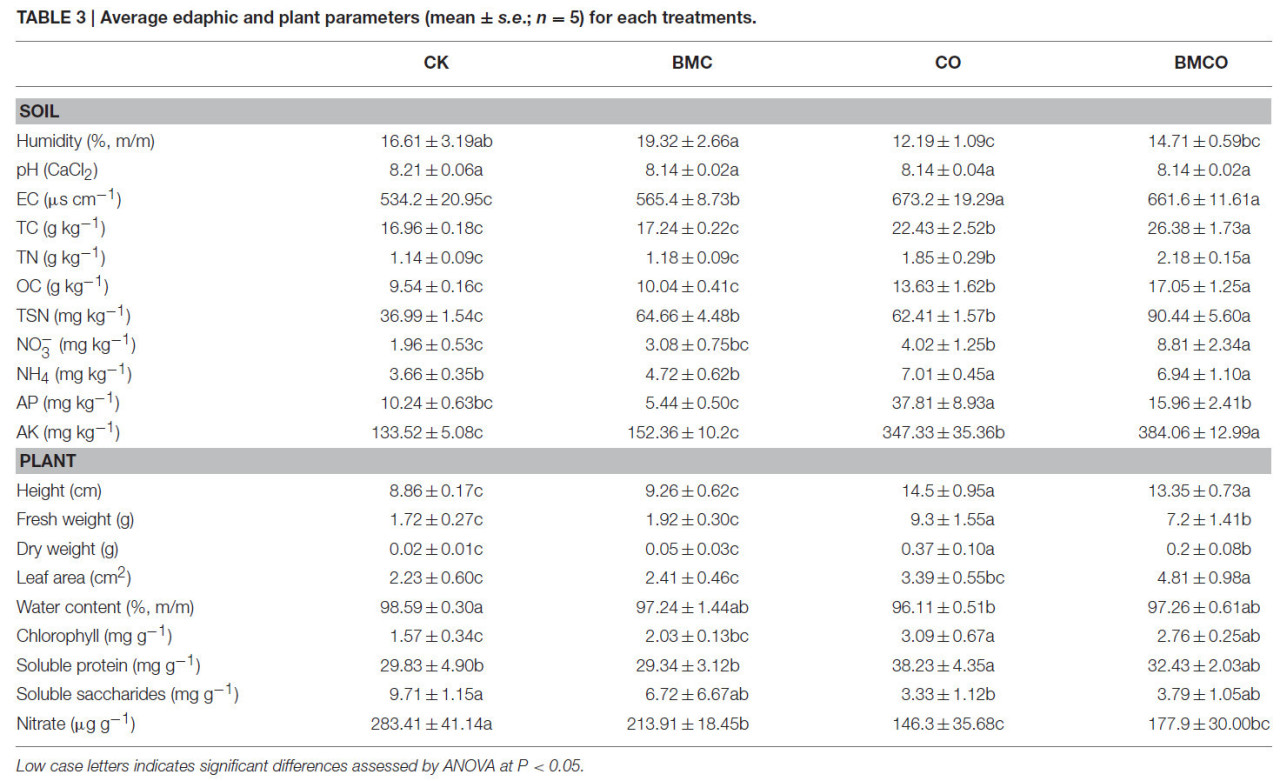
Таблица 3. Средние почвенные и растительные параметры (среднее ± стандартная ошибка; n = 5) для каждой обработки.
Строчные буквы указывают на значимые различия, оценённые с помощью ANOVA при P < 0,05.
Что касается растительных параметров, обработки CO и BMCO приводили к существенному увеличению урожайности (по высоте и массе) пекинской капусты по сравнению с обработками CK и БМК. Однако, что удивительно, комбинация БМК и компоста (обработка BMCO) приводила к дальнейшему увеличению площади листьев, достигая значений до 4,81 ± 0,98 см², что не достигалось при добавлении только БМК или только компоста (Таблица 3). Добавление только компоста вызывало снижение содержания растворимых сахаров и нитратов, но увеличение растворимого белка и хлорофилла по сравнению с контролем. Добавление БМК в почву самостоятельно или в почву, обработанную компостом, оказывало незначительное влияние на содержание хлорофилла, растворимых сахаров и белка. Однако БМК заметно снижал содержание нитратов в пекинской капусте при использовании самостоятельно.
Влияние компоста и биоугле-минерального комплекса на бактериальное разнообразие
Учитывая приведённые выше результаты, добавление компоста и БМК (по отдельности или в комбинации) оказывало явные аддитивные и синергетические эффекты на содержание питательных веществ в почве. Мы выдвинули гипотезу, что это может быть связано с изменениями метаболических процессов, происходящих в почве, и поэтому исследовали состав её бактериального сообщества. Всего было получено 1 174 380 высококачественных последовательностей гена 16S рРНК из 32 образцов, охватывающих три повторности образцов 1-го дня и пять повторностей образцов 40-го дня почвы для каждой из четырёх обработок. После субдискретизации и кластеризации было получено 10 509 операционных таксономических единиц (OTU) при 97% идентичности (примерно соответствует видам), при этом количество OTU варьировало от 519 до 1940 на образец. Полнота покрытия Гуда для наблюдаемых OTU составила 89,2 ± 5,5% при глубине секвенирования 5793 прочтений, что указывает на хорошую репрезентативность выборки сообщества.
Добавление компоста (обработки CO и BMCO) значительно снижало выравненность бактериального сообщества в образцах 1-го дня (P < 0,001, Таблица S2). БМК вызывал небольшое снижение бактериального богатства в начале эксперимента, тогда как CO не влиял на микробное богатство (P = 0,604, Таблица S2). Интересно, что бактериальное богатство и выравненность всех обработок достигли одинакового уровня после 40 дней выращивания, при этом богатство почти удвоилось по сравнению с началом вегетационного опыта (P < 0,001).
Состав бактериальных сообществ в значительной степени определялся факторами CT и CO (Рисунок 1C). Первая ось PCoA разделяла образцы по двум временным точкам (т.е. День 1 и 40), тогда как влияние компоста проявлялось вдоль второй оси PCoA. Повторные образцы в день 1 оказались более разрозненными, чем в день 40, что подтверждалось анализом PERMDISP, показавшим, что среднее расстояние до центроида обработок для образцов 1-го дня было на 27,6% больше, чем для образцов 40-го дня (P < 0,001, Таблица S3). Кроме того, дисперсия, связанная с CO, была незначительной (P = 0,962, Таблица S3), что позволяет предположить, что различия в бактериальном β-разнообразии между группами с компостом (CO и BMCO) и без него (CK и БМК) были обусловлены главным образом несходством между группами, а не дисперсией внутри групп.
Основной тест PERMANOVA показал значительный эффект на β-разнообразие, связанный со взаимодействием факторов CO и CT (VC = 19,77, P < 0,001), и меньший эффект, связанный со взаимодействием факторов БМК и CT (VC = 7,74, P = 0,012). Попарное сравнение в пределах уровней CT показало, что CO сохранял значимый эффект с течением времени (Таблица 4). Это подтверждает представление о том, что компост привносил в почву специфические бактериальные сообщества в начале вегетационного опыта и значительно трансформировал исходные бактериальные сообщества почвы в течение периода выращивания. Эффект БМК был относительно небольшим по сравнению с CO (Таблица 4). Тем не менее, наблюдалось явное различие между обработками CO и BMCO в день 40 (Рисунок 1C), что указывает на дополнительное изменение бактериальных сообществ поверх изменений, вызванных компостом.
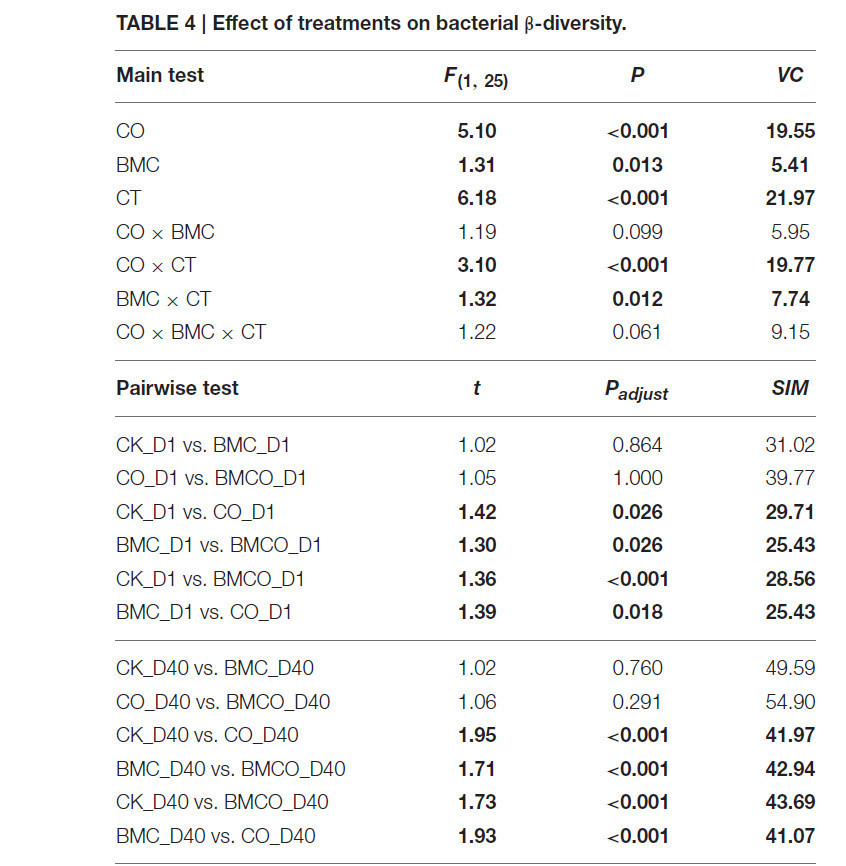
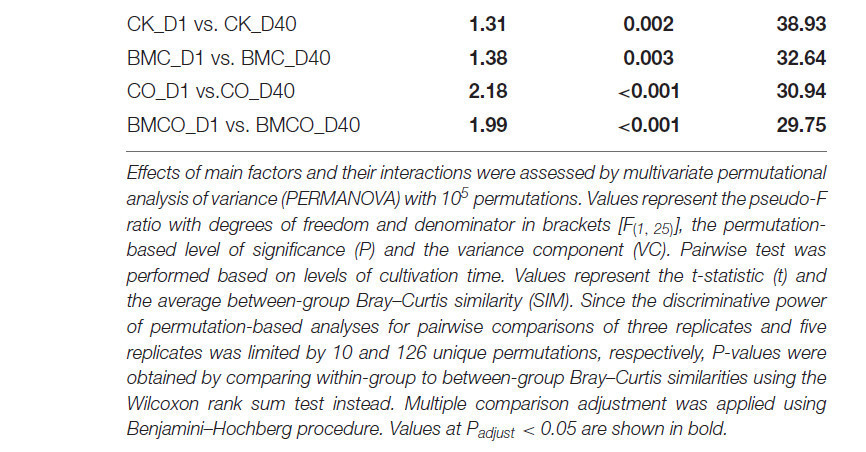
Таблица 4. Влияние обработок на бактериальное β-разнообразие.
Эффекты основных факторов и их взаимодействий оценивались с помощью многомерного пермутационного дисперсионного анализа (PERMANOVA) с 10⁵ перестановками. Значения представляют псевдо-F отношение со степенями свободы и знаменателем в скобках [F(1, 25)], пермутационный уровень значимости (P) и компонент дисперсии (VC). Попарный тест выполнялся на основе уровней времени культивирования. Значения представляют t-статистику (t) и среднее сходство Брея–Кёртиса между группами (SIM). Поскольку различительная способность пермутационных анализов для попарных сравнений трёх повторностей и пяти повторностей была ограничена 10 и 126 уникальными перестановками соответственно, P-значения были получены путём сравнения внутригрупповых и межгрупповых сходств Брея–Кёртиса с использованием рангового критерия Уилкоксона. Корректировка для множественных сравнений выполнялась с использованием процедуры Бенджамини–Хохберга. Значения при Padjust < 0,05 выделены жирным шрифтом.
Связи между почвенными переменными и бактериальными сообществами
Как почвенные параметры, так и бактериальные сообщества были значительно подвержены влиянию внесения компоста и БМК через 40 дней (см. разделы «Влияние компоста и биоугле-минерального комплекса на почвенные и растительные переменные» и «Влияние компоста и биоугле-минерального комплекса на бактериальное разнообразие»), и поэтому мы исследовали корреляции между почвенными переменными и микробным β-разнообразием с использованием метода дистанционной линейной регрессии. Маргинальные тесты показали, что все переменные (кроме pH) имели значимые корреляции с бактериальными сообществами (Таблица 5). Кроме того, поскольку многие почвенные переменные могут объяснять одну и ту же часть вариации в бактериальном β-разнообразии, были выполнены последовательные тесты для проверки остаточной вариации с учётом других переменных (Mcardle and Anderson, 2001). Это показало, что все переменные вместе объясняли 70,3% дисперсии в бактериальном β-разнообразии. EC вносил наибольший вклад, объясняя 22,3% вариации бактериального β-разнообразия (P < 0,05), влажность и отношение C:N объясняли дополнительные 5,4% (P = 0,02) и 5,2% (P = 0,05) соответственно. Эти переменные составили решение BEST (т.е. наилучшую подогнанную модель). Анализ dbRDA проиллюстрировал адекватность модели, состоящей из этих трёх переменных, для прогнозирования закономерностей бактериального β-разнообразия (Рисунок S1). Различия в бактериальном β-разнообразии, связанные с CO, разделялись по EC и C:N, тогда как различия, связанные с БМК, разделялись по влажности почвы.
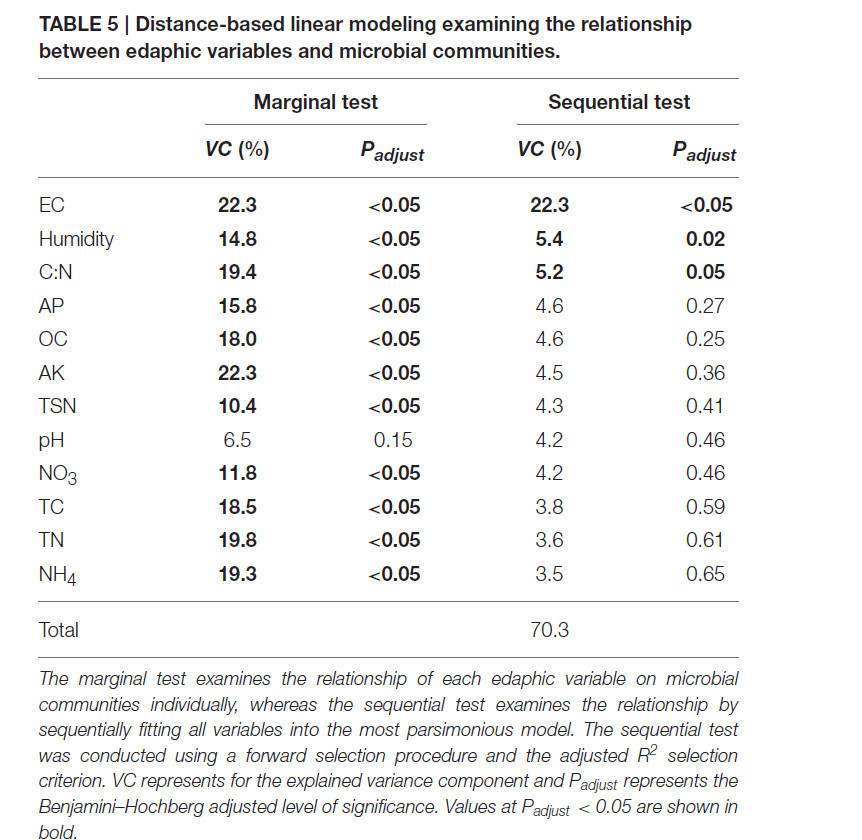
Таблица 5. Дистанционное линейное моделирование для изучения взаимосвязи между почвенными переменными и микробными сообществами.
Маргинальный тест проверяет взаимосвязь каждой почвенной переменной с микробными сообществами по отдельности, тогда как последовательный тест проверяет взаимосвязь путём последовательного включения всех переменных в наиболее экономную модель. Последовательный тест проводился с использованием процедуры прямого отбора и критерия выбора скорректированного R². VC представляет объяснённый компонент дисперсии, а Padjust представляет скорректированный по Бенджамини–Хохбергу уровень значимости. Значения при Padjust < 0,05 выделены жирным шрифтом.
Таксономический состав и таксоны, ассоциированные с обработками
Учитывая чёткое различие бактериальных сообществ между обработками и их корреляцию с почвенными параметрами, мы далее исследовали, какие конкретные микробные таксоны присутствовали и были активны в каждой обработке. Proteobacteria, Actinobacteria и Bacteroidetes были тремя наиболее доминирующими филумами (Рисунок 2A), на которые приходилось 73,9% бактериального разнообразия, наблюдаемого во всех образцах. Филумы Actinobacteria, Bacteroidetes и Firmicutes были более обильны в день 1 в обработках с компостом (CO и BMCO) по сравнению с обработками без компоста (CK и БМК). Через 40 дней филумный состав стал более сходным между обработками (Рисунок 2A), что согласуется с измерениями α-разнообразия (см. Таблицу S2). Более значительные изменения состава за 40-дневный вегетационный период были обнаружены в обработке CO по сравнению с CK на уровне филумов, при этом обработка CK демонстрировала 92,4% сходства, а обработка CO — только 76,9% между началом и концом вегетационного опыта. Это позволяет предположить, что обработки с компостом характеризовались относительно высокой оборачиваемостью бактериальных сообществ по сравнению с необработанной почвой.
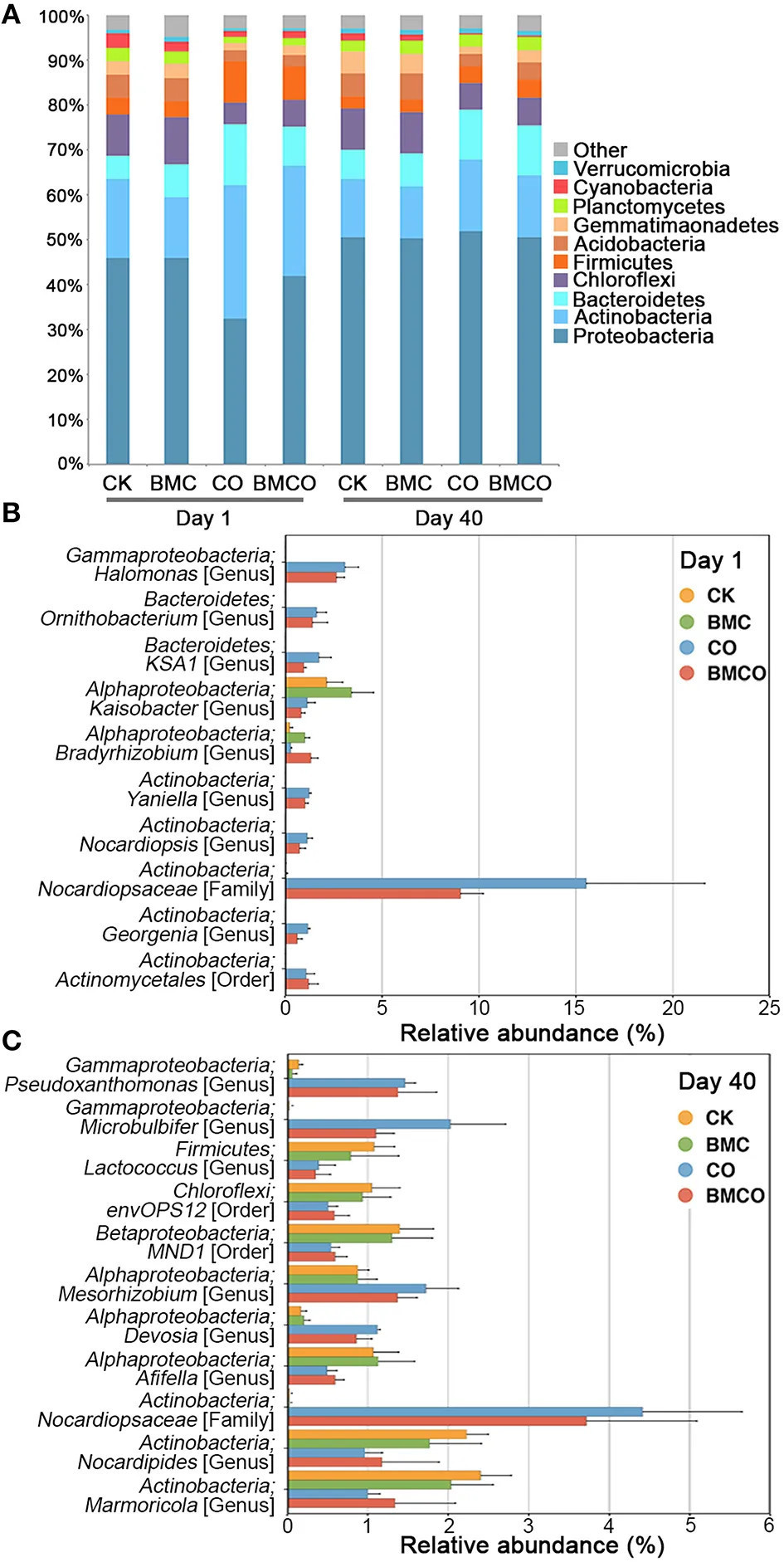
Рисунок 2. Состав бактериальных сообществ. (A) Распределение 10 наиболее обильных филумов по обработкам в день 1 и день 40. (B) Средняя относительная численность 10 наиболее обильных OTU, значительно различавшихся при сравнении обработок с компостом и без него (CK и БМК против CO и BMCO) в день 1, n = 5. (C) Средняя относительная численность 10 наиболее обильных OTU, значительно различавшихся при сравнении обработок с компостом и без него (CK и БМК против CO и BMCO) в день 40, n = 5. P < 0,05 с корректировкой Бенджамини–Хохберга для множественных сравнений.
Далее мы проанализировали профили сообществ на основе OTU для дней 1 и 40 отдельно, чтобы дополнительно определить различия между обработками. В начале вегетационного опыта внесение компоста привнесло в почву 59 новых OTU, которые главным образом относились к филумам Actinobacteria (49,2%), Proteobacteria (22,8%), Bacteroidetes (16,5%) и Firmicutes (9,5%). Это внесение новых бактерий было дополнительно проиллюстрировано при рассмотрении только 10 наиболее обильных OTU (см. Рисунок 2B). Например, OTU, отнесённая к семейству Nocardiopsaceae (филум Actinobacteria), составляла от 9 до 15% бактериальных сообществ в обработках, получавших компост, но практически отсутствовала в почве без добавок. Однако через 40 дней другие OTU вносили наибольший вклад в различия между обработками, включая OTU, принадлежащие к родам Pseudoxanthomonas, Microbulbifer, Mesorhizobium и Devosia (Рисунок 2C). Обработка BMCO показывала значительное увеличение OTU, отнесённых к таксонам Ellin6067, MND1, Ellin6075 и Cytophagaceae, по сравнению с обработкой CO (Рисунок S2).
Индикаторный видовой анализ дополнительно идентифицировал 144 OTU, ассоциированные с обработками, которые составляли 21,2% всех последовательностей. Было обнаружено 7, 10, 18 и 3 OTU, ассоциированных с обработками CK, БМК, CO и BMCO соответственно (Рисунок 3 и Таблица S4 для подробностей). Ни одна OTU не была ассоциирована с комбинациями между обработками CK и CO, что указывает на различную биотическую среду этих двух обработок. Кроме того, многие OTU были привнесены в почву через компост, но истощались (или даже исчезали) со временем, в то время как несколько автохтонных OTU стимулировались в обработках CO и BMCO (Рисунок 3).

Рисунок 3. Бактериальные операционные таксономические единицы (OTU), идентифицированные индикаторным анализом как индикаторные виды для каждой обработки или комбинации обработок. Тепловая карта в левой части панели показывает относительную численность (>1% хотя бы в одном образце) OTU во всех образцах. OTU кластеризованы с использованием ненаправленной иерархической кластеризации на основе относительной численности. Зелёная часть древа охватывает OTU, привнесённые компостом, тогда как красная часть охватывает автохтонные OTU почвы, стимулируемые компостом. Сила ассоциации представлена в средней части панели, где «сила» для данной OTU определяется как её предпочтение соответствующей обработке или комбинации обработок (−1, избегание участка; +1, преобладание на участке). Буквы в квадратных скобках обозначают OTU, имевшие значимую силу ассоциации с соответствующей обработкой или комбинацией обработок (Padjust < 0,05). Названия OTU отформатированы как: название уровня класса; название самого низкого классифицированного уровня [самый низкий классифицированный уровень]. Полный список индикаторных OTU приведён в Таблице S4.
Обсуждение
В этом исследовании мы стремились определить влияние, которое компост и биоугле-минеральный комплекс (БМК) оказывают на бактериальные сообщества в почве и на выращивание пекинской капусты. В целом, результаты показывают, что почва, растения и бактериальные сообщества были явно подвержены влиянию внесения компоста или биоугле-минерального комплекса как по отдельности, так и в комбинации (Рисунок 1). Компост был основным фактором, определяющим почвенную среду, рост растений и микробиоту. В частности, компост добавлял в почву как высокие уровни питательных веществ, так и специфические микроорганизмы, но относительная численность привнесённых бактерий снижалась после 40 дней выращивания, и вместо них в сообществе доминировали другие почвенные микроорганизмы (Рисунки 2, 3). Было показано, что БМК взаимодействует с компостом синергетически как в отношении некоторых почвенных параметров (т.е. TC, TN, OC, NO⁻₃ и AK), так и свойств растений (т.е. площадь листьев и содержание нитратов, Таблица 3). Мы также идентифицировали ряд сильных таксонов, ассоциированных с обработками, которые могут играть ключевую роль в обеспечении положительных результатов для почвенной и растительной среды.
Влияние компоста на процессы питания в почве
Типичная норма внесения (т.е. 28,5 т га⁻¹; Wong и др., 1999), использованная здесь для компоста, оказывала явное существенное влияние на почвенную среду. Почвенные микроорганизмы являются первыми респондентами и деструкторами почвенных питательных веществ, и наш анализ показал, что добавление компоста приводило к существенным изменениям в структуре и составе бактериального сообщества (Рисунок 1C; Рисунок S1; Таблица S2). Это согласуется с недавними исследованиями, показывающими, что компост влияет на почвенное биоразнообразие (Mäder и др., 2002; Hartmann и др., 2015). Следовательно, компост не только привносил питательные вещества в почвенную среду, но и стимулировал рост и оборачиваемость бактерий в почве. Таким образом, происходила трансформация микробных процессов в почве, что впоследствии могло повлиять на уровни питательных веществ и продуктивность растений.
Мы наблюдали значимые различия в OTU, отнесённых к филумам Chloroflexi, Acidobacteria, Actinobacteria, Bacteroidetes и Proteobacteria, между обработками с компостом и без него (Рисунки 2, 3). Chloroflexi и Acidobacteria, которые, как было ранее показано, являются медленнорастущими бактериями (Davis и др., 2011) и обычно предпочитают олиготрофные условия (Fierer и др., 2012; Hartmann и др., 2015), были сильно ассоциированы с обработками БМК и CK (Таблица S4). OTU, обильные в обработках CO и BMCO в пределах филума Actinobacteria, были отнесены к родам Streptomyces, Phytohabitans, Aeromicrobium, Nocardioides и Pimelobacter, которые известны как деструкторы лигноцеллюлозы и играют важные роли в обороте органического вещества (Abdulla and El-Shatoury, 2007). Кроме того, OTU в пределах семейств Cytophagaceae и Chitinophagaceae (филум Bacteroidetes), которые известны своей способностью разлагать сложные углеводы (McBride и др., 2014; Rosenberg, 2014), были ассоциированы с компостом (Таблица S4). OTU, стимулируемые компостом в классе Gammaproteobacteria, принадлежали к родам Microbulbifer, Lysobacter и Pseudoxanthomonas (Рисунок 3). Microbulbifer spp. выделяют гидролитические ферменты для расщепления целлюлозы, хитина и желатина (González и др., 1997). Lysobacter spp. являются известными агентами биоконтроля, продуцирующими несколько антибактериальных природных продуктов (Xie и др., 2012). Pseudoxanthomonas spp., как было показано, способны разлагать различные органические соединения (Kim и др., 2008).
Среди Alphaproteobacteria в общей сложности 14 OTU, ассоциированных с обработками, для четырёх обработок были классифицированы как относящиеся к порядку Rhizobiales, и они имеют различные предпочтения к определённым обработкам и метаболическим процессам. Из шести OTU, обогащённых в обработках CK и БМК, пять были отнесены к родам Rhodoplanes, Hyphomicrobium и Pedomicrobium, которые известны как хемоорганотрофы. Представители этих родов обычно используют простые углеродные субстраты и минеральные соли и адаптированы к бедным питательными веществами местообитаниям (Oren and Xu, 2014). Кроме того, Pedomicrobium spp. и Hyphomicrobium spp. известны как простековые бактерии, обладающие селективным преимуществом в олиготрофных условиях (Semenov and Staley, 1992). Напротив, восемь OTU, ассоциированных с обработками CO и BMCO, были отнесены к родам Devosia, Mesorhizobium, Agrobacterium и Shinella. Хемогетеротрофные Devosia spp. обладают генами для азотфиксации и клубенькообразования (Rivas и др., 2002; Bautista и др., 2010), тогда как род Shinella был обнаружен в копиотрофных условиях (Alves и др., 2014), включая корневые клубеньки травянистых бобовых (Lin и др., 2008). Mesorhizobium spp. и Agrobacterium spp. являются хорошо известными диазотрофами (Lippincott и др., 1981) и обладают способностью стимулировать рост растений (Hao и др., 2012).
В совокупности эти данные показывают, что необработанная почва и почва, обработанная небольшими количествами (1,5 т га⁻¹) относительно трудноразлагаемого БМК, поддерживали бактериальные сообщества, имеющие олиготрофный характер. Это указывает на то, что природная почва, использованная для выращивания пекинской капусты, была лимитирована по питательным веществам, что также отражалось в низкой урожайности растений и высоком содержании растворимых сахаров (Roitsch, 1999). Добавление компоста, независимо от присутствия БМК, преодолевает эти ограничения и способствует росту многих бактерий, которые не только адаптированы к более копиотрофным условиям, но и обладают такими характеристиками, как азотфиксация, которые полезны для роста пекинской капусты. Следовательно, увеличение урожайности, наблюдаемое здесь, вероятно, было результатом не только дополнительных питательных веществ, но и стимуляции полезных микробных организмов и процессов.
Кроме того, OTU, отнесённая к Nocardiopsaceae, была наиболее обильным таксоном, привнесённым компостом, и составляла 15,52 ± 6,12 и 9,05 ± 1,17% от общей относительной численности в обработках CO и BMCO в день 1 соответственно. Её численность значительно снизилась в течение 40 дней выращивания (4,1 ± 1,23 и 3,72 ± 1,37% для обработок CO и BMCO соответственно). Репрезентативная последовательность для этой OTU Nocardiopsaceae показала 100% идентичность с последовательностью гена 16S рРНК изолята из кожных поражений крупного рогатого скота (Genbank JQ031766) и 96% идентичность с геном 16S рРНК Nocardiopsis nikkonensis, выделенного из зрелого компоста (Yamamura и др., 2010). Было обнаружено четыре дополнительных OTU, отнесённых к родам Yaniella, Georgenia, Halomonas и Ornithobacterium, привнесённых компостом, но значительно снизившихся к 40-му дню. Репрезентативные последовательности OTU Yaniella, Georgenia и Halomonas оказались наиболее близки к культивируемым штаммам из галофильных сред (Vreeland и др., 1980; Li и др., 2004; Tang и др., 2010). OTU Ornithobacterium была близка к штаммам смертельного патогена для домашней птицы (Hafez, 2002). Это показывает, что компост привносит в почву разнообразные микроорганизмы, включая потенциальных патогенов из животного сырья. Однако эти микроорганизмы, происходящие из компостного сырья (т.е. куриного помёта), снижались в численности, что, вероятно, связано с воздействием различных условий окружающей среды (таких как температура, влажность, солёность или доступность питательных веществ), присутствующих в почве (Sun и др., 2015).
Аддитивные и синергетические эффекты биоугле-минерального комплекса с компостом
Сам по себе БМК был недостаточен для достижения такой же урожайности растений, как обработка CO, но мы обнаружили ряд аддитивных и синергетических эффектов при комбинировании БМК и компоста. Особый интерес представляет то, что обработка BMCO имела повышенные значения TC, TN, OC, TSN, NO⁻₃ и AK, часто превышающие уровень, ожидаемый от простого суммирования отдельных эффектов CO и БМК. Например, уровень NO⁻₃ в обработке BMCO был в два раза выше (8,81 ± 2,34 мг кг⁻¹), чем в обработке CO (4,02 ± 1,25 мг кг⁻¹), и не мог быть объяснён простым учётом эффекта только БМК (3,08 ± 0,75 мг кг⁻¹) и почвенного контроля (1,96 ± 0,53 мг кг⁻¹). Почвенные нитраты являются важным фактором для роста растений и, как было показано, стимулируют рост листьев (Guo и др., 2007). Таким образом, мы предполагаем, что увеличение размера листьев, наблюдаемое в нашем эксперименте с пекинской капустой, было прямым следствием повышенного уровня нитратов, вызванного синергетической активностью между БМК и компостом. Аналогичным образом, мы обнаружили, что чистый прирост TC и TN в обработках CO и BMCO по сравнению с CK был в среднем в три и два раза больше, чем можно было бы объяснить углеродом и азотом, внесёнными с компостом и БМК плюс компост (Таблицы 1, 2). Это увеличение потенциально может быть объяснено обогащением определёнными типами бактерий, такими как разлагающие (например, Streptomyces spp. или Cytophagaceae) или азотфиксирующие (например, Mesorhizobium spp. или Agrobacterium spp.) таксоны (см. выше). Хотя это увеличение в обработке BMCO не дало немедленных преимуществ для роста растений в короткие сроки нашего эксперимента по выращиванию, мы ожидаем, что более длительные испытания, особенно с несколькими вегетационными сезонами, покажут устойчивое увеличение почвенных питательных веществ и, следовательно, вероятно, устойчивую продуктивность.
Одно из объяснений повышенных значений нитратов, наблюдаемых в обработках BMCO, заключается в лежащих в основе микробных процессах. Мы обнаружили, что OTU таксонов Ellin6067, MND1 и Ellin6075 были значительно более обильны в обработках BMCO по сравнению с CO в день 40 (Рисунок S2). Ellin6067 spp. описывается как предполагаемая аммоний-окисляющая бактерия (Xia и др., 2005), а MND1 spp. и Ellin6075 spp. способны к нитрификации (Chuang и др., 2007; Cao и др., 2015). Это означает, что процессы нитрификации были усилены в обработке BMCO. Мы дополнительно исследовали это, суммируя все известные нитрифицирующие бактерии (используя ключевое слово «нитро» в таксономическом названии). В день 1 почва содержала относительную численность 0,78 ± 0,11% для этих суммарных нитрифицирующих бактерий. Через 40 дней БМК оказывал значимое влияние на относительную численность нитрифицирующих бактерий (P = 0,007), при этом обработка BMCO имела наибольшее количество нитрифицирующих бактерий (1,10 ± 0,07%), за ней следовали БМК (1,04 ± 0,04%), CK (0,95 ± 0,09%) и CO (0,79 ± 0,05%). Усиленная нитрифицирующая активность, о чём свидетельствует относительно более высокое количество нитрифицирующих бактерий, приводила бы к более высокому уровню нитратов (Рисунок 3), а это, в свою очередь, могло бы приводить к увеличению размера листьев пекинской капусты. Таким образом, мы предполагаем, что в нашем случае увеличение площади листьев пекинской капусты было косвенным следствием стимуляции нитрификации БМК в почвах, обработанных компостом.
Мы также наблюдали, что содержание доступного фосфата снижалось в обработке BMCO по сравнению с обработкой CO (Таблица 3). Ортофосфат (основная форма доступного фосфата) является вторым по значимости лимитирующим макроэлементом (после азота) для роста растений (Schachtman и др., 1998), и сельскохозяйственные культуры имеют высокую потребность в фосфатных удобрениях при выращивании на слабощелочных почвах (Lucas and Davis, 1961). Поэтому вероятно, что относительно высокий pH компоста и ионы металлов (Ca²⁺, Mg²⁺, Fe³⁺, Al³⁺) на БМК (Таблица 1) создавали условия для иммобилизации ортофосфата, что приводило к низкому содержанию доступного фосфата и, следовательно, ограничивало рост растений (Cao and Harris, 2010). Однако этот эффект может быть лишь краткосрочным, поскольку было обнаружено, что биоуголь из твёрдых пород древесины, имеющий свойства, сходные с использованным здесь биоуглём, за исключением минерального покрытия, способствует росту фосфор-мобилизующих бактерий и последующему росту райграса пастбищного Lolium perenne (Fox и др., 2014). Поскольку наш вегетационный опыт длился всего 40 дней, такие положительные эффекты микробной фосфатмобилизации могли не успеть проявиться в достаточной степени.
Заключение
Это исследование предоставляет доказательства того, что БМК может действовать синергетически с компостом, опосредуя микробные процессы, которые приводят к изменениям в почвенных циклах питательных веществ, что, в свою очередь, может влиять на агрономические свойства. Основываясь на наших наблюдениях, мы предлагаем модель, в которой большое количество питательных веществ, обеспечиваемое компостом, ускоряет почвенные процессы, такие как оборачиваемость питательных веществ и бактериальных сообществ. Затем эти процессы могут контролироваться (или направляться) добавлением БМК, сложные химические свойства и поверхностные структуры которого, как известно, влияют на физико-химические свойства почвенной среды (например, влажность, Рисунок S1), а также на микробный метаболизм и разнообразие (Joseph и др., 2013). Таким образом, разработка БМК в будущем для соответствия конкретным органическим удобрениям и почвенным параметрам могла бы обеспечить микробно-опосредованные превращения питательных веществ с улучшенной сельскохозяйственной продуктивностью. Комбинации БМК и компоста, которые обеспечивают сниженные уровни микробно-опосредованных выбросов N₂O и метана, также потенциально могут быть разработаны, аналогично тому, что недавно наблюдалось для добавления обычного биоугля в сельскохозяйственные почвы (Harter и др., 2014; Ho и др., 2015).
Дополнительные материалы
Дополнительные материалы к этой статье можно найти онлайн по адресу: http://journal.frontiersin.org/article/10.3389/fmicb.2016.00372
Ссылки
1. Abdulla H. M., El-Shatoury S. A. (2007). Actinomycetes in rice straw decomposition. Waste Manage. 27, 850–853. doi: 10.1016/j.wasman.2006.06.006. CrossRef. Google Scholar.
2. Alves L. M. C., De Souza J. A. M., De Mello Varani A., De Macedo Lemos E. G. (2014). The family Rhizobiaceae, in The Prokaryotes, eds Rosenberg E., DeLong E. F., Lory S., Stackebrandt E., Thompson F. (Berlin; Heidelberg: Springer), 419–437. doi: 10.1007/978-3-642-30197-1_297. CrossRef. Google Scholar.
3. Anderson M. J., Gorley R. N., Clarke K. R. (2008). PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods. Plymouth, UK: PRIMER-E Ltd. Google Scholar.
4. Bautista V. V., Monsalud R. G., Yokota A. (2010). Devosia yakushimensis sp. nov., isolated from root nodules of pueraria lobata (willd.) ohwi. Int. J. Syst. Evol. Microbiol. 60, 627–632. doi: 10.1099/ijs.0.011254-0. CrossRef. Google Scholar.
5. Berry P., Sylvester-Bradley R., Philipps L., Hatch D., Cuttle S., Rayns F., et al. (2002). Is the productivity of organic farms restricted by the supply of available nitrogen? Soil Use Manage. 18, 248–255. doi: 10.1079/SUM2002129. CrossRef. Google Scholar.
6. Brown J. R. (1998). Recommended Chemical Soil Test Procedures for the North Central Region. Missouri Agricultural Experiment Station, University of Missouri--Columbia. Google Scholar.
7. Cao L. T. T., Kodera H., Abe K., Imachi H., Aoi Y., Kindaichi T., et al. (2015). Biological oxidation of Mn (II) coupled with nitrification for removal and recovery of minor metals by downflow hanging sponge reactor. Water Res. 68, 545–553. doi: 10.1016/j.watres.2014.10.002. CrossRef. Google Scholar.
8. Cao X., Harris W. (2010). Properties of dairy-manure-derived biochar pertinent to its potential use in remediation. Bioresour. Technol. 101, 5222–5228. doi: 10.1016/j.biortech.2010.02.052. CrossRef. Google Scholar.
9. Chan K., Van Zwieten L., Meszaros I., Downie A., Joseph S. (2008). Agronomic values of greenwaste biochar as a soil amendment. Soil Res. 45, 629–634. doi: 10.1071/SR07109. CrossRef. Google Scholar.
10. Chia C. H., Munroe P., Joseph S., Lin Y. (2010). Microscopic characterisation of synthetic Terra Preta. Soil Res. 48, 593–605. doi: 10.1071/SR10012. CrossRef. Google Scholar.
11. Chia C. H., Singh B. P., Joseph S., Graber E. R., Munroe P. (2014). Characterization of an enriched biochar. J. Anal. Appl. Pyrolysis 108, 26–34. doi: 10.1016/j.jaap.2014.05.021. CrossRef. Google Scholar.
12. Chia C., Munroe P., Joseph S. D., Lin Y., Lehmann J., Muller D., et al. (2012). Analytical electron microscopy of black carbon and microaggregated mineral matter in Amazonian dark Earth. J. Microsc. (Oxford) 245, 129–139. doi: 10.1111/j.1365-2818.2011.03553.x. CrossRef. Google Scholar.
13. Chuang H.-P., Imachi H., Tandukar M., Kawakami S., Harada H., Ohashi A. (2007). Microbial community that catalyzes partial nitrification at low oxygen atmosphere as revealed by 16S rRNA and amoA genes. J. Biosci. Bioeng. 104, 525–528. doi: 10.1263/jbb.104.525. CrossRef. Google Scholar.
14. Clarke K. R., Warwick R. M. (2001). Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 2nd Edn. Plymouth: PRIMER-E Ltd., 172. Google Scholar.
15. Corwin D., Lesch S. (2005). Apparent soil electrical conductivity measurements in agriculture. Comput. Electron. Agric. 46, 11–43. doi: 10.1016/j.compag.2004.10.005. CrossRef. Google Scholar.
16. Davis K. E., Sangwan P., Janssen P. H. (2011). Acidobacteria, Rubrobacteridae and Chloroflexi are abundant among very slow-growing and mini-colony-forming soil bacteria. Environ. Microbiol. 13, 798–805. doi: 10.1111/j.1462-2920.2010.02384.x. CrossRef. Google Scholar.
17. De Cáceres M., Legendre P. (2009). Associations between species and groups of sites: indices and statistical inference. Ecology 90, 3566–3574. doi: 10.1890/08-1823.1. CrossRef. Google Scholar.
18. De Cáceres M., Legendre P., Moretti M. (2010). Improving indicator species analysis by combining groups of sites. Oikos 119, 1674–1684. doi: 10.1111/j.1600-0706.2010.18334.x. CrossRef. Google Scholar.
19. Edgar R. C., Haas B. J., Clemente J. C., Quince C., Knight R. (2011). UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 27, 2194–2200. doi: 10.1093/bioinformatics/btr381. CrossRef. Google Scholar.
20. Fierer N., Lauber C. L., Ramirez K. S., Zaneveld J., Bradford M. A., Knight R. (2012). Comparative metagenomic, phylogenetic and physiological analyses of soil microbial communities across nitrogen gradients. ISME J. 6, 1007–1017. doi: 10.1038/ismej.2011.159. CrossRef. Google Scholar.
21. Fox A., Kwapinski W., Griffiths B. S., Schmalenberger A. (2014). The role of sulfur-and phosphorus-mobilizing bacteria in biochar-induced growth promotion of Lolium perenne. FEMS Microbiol. Ecol. 90, 78–91. doi: 10.1111/1574-6941.12374. CrossRef. Google Scholar.
22. Glaser B., Balashov E., Haumaier L., Guggenberger G., Zech W. (2000). Black carbon in density fractions of anthropogenic soils of the Brazilian Amazon region. Org. Geochem. 31, 669–678. doi: 10.1016/S0146-6380(00)00044-9. CrossRef. Google Scholar.
23. González J., Mayer F., Moran M. A., Hodson R. E., Whitman W. (1997). Microbulbifer hydrolyticus gen. nov., sp. nov., and Marinobacterium georgiense gen. nov., sp. nov., two marine bacteria from a lignin-rich pulp mill waste enrichment community. Int. J. Syst. Bacteriol. 47, 369–376. doi: 10.1099/00207713-47-2-369. CrossRef. Google Scholar.
24. Grossman J. M., O'neill B. E., Tsai S. M., Liang B., Neves E., Lehmann J., et al. (2010). Amazonian anthrosols support similar microbial communities that differ distinctly from those extant in adjacent, unmodified soils of the same mineralogy. Microb. Ecol. 60, 192–205. doi: 10.1007/s00248-010-9689-3. CrossRef. Google Scholar.
25. Guo S., Zhou Y., Shen Q., Zhang F. (2007). Effect of ammonium and nitrate nutrition on some physiological processes in higher plants-growth, photosynthesis, photorespiration, and water relations. Plant Biol. 9, 21–29. doi: 10.1055/s-2006-924541. CrossRef. Google Scholar.
26. Hafez H. M. (2002). Diagnosis of Ornithobacterium rhinotracheale. Int. J. Poult. Sci. 1, 114–118. doi: 10.3923/ijps.2002.114.118. CrossRef. Google Scholar.
27. Hao X., Lin Y., Johnstone L., Baltrus D. A., Miller S. J., Wei G., et al. (2012). Draft genome sequence of plant growth-promoting rhizobium Mesorhizobium amorphae, isolated from zinc-lead mine tailings. J. Bacteriol. 194, 736–737. doi: 10.1128/JB.06475-11. CrossRef. Google Scholar.
28. Harter J., Krause H.-M., Schuettler S., Ruser R., Fromme M., Scholten T., et al. (2014). Linking N2O emissions from biochar-amended soil to the structure and function of the N-cycling microbial community. ISME J. 8, 660–674. doi: 10.1038/ismej.2013.160. CrossRef. Google Scholar.
29. Hartmann M., Frey B., Mayer J., Mäder P., Widmer F. (2015). Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 9, 1177–1194. doi: 10.1038/ismej.2014.210. CrossRef. Google Scholar.
30. Ho A., Reim A., Kim S. Y., Meima-Franke M., Termorshuizen A., Boer W., et al. (2015). Unexpected stimulation of soil methane uptake as emergent property of agricultural soils following bio-based residue application. Glob. Change Biol. 21, 3864–3879. doi: 10.1111/gcb.12974. CrossRef. Google Scholar.
31. Jones J. B., Jr (2001). Laboratory Guide for Conducting Soil Tests and Plant Analysis. Boca Raton, FL: CRC Press. Google Scholar.
32. Joseph S., Graber E., Chia C., Munroe P., Donne S., Thomas T., et al. (2013). Shifting paradigms: development of high-efficiency biochar fertilizers based on nano-structures and soluble components. Carbon Manage. 4, 323–343. doi: 10.4155/cmt.13.23. CrossRef. Google Scholar.
33. Kim J. M., Le N. T., Chung B. S., Park J. H., Bae J.-W., Madsen E. L., et al. (2008). Influence of soil components on the biodegradation of benzene, toluene, ethylbenzene, and o-, m-, and p-xylenes by the newly isolated bacterium Pseudoxanthomonas spadix BD-a59. Appl. Environ. Microbiol. 74, 7313–7320. doi: 10.1128/AEM.01695-08. CrossRef. Google Scholar.
34. Kozich J. J., Westcott S. L., Baxter N. T., Highlander S. K., Schloss P. D. (2013). Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 79, 5112–5120. doi: 10.1128/AEM.01043-13. CrossRef. Google Scholar.
35. Li F., Cao X., Zhao L., Wang J., Ding Z. (2014). Effects of mineral additives on biochar formation: carbon retention, stability, and properties. Environ. Sci. Technol. 48, 11211–11217. doi: 10.1021/es501885n. CrossRef. Google Scholar.
36. Li H., Sun Q., Zhao S., Zhang W. (2000). Principles and Techniques of Plant Physiological Biochemical Experiment. Beijing: Higher Education, 130–194. Google Scholar.
37. Li W.-J., Chen H.-H., Xu P., Zhang Y.-Q., Schumann P., Tang S.-K., et al. (2004). Yania halotolerans gen. nov., sp. nov., a novel member of the suborder micrococcineae from saline soil in china. Int. J. Syst. Evol. Microbiol. 54, 525–531. doi: 10.1099/ijs.0.02875-0. CrossRef. Google Scholar.
38. Lin D. X., Wang E. T., Tang H., Han T. X., He Y. R., Guan S. H., et al. (2008). Shinella kummerowiae sp. nov., a symbiotic bacterium isolated from root nodules of the herbal legume Kummerowia stipulacea. Int. J. Syst. Evol. Microbiol. 58, 1409–1413. doi: 10.1099/ijs.0.65723-0. CrossRef. Google Scholar.
39. Lin Y., Munroe P., Joseph S., Ziolkowski A., Van Zwieten L., Kimber S., et al. (2013). Chemical and structural analysis of enhanced biochars: thermally treated mixtures of biochar, chicken litter, clay and minerals. Chemosphere 91, 35–40. doi: 10.1016/j.chemosphere.2012.11.063. CrossRef. Google Scholar.
40. Lippincott J. A., Lippincott B. B., Starr M. P. (1981). The genus Agrobacterium, in The Prokaryotes, eds Starr M. P., Heinz Stolp, Trüper H. G., Balows A., Schlegel H. G. (Berlin; Heidelberg: Springer), 842–855. doi: 10.1007/978-3-662-13187-9_68. CrossRef. Google Scholar.
41. Lucas R., Davis J. F. (1961). Relationships between pH values of organic soils and availabilities of 12 plant nutrients. Soil Sci. 92, 177–182. doi: 10.1097/00010694-196109000-00005. CrossRef. Google Scholar.
42. Mäder P., Fliessbach A., Dubois D., Gunst L., Fried P., Niggli U. (2002). Soil fertility and biodiversity in organic farming. Science 296, 1694–1697. doi: 10.1126/science.1071148. CrossRef. Google Scholar.
43. McArdle B. H., Anderson M. J. (2001). Fitting multivariate models to community data: a comment on distance-based redundancy analysis. Ecology 82, 290–297. doi: 10.1890/0012-9658(2001)082[0290:FMMTCD]2.0.CO;2. CrossRef. Google Scholar.
44. McBride M. J., Liu W., Lu X., Zhu Y., Zhang W. (2014). The family Cytophagaceae, in The Prokaryotes, eds Rosenberg E., DeLong E. F., Lory S., Stackebrandt E., Thompson F. (Berlin; Heidelberg: Springer), 577–593. doi: 10.1007/978-3-642-38954-2_382. CrossRef. Google Scholar.
45. Nielsen S., Minchin T., Kimber S., Van Zwieten L., Gilbert J., Munroe P., et al. (2014). Comparative analysis of the microbial communities in agricultural soil amended with enhanced biochars or traditional fertilisers. Agric. Ecosyst. Environ. 191, 73–82. doi: 10.1016/j.agee.2014.04.006. CrossRef. Google Scholar.
46. O'Neill B., Grossman J., Tsai M. T., Gomes J., Lehmann J., Peterson J., et al. (2009). Bacterial community composition in Brazilian anthrosols and adjacent soils characterized using culturing and molecular identification. Microb. Ecol. 58, 23–35. doi: 10.1007/s00248-009-9515-y. CrossRef. Google Scholar.
47. Olsen S. R. (1954). Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate. Washington, DC: U.S. Dept. of Agriculture. Google Scholar.
48. Oren A., Xu X.-W. (2014). The family Hyphomicrobiaceae, in The Prokaryotes, eds Rosenberg E., DeLong E. F., Lory S., Stackebrandt E., Thompson F. (Berlin; Heidelberg: Springer), 247–281. doi: 10.1007/978-3-642-30197-1_257. CrossRef. Google Scholar.
49. Parks D. H., Tyson G. W., Hugenholtz P., Beiko R. G. (2014). STAMP: statistical analysis of taxonomic and functional profiles. Bioinformatics 30, 3123–3124. doi: 10.1093/bioinformatics/btu494. CrossRef. Google Scholar.
50. Pruesse E., Quast C., Knittel K., Fuchs B. M., Ludwig W., Peplies J., et al. (2007). SILVA: a comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 35, 7188–7196. doi: 10.1093/nar/gkm864. CrossRef. Google Scholar.
51. Ramankutty N., Rhemtulla J. (2012). Can intensive farming save nature? Front. Ecol. Environ. 10, 455. doi: 10.1890/1540-9295-10.9.455. CrossRef. Google Scholar.
52. Reeder J., Knight R. (2009). The 'rare biosphere': a reality check. Nat. Methods 6, 636–637. doi: 10.1038/nmeth0909-636. CrossRef. Google Scholar.
53. Rigby D., Cáceres D. (2001). Organic farming and the sustainability of agricultural systems. Agric. Syst. 68, 21–40. doi: 10.1016/S0308-521X(00)00060-3. CrossRef. Google Scholar.
54. Rivas R., Velázquez E., Willems A., Vizcaíno N., Subba-Rao N. S., Mateos P. F., et al. (2002). A new species of Devosia that forms a unique nitrogen-fixing root-nodule symbiosis with the aquatic legume Neptunia natans (Lf) Druce. Appl. Environ. Microbiol. 68, 5217–5222. doi: 10.1128/AEM.68.11.5217-5222.2002. CrossRef. Google Scholar.
55. Roitsch T. (1999). Source-sink regulation by sugar and stress. Curr. Opin. Plant Biol. 2, 198–206. doi: 10.1016/S1369-5266(99)80036-3. CrossRef. Google Scholar.
56. Rosenberg E. (2014). The family Chitinophagaceae, in The Prokaryotes, eds Rosenberg E., DeLong E. F., Lory S., Stackebrandt E., Thompson F. (Berlin; Heidelberg: Springer), 493–495. doi: 10.1007/978-3-642-38954-2_137. CrossRef. Google Scholar.
57. Schachtman D. P., Reid R. J., Ayling S. M. (1998). Phosphorus uptake by plants: from soil to cell. Plant Physiol. 116, 447–453. doi: 10.1104/pp.116.2.447. CrossRef. Google Scholar.
58. Semenov A., Staley J. T. (1992). Ecology of polyprosthecate bacteria, in Advances in Microbial Ecology, ed Marshall K. C. (Boston, MA: Springer), 339–382. doi: 10.1007/978-1-4684-7609-5_7. CrossRef. Google Scholar.
59. Seufert V., Ramankutty N., Foley J. A. (2012). Comparing the yields of organic and conventional agriculture. Nature 485, 229–232. doi: 10.1038/nature11069. CrossRef. Google Scholar.
60. Sun R., Zhang X.-X., Guo X., Wang D., Chu H. (2015). Bacterial diversity in soils subjected to long-term chemical fertilization can be more stably maintained with the addition of livestock manure than wheat straw. Soil Biol. Biochem. 88, 9–18. doi: 10.1016/j.soilbio.2015.05.007. CrossRef. Google Scholar.
61. Tang S.-K., Wang Y., Lee J.-C., Lou K., Park D.-J., Kim C.-J., et al. (2010). Georgenia halophila sp. nov., a halophilic actinobacterium isolated from a salt lake. Int. J. Syst. Evol. Microbiol. 60, 1317–1421. doi: 10.1099/ijs.0.014993-0. CrossRef. Google Scholar.
62. Unc A., Goss M. J. (2004). Transport of bacteria from manure and protection of water resources. Appl. Soil Ecol. 25, 1–18. doi: 10.1016/j.apsoil.2003.08.007. CrossRef. Google Scholar.
63. Vreeland R., Litchfield C., Martin E., Elliot E. (1980). Halomonas elongata, a new genus and species of extremely salt-tolerant bacteria. Int. J. Syst. Bacteriol. 30, 485–495. doi: 10.1099/00207713-30-2-485. CrossRef. Google Scholar.
64. Walkley A., Black I. A. (1934). An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 37, 29–38. doi: 10.1097/00010694-193401000-00003. CrossRef. Google Scholar.
65. Wickham H. (2009). ggplot2: Elegant Graphics for Data Analysis. New York, NY: Springer Science & Business Media. Google Scholar.
66. Wilcoxon F. (1945). Individual comparisons by ranking methods. Biometrics Bull. 1, 80–83. doi: 10.2307/3001968. CrossRef. Google Scholar.
67. Wong J., Ma K., Fang K., Cheung C. (1999). Utilization of a manure compost for organic farming in Hong Kong. Bioresour. Technol. 67, 43–46. doi: 10.1016/S0960-8524(99)00066-8. CrossRef. Google Scholar.
68. Xia S., Shi Y., Fu Y., Ma X. (2005). DGGE analysis of 16S rDNA of ammonia-oxidizing bacteria in chemical--biological flocculation and chemical coagulation systems. Appl. Microbiol. Biotechnol. 69, 99–105. doi: 10.1007/s00253-005-0035-5. CrossRef. Google Scholar.
69. Xie Y., Wright S., Shen Y., Du L. (2012). Bioactive natural products from Lysobacter. Nat. Prod. Rep. 29, 1277–1287. doi: 10.1039/c2np20064c. CrossRef. Google Scholar.
70. Yamamura H., Ohkubo S.-Y., Ishida Y., Otoguro M., Tamura T., Hayakawa M. (2010). Nocardiopsis nikkonensis sp. nov., isolated from a compost sample. Int. J. Syst. Evol. Microbiol. 60, 2967–2971. doi: 10.1099/ijs.0.016956-0. CrossRef. Google Scholar.

Ye J, Zhang R, Nielsen S, Joseph SD, Huang D and Thomas T (2016) A Combination of Biochar–Mineral Complexes and Compost Improves Soil Bacterial Processes, Soil Quality, and Plant Properties. Front. Microbiol. 7:372. doi: 10.3389/fmicb.2016.00372
Перевод статьи «A Combination of Biochar–Mineral Complexes and Compost Improves Soil Bacterial Processes, Soil Quality, and Plant Properties» авторов Ye J, Zhang R, Nielsen S, Joseph SD, Huang D and Thomas T., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык
Фото: Designed by Freepik























































































































Комментарии (0)