Недорогая и эффективная система фермерского разведения Spongia anclotea
Губки в настоящее время востребованы из-за их коммерческого, промышленного и фармацевтического значения. Это привело к росту спроса на мировых рынках, что, в свою очередь, вызвало неконтролируемый и чрезмерный вылов, который наряду со вспышками заболеваний поставил под угрозу дикие популяции, причём ряд местообитаний был полностью разграблен. Аквакультура фрагментов губок представляет собой альтернативу сбору диких особей, поскольку регенерация фрагментов проста, дешева и эффективна. В качестве объекта нашего исследования мы выбрали Spongia anclotea, обычный вид для тропической западной Атлантики, из-за его высокого спроса на рынке косметики, средств по уходу за телом и фармацевтических препаратов.
Аннотация
Мы организовали низкозатратную ферму по разведению губок из недорогих материалов, доступную для местных сообществ и не требующую значительного обслуживания. В течение четырёхлетнего периода изучались рост и выживаемость 384 из 2304 случайно выбранных губок. По окончании исследования объём губок увеличился в среднем на 380% ± 275% от первоначального (с максимумом в 1480%), а выживаемость составила 87,5% (смертность и отсоединение в течение всего периода учитывались как гибель). Выращенные на нашей ферме губки отличались более высоким качеством и лучшей формой по сравнению с дикими особями, собранными в этом же районе, а после шести лет культивирования (4 года настоящего исследования + 2 года прогнозируемого роста на основе наших данных) это улучшенное качество могло бы обеспечить более высокую прибыль (618 долл. США против 547 долл. США соответственно для стандартного размера партии). Данная экспериментальная установка может считаться хорошей альтернативой сбору диких губок и выгодной экономической возможностью для развивающихся стран.
1 Введение
Губки (тип Porifera) представляют собой монофилетическую группу (Wörheide et al., 2012), насчитывающую около 9 500 описанных видов (de Voogd et al., 2024). Исторически, начиная с эпохи Римской империи, небольшая часть губок, относящихся к отряду Dictyoceratida (семейство Spongiidae), характеризующихся высококачественным спонгиновым скелетом благодаря почти полному отсутствию спикул, большой водоудерживающей способностью, эластичностью и устойчивостью к износу, добывалась для косметических целей в Средиземном море (Pronzato и Manconi, 2008; Ehrlich et al., 2018; Jesionowski et al., 2018). В последнее время было показано, что губки могут быть полезны и рентабельны в различных областях, таких как биомедицина и фармацевтика — благодаря открытию и применению их вторичных метаболитов (Faulkner et al., 1993; Amigó et al., 2008; Indraningrat et al., 2016; Abdelaleem et al., 2020; Pawlik и McMurray, 2020), биомониторинг и биоремедиация — благодаря их способности фильтровать морскую воду, удаляя и накапливая различные классы загрязнителей (Patel et al., 1985; Olesen и Weeks, 1994; Hansen et al., 1995; Philp, 1999; Perez et al., 2002, 2003, 2005; Milanese et al., 2003; Cebrian et al., 2007; Santos-Gandelman et al., 2014; Orani et al., 2018), а также аквакультура — как в качестве целевых фермерских видов, так и в качестве инструмента фильтрации воды для биоремедиации и снижения экологического воздействия фермерства в интегрированных многотрофических аквакультурных системах (IMTA) (Longo et al., 2016, 2020; Giangrande et al., 2020; Gökalp et al., 2021; Aguilo-Arce et al., 2023). Этот рост спроса сделал губок целевым объектом на мировых рынках. Хотя данные до 1900 года для промысловых губок не являются надежными, пик добычи в 2 миллиона губок был зарегистрирован в 1902 году (Sella, 1912), а спрос в начале XX века оценивался в 30 тонн в год. Добыча диких губок со временем увеличилась: более 100 тонн в год в Англии и Германии в 1930-х годах (Arndt, 1938), 200 тонн в год в 1985 году и до около 2000 тонн в 2003 году (FAO Fishery Information and Unit (FAO-FIDI), 2005).
С конца XX века новые открытия о применении губок создали чрезмерную промысловую нагрузку на эти организмы, и вместе со вспышками заболеваний (Webster, 2007) дикие популяции оказались под серьезной угрозой, а в ряде районов наблюдается значительное снижение как численности губок, так и видового богатства (Croft, 1990; Pronzato, 2003; Bertolino et al., 2017).
Эта ситуация привела к активизации усилий по аквакультуре губок по всему миру — от Средиземного моря до Атлантического и Тихого океанов (Cahn, 1948; Storr, 1957, 1964; Croft, 1990; Handley et al., 2003). Исследователи пытались разработать новые методы фермерства, направленные на максимизацию эффективности и снижение затрат. Со временем оценивались эффекты новых фермерских методов (таких как клетки, ограждения, подвесные линии, веревки, сетчатые карманы, шипы и пластиковые штифты (Duckworth et al., 2007; De Caralt et al., 2010; Osinga et al., 2010; Zea et al., 2010; Page et al., 2011; Betanzos-Vega et al., 2019)) на рост, выживаемость и продукцию вторичных метаболитов губок, наряду с другими испытаниями по интегрированной марикультуре с размещением губок в ассоциации с системами аквакультуры рыб и мидий (Gökalp et al., 2019, 2021; Giangrande et al., 2020; Li et al., 2023). На сегодняшний день общепринятой является идея, что аквакультура губок является видоспецифичной, и каждая стратегия фермерства должна быть адаптирована к потребностям целевого организма (Duckworth, 2009; Schippers et al., 2012). В настоящее время одним из наиболее распространенных методов фермерства является выращивание фрагментов, взятых от природных особей, с использованием замечательной регенеративной способности этих организмов. Эта практика впервые была применена в Средиземном море (Cavolini, 1785, 1853), затем распространилась в Америку, Азию и Океанию (Duckworth et al., 1997, 2007; Louden et al., 2007). Аквакультура фрагментов губок открыла новые возможности для развивающихся стран, которые сталкивались с экологическими проблемами, связанными с прошлым выловом природных особей. Фактически, этот метод регенерации фрагментов был признан простым, быстрым, недорогим и эффективным (Ayling, 1983; Duckworth, 2003). При правильных протоколах и условиях одна особь может служить «источником» для нескольких поколений (Osinga et al., 1999; Duckworth, 2001; Belarbi, 2003; Louden et al., 2007), что сокращает потребность в природных донорах (Duckworth, 2009). Эти данные делают фермерское выращивание губок беспроигрышным выбором для сообществ в развивающихся странах, поскольку оно не требует дорогостоящих технологий и инструментов, легко в эксплуатации, рентабельно и, с экологической точки зрения, может снизить антропогенную нагрузку на природные запасы (Yi et al., 2005; Betanzos-Vega et al., 2019).
Род Spongia высоко ценится на рынках благодаря своим специфическим характеристикам, связанным с уходом за телом, включая тонкую текстуру и водоабсорбцию (Verdenal и Verdenal, 1987). Хорошо известный вид Spongia pertusa, ныне признанный как Spongia (Spongia) anclotea de Laubenfels & Storr, 1958, характерен для атлантического побережья Америки от Флориды до Бразилии. Этот вид был выбран из-за его высокой рыночной стоимости для косметики и ухода за телом, а также из-за недавно открытых фармацевтических применений его вторичных метаболитов — как противогрибкового средства против патогенов человека и как ингибиторов пролиферации CSC-подобных клеток рака молочной железы (Osinga et al., 1999; Tang et al., 2018, 2022; Tian et al., 2023). Таким образом, улучшенное понимание его биологии и физиологии имеет основополагающее значение для успешного фермерского выращивания. Насколько нам известно, только одно исследование Oronti et al. (2012) провело краткосрочное предварительное исследование роста и выживаемости S. anclotea на Багамах. Наше исследование является первым долгосрочным исследованием, посвященным адаптивности этого вида к фермерству. В этой работе мы ставили целью исследовать регенерацию и скорость роста S. anclotea, а также выживаемость и отсоединение на экспериментальном участке в Кейп-Элеутера, Багамы. Затем была оценена производительность фермы (в долларах США) и доход от продажи выращенных губок был сравнен с прибылью от продажи диких губок аналогичного размера, собранных в районе исследования. Это позволило нам оценить фактическую применимость и преимущества данной системы фермерства для помощи экономике местных сообществ и снижения нагрузки на природные популяции губок.
2 Материалы и методы
2.1 Район исследования
Исследование проводилось в бухте Кейп-Элеутера (Багамы), недалеко от школы и института Кейп-Элеутера (Рисунок 1). Ферма была установлена в дноуглубленном канале (глубина 3 м) в 15 м от берега. Район был выбран на основе предыдущего пилотного исследования (Oronti et al., 2012), в котором фермерское выращивание Spongia anclotea в этом месте показало более высокие темпы роста и меньшую смертность по сравнению с другими участками в этом районе, вероятно, из-за более высокого наземного стока питательных веществ и меньшей волновой энергии и турбулентности воды. Кроме того, близость к берегу сделала установку, отбор проб и анализ значительно проще по сравнению с удаленными от берега районами.
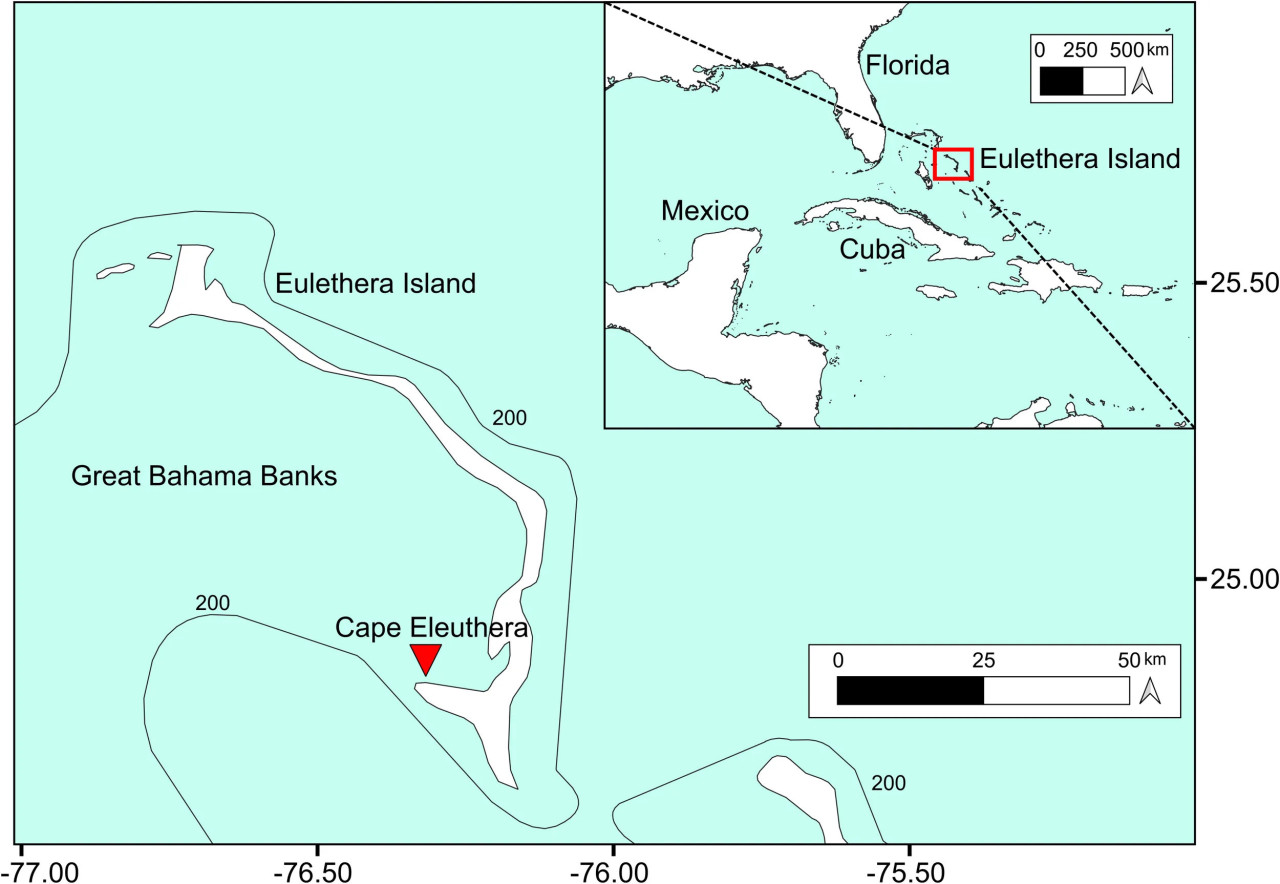
Рисунок 1 Район исследования. Красный треугольник указывает на место установки фермы Spongia anclotea в Кейп-Элеутера, остров Элеутера, Багамы.
2.2 Экспериментальная установка
Всего 8 фермерских ячеек (A-B-C-D-E-F-G-H) были построены в ноябре 2010 года по схеме, представленной на Рисунке 2, на глубине 3 м. Ячейка X уже существовала с предыдущего пилотного исследования (Oronti et al., 2012) и была заполнена более старыми губками, поэтому она не рассматривалась в этом исследовании. Каждая ячейка включала 6 прядей, состоящих из 5 модулей, расположенных на расстоянии 105 см друг от друга. Для этого на дно были уложены 240 бетонных блоков (по 15 кг каждый), каждый из которых имел вертикальный железный стержень (1 метр длиной, 19 мм диаметром), к которому на разной высоте (15 см и 45 см от верхней части стержня) крепились две битумные нейлоновые веревки, натянутые между стержнями одной пряди.
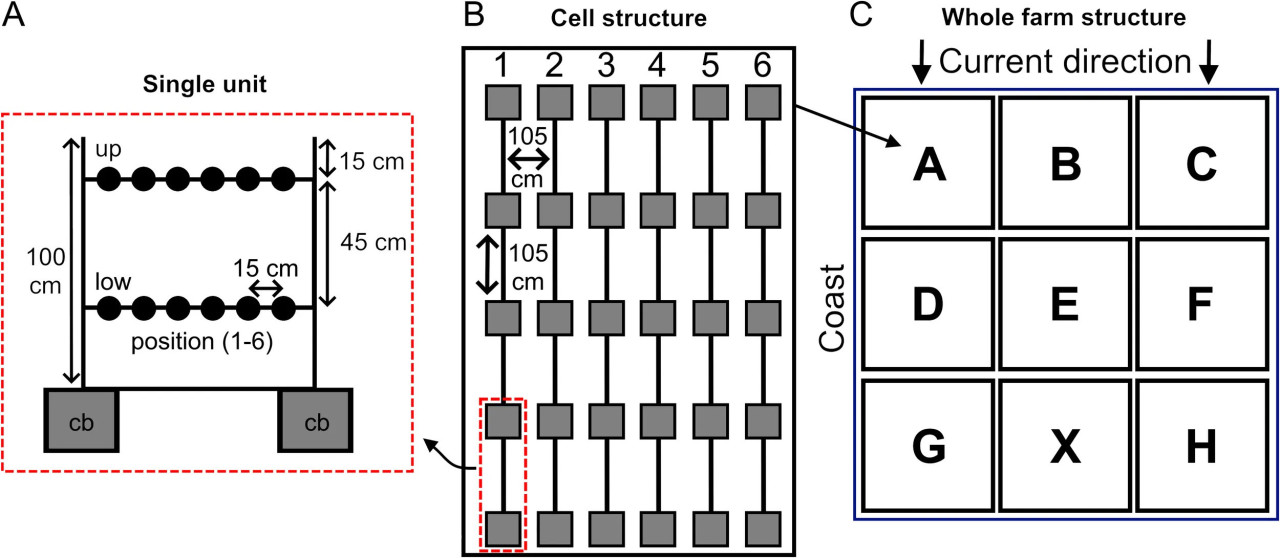
Рисунок 2 Схема фермы губок. (A) Детали одного модуля пряди (полная прядь состоит из 4 модулей). Два бетонных блока (cb) поддерживают два железных стержня, между которыми натянуты две нейлоновые веревки; по 6 губок (черные кружки) на каждой веревке на двух глубинах, верхней и нижней, размещены (всего 24 на полной пряди). (B) Строение одной ячейки из шести прядей (1-6), вид сверху: размеры приблизительно 4 x 5 метров в длину и ширину соответственно. (C) Общая карта фермы вид сверху, показаны все ячейки (A–H). Левая сторона находится ближе к береговой линии, правая сторона обращена к открытому морю. Основное морское течение в этом районе направлено вниз по схеме.
Верхние части (2/3 общего объема) донорских губок из этого района (по оценкам, от 576 до 768 природных особей, объем экспланта составлял от 450 до 600 см³ на донорскую губку) были срезаны ножом для получения фрагментов, оставшиеся 1/3 были оставлены прикрепленными к субстрату для восстановления, как предполагали Duckworth и Battershill (2003). Затем части губок были разделены, с обеспечением того, чтобы по крайней мере одна сторона имела неповрежденную пинакодерму, на множество более мелких субсферических фрагментов (обычно 3-4 фрагмента на донорскую губку). На каждом этапе обработки губки всегда оставались под водой, чтобы избежать воздействия воздуха и минимизировать стресс. Фрагменты затем привязывались пластиковыми стяжками на расстоянии 15 см друг от друга, всего 48 особей на прядь, пронумерованных от 1 до 24 вверх для верхней нити и от 1 до 24 вниз для нижней. Ориентация прядей была установлена параллельно местному течению воды, чтобы минимизировать гидродинамический стресс.
Для анализа роста, из-за большого количества образцов, обрабатывалась только одна случайно выбранная прядь на ячейку, всего 2304 образца (6 прядей x 2 ряда x 24 образца x 8 ячеек).
Эксперимент продолжался с 2010 по 2014 год. Он начался в ноябре 2010 года (установка и первые измерения) с последующими измерениями в июле 2011 года, январе 2012 года, июле 2012 года, феврале 2013 года и сентябре 2014 года. Размеры измерялись под водой с помощью прецизионного штангенциркуля с оценкой длины трех основных осей губок (длина, ширина, высота: x, y, z). Объемы рассчитывались с использованием эллипсоида в качестве эталонной формы (Формула 1), из-за его сходства с формами образцов.
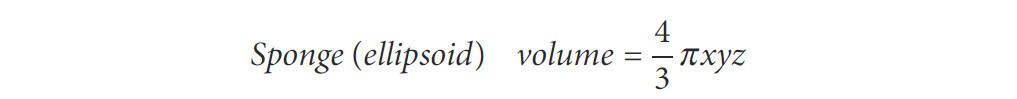
Формула 1. Объем эллипсоида.
Скорость роста, выраженная в процентах, рассчитывалась по Формуле 2.
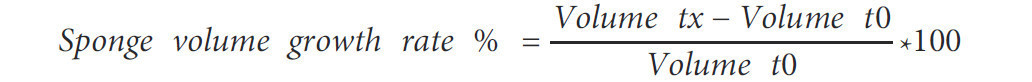
Формула 2. Скорость роста губки. Объем в момент tx показывает объем образца, измеренный в один из пяти моментов отбора проб (t1-t5), в то время как объем в момент t0 представляет собой начальное измерение объема этого образца.
Уровень выживаемости оценивался с учетом как гибели, так и отсоединения губок, поскольку оба исхода приводили к потере особи для финальной продажи.
2.3 Экономический анализ
Были оценены общие затраты на первоначальную установку и ежегодное обслуживание, включая материалы (бетонные блоки, железные стержни, битумные нейлоновые веревки, пластиковые стяжки) и рабочую силу (подготовка конструкций с аквалангом, мониторинг и измерение губок). По окончании исследования качество и рыночная стоимость (оптовая цена) выращенных губок были изучены экспертами в этой области из фермы губок Spugnificio Rosenfield в Мудже, Италия, профессиональная деятельность которой заключается в импорте оптом фермерских или выловленных губок, их обработке и моделировании для получения высоко востребованных сферических форм и последующей перепродаже. Качество губок в первую очередь зависит от формы и текстуры их скелета. Дикие губки, растущие на субстрате, развивают неправильную морфологию и текстуру, которые менее коммерчески привлекательны. Напротив, губки, выращенные на подвесных линиях без пространственных ограничений, растут свободно во всех направлениях, что приводит к более сферической форме и однородной текстуре, которые, вероятно, будут определены как 1-й сорт и пользуются высоким спросом на рынке. Наконец, потенциальная прибыль для фермеров (доходы минус затраты) была оценена и сравнена со средней прибылью от продажи собранных диких губок аналогичного размера. Были сделаны две оценки: на t4 (3 года), когда губки достигли коммерческого размера, и прогнозируемый t6 (4 года текущего исследования плюс 2 года прогнозируемого роста на основе наших данных, при предположении постоянного роста и отсутствия потенциальных стрессовых событий), когда производительность фермы стабилизируется. Количество товарных губок было оценено на основе выживших особей на t4. На t6 разница в количестве не предполагалась, исходя из предпосылки, что в действующей ферме (в отличие от нашего исследования, где мертвые/отсоединившиеся губки намеренно не заменялись) пустые места постоянно заполняются, что минимизирует потерю особей. Мы рассматривали в качестве дохода только оптовую цену губок, исключая дополнительные затраты для покупателей, поскольку они не имели отношения к фокусу исследования на экономической осуществимости фермы. Аналогично, для собранных губок мы решили рассматривать в качестве прибыли оптовую цену для покупателей. Мы исключили наличие значительных дополнительных расходов для сборщиков губок, поскольку большая часть сбора губок в Карибском бассейне осуществляется с помощью «багров и крюков» или ныряния с задержкой дыхания на мелководье с небольшими кустарными лодками в качестве поддержки, с почти отсутствующими расходами и оборудованием. На Багамах сбор губок в сельских местных сообществах является эпизодической деятельностью, осуществляемой рыбаками. По этой причине стоимость лодки не учитывалась, поскольку она уже покрывается их основными рыболовными операциями и не представляет дополнительных расходов. Поскольку исследование закончилось в 2014 году, цены были обновлены не простой конвертацией прошлой и текущей глобальной стоимости доллара. Мы исследовали текущую местную стоимость на Багамах каждого компонента как для затрат, так и для доходов и сообщили их в американских долларах (USD).
2.4 Данные о переменных окружающей среды из спутниковых данных и анализ аномалий
Чтобы предоставить общий обзор физических условий во время эксперимента, мы сообщили о сезонных вариациях и аномалиях нескольких переменных окружающей среды в течение периода исследования. Спутниковые данные о температуре морской воды, хлорофилле-a, кислороде, солености, нитратах, фосфатах, силикатах и биомассе для экспериментального участка и временного периода были получены из продукта Global Ocean Physics Reanalysis (doi.org/10.48670/moi-00021), набор данных "cmems_mod_glo_phy_my_0.083deg_P1D-m", и продукта Global Ocean Biogeochemistry Hindcast (https://doi.org/10.48670/moi-00019), набор данных "cmems_mod_glo_bgc_my_0.25deg_P1D-m", в Copernicus Marine Service. Детали описаны в Дополнительных данных 1.
Данные о температуре (с 1987 по 2021 год) были проанализированы с использованием пакета R "heatwaveR" для оценки морских волн тепла "MHW" и холодных периодов "MCS" в течение периода исследования, детали протокола описаны в Дополнительных данных 2. Следуя той же процедуре, аномалии выше 90-го процентиля и ниже 10-го процентиля были также обнаружены для каждой другой переменной (в этом случае климатология рассчитывалась за период 1993-2021 годов из-за ограниченной протяженности доступных наборов данных).
2.5 Статистический анализ
Обобщенные линейные смешанные модели (GLMM) были запущены для изучения статистических различий в скорости роста между (1) рядами (верхний-нижний), (2) позициями (1-24), (3) ячейками (A-H) и их взаимодействием со временем (Дополнительная таблица 1). Для (1) мы задали GLMM со скоростью роста как зависимой переменной, взаимодействием ряда и времени как фиксированными факторами и идентификатором образца как случайным фактором. Для (2) была задана GLMM со скоростью роста как зависимой переменной, взаимодействием позиции и времени как фиксированными факторами и идентификатором образца как случайным фактором. Для (3) была задана GLMM со скоростью роста как зависимой переменной, взаимодействием ячейки и времени как фиксированными факторами и идентификатором образца как случайным фактором. Для оценки различий в выживаемости (гибель + отсоединение) губок между рядами, позициями и ячейками были заданы три биномиальные GLMM с живыми/мертвыми организмами (как 0/1) в качестве зависимой переменной (Дополнительная таблица 1). Как и для скорости роста, первая модель включала взаимодействие ряда и времени как фиксированные факторы и идентификатор образца как случайный фактор. Вторая модель включала взаимодействие позиции и времени как фиксированные факторы и идентификатор образца как случайный фактор. Третья модель включала взаимодействие ячеек и времени как фиксированные факторы и идентификатор образца как случайный фактор. Для всех моделей попарные сравнения (тест Тьюки) проводились при обнаружении значительных различий. Статистические модели были разработаны в среде R v.3.6.2 (R Core Team, 2021) с использованием пакета stats (R Core Team, 2021), lme4 (Bates et al., 2015) и emmeans (Lenth, 2023).
3 Результаты
Всего было проанализировано 384 губки 6 раз в течение 4-летнего периода, всего 2304 измерения (1152 верхних и 1152 нижних) (Рисунки 3B–F). Интервалы отбора проб: ноябрь 2010, июль 2011, январь 2012, июль 2012, февраль 2013 и сентябрь 2014.

Рисунок 3 Экспериментальная установка. (A) дикий экземпляр Spongia anclotea, (B) t0, оранжевый цвет указывает на свежие поверхности срезов, (C, D) общий вид района фермы и увеличенное изображение отсоединившейся особи на t1, (E) t4, (F) особи на финальном времени отбора t5.
3.1 Анализ роста и смертности
Средняя скорость увеличения объема губок на t5 составила 380% ± 275%, с минимальным индивидуальным значением, зарегистрированным в ячейке C (-80%), и максимальным показателем в ячейке D (1480%) (Таблица 1).
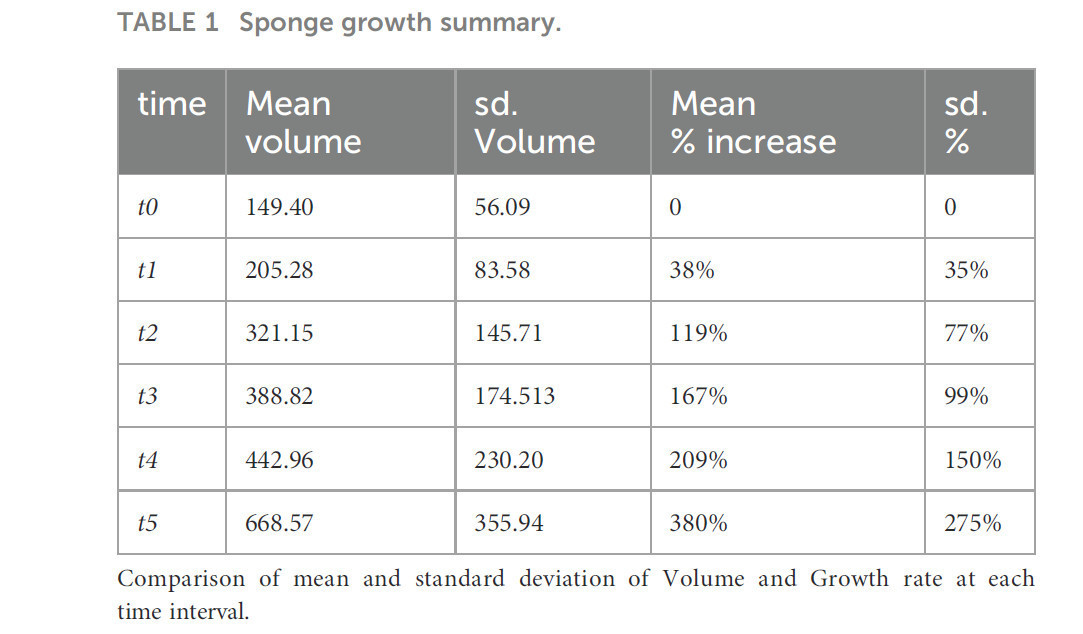
Таблица 1 Сводка по росту губок.
Сравнение средних значений и стандартных отклонений объема и скорости роста на каждом временном интервале.
Анализируя различные части нашей фермерской конструкции, различий в росте между верхними и нижними рядами ячеек не наблюдалось на каждом временном интервале, и различий между позициями 1-24 на двух рядах обнаружено не было (Дополнительная таблица 1). Рассматривая весь период t0-t5, различий в скорости роста между ячейками не наблюдалось. На t5 различия наблюдались только между ячейками C-D и C-G (Рисунок 4; Дополнительная таблица 1).
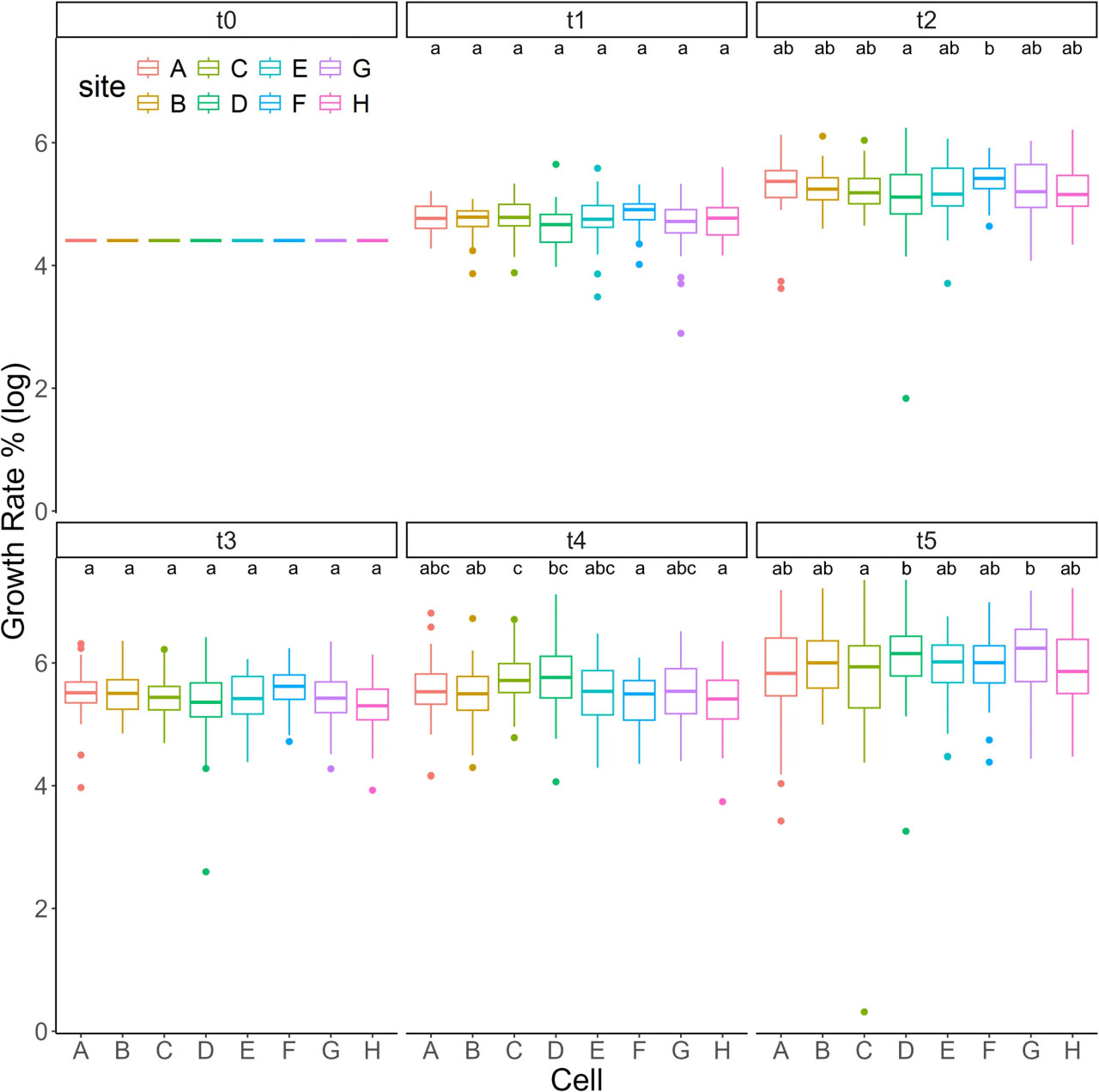
Рисунок 4 Анализ роста. Рост (логарифмическая шкала) S. anclotea для каждой фермерской ячейки (A–H) в разные временные интервалы (t0-t5).
В общей сложности 54 из 384 губок (14%) были потеряны из-за гибели или отсоединения в течение исследования, без статистических различий между рядами, позициями и ячейками (Рисунок 5; Дополнительная таблица 1).
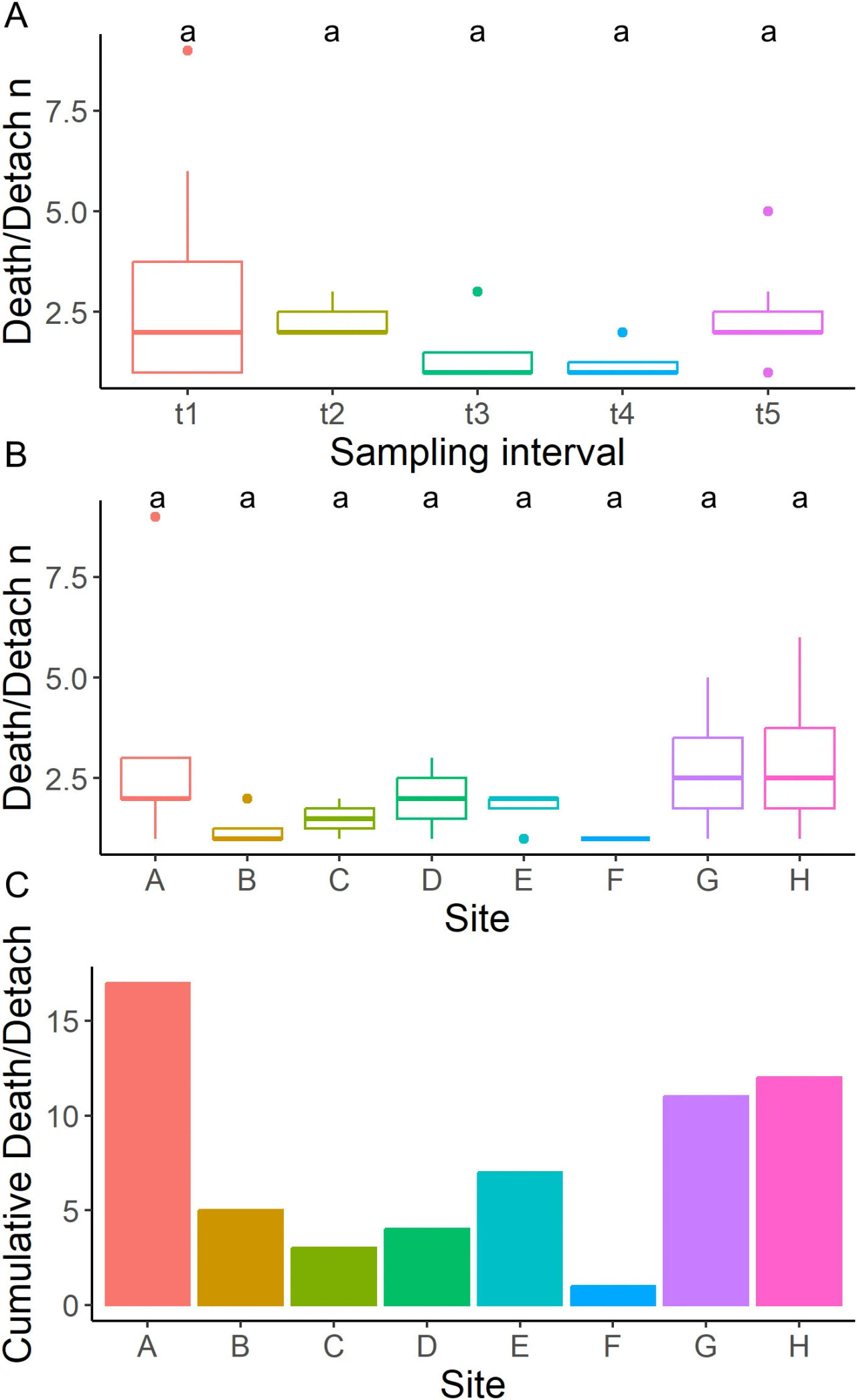
Рисунок 5 Анализ смертности. (A) ящичковая диаграмма отсоединившихся/погибших особей для каждого временного интервала, (B) ящичковая диаграмма отсоединившихся/погибших особей для каждой фермерской ячейки, (C) общее количество отсоединившихся/погибших особей для каждой фермерской ячейки.
По личному наблюдению авторов, начиная с t0, песчано-оранжевые поверхности срезов фрагментов покрывались регенерированной пинакодермой в течение нескольких недель. Мы наблюдали, что способность к самоочищению, столь очевидная у диких экземпляров (Рисунок 3A), была первоначально ослаблена у пересаженных фрагментов, которые имели тенденцию покрываться как осадками, так и эпибионтами, и восстанавливалась только через один или два года после начала культивирования. Интересно, что это восстановление соответствовало восстановлению изначально утраченных, вероятно, из-за срезки и пересадки, эндобионтных организмов, таких как черви, креветки и т.д. На сегодняшний день роль организмов, живущих внутри губок, и природа их ассоциаций все еще обсуждаются; если выгоды для эндобионтов (пища, убежище) оценить легче, то выгода для губки не была определена (Westinga и Hoetjes, 1981; Wulff, 2006; Martin и Britayev, 2018; Goren et al., 2021; Lira et al., 2024). Здесь мы могли бы предположить, как личное размышление, основанное на качественных наблюдениях, потенциальную роль этих организмов в очистке поверхности губок.
3.2 Переменные окружающей среды и аномалии
Волны тепла и холодные периоды, а также аномалии Chl-a, кислорода, солености, нитратов, фосфатов, силикатов и биомассы, рассчитанные за период исследования, представлены на Рисунке 6 и Дополнительном Рисунке 1. Климатические условия в этом районе были довольно стабильными для каждой переменной в течение исследования, без сопутствующих аномалий, за исключением октября/ноября 2012 года, в период t3-t4, когда произошло интенсивное падение температуры от 90-го процентиля до события холодного периода (ниже 10-го процентиля). Одновременно наблюдалась аномалия солености ниже 10-го процентиля и аномальные пики выше 90-го процентиля первичной продуктивности и биомассы в этом районе. Эти аномалии совпали с ураганом Сэнди, обрушившимся на Багамы в конце октября 2012 года. Нитраты, фосфаты и силикаты не показали связанных с ними аномалий.
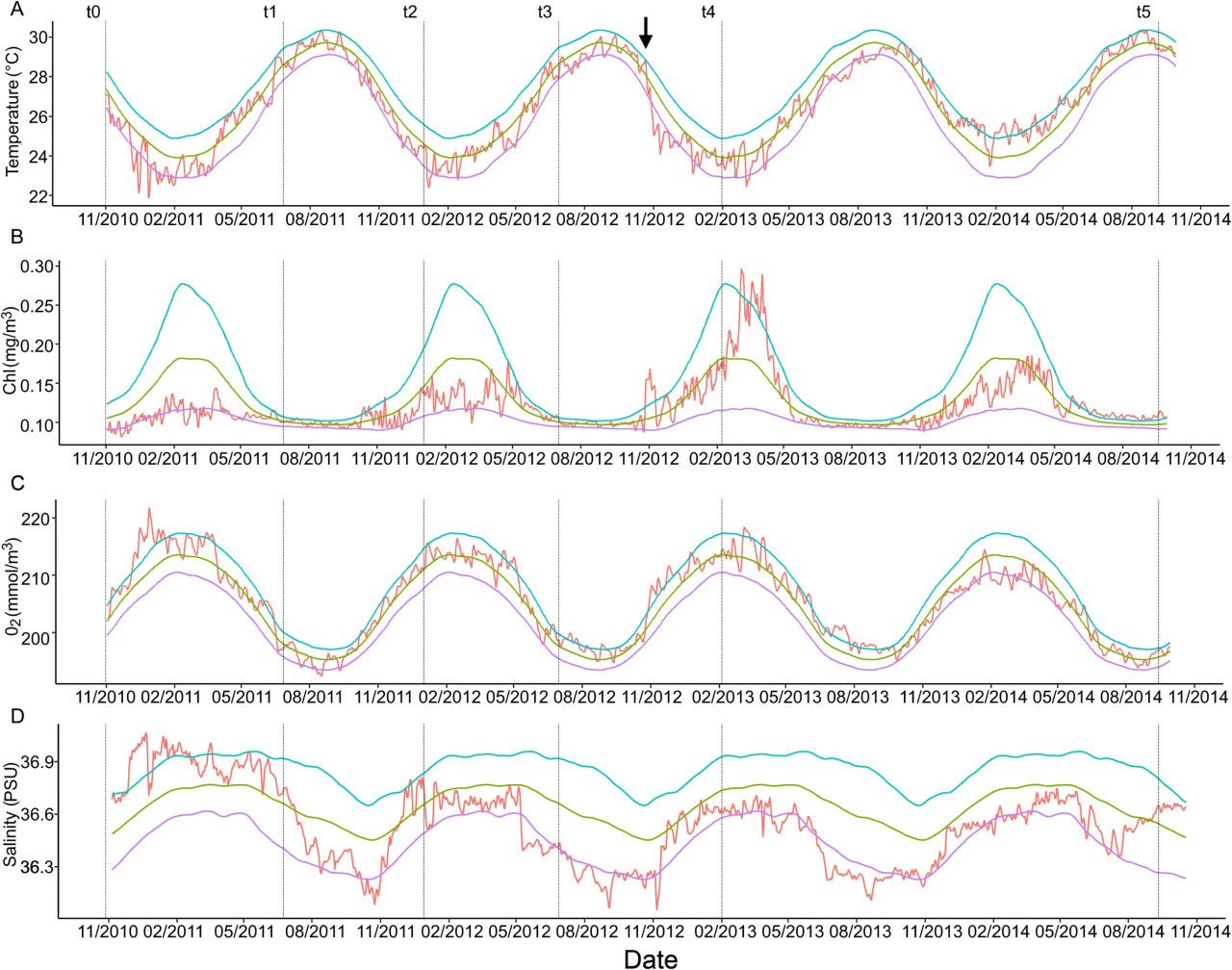
Рисунок 6 Аномалии факторов окружающей среды. Красная линия — наблюдаемое значение переменной, зеленый — климатология, синяя и фиолетовая линии показывают 90-й и 10-й процентили для каждой переменной. (A) Температура, (B) Хлорофилл-a, (C) Кислород и (D) Соленость. Пунктирные линии — временные интервалы t1-t5. Черная стрелка указывает на прибытие урагана Сэнди в район исследования.
3.3 Экономический анализ
Эксперты фермы губок Spugnificio Rosenfield из Муджи, в лице Елены Песле, заявили, что губки достигли продажного размера на t4 и определили их качество как первый сорт для 80% губок, остальные 20% — второй сорт. Под их наблюдением мы оценили затраты (все значения обновлены до 2023 года) на первоначальную установку в 387 долларов плюс 15 долларов на ежегодные работы по обслуживанию, в общей сложности 432 доллара на t4. Обслуживание заключалось в основном в проверке целостности конструкции и очистке веревок от обрастания — работы, которые могут выполняться операторами фермы на ежемесячной основе, без необходимости привлечения внешних экспертов и дополнительных затрат (Таблица 2). Стандартная партия собранных диких губок товарного размера, обычно состоящая из 50% первого сорта и 50% второго сорта, стоит 1 доллар за губку, в то время как фермерская высококачественная партия того же размера стоит около 2-2,5 доллара за губку. На t4 выжило 348 из 384 губок, и это число было использовано для анализа. Основываясь на наших результатах по росту и выживаемости фермерских губок, на гипотетическом прогнозируемом t6 (при предположении постоянного роста и отсутствия потенциальных стрессовых событий) прибыль от выращенных особей Spongia anclotea превысит прибыль от сбора диких губок (618 долларов против 547 долларов) (Таблица 3).
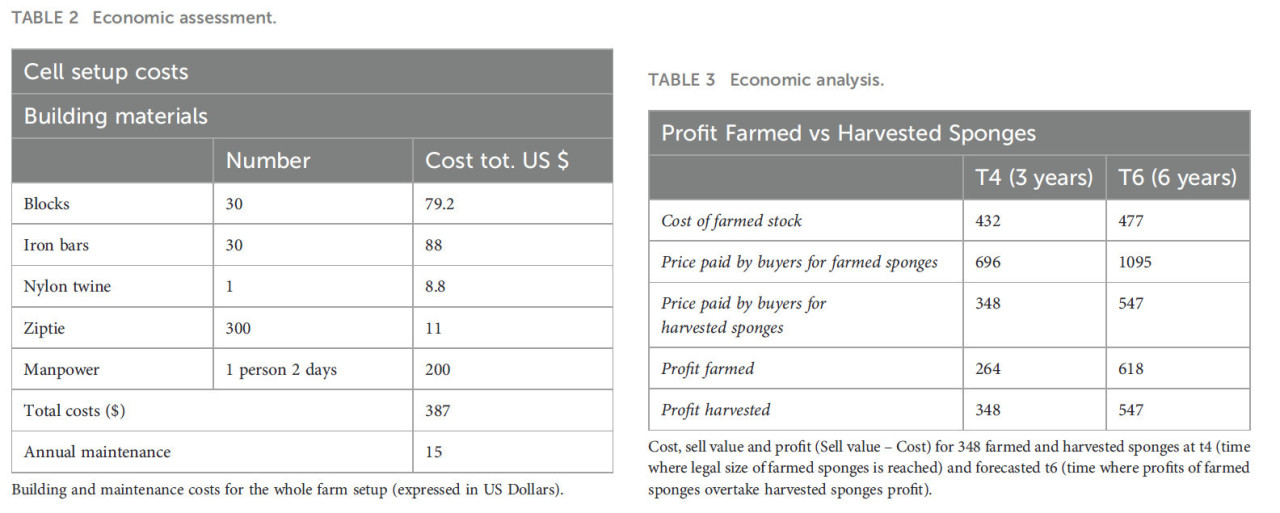
Таблица 2 Экономическая оценка.
Затраты на строительство и обслуживание всей фермерской установки (выражены в долларах США).
Таблица 3 Экономический анализ.
Затраты, стоимость продажи и прибыль (Стоимость продажи – Затраты) для 348 фермерских и выловленных губок на t4 (время, когда фермерские губки достигают товарного размера) и прогнозируемом t6 (время, когда прибыль фермерских губок превышает прибыль выловленных губок).
4 Обсуждение
Морские губки являются фундаментальным компонентом бентической экосистемы благодаря их численности, функциям и услугам, таким как предоставление убежищ (Herrnkind et al., 1997; Coppock et al., 2022), биоремедиация (Amato et al., 2024) и фильтрация воды (Dayton et al., 1974; Ayling, 1981; Costello и Myers, 1987; Maldonado et al., 2012, 2017; De Goeij et al., 2013; Pawlik и McMurray, 2020). В коммерческом отношении спрос на губки постоянно растет для косметических, промышленных, фармацевтических применений, и поставки от природных популяций, которые в основном расположены в развивающихся странах и подвергаются чрезмерному стрессу, становятся все менее способными удовлетворять спрос. На современном уровне знаний фермерское выращивание губок требует простых установок и недорогих инструментов (Verdenal, 1990; Duckworth и Battershill, 2003; Page et al., 2005), что может принести пользу местным сообществам в развивающихся странах, у которых нет крупных капиталов для инвестиций. По этим причинам в нашем исследовании мы решили применить структуру фермерского выращивания губок, которая потенциально отвечает этим потребностям и может быть собрана из простых материалов (бетонные блоки, железные стержни и нейлоновые веревки), требует низких затрат на обработку и обслуживание, обеспечивая при этом эффективный рост губок, как уже наблюдалось во многих исследованиях (Corriero et al., 2004; Friday, 2011; Çelik et al., 2011; Maslin et al., 2021; Bierwirth et al., 2022).
Двумя наиболее важными параметрами в фермерском выращивании губок являются скорость роста и выживаемость (Duckworth, 2009; Santiago et al., 2019; Mohite et al., 2020; Ou et al., 2020). Глядя на наши результаты, значительных различий ни в росте, ни в смертности не было обнаружено для двух глубин, позиции на пряди и ячейки. С конструктивной точки зрения, эти однородные результаты указывают на то, что наша фермерская установка является надежной и эффективной средой фермерства, где, по крайней мере для этого вида, разные глубины фермерства не влияют на рост губок, и так же не влияет позиция на веревке или позиция ячейки. Согласно Duckworth et al. (2007), объем фермерских губок должен увеличиваться на 100% в год, чтобы считаться рентабельным. В нашем исследовании на t5 (4 года) средняя скорость увеличения объема составила 380% (индивидуальные значения варьировались от минимальных -80% до максимальных 1480%). Этот широкий диапазон результатов может быть связан с изменчивостью состояния здоровья, устойчивости и резилентности среди многочисленных донорских особей. Среди донорских губок некоторые особи могли уже находиться в неблагополучном состоянии здоровья или быть менее генетически одаренными, что приводило к уменьшению объема с течением времени, в то время как другие, более приспособленные к росту, демонстрировали наивысшие значения. Однако по сравнению с другими таксонами данные о скорости роста и сокращения объема губок в литературе скудны и ограничены несколькими видами (Ayling, 1983; Barthel, 1986; Hoppe, 1988; Garrabou и Zabala, 2001; Duckworth et al., 2007; De Caralt et al., 2010; Osinga et al., 2010; Zea et al., 2010; Page et al., 2011; Padiglia et al., 2018; Gökalp et al., 2019; Li et al., 2023). Более того, в условиях этого дефицита данных рост фермерских губок был признан видоспецифичным, в значительной степени вариабельным [от отрицательного роста (Hoppe, 1988; De Caralt et al., 2008; Page et al., 2011; Di Camillo et al., 2012) до 2000% увеличения в год (Pronzato и Manconi, 2008; Page et al., 2011; Schippers et al., 2012)] и легко подверженным влиянию нескольких факторов, таких как сезонность, температура воды, прозрачность воды и течения (Barthel и Theede, 1986). Таким образом, четкая картина ростовых показателей в зависимости от разных методов фермерства недоступна (Corriero et al., 2004). Единственное исследование по Spongia anclotea было проведено Oronti et al. (2012), которые сообщили о скорости роста 6,2 см³/месяц в течение трехлетнего периода. Мы зарегистрировали лучшую скорость роста (11,28 см³/месяц), но следует учитывать, что меньший начальный размер фрагментов губок в исследовании Oronti et al. (2012) мог повлиять на скорость роста. В пределах рода Spongia, Çelik et al. (2011) исследовали образцы Spongia officinalis, выращенные на линиях, и зарегистрировали скорости роста 5-17% за два года. Наша система фермерства S. anclotea из Кейп-Элеутера показала лучшие результаты. Было продемонстрировано, что факторы окружающей среды, такие как сезонные вариации температуры воды, потенциально влияют на рост губок. В местах с выраженной сезонностью, таких как умеренные зоны, рост выше в теплый сезон (Handley et al., 2003; Kelly et al., 2004; Page et al., 2005). Аналогично, исследования в Австралии нашли положительную взаимосвязь между скоростью роста и температурой (Duckworth et al., 2007). Другие исследования, напротив, не обнаружили влияния сезонности на рост губок (Ayling, 1983; Hoppe, 1988; Costa et al., 2015). Поскольку Багамы находятся в полутропической зоне, температурный диапазон в нашем районе исследования может быть слишком ограничен, чтобы значительно влиять на скорость роста губок, или S. anclotea может быть одним из тех видов, которые не особенно подвержены влиянию изменений водных условий. Однако в период t3-t4 наблюдалось падение роста в некоторых фермерских ячейках. В то время в Кейп-Элеутера произошел длительный холодный период, вызванный воздействием урагана Сэнди на этот район, что потенциально замедлило рост губок. Учитывая недостаток исследований по росту губок, наши результаты могут помочь заполнить этот пробел в науке о губках.
В аквакультуре другой важной проблемой является смертность организмов, поскольку она означает полную потерю времени и денежных средств. В нашем исследовании смертность была ограниченной: только 54 из 384 губок погибли (или были потеряны) в течение исследования. Verdenal (1990) утверждал, что хорошая ферма должна давать около 90% выживаемости организмов в год. Наша система фермерства показала итоговые 86% за трехлетний период, что соответствует стандарту. Этот результат примечателен по сравнению с показателями выживаемости, сообщаемыми в литературе, которые варьировались от 0 до 100% выживаемости в различных методах фермерства и видах (Osinga, 2010; Gökalp et al., 2019, 2020; Bierwirth et al., 2022). Oronti et al. (2012) зарегистрировали 12,5% смертность для S. anclotea за три года фермерства. Более высокая смертность (14%), которую мы наблюдали, может быть связана с большей продолжительностью нашего исследования. Для S. officinalis, Çelik et al. (2011) сообщили о выживаемости 82-85% образцов, выращенных с помощью метода линий, аналогичного нашему. Из-за отсутствия существующих литературных данных по S. anclotea мы не смогли определить, представляют ли наши результаты по росту и выживаемости просто внутреннюю скорость роста этого вида или на них влияет методология фермерства и местоположение. На нашей ферме большинство потерянных губок были связаны с отсоединением. Гибель губок наблюдалась только в течение первых 12-18 месяцев после пересадки. Мы решили в научных целях не добавлять новые губки на пустые места, но в контексте экономической деятельности было бы довольно легко заполнить пустоты новыми губками, собранными в этом районе (иногда отсоединившиеся губки все еще находятся вблизи ячеек), и поддерживать ферму полностью работоспособной.
Еще одним ключевым фактором для скорости роста и выживаемости является гидродинамика района. Было показано, что скорость роста фермерских губок увеличивается с потоком воды (Wilkinson и Vacelet, 1979), который предоставляет больше пищевых частиц, пока он не становится достаточно сильным, чтобы вызвать физическое повреждение тканей губок (Duckworth et al., 1997; Bannister et al., 2007). Duckworth (2003) обнаружил, что особи Latrunculia wellingtonensis, выращенные в районах с сильным движением воды, выросли в три раза больше по сравнению с районами со слабыми течениями. И наоборот, в районах, где сильные течения вызывают чрезмерное взвешивание или где слабые течения откладывают большие количества частиц, губки могут быть погребены, что имеет пагубное влияние на их способность фильтровать воду и, следовательно, на их выживание (Reiswig, 1971; Osinga et al., 2001; Gökalp et al., 2020). Во время нашего исследования повреждение сильными течениями и повышенное взвешивание, вызванные ураганом Сэнди, могли объяснить слабый рост, наблюдаемый в период t3-t4. Кроме того, зарегистрированный пик Chl-a (скорее всего, цветение водорослей) мог дополнительно нарушить фильтрационную способность губок, приводя к еще более сильному состоянию стресса. За исключением потенциального острого воздействия урагана, других свидетельств влияния гидродинамики не наблюдалось, поскольку рост и смертность губок в различных позициях ячеек и, следовательно, различной подверженности течениям, не показали четкой картины потенциальных взаимодействий. Это предполагает, с одной стороны, что S. anclotea может быть толерантна к средним течениям и взвешиванию, и в то же время, что выбор района (у берега, не подверженного сильным течениям и волнам) и конструкция фермы губок были эффективны для этого вида. Таким образом, это подтверждает, что поиск правильного местоположения и планирование продуманной конструкции фермы является ключевым фактором максимизации общей эффективности и производительности.
Следующим шагом этого исследования была оценка коммерческого качества фермерских губок. Эксперты из фермы губок Spugnificio Rosenfeld в Мудже заявили, что большинство губок уже превышали минимальный коммерческий размер на t4 и что около 80% из них были определены как губки первого сорта, остальные 20% — второго сорта. На t4 общая расчетная прибыль для фермеров за 348 губок составила 264 доллара, в то время как стандартная партия природных выловленных губок (обычно состоящая из 50-50 губок 1-го и 2-го сорта) обычно стоит 348 долларов. В более долгосрочной перспективе, 6 лет, рентабельность нашей фермы превысила стандартную партию при одинаковом объеме губок (618 долларов против 547 долларов). Следует учитывать, что время, необходимое для достижения коммерческого размера, и соответствующий t6 могли бы быть легко сокращены за счет использования начальных фрагментов большего размера. Здесь, поскольку это было пилотное исследование, мы решили получать 3-4 маленьких фрагмента от одной донорской губки. Просто начиная с фрагментов вдвое большего размера, время фермерства сократилось бы вдвое с соответствующими экономическими выгодами для производителей и предотвращением проблем со смертностью при меньших размерах.
В качестве заключения, наши результаты предполагают, что эта система фермерства является действительной альтернативой выловленным губкам, поскольку ее рентабельность в кратко-/среднесрочной перспективе выше. В дополнение к экономическим и коммерческим аспектам, наши результаты также значимы с экологической точки зрения, поскольку поиск новых источников губок, таких как аквакультура, может быть ключевым инструментом для снижения нагрузки на их популяции. Кроме того, наличие открытых морских ферм губок, действующих как районы высокоплотного размножения, может предоставить значительное количество фрагментов, пропагул и личинок для окружающих регионов, способствуя пополнению диких популяций, подверженных промысловому давлению и событиям естественной смертности (Leong и Pawlik, 2010; Pawlik, 2011). С другой стороны, долгосрочное фермерство может увеличить риск ограниченного локального размножения с последующей генетической деградацией с течением времени, что ухудшит их приспособленность и выживаемость (Pérez-Portela et al., 2014). Кроме того, высокая плотность особей может способствовать вспышкам болезней губок, хищников и паразитов, как наблюдалось по всему миру на различных видах и в различных местах (Webster, 2007; Maldonado et al., 2010; Page et al., 2011; Wulff, 2012; Choudhury et al., 2015), которые могут затем распространиться и нанести вред диким популяциям.
Необходимы дальнейшие исследования для проверки эффективности этой фермерской техники в долгосрочной перспективе как на S. anclotea, так и на других коммерчески важных видах губок. Это позволит получить более четкую картину всех потенциальных выгод, как экономических, так и экологических, или неожиданных негативных последствий для экосистем и экономики местных сообществ по всему миру.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти онлайн по адресу: https://www.frontiersin.org/articles/10.3389/fmars.2025.1519832/full#supplementary-material
Ссылки
1. Abdelaleem E. R., Samy M. N., Desoukey S. Y., Liu M., Quinn R. J., Abdelmohsen U. R. (2020). Marine natural products from sponges (Porifera) of the order Dictyoceratida, (2013 to 2019); a promising source for drug discovery. RSC Adv. 10, 34959–34976. doi: 10.1039/D0RA04408C. CrossRef. Google Scholar.
2. Aguilo-Arce J., Ferriol P., Trani R., Puthod P., Pierri C., Longo C. (2023). Sponges as emerging by-product of integrated multitrophic aquaculture (IMTA). J. Mar. Sci. Eng. 11, 80. doi: 10.3390/jmse11010080. CrossRef. Google Scholar.
3. Amato A., Esposito R., Federico S., Pozzolini M., Giovine M., Bertolino M. et al. (2024). Marine sponges as promising candidates for integrated aquaculture combining biomass increase and bioremediation: an updated review. Front. Mar. Sci. 10. doi: 10.3389/fmars.2023.1234225. CrossRef. Google Scholar.
4. Amigó M., Payá M., Braza-Boïls A., De Rosa S., Terencio M. C. (2008). Avarol inhibits TNF-α generation and NF-κB activation in human cells and in animal models. Life Sci. 82, 256–264. doi: 10.1016/j.lfs.2007.11.017. CrossRef. Google Scholar.
5. Arndt W. (1938). "Schwamme," in Die Rohstoffe des Tierreichs 1, 2 Hälfte, Gebr. Eds. Arndt W., Pax F. (Borntraeger, Berlin), 1577–2000. Google Scholar.
6. Ayling A. L. (1983). Growth and regeneration rates in thinly encrusting demospongiae from temperate waters. Biol. Bull. 165, 343–352. doi: 10.2307/1541200. CrossRef. Google Scholar.
7. Ayling A. M. (1981). The role of biological disturbance in temperate subtidal encrusting communities. Ecology 62, 830–847. doi: 10.2307/1937749. CrossRef. Google Scholar.
8. Bannister R. J., Brinkman R., Wolff C., Battershill C., De Nys R. (2007). The distribution and abundance of dictyoceratid sponges in relation to hydrodynamic features: identifying candidates and environmental conditions for sponge aquaculture. Mar. Freshw. Res. 58, 624. doi: 10.1071/MF07011. CrossRef. Google Scholar.
9. Barthel D. (1986). On the ecophysiology of the sponge Halichondria panicea in Kiel Bight. I. Substrate specificity, growth and reproduction. Mar. Ecol. Prog. Ser. 32, 291–298. doi: 10.3354/meps032291. CrossRef. Google Scholar.
10. Barthel D., Theede H. (1986). A new method for the culture of marine sponges and its application for experimental studies. Ophelia 25, 75–82. doi: 10.1080/00785326.1986.10429715. CrossRef. Google Scholar.
11. Bates D., Mächler M., Bolker B., Walker S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Software 67, 1–48. doi: 10.18637/jss.v067.i01. CrossRef. Google Scholar.
12. Belarbi E. (2003). Producing drugs from marine sponges. Biotechnol. Adv. 21, 585–598. doi: 10.1016/S0734-9750(03)00100-9. CrossRef. Google Scholar.
13. Bertolino M., Costa G., Carella M., Cattaneo-Vietti R., Cerrano C., Pansini M. et al. (2017). The dynamics of a Mediterranean coralligenous sponge assemblage at decennial and millennial temporal scales. PloS One 12, e0177945. doi: 10.1371/journal.pone.0177945. CrossRef. Google Scholar.
14. Betanzos-Vega A., Mazón-Suástegui J. M., Formoso-García M., Avilés-Quevedo M. A. (2019). "Sponge fishery and aquaculture in Cuba: impacts and challenges," in Invertebrates - Ecophysiology and Management. Eds. Ray S., Diarte-Plata G., Escamilla-Montes R. (London, UK: IntechOpen), 103–117. doi: 10.5772/intechopen.84785. CrossRef. Google Scholar.
15. Bierwirth J., Mantas T. P., Villechanoux J., Cerrano C. (2022). Restoration of marine sponges—What can we learn from over a century of experimental cultivation? Water 14, 1055. doi: 10.3390/w14071055. CrossRef. Google Scholar.
16. Cahn A. R. (1948). Japanese sponge culture experiments in the South Pacific Islands, General Headquarters, Supreme Commander for the Allied Powers, National Resources Section. (Washington DC: U.S Fish and Wildlife Service, U.S Department of the Interior). Google Scholar.
17. Cavolini F. (1785). Memorie per servire alla storia de'polipi marini (Napoli, Italy: Wentworth Press). Google Scholar.
18. Cavolini F. (1853). Memorie postume sceverate dalle schede autografice di Filippo Cavolini per cura ed a spese di SD Chiaie. Tip. delle Streghe. (Benevento, Italy: Tipografia delle streghe). Google Scholar.
19. Cebrian E., Uriz M.-J., Turon X. (2007). Sponges as biomonitors of heavy metals in spatial and temporal surveys in northwestern Mediterranean: multispecies comparison. Environ. Toxicol. Chem. Int. J. 26, 2430–2439. doi: 10.1897/07-292.1. CrossRef. Google Scholar.
20. Çelik İ., Cirik Ş., Altιnağaç U., Ayaz A., Çelik P., Tekeşoğlu H. et al. (2011). Growth performance of bath sponge (Spongia officinalis Linnaeus 1759) farmed on suspended ropes in the Dardanelles (Turkey): Growth performance of bath sponge (Spongia officinalis) in the Dardanelles (Turkey). Aquac. Res. 42, 1807–1815. doi: 10.1111/j.1365-2109.2010.02781.x. CrossRef. Google Scholar.
21. Choudhury J. D., Pramanik A., Webster N. S., Llewellyn L. E., Gachhui R., Mukherjee J. (2015). The pathogen of the Great Barrier Reef sponge Rhopaloeides odorabile is a new strain of Pseudoalteromonas agarivorans containing abundant and diverse virulence-related genes. Mar. Biotechnol. 17, 463–478. doi: 10.1007/s10126-015-9627-y. CrossRef. Google Scholar.
22. Coppock A. G., Kingsford M. J., Battershill C. N., Jones G. P. (2022). Significance of fish–sponge interactions in coral reef ecosystems. Coral Reefs 41, 1285–1308. doi: 10.1007/s00338-022-02253-8. CrossRef. Google Scholar.
23. Corriero G., Longo C., Mercurio M., Nonnis Marzano C., Lembo G., Spedicato M. T. (2004). Rearing performance of Spongia officinalis on suspended ropes off the Southern Italian Coast (Central Mediterranean Sea). Aquaculture 238, 195–205. doi: 10.1016/j.aquaculture.2004.04.030. CrossRef. Google Scholar.
24. Costa M. F. D. B., Mansur K. F. R., Leite F. P. P. (2015). Temporal variation of the gammaridean fauna (Crustacea, Amphipoda) associated with the sponge Mycale angulosa (Porifera, Demospongiae) in southeastern Brazil. Nauplius 23, 79–87. doi: 10.1590/S0104-64972015002312. CrossRef. Google Scholar.
25. Costello M. J., Myers A. A. (1987). Amphipod fauna of the sponges Halichondria panicea and Hymeniacidon perleve in Lough Hine, Ireland. Mar. Ecol. Prog. Ser. 41, 115–121. doi: 10.3354/meps041115. CrossRef. Google Scholar.
26. Croft R. A. (1990). Recommendations for Establishing a Commercial Sponge Industry Within the Pohnpei Region (Waimanalo, Hawaii: US Department of Agriculture, Center for Tropical and Subtropical Aquaculture). Google Scholar.
27. Dayton P. K., Robilliard G. A., Paine R. T., Dayton L. B. (1974). Biological accommodation in the benthic community at McMurdo Sound, Antarctica. Ecol. Monogr. 44, 105–128. doi: 10.2307/1942321. CrossRef. Google Scholar.
28. De Caralt S., Sánchez-Fontenla J., Uriz M. J., Wijffels R. H. (2010). In situ aquaculture methods for Dysidea avara (Demospongiae, Porifera) in the Northwestern Mediterranean. Mar. Drugs 8, 1731–1742. doi: 10.3390/md8061731. CrossRef. Google Scholar.
29. De Caralt S., Uriz M. J., Wijffels R. H. (2008). Grazing, differential size-class dynamics and survival of the Mediterranean sponge Corticium candelabrum. Mar. Ecol. Prog. Ser. 360, 97–106. doi: 10.3354/meps07365. CrossRef. Google Scholar.
30. De Goeij J. M., Van Oevelen D., Vermeij M. J. A., Osinga R., Middelburg J. J., De Goeij A. F. P. M. et al. (2013). Surviving in a marine desert: the sponge loop retains resources within coral reefs. Science 342, 108–110. doi: 10.1126/science.1241981. CrossRef. Google Scholar.
31. de Voogd N., Alvarez B., Boury-Esnault N., Cárdenas P., Díaz M.-C., Dohrmann M. et al. (2024). World Porifera Database. doi: 10.14284/359. CrossRef. Google Scholar.
32. Di Camillo C., Coppari M., Bartolucci I., Bo M., Betti F., Bertolino M. et al. (2012). Temporal variations in growth and reproduction of Tedania anhelans and Chondrosia reniformis in the North Adriatic Sea. Anc. Anim. New Chall. Dev. Sponge Res. 219, 299–313. doi: 10.1007/s10750-011-0877-z. CrossRef. Google Scholar.
33. Duckworth A. (2001). Farming sponges for chemicals with pharmaceutical potential. World Aquac. Magazine 14, 16–18. Google Scholar.
34. Duckworth A. R. (2003). Effect of wound size on the growth and regeneration of two temperate subtidal sponges. J. Exp. Mar. Biol. Ecol. 287, 139–153. doi: 10.1016/S0022-0981(02)00552-X. CrossRef. Google Scholar.
35. Duckworth A. (2009). Farming sponges to supply bioactive metabolites and bath sponges: A review. Mar. Biotechnol. 11, 669–679. doi: 10.1007/s10126-009-9213-2. CrossRef. Google Scholar.
36. Duckworth A., Battershill C. (2003). Sponge aquaculture for the production of biologically active metabolites: the influence of farming protocols and environment. Aquaculture 221, 311–329. doi: 10.1016/S0044-8486(03)00070-X. CrossRef. Google Scholar.
37. Duckworth A. R., Battershill C. N., Bergquist P. R. (1997). Influence of explant procedures and environmental factors on culture success of three sponges. Aquaculture 156, 251–267. doi: 10.1016/S0044-8486(97)00131-2. CrossRef. Google Scholar.
38. Duckworth A. R., Wolff C., Evans-Illidge E. (2007). "Developing methods for commercially farming bath sponges in tropical Australia," in Porifera Res.-Biodivers. Innov. Sustain. Eds. Custodio M. R., Lobo-Hajdu G., Hajdu E., Muricy G. (Museu Nacional, Rio de Janeiro, Brazil). Google Scholar.
39. Ehrlich H., Wysokowski M., Żółtowska-Aksamitowska S., Petrenko I., Jesionowski T. (2018). Collagens of Poriferan origin. Mar. Drugs 16, 79. doi: 10.3390/md16030079. CrossRef. Google Scholar.
40. FAO Fishery Information, DUnit (FAO-FIDI), S. (2005). Collation, Analysis and Dissemination of Global and Regional Fishery Statistics. (Rome, Italy: Food and Agriculture Organization of the United Nations (FAO)). Google Scholar.
41. Faulkner D., Hai-yin H., Unson M., Bewley C., Garson M. (1993). New metabolites from marine sponges: are symbionts important? Gazzetta Chim. Ital. 123, 301–307. Google Scholar.
42. Friday S. (2011). A Study of Sponge Aquaculture in Jambiani: Is Shallow Farming Feasible? (Brattleboro, USA: Independent Study Project (ISP) Collection). Google Scholar.
43. Garrabou J., Zabala M. (2001). Growth dynamics in four mediterranean demosponges. Estuar. Coast. Shelf Sci. 52, 293–303. doi: 10.1006/ecss.2000.0699. CrossRef. Google Scholar.
44. Giangrande A., Pierri C., Arduini D., Borghese J., Licciano M., Trani R. et al. (2020). An innovative IMTA system: Polychaetes, sponges and macroalgae co-cultured in a Southern Italian in-shore mariculture plant (Ionian Sea). J. Mar. Sci. Eng. 8, 733. doi: 10.3390/jmse8100733. CrossRef. Google Scholar.
45. Gökalp M., Kooistra T., Rocha M. S., Silva T. H., Osinga R., Murk A. J. et al. (2020). The effect of depth on the morphology, bacterial clearance, and respiration of the mediterranean sponge chondrosia reniformis (Nardo 1847). Mar. Drugs 18, 358. doi: 10.3390/md18070358. CrossRef. Google Scholar.
46. Gökalp M., Mes D., Nederlof M., Zhao H., Merijn de Goeij J., Osinga R. (2021). The potential roles of sponges in integrated mariculture. Rev. Aquac. 13, 1159–1171. doi: 10.1111/raq.12516. CrossRef. Google Scholar.
47. Gökalp M., Wijgerde T., Sarà A., De Goeij J. M., Osinga R. (2019). Development of an integrated mariculture for the collagen-rich sponge chondrosia reniformis. Mar. Drugs 17, 29. doi: 10.3390/md17010029. CrossRef. Google Scholar.
48. Goren L., Idan T., Shefer S., Ilan M. (2021). Sponge-associated polychaetes: not a random assemblage. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.695163. CrossRef. Google Scholar.
49. Handley S., Kelly S., Kelly M. (2003). Non-destructive video image analysis method for measuring growth in sponge farming: Preliminary results from the New Zealand bath-sponge Spongia (Heterofibria) manipulatus. N. Z. J. Mar. Freshw. Res. 37, 613–621. doi: 10.1080/00288330.2003.9517192. CrossRef. Google Scholar.
50. Hansen I. V., Weeks J. M., Depledge M. H. (1995). Accumulation of copper, zinc, cadmium and chromium by the marine sponge Halichondria panicea Pallas and the implications for biomonitoring. Mar. pollut. Bull. 31, 133–138. doi: 10.1016/0025-326X(94)00228-2. CrossRef. Google Scholar.
51. Herrnkind W. F., Butler M. J. IV, Hunt J. H., Childress M. (1997). Role of physical refugia: implications from a mass sponge die-off in a lobster nursery in Florida. Mar. Freshw. Res. 48, 759–770. doi: 10.1071/MF97193. CrossRef. Google Scholar.
52. Hoppe W. F. (1988). Growth, regeneration and predation in three species of large coral reef sponges. Mar. Ecol. Prog. Ser. Oldendorf 50, 117–125. doi: 10.3354/meps050117. CrossRef. Google Scholar.
53. Indraningrat A., Smidt H., Sipkema D. (2016). Bioprospecting sponge-associated microbes for antimicrobial compounds. Mar. Drugs 14, 87. doi: 10.3390/md14050087. CrossRef. Google Scholar.
54. Jesionowski T., Norman M., Żółtowska-Aksamitowska S., Petrenko I., Joseph Y., Ehrlich H. (2018). Marine spongin: naturally prefabricated 3D scaffold-based biomaterial. Mar. Drugs 16, 88. doi: 10.3390/md16030088. CrossRef. Google Scholar.
55. Kelly M., Handley S., Page M., Butterfield P., Hartill B., Kelly S. (2004). Aquaculture trials of the New Zealand bath-sponge Spongia (Heterofibria) manipulatus using lanterns. N. Z. J. Mar. Freshw. Res. 38, 231–241. doi: 10.1080/00288330.2004.9517233. CrossRef. Google Scholar.
56. Lenth R. V. (2023). emmeans: Estimated Marginal Means, aka Least-Squares Means. Available online at: https://CRAN.R-project.org/package=emmeans. Google Scholar.
57. Leong W., Pawlik J. R. (2010). Fragments or propagules? Reproductive tradeoffs among Callyspongia spp. from Florida coral reefs. Oikos 119, 1417–1422. doi: 10.1111/j.1600-0706.2010.18092.x. CrossRef. Google Scholar.
58. Li Z., Xia G., Ou H., Chen M., You Y., Chen J. (2023). Large-scale mariculture of the sponge Haliclona simulans on floating rafts in Zhao'an Bay, Fujian Province, China. Aquaculture 563, 738893. doi: 10.1016/j.aquaculture.2022.738893. CrossRef. Google Scholar.
59. Lira A. L., de O., Craveiro N., Paresque K., Fukuda M. V., Rosa Filho J. S. (2024). Ecological review of the Syllidae (Annelida) associated with sponges (Porifera), including the description of a new species from northeastern Brazil. Ocean Coast. Res. 72, e24039. doi: 10.1590/2675-2824072.23117. CrossRef. Google Scholar.
60. Longo C., Cardone F., Corriero G., Licciano M., Pierri C., Stabili L. (2016). The co-occurrence of the demosponge Hymeniacidon perlevis and the edible mussel Mytilus galloprovincialis as a new tool for bacterial load mitigation in aquaculture. Environ. Sci. pollut. Res. 23, 3736–3746. doi: 10.1007/s11356-015-5587-z. CrossRef. Google Scholar.
61. Longo C., Scrascia M., Trani R., Pierri C., Cariglia A., Cariglia F. et al. (2020). "Assesment of sponge mariculture potential in polyculture system in Manfredonia Gulf toward the IMTA implementation," in Proceedings of the Aquafarm Novelfarm, Pordenone, Italy, 19–20 February 2020. Google Scholar.
62. Louden D., Inderbitzin S., Peng Z., De Nys R. (2007). Development of a new protocol for testing bath sponge quality. Aquaculture 271, 275–285. doi: 10.1016/j.aquaculture.2007.06.010. CrossRef. Google Scholar.
63. Maldonado M., Aguilar R., Bannister R. J., Bell J. J., Conway K. W., Dayton P. K. et al. (2017). "Sponge grounds as key marine habitats: A synthetic review of types, structure, functional roles, and conservation concerns," in Marine Animal Forests: The Ecology of Benthic Biodiversity Hotspots. Eds. Rossi S., Bramanti L., Gori A., Orejas Saco del Valle C. (Springer International Publishing, Cham), 1–39. doi: 10.1007/978-3-319-17001-5_24-1. CrossRef. Google Scholar.
64. Maldonado M., Ribes M., Van Duyl F. C. (2012). "Nutrient fluxes through sponges," in Advances in Marine Biology (Elsevier), 113–182. doi: 10.1016/B978-0-12-394283-8.00003-5. CrossRef. Google Scholar.
65. Maldonado M., Sánchez-Tocino L., Navarro C. (2010). Recurrent disease outbreaks in corneous demosponges of the genus Ircinia: epidemic incidence and defense mechanisms. Mar. Biol. 157, 1577–1590. doi: 10.1007/s00227-010-1431-7. CrossRef. Google Scholar.
66. Martin D., Britayev T. A. (2018). "Symbiotic polychaetes revisited: an update of the known species and relationships, (1998–2017)," in Oceanography and marine Biology (Boca Raton, USA: CRC Press), 371–447. doi: 10.1201/9780429454455-6. CrossRef. Google Scholar.
67. Maslin M., Gaertner-Mazouni N., Debitus C., Joy N., Ho R. (2021). Marine sponge aquaculture towards drug development: An ongoing history of technical, ecological, chemical considerations and challenges. Aquac. Rep. 21, 100813. doi: 10.1016/j.aqrep.2021.100813. CrossRef. Google Scholar.
68. Milanese M., Chelossi E., Manconi R., Sara A., Sidri M., Pronzato R. (2003). The marine sponge Chondrilla nucula Schmidt 1862 as an elective candidate for bioremediation in integrated aquaculture. Biomol. Eng. 20, 363–368. doi: 10.1016/s1389-0344(03)00052-2. CrossRef. Google Scholar.
69. Mohite S., Mahakal B., Bhatkar H., Patil S. (2020). Culture of marine sponges under controlled conditions. J. Exp. Zool. India. 23, 753–757. Google Scholar.
70. Olesen T., Weeks J. (1994). Accumulation of Cd by the marine sponge Halichondria panicea Pallas: effects upon filtration rate and its relevance for biomonitoring. Bull. Environ. Contam. Toxicol. 52, 722–728. doi: 10.1007/BF00195494. CrossRef. Google Scholar.
71. Orani A. M., Barats A., Zitte W., Morrow C., Thomas O. P. (2018). Comparative study on the bioaccumulation and biotransformation of arsenic by some northeastern Atlantic and northwestern Mediterranean sponges. Chemosphere 201, 826–839. doi: 10.1016/j.chemosphere.2018.03.078. CrossRef. Google Scholar.
72. Oronti A., Danylchuk A. J., Elmore C. E., Auriemma R., Pesle G. (2012). Assessing the feasibility of sponge aquaculture as a sustainable industry in The Bahamas. Aquac. Int. 20, 295–303. doi: 10.1007/s10499-011-9457-5. CrossRef. Google Scholar.
73. Osinga R. (2010). Diet Composition of Two Temperate Calcareous Sponges: Leucosolenia eChinata and Leucetta sp. from the Wellington South Coast, New Zealand. Open Mar. Biol. J. 4, 74–81. doi: 10.2174/1874450801004010074. CrossRef. Google Scholar.
74. Osinga R., Kleijn R., Groenendijk E., Niesink P., Tramper J., Wijffels R. H. (2001). Development of In Vivo Sponge Cultures: Particle Feeding by the Tropical Sponge Pseudosuberites aff. andrewsi. Mar. Biotechnol. 3, 0544–0554. doi: 10.1007/s1012601-0078-2. CrossRef. Google Scholar.
75. Osinga R., Sidri M., Cerig E., Gokalp S. Z., Gokalp M. (2010). Sponge aquaculture trials in the East-Mediterranean Sea: new approaches to earlier ideas. Open Mar. Biol. J. 4, 74–81. doi: 10.2174/1874450801004010074. CrossRef. Google Scholar.
76. Osinga R., Tramper J., Wijffels R. H. (1999). Cultivation of marine sponges. Mar. Biotechnol. 1, 509–532. doi: 10.1007/PL00011807. CrossRef. Google Scholar.
77. Ou H., Zhai J., Wang D., Zhao J., Chen M., Ding S. et al. (2020). Cultivation of sponge Haliclona simulans juveniles in a floating sea raft. Aquaculture 529, 735660. doi: 10.1016/j.aquaculture.2020.735660. CrossRef. Google Scholar.
78. Padiglia A., Ledda F. D., Padedda B. M., Pronzato R., Manconi R. (2018). Long-term experimental in situ farming of Crambe crambe (Demospongiae: Poecilosclerida). PeerJ 6, e4964. doi: 10.7717/peerj.4964. CrossRef. Google Scholar.
79. Page M. J., Handley S. J., Northcote P. T., Cairney D., Willan R. C. (2011). Successes and pitfalls of the aquaculture of the sponge Mycale hentscheli. Aquaculture 312, 52–61. doi: 10.1016/j.aquaculture.2010.12.006. CrossRef. Google Scholar.
80. Page M. J., Northcote P. T., Webb V. L., Mackey S., Handley S. J. (2005). Aquaculture trials for the production of biologically active metabolites in the New Zealand sponge Mycale hentscheli (Demospongiae: Poecilosclerida). Aquaculture 250, 256–269. doi: 10.1016/j.aquaculture.2005.04.069. CrossRef. Google Scholar.
81. Patel B., Balani M., Patel S. (1985). Sponge 'sentinel'of heavy metals. Sci. Total Environ. 41, 143–152. doi: 10.1016/0048-9697(85)90184-6. CrossRef. Google Scholar.
82. Pawlik J. R. (2011). The chemical ecology of sponges on Caribbean reefs: natural products shape natural systems. BioScience 61, 888–898. doi: 10.1525/bio.2011.61.11.8. CrossRef. Google Scholar.
83. Pawlik J. R., McMurray S. E. (2020). The emerging ecological and biogeochemical importance of sponges on coral reefs. Annu. Rev. Mar. Sci. 12, 315–337. doi: 10.1146/annurev-marine-010419-010807. CrossRef. Google Scholar.
84. Perez T., Longet D., Schembri T., Rebouillon P., Vacelet J. (2005). Effects of 12 years' operation of a sewage treatment plant on trace metal occurrence within a Mediterranean commercial sponge (Spongia officinalis, Demospongiae). Mar. pollut. Bull. 50, 301–309. doi: 10.1016/j.marpolbul.2004.11.001. CrossRef. Google Scholar.
85. Perez T., Sarrazin L., Rebouillon P., Vacelet J. (2002). First evidences of surfactant biodegradation by marine sponges (Porifera): an experimental study with a linear alkylbenzenesulfonate. Hydrobiologia 489, 225–233. doi: 10.1023/A:1023217218585. CrossRef. Google Scholar.
86. Perez T., Wafo E., Fourt M., Vacelet J. (2003). Marine sponges as biomonitor of polychlorobiphenyl contamination: concentration and fate of 24 congeners. Environ. Sci. Technol. 37, 2152–2158. doi: 10.1021/es026234v. CrossRef. Google Scholar.
87. Pérez-Portela R., Noyer C., Becerro M. (2014). Genetic structure and diversity of the endangered bath sponge Spongia lamella. Aquat. Conserv. Mar. Freshw. Ecosyst. 25, 365–379. doi: 10.1002/aqc2423. CrossRef. Google Scholar.
88. Philp R. (1999). Cadmium content of the marine sponge Microciona prolifera, other sponges, water and sediment from the eastern Florida panhandle: possible effects on Microciona cell aggregation and potential roles of low pH and low salinity. Comp. Biochem. Physiol. C Pharmacol. Toxicol. Endocrinol. 124, 41–49. doi: 10.1016/S0742-8413(99)00046-8. CrossRef. Google Scholar.
89. Pronzato R. (2003). Mediterranean Sponge Fauna: a biological, historical and cultural heritage. Biogeogr. – J. Integr. Biogeogr. 24, 91–99. doi: 10.21426/B6110118. CrossRef. Google Scholar.
90. Pronzato R., Manconi R. (2008). Mediterranean commercial sponges: over 5000 years of natural history and cultural heritage. Mar. Ecol. 29, 146–166. doi: 10.1111/j.1439-0485.2008.00235.x. CrossRef. Google Scholar.
91. R Core Team (2021). R: A Language and Environment for Statistical Computing (Vienna, Austria: R Foundation for Statistical Computing). Available at: https://www.R-project.org/. Google Scholar.
92. Reiswig H. M. (1971). Particle feeding in natural populations of three marine demosponges. Biol. Bull. 141, 568–591. doi: 10.2307/1540270. CrossRef. Google Scholar.
93. Santiago V. S., Manzano G. G., Yu C. C., Aliño P. M., Salvador-Reyes L. A. (2019). Mariculture potential of renieramycin-producing Philippine blue sponge Xestospongia sp. (Porifera: Haplosclerida). Aquaculture 502, 356–364. doi: 10.1016/j.aquaculture.2018.12.059. CrossRef. Google Scholar.
94. Santos-Gandelman J. F., Cruz K., Crane S., Muricy G., Giambiagi-deMarval M., Barkay T. et al. (2014). Potential application in mercury bioremediation of a marine sponge-isolated Bacillus cereus strain Pj1. Curr. Microbiol. 69, 374–380. doi: 10.1007/s00284-014-0597-5. CrossRef. Google Scholar.
95. Schippers K. J., Sipkema D., Osinga R., Smidt H., Pomponi S. A., Martens D. E. et al. (2012). Cultivation of sponges, sponge cells and symbionts: achievements and future prospects. Adv. Mar. Biol. 62, 273–337. doi: 10.1016/B978-0-12-394283-8.00006-0. CrossRef. Google Scholar.
96. Sella M. (1912). La pesca delle Spugne nella Libia Vol. 13 (Venezia: Memorie del Regio Comitato Talassografico Italiano), 1–153. Google Scholar.
97. Storr J. F. (1957). The sponge industry of Florida (Miami, USA: State of Florida, Board of Conservation). Educational Series No. 9. Google Scholar.
98. Storr J. F. (1964). Ecology of the Gulf of Mexico commercial sponges and its relation to the fishery (United States Fish and Wildlife Service). Special Scientific Report-Fisheries No. 466. Google Scholar.
99. Tang J., Wu W., Yang F., Liu L., Yang Z., Liu L. et al. (2018). Marine sponge-derived smenospongine preferentially eliminates breast cancer stem-like cells via p38/AMPK α pathways. Cancer Med. 7, 3965–3976. doi: 10.1002/cam4.1640. CrossRef. Google Scholar.
100. Tang W.-Z., Zhao H.-M., Tian Y., Dai S.-W., Zhang A., Lin H.-W. et al. (2022). Merosesquiterpenes from the marine sponge Spongia pertusa Esper and their antifungal activities. Tetrahedron Lett. 93, 153690. doi: 10.1016/j.tetlet.2022.153690. CrossRef. Google Scholar.
101. Tian X.-H., Hong L.-L., Jiao W.-H., Lin H.-W. (2023). Natural sesquiterpene quinone/quinols: Chemistry, biological activity, and synthesis. Nat. Prod. Rep. 40, 718–749. doi: 10.1039/D2NP00045H. CrossRef. Google Scholar.
102. Verdenal B. (1990). Sponge culture on vertical ropes in the Northwestern Mediterranean Sea. New Perspect. Sponge Biol. 1, 416–424. Google Scholar.
103. Verdenal B., Verdenal M. (1987). Evaluation de l'intérêt economique de la culture d'eponges commerciales sur les côtes méditerranéennes françaises. Aquaculture 64, 9–29. doi: 10.1016/0044-8486(87)90202-X. CrossRef. Google Scholar.
104. Webster N. S. (2007). Sponge disease: a global threat? Environ. Microbiol. 9, 1363–1375. doi: 10.1111/j.1462-2920.2007.01303.x. CrossRef. Google Scholar.
105. Westinga E., Hoetjes P. (1981). The intrasponge fauna of Spheciospongia vesparia (Porifera, Demospongiae) at Curaçao and Bonaire. Mar. Biol. 62, 139–150. doi: 10.1007/BF00388176. CrossRef. Google Scholar.
106. Wilkinson C. R., Vacelet J. (1979). Transplantation of marine sponges to different conditions of light and current. J. Exp. Mar. Biol. Ecol. 37, 91–104. doi: 10.1016/0022-0981(79)90028-5. CrossRef. Google Scholar.
107. Wörheide G., Dohrmann M., Erpenbeck D., Larroux C., Maldonado M., Voigt O. et al. (2012). "Chapter one - deep phylogeny and evolution of sponges (Phylum porifera)," in Advances in Sponge Science: Phylogeny, Systematics, Ecology. Eds. Becerro M. A., Uriz M. J., Maldonado M., Turon X. (Academic Press), 1–78. doi: 10.1016/B978-0-12-387787-1.00007-6. CrossRef. Google Scholar.
108. Wulff J. (2006). Ecological interactions of marine sponges. Can. J. Zool. 84, 146–166. doi: 10.1139/Z06-019. CrossRef. Google Scholar.
109. Wulff J. (2012). "Chapter four - ecological interactions and the distribution, abundance, and diversity of sponges," in Advances in Sponge Science: Phylogeny, Systematics, Ecology. Eds. Becerro M. A., Uriz M. J., Maldonado M., Turon X. (Academic Press), 273–344. doi: 10.1016/B978-0-12-387787-1.00003-9. CrossRef. Google Scholar.
110. Yi Q., Wei Z., Hua L., Xingju Y., Meifang J. (2005). Cultivation of marine sponges. Chin. J. Oceanol. Limnol. 23, 194–198. doi: 10.1007/BF02894238. CrossRef. Google Scholar.
111. Zea S., Castellanos L., Valderrama K., Puentes C., Gomez-Leon J., Pomponi S. (2010). "Potential of antitumoral (þ)-discodermalide production by the Caribbean sponge Discodermia dissoluta," in VIII World Sponge Conference, (Girona, Spain: World Sponge Conference). Google Scholar.


Motta G, Auriemma R, Brooks A, Novak M, Pesle E, Terlizzi A and Avian M (2025) A cheap and efficient system for Spongia anclotea farming in the Bahamas. Front. Mar. Sci. 12:1519832. doi: 10.3389/fmars.2025.1519832
Перевод статьи «A cheap and efficient system forSpongia ancloteafarming in the Bahamas» авторов Motta G, Auriemma R, Brooks A, Novak M, Pesle E, Terlizzi A and Avian M., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык
Фото: Продажа натуральных губок, остров Сими, Греция/wikipedia






















































































































Комментарии (0)