454-пиросеквенирование выявляет изменчивое грибное разнообразие в различных системах земледелия
Оазисные системы земледелия широко распространены в некоторых частях мира, особенно на Аравийском полуострове и в ряде стран Африки. В Омане большинство хозяйств ведётся по полуоазисной системе (SOF), предполагающей выращивание нескольких культур — преимущественно для личного потребления, но также и для местного рынка. Целью настоящего исследования было изучение разнообразия грибов методом пиросеквенирования в почвах фермы, использующей систему SOF и занятой под финиковые пальмы, кислые лаймы и огурцы. Полученные данные сравнивались с результатами с органической фермы (OR), где выращивались огурцы и томаты.
Аннотация
Выявлена вариабельность грибного разнообразия в зависимости от культуры в пределах одного хозяйства. Число OTU, оценки богатства по Chao1 и значения индекса Шеннона показали, что почвы под финиковыми пальмами и лаймами характеризуются более высоким разнообразием грибов по сравнению с почвой под огурцами (в системе SOF). Кроме того, эти же показатели свидетельствуют о более высоком уровне грибного разнообразия в ризосфере огурцов на органической ферме по сравнению с SOF. В большинстве проб как с органической, так и с полуоазисной фермы доминировал отдел Ascomycota; среди других преобладающих отделов — Microsporidia, Chytridiomycota и Basidiomycota. Наблюдаемые различия в уровне грибного разнообразия внутри системы SOF, по-видимому, обусловлены различиями в агротехнических практиках, применяемых для каждой из культур.
Введение
Почва является резервуаром тысяч видов грибов и бактерий, которые играют важную роль в естественных и обрабатываемых сельскохозяйственных почвах (Abed и др., 2013; Al-Sadi и др., 2015b; Kaisermann и др., 2015; Tardy и др., 2015). Грибы являются наиболее доминирующими эукариотическими видами с точки зрения биомассы в почве. Грибы играют важную роль как разлагатели, участники круговорота питательных веществ, агрегаторы почвы, патогены и микоризные симбионты (Guo и др., 2015; Thomson и др., 2015; Stott и Taylor, 2016).
Изменения в землепользовании и агротехнических приёмах привели к снижению качества почвы, плодородия и продуктивности (Cherubin и др., 2015; Price и др., 2015). Продуктивность и здоровье почв в некоторой степени зависят от процессов, осуществляемых почвенными микробными сообществами (Guo и др., 2015; Heilmann-Clausen и др., 2015; Stott и Taylor, 2016). Интенсивное применение неорганических удобрений и пестицидов может влиять на почвенные микробные популяции и приводить к снижению микробного разнообразия или изменениям в микробных сообществах (Esmaeili Taheri и др., 2015; Filimon и др., 2015; Pose-Juan и др., 2015; Rangel и др., 2015).
Оазисная система земледелия была распространённой практикой на Аравийском полуострове на протяжении многих тысяч лет. Эта система характеризуется выращиванием нескольких культур, главным образом финиковых пальм, вокруг источника воды. Источник воды, финиковые пальмы и другие культуры служили для создания благоприятной среды для людей, где совместно проживало несколько человек, принадлежащих к одному или разным племенам (Kharusi и Salman, 2015). Эта система трансформировалась в полуоазисную, традиционную систему ведения сельского хозяйства в большинстве районов выращивания на Аравийском полуострове, особенно в Омане, где фермеры продолжают выращивать несколько культур на участках, орошаемых проточной водой или подземными водами, извлекаемыми из скважин (Al-Sadi и др., 2013; Yaish и Kumar, 2015). В наши дни на большинстве ферм Омана выращивают как минимум две или более из следующих культур: финиковые пальмы, кислые лаймы, манго и огурцы (Al-Sadi и др., 2013). На этих фермах обычно используются животные навозы, особенно под финиковые пальмы и цитрусовые (Al-Sadi и др., 2011, 2015b). Однако для овощных культур, главным образом огурцов, вносятся питательные субстраты, компосты и различные неорганические удобрения (Al-Mazroui и Al-Sadi, 2015). Кроме того, заражение огурцов патогенными грибами делает необходимым частое применение системных и контактных фунгицидов (Al-Sadi, 2012). С другой стороны, в некоторых хозяйствах страны наблюдается переход к использованию органического земледелия, однако темпы роста этого сектора очень низкие (Al-Mazroui и Al-Sadi, 2015).
Предыдущие исследования показали, что численность и структура популяций почвенной флоры и фауны могут подвергаться влиянию ряда факторов, включая приёмы возделывания, виды растений, а также внесение органических и неорганических удобрений и пестицидов (DeAngelis и др., 2015; Matsushita и др., 2015; Tardy и др., 2015; Van Geel и др., 2015; Coleman-Derr и др., 2016). Однако в этой части мира имеется мало информации о влиянии систем возделывания на грибное разнообразие. Кроме того, мало известно об уровне грибного разнообразия между различными культурами в пределах одной фермы.
Данное исследование было направлено на изучение грибного разнообразия в полуоазисной и органической системах. Конкретными задачами являлись: (1) исследовать уровень грибного разнообразия в полуоазисной системе земледелия (SOF) и (2) охарактеризовать степень грибного разнообразия по отношению к органической системе и в ризосфере различных культур. Изучение динамики микробных популяций в системах земледелия может помочь предоставить информацию о состоянии здоровья почв в полуоазисных системах и способствовать переходу к более органическим коммерческим системам ведения сельского хозяйства в развивающихся странах.
Материалы и методы
Сбор образцов
В исследовании изучалось грибное разнообразие в полуоазисной системе земледелия в Омане в сравнении с органической системой. Эксперимент был проведён на одной органической и одной полуоазисной ферме. Подробная информация о местоположении, культурах и погодных условиях представлена в Таблице 1.
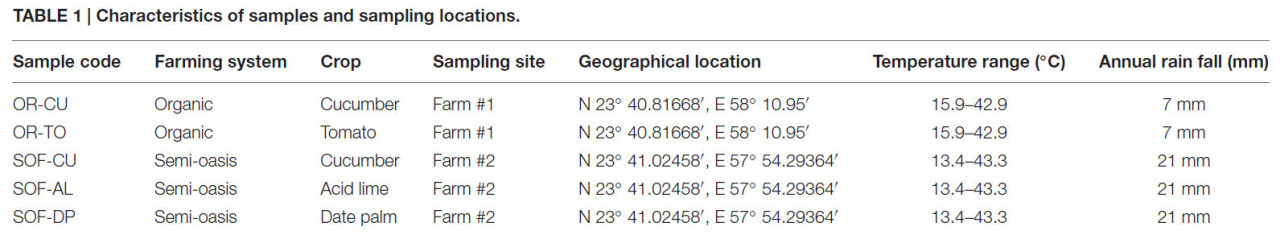
Таблица 1 Характеристика образцов и мест отбора проб.
Образцы почвы были собраны в период с сентября по ноябрь 2013 года из ризосферы огурца и томата на органической ферме, а также из ризосферы огурца, кислого лайма и финиковой пальмы на полуоазисной ферме. Ризосферной почвой в данном исследовании считалась почва в пределах 0–3 см от корней растений. Огурец и томат выращивались на органической ферме как минимум последние 8 лет. Финиковые пальмы и кислые лаймы на полуоазисной ферме имели возраст 9–12 лет. Они удобрялись животным навозом и никогда не подвергались фунгицидной обработке почвы. Огурцы на полуоазисной ферме удобрялись неорганическими удобрениями и животными навозами. Почвы под огурцами на полуоазисной ферме в течение последних 5 лет получали обработки мефеноксамом, гимексазолом и тиофанат-метилом.
Каждый образец почвы объёмом приблизительно 1 кг отбирался из трёх мест в верхнем 5–15 см слое вблизи (0–3 см) активных питающих корней каждой культуры. Образцы почвы отбирались из ризосферы трёх случайно выбранных растений каждой культуры, выращиваемых на разных участках в пределах фермы. Образцы почвы помещались в стерильные пластиковые пакеты и затем транспортировались в Лабораторию фитопатологии Университета Султана Кабуса (Оман). Каждый образец был подвергнут химическому и физическому анализу. Почва для анализа ДНК была измельчена с жидким азотом и затем хранилась при температуре -80°C до выделения ДНК.
Физико-химические свойства почвы
Образцы почвы были высушены на воздухе, измельчены и затем просеяны через сито с размером ячеек 2 мм для удаления корней и растительных остатков. После этого просеянная почва хранилась в пластиковых пробирках до проведения анализа. Для каждого образца почвы были определены различные физико-химические параметры. Текстура почвы определялась с помощью ареометрического метода (Gee и Bauder, 1986). Электропроводность (EC) и pH определялись с использованием измерителей EC и pH (Zhang и др., 2005). Содержание калия (K) измерялось пламенно-фотометрическим методом (пламенный фотометр Sheerwood 450), в то время как концентрация фосфора (P) определялась с использованием масс-спектрометра с индуктивно связанной плазмой (Perkin Elmer, США). Общий неорганический углерод (TIC) и общий органический углерод (TOC) анализировались с использованием анализатора общего органического углерода (TOC-V, Shimadzu, Япония). Анализ азота проводился путём смешивания 0,5 г образца почвы с одной таблеткой катализатора Kjeltab в 10 мл серной кислоты с последующим нагреванием при 420°C в течение 20–30 минут. Затем раствор охлаждали, после чего проводили анализ общего азота с использованием анализатора Kjeltec (FOSS TECATOR, Швеция).
Анализ методом пиросеквенирования
ДНК выделяли из образцов почвы в соответствии с протоколом Volossiouk и др. (1995) с некоторыми модификациями. Почву измельчали с жидким азотом в течение 5 минут, затем 0,05 г каждого образца переносили в пробирку Эппендорфа объёмом 1,5 мкл. Затем добавляли 125 мкл обезжиренного молока и инкубировали смесь в термостате при 65°C в течение 1 часа. Смесь центрифугировали, и супернатант переносили в новую пробирку Эппендорфа. После этого к супернатанту добавляли 500 мкл SDS-экстракционного буфера (0,3% SDS, 140 мM NaCl, 50 мM NaAc, pH 5,1), после чего добавляли один объём смеси фенол:хлороформ:изоамиловый спирт (25:24:1). Суспензию центрифугировали, и супернатант переносили в новую пробирку Эппендорфа. Затем к супернатанту добавляли 0,6 объёма изопропанола и 10 мкл NaAc, инкубировали при -20°C в течение ночи и затем центрифугировали. Осадок промывали 600 мкл этанола, высушивали и наконец ресуспендировали в TE-буфере. Качество и количество ДНК оценивали с использованием спектрофотометра Nano Drop (Thermo Scientific, США).
Выделение ДНК проводилось из трёх повторных образцов, отобранных от каждой культуры. Затем три повторных образца ДНК от каждой культуры были объединены в композитный образец. Для амплификации области ITS2 использовали праймеры ITS1F и ITS2aR. Образцы были отправлены в Исследовательскую и испытательную лабораторию (RTL, Лаббок, Техас, США) для проведения меченого 454-пиросеквенирования (Dowd и др., 2008a,b).
Полученные последовательности длиной менее 300 пар оснований были исключены из дальнейшего анализа, а остальные были проверены на высокое качество с использованием RDP версия 9 (Cole и др., 2009). Концевые участки низкого качества и метки были удалены, и последовательности были проверены на наличие химер с использованием программного обеспечения UCHIME для обнаружения химер (Edgar и др., 2011). Полученные последовательности анализировались с использованием алгоритма BLASTn.NET путём сравнения с высококачественными последовательностями из NCBI, и результаты были валидированы на основе методов таксономического расстояния (Dowd и др., 2005, 2008a,b). Из 1980–14189 необработанных прочтений для семи образцов было получено 1230–11829 прочтений после фильтрации при пороге сходства 97%.
Дальнейший анализ данных пиросеквенирования проводился с использованием программного обеспечения R (R Development Core Team, 2011). Был построен график кривой разрежения, показывающий количество OTU в зависимости от количества последовательностей. Богатство оценивалось с использованием оценщика богатства Chao1 по формуле: Schao1 = Sobs + n1 (n1-1)/2 (n2 + 1), где Sobs — количество наблюдаемых видов/OTU, а ni — количество OTU с численностью i. Индекс Шеннона рассчитывался по формуле H′ = -Σpiln(pi), где Pi — доля филотипов, а i — общее количество всех филотипов в образце. Расстояния UniFrac и Брея–Кёртиса рассчитывались с использованием пакета phyloseq в R. Был проведён анализ главных координат (PCoA) и построены графики на основе взвешенных и невзвешенных расстояний UniFrac и расстояний Брея–Кёртиса. Была построена тепловая карта относительной численности наиболее доминирующих родов грибов, и образцы были отсортированы на основе расстояний Брея–Кёртиса, взвешенных и невзвешенных расстояний UniFrac.
Статистический анализ
Для выявления различий между почвами из разных систем возделывания использовался критерий Стьюдентизированного размаха Тьюки (SAS, SAS Institute Inc., США). Также был проведён корреляционный анализ с использованием SAS.
Результаты
Физико-химические свойства почвы
Почвы из-под различных культур и систем земледелия показали вариабельность физико-химических свойств (Таблица 2). Почвы органической фермы были супесчаными, в то время как почвы с полуоазисной фермы были песчаными и супесчаными. pH большинства почв был нейтральным или щелочным (pH = 7,7–8,4), в то время как электропроводность варьировала от 1 до 10,54 мСм. Уровень неорганического углерода был значительно выше в почвах с полуоазисной фермы по сравнению с органической, в то время как органический углерод был значительно выше в почве из-под финиковых пальм по сравнению со всеми остальными почвами (P < 0,05). Уровни азота, фосфора и калия варьировали в зависимости от системы земледелия и культуры (Таблица 2).
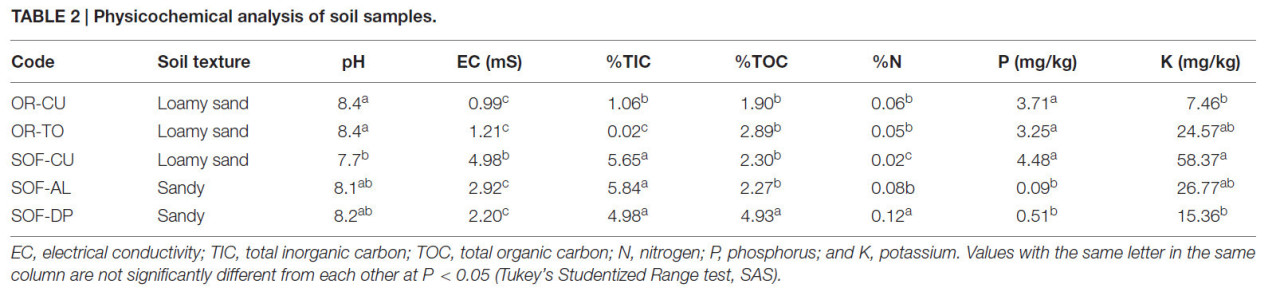
Таблица 2 Физико-химический анализ почвенных образцов.
EC — электропроводность; TIC — общий неорганический углерод; TOC — общий органический углерод; N — азот; P — фосфор; K — калий. Значения с одинаковой буквой в одном столбце достоверно не различаются при P < 0,05 (критерий Стьюдентизированного размаха Тьюки, SAS).
Взаимосвязь между культурами и грибным разнообразием
Анализ грибного разнообразия в полуоазисной системе, где выращиваются финиковая пальма, кислый лайм и огурец, показал наличие дифференцированного разнообразия среди трёх культур. Оценки богатства по Chao1 показали, что в ризосфере трёх культур было обнаружено в общей сложности 23, 19 и 14 OTU соответственно (Рисунки 1 и 2). Оценки индекса Шеннона показали, что наибольшее значение имеет финиковая пальма — 2,50, затем кислый лайм — 2,22 и огурец — 1,25 (Рисунок 2).
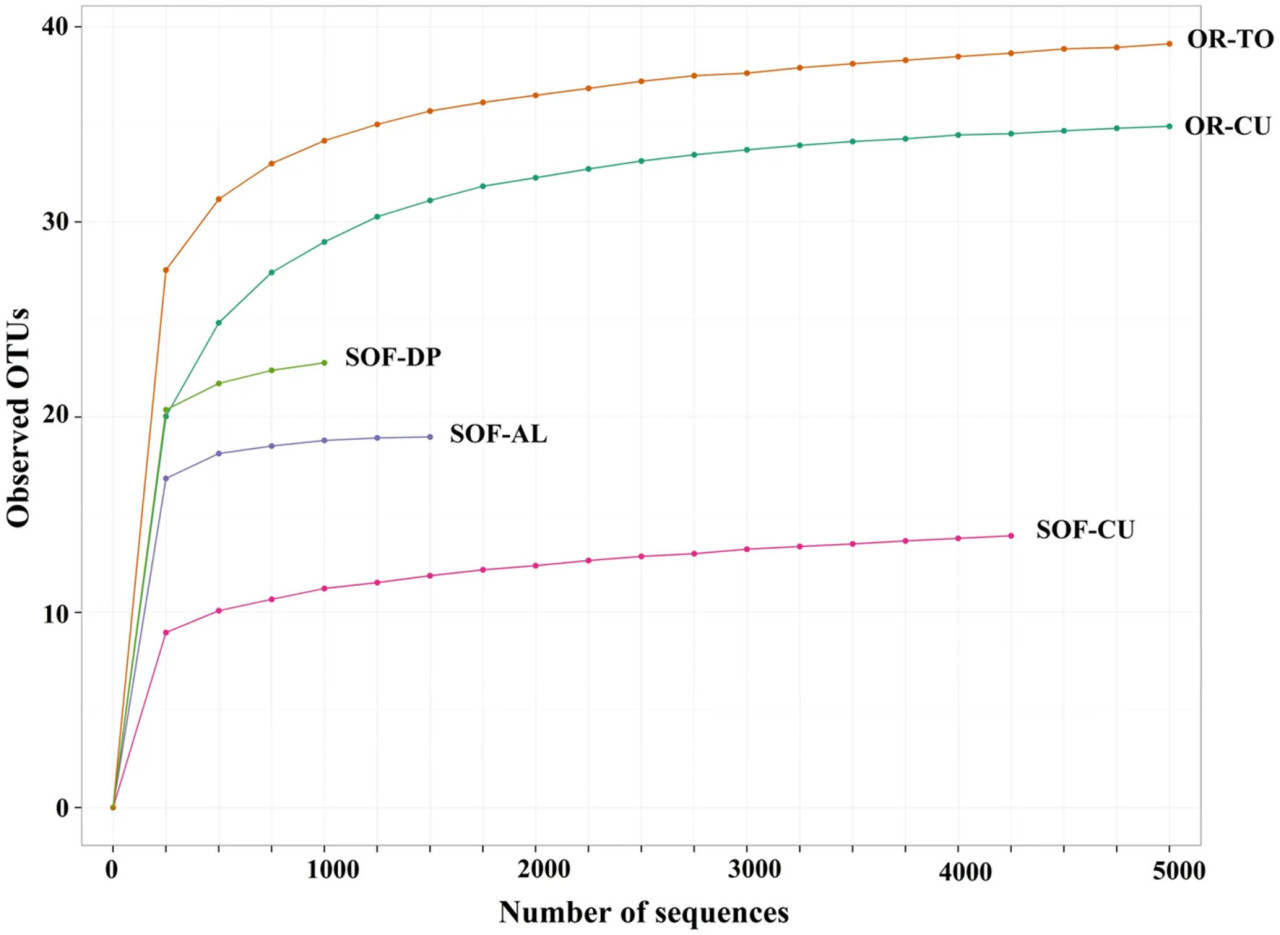
РИСУНОК 1 Кривая разрежения наблюдаемых OTU двух почвенных образцов, полученных от томата (OR-TO) и огурца (OR-CU), выращенных на органической ферме, и трёх почвенных образцов, полученных от огурца (SOF-CU), кислого лайма (SOF-AL) и финиковой пальмы (SOF-DP), выращенных на полуоазисной ферме.
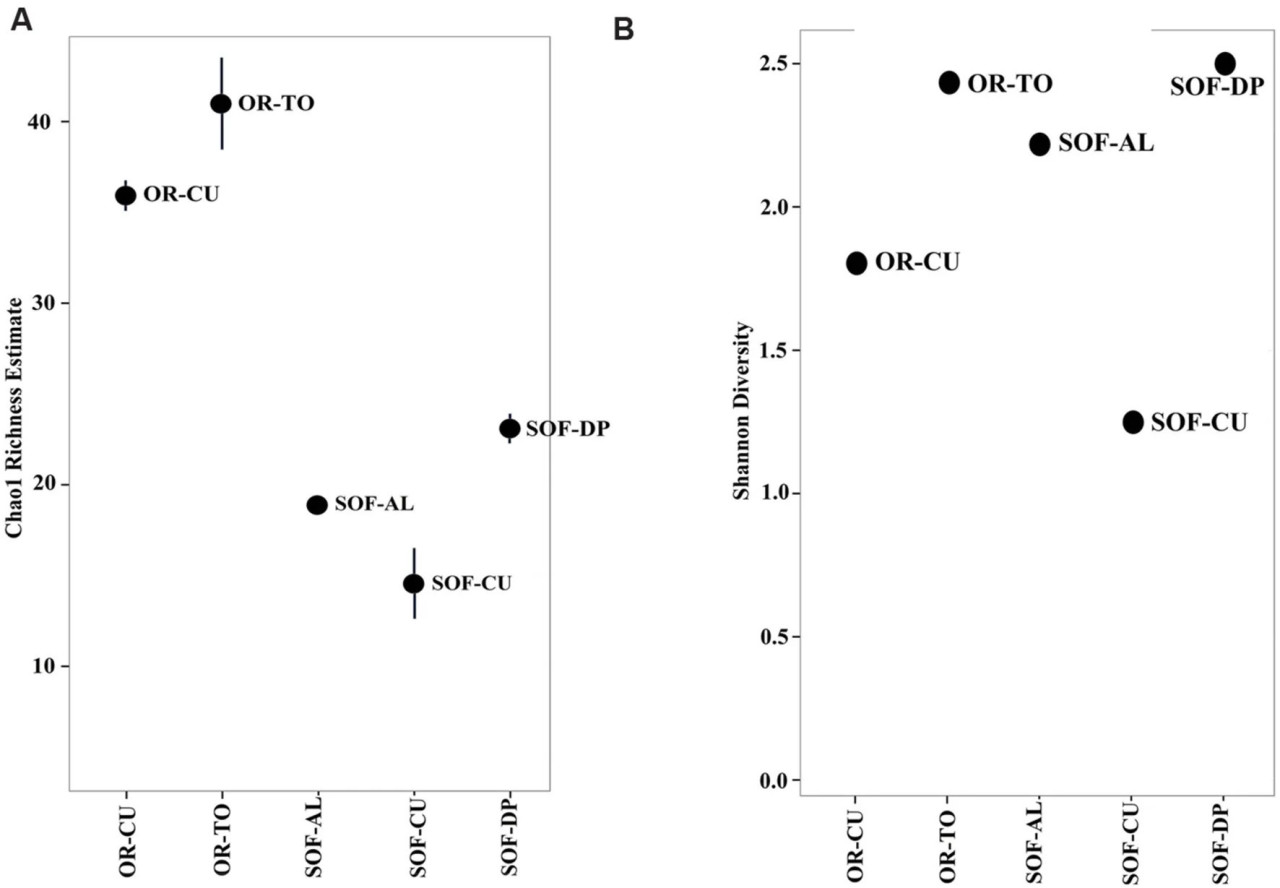
РИСУНОК 2 Богатство по Chao1 (A) и разнообразие по Шеннону (B) в общих данных микробиома пяти почвенных образцов, полученных с органической фермы (OR) и полуоазисной фермы (SOF). Коды образцов описаны в Таблице 1.
Ascomycota был наиболее доминирующим типом, присутствующим в почве из-под трёх культур. Другими доминирующими типами были Microsporidia, Basidiomycota и Chytridiomycota (Рисунок 3). Классы Dothideomycetes, Microsporidetes и Sordariomycetes были наиболее распространёнными в почвах из-под трёх культур (Рисунок 3). Другими распространёнными классами были Eurotiomycetes, Leotiomycetes и Chytridiomycetes. Кислый лайм имел семь общих классов с финиковой пальмой, в то время как огурец имел пять общих классов с финиковой пальмой. Количество общих классов между кислым лаймом и огурцом составило четыре. Cladosporium и Systenostrema были обнаружены в почвах из-под трёх культур, в то время как другие виды грибов были обнаружены в почве из-под одной-двух культур. Systenostrema имел высокую численность в почве из-под огурца (Рисунок 4).
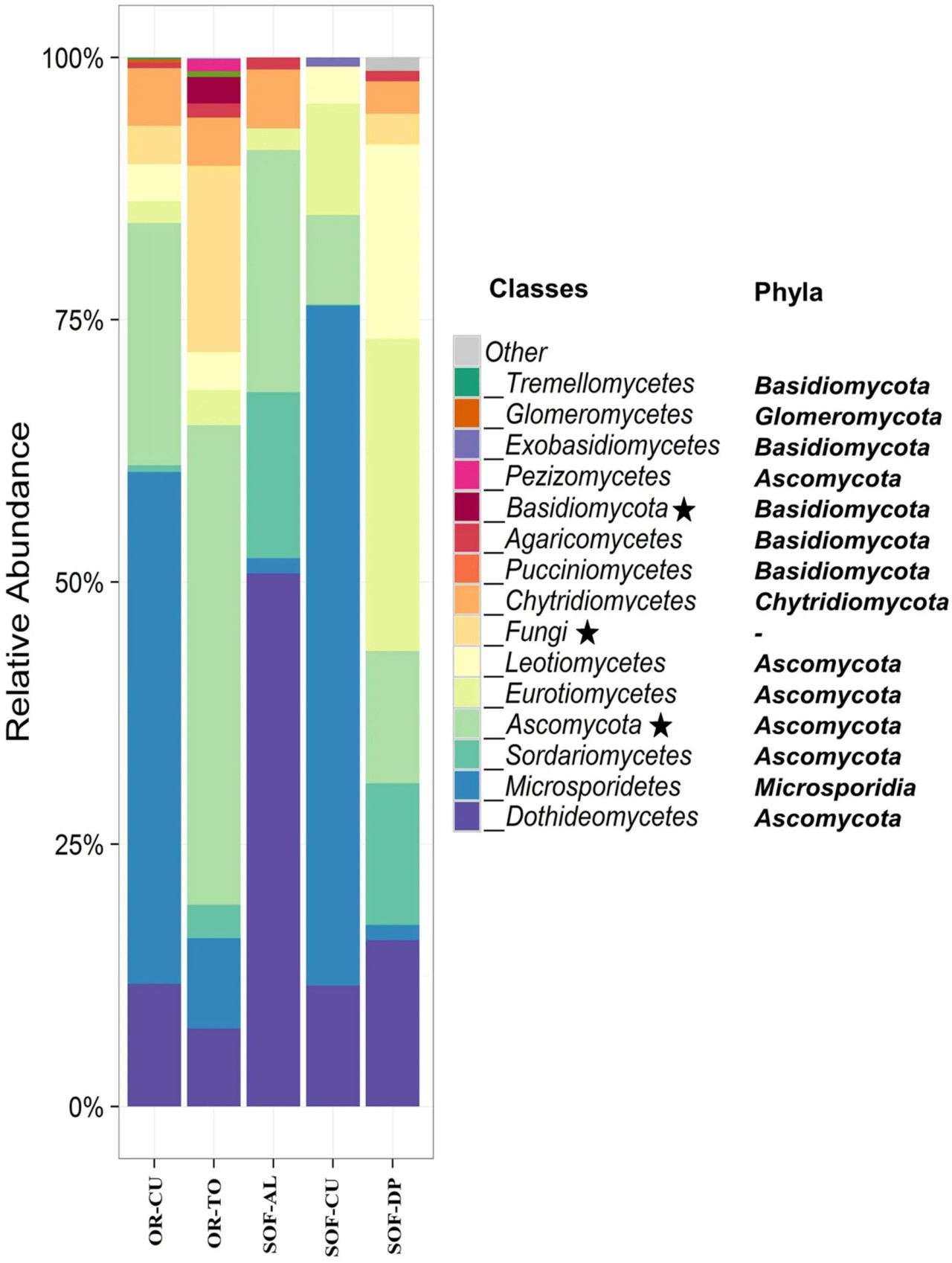
РИСУНОК 3 Относительная численность грибных сообществ на уровне классов в органической (OR) и полуоазисной (SOF) системах земледелия. Звёздочки в легенде указывают на таксономические единицы, которые не удалось разрешить до уровня класса. Типы различных классов указаны на рисунке. Коды образцов описаны в Таблице 1.
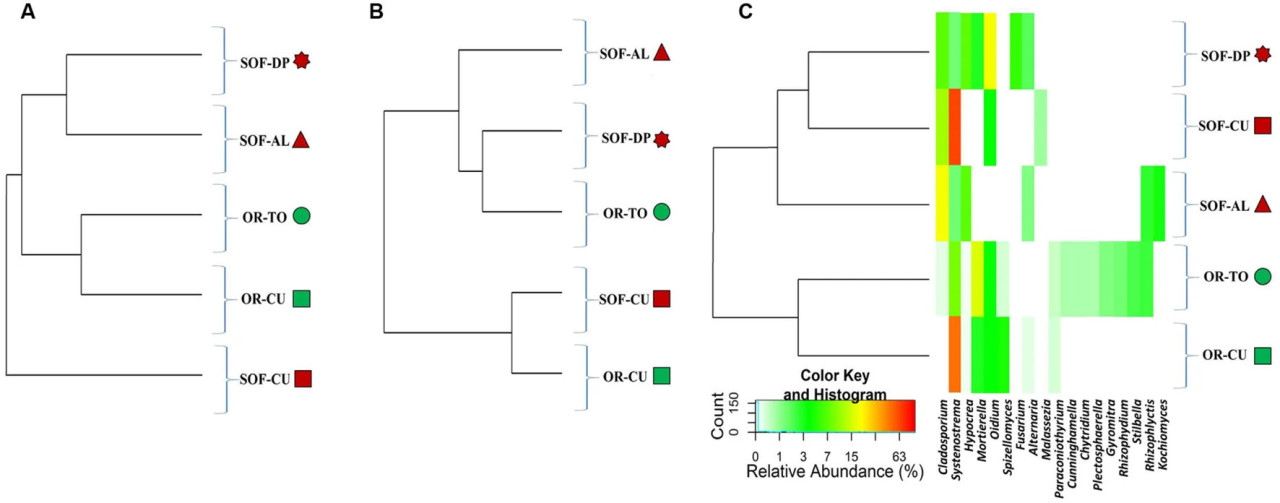
РИСУНОК 4 Тепловая карта относительной численности 18 доминирующих родов грибов в пяти образцах. Образцы отсортированы на основе расстояний Брея–Кёртиса (A), взвешенных расстояний Unifrac (B) и невзвешенных расстояний Unifrac (C). Цветные фигуры рядом с кодами образцов указывают на источник почв (зелёный — органический, тёмно-красный — полуоазисный). Роды грибов описаны в (C), а коды образцов описаны в Таблице 1.
Грибное разнообразие на органической и полуоазисной фермах
Более высокий уровень грибного разнообразия наблюдался в ризосфере огурцов, выращенных на органической ферме, по сравнению с полуоазисной. Значения индекса Шеннона и богатства по Chao1 составляли 1,80 и 36 для почвы из-под огурца на органической ферме по сравнению с 1,25 и 14 для почвы с полуоазисной фермы соответственно (Рисунок 2). Томат, выращенный на органической ферме, также имел относительно высокие значения разнообразия по Шеннону и богатства (Рисунок 2).
Тип Ascomycota был наиболее доминирующим типом в большинстве почвенных образцов из-под огурца на органической и полуоазисной фермах, а также из-под органического томата. Другие типы включали Microsporidia, Chytridiomycota и Basidiomycota. Наши результаты показали, что Microsporidetes был основным классом в огурце, выращенном на органической и полуоазисной фермах (Рисунок 3).
Анализ грибного разнообразия в почвах органической и полуоазисной ферм с использованием невзвешенных расстояний UniFrac показал разделение органических систем от полуоазисных (Рисунки 4 и 5). С другой стороны, взвешенный UniFrac разделил огурец, выращенный на органической и полуоазисной фермах, от других культур, в то время как Брей–Кёртис также отделил органические огурец и томат от других, с близкой связью с SOF-DP/AL (Рисунки 4 и 5). Дальнейшие статистические анализы не могли быть проведены из-за использования одного технического и одного биологического повтора для каждого образца. Корреляции между оценками разнообразия и физико-химическими свойствами почвы обнаружено не было (P > 0,05).
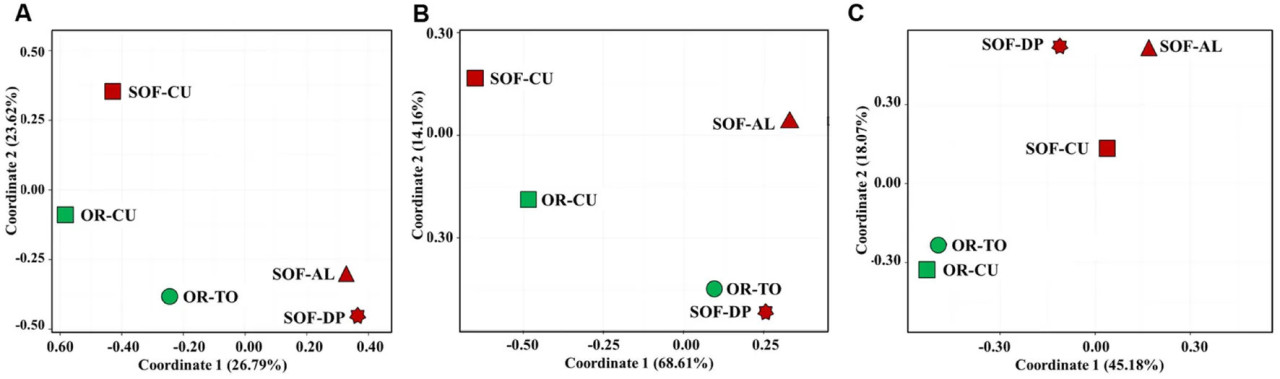
РИСУНОК 5 Анализ главных координат грибного разнообразия в пяти почвенных образцах на основе расстояний Брея–Кёртиса (A), взвешенных расстояний Unifrac (B) и невзвешенных расстояний Unifrac (C). Зелёный и тёмно-красный цвета указывают на органическую и полуоазисную системы земледелия соответственно. Коды образцов описаны в Таблице 1.
Обсуждение
Было установлено, что полуоазисная система земледелия в Омане характеризуется дифференцированным уровнем грибного разнообразия. Уровень грибного разнообразия был выше в почвах из-под финиковых пальм и цитрусовых по сравнению с почвами из-под огурцов. Это первое исследование на основе 454-пиросеквенирования, которое было направлено на изучение грибного разнообразия в полуоазисной системе. Более низкий уровень грибного разнообразия в ризосфере огурца может быть связан с применением фунгицидов на этой культуре. Огурцы сильно поражаются увяданием и другими корневыми заболеваниями, что делает необходимым частое применение мефеноксама, тиофанат-метила, гимексазола и других фунгицидов (Al-Sadi, 2012; Al-Sadi и др., 2012, 2015a). Поэтому вероятно, что применение фунгицидов могло привести к снижению грибных популяций в ризосфере огурцов. С другой стороны, применение фунгицидов для финиковых пальм и кислого лайма в Омане минимально или вообще отсутствует на многих фермах из-за ограниченного количества грибных заболеваний, поражающих эти две культуры, особенно финиковые пальмы. Кроме того, обработка почвы, которая, как сообщалось, снижает грибное разнообразие (Tardy и др., 2015), часто практикуется для почв под огурцами, что не характерно для финиковых пальм и кислого лайма.
Предыдущие исследования показали, что микробное разнообразие значительно увеличивается на фермах, которые используют органический навоз, по сравнению с химическим удобрением (Qiu и др., 2012; Zhang и др., 2012). Известно, что органические навозы характеризуются высоким грибным разнообразием (Al-Mazroui и Al-Sadi, 2015; Al-Sadi и др., 2015b). Применение животных навозов вместо минеральных удобрений для финиковых пальм и кислого лайма могло способствовать повышению разнообразия грибов в почве из-под этих культур. Также очевидно, что тип выращиваемых культур повлиял на уровень грибного разнообразия и на взаимодействие между этими культурами и грибами (Badri и Vivanco, 2009; Sugiyama и др., 2010).
Количество OTU, разнообразие по Шеннону и оценка Chao1 показали, что уровень грибного разнообразия в почве из-под органически выращенного огурца был выше по сравнению с огурцом, выращенным с использованием неорганических удобрений. Это подтверждает результаты предыдущих исследований о более высоком грибном разнообразии в органических почвах (Sugiyama и др., 2010). Это может быть частично связано с использованием органических навозов, а также с тем, что химические фунгициды не применяются в почвах на органических фермах. Кроме того, органические компосты, которые часто вносятся на эти фермы, обычно обрабатываются некоторыми видами грибов для целей биоконтроля, что может объяснять более высокий уровень грибного разнообразия на этих фермах (Al-Sadi и др., 2015b; Kohler и др., 2015; On и др., 2015).
Анализ грибного разнообразия с использованием невзвешенного UniFrac, взвешенного UniFrac и Брея–Кёртиса показал кластеризацию различных почвенных образцов в разные группы под влиянием либо систем земледелия, либо видов растений. Невзвешенный анализ UniFrac разделил почвы в зависимости от систем земледелия, то есть различные системы земледелия обусловили качественное различие в грибном разнообразии. Однако взвешенный UniFrac показал кластеризацию на основе видов растений, что свидетельствует о наличии некоторых доминирующих таксонов в огурце, которым, по-видимому, является Systenostrema (Microsporidetes). Брей–Кёртис не учитывает таксономическое родство между OTU, в то время как UniFrac учитывает. Это обстоятельство привело к более тесной кластеризации между OR-TO с OR-CU, SOF-AL и SOF-DP, но не повлияло на сходство между SOF-CU и OR-CU (Рисунок 5A). Это показывает, что OTU, которые обусловили различие между OR-TO/CU и SOF-AL/DP, обнаруженное с помощью Брея–Кёртиса, каким-то образом филогенетически связаны. Кроме того, общая дисперсия, объясняемая координатами 1 и 2, выше во взвешенном UniFrac (~83%) по сравнению с невзвешенным (~63%) и Бреем–Кёртисом (~50%). Это может свидетельствовать о том, что виды растений и/или связанные с ними агротехнические приёмы определяют структуру грибного сообщества, хотя системы земледелия оказывают сильное влияние на качественную структуру. В другом исследовании на видах Agave Coleman-Derr и др. (2016) предположили, что биогеография вида-хозяина является основным детерминантом грибного микробиома.
Тип Ascomycota был наиболее доминирующим типом в большинстве образцов с органической и полуоазисной ферм. Этот тип широко распространён в различных почвах по всему миру (Qiu и др., 2012; Abed и др., 2013; Al-Sadi и др., 2015b). Это главным образом связано с тем, что Ascomycota содержит несколько пигментированных видов, которые обычно лучше переносят более высокие температуры, и потому что многие виды этого типа продуцируют обильные споры (Abed и др., 2013). Типы Microsporidia, Chytridiomycota и Basidiomycota также были распространены в некоторых почвах ферм. Различия в видовом составе между различными почвами могут быть связаны с различными системами выращивания, агротехническими приёмами и различными культурами.
Физико-химический анализ почв показал вариации pH почвы, содержания углерода и питательных веществ от одной почвы к другой. Уровни органического углерода были одинаковыми или более низкими в органических почвах по сравнению с почвами полуоазисной системы. Предыдущие исследования показали, что органические системы не всегда увеличивают содержание органического вещества в почве по сравнению с традиционными системами (Li и др., 2012). Несколько более низкий pH почвы в полуоазисной системе по сравнению с органической согласуется с предыдущими исследованиями (Li и др., 2012). На pH почвы и содержание питательных веществ обычно влияют растительность, тип почвы, углерод и азот (Barak и др., 1997; Kuramae и др., 2012).
Заключение
Наше исследование показало наличие дифференцированных уровней грибного разнообразия, которые, предположительно, связаны с системами земледелия, видами растений или агротехническими приёмами. Полуоазисная система, по-видимому, характеризуется высоким уровнем грибного разнообразия, особенно под культурами, которые выращиваются полуорганическим способом. Ожидается, что это будет отражаться на здоровье и качестве почвы, особенно если грибные сообщества представлены видами, стимулирующими рост растений или подавляющими болезни. Тенденция к производству огурцов в этих системах, по-видимому, оказывает влияние на уровень грибного разнообразия, возможно, из-за интенсивного применения фунгицидов. Текущий переход к использованию пестицидов в полуоазисных системах должен учитывать негативное влияние химических веществ на разнообразие полезных грибов.
Ссылки
1. Abed R. M. M., Al-Sadi A. M., Al-Shihi M. A., Al-Hinai S., Robinson M. D. (2013). Diversity of free-living and lichenized fungal communities in biological desert crusts of the Sultanate of Oman and their role in improving soil properties. Soil Biol. Biochem. 57, 695–705. doi: 10.1016/j.soilbio.2012.07.023. CrossRef. Google Scholar.
2. Al-Mazroui S. S., Al-Sadi A. M. (2015). Highly variable fungal diversity and the occurrence of potentially plant pathogenic fungi in potting media, organic fertilizers and composts originating from 14 countries. J. Plant Pathol. 97, 529–534. doi: 10.4454/JPP.V97I3.033. CrossRef. Google Scholar.
3. Al-Sadi A. M. (2012). Efficacy of mefenoxam is affected by a lag period between application and inactivation of Pythium species. Phytopathol. Mediterran. 51, 292–297. Google Scholar.
4. Al-Sadi A. M., Al-Ghaithi A. G., Al-Balushi Z. M., Al-Jabri A. H. (2012). Analysis of diversity in Pythium aphanidermatum populations from a single greenhouse reveals phenotypic and genotypic changes over 2006 to 2011. Plant Dis. 96, 852–858. doi: 10.1094/PDIS-07-11-0624. CrossRef. Google Scholar.
5. Al-Sadi A. M., Al-Masoodi R. S., Al-Ismaili M., Al-Mahmooli I. H. (2015a). Population structure and development of resistance to hymexazol among Fusarium solani populations from date palm, Citrus and cucumber. J. Phytopathol. 163, 947–955. doi: 10.1111/jph.12397. CrossRef. Google Scholar.
6. Al-Sadi A. M., Al-Mazroui S. S., Phillips A. (2015b). Evaluation of culture-based techniques and 454 pyrosequencing for the analysis of fungal diversity in potting media and organic fertilizers. J. Appl. Microbiol. 119, 500–509. doi: 10.1111/jam.12854. CrossRef. Google Scholar.
7. Al-Sadi A. M., Al-Said F. A., Al-Jabri A. H., Al-Mahmooli I. H., Al-Hinai A. H., de Cock A. W. A. M. (2011). Occurrence and characterization of fungi and oomycetes transmitted via potting mixtures and organic manures. Crop Prot. 30, 38–44. doi: 10.1016/j.cropro.2010.09.015. CrossRef. Google Scholar.
8. Al-Sadi A. M., Al-Wehaibi A. N., Al-Shariqi R. M., Al-Hammadi M. S., Al-Hosni I. A., Al-Mahmooli I. H., et al. (2013). Population genetic analysis reveals diversity in Lasiodiplodia species infecting date palm, Citrus, and mango in Oman and the UAE. Plant Dis. 97, 1363–1369. doi: 10.1094/PDIS-03-13-0245-RE. CrossRef. Google Scholar.
9. Badri D. V., Vivanco J. M. (2009). Regulation and function of root exudates. Plant Cell Environ. 32, 666–681. doi: 10.1111/j.1365-3040.2009.01926.x. CrossRef. Google Scholar.
10. Barak P., Jobe B. O., Krueger A. R., Peterson L. A., Laird D. A. (1997). Effects of long-term soil acidification due to nitrogen fertilizer inputs in Wisconsin. Plant Soil 197, 61–69. doi: 10.1023/a:1004297607070. CrossRef. Google Scholar.
11. Cherubin M. R., Franco A. L. C., Cerri C. E. P., Oliveira D. M. S., Davies C. A., Cerri C. C. (2015). Sugarcane expansion in Brazilian tropical soils-Effects of land use change on soil chemical attributes. Agricult. Ecosyst. Environ. 211, 173–184. doi: 10.1016/j.agee.2015.06.006. CrossRef. Google Scholar.
12. Cole J. R., Wang Q., Cardenas E., Fish J., Chai B., Farris R. J., et al. (2009). The Ribosomal Database Project: improved alignments and new tools for rRNA analysis. Nucleic Acids Res. 37, D141–D145. doi: 10.1093/nar/gkn879. CrossRef. Google Scholar.
13. Coleman-Derr D., Desgarennes D., Fonseca-Garcia C., Gross S., Clingenpeel S., Woyke T., et al. (2016). Plant compartment and biogeography affect microbiome composition in cultivated and native Agave species. New Phytol. 209, 798–811. doi: 10.1111/nph.13697. CrossRef. Google Scholar.
14. DeAngelis K. M., Pold G., Topçuoglu B. D., van Diepen L. T. A., Varney R. M., Blanchard J. L., et al. (2015). Long-term forest soil warming alters microbial communities in temperate forest soils. Front. Microbiol. 6:104. doi: 10.3389/fmicb.2015.00104. CrossRef. Google Scholar.
15. Dowd S. E., Callaway T. R., Wolcott R. D., Sun Y., McKeehan T., Hagevoort R. G., et al. (2008a). Evaluation of the bacterial diversity in the feces of cattle using 16S rDNA bacterial tag-encoded FLX amplicon pyrosequencing (bTEFAP). BMC Microbiol. 8:125. doi: 10.1186/1471-2180-8-125. CrossRef. Google Scholar.
16. Dowd S. E., Sun Y., Wolcott R. D., Domingo A., Carroll J. A. (2008b). Bacterial tag encoded FLX amplicon pyrosequencing (bTEFAP) for microbiome studies:bacterial diversity in the ileum of newly weaned Salmonella-infected pigs. Foodborne Pathog. Dis. 5, 459–472. doi: 10.1089/fpd.2008.0107. CrossRef. Google Scholar.
17. Dowd S. E., Zaragoza J., Rodriguez J. R., Oliver M. J., Payton P. R. (2005). Windows.NET network distributed basic local alignment search toolkit (W.ND-BLAST). BMC Bioinformat. 6:93. doi: 10.1186/1471-2105-6-93. CrossRef. Google Scholar.
18. Edgar R. C., Haas B. J., Clemente J. C., Quince C., Knight R. (2011). UCHIME improves sensitivity and speed of chimera detection. Oxford J. Bioinformat. 27, 2194–2200. doi: 10.1093/bioinformatics/btr381. CrossRef. Google Scholar.
19. Esmaeili Taheri A., Hamel C., Gan Y. (2015). Pyrosequencing reveals the impact of foliar fungicide application to chickpea on root fungal communities of durum wheat in subsequent year. Fungal Ecol. 15, 73–81. doi: 10.1016/j.funeco.2015.03.005. CrossRef. Google Scholar.
20. Filimon M. N., Voia S. O., Popescu R., Dumitrescu G., Ciochina L. P., Mituletu M., et al. (2015). The effect of some insecticides on soil microorganisms based on enzymatic and bacteriological analyses. Roman. Biotechnolog. Lett. 20, 10439–10447. Google Scholar.
21. Gee G. W., Bauder J. W. (1986). "Particle size analysis," in Methods of Soil Analysis, Part-I, Physical and Mineralogical Methods, ed. Klute A. (Madison, WI: American Society of Agronomy), 383–411. Google Scholar.
22. Guo X., Petermann J. S., Schittko C., Wurst S. (2015). Independent role of belowground organisms and plant cultivar diversity in legume-grass communities. Appl. Soil Ecol. 95, 1–8. doi: 10.1016/j.apsoil.2015.05.010. CrossRef. Google Scholar.
23. Heilmann-Clausen J., Barron E. S., Boddy L., Dahlberg A., Griffith G. W., Nordén J., et al. (2015). A fungal perspective on conservation biology. Conservation Biol. 29, 61–68. doi: 10.1111/cobi.12388. CrossRef. Google Scholar.
24. Kaisermann A., Maron P. A., Beaumelle L., Lata J. C. (2015). Fungal communities are more sensitive indicators to non-extreme soil moisture variations than bacterial communities. Appl. Soil Ecol. 86, 158–164. doi: 10.1016/j.apsoil.2014.10.009. CrossRef. Google Scholar.
25. Kharusi N. S., Salman A. (2015). In search of water: hydrological terms in Oman's toponyms. Names 63, 16–29. doi: 10.1179/0027773814Z.00000000094. CrossRef. Google Scholar.
26. Kohler J., Caravaca F., Azcón R., Díaz G., Roldán A. (2015). The combination of compost addition and arbuscular mycorrhizal inoculation produced positive and synergistic effects on the phytomanagement of a semiarid mine tailing. Sci. Total Environ. 514, 42–48. doi: 10.1016/j.scitotenv.2015.01.085. CrossRef. Google Scholar.
27. Kuramae E. E., Yergeau E., Wong L. C., Pijl A. S., Van Veen J. A., Kowalchuk G. A. (2012). Soil characteristics more strongly influence soil bacterial communities than land-use type. FEMS Microbiol. Ecol. 79, 12–24. doi: 10.1111/j.1574-6941.2011.01192.x. CrossRef. Google Scholar.
28. Li R., Khafipour E., Krause D. O., Entz M. H., de Kievit T. R., Fernando W. G. D. (2012). Pyrosequencing reveals the influence of organic and conventional farming systems on bacterial communities. PLoS ONE 7:e51897. doi: 10.1371/journal.pone.0051897. CrossRef. Google Scholar.
29. Matsushita Y., Bao Z., Kurose D., Okada H., Takemoto S., Sawada A., et al. (2015). Community structure, diversity, and species dominance of bacteria, fungi, and nematodes from naturally and conventionally farmed soil: a case study on Japanese apple orchards. Organ. Agricult. 5, 11–28. doi: 10.1007/s13165-015-0096-4. CrossRef. Google Scholar.
30. On A., Wong F., Ko Q., Tweddell R. J., Antoun H., Avis T. J. (2015). Antifungal effects of compost tea microorganisms on tomato pathogens. Biol. Cont. 80, 63–69. doi: 10.1016/j.biocontrol.2014.09.017. CrossRef. Google Scholar.
31. Pose-Juan E., Sánchez-Martín M. J., Herrero-Hernández E., Rodríguez-Cruz M. S. (2015). Application of mesotrione at different doses in an amended soil: dissipation and effect on the soil microbial biomass and activity. Sci. Total Environ. 536, 31–38. doi: 10.1016/j.scitotenv.2015.07.039. CrossRef. Google Scholar.
32. Price G. W., Astatkie T., Gillis J. D., Liu K. (2015). Long-term influences on nitrogen dynamics and pH in an acidic sandy soil after single and multi-year applications of alkaline treated biosolids. Agricult. Ecosys. Environ. 208, 1–11. doi: 10.1016/j.agee.2015.04.010. CrossRef. Google Scholar.
33. Qiu M., Zhang R., Xue C., Zhang S., Li S., Zhang N., et al. (2012). Application of bio-organic fertilizer can control Fusarium wilt of cucumber plants by regulating microbial community of rhizosphere soil. Biol. Fertil. Soils 48, 807–816. doi: 10.1002/jsfa.5653. CrossRef. Google Scholar.
34. R Development Core Team (2011). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. Google Scholar.
35. Rangel D. E. N., Alder-Rangel A., Dadachova E., Finlay R. D., Kupiec M., Dijksterhuis J., et al. (2015). Fungal stress biology: a preface to the fungal stress responses special edition. Curr. Genet. 61, 231–238. doi: 10.1007/s00294-015-0500-3. CrossRef. Google Scholar.
36. Stott M. B., Taylor M. W. (2016). Microbial ecology research in New Zealand. N. Z. J. Ecol. 40, 12–28. Google Scholar.
37. Sugiyama A., Vivanco J. M., Jayanty S. S., Manter D. K. (2010). Pyrosequencing assessment of soil microbial communities in organic and conventional potato farms. Plant Dis. 94, 1329–1335. doi: 10.1094/PDIS-02-10-0090. CrossRef. Google Scholar.
38. Tardy V., Spor A., Mathieu O., Lévèque J., Terrat S., Plassart P., et al. (2015). Shifts in microbial diversity through land use intensity as drivers of carbon mineralization in soil. Soil Biol. Biochemist. 90, 204–213. doi: 10.1016/j.soilbio.2015.08.010. CrossRef. Google Scholar.
39. Thomson B. C., Tisserant E., Plassart P., Uroz S., Griffiths R. I., Hannula S. E., et al. (2015). Soil conditions and land use intensification effects on soil microbial communities across a range of European field sites. Soil Biol. Biochemist. 88, 403–413. doi: 10.1016/j.soilbio.2015.06.012. CrossRef. Google Scholar.
40. Van Geel M., Ceustermans A., Van Hemelrijck W., Lievens B., Honnay O. (2015). Decrease in diversity and changes in community composition of arbuscular mycorrhizal fungi in roots of apple trees with increasing orchard management intensity across a regional scale. Mol. Ecol. 24, 941–952. doi: 10.1111/mec.13079. CrossRef. Google Scholar.
41. Volossiouk T., Robb E. J., Nazar R. N. (1995). Direct DNA extraction for PCR-mediated assays of soil organisms. Appl. Environ. Microbiol. 61, 3972–3976. Google Scholar.
42. Yaish M. W., Kumar P. P. (2015). Salt tolerance research in date palm tree (Phoenix dactylifera L.), past, present, and future perspectives. Front. Plant Sci. 6:348. doi: 10.3389/fpls.2015.00348. CrossRef. Google Scholar.
43. Zhang H., Schroder L. L., Pittman J. J., Wang J. J., Payton M. E. (2005). Soil salinity using saturated paste and 1;1 soil and water extracts. Soil Sci. Soc. Am. J. 69, 1146–1151. doi: 10.2136/sssaj2004.0267. CrossRef. Google Scholar.
44. Zhang Q. C., Shamsi I. H., Xu D. T., Wang G. H., Lin X. Y., Jilani G., et al. (2012). Chemical fertilizer and organic manure inputs in soil exhibit a vice versa pattern of microbial community structure. Appl. Soil Ecol. 57, 1–8. doi: 10.1016/j.apsoil.2012.02.012. CrossRef. Google Scholar.

Kazeeroni EA and Al-Sadi AM (2016) 454-Pyrosequencing Reveals Variable Fungal Diversity Across Farming Systems. Front. Plant Sci. 7:314. doi: 10.3389/fpls.2016.00314
Перевод статьи «454-Pyrosequencing Reveals Variable Fungal Diversity Across Farming Systems» авторов Kazeeroni EA and Al-Sadi AM., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык
Фото: Designed by Freepik
















































































































Комментарии (0)