Аспекты безопасности продукции, получаемой с использованием растений-продуцентов рекомбинантных белков
В данном обзоре рассматриваются наиболее релевантные хост-системы с точки зрения их токсикологического профиля, включая наличие вторичных метаболитов, а также риски, возникающие из-за сохранения этих веществ после этапов очистки и выделения готового продукта
Аннотация
Получение рекомбинантных белков в растениях продвигается с 1990-х годов как быстрая, экономически эффективная и (что самое главное) безопасная альтернатива культивированию бактерий или клеток животных для производства биологических фармацевтических препаратов. Многочисленные виды растений были исследованы на предмет производства широкого спектра кандидатов в лекарственные средства на основе белков. Присущая этим продуктам безопасность часто подчеркивается как преимущество данного подхода, поскольку растительные вирусы не реплицируются в организме человека, и наоборот. Однако при рассмотрении других патогенов требуется более детальный анализ этого принципа, поскольку токсичные соединения представляют риск даже в отсутствие репликации. Аналогичным образом необходимо оценивать риски, связанные с растением-хозяином (например, наличие токсичных вторичных метаболитов) и подходом к производству (например, транзиентная экспрессия на основе бактериальной инфильтрации значительно увеличивает нагрузку эндотоксинов). В данном обзоре рассматриваются наиболее релевантные хост-системы с точки зрения их токсикологического профиля, включая наличие вторичных метаболитов, а также риски, возникающие из-за сохранения этих веществ после этапов очистки и выделения готового продукта. Аналогично мы обсуждаем ряд растительных патогенов и переносчиков заболеваний, которые могут влиять на безопасность продукта, например, из-за высвобождения токсинов. Также рассматривается способность технологических стадий очистки удалять загрязнители и связанные с процессом токсичные примеси, такие как эндотоксины. Данный обзор продукции, полученной на растительной основе, с акцентом на аспекты безопасности, содержит рекомендации, которые позволят заинтересованным сторонам выбрать наиболее подходящие стратегии для разработки технологического процесса.
1 Введение
Растения и растительные клетки могут использоваться для производства активных фармацевтических ингредиентов, включая кандидаты в лекарственные препараты на основе низкомолекулярных соединений и рекомбинантные белки (Eidenberger и др., 2023). Хотя рекомбинантные белки могут продуцироваться во многих различных хост-системах, посттрансляционные модификации, осуществляемые растениями (особенно гликозилирование), могут обеспечивать превосходную активность продукта (Tekoah и др., 2013; Gengenbach и др., 2019), или они могут быть гуманизированы с использованием современных инструментов генетической инженерии (Strasser и др., 2008; Jansing и др., 2018). Те же инструменты могут использоваться для модификации видов растений-хозяев, таких как табак (Nicotiana tabacum) (Menary и др., 2020a), превращая их в хозяев-продуцентов, оптимизированных для производства биологических препаратов (Fraser и др., 2020; Buyel и др., 2021; Huang и Puchta, 2021; Uranga и др., 2021). Одним из примеров такого подхода является модификация метаболизма табака для устранения биосинтеза никотина (Schachtsiek и Stehle, 2019). Производственная стратегия может быть адаптирована для приоритизации скорости (транзиентная экспрессия) или масштабируемости (трансгенные растения) в соответствии с требованиями к конкретным продуктам и рыночными ожиданиями (Buyel и др., 2017; Tusé и др., 2020). После выбора идеального хозяина и производственной стратегии могут быть подобраны платформенные технологии нисходящей обработки для обеспечения высокой чистоты продукта (Buyel и др., 2015a; Ma и др., 2015), включая соблюдение надлежащей производственной практики даже при использовании базовых сооружений для культивирования, таких как теплицы (Ma и др., 2015; Ward и др., 2021). Количество специализированных этапов удаления вирусов в процессах PMF часто ниже по сравнению с процессами на основе клеток млекопитающих, поскольку растительные клетки не поддерживают репликацию вирусов человека (Commandeur и Twyman, 2005; Ma и др., 2015).
Эти принципы свидетельствуют, что растения и растительные клетки могли бы широко использоваться для производства безопасных биологических препаратов в соответствии с нормативными требованиями и производственными стандартами (Hundleby и др., 2022). Тем не менее, на сегодняшний день одобрено лишь небольшое количество продуктов PMF, и с учетом разнообразия задействованных производственных платформ каждый из них может рассматриваться как уникальный. Напротив, микробные и животные клетки использовались для производства многих различных одобренных рекомбинантных биологических белков (Walsh и Walsh, 2022). Поэтому важно выявить ключевые факторы для разработки экономически эффективных, масштабируемых, устойчивых и, особенно, безопасных производственных процессов на растительной основе для биологических белков, что в конечном итоге позволит промышленности внедрить эту технологию без каких-либо ограничений (Menary и др., 2020b).
В данном обзоре рассматриваются аспекты безопасности PMF, охватывающие широкий спектр видов растений (хозяев), процессов и продуктов (Рисунок 1) и связанных с ними рисков (Таблица 1). Сначала мы рассматриваем влияние выбора хозяина, который определяет, необходимо ли учитывать наличие токсичных метаболитов и белков. Далее мы рассматриваем производственные процессы, включая условия культивирования растений, стратегии экспрессии и операции очистки. Затем мы переходим к продукту и его модификации в растении, что связывает нас с выбором хозяина. В заключении мы оцениваем потенциал селекции и генетической инженерии для решения некоторых ключевых проблем безопасности. Данная статья не рассматривает экологическую или производственную безопасность PMF (Knödler и др., 2023a), например, высвобождение трансгенной пыльцы в окружающую среду, что обсуждалось в других работах (Commandeur и Twyman, 2005). Хотя основное внимание в данном обзоре уделяется биологическим препаратам, аналогичные соображения применимы к таким продуктам, как пищевые и кормовые добавки, хотя с различиями в способе производства и использования. Например, пищевые и кормовые добавки обычно производятся в большем масштабе, чем фармацевтические препараты, и должны сохранять функциональность после пероральной доставки (Barzee и др., 2022), тогда как фармацевтические препараты могут быть разработаны для многих различных способов доставки, включая пероральный, внутривенный и внутримышечный. Сходства между фармацевтическими PMF и не фармацевтическими применениями выделяются там, где это уместно.
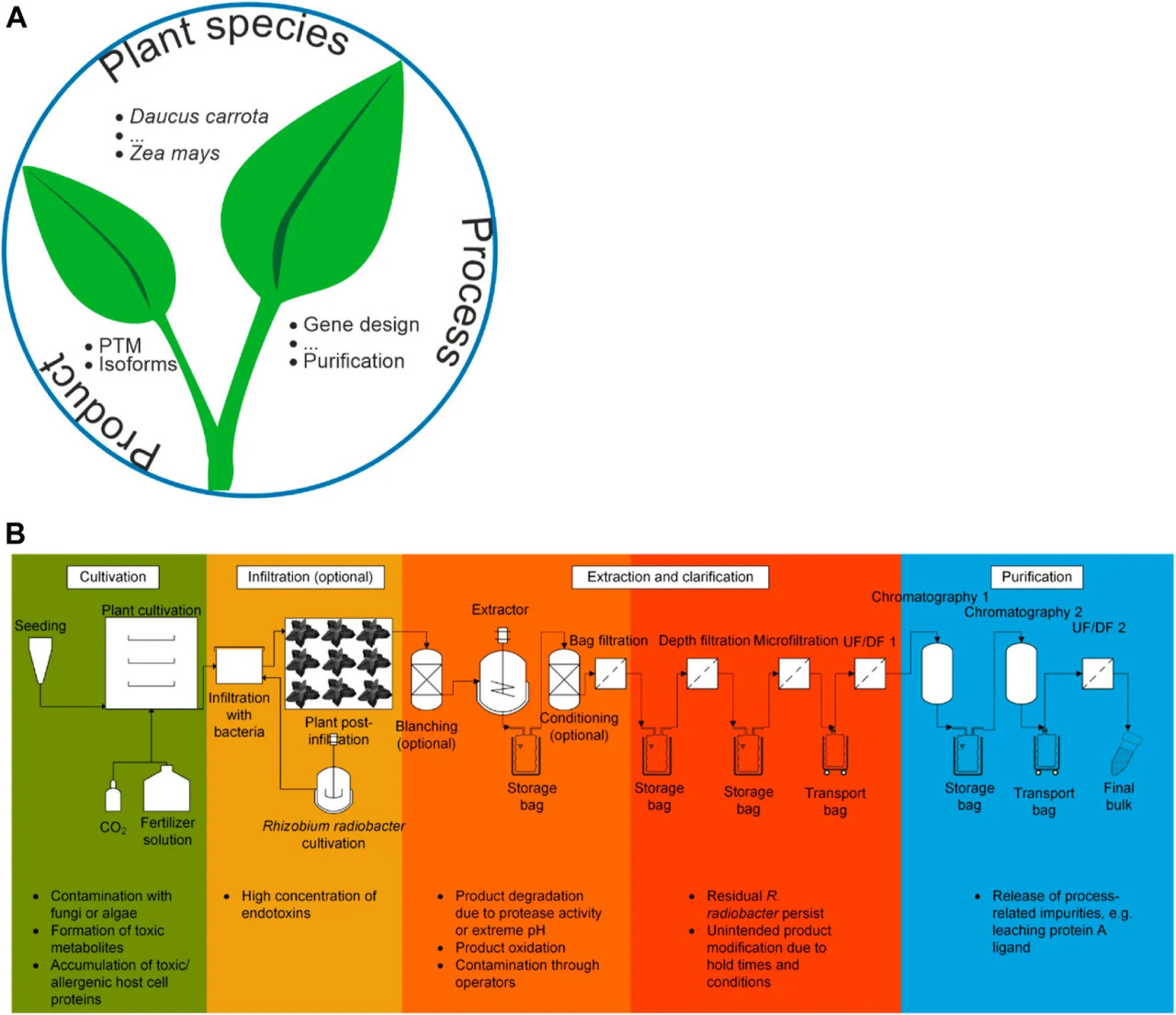
РИСУНОК 1 Аспекты безопасности продукции в молекулярном фарминге растений. (A) Три основных аспекта, определяющих конечную безопасность продукта, обсуждаемые в данном обзоре, включая некоторые примеры. (B) Обобщенная технологическая схема производства рекомбинантных белков в растениях. Культивирование изображено как (полностью) контролируемая система выращивания (Раздел 3.3.2), но могут использоваться и другие условия, например, теплицы. Схема может быть адаптирована для стратегии трансгенной экспрессии путем исключения этапов инфильтрации и культивирования Rhizobium radiobacter. Она также может быть преобразована в процесс суспензионной культуры растительных клеток путем замены культивирования целых растений на последовательность биореакторов. В последнем случае инфильтрация все еще может быть актуальной, если для экспрессии используются пакеты растительных клеток (Rademacher и др., 2019). Некоторые потенциальные факторы риска выделены на каждом этапе процесса.
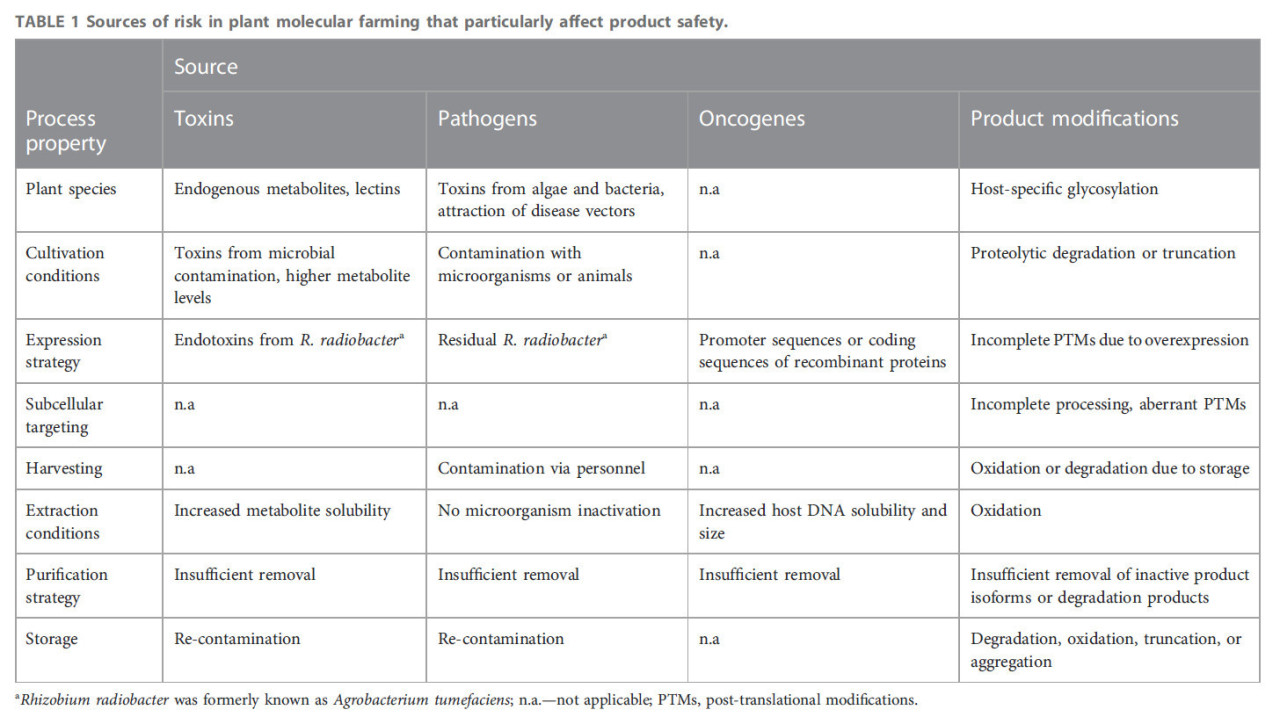
ТАБЛИЦА 1 Источники риска в молекулярном фарминге растений, которые особенно влияют на безопасность продукции.
2 Аспекты безопасности, связанные с хозяином, в молекулярном фарминге растений
2.1 Вредные метаболиты и белки, специфичные для хозяина
2.1.1 Низкомолекулярные метаболиты
Компоненты клеток хозяина определяются как примеси, связанные с процессом, во всех системах экспрессии (Argentine и др., 2007; Arfi и др., 2016; Jones и др., 2021). В некоторых случаях такие молекулы являются непосредственно токсичными, например, липополисахариды, известные как эндотоксины, продуцируемые грамотрицательными бактериями (Раздел 3.1) (Serdakowski London и др., 2012). В отличие от этих крупных компонентов клеточной стенки, которые легко обнаруживаются в специфических анализах, растения и растительные клетки также содержат разнообразные метаболиты с широким динамическим диапазоном концентраций, включая пигменты (например, хлорофилл) и полифенолы (Moore и др., 2014; Wang и др., 2019). Конкретная цель или польза этих сложных низкомолекулярных соединений может быть не очевидна, но они часто являются внутренне биоактивными (Acamovic и Brooker, 2005; Wink, 2009; Napagoda и др., 2022). Соответственно, они используются в качестве пищевых добавок, косметических ингредиентов и фармацевтических препаратов, например, для извлечения противоракового препарата паклитаксела из лекарственных растений (Pereira и др., 2012; Buyel, 2018) и производных клеточных культур (Ochoa-Villarreal и др., 2016). Однако, когда такие биоактивные соединения присутствуют в хозяевах PMF, используемых для производства рекомбинантных белков, они рассматриваются как примеси, которые должны быть удалены в процессе очистки (Таблица 2). Например, никотин очищается из табака для использования в качестве фармацевтического препарата, включая никотинозаместительную терапию и лечение легких когнитивных нарушений (Sanchez-Ramos, 2020; Kheawfu и др., 2021), но когда табак используется для производства рекомбинантных моноклональных антител, никотин является нежелательной примесью (Ma и др., 2015).
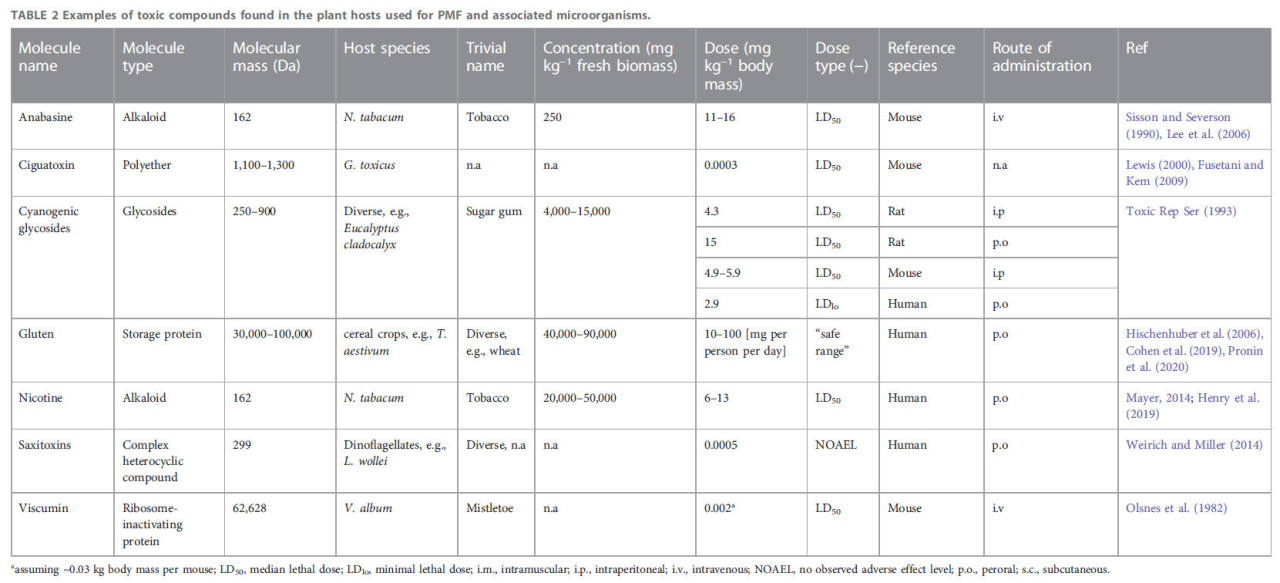
ТАБЛИЦА 2 Примеры токсичных соединений, обнаруженных в растениях-хозяевах, используемых для PMF, и связанных с ними микроорганизмах.
Пасленовые растения, такие как табак, перец (Capsicum annuum), картофель (Solanum tuberosum) и томат (Solanum lycopersicum), привлекательны для применений PMF, поскольку они производят большое количество биомассы [например, 100 000–500 000 кг га⁻¹ год⁻¹ для табака (Stoger и др., 2002; Huebbers и Buyel, 2021)]. Однако они также содержат нежелательные или даже токсичные алкалоиды, такие как капсаицин, соланин, анабазин и никотин (Green и др., 2013; Stephan и др., 2017; Günthardt и др., 2018). Последний имеет расчетную среднюю летальную дозу 6–13 мг кг⁻¹ массы тела у человека (пероральное поступление; внутривенное, вероятно, меньше) (Mayer, 2014), но концентрации всего 0,025 мг кг⁻¹ могут вызывать биологические реакции, такие как изменение крутящего момента разгибателя голени (Mündel и др., 2017). В случае анабазина основную озабоченность вызывает тератогенный потенциал, а не острая токсичность (Keeler и др., 1984; Green и др., 2013), хотя трудно подобрать подходящую модельную систему (Welch и др., 2014).
Аналогичным образом, цианогенные гликозиды являются соединениями, происходящими от аминокислот, которые обнаруживаются у многих растений, включая сельскохозяйственные культуры, на разных стадиях их жизненного цикла, в зависимости от обеспечения питательными веществами (Gleadow и Møller, 2014; Lechtenberg и др., 2005-2010). Эта сложная группа молекул, вероятно, возникла как защита от травоядных, и их токсичность обусловлена высвобождением циановодорода при контакте с определенными β-глюкозидазами. В зависимости от вида растения и ткани цианогенные гликозиды могут присутствовать в концентрациях до ∼8 г кг⁻¹ сухого растительного вещества. Например, их концентрация в молодых листьях Eucalyptus cladocalyx вдвое выше, чем в старых листьях (Gleadow и Woodrow, 2000). Аналогичные концентрации обнаружены у бамбука (Bambusa vulgaris, 1–8 г кг⁻¹) (Nyirenda и др., 2021). Некоторые растительные соединения являются еще более токсичными, включая сапонины и гликоалкалоиды (Wink, 2009; Napagoda и др., 2022; Rasool и др., 2022).
Поэтому необходимо удалять такие метаболиты в процессе очистки продукта, снижая их концентрацию не только ниже уровня токсичности, но и ниже уровня минимального эффекта, который может быть неизвестен или трудноопределим. Кроме того, конкретное соединение в растительном экстракте, которое вызывает определенную биологическую реакцию (например, нарушение иммунных ответов), может быть еще не известно (Harwanto и др., 2022; Urbański и др., 2023). Создание и обновление систематических баз данных растительных токсинов (Günthardt и др., 2018) может помочь рационально определить риски, связанные с определенными растениями-хозяевами. Соответствующие количественные анализы также необходимы для успешной, целенаправленной и рациональной разработки безопасных процессов.
Одним из практических примеров удаления метаболитов является производство моноклональных антител в табаке для клинических испытаний на людях. Никотин был удален ниже предела обнаружения с помощью простого двухстадийного процесса очистки, состоящего из аффинной хроматографии на смоле белка А и полирующей стадии с использованием керамического гидроксиапатита (Ma и др., 2015). Это стало возможным главным образом потому, что размер (или масса) продукта — моноклонального антитела — и никотина (то есть три порядка величины), а также их поверхностные свойства существенно различаются (например, с точки зрения заряда и гидрофобности). Аналогичные результаты были получены и другими исследователями (Fu и др., 2010). Эффективное разделение может быть более сложной задачей, если продукт также является низкомолекулярным соединением, особенно если физико-химические свойства продукта и примесей сходны (например, с точки зрения растворимости). В частности, это исключило бы использование мембранных аппаратов, таких как ультрафильтрация/диафильтрация, которые могут удалять низкомолекулярные примеси при операциях замены буфера при очистке более крупных белков (Opdensteinen и др., 2018). Поэтому полезно выбирать растения-хозяева, в которых либо нет известных токсичных метаболитов, либо такие метаболиты легко отделяются от продукта (Таблица 3). Соответственно, несколько пищевых растений или полученных из них клеточных культур использовались для производства безопасных биологических препаратов, включая морковь (Daucus carota), салат-латук (Lactuca sativa), кукурузу (Zea mays), ячмень (Hordeum vulgare) и рис (Oryza sativa) (Xu и др., 2011; Grabowski и др., 2014; Mirzaee и др., 2022; Ganesan и др., 2023). Но даже продовольственные культуры могут содержать низкие концентрации токсичных алкалоидов, которые необходимо удалять при переработке, например, лупинин из люпина (Lupinus mutabilis) (Kaiser и др., 2020; Griffiths и др., 2021). Традиционная селекция и генетическая модификация могут быть использованы для истощения или даже полного удаления таких метаболитов, как обсуждается более подробно ниже (см. Раздел 5). В целом, риск, связанный с низкомолекулярными примесями растительного происхождения, невелик, если продуктом является рекомбинантный белок, поскольку схемы очистки обычно включают этапы разделения по размеру для удаления агрегатов белков и продуктов деградации, и эти этапы также обеспечивают удаление алкалоидов и других биоактивных метаболитов.
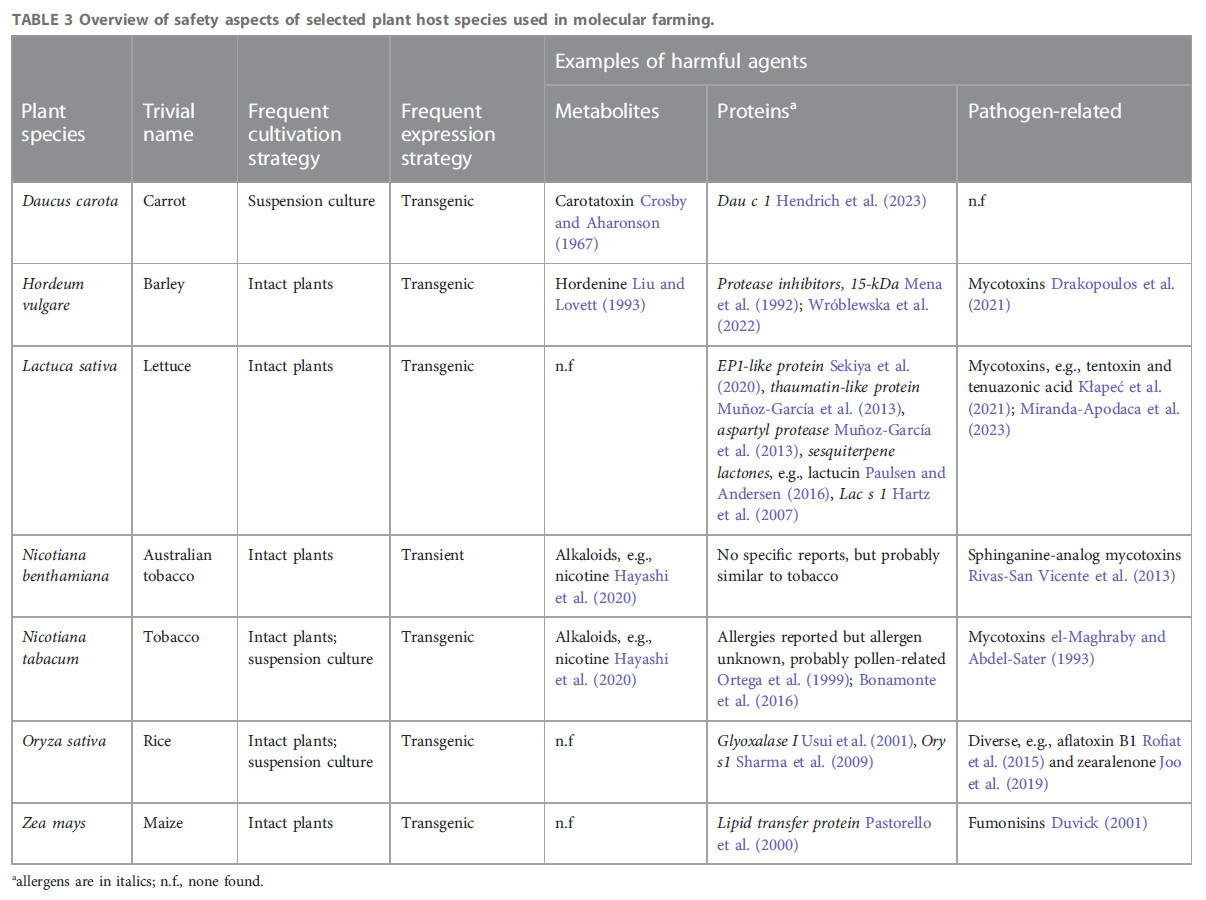
ТАБЛИЦА 3 Обзор аспектов безопасности некоторых видов растений-хозяев, используемых в молекулярном фарминге.
2.1.2 Белки растительных клеток-хозяев
Растения содержат не только токсичные метаболиты, но и некоторые вредные белки. Наиболее токсичными белками являются рибосом-инактивирующие токсины, такие как рицин или висцумин, но растения, продуцирующие такие токсины, не используются в качестве хозяев для PMF (Olsnes и др., 1982; Worbs и др., 2011). Однако другие лектины, такие как агглютинин рисовых отрубей (UniProt ID Q0JF21; ∼22 кДа) или лектин гороха (Pisum sativum) (UniProt ID P02867; ∼30 кДа), присутствуют в продовольственных культурах, используемых в PMF (Miyoshi и др., 2001; Kabir и др., 2013). Эти белки могут останавливать клеточный цикл, ингибировать пролиферацию или вызывать апоптоз у животных и поэтому представляют собой соответствующий риск для безопасности, который должен контролироваться (Jiang и др., 2015). Из-за своего размера они могут соочищаться с такими продуктами, как циановирин-N (∼11 кДа) (Opdensteinen и др., 2018), но должны легко отделяться от крупных белков, таких как антитела (∼150 кДа) (Ma и др., 2015). Однако углевод-связывающая активность многих лектинов заставляет их связываться с гликозилированными белками-мишенями, что может приводить к соочищению. Аналогичные неспецифические взаимодействия были зарегистрированы между белками клеток-хозяев яичника китайского хомячка и моноклональными антителами (Li, 2022). Условия, подавляющие такие взаимодействия, должны быть определены в ходе разработки нисходящего процесса.
Присутствие глютенов представляет собой еще один белковый риск, который особенно актуален при использовании зерновых культур в качестве хозяев для PMF (Ito, 2015; Abedi и Pourmohammadi, 2020). Глютены представляют собой разнообразные белки, которые могут быть классифицированы как глютенины или глиадины (также известные как осборновские фракции) (Osborne, 1907; Biesiekierski, 2017). Эти белки сами по себе не токсичны, но они присутствуют в гораздо более высоких концентрациях, чем большинство токсичных белков, и являются сильными аллергенами. В пшенице (Triticum aestivum) содержится 40–90 г глютена на килограмм пшеничной муки (Pronin и др., 2020). Глютены могут вызывать иммунные ответы при концентрациях ∼12 мг кг⁻¹ массы тела у человека (Lähdeaho и др., 2011; Cabanillas, 2020; Taraghikhah и др., 2020). В то время как некоторая толерантность может быть выработана у пациентов с целиакией (Elli и др., 2020), был предложен порог безопасности 10–100 мг на человека в день (Hischenhuber и др., 2006; Cohen и др., 2019). Важно, что глютены растворимы в воде и накапливаются в семенах, куда в зерновых культурах обычно направляются рекомбинантные белки, поскольку это повышает стабильность продукта (Tosi и др., 2011; Arcalis и др., 2014). Поэтому концентрация этих аллергенов в первичных экстрактах семян высока. Глютены термостабильны (Biesiekierski, 2017) и имеют молекулярную массу от ∼30 до >100 кДа (Tosi и др., 2011), поэтому их может быть трудно отделить от белков-мишеней с помощью бланширования/нагревания (Buyel и др., 2016) или ультрафильтрации/диафильтрации (Opdensteinen и др., 2018). Хотя присутствие глютена создает проблемы с точки зрения разработки нисходящего процесса, общее влияние на безопасность невелико. В частности, глютены легко обнаруживаются (Schubert-Ullrich и др., 2009), и фармацевтические белки должны иметь чистоту более 95% (Jin и др., 2018). В маловероятном случае, если продукт PMF содержит 5% глютена и требуются большие дозы продукта (например, 0,05 г антител к вирусу Эбола на килограмм массы тела каждые 3 дня (Davey и др., 2016)), пациент массой 70 кг будет подвергаться воздействию в среднем ∼175 мг глютена в день, что примерно вдвое превышает безопасный порог. Хотя маловероятно, что одно соединение будет составлять все примеси в продукте, эта оценка подчеркивает важность удаления даже соединений, которые могут рассматриваться как умеренный риск для безопасности, таких как аллергены. Это особенно актуально в случаях, когда требуются высокие дозы продукта (до нескольких граммов на человека), как это может быть при постэкспозиционной профилактике, лечении острых заболеваний (Taylor и др., 2021; Hwang и др., 2022) или противораковой терапии (Hendrikx и др., 2017). Это также актуально для неантительных продуктов, которые не могут быть захвачены аффинной хроматографией, и не фармацевтических продуктов, таких как пищевые добавки, где любая форма хроматографии была бы слишком дорогой.
Важно, что количество, распространенность и активность опасных белков могут быть снижены, в некоторых случаях ниже уровня обнаружения, с помощью разработки процесса (см. Раздел 3.4) и стратегий генетической инженерии (см. Раздел 5). Например, большинство белков растительных клеток-хозяев может быть удалено с помощью анионообменной хроматографии (Buyel и Fischer, 2014a; Bernau и др., 2022).
2.2 Загрязнение переносчиками заболеваний и продуктами патогенов растений
В дополнение к вредным молекулам, продуцируемым растениями, хозяева PMF также могут привлекать вредителей и патогены, которые могут непосредственно наносить вред человеку или продуцировать токсичные белки и метаболиты, что является активной областью исследований в контексте безопасности пищевых продуктов (Fletcher и др., 2013; Sobiczewski и Iakimova, 2022). Например, грибы, поражающие зерновые, продуцируют (спорыньи) алкалоиды и канцерогенные микотоксины (Hulvová и др., 2013; Florea и др., 2017; Sweany и др., 2022), последние включают афлатоксин B1, который токсичен при микромолярных концентрациях (Bianco и др., 2012; Marchese и др., 2018). Аналогично, прокариотические сине-зеленые водоросли (цианопрокариоты), такие как Lyngbya wollei, и эукариотические зеленые водоросли (хлорофиты), такие как динофлагелляты (например, Ostreopsis siamensis и Gambierdiscus toxicus), могут колонизировать среду обитания человека (Hofbauer, 2021), такую как личные аквариумы, ирригационные/дренажные канавы или заливные столы (см. Раздел 3.3) и соответствующие резервуары с удобрениями. Водоросли могут распространяться по воздуху, а также размножаться в почве или на блоках каменной ваты, часто используемых для поддержки роста растений в PMF. Риск для биопроизводства отражает способность водорослей продуцировать аллергены и токсины, такие как цигуатоксин и майтотоксин (оба из G. toxicus), которые вызывают диарею и рвоту у людей (Friedman и др., 2017; Hofbauer, 2021) или даже смерть (Ohizumi и Yasumoto, 1983). В частности, LD50 цигуатоксина у мышей составляет ∼250 нг кг⁻¹ при внутрибрюшинном введении (Lewis, 2000), а майтотоксин имеет минимальную летальную дозу ∼170 нг кг⁻¹ (Bagnis и др., 1980; Ohizumi и Yasumoto, 1983). Аналогичным образом, цианопрокариоты продуцируют сакситоксины, такие как токсин-1 L. wollei, с уровнем отсутствия наблюдаемого неблагоприятного эффекта ∼500 нг кг⁻¹ массы тела после перорального поступления у человека (Weirich и Miller, 2014). Для внутривенных фармацевтических препаратов NOAEL, вероятно, будет ниже.
Некоторые грибковые (Alternaria infectoria) и бактериальные (Erwinia persinicus) патогены растений могут вызывать оппортунистические инфекции у людей. Например, Rhizobium radiobacter, широко используемый для транзиентной экспрессии в приложениях PMF, может вызывать бактериемию и кератит, как недавно было рассмотрено (Kim и др., 2020). Однако число зарегистрированных случаев крайне мало (<50 в доступной литературе), несмотря на повсеместное присутствие этого вида в почве и искусственных средах, таких как лаборатории (Dessaux и Faure, 2018; Zhu и др., 2020). Кроме того, большинство пациентов, страдающих от спорадического заболевания, которое, по-видимому, было связано с инфекцией R. radiobacter, были иммунокомпрометированы, или место инфекции было связано с хирургическим вмешательством или глазом (Kim и др., 2020), где адаптивная иммунная система особенно слаба (Akpek и Gottsch, 2003). Соответственно, риск заражения патогенами растений для человека представляется минимальным, особенно учитывая, что фармацевтические продукты проходят (несколько) этапов стерилизующей фильтрации или даже более строгого разделения по размеру (например, ультрафильтрация/диафильтрация или гель-проникающая хроматография), которые удалят любые неповрежденные клетки (Holtz и др., 2015; Ma и др., 2015). Эндотоксины представляют собой более актуальный, связанный с процессом риск, специфически ассоциированный с R. radiobacter и транзиентной экспрессией (см. Раздел 3.2).
В отличие от таких бактерий, растительные вирусы не инфицируют и не реплицируются в клетках человека и поэтому вряд ли вызывают заболевания. Растительные вирусы могут обнаруживаться в ассоциации с человеком, но связь, как полагают, является косвенной — например, РНК вируса табачной мозаики была обнаружена в слюне человека, но ее присутствие было связано с курением, а не с инфекцией (Balique и др., 2012). Аналогично, ДНК табака была обнаружена у пациентов с вентилятор-ассоциированной пневмонией, которые были курильщиками (Bousbia и др., 2010).
Растения также могут привлекать насекомых, которые могут быть переносчиками заболеваний человека. Например, летучие вещества (особенно терпеноиды), выделяемые некоторыми видами растений, могут привлекать комаров, таких как Anopheles gambiae (Nyasembe и др., 2012; Nikbakhtzadeh и др., 2014), ключевого переносчика малярии. Кроме того, виды растений, которыми питаются комары, также могут влиять на жизнеспособность Plasmodium falciparum (Hien и др., 2016), одного из паразитов, вызывающих малярию. В частности, комары, питавшиеся плодами Mangifera indica вместо черенков Thevetia neriifolia или глюкозного контроля, имели ∼50% меньшую вероятность выжить в течение 7 дней, а среднее число развивающихся ооцист в кишечнике инфицированных самок комаров было снижено на ∼60%. Некоторые виды растений или условия культивирования также могут привлекать грызунов, переносящих патогены.
Риск, связанный с патогенами растений, насекомыми и другими животными в контексте PMF, невелик. Например, культуры PMF, такие как табак, продуцируют лишь низкие концентрации терпеноидов, которые вряд ли привлекут комаров (Lücker и др., 2004). В зонах умеренного климата такие переносчики болезней в любом случае вряд ли будут присутствовать в окрестностях производственной площадки. В целом, условия культивирования в PMF не благоприятствуют многим из вышеупомянутых патогенов, а естественный микробиом растений может снижать приспособленность таких патогенов, как P. falciparum (Bassene и др., 2020). Как обсуждается ниже, существует множество вариантов конструкции оборудования и процесса, позволяющих минимизировать или даже исключить риски, связанные с вредителями и патогенами, включая ультрафиолетовые лампы или генераторы озона для инактивации водорослей и бактерий в ирригационных системах или переносимых персоналом, контролируемую среду и ловушки (например, мышеловки и желтые клеевые ловушки для насекомых) для защиты зоны культивирования от патогенов, распространяемых животными.
3 Потенциальные риски, возникающие при разработке биопроцесса
Решения, принимаемые в процессе разработки процесса, могут значительно влиять на безопасность продукта. Некоторые этапы процесса непосредственно направлены на обеспечение безопасности, включая этапы выдерживания при низком pH для инактивации вирусов (Mazzer и др., 2015), но другие решения могут иметь непреднамеренные последствия, и их следует избегать или смягчать.
3.1 Элементы экспрессионной кассеты
Онкогены или их части могут использоваться в качестве продуктов или в качестве строительных блоков для экспрессионных векторов, включая регуляторные элементы для усиления накопления продукта. В качестве примера, связанного с продуктом, онкогенный белок E7 вируса папилломы человека связывается с белком ретинобластомы и необходим для поддержания жизнеспособности папилломавирус-индуцированных опухолей, как обнаружено в широко используемой клеточной линии HeLa (Nishimura и др., 2006). Однако белок E7 также продуцировался в растениях (и многих других хост-системах) в качестве кандидата вакцины для лечения инфекций, вызванных вирусом папилломы человека 16 (Venuti и др., 2009; Buyel и др., 2012). Соответственно, продукт вакцины потенциально может содержать остаточную ДНК клеток-хозяев, включая последовательности, кодирующие онкогенный рекомбинантный белок. В конкретном случае E7 кодирующая последовательность была мутирована, чтобы сделать белок нетуморогенным и, таким образом, снизить этот риск (Smahel и др., 2001). Однако такие решения требуют точного знания сайтов связывания/взаимодействия белка, которое может быть не всегда доступно.
Риски, связанные с такой онкогенной ДНК, также могут включать регуляторные последовательности, и вероятность того, что остаточная ДНК может трансформировать клетки животных и в конечном итоге вызвать развитие опухолей, обсуждалась (Peden и др., 2006). Были созданы модели для оценки связанного риска (Yang и др., 2010), который, по-видимому, является незначительным, основываясь на данных многочисленных исследований (Palladino и др., 1987; Dortant и др., 1997). В частности, было признано необходимым 1–10 г остаточной ДНК хозяина для индукции опухоли (Sheng и др., 2008), тогда как нормативный порог для геномной ДНК клеток-хозяев в фармацевтических продуктах составляет ∼0,1–1,0 нг на дозу (Wang и др., 2012). Учитывая, что доступны чувствительные методы обнаружения на основе ПЦР и что ДНК является высокозаряженным полимером, который может быть эффективно удален с помощью анионообменной хроматографии (Stone и др., 2018), риск, связанный с остаточной ДНК, невелик. При необходимости этап ферментативной обработки может дополнительно снизить нагрузку остаточной ДНК и, таким образом, повысить безопасность продукта (Kawka и др., 2021).
3.2 Стратегия экспрессии и эндотоксины
Эндотоксины являются хорошо известным фактором риска в биопроизводстве, поскольку они являются сильными активаторами и модуляторами иммунной системы человека, приводя к септическому шоку (Opal, 2010). Такие токсины в изобилии присутствуют в процессах, где в качестве хозяев используются грамотрицательные бактерии (Petsch и Anspach, 2000), но они также актуальны в PMF из-за преднамеренного использования бактерий для переноса генов, а также из-за наличия случайных бактерий на поверхности или внутри растительных тканей. В масштабируемой трансгенной системе (Buyel и др., 2017) нагрузка эндотоксинов обычно низка, поскольку бактерии, используемые для переноса генов, уничтожаются после стабильной интеграции трансгенов в ядерный или пластидный геном растения (Herrera-Estrella и др., 2005). Поэтому единственными присутствующими грамотрицательными бактериями будут те, которые естественным образом встречаются на поверхности растений, например, Pseudomonas spp. (Compant и др., 2019). В отличие от этого, транзиентная экспрессия (Tusé и др., 2020) требует инфильтрации растений грамотрицательной бактерией R. radiobacter (Spiegel и др., 2019), а стресс, вовлеченный в этот процесс, также стимулирует продукцию эндотоксинов, а также синтез вторичных метаболитов в растениях (Buyel и др., 2015b). Соответственно, концентрация эндотоксинов может увеличиваться в 200 раз до ∼10⁴ ЕД на миллиграмм общего белка (Arfi и др., 2016), что составляет ∼3 × 10⁴ ЕД мл⁻¹. Это находится в том же диапазоне, что и лизат Escherichia coli (10³–10⁵ ЕД мл⁻¹) (Szermer-Olearnik и Boratyński, 2015), и превышает нормативный порог в 5 ЕД кг⁻¹ массы тела ч⁻¹ (Hirayama и Sakata, 2002), даже предполагая микрограммовые дозы белка (Cummings и др., 2014). Соответственно, эндотоксины должны быть удалены, особенно когда продукт производится с помощью транзиентной экспрессии. Для этой цели могут использоваться специализированные методы, такие как фазовое разделение (Aida и Pabst, 1990), аффинная сорбция (Anspach, 2001), хроматография (Serdakowski London и др., 2012) и ультрафильтрация/диафильтрация (Jang и др., 2009), как было продемонстрировано для ряда растительных экстрактов (Arfi и др., 2016). Однако многие из этих этапов обычно включены в стандартные схемы нисходящей обработки (Ma и др., 2015; Opdensteinen и др., 2018; Knödler и др., 2023b), поэтому дополнительные усилия для удаления эндотоксинов обычно не требуются, при условии тщательного контроля процесса.
Гибридным подходом является использование индуцируемой экспрессии трансгенов (Mortimer и др., 2015; Hahn-Löbmann и др., 2019). Эта стратегия имеет те же сроки разработки, что и трансгенный подход, но обеспечивает ограниченную по времени экспрессию продукта, тем самым минимизируя токсическое влияние последнего на развитие и рост растений. С точки зрения безопасности продукта, она аналогична трансгенным растениям (низкий уровень эндотоксинов и отсутствие R. radiobacter), а индуктор должен быть выбран так, чтобы обеспечить эффективность (активность при низких концентрациях), легкое удаление (например, в идеале низкомолекулярное соединение, такое как этанол) и отсутствие токсичности.
3.3 Условия культивирования
Важно, что независимо от стратегии экспрессии и условий культивирования, продукты PMF могут производиться без компонентов животного происхождения, поскольку для культивирования растений, растительных клеток и R. radiobacter могут использоваться определенные или веганские удобрения/среды (Houdelet и др., 2017; Leth и McDonald, 2017; Geng и др., 2019; Kang и др., 2022). Таким образом, можно исключить загрязнение патогенами и вредными агентами, такими как прионы, в субстратах, что повышает безопасность продуктов PMF.
3.3.1 Суспензионные культуры растительных клеток
Биореакторы, подходящие для обычных культур микробных и клеток млекопитающих, также могут использоваться для растительных клеток [и даже растительных тканей и целых растений (Murthy и др., 2023)] с небольшими модификациями или без них (Holland и др., 2013). Эти реакторы обеспечивают высокую степень изоляции процесса и минимизируют или даже устраняют некоторые из рисков, обсуждаемых выше (Huang и McDonald, 2012). Например, бактерии, колонизирующие поверхности растений, не будут обнаружены в биореакторе. Тем не менее, необходимо соблюдать осторожность при инокуляции и сборе урожая из реакторов, особенно когда большие объемы (т.е. несколько литров) обрабатываются на поздних стадиях типичной посевной цепочки реакторов, поскольку стерильность может быть трудно обеспечить, как хорошо известно для других биопроцессов (Müller и др., 2022). Риск загрязнения может быть снижен, если, например, используются реакторы с орбитальным встряхиванием (Raven и др., 2014) или аэрлифтные/барботажные реакторы (Wilson и Roberts, 2012), поскольку они содержат меньше движущихся частей, уплотнений и фитингов, чем реакторы с механическим перемешиванием (Werner и др., 2018). Аналогично, одноразовые реакторы могут снизить риски перекрестного загрязнения продуктами (Raven и др., 2011). Использование фотобиореакторов также повышает безопасность, поскольку автотрофное культивирование растительных клеток в таких реакторах не требует органических источников углерода в культуральной среде (Legrand и др., 2021), фактически лишая ее субстрата, необходимого для роста многих загрязняющих бактерий, дрожжей и грибов (хотя фототрофные бактерии и водоросли остаются риском загрязнения). Культивирование растительных клеток в фотобиореакторах также требует накопления хлорофиллов и других пигментов, и эти соединения могут непреднамеренно взаимодействовать с молекулами продукта (см. Раздел 3.4.2). Другим недостатком фотобиореакторов является то, что они обычно используют нестандартные конструкции, например, трубчатую геометрию, для обеспечения достаточного освещения (Chanquia и др., 2022).
Растительные клетки в суспензии часто имеют тенденцию прилипать даже к поверхностям из нержавеющей стали в биореакторе (Holland и др., 2017). Это не только ограничивает время работы биопроцесса, но и может препятствовать клеточному метаболизму, ограничивая поступление кислорода и/или питательных веществ в образующихся клеточных агрегатах. Эти неоптимальные условия могут приводить к клеточному стрессу, аутофагии (например, пероксисом) и гибели клеток (Voitsekhovskaja и др., 2014; Tyutereva и др., 2018; Ma и др., 2022), что может вызывать (частичную) деградацию или модификацию продукта, в конечном итоге увеличивая гетерогенность продукта и снижая активность. Кроме того, если клетки начинают разлагаться в этих поверхностных агрегатах, а затем снова попадают в объемную ферментационную среду, высвобождаемые ими молекулы могут запускать нежелательные сигнальные каскады в живых клетках, снижая общую продуктивность (Salguero-Linares и Coll, 2023). Поэтому разработка линий производственных клеток и протоколы культивирования должны быть сосредоточены на низкой адгезии и низкой агрегации, а также на стратегиях мониторинга для обеспечения того, чтобы качество продукта не было скомпрометировано.
3.3.2 Культивирование целых растений
Культивирование растений в открытом поле в настоящее время подходит только для низкомолекулярных фармацевтических продуктов, таких как морфин, который экстрагируется из маковой соломы (Krikorian и Ledbetter, 1975). Эти молекулы имеют простую структуру, хорошо определенную конформацию и обычно выделяются с использованием органических растворителей, которые также действуют как дезинфицирующие средства (Kuyukina и др., 2014). Поэтому контроль качества продукта прост (например, анализ ЖХ-МС), а любые загрязнители эффективно удаляются жесткими условиями экстракции.
Напротив, на сегодняшний день белковые фармацевтические препараты производились в растениях, выращенных в закрытых помещениях. Это позволяет избежать любых непредсказуемых воздействий переменной внешней среды и обеспечивает соблюдение требований GMP. В частности, экстракция рекомбинантных белков обычно основана на водных буферах (Buyel и др., 2015a), которые не инактивируют патогены, занесенные насекомыми-вредителями и грызунами (см. Раздел 2.2). Дополнительными факторами, препятствующими культивированию в открытом поле, являются ионы тяжелых металлов, пестициды и антропогенные токсичные загрязнители, которые могут загрязнять почвы (Sigmund и др., 2022; D'Angelo и др., 2013) и растительные ткани, и потенциально конечный продукт (Zeng и др., 2019; Zhang и др., 2020).
Такие риски могут быть предотвращены, если растения культивируются в теплицах, о чем сообщалось для нескольких процессов, соответствующих GMP, по производству моноклональных антител и вакцин (Ma и др., 2015; Ward и др., 2021). Кроме того, в таких условиях могут использоваться хорошо определенные субстраты для роста, такие как блоки каменной ваты, в сочетании с автоматизированными гидропонными системами орошения. Закрытая среда облегчает эффективную борьбу с вредителями и позволяет строго регламентировать доступ персонала, что минимизирует риск загрязнения патогенами. Однако контроль процесса в такой среде все еще ограничен. Например, выходы одного и того же антитела могут колебаться от 2 до 6 г на партию растений массой 200 кг из-за сезонных эффектов и переменной погоды (Sack и др., 2015). Более важно, что целостность продукта может быть нарушена из-за активности протеаз, например, в условиях интенсивного освещения или высокой температуры, которые не могут быть смягчены климат-контролем (Knödler и др., 2019). Даже если климат-контроль поддерживает условия культивирования в пределах спецификаций, листья могут становиться горячее, чем окружающая среда, из-за интенсивной инсоляции (Huebbers и Buyel, 2021). Интенсивный свет также может вызывать синтез потенциально вредных метаболитов (Buyel и др., 2015b; Thoma и др., 2020) (см. Раздел 2.1), которые необходимо удалять в ходе нисходящей обработки. Поскольку теплицы обычно являются нестерильными средами, вероятно, что водоросли начнут расти на поверхностях и в растворе удобрений, особенно если резервуары и канавы/заливные столы не покрыты должным образом. Как обсуждалось выше, эти прокариотические и эукариотические водоросли могут быть вредными или выделять токсичные соединения (см. Раздел 2.2). Поэтому для снижения воздействия водорослей следует устанавливать ультрафиолетовые лампы или генераторы озона (Sharrer и Summerfelt, 2007).
Закрытые культивационные сооружения обеспечивают еще более высокую степень контроля процесса, чем теплицы. Эти сооружения предназначены для устранения любого воздействия окружающей среды на рост растений путем обеспечения полного искусственного климата: температуры, влажности и орошения, а также света и, возможно, газового состава (Farhangi и др., 2023). Терминология, используемая для закрытых культивационных сооружений, может вводить в заблуждение и быть неоднозначной. Например, их часто называют «вертикальными фермами», потому что несколько вертикально штабелированных слоев культивирования могут повысить экономическую эффективность, но также могут использоваться и однослойные конструкции (Huebbers и Buyel, 2021). Альтернативный термин «закрытое хозяйство» или «закрытое сельское хозяйство» также неточен, поскольку это может быть расширено и на теплицы. Поэтому более точным термином могут быть (полностью) «контролируемые системы выращивания».
Независимо от терминологии, цифровая интеграция обеспечивает контроль над отдельными параметрами, такими как состав удобрений и свет (Huebbers и Buyel, 2021; Kaur и др., 2023), но требует датчиков или даже сетей датчиков, учитывающих дискретные характеристики отдельных растений (Huebbers и Buyel, 2021). Например, метаболитный и липидный состав растений может модулироваться путем выбора определенных длин волн света для освещения (Rihan и др., 2022), и, таким образом, может помочь снизить концентрацию потенциально вредных метаболитов, таких как алкалоиды (см. Раздел 2.1.1). Кроме того, CEGS могут быть полностью автоматизированы, так что вмешательство человека и, в конечном итоге, риск загрязнения патогенами человека минимизируется (Wirz и др., 2012; Huebbers и Buyel, 2021; Ren и др., 2023). Высокая степень автоматизации/механизации в таких системах и близость соответствующих устройств и растений увеличивают вероятность контакта продукта с вспомогательными и эксплуатационными материалами, такими как смазки. Поэтому все устройства должны быть спроектированы так, чтобы минимизировать такие риски. Это включает выбор соответствующих строительных материалов, включая стали, совместимые с пищевыми или фармацевтическими применениями, и пластики, лишенные выщелачиваемых веществ (Jenke, 2002; Cuadros-Rodríguez и др., 2020; Zimmermann и др., 2021).
CEGS в целом являются наиболее безопасной средой для производства биологических белков с помощью PMF. Эта технология масштабируема (например, несколько сотен килограммов биомассы могут быть переработаны в неделю (Holtz и др., 2015)), но высокие инвестиционные и энергетические затраты остаются существенным узким местом (Huebbers и Buyel, 2021; van Delden и др., 2021). Это особенно актуально, если продукты PMF не предназначены для фармацевтического применения, где ценовое давление на производство выше, и инфраструктура с низкими инвестициями может быть единственным вариантом для создания экономически жизнеспособных процессов.
3.4 Экстракция и очистка продукта
Хотя очистка продукта является ключевым требованием GMP в PMF, по крайней мере, для продуктов, которые будут вводиться инъекционно, очистка также создает некоторые риски, которые должны быть смягчены в процессе разработки процесса. Типичная последовательность нисходящей обработки в PMF начинается со сбора урожая и дополнительной кондиционирования (например, промывки (Ma и др., 2015) или бланширования (Buyel и др., 2014a); Рисунок 1B). Затем следует начальная экстракция, которая может включать дальнейшие этапы кондиционирования, такие как регулировка pH или флокуляция (Buyel и Fischer, 2014b; Buyel и Fischer, 2014c; Buyel и Fischer, 2014d). Следующей основной операцией является осветление, которое обычно включает несколько этапов фильтрации (Buyel и Fischer, 2014e), ведущее к очистке продукта с помощью двухфазной экстракции (Platis и др., 2008), мембранного разделения (Opdensteinen и др., 2018), хроматографии (Buyel и др., 2012) или их комбинаций. Общий риск заключается в том, что последовательность нисходящих операций не обеспечивает необходимую чистоту из-за недостаточного удаления примесей, связанных с процессом и/или продуктом, но каждая нисходящая операция создает специфические риски для безопасности продукта, которые должны контролироваться и минимизироваться в ходе разработки процесса.
3.4.1 Сбор урожая и кондиционирование
Ручные процессы сбора урожая несут в себе внутренний риск для безопасности, поскольку операторы вступают в тесный контакт с растительной биомассой, содержащей фармацевтический продукт, и могут переносить патогены. Поэтому средства индивидуальной защиты (в данном случае с точки зрения защиты собранной биомассы и продукта) должны всегда использоваться, включая халаты, перчатки, сетки для волос и маски. Кроме того, состояние здоровья операторов должно контролироваться, и персонал следует поощрять сообщать о любых признаках заболевания, чтобы можно было провести замену и/или изменение графика работы. Хотя это обычные процедуры для процессов, соответствующих GMP, на основе клеточных культур, они также важны в контексте целых растений, поскольку руководящие принципы Международной конференции по гармонизации (ICH), такие как Q7¹ (Руководство по надлежащей производственной практике для активных фармацевтических ингредиентов), применяются только к этапам после сбора растений и начальной экстракции. Несмотря на то, что растения не поддерживают репликацию вирусов человека, как обсуждалось выше, сбор целых растений и их частей, таких как листья, в идеале должен быть полностью автоматизирован, как это реализовано в некоторых CEGS, в частности, в закрытых вертикальных фермах (Wirz и др., 2012), поскольку это минимизирует риски загрязнения, связанные с операторами.
В некоторых процессах собранная биомасса подвергается термической предварительной обработке, описываемой как бланширование, при которой растительная биомасса погружается в горячий (50°C–90°C) (Buyel и др., 2014a), возможно, слегка кислый буфер (Opdensteinen и др., 2020), что удаляет 50%–95% белков клеток-хозяев табака (Buyel и др., 2016). Это является преимуществом с точки зрения безопасности продукта, но риски включают частичную или полную необратимую денатурацию продукта, сопровождающуюся изменением активности. Неправильно разработанный этап бланширования может даже увеличить активность протеаз и деградацию продукта (Menzel и др., 2016; Menzel и др., 2018). Поэтому такие этапы кондиционирования должны быть тщательно разработаны и реализованы, включая перекрестную проверку активности продукта в подходящих анализах.
3.4.2 Экстракция и дальнейшее кондиционирование
Ручной сбор урожая обычно сопровождается ручной передачей биомассы в экстракционное устройство, поэтому защитные меры, обсуждаемые выше, должны соблюдаться и для этого последующего этапа. Биологические препараты обычно экстрагируются из растений и растительных клеток путем гомогенизации биомассы в присутствии буфера (Buyel и др., 2015a). Последний контролирует pH и окислительно-восстановительные условия для стабилизации продукта и предотвращения нежелательной модификации окислением или реакцией с растительными пигментами и фенольными соединениями. Окисление и другие нежелательные реакции также могут подавляться экстракцией в атмосфере азота (Ma и др., 2015). Ингибиторы протеаз могут добавляться при экстракции в малом масштабе (Menzel и др., 2016) или коэкспрессироваться в растительных клетках (что также увеличивает накопление продукта (Jutras и др., 2016; Grosse-Holz и др., 2018)), но целостность продукта может быть поддержана просто за счет охлаждения всех буферов до ∼10°C (Ma и др., 2015). Охлаждение буфера полезно, поскольку в процесс ничего не добавляется, но для этого этапа потребуется дополнительное оборудование. Напротив, любые ингибиторы являются дополнительными загрязнителями, которые необходимо удалять впоследствии. То же самое справедливо для методов экстракции, которые вообще не требуют буферов, таких как использование шнекового пресса (Buyel и Fischer, 2014d). Однако условия в получающемся зеленом соке могут отрицательно влиять на продукт и его активность, например, из-за низкого pH (например, ∼5,5 в случае табака). Как и в случае с бланшированием, внедрение таких методов должно сопровождаться тщательной оценкой влияния на стабильность и активность продукта.
Затем экстракт может быть кондиционирован для облегчения последующего осветления и очистки путем регулировки pH, а также добавления флокулянтов и/или фильтрующих добавок. Последние часто представляют собой крупные инертные целлюлозные волокна, которые легко удаляются на последующих этапах очистки (Buyel и др., 2014b), тогда как флокулянты являются высокозаряженными полимерами (Buyel и Fischer, 2014c), которые могут связываться с белками (Jurjevec и др., 2023) и ускользать от обнаружения. Поэтому для фармацевтических продуктов следует использовать самые высокие степени чистоты, чтобы обеспечить безопасность продукта. Хотя не фармацевтические продукты, такие как пищевые добавки, также должны соответствовать надлежащей производственной практике (Manning, 2018), требования к чистоте и безопасности обычно менее строгие, и это следует учитывать при выборе реагентов, чтобы согласовать качество сырья и требования безопасности.
3.4.3 Осветление и очистка
Этапы осветления (в основном фильтрация) и очистки обычно удаляют частицы, включая вирусы, а также растворимые компоненты клеток-хозяев, такие как белки. Поэтому обе операции внутренне повышают безопасность биологических препаратов. Тем не менее, процессы на основе бактерий или клеток животных-хозяев показали, что оба этапа также создают риски с точки зрения безопасности продукта. Например, оборудование может выделять выщелачиваемые и экстрагируемые вещества, и его следует выбирать для минимизации этих рисков, принимая во внимание свойства растительных экстрактов, такие как присутствие фенольных соединений. В частности, белок А, распространенный аффинный лиганд для захвата антител, может обнаруживаться как примесь, связанная с процессом, во фракциях элюата антител (Carter-Franklin и др., 2007), а воздействие фенолов может приводить к необратимому обесцвечиванию хроматографических смол.
Другим основным риском для безопасности продукта является время выдержки, необходимое в процессе обработки экстракта. Например, даже во многих непрерывных процессах используются промежуточные накопительные емкости, которые обеспечивают временной буфер между отдельными этапами нисходящей обработки (например, двумя последовательными фильтрами) для компенсации колебаний объемных потоков. Режим потока в таких емкостях далек от идеального поршневого потока и создает широкое распределение времени пребывания (Sencar и др., 2020; Lali и др., 2022). Поэтому часть продукта будет задерживаться в процессе существенно дольше, чем можно было бы ожидать, исходя из среднего времени пребывания. Это критично на ранних этапах очистки, когда протеазы или оксидазы клеток-хозяев все еще обильны и могут воздействовать на продукт и модифицировать его, потенциально нарушая его активность и безопасность. Охлаждение промежуточных продуктов может снизить такую нежелательную ферментативную активность, но требует дополнительного оборудования. Полностью непрерывный процесс без накопительных емкостей требует сложного управления процессом и уязвим для ошибок, которые создают новые риски для безопасности продукта. Поэтому комплексное управление рисками необходимо в ходе разработки процесса (Sparrow и др., 2013; Zalai и др., 2013; Qiu и др., 2015; Luo и др., 2021).
4 Свойства продукта, значимые для безопасности
Целевой профиль биологических препаратов, производимых с помощью PMF, основан на тех же аспектах, которые предусмотрены для других биопроцессов, таких как эффективность и безопасность². Оба зависят от молекулярных свойств продукта, таких как целостность последовательности, фолдинг и PTMs, такие как дисульфидные связи, фосфорилирование и гликозилирование. Последнее может влиять на фолдинг, но в контексте безопасности продукта основной проблемой является иммуногенность, поскольку N-связанные гликаны растений отличаются от таковых у млекопитающих в нескольких фундаментальных аспектах, включая присутствие остатков ксилозы (не встречающихся у млекопитающих) и связывание фукозы через α3-гликозидную связь (у млекопитающих обнаруживается α6-связь) (Strasser, 2016). Кроме того, O-связанные гликаны у растений в основном обнаруживаются на остатках гидроксипролина, тогда как серин и треонин являются предпочтительными мишенями у млекопитающих. Эти неродные гликаны могут вызывать иммунные ответы, когда рекомбинантные белки человека, продуцируемые с помощью PMF, вводятся инъекционно (Bardor и др., 2003; Jin и др., 2008). Однако нет никаких доказательств того, что образование антител к гликанам является вредным (Shaaltiel и Tekoah, 2016; Rup и др., 2017). Ситуация может быть иной у пациентов с аллергией в анамнезе (Schwestka и др., 2021), которые могут быть особенно чувствительны к гликановым структурам растительного происхождения. Аналогично, люди с аллергией на белки яйца, включая гликопротеины, такие как овотрансферрин и овомукоид (Hwang и др., 2014), могут испытывать легкие, но нежелательные побочные эффекты, похожие на аллергию (не анафилаксию), при получении вакцин против гриппа, произведенных в яйцах (James и др., 1998; Gruenberg и Shaker, 2011). Для решения этой проблемы были разработаны различные линии растений (клеток), у которых отсутствуют растительные гликозилтрансферазы (Strasser и др., 2004; Jansing и др., 2018), а в некоторых случаях также интегрированы ферменты человека, чтобы сделать гликаны не только совместимыми с человеком, но и полностью гуманизированными (Castilho и др., 2013; Montero-Morales и Steinkellner, 2018). Соответствующие продукты PMF могут достигать большей активности, как сообщалось по крайней мере для одного кандидата вакцины (Pantazica и др., 2023).
5 Цели селекции и генетической инженерии для решения аспектов безопасности
Селекция и генетическая инженерия могут использоваться для снижения некоторых из вышеупомянутых рисков безопасности, и не только путем модификации гликанов. В частности, протеазы растений могут быть инактивированы для повышения целостности продукта (например, для минимизации деградации и агрегации), или они могут быть направленно экспрессированы для обеспечения точной обработки, такой как удаление лидерных последовательностей, как было продемонстрировано для трансформирующего фактора роста β1 (Goulet и др., 2012; Wilbers и др., 2016). Аналогично, ферментные каскады, синтезирующие токсичные соединения, могут быть прерваны, как было продемонстрировано на примере создания безникотинового табака (Schachtsiek и Stehle, 2019). Можно также учиться у других хост-систем и нокаутировать белки клеток-хозяев, которые трудно удалить в ходе нисходящей обработки (Chiu и др., 2017), включая, но не ограничиваясь токсичными или аллергенными, как обсуждалось выше. Эти и другие варианты, такие как использование шаперонов для обеспечения правильного фолдинга белка или модификации для предотвращения окисления, все из которых улучшают показатели видов-хозяев с точки зрения выхода, активности и безопасности продукта, были подробно рассмотрены в других работах (Buyel и др., 2021; Singh и др., 2021). Система CRISPR/Cas9 и ее регуляторные последствия были тщательно оценены в контексте PMF (Bortesi и Fischer, 2015; Eckerstorfer и др., 2019; Fiaz и др., 2021).
Важно, что такие улучшающие безопасность шаги генетической инженерии должны быть сбалансированы, например, с жизнеспособностью и продуктивностью полученной линии растений (клеток). Например, желательно нокаутировать протеазы, как обсуждалось выше, поскольку они могут вызывать (частичную) деградацию продукта PMF, тем самым снижая его активность (Donini и др., 2015; Mandal и др., 2016; Menzel и др., 2018). Однако протеазы выполняют важные биологические функции и могут быть необходимы для прорастания и роста растений (Martinez и др., 2019; van der Hoorn и Klemenčič, 2021), что важно для достижения высокого выхода продукта. Поэтому в некоторых случаях процессная инженерия, а не генетическая инженерия, может быть более подходящей.
Кроме того, одиночных нокаутов может быть недостаточно из-за избыточности в метаболических путях. Например, путь биосинтеза морфина разветвляется, когда достигает промежуточного тебаина, который может быть преобразован в морфин через кодеинон или морфинон (Ziegler и др., 2009; Onoyovwe и др., 2013). Поэтому по крайней мере один фермент в каждой ветви должен быть нокаутирован, чтобы полностью заблокировать синтез морфина. В этом контексте инактивация определенного ферментного каскада может привести к перенаправлению метаболического потока на другие метаболиты, которые также могут быть токсичными.
6 Заключение и перспективы
Растения, как и все другие биологические хозяева, представляют определенные риски для безопасности продукта из-за своих природных компонентов (например, токсичных метаболитов), условий культивирования и различий в PTMs. Важно контролировать эти риски при эксплуатации производственных процессов, соответствующих GMP, и внедрять соответствующую стратегию управления рисками. Такая стратегия должна быть сосредоточена на выявлении и приоритизации рисков на основе специфики конкретного процесса, например, белковых (Bracewell и др., 2015) или низкомолекулярных примесей (Luo и др., 2021). Поскольку приоритизация будет зависеть от свойств продукта и характеристик производственного процесса, нецелесообразно давать общие рекомендации, за исключением установленных концепций и эвристик, таких как анализ видов и критичности отказов или анализ рисков и критических контрольных точек, как изложено в руководящих принципах ICH Q9³. Однако такая оценка рисков могла бы значительно выиграть от структурированных и курируемых баз данных, которые агрегируют, например, информацию о фитотоксинах (Günthardt и др., 2018), поскольку знания и, в конечном счете, безопасность продукта будут расти по мере разработки все новых и новых процессов. Кроме того, общие знания о таких примесях и загрязнителях (т.е. исключая конкретные этапы процесса или условия) должны носить доконкурентный характер и, следовательно, могут быть раскрыты вовлеченными компаниями. Финансирование курирования и поддержания такой базы данных, скорее всего, станет узким местом.
При рассмотрении отдельных аспектов безопасности, обсуждаемых в Разделах 2–5, ни один из рисков не является достаточно серьезным, чтобы препятствовать использованию растений для применений PMF. Действительно, такие риски легко смягчаются путем внедрения установленных принципов управления рисками и разработки процесса. В целом, растения можно рассматривать как безопасные хост-системы для PMF, а выбор продовольственных или кормовых культур позволяет исключить многие риски, связанные с хозяевами, продуцирующими внутренне токсичные компоненты.
Сокращения
CEGS — контролируемые системы выращивания; CHO — яичник китайского хомячка; HCP — белок клетки-хозяина; PMF — молекулярный фарминг растений; PTM — посттрансляционная модификация.
Сноски
1. https://www.ema.europa.eu/en/documents/scientific-guideline/ich-q-7-good-manufacturing-practice-active-pharmaceutical-ingredients-step-5_en.pdf
2. https://www.who.int/observatories/global-observatory-on-health-research-and-development/analyses-and-syntheses/target-product-profile/links-to-who-tpps-and-ppcs
3. https://www.ema.europa.eu/en/documents/scientific-guideline/international-conference-harmonisation-technical-requirements-registration-pharmaceuticals-human-use_en-3.pdf
Ссылки
1. Abedi E., Pourmohammadi K. (2020). The effect of redox agents on conformation and structure characterization of gluten protein: An extensive review. Food Sci. Nutr. 8 (12), 6301–6319. [PMID: 33312518]. doi: 10.1002/fsn3.1937. CrossRef. Google Scholar.
2. Acamovic T., Brooker J. D. (2005). Biochemistry of plant secondary metabolites and their effects in animals. Proc. Nutr. Soc. 64 (3), 403–412. [PMID: 16048675]. doi: 10.1079/PNS2005449. CrossRef. Google Scholar.
3. Aida Y., Pabst M. J. (1990). Removal of endotoxin from protein solutions by phase-separation using Triton X-114. J. Immunol. Methods 132 (2), 191–195. doi: 10.1016/0022-1759(90)90029-U. CrossRef. Google Scholar.
4. Akpek E. K., Gottsch J. D. (2003). Immune defense at the ocular surface. Eye (Lond) 17 (8), 949–956. [PMID: 14631402]. doi: 10.1038/sj.eye.6700617. CrossRef. Google Scholar.
5. Manning L. (2018). Food & drink - good manufacturing practice: A guide to its responsible management. Seventh edition. Oxford, London: Wiley; Institute of Food Science and Technology. Google Scholar.
6. Anspach F. B. (2001). Endotoxin removal by affinity sorbents. J. Biochem. Biophysical Methods 49 (1-3), 665–681. [PMID: 11694310]. doi: 10.1016/S0165-022X(01)00228-7. CrossRef. Google Scholar.
7. Arcalis E., Ibl V., Peters J., Melnik S., Stoger E. (2014). The dynamic behavior of storage organelles in developing cereal seeds and its impact on the production of recombinant proteins. Front. plant Sci. 5 (439), 439–512. [PMID: 25232360]. doi: 10.3389/fpls.2014.00439. CrossRef. Google Scholar.
8. Arfi Z. A., Hellwig S., Drossard J., Fischer R., Buyel J. F. (2016). Polyclonal antibodies for specific detection of tobacco host cell proteins can be efficiently generated following RuBisCO depletion and the removal of endotoxins. Biotechnol. J. 11 (4), 507–518. doi: 10.1002/biot.201500271. CrossRef. Google Scholar.
9. Argentine M. D., Owens P. K., Olsen B. A. (2007). Strategies for the investigation and control of process-related impurities in drug substances. Adv. Drug Deliv. Rev. 59 (1), 12–28. [PMID: 17189658]. doi: 10.1016/j.addr.2006.10.005. CrossRef. Google Scholar.
10. Bagnis R., Chanteau S., Chungue E., Hurtel J. M., Yasumoto T., Inoue A. (1980). Origins of ciguatera fish poisoning: A new dinoflagellate, Gambierdiscus toxicus adachi and fukuyo, definitively involved as a causal agent. Toxicon 18 (2), 199–208. [PMID: 7190330]. doi: 10.1016/0041-0101(80)90074-4. CrossRef. Google Scholar.
11. Balique F., Colson P., Raoult D. (2012). Tobacco mosaic virus in cigarettes and saliva of smokers. J. Clin. Virol. 55 (4), 374–376. [PMID: 22959216]. doi: 10.1016/j.jcv.2012.08.012. CrossRef. Google Scholar.
12. Bardor M., Faveeuw C., Fitchette A. C. (2003). Immunoreactivity in mammals of two typical plant glyco-epitopes, core alpha(1,3)-fucose and core xylose. Glycobiology 13 (6), 427–434. doi: 10.1093/glycob/cwg024. CrossRef. Google Scholar.
13. Barzee T. J., El Mashad H. M., Cao L., Chio A., Pan Z., Zhang R. (2022). Cell‐cultivated food production and processing: A review. Food Bioeng. 1 (1), 4–25. doi: 10.1002/fbe2.12009. CrossRef. Google Scholar.
14. Bassene H., Niang E. H. A., Fenollar F., Doucoure S., Faye O., Raoult D. et al (2020). Role of plants in the transmission of Asaia sp., which potentially inhibit the Plasmodium sporogenic cycle in Anopheles mosquitoes. Sci. Rep. 10 (1), 7144. [PMID: 32346047]. doi: 10.1038/s41598-020-64163-5. CrossRef. Google Scholar.
15. Bernau C. R., Knödler M., Emonts J., Jäpel R. C., Buyel J. F. (2022). The use of predictive models to develop chromatography-based purification processes. Front. Bioeng. Biotechnol. 10, 1009102. doi: 10.3389/fbioe.2022.1009102. CrossRef. Google Scholar.
16. Bianco G., Russo R., Marzocco S., Velotto S., Autore G., Severino L. (2012). Modulation of macrophage activity by aflatoxins B1 and B2 and their metabolites aflatoxins M1 and M2. Toxicon 59 (6), 644–650. [PMID: 22402176]. doi: 10.1016/j.toxicon.2012.02.010. CrossRef. Google Scholar.
17. Biesiekierski J. R. (2017). What is gluten? J. Gastroenterol. Hepatol. 32 (1), 78–81. [PMID: 28244676]. doi: 10.1111/jgh.13703. CrossRef. Google Scholar.
18. Bonamonte D., Vestita M., Filoni A., Mastrolonardo M., Angelini G., Foti C. (2016). Tobacco-induced contact dermatitis. Eur. J. Dermatol 26 (3), 223–231. [PMID: 27020490]. doi: 10.1684/ejd.2016.2771. CrossRef. Google Scholar.
19. Bortesi L., Fischer R. (2015). The CRISPR/Cas9 system for plant genome editing and beyond. Biotechnol. Adv. 33 (1), 41–52. [PMID: 25536441]. doi: 10.1016/j.biotechadv.2014.12.006. CrossRef. Google Scholar.
20. Bousbia S., Papazian L., La Scola B., Raoult D. (2010). Detection of plant DNA in the bronchoalveolar lavage of patients with ventilator-associated pneumonia. PLoS ONE 5 (6), e11298. [PMID: 20585574]. doi: 10.1371/journal.pone.0011298. CrossRef. Google Scholar.
21. Bracewell D. G., Francis R., Smales C. M. (2015). The future of host cell protein (HCP) identification during process development and manufacturing linked to a risk-based management for their control. Biotechnol. Bioeng. 112 (9), 1727–1737. doi: 10.1002/bit.25628. CrossRef. Google Scholar.
22. Buyel J. F., Bautista J. A., Fischer R., Yusibov V. M. (2012). Extraction, purification and characterization of the plant-produced HPV16 subunit vaccine candidate E7 GGG. J. Chromatogr. B 880 (1), 19–26. [PMID: 22134037]. doi: 10.1016/j.jchromb.2011.11.010. CrossRef. Google Scholar.
23. Buyel J. F., Buyel J. J., Haase C., Fischer R. (2015b). The impact of Pseudomonas syringae type III effectors on transient protein expression in tobacco. Plant Biol. (Stuttg) 17 (2), 484–492. [PMID: 25243954]. doi: 10.1111/plb.12264. CrossRef. Google Scholar.
24. Buyel J. F., Fischer R. (2014d). A juice extractor can simplify the downstream processing of plant-derived biopharmaceutical proteins compared to blade-based homogenizers. Process Biochem. 50 (5), 859–866. doi: 10.1016/j.procbio.2015.02.017. CrossRef. Google Scholar.
25. Buyel J. F., Fischer R. (2014c). Flocculation increases the efficacy of depth filtration during the downstream processing of recombinant pharmaceutical proteins produced in tobacco. Plant Biotechnol. J. 12 (2), 240–252. [PMID: 24165151]. doi: 10.1111/pbi.12132. CrossRef. Google Scholar.
26. Buyel J. F., Fischer R. (2014a). Generic chromatography-based purification strategies accelerate the development of downstream processes for biopharmaceutical proteins produced in plants. Biotechnol. J. 9 (4), 566–577. [PMID: 24478119]. doi: 10.1002/biot.201300548. CrossRef. Google Scholar.
27. Buyel J. F., Fischer R. (2014e). Scale-down models to optimize a filter train for the downstream purification of recombinant pharmaceutical proteins produced in tobacco leaves. Biotechnol. J. 9 (3), 415–425. [PMID: 24323869]. doi: 10.1002/biot.201300369. CrossRef. Google Scholar.
28. Buyel J. F., Fischer R. (2014b). Synthetic polymers are more effective than natural flocculants for the clarification of tobacco leaf extracts. J. Biotechnol. 195, 37–42. doi: 10.1016/j.jbiotec.2014.12.018. CrossRef. Google Scholar.
29. Buyel J. F., Gruchow H. M., Boes A., Fischer R. (2014a). Rational design of a host cell protein heat precipitation step simplifies the subsequent purification of recombinant proteins from tobacco. Biochem. Eng. J. 88 (15), 162–170. doi: 10.1016/j.bej.2014.04.015. CrossRef. Google Scholar.
30. Buyel J. F., Hubbuch J., Fischer R. (2016). Comparison of tobacco host cell protein removal methods by blanching intact plants or by heat treatment of extracts. Jove-J Vis. Exp. e54343 (114), 54343. doi: 10.3791/54343. CrossRef. Google Scholar.
31. Buyel J. F., Opdensteinen P., Fischer R. (2014b). Cellulose-based filter aids increase the capacity of depth filters during the downstream processing of plant-derived biopharmaceutical proteins. Biotechnol. J. 10 (4), 584–591. doi: 10.1002/biot.201400611. CrossRef. Google Scholar.
32. Buyel J. F. (2018). Plants as sources of natural and recombinant anti-cancer agents. Biotechnol. Adv. 36 (2), 506–520. [PMID: 29408560]. doi: 10.1016/j.biotechadv.2018.02.002. CrossRef. Google Scholar.
33. Buyel J. F., Stöger E., Bortesi L. (2021). Targeted genome editing of plants and plant cells for biomanufacturing. Transgenic Res. 30 (4), 401–426. [PMID: 33646510]. doi: 10.1007/s11248-021-00236-z. CrossRef. Google Scholar.
34. Buyel J. F., Twyman R. M., Fischer R. (2015a). Extraction and downstream processing of plant-derived recombinant proteins. Biotechnol. Adv. 33 (6), 902–913. [PMID: 25922318]. doi: 10.1016/j.biotechadv.2015.04.010. CrossRef. Google Scholar.
35. Buyel J. F., Twyman R. M., Fischer R. (2017). Very-large-scale production of antibodies in plants: The biologization of manufacturing. Biotechnol. Adv. 35 (4), 458–465. [PMID: 28347720]. doi: 10.1016/j.biotechadv.2017.03.011. CrossRef. Google Scholar.
36. Cabanillas B. (2020). Gluten-related disorders: Celiac disease, wheat allergy, and nonceliac gluten sensitivity. Crit. Rev. Food Sci. Nutr. 60 (15), 2606–2621. [PMID: 31402697]. doi: 10.1080/10408398.2019.1651689. CrossRef. Google Scholar.
37. Carter-Franklin J. N., Victa C., McDonald P., Fahrner R. (2007). Fragments of protein A eluted during protein A affinity chromatography. J. Chromatogr. A 1163 (1-2), 105–111. [PMID: 17643441]. doi: 10.1016/j.chroma.2007.06.012. CrossRef. Google Scholar.
38. Castilho A., Neumann L., Gattinger P., Strasser R., Vorauer-Uhl K., Sterovsky T. et al (2013). Generation of biologically active multi-sialylated recombinant human EPOFc in plants. PloS one 8 (1), e54836. [PMID: 23372778]. doi: 10.1371/journal.pone.0054836. CrossRef. Google Scholar.
39. Chanquia S. N., Vernet G., Kara S. (2022). Photobioreactors for cultivation and synthesis: Specifications, challenges, and perspectives. Eng. Life Sci. 22 (12), 712–724. [PMID: 36514531]. doi: 10.1002/elsc.202100070. CrossRef. Google Scholar.
40. Chiu J., Valente K. N., Levy N. E., Min L., Lenhoff A. M., Lee K. H. (2017). Knockout of a difficult-to-remove CHO host cell protein, lipoprotein lipase, for improved polysorbate stability in monoclonal antibody formulations. Biotechnol. Bioeng. 114 (5), 1006–1015. [PMID: 27943242]. doi: 10.1002/bit.26237. CrossRef. Google Scholar.
41. Cohen I. S., Day A. S., Shaoul R. (2019). Gluten in celiac disease-more or less? Rambam Maimonides Med. J. 10 (1), e0007. [PMID: 30720425]. doi: 10.5041/RMMJ.10360. CrossRef. Google Scholar.
42. Commandeur U., Twyman R. M. (2005). "Biosafety aspects of molecular farming in plants," in Molecular farming (Weinheim, Germany: Wiley-VCH Verlag GmbH & Co. KGaA), 251–266. Google Scholar.
43. Compant S., Samad A., Faist H., Sessitsch A. (2019). A review on the plant microbiome: Ecology, functions, and emerging trends in microbial application. J. Adv. Res. 19, 29–37. [PMID: 31341667]. doi: 10.1016/j.jare.2019.03.004. CrossRef. Google Scholar.
44. Crosby D. G., Aharonson N. (1967). The structure of carotatoxin, a natural toxicant from carrot. Tetrahedron 23 (1), 465–472. [PMID: 6037290]. doi: 10.1016/S0040-4020(01)83330-5. CrossRef. Google Scholar.
45. Cuadros-Rodríguez L., Lazúen-Muros M., Ruiz-Samblás C., Navas-Iglesias N. (2020). Leachables from plastic materials in contact with drugs. State of the art and review of current analytical approaches. Int. J. Pharm. 583, 119332. [PMID: 32360549]. doi: 10.1016/j.ijpharm.2020.119332. CrossRef. Google Scholar.
46. Cummings J. F., Guerrero M. L., Moon J. E., Waterman P., Nielsen R. K., Jefferson S. et al (2014). Safety and immunogenicity of a plant-produced recombinant monomer hemagglutinin-based influenza vaccine derived from influenza A (H1N1)pdm09 virus: A phase 1 dose-escalation study in healthy adults. Vaccine 32 (19), 2251–2259. doi: 10.1016/j.vaccine.2013.10.017. CrossRef. Google Scholar.
47. D'Angelo E. (2013). "Methods for determining toxic organic compounds in wetlands," in Methods in biogeochemistry of wetlands. Editors DeLaune R., Reddy K., Richardson C. (Madison, WI, USA: John Wiley & Sons, Inc), 815–839. Google Scholar.
48. Davey R. T., Dodd L., Proschan M. A. (2016). A randomized, controlled trial of ZMapp for ebola virus infection. N. Engl. J. Med. 375 (15), 1448–1456. [PMID: 27732819]. doi: 10.1056/NEJMoa1604330. CrossRef. Google Scholar.
49. Dessaux Y., Faure D. (2018). Niche construction and exploitation by Agrobacterium: How to survive and face competition in soil and plant habitats. Curr. Top. Microbiol. 418, 55–86. [PMID: 29556826]. doi: 10.1007/82_2018_83. CrossRef. Google Scholar.
50. Donini M., Lombardi R., Lonoce C., Di Carli M., Marusic C., Morea V. et al (2015). Antibody proteolysis: A common picture emerging from plants. Bioengineered 6 (5), 299–302. [PMID: 26186119]. doi: 10.1080/21655979.2015.1067740. CrossRef. Google Scholar.
51. Dortant P. M., Claassen I. J., van Kreyl C. F., van Steenis G., Wester P. W. (1997). Risk assessment on the carcinogenic potential of hybridoma cell DNA: Implications for residual contaminating cellular DNA in biological products. Biologicals 25 (4), 381–390. [PMID: 9467034]. doi: 10.1006/biol.1997.0114. CrossRef. Google Scholar.
52. Drakopoulos D., Sulyok M., Krska R., Logrieco A. F., Vogelgsang S. (2021). Raised concerns about the safety of barley grains and straw: A Swiss survey reveals a high diversity of mycotoxins and other fungal metabolites. Food control. 125, 107919. doi: 10.1016/j.foodcont.2021.107919. CrossRef. Google Scholar.
53. Duvick J. (2001). Prospects for reducing fumonisin contamination of maize through genetic modification. Environ. Health Persp 109, 337–342. [PMID: 11359705]. doi: 10.1289/ehp.01109s2337. CrossRef. Google Scholar.
54. Eckerstorfer M. F., Engelhard M., Heissenberger A., Simon S., Teichmann H. (2019). Plants developed by new genetic modification techniques-comparison of existing regulatory frameworks in the EU and non-EU countries. Front. Bioeng. Biotechnol. 7, 26. [PMID: 30838207]. doi: 10.3389/fbioe.2019.00026. CrossRef. Google Scholar.
55. Eidenberger L., Kogelmann B., Steinkellner H. (2023). Plant-based biopharmaceutical engineering. Nat. Rev. Bioeng. 1, 426–439. doi: 10.1038/s44222-023-00044-6. CrossRef. Google Scholar.
56. el-Maghraby O. M., Abdel-Sater M. A. (1993). Mycoflora and natural occurrence of mycotoxins in tobacco from cigarettes in Egypt. Zentralbl Mikrobiol. 148 (4), 253–264. [PMID: 8368025]. doi: 10.1016/s0232-4393(11)80101-0. CrossRef. Google Scholar.
57. Elli L., Bascuñán K., Di Lernia L., Bardella M. T., Doneda L., Soldati L. et al (2020). Safety of occasional ingestion of gluten in patients with celiac disease: A real-life study. BMC Med. 18 (1), 42. [PMID: 32172690]. doi: 10.1186/s12916-020-1511-6. CrossRef. Google Scholar.
58. Farhangi H., Mozafari V., Roosta H. R., Shirani H., Farhangi M. (2023). Optimizing growth conditions in vertical farming: Enhancing lettuce and basil cultivation through the application of the taguchi method. Sci. Rep. 13 (1), 6717. [PMID: 37185295]. doi: 10.1038/s41598-023-33855-z. CrossRef. Google Scholar.
59. Fiaz S., Aslam Khan S., Anis G. B. (2021). Application of genome engineering methods for quality improvement in important crops. In: Upadhyay S., Genome engineering for crop improvement. 1st edition. [Erscheinungsort nicht ermittelbar], Boston, MA: Wiley, 43–68. Google Scholar.
60. Fletcher J., Leach J. E., Eversole K., Tauxe R. (2013). Human pathogens on plants: Designing a multidisciplinary strategy for research. Phytopathology 103 (4), 306–315. [PMID: 23406434]. doi: 10.1094/phyto-09-12-0236-ia. CrossRef. Google Scholar.
61. Florea S., Panaccione D. G., Schardl C. L. (2017). Ergot alkaloids of the family clavicipitaceae. Phytopathology 107 (5), 504–518. [PMID: 28168931]. doi: 10.1094/phyto-12-16-0435-rvw. CrossRef. Google Scholar.
62. Fraser P. D., Aharoni A., Hall R. D., Huang S., Giovannoni J. J., Sonnewald U. et al (2020). Metabolomics should be deployed in the identification and characterization of gene-edited crops. Plant J. 102 (5), 897–902. [PMID: 31923321]. doi: 10.1111/tpj.14679. CrossRef. Google Scholar.
63. Friedman M. A., Fernandez M., Backer L. C., Dickey R., Bernstein J., Schrank K. et al (2017). An updated review of ciguatera fish poisoning: Clinical, epidemiological, environmental, and public health management. Mar. Drugs 15 (3), 72. [PMID: 28335428]. doi: 10.3390/md15030072. CrossRef. Google Scholar.
64. Fu H., Machado P. A., Hahm T. S., Kratochvil R. J., Wei C. I., Lo Y. M. (2010). Recovery of nicotine-free proteins from tobacco leaves using phosphate buffer system under controlled conditions. Bioresour. Technol. 101 (6), 2034–2042. doi: 10.1016/j.biortech.2009.10.045. CrossRef. Google Scholar.
65. Fusetani N., Kem W. (Editors) (2009). Marine toxins as research tools (Berlin: Springer). Google Scholar.
66. Ganesan P. K., Kulchar R. J., Kaznica P., Montoya-Lopez R., Green B. J., Streatfield S. J. et al (2023). Optimization of biomass and target protein yield for Phase III clinical trial to evaluate Angiotensin Converting Enzyme 2 expressed in lettuce chloroplasts to reduce SARS-CoV-2 infection and transmission. Plant Biotechnol. J. 21 (2), 244–246. [PMID: 36314106]. doi: 10.1111/pbi.13954. CrossRef. Google Scholar.
67. Geng Y., Cao G., Wang L., Wang S. (2019). Effects of equal chemical fertilizer substitutions with organic manure on yield, dry matter, and nitrogen uptake of spring maize and soil nitrogen distribution. PLoS ONE 14 (7), e0219512. [PMID: 31287845]. doi: 10.1371/journal.pone.0219512. CrossRef. Google Scholar.
68. Gengenbach B. B., Keil L. L., Opdensteinen P., Müschen C. R., Melmer G., Lentzen H. et al (2019). Comparison of microbial and transient expression (tobacco plants and plant-cell packs) for the production and purification of the anticancer mistletoe lectin viscumin. Biotechnol. Bioeng. 116 (9), 2236–2249. [PMID: 31140580]. doi: 10.1002/bit.27076. CrossRef. Google Scholar.
69. Gleadow R. M., Møller B. L. (2014). Cyanogenic glycosides: Synthesis, physiology, and phenotypic plasticity. Annu. Rev. Plant Biol. 65, 155–185. [PMID: 24579992]. doi: 10.1146/annurev-arplant-050213-040027. CrossRef. Google Scholar.
70. Gleadow R. M., Woodrow I. E. (2000). Temporal and spatial variation in cyanogenic glycosides in Eucalyptus cladocalyx. Tree Physiol. 20 (9), 591–598. [PMID: 12651423]. doi: 10.1093/treephys/20.9.591. CrossRef. Google Scholar.
71. Goulet C., Khalf M., Sainsbury F., D'Aoust M. A., Michaud D. (2012). A protease activity-depleted environment for heterologous proteins migrating towards the leaf cell apoplast. Plant Biotechnol. J. 10 (1), 83–94. doi: 10.1111/j.1467-7652.2011.00643.x. CrossRef. Google Scholar.
72. Grabowski G. A., Golembo M., Shaaltiel Y. (2014). Taliglucerase alfa: An enzyme replacement therapy using plant cell expression technology. Mol. Genet. Metab. 112 (1), 1–8. doi: 10.1016/j.ymgme.2014.02.011. CrossRef. Google Scholar.
73. Green B. T., Lee S. T., Welch K. D., Panter K. E. (2013). Plant alkaloids that cause developmental defects through the disruption of cholinergic neurotransmission. Birth Defects Res. C Embryo Today 99 (4), 235–246. [PMID: 24339035]. doi: 10.1002/bdrc.21049. CrossRef. Google Scholar.
74. Griffiths M. R., Strobel B. W., Hama J. R., Cedergreen N. (2021). Toxicity and risk of plant-produced alkaloids to Daphnia magna. Environ. Sci. Eur. 33 (1), 10. doi: 10.1186/s12302-020-00452-0. CrossRef. Google Scholar.
75. Grosse-Holz F., Madeira L., Zahid M. A., Songer M., Kourelis J., Fesenko M. et al (2018). Three unrelated protease inhibitors enhance accumulation of pharmaceutical recombinant proteins in Nicotiana benthamiana. Plant Biotechnol. J. 16 (10), 1797–1810. [PMID: 29509983]. doi: 10.1111/pbi.12916. CrossRef. Google Scholar.
76. Gruenberg D. A., Shaker M. S. (2011). An update on influenza vaccination in patients with egg allergy. Curr. Opin. Pediatr. 23 (5), 566–572. [PMID: 21881505]. doi: 10.1097/MOP.0b013e32834ac7a3. CrossRef. Google Scholar.
77. Günthardt B. F., Hollender J., Hungerbühler K., Scheringer M., Bucheli T. D. (2018). Comprehensive toxic plants-phytotoxins database and its application in assessing aquatic micropollution potential. J. Agric. Food Chem. 66 (29), 7577–7588. [PMID: 29944838]. doi: 10.1021/acs.jafc.8b01639. CrossRef. Google Scholar.
78. Hahn-Löbmann S., Stephan A., Schulz S., Schneider T., Shaverskyi A., Tusé D. et al (2019). Colicins and salmocins - new classes of plant-made non-antibiotic food antibacterials. Front. Plant Sci. 10, 437. [PMID: 31024601]. doi: 10.3389/fpls.2019.00437. CrossRef. Google Scholar.
79. Hartz C., Del San Miguel-Moncín M. M., Cisteró-Bahíma A., Fötisch K., Metzner K. J., Fortunato D. et al (2007). Molecular characterisation of Lac s 1, the major allergen from lettuce (Lactuca sativa). Mol. Immunol. 44 (11), 2820–2830. [PMID: 17349693]. doi: 10.1016/j.molimm.2007.01.030. CrossRef. Google Scholar.
80. Harwanto D., Negara B. F. S. P., Tirtawijaya G., Meinita M. D. N., Choi J-S. (2022). Evaluation of toxicity of crude phlorotannins and phloroglucinol using different model organisms. Toxins (Basel) 14 (5), 312. [PMID: 35622559]. doi: 10.3390/toxins14050312. CrossRef. Google Scholar.
81. Hayashi S., Watanabe M., Kobayashi M., Tohge T., Hashimoto T., Shoji T. (2020). Genetic manipulation of transcriptional regulators alters nicotine biosynthesis in tobacco. Plant Cell Physiol. 61 (6), 1041–1053. [PMID: 32191315]. doi: 10.1093/pcp/pcaa036. CrossRef. Google Scholar.
82. Hendrich J. M., Wangorsch A., Rödel K., Jacob T., Mahler V., Wöhrl B. M. (2023). Allergenicity and IgE recognition of new dau c 1 allergens from carrot. Mol. Nutr. Food Res. 67 (3), e2200421. [PMID: 36458641]. doi: 10.1002/mnfr.202200421. CrossRef. Google Scholar.
83. Hendrikx J. J. M. A., Haanen J. B. A. G., Voest E. E., Schellens J. H. M., Huitema A. D. R., Beijnen J. H. (2017). Fixed dosing of monoclonal antibodies in oncology. Oncologist 22 (10), 1212–1221. [PMID: 28754722]. doi: 10.1634/theoncologist.2017-0167. CrossRef. Google Scholar.
84. Henry J. B., Vann M. C., Lewis R. S. (2019). Agronomic practices affecting nicotine concentration in flue-cured tobacco: A review. Agron. J. 111 (6), 3067–3075. doi: 10.2134/agronj2019.04.0268. CrossRef. Google Scholar.
85. Herrera-Estrella L., Simpson J., Martinez-Trujillo M. (2005). Transgenic plants: An historical perspective. Methods Mol. Biol. 286, 003–032. [PMID: 15310910]. doi: 10.1385/1-59259-827-7:003. CrossRef. Google Scholar.
86. Hien D. F. D. S., Dabiré K. R., Roche B., Diabaté A., Yerbanga R. S., Cohuet A. et al (2016). Plant-Mediated effects on mosquito capacity to transmit human malaria. PLoS Pathog. 12 (8), e1005773. [PMID: 27490374]. doi: 10.1371/journal.ppat.1005773. CrossRef. Google Scholar.
87. Hirayama C., Sakata M. (2002). Chromatographic removal of endotoxin from protein solutions by polymer particles. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 781 (1-2), 419–432. [PMID: 12450672]. doi: 10.1016/S1570-0232(02)00430-0. CrossRef. Google Scholar.
88. Hischenhuber C., Crevel R., Jarry B., Maki M., Moneret-Vautrin D. A., Romano A. et al (2006). Review article: Safe amounts of gluten for patients with wheat allergy or coeliac disease. Aliment. Pharmacol. Ther. 23 (5), 559–575. [PMID: 16480395]. doi: 10.1111/j.1365-2036.2006.02768.x. CrossRef. Google Scholar.
89. Hofbauer W. K. (2021). Toxic or otherwise harmful algae and the built environment. Toxins (Basel) 13 (7), 465. [PMID: 34209446]. doi: 10.3390/toxins13070465. CrossRef. Google Scholar.
90. Holland T., Blessing D., Hellwig S., Sack M. (2013). The in-line measurement of plant cell biomass using radio frequency impedance spectroscopy as a component of process analytical technology. Biotechnol. J. 8 (10), 1231–1240. doi: 10.1002/biot.201300125. CrossRef. Google Scholar.
91. Holland T., Buyel J. F. (2017). "Bioreactor-based production of glycoproteins in plant cell suspension cultures," in Recombinant glycoprotein production. Editors Picanço e Castro, V., Swiech K. (New York: Springer), 129–146. Google Scholar.
92. Holtz B. R., Berquist B. R., Bennett L. D., Kommineni V. J., Munigunti R. K., White E. L. et al (2015). Commercial-scale biotherapeutics manufacturing facility for plant-made pharmaceuticals. Plant Biotechnol. J. 13 (8), 1180–1190. [PMID: 26387511]. doi: 10.1111/pbi.12469. CrossRef. Google Scholar.
93. Houdelet M., Galinski A., Holland T., Wenzel K., Schillberg S., Buyel J. F. (2017). Animal component-free Agrobacterium tumefaciens cultivation media for better GMP-compliance increases biomass yield and pharmaceutical protein expression in Nicotiana benthamiana. Biotechnol. J. 12 (4), 1600721. [PMID: 28221723]. doi: 10.1002/biot.201600721. CrossRef. Google Scholar.
94. Huang T-K., Puchta H. (2021). Novel CRISPR/cas applications in plants: From prime editing to chromosome engineering. Transgenic Res. 30 (4), 529–549. [PMID: 33646511]. doi: 10.1007/s11248-021-00238-x. CrossRef. Google Scholar.
95. Huang T. K., McDonald K. A. (2012). Bioreactor systems for in vitro production of foreign proteins using plant cell cultures. Biotechnol. Adv. 30 (2), 398–409. [PMID: 21846499]. doi: 10.1016/j.biotechadv.2011.07.016. CrossRef. Google Scholar.
96. Huebbers J. W., Buyel J. F. (2021). On the verge of the market - plant factories for the automated and standardized production of biopharmaceuticals. Biotechnol. Adv. 46, 107681. [PMID: 33326816]. doi: 10.1016/j.biotechadv.2020.107681. CrossRef. Google Scholar.
97. Hulvová H., Galuszka P., Frébortová J., Frébort I. (2013). Parasitic fungus Claviceps as a source for biotechnological production of ergot alkaloids. Biotechnol. Adv. 31 (1), 79–89. [PMID: 22261014]. doi: 10.1016/j.biotechadv.2012.01.005. CrossRef. Google Scholar.
98. Hundleby P. A. C., D'Aoust M-A., Finkle C., Atkins J., Twyman R. M. (2022). Regulation of molecular farming products. Methods Mol. Biol. 2480, 313–333. [PMID: 35616870]. doi: 10.1007/978-1-0716-2241-4_17. CrossRef. Google Scholar.
99. Hwang H. S., Kim B. S., Park H., Park H-Y., Choi H-D., Kim H. H. (2014). Type and branched pattern of N-glycans and their structural effect on the chicken egg allergen ovotransferrin: A comparison with ovomucoid. Glycoconj. J. 31 (1), 41–50. [PMID: 24014058]. doi: 10.1007/s10719-013-9498-2. CrossRef. Google Scholar.
100. Hwang Y-C., Lu R-M., Su S-C., Chiang P. Y., Ko S. H., Ke F. Y. et al (2022). Monoclonal antibodies for COVID-19 therapy and SARS-CoV-2 detection. J. Biomed. Sci. 29 (1), 1. [PMID: 34983527]. doi: 10.1186/s12929-021-00784-w. CrossRef. Google Scholar.
101. Ito K. (2015). Grain and legume allergy. Chem. Immunol. Allergy 101, 145–151. [PMID: 26022874]. doi: 10.1159/000375468. CrossRef. Google Scholar.
102. James J. M., Zeiger R. S., Lester M. R., Fasano M. B., Gern J. E., Mansfield L. E. et al (1998). Safe administration of influenza vaccine to patients with egg allergy. J. Pediatr. 133 (5), 624–628. [PMID: 9821418]. doi: 10.1016/s0022-3476(98)70101-5. CrossRef. Google Scholar.
103. Jang H., Kim H-S., Moon S-C., Lee Y. R., Yu K. Y., Lee B. K. et al (2009). Effects of protein concentration and detergent on endotoxin reduction by ultrafiltration. Biochem. Mol. Biol. Rep. 42 (7), 462–466. [PMID: 19643046]. doi: 10.5483/bmbrep.2009.42.7.462. CrossRef. Google Scholar.
104. Jansing J., Sack M., Augustine S. M., Fischer R., Bortesi L. (2018). CRISPR/Cas9-mediated knockout of six glycosyltransferase genes in Nicotiana benthamiana for the production of recombinant proteins lacking beta-1,2-xylose and core alpha-1,3-fucose. Plant Biotechnol. J. 17, 350–361. [PMID: 29969180]. doi: 10.1111/pbi.12981. CrossRef. Google Scholar.
105. Jenke D. (2002). Extractable/leachable substances from plastic materials used as pharmaceutical product containers/devices. Pda J. Pharm. Sci. Tech. 56 (6), 332–371. [PMID: 12497770]. Google Scholar.
106. Jiang Q. L., Zhang S., Tian M., Xie T., Chen D. Y. (2015). Plant lectins, from ancient sugar-binding proteins to emerging anti-cancer drugs in apoptosis and autophagy. Cell Prolif. 48 (1), 17–28. [PMID: 25488051]. doi: 10.1111/cpr.12155. CrossRef. Google Scholar.
107. Jin C., Altmann F., Strasser R., Mach L., Schahs M., Kunert R. et al (2008). A plant-derived human monoclonal antibody induces an anti-carbohydrate immune response in rabbits. Glycobiology 18 (3), 235–241. [PMID: 18203810]. doi: 10.1093/glycob/cwm137. CrossRef. Google Scholar.
108. Jin J., Tarrant R. D., Bolam E. J., Angell-Manning P., Soegaard M., Pattinson D. J. et al (2018). Production, quality control, stability, and potency of cGMP-produced Plasmodium falciparum RH5.1 protein vaccine expressed in Drosophila S2 cells. NPJ Vaccines 3, 32. [PMID: 30131879]. doi: 10.1038/s41541-018-0071-7. CrossRef. Google Scholar.
109. Jones M., Palackal N., Wang F., Gaza-Bulseco G., Hurkmans K., Zhao Y. et al (2021). High-risk" host cell proteins (HCPs): A multi-company collaborative view. Biotechnol. Bioeng. 118 (8), 2870–2885. [PMID: 33930190]. doi: 10.1002/bit.27808. CrossRef. Google Scholar.
110. Joo Y., Ok H. E., Kim J., Lee S. Y., Jang S. K., Park K. H. et al (2019). A statistical model for determining zearalenone contamination in rice (Oryza sativa L) at harvest and its prediction under different climate change scenarios in South Korea. Appl. Biol. Chem. 62 (1), 38. doi: 10.1186/s13765-019-0447-z. CrossRef. Google Scholar.
111. Jurjevec A., Brocard C., Striedner G., Cserjan-Puschmann M., Hahn R. (2023). Polyethyleneimine efficiently extracts recombinant cytoplasmatic green fluorescent protein produced in Escherichia coli with high purity. J. Biotechnol. 371-372, 41–49. [PMID: 37285941]. doi: 10.1016/j.jbiotec.2023.06.001. CrossRef. Google Scholar.
112. Jutras P. V., Marusic C., Lonoce C., Deflers C., Goulet M. C., Benvenuto E. et al (2016). An accessory protease inhibitor to increase the yield and quality of a tumour-targeting mAb in nicotiana benthamiana leaves. PloS one 11 (11), e0167086. [PMID: 27893815]. doi: 10.1371/journal.pone.0167086. CrossRef. Google Scholar.
113. Kabir S. R., Nabi M. M., Haque A., Rokon U. Z., Mahmud Z. H., Reza M. A. (2013). Pea lectin inhibits growth of Ehrlich ascites carcinoma cells by inducing apoptosis and G2/M cell cycle arrest in vivo in mice. Phytomedicine 20 (14), 1288–1296. [PMID: 23867650]. doi: 10.1016/j.phymed.2013.06.010. CrossRef. Google Scholar.
114. Kaiser N., Douches D., Dhingra A., Glenn K. C., Herzig P. R., Stowe E. C. et al (2020). The role of conventional plant breeding in ensuring safe levels of naturally occurring toxins in food crops. Trends Food Sci. Technol. 100, 51–66. doi: 10.1016/j.tifs.2020.03.042. CrossRef. Google Scholar.
115. Kang S-M., Adhikari A., Bhatta D., Gam H. J., Gim M. J., Son J. I. et al (2022). Comparison of effects of chemical and food waste-derived fertilizers on the growth and nutrient content of lettuce (Lactuca sativa L). Resources 11 (2), 21. doi: 10.3390/resources11020021. CrossRef. Google Scholar.
116. Kaur G., Upadhyaya P., Chawla P. (2023). Comparative analysis of IoT-based controlled environment and uncontrolled environment plant growth monitoring system for hydroponic indoor vertical farm. Environ. Res. 222, 115313. [PMID: 36709025]. doi: 10.1016/j.envres.2023.115313. CrossRef. Google Scholar.
117. Kawka K., Wilton A. N., Madadkar P., Medina M. F. C., Lichty B. D., Ghosh R. et al (2021). Integrated development of enzymatic DNA digestion and membrane chromatography processes for the purification of therapeutic adenoviruses. Sep. Purif. Technol. 254, 117503. doi: 10.1016/j.seppur.2020.117503. CrossRef. Google Scholar.
118. Keeler R. F., Crowe M. W., Lambert E. A. (1984). Teratogenicity in swine of the tobacco alkaloid anabasine isolated from Nicotiana glauca. Teratology 30 (1), 61–69. [PMID: 6484853]. doi: 10.1002/tera.1420300109. CrossRef. Google Scholar.
119. Kheawfu K., Kaewpinta A., Chanmahasathien W., Rachtanapun P., Jantrawut P. (2021). Extraction of nicotine from tobacco leaves and development of fast dissolving nicotine extract film. Membr. (Basel) 11 (6), 403. [PMID: 34071396]. doi: 10.3390/membranes11060403. CrossRef. Google Scholar.
120. Kim J-S., Yoon S-J., Park Y-J., Kim S-Y., Ryu C-M. (2020). Crossing the kingdom border: Human diseases caused by plant pathogens. Environ. Microbiol. 22 (7), 2485–2495. [PMID: 32307848]. doi: 10.1111/1462-2920.15028. CrossRef. Google Scholar.
121. Kłapeć T., Wójcik-Fatla A., Farian E., Kowalczyk K., Cholewa G., Cholewa A. et al (2021). Levels of filamentous fungi and selected mycotoxins in leafy and fruit vegetables and analysis of their potential health risk for consumers. Ann. Agric. Environ. Med. 28 (4), 585–594. [PMID: 34969215]. doi: 10.26444/aaem/143031. CrossRef. Google Scholar.
122. Knödler M., Opdensteinen P., Sankaranarayanan R. A., Morgenroth A., Buhl E. M., Mottaghy F. M. et al (2023b). Simple plant-based production and purification of the assembled human ferritin heavy chain as a nanocarrier for tumor-targeted drug delivery and bioimaging in cancer therapy. Biotechnol. Bioeng. 120 (4), 1038–1054. [PMID: 36539373]. doi: 10.1002/bit.28312. CrossRef. Google Scholar.
123. Knödler M., Reunious P. W., Buyel J. F. (2023a). Risk assessment and bioburden evaluation of Agrobacterium tumefaciens-mediated transient protein expression in plants using the CaMV35S promoter. Bmc Biotechnol. 23 (1), 14. [PMID: 37286972]. doi: 10.1186/s12896-023-00782-w. CrossRef. Google Scholar.
124. Knödler M., Rühl C., Emonts J., Buyel J. F. (2019). Seasonal weather changes affect the yield and quality of recombinant proteins produced in transgenic tobacco plants in a greenhouse setting. Front. plant Sci. 10, 1245. doi: 10.3389/fpls.2019.01245. CrossRef. Google Scholar.
125. Krikorian A. D., Ledbetter M. C. (1975). Some observations on the cultivation of opium poppy (Papaver somniferum L) for its latex. Bot. Rev. 41 (1), 30–103. [PMID: 11631688]. doi: 10.1007/BF02860836. CrossRef. Google Scholar.
126. Kuyukina M. S., Ivshina I. B., Korshunova I. O., Rubtsova E. V. (2014). Assessment of bacterial resistance to organic solvents using a combined confocal laser scanning and atomic force microscopy (CLSM/AFM). J. Microbiol. Methods 107, 23–29. [PMID: 25193441]. doi: 10.1016/j.mimet.2014.08.020. CrossRef. Google Scholar.
127. Lähdeaho M-L., Mäki M., Laurila K., Huhtala H., Kaukinen K. (2011). Small-bowel mucosal changes and antibody responses after low- and moderate-dose gluten challenge in celiac disease. BMC Gastroenterol. 11, 129. [PMID: 22115041]. doi: 10.1186/1471-230X-11-129. CrossRef. Google Scholar.
128. Lali N., Satzer P., Jungbauer A. (2022). Residence time distribution in counter-current protein A affinity chromatography using an inert tracer. J. Chromatogr. A 1683, 463530. [PMID: 36206581]. doi: 10.1016/j.chroma.2022.463530. CrossRef. Google Scholar.
129. Lechtenberg M. (2005-2010). "Cyanogenic glycosides and biogenetically related compounds in higher plants and animals," in Encyclopedia of life sciences. Editors Wiley, John Sons L. (London, New York: John Wiley & Sons), 1–18. Google Scholar.
130. Lee S. T., Wildeboer K., Panter K. E., Kem W. R., Gardner D. R., Molyneux R. J. et al (2006). Relative toxicities and neuromuscular nicotinic receptor agonistic potencies of anabasine enantiomers and anabaseine. Neurotoxicol Teratol. 28 (2), 220–228. [PMID: 16488116]. doi: 10.1016/j.ntt.2005.12.010. CrossRef. Google Scholar.
131. Legrand J., Artu A., Pruvost J. (2021). A review on photobioreactor design and modelling for microalgae production. React. Chem. Eng. 6 (7), 1134–1151. doi: 10.1039/D0RE00450B. CrossRef. Google Scholar.
132. Leth I. K., McDonald K. A. (2017). Media development for large scale Agrobacterium tumefaciens culture. Biotechnol. Prog. 33 (5), 1218–1225. [PMID: 28556626]. doi: 10.1002/btpr.2504. CrossRef. Google Scholar.
133. Lewis R. J. (2000). Ion channel toxins and therapeutics: From cone snail venoms to ciguatera. Ther. Drug Monit. 22 (1), 61–64. [PMID: 10688261]. doi: 10.1097/00007691-200002000-00013. CrossRef. Google Scholar.
134. Li Y. (2022). Two major mechanisms contributing to copurification of CHO host cell proteins and strategies to minimize their negative impact. Protein Expr. Purif. 197, 106113. [PMID: 35598695]. doi: 10.1016/j.pep.2022.106113. CrossRef. Google Scholar.
135. Liu D. L., Lovett J. V. (1993). Biologically active secondary metabolites of barley. II. Phytotoxicity of barley allelochemicals. J. Chem. Ecol. 19 (10), 2231–2244. [PMID: 24248572]. doi: 10.1007/BF00979660. CrossRef. Google Scholar.
136. Lücker J., Schwab W., van Hautum B., Blaas J., van der Plas L. H. W., Bouwmeester H. J. et al (2004). Increased and altered fragrance of tobacco plants after metabolic engineering using three monoterpene synthases from lemon. Plant Physiol. 134 (1), 510–519. [PMID: 14718674]. doi: 10.1104/pp.103.030189. CrossRef. Google Scholar.
137. Luo H., Li Y., Robbins D., Wang S., Xi G., Cox M. et al (2021). Safety risk management for low molecular weight process-related impurities in monoclonal antibody therapeutics: Categorization, risk assessment, testing strategy, and process development with leveraging clearance potential. Biotechnol. Prog. 37 (3), e3119. [PMID: 33373106]. doi: 10.1002/btpr.3119. CrossRef. Google Scholar.
138. Ma J., Rukh G., Ruan Z., Xie X., Ye Z., Liu D. (2022). Effects of hypoxia stress on growth, root respiration, and metabolism of phyllostachys praecox. Life (Basel) 12 (6), 808. [PMID: 35743839]. doi: 10.3390/life12060808. CrossRef. Google Scholar.
139. Ma J. K., Drossard J., Lewis D., Altmann F., Boyle J., Christou P. et al (2015). Regulatory approval and a first-in-human phase I clinical trial of a monoclonal antibody produced in transgenic tobacco plants. Plant Biotechnol. J. 13 (8), 1106–1120. [PMID: 26147010]. doi: 10.1111/pbi.12416. CrossRef. Google Scholar.
140. Mandal M. K., Ahvari H., Schillberg S., Schiermeyer A. (2016). Tackling unwanted proteolysis in plant production hosts used for molecular farming. Front. plant Sci. 7, 267. [PMID: 27014293]. doi: 10.3389/fpls.2016.00267. CrossRef. Google Scholar.
141. Marchese S., Polo A., Ariano A., Velotto S., Costantini S., Severino L. (2018). Aflatoxin B1 and M1: Biological properties and their involvement in cancer development. Toxins (Basel) 10 (6), 214. [PMID: 29794965]. doi: 10.3390/toxins10060214. CrossRef. Google Scholar.
142. Martinez M., Gómez-Cabellos S., Giménez M. J., Barro F., Diaz I., Diaz-Mendoza M. (2019). Plant proteases: From key enzymes in germination to allies for fighting human gluten-related disorders. Front. Plant Sci. 10, 721. [PMID: 31191594]. doi: 10.3389/fpls.2019.00721. CrossRef. Google Scholar.
143. Mayer B. (2014). How much nicotine kills a human? Tracing back the generally accepted lethal dose to dubious self-experiments in the nineteenth century. Arch. Toxicol. 88 (1), 5–7. [PMID: 24091634]. doi: 10.1007/s00204-013-1127-0. CrossRef. Google Scholar.
144. Mazzer A. R., Perraud X., Halley J., O'Hara J., Bracewell D. G. (2015). Protein A chromatography increases monoclonal antibody aggregation rate during subsequent low pH virus inactivation hold. J. Chromatogr. A 1415, 83–90. [PMID: 26346187]. doi: 10.1016/j.chroma.2015.08.068. CrossRef. Google Scholar.
145. Mena M., Sanchez-Monge R., Gomez L., Salcedo G., Carbonero P. (1992). A major barley allergen associated with baker's asthma disease is a glycosylated monomeric inhibitor of insect alpha-amylase: cDNA cloning and chromosomal location of the gene. Plant Mol. Biol. 20 (3), 451–458. [PMID: 1421148]. doi: 10.1007/BF00040604. CrossRef. Google Scholar.
146. Menary J., Amato M., Sanchez A. C., Hobbs M., Pacho A., Fuller S. S. (2020a). New hope for a "cursed" crop? Understanding stakeholder attitudes to plant molecular farming with modified tobacco in europe. Front. Plant Sci. 11, 791. [PMID: 32595677]. doi: 10.3389/fpls.2020.00791. CrossRef. Google Scholar.
147. Menary J., Hobbs M., Mesquita de Albuquerque S., Pacho A., Drake P. M. W., Prendiville A. et al (2020b). Shotguns vs Lasers: Identifying barriers and facilitators to scaling-up plant molecular farming for high-value health products. PloS one 15 (3), e0229952. [PMID: 32196508]. doi: 10.1371/journal.pone.0229952. CrossRef. Google Scholar.
148. Menzel S., Holland T., Boes A., Spiegel H., Fischer R., Buyel J. F. (2018). Downstream processing of a plant-derived malaria transmission-blocking vaccine candidate. Protein Expres Purif. 152, 122–130. [PMID: 30059744]. doi: 10.1016/j.pep.2018.07.012. CrossRef. Google Scholar.
149. Menzel S., Holland T., Boes A., Spiegel H., Bolzenius J., Fischer R. et al (2016). Optimized blanching reduces the host cell protein content and substantially enhances the recovery and stability of two plant-derived malaria vaccine candidates. Front. plant Sci. 7 (159), 159–215. doi: 10.3389/fpls.2016.00159. CrossRef. Google Scholar.
150. Miranda-Apodaca J., Artetxe U., Aguado I., Martin-Souto L., Ramirez-Garcia A., Lacuesta M. et al (2023). Stress response to climate change and postharvest handling in two differently pigmented lettuce genotypes: Impact on Alternaria alternata invasion and mycotoxin production. Plants (Basel) 12 (6), 1304. [PMID: 36986993]. doi: 10.3390/plants12061304. CrossRef. Google Scholar.
151. Mirzaee M., Osmani Z., Frébortová J., Frébort I. (2022). Recent advances in molecular farming using monocot plants. Biotechnol. Adv. 58, 107913. [PMID: 35081454]. doi: 10.1016/j.biotechadv.2022.107913. CrossRef. Google Scholar.
152. Miyoshi N., Koyama Y., Katsuno Y., Hayakawa S., Mita T., Ohta T. et al (2001). Apoptosis induction associated with cell cycle dysregulation by rice bran agglutinin. J. Biochem. 130 (6), 799–805. [PMID: 11726280]. doi: 10.1093/oxfordjournals.jbchem.a003051. CrossRef. Google Scholar.
153. Montero-Morales L., Steinkellner H. (2018). Advanced plant-based glycan engineering. Front. Bioeng. Biotechnol. 6, 81. [PMID: 29963553]. doi: 10.3389/fbioe.2018.00081. CrossRef. Google Scholar.
154. Moore B., Andrew R. L., Kulheim C., Foley W. J. (2014). Explaining intraspecific diversity in plant secondary metabolites in an ecological context. New Phytol. 201 (3), 733–750. doi: 10.1111/Nph.12526. CrossRef. Google Scholar.
155. Mortimer C. L., Dugdale B., Dale J. L. (2015). Updates in inducible transgene expression using viral vectors: From transient to stable expression. Curr. Opin. Biotechnol. 32, 85–92. [PMID: 25437638]. doi: 10.1016/j.copbio.2014.11.009. CrossRef. Google Scholar.
156. Müller J., Ott V., Eibl D., Eibl R. (2022). Seed train intensification using an ultra-high cell density cell banking process. Processes 10 (5), 911. doi: 10.3390/pr10050911. CrossRef. Google Scholar.
157. Mündel T., Machal M., Cochrane D. J., Barnes M. J. (2017). A randomised, placebo-controlled, crossover study investigating the effects of nicotine gum on strength, power and anaerobic performance in nicotine-naïve, active males. Sports Med. Open 3 (1), 5. [PMID: 28092056]. doi: 10.1186/s40798-016-0074-8. CrossRef. Google Scholar.
158. Muñoz-García E., Luengo-Sánchez O., Haroun-Díaz E., Maroto A. S., Palacín A., Díaz -Perales A. et al (2013). Identification of thaumatin-like protein and aspartyl protease as new major allergens in lettuce (Lactuca sativa). Mol. Nutr. Food Res. 57 (12), 2245–2252. [PMID: 23983075]. doi: 10.1002/mnfr.201300139. CrossRef. Google Scholar.
159. Murthy H. N., Joseph K. S., Paek K. Y., Park S. Y. (2023). Bioreactor systems for micropropagation of plants: Present scenario and future prospects. Front. Plant Sci. 14, 1159588. [PMID: 37152119]. doi: 10.3389/fpls.2023.1159588. CrossRef. Google Scholar.
160. Napagoda M. (2022). "9 Poisonous plants and their toxic metabolites," in Chemistry of natural products. Editors Napagoda M., Jayasinghe L. (Berlin, Germany: De Gruyter), 173–188. Google Scholar.
161. Nikbakhtzadeh M. R., Terbot J. W., Otienoburu P. E., Foster W. A. (2014). Olfactory basis of floral preference of the malaria vector Anopheles gambiae (Diptera: Culicidae) among common african plants. J. Vector Ecol. 39 (2), 372–383. [PMID: 25424267]. doi: 10.1111/jvec.12113. CrossRef. Google Scholar.
162. Nishimura A., Nakahara T., Ueno T., Sasaki K., Yoshida S., Kyo S. et al (2006). Requirement of E7 oncoprotein for viability of HeLa cells. Microbes Infect. 8 (4), 984–993. [PMID: 16500131]. doi: 10.1016/j.micinf.2005.10.015. CrossRef. Google Scholar.
163. Nyasembe V. O., Teal P. E. A., Mukabana W. R., Tumlinson J. H., Torto B. (2012). Behavioural response of the malaria vector Anopheles gambiae to host plant volatiles and synthetic blends. Parasit. Vectors 5, 234. [PMID: 23069316]. doi: 10.1186/1756-3305-5-234. CrossRef. Google Scholar.
164. Nyirenda K. K. (2021). "Toxicity potential of cyanogenic glycosides in edible plants," in Medical toxicology. Editors Erkekoglu P., Ogawa T. (London, UK: IntechOpen). Google Scholar.
165. Ochoa-Villarreal M., Howat S., Hong S., Jang M. O., Jin Y. W., Lee E. K. et al (2016). Plant cell culture strategies for the production of natural products. Bmb Rep. 49 (3), 149–158. [PMID: 26698871]. doi: 10.5483/bmbrep.2016.49.3.264. CrossRef. Google Scholar.
166. Ohizumi Y., Yasumoto T. (1983). Contraction and increase in tissue calcium content induced by maitotoxin, the most potent known marine toxin, in intestinal smooth muscle. Br. J. Pharmacol. 79 (1), 3–5. [PMID: 6871549]. doi: 10.1111/j.1476-5381.1983.tb10485.x. CrossRef. Google Scholar.
167. Olsnes S., Stirpe F., Sandvig K., Pihl A. (1982). Isolation and characterization of viscumin, a toxic lectin from Viscum album L. (mistletoe). J. Biol. Chem. 257 (22), 13263–13270. [PMID: 7142144]. doi: 10.1016/S0021-9258(18)33440-9. CrossRef. Google Scholar.
168. Onoyovwe A., Hagel J. M., Chen X., Khan M. F., Schriemer D. C., Facchini P. J. (2013). Morphine biosynthesis in opium poppy involves two cell types: Sieve elements and laticifers. Plant Cell 25 (10), 4110–4122. [PMID: 24104569]. doi: 10.1105/tpc.113.115113. CrossRef. Google Scholar.
169. Opal S. M. (2010). Endotoxins and other sepsis triggers. Contrib. Nephrol. 167, 14–24. [PMID: 20519895]. doi: 10.1159/000315915. CrossRef. Google Scholar.
170. Opdensteinen P., Clodt J. I., Müschen C. R., Filiz V., Buyel J. F. (2018). A combined ultrafiltration/diafiltration step facilitates the purification of cyanovirin-N from transgenic tobacco extracts. Front. Bioeng. Biotechnol. 6, 206. [PMID: 30687700]. doi: 10.3389/fbioe.2018.00206. CrossRef. Google Scholar.
171. Opdensteinen P., Lobanov A., Buyel J. F. (2020). A combined pH and temperature precipitation step facilitates the purification of tobacco‐derived recombinant proteins that are sensitive to extremes of either parameter. Biotechnol. J. 16, 2000340. In preparation. doi: 10.1002/biot.202000340. CrossRef. Google Scholar.
172. Ortega N., Quiralte J., Blanco C., Castillo R., Alvarez M. J., Carrillo T. (1999). Tobacco allergy: Demonstration of cross-reactivity with other members of solanaceae family and mugwort pollen. Ann. Allergy Asthma Immunol. 82 (2), 194–197. [PMID: 10071524]. doi: 10.1016/S1081-1206(10)62596-3. CrossRef. Google Scholar.
173. Osborne T. B. (1907). The proteins of the wheat kernel. Washington: Carnegie Inst. Google Scholar.
174. Palladino M. A., Levinson A. D., Svedersky L. P., Obijeski J. F. (1987). Safety issues related to the use of recombinant DNA-derived cell culture products. I. Cellular components. Dev. Biol. Stand 66, 13–22. [PMID: 3582742]. Google Scholar.
175. Pantazica A-M., van Eerde A., Dobrica M-O., Caras I., Ionescu I., Costache A. et al (2023). The "humanized" N-glycosylation pathway in CRISPR/Cas9-edited Nicotiana benthamiana significantly enhances the immunogenicity of a S/preS1 Hepatitis B Virus antigen and the virus-neutralizing antibody response in vaccinated mice. Plant Biotechnol. J. 21 (6), 1176–1190. [PMID: 36779605]. doi: 10.1111/pbi.14028. CrossRef. Google Scholar.
176. Pastorello E. A., Farioli L., Pravettoni V., Ispano M., Scibola E., Trambaioli C. et al (2000). The maize major allergen, which is responsible for food-induced allergic reactions, is a lipid transfer protein. J. Allergy Clin. Immunol. 106 (4), 744–751. [PMID: 11031346]. doi: 10.1067/mai.2000.108712. CrossRef. Google Scholar.
177. Paulsen E., Andersen K. E. (2016). Lettuce contact allergy. Contact Dermat. 74 (2), 67–75. [PMID: 26289653]. doi: 10.1111/cod.12458. CrossRef. Google Scholar.
178. Peden K., Sheng L., Pal A., Lewis A. (2006). Biological activity of residual cell-substrate DNA. Dev. Biol. 123, 45–53. [PMID: 16566436]. Google Scholar.
179. Pereira D. M., Valentao P., Correia-da-Silva G., Teixeira N., Andrade P. B. (2012). Plant secondary metabolites in cancer chemotherapy: Where are we? Curr. Pharm. Biotechno 13 (5), 632–650. [PMID: 22122478]. doi: 10.2174/138920112799857530. CrossRef. Google Scholar.
180. Petsch D., Anspach F. B. (2000). Endotoxin removal from protein solutions. J. Biotechnol. 76 (2-3), 97–119. [PMID: 10656326]. doi: 10.1016/S0168-1656(99)00185-6. CrossRef. Google Scholar.
181. Platis D., Drossard J., Fischer R., Ma J. K. C., Labrou N. E. (2008). New downstream processing strategy for the purification of monoclonal antibodies from transgenic tobacco plants. J. Chromatogr. A 1211 (1-2), 80–89. doi: 10.1016/j.chroma.2008.09.103. CrossRef. Google Scholar.
182. Pronin D., Börner A., Weber H., Scherf K. A. (2020). Wheat (Triticum aestivum L) breeding from 1891 to 2010 contributed to increasing yield and glutenin contents but decreasing protein and gliadin contents. J. Agric. Food Chem. 68 (46), 13247–13256. [PMID: 32648759]. doi: 10.1021/acs.jafc.0c02815. CrossRef. Google Scholar.
183. Qiu J., Li K., Miller K., Raghani A. (2015). Risk-based strategy to determine testing requirement for the removal of residual process reagents as process-related impurities in bioprocesses. Pda J. Pharm. Sci. Tech. 69 (3), 334–345. [PMID: 26048741]. doi: 10.5731/pdajpst.2015.01056. CrossRef. Google Scholar.
184. Rademacher T., Sack M., Blessing D., Fischer R., Holland T., Buyel J. F. (2019). Plant cell packs: A scalable platform for recombinant protein production and metabolic engineering. Plant Biotechnol. J. 17 (8), 1560–1566. [PMID: 30672078]. doi: 10.1111/pbi.13081. CrossRef. Google Scholar.
185. Rasool F., Nizamani Z. A., Ahmad K. S., Parveen F., Khan S. A., Sabir N. (2022). Phytotoxicological study of selected poisonous plants from Azad Jammu & Kashmir. PLoS ONE 17 (5), e0263605. [PMID: 35544538]. doi: 10.1371/journal.pone.0263605. CrossRef. Google Scholar.
186. Raven N., Rasche S., Kuehn C., Anderlei T., Klöckner W., Schuster F. et al (2014). Scaled-up manufacturing of recombinant antibodies produced by plant cells in a 200-L orbitally-shaken disposable bioreactor. Biotechnol. Bioeng. 112, 308–321. [PMID: 25117428]. doi: 10.1002/bit.25352. CrossRef. Google Scholar.
187. Raven N., Schillberg S., Kirchhoff J., Brändli J., Imseng N., Eibl R. (2011). "Growth of BY-2 suspension cells and plantibody production in single-use bioreactors," in Single-use technology in biopharmaceutical manufacture. Editors Eibl R., Eibl D. (Hoboken, NY, USA: John Wiley & Sons), 252–259. Google Scholar.
188. Ren G., Wu H., Bao A., Lin T., Ting K-C., Ying Y. (2023). Mobile robotics platform for strawberry temporal-spatial yield monitoring within precision indoor farming systems. Front. Plant Sci. 14, 1162435. [PMID: 37180389]. doi: 10.3389/fpls.2023.1162435. CrossRef. Google Scholar.
189. Rihan H. Z., Aljafer N., Jbara M., McCallum L., Lengger S., Fuller M. P. (2022). The impact of LED lighting spectra in a plant factory on the growth, physiological traits and essential oil content of lemon balm (melissa officinalis). Plants (Basel) 11 (3), 342. [PMID: 35161322]. doi: 10.3390/plants11030342. CrossRef. Google Scholar.
190. Rivas-San Vicente M., Larios-Zarate G., Plasencia J. (2013). Disruption of sphingolipid biosynthesis in Nicotiana benthamiana activates salicylic acid-dependent responses and compromises resistance to Alternaria alternata f. sp. lycopersici. Planta 237 (1), 121–136. [PMID: 22990908]. doi: 10.1007/s00425-012-1758-z. CrossRef. Google Scholar.
191. Rofiat A-S., Fanelli F., Atanda O., Sulyok M., Cozzi G., Bavaro S. et al (2015). Fungal and bacterial metabolites associated with natural contamination of locally processed rice (Oryza sativa L) in Nigeria. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 32 (6), 950–959. [PMID: 25767914]. doi: 10.1080/19440049.2015.1027880. CrossRef. Google Scholar.
192. Rup B., Alon S., Amit-Cohen B. C., Brill Almon E., Chertkoff R., Tekoah Y. et al (2017). Immunogenicity of glycans on biotherapeutic drugs produced in plant expression systems-The taliglucerase alfa story. PloS one 12 (10), e0186211. [PMID: 29088235]. doi: 10.1371/journal.pone.0186211. CrossRef. Google Scholar.
193. Sack M., Rademacher T., Spiegel H., Boes A., Hellwig S., Drossard J. et al (2015). From gene to harvest: Insights into upstream process development for the GMP production of a monoclonal antibody in transgenic tobacco plants. Plant Biotechnol. J. 13 (8), 1094–1105. doi: 10.1111/pbi.12438. CrossRef. Google Scholar.
194. Salguero-Linares J., Coll N. S. (2023). Cell death as a defense strategy against pathogens in plants and animals. PLoS Pathog. 19 (4), e1011253. [PMID: 37023043]. doi: 10.1371/journal.ppat.1011253. CrossRef. Google Scholar.
195. Sanchez-Ramos J. R. (2020). The rise and fall of tobacco as a botanical medicine. J. Herb. Med. 22, 100374. [PMID: 32834941]. doi: 10.1016/j.hermed.2020.100374. CrossRef. Google Scholar.
196. Schachtsiek J., Stehle F. (2019). Nicotine-free, nontransgenic tobacco (Nicotiana tabacum l) edited by CRISPR-Cas9. Plant Biotechnol. J. 17 (12), 2228–2230. [PMID: 31206994]. doi: 10.1111/pbi.13193. CrossRef. Google Scholar.
197. Schubert-Ullrich P., Rudolf J., Ansari P., Galler B., Führer M., Molinelli A. et al (2009). Commercialized rapid immunoanalytical tests for determination of allergenic food proteins: An overview. Anal. Bioanal. Chem. 395 (1), 69–81. [PMID: 19308361]. doi: 10.1007/s00216-009-2715-y. CrossRef. Google Scholar.
198. Schwestka J., König-Beihammer J., Shin Y-J., Vavra U., Kienzl N. F., Grünwald-Gruber C. et al (2021). Impact of specific N-glycan modifications on the use of plant-produced SARS-CoV-2 antigens in serological assays. Front. Plant Sci. 12, 747500. [PMID: 34646292]. doi: 10.3389/fpls.2021.747500. CrossRef. Google Scholar.
199. Sekiya R., Nagano T., Moriyama T., Kishi T., Shinke H., Yano E. et al (2020). Occupational respiratory allergy to lettuce in lettuce farmers. Clin. Exp. Allergy 50 (8), 932–941. [PMID: 32542808]. doi: 10.1111/cea.13682. CrossRef. Google Scholar.
200. Sencar J., Hammerschmidt N., Jungbauer A. (2020). Modeling the residence time distribution of integrated continuous bioprocesses. Biotechnol. J. 15 (8), e2000008. [PMID: 32302457]. doi: 10.1002/biot.202000008. CrossRef. Google Scholar.
201. Serdakowski London A., Kerins B., Tschantz W. R., Mackay K. (2012). Endotoxin removal and prevention for pre-clinical biologics production. Biotechnol. J. 7 (12), 1509–1516. [PMID: 23081824]. doi: 10.1002/biot.201200220. CrossRef. Google Scholar.
202. Shaaltiel Y., Tekoah Y. (2016). Plant specific N-glycans do not have proven adverse effects in humans. Nat. Biotechnol. 34 (7), 706–708. [PMID: 27404878]. doi: 10.1038/nbt.3556. CrossRef. Google Scholar.
203. Sharma R., Singh A. K., Umashankar V. (2009). Characterization of allergenic epitopes of Ory s1 protein from Oryza sativa and its homologs. Bioinformation 4 (1), 12–18. [PMID: 20011147]. doi: 10.6026/97320630004012. CrossRef. Google Scholar.
204. Sharrer M. J., Summerfelt S. T. (2007). Ozonation followed by ultraviolet irradiation provides effective bacteria inactivation in a freshwater recirculating system. Aquac. Eng. 37 (2), 180–191. doi: 10.1016/j.aquaeng.2007.05.001. CrossRef. Google Scholar.
205. Sheng L., Cai F., Zhu Y., Pal A., Athanasiou M., Orrison B. et al (2008). Oncogenicity of DNA in vivo: Tumor induction with expression plasmids for activated H-ras and c-myc. Biologicals 36 (3), 184–197. [PMID: 18218323]. doi: 10.1016/j.biologicals.2007.11.003. CrossRef. Google Scholar.
206. Sigmund G., Arp H. P. H., Aumeier B. M., Bucheli T. D., Chefetz B., Chen W. et al (2022). Sorption and mobility of charged organic compounds: How to confront and overcome limitations in their assessment. Environ. Sci. Technol. 56 (8), 4702–4710. [PMID: 35353522]. doi: 10.1021/acs.est.2c00570. CrossRef. Google Scholar.
207. Singh A. A., Pillay P., Tsekoa T. L. (2021). Engineering approaches in plant molecular farming for global health. Vaccines (Basel) 9 (11), 1270. [PMID: 34835201]. doi: 10.3390/vaccines9111270. CrossRef. Google Scholar.
208. Sisson V. A., Severson R. F. (1990). Alkaloid composition of the nicotiana species. Contributions Tob. Nicotine Res. 14 (6), 327–339. doi: 10.2478/cttr-2013-0610. CrossRef. Google Scholar.
209. Smahel M., Sima P., Ludvikova V., Vonka V. (2001). Modified HPV16 E7 genes as DNA vaccine against E7-containing oncogenic cells. Virology 281 (2), 231–238. doi: 10.1006/viro.2000.0794. CrossRef. Google Scholar.
210. Sobiczewski P., Iakimova E. T. (2022). Plant and human pathogenic bacteria exchanging their primary host environments. J. Hortic. Res. 30 (1), 11–30. doi: 10.2478/johr-2022-0009. CrossRef. Google Scholar.
211. Sparrow P., Broer I., Hood E., Eversole K., Hartung F., Schiemann J. (2013). Risk assessment and regulation of molecular farming - a comparison between europe and us. Curr. Pharm. Des. 19 (31), 5513–5530. doi: 10.2174/1381612811319310007. CrossRef. Google Scholar.
212. Spiegel H., Boes A., Perales Morales C., Rademacher T., Buyel J. F. (2019). Ready to use stocks of Agrobacterium tumefaciens can simplify process development for the production of recombinant proteins by transient expression in plants. Biotechnol. J. 14, e1900113. [PMID: 31218827]. doi: 10.1002/biot.201900113. CrossRef. Google Scholar.
213. Stephan A., Hahn-Löbmann S., Rosche F., Buchholz M., Giritch A., Gleba Y. (2017). Simple purification of nicotiana benthamiana-produced recombinant colicins: High-yield recovery of purified proteins with minimum alkaloid content supports the suitability of the host for manufacturing food additives. Int. J. Mol. Sci. 19 (1), 95. [PMID: 29286298]. doi: 10.3390/ijms19010095. CrossRef. Google Scholar.
214. Stoger E., Sack M., Perrin Y., Vaquero C., Torres E., Twyman R. M. et al (2002). Practical considerations for pharmaceutical antibody production in different crop systems. Mol. Breed. 9 (3), 149–158. doi: 10.1023/A:1019714614827. CrossRef. Google Scholar.
215. Stone M. C., Borman J., Ferreira G., Robbins P. D. (2018). Effects of pH, conductivity, host cell protein, and DNA size distribution on DNA clearance in anion exchange chromatography media. Biotechnol. Prog. 34 (1), 141–149. [PMID: 28884511]. doi: 10.1002/btpr.2556. CrossRef. Google Scholar.
216. Strasser R., Altmann F., Mach L., Glössl J., Steinkellner H. (2004). Generation of Arabidopsis thaliana plants with complex N-glycans lacking β1,2-linked xylose and core α1,3-linked fucose. FEBS Lett. 561 (1-3), 132–136. doi: 10.1016/S0014-5793(04)00150-4. CrossRef. Google Scholar.
217. Strasser R., Stadlmann J., Schahs M., Stiegler G., Quendler H., Mach L. et al (2008). Generation of glyco-engineered Nicotiana benthamiana for the production of monoclonal antibodies with a homogeneous human-like N-glycan structure. Plant Biotechnol. J. 6 (4), 392–402. doi: 10.1111/j.1467-7652.2008.00330.x. CrossRef. Google Scholar.
218. Strasser R. (2016). Plant protein glycosylation. Glycobiology 26 (9), 926–939. [PMID: 26911286]. doi: 10.1093/glycob/cww023. CrossRef. Google Scholar.
219. Sweany R. R., Breunig M., Opoku J., Clay K., Spatafora J. W., Drott M. T. et al (2022). Why do plant-pathogenic fungi produce mycotoxins? Potential roles for mycotoxins in the plant ecosystem. Phytopathology 112 (10), 2044–2051. [PMID: 35502928]. doi: 10.1094/phyto-02-22-0053-sym. CrossRef. Google Scholar.
220. Szermer-Olearnik B., Boratyński J. (2015). Removal of endotoxins from bacteriophage preparations by extraction with organic solvents. PLoS ONE 10 (3), e0122672. [PMID: 25811193]. doi: 10.1371/journal.pone.0122672. CrossRef. Google Scholar.
221. Taraghikhah N., Ashtari S., Asri N., Shahbazkhani B., Al-Dulaimi D., Rostami-Nejad M. et al (2020). An updated overview of spectrum of gluten-related disorders: Clinical and diagnostic aspects. BMC Gastroenterol. 20 (1), 258. [PMID: 32762724]. doi: 10.1186/s12876-020-01390-0. CrossRef. Google Scholar.
222. Taylor P. C., Adams A. C., Hufford M. M., La Torre I. de, Winthrop K., Gottlieb R. L. (2021). Neutralizing monoclonal antibodies for treatment of COVID-19. Nat. Rev. Immunol. 21 (6), 382–393. [PMID: 33875867]. doi: 10.1038/s41577-021-00542-x. CrossRef. Google Scholar.
223. Tekoah Y., Tzaban S., Kizhner T., Hainrichson M., Gantman A., Golembo M. et al (2013). Glycosylation and functionality of recombinant beta-glucocerebrosidase from various production systems. Biosci. Rep. 33, e00071–U272. doi: 10.1042/Bsr20130081. CrossRef. Google Scholar.
224. Thoma F., Somborn-Schulz A., Schlehuber D., Keuter V., Deerberg G. (2020). Effects of light on secondary metabolites in selected leafy greens: A review. Front. Plant Sci. 11, 497. [PMID: 32391040]. doi: 10.3389/fpls.2020.00497. CrossRef. Google Scholar.
225. Tosi P., Gritsch C. S., He J., Shewry P. R. (2011). Distribution of gluten proteins in bread wheat (Triticum aestivum) grain. Ann. Bot. 108 (1), 23–35. [PMID: 21693664]. doi: 10.1093/aob/mcr098. CrossRef. Google Scholar.
226. Toxic Rep Ser (1993). NTP toxicity studies of sodium cyanide (CAS No. 143-33-9) administered by dosed water to F344/N rats and B6C3F1 mice. Toxic. Rep. Ser. 37, 1. [PMID: 11965236]. Google Scholar.
227. Tusé D., Nandi S., McDonald K. A., Buyel J. F. (2020). The emergency response capacity of plant-based biopharmaceutical manufacturing-what it is and what it could Be. Front. Plant Sci. 11, 594019. [PMID: 33193552]. doi: 10.3389/fpls.2020.594019. CrossRef. Google Scholar.
228. Tyutereva E. V., Dobryakova K. S., Schiermeyer A., Shishova M. F., Pawlowski K., Demidchik V. et al (2018). The levels of peroxisomal catalase protein and activity modulate the onset of cell death in tobacco BY-2 cells via reactive oxygen species levels and autophagy. Funct. Plant Biol. 45 (2), 247–258. [PMID: 32291039]. doi: 10.1071/FP16418. CrossRef. Google Scholar.
229. Uranga M., Vazquez-Vilar M., Orzáez D., Daròs J-A. (2021). CRISPR-Cas12a genome editing at the whole-plant level using two compatible RNA virus vectors. CRISPR J. 4 (5), 761–769. [PMID: 34558964]. doi: 10.1089/crispr.2021.0049. CrossRef. Google Scholar.
230. Urbański A., Konopińska N., Bylewska N., Gmyrek R., Spochacz-Santoro M., Bufo S. A. et al (2023). Solanum nigrum fruit extract modulates immune system activity of mealworm beetle, Tenebrio molitor L. Tenebrio Molit. L. Toxins (Basel) 15 (1), 68. [PMID: 36668887]. doi: 10.3390/toxins15010068. CrossRef. Google Scholar.
231. Usui Y., Nakase M., Hotta H., Urisu A., Aoki N., Kitajima K. et al (2001). A 33-kDa allergen from rice (oryza sativa L.japonica). J. Biol. Chem. 276 (14), 11376–11381. [PMID: 11139585]. doi: 10.1074/jbc.M010337200. CrossRef. Google Scholar.
232. van Delden S. H., SharathKumar M., Butturini M., Graamans L. J. A., Heuvelink E., Kacira M. et al (2021). Current status and future challenges in implementing and upscaling vertical farming systems. Nat. Food 2, 944–956. doi: 10.1038/s43016-021-00402-w. CrossRef. Google Scholar.
233. van der Hoorn R. A. L., Klemenčič M. (2021). Plant proteases: From molecular mechanisms to functions in development and immunity. J. Exp. Bot. 72 (9), 3337–3339. [PMID: 33847361]. doi: 10.1093/jxb/erab129. CrossRef. Google Scholar.
234. Venuti A., Massa S., Mett V., Vedova L. D., Paolini F., Franconi R. et al (2009). An E7-based therapeutic vaccine protects mice against HPV16 associated cancer. Vaccine 27 (25-26), 3395–3397. doi: 10.1016/j.vaccine.2009.01.068. CrossRef. Google Scholar.
235. Voitsekhovskaja O. V., Schiermeyer A., Reumann S. (2014). Plant peroxisomes are degraded by starvation-induced and constitutive autophagy in tobacco BY-2 suspension-cultured cells. Front. Plant Sci. 5, 629. [PMID: 25477890]. doi: 10.3389/fpls.2014.00629. CrossRef. Google Scholar.
236. Walsh G., Walsh E. (2022). Biopharmaceutical benchmarks 2022. Nat. Biotechnol. 40 (12), 1722–1760. [PMID: 36471135]. doi: 10.1038/s41587-022-01582-x. CrossRef. Google Scholar.
237. Wang S., Alseekh S., Fernie A. R., Luo J. (2019). The structure and function of major plant metabolite modifications. Mol. Plant 12 (7), 899–919. [PMID: 31200079]. doi: 10.1016/j.molp.2019.06.001. CrossRef. Google Scholar.
238. Wang X., Morgan D. M., Wang G., Mozier N. M. (2012). Residual DNA analysis in biologics development: Review of measurement and quantitation technologies and future directions. Biotechnol. Bioeng. 109 (2), 307–317. [PMID: 21956148]. doi: 10.1002/bit.23343. CrossRef. Google Scholar.
239. Ward B. J., Gobeil P., Séguin A., Atkins J., Boulay I., Charbonneau P. Y. et al (2021). Phase 1 randomized trial of a plant-derived virus-like particle vaccine for COVID-19. Nat. Med. 27 (6), 1071–1078. [PMID: 34007070]. doi: 10.1038/s41591-021-01370-1. CrossRef. Google Scholar.
240. Weirich C. A., Miller T. R. (2014). Freshwater harmful algal blooms: Toxins and children's health. Curr. Probl. Pediatr. Adolesc. Health Care 44 (1), 2–24. [PMID: 24439026]. doi: 10.1016/j.cppeds.2013.10.007. CrossRef. Google Scholar.
241. Welch K. D., Lee S. T., Panter K. E., Gardner D., Knoppel E., Green B. et al (2014). Studies on the teratogenicity of anabasine in a rat model. Toxicon 87, 32–37. [PMID: 24905648]. doi: 10.1016/j.toxicon.2014.05.018. CrossRef. Google Scholar.
242. Werner S., Maschke R. W., Eibl D., Eibl R. (2018). Bioreactor technology for sustainable production of plant cell-derived products. In: Bley T., Pavlov A. editors. Bioprocessing of plant in vitro systems. Cham: Springer International Publishing, 413–432. Google Scholar.
243. Wilbers R. H., Westerhof L. B., van Raaij D. R., Adrichem M., Prakasa A. D., Lozano-Torres J. L. et al (2016). Co-expression of the protease furin inNicotiana benthamianaleads to efficient processing of latent transforming growth factor-β1 into a biologically active protein. Plant Biotechnol. J. 14 (8), 1695–1704. [PMID: 26834022]. doi: 10.1111/pbi.12530. CrossRef. Google Scholar.
244. Wilson S. A., Roberts S. C. (2012). Recent advances towards development and commercialization of plant cell culture processes for the synthesis of biomolecules. Plant Biotechnol. J. 10 (3), 249–268. doi: 10.1111/j.1467-7652.2011.00664.x. CrossRef. Google Scholar.
245. Wink M. (2009). Mode of action and toxicology of plant toxins and poisonous plants. Mitt. Julius Kühn-Inst 421, 93–112. Google Scholar.
246. Wirz H., Sauer-Budge A. F., Briggs J., Sharpe A., Shu S. D., Sharon A. (2012). Automated production of plant-based vaccines and pharmaceuticals. J. Laboratory Automation 17 (6), 449–457. doi: 10.1177/2211068212460037. CrossRef. Google Scholar.
247. Worbs S., Kohler K., Pauly D., Avondet M. A., Schaer M., Dorner M. B. et al (2011). Ricinus communis intoxications in human and veterinary medicine-a summary of real cases. Toxins (Basel) 3 (10), 1332–1372. [PMID: 22069699]. doi: 10.3390/toxins3101332. CrossRef. Google Scholar.
248. Wróblewska B., Kubicka E., Semenowicz E., Ogrodowczyk A., Mikołajczyk A., Złotkowska D. (2022). The immune system response to 15-kDa barley protein: A mouse model study. Nutrients 14 (20), 4371. [PMID: 36297055]. doi: 10.3390/nu14204371. CrossRef. Google Scholar.
249. Xu X., Gan Q., Clough R. C., Pappu K. M., Howard J. A., Baez J. A. et al (2011). Hydroxylation of recombinant human collagen type I alpha 1 in transgenic maize co-expressed with a recombinant human prolyl 4-hydroxylase. Bmc Biotechnol. 11, 69. [PMID: 21702901]. doi: 10.1186/1472-6750-11-69. CrossRef. Google Scholar.
250. Yang H., Zhang L., Galinski M. (2010). A probabilistic model for risk assessment of residual host cell DNA in biological products. Vaccine 28 (19), 3308–3311. [PMID: 20226252]. doi: 10.1016/j.vaccine.2010.02.099. CrossRef. Google Scholar.
251. Zalai D., Dietzsch C., Herwig C. (2013). Risk-based process development of biosimilars as part of the quality by design paradigm. Pda J. Pharm. Sci. Tech. 67 (6), 569–580. [PMID: 24265299]. doi: 10.5731/pdajpst.2013.00943. CrossRef. Google Scholar.
252. Zeng S., Ma J., Yang Y., Zhang S., Liu G-J., Chen F. (2019). Spatial assessment of farmland soil pollution and its potential human health risks in China. Sci. Total Environ. 687, 642–653. [PMID: 31220718]. doi: 10.1016/j.scitotenv.2019.05.291. CrossRef. Google Scholar.
253. Zhang Z., Wu X., Wu Q., Huang X., Zhang J., Fang H. (2020). Speciation and accumulation pattern of heavy metals from soil to rice at different growth stages in farmland of southwestern China. Environ. Sci. Pollut. Res. Int. 27 (28), 35675–35691. [PMID: 32601861]. doi: 10.1007/s11356-020-09711-2. CrossRef. Google Scholar.
254. Zhu X., Li X., Wang W., Ning K. (2020). Bacterial contamination screening and interpretation for biological laboratory environments. Med. Microecology 5, 100021. doi: 10.1016/j.medmic.2020.100021. CrossRef. Google Scholar.
255. Ziegler J., Facchini P. J., Geissler R., Schmidt J., Ammer C., Kramell R. et al (2009). Evolution of morphine biosynthesis in opium poppy. Phytochemistry 70 (15-16), 1696–1707. [PMID: 19665152]. doi: 10.1016/j.phytochem.2009.07.006. CrossRef. Google Scholar.
256. Zimmermann L., Bartosova Z., Braun K., Oehlmann J., Völker C., Wagner M. (2021). Plastic products leach chemicals that induce in vitro toxicity under realistic use conditions. Environ. Sci. Technol. 55 (17), 11814–11823. [PMID: 34488348]. doi: 10.1021/acs.est.1c01103. CrossRef. Google Scholar.

Buyel JF (2023) Product safety aspects of plant molecular farming. Front. Bioeng. Biotechnol. 11:1238917. doi: 10.3389/fbioe.2023.1238917
Перевод статьи «Product safety aspects of plant molecular farming.» автора Buyel JF, оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык
Фото: Designed by Freepik

















































































































Комментарии (0)