Как органическое земледелие помогает перцу побеждать фитофтороз с помощью полезных бактерий
Почвенные заболевания часто менее выражены в органических хозяйствах, возможно, благодаря привлечению полезных микроорганизмов сельскохозяйственными культурами. В данном исследовании изучалась супрессивность (способность подавлять болезнь) почв из систем органического, интегрированного и традиционного земледелия в отношении фитофтороза перца (Phytophthora capsici) в условиях камерных вегетационных опытов.
Аннотация
Заболеваемость была на 41,3 % и 34,1 % ниже в почве из органической системы земледелия по сравнению с почвой из интегрированной или традиционной систем соответственно. Бета-разнообразие ризосферных микробных сообществ различалось между вариантами, при этом в органической почве наблюдалось обогащение такими таксонами, как Bacillus, Sporosarcina, Acidobacteria групп Gp5, Gp6, Gp22 и Ignavibacterium. Культурально-зависимый анализ показал, что 50,3 % in vitro антагонистов P. capsici, выделенных из ризосферы здоровых растений перца, принадлежали к роду Bacillus. Комплексный анализ антагонистов in vitro и бактериального разнообразия показал, что количество антагонистических Bacillus было выше в ризосфере перца, обработанного органической почвой. Микробный консорциум из 18 in vitro антагонистов рода Bacillus значительно повысил супрессивность почвы из интегрированной системы земледелия в отношении фитофтороза перца. В целом, микробиом почвы в условиях долгосрочной органической системы земледелия в большей степени подавлял фитофтороз, что, вероятно, обусловлено антагонизмом Bacillus в ризосфере. Данное исследование даёт представление об управлении микробиомом для подавления болезней в тепличных условиях.
Введение
Растущее количество данных свидетельствует о том, что почвенные микробиомы или микробиомы, ассоциированные с сельскохозяйственными культурами, играют ключевую роль в здоровье растений (Gómez Expósito и др., 2017). Исследования бактериального разнообразия показывают, что такие таксономические группы, как Bacteroidetes (Yin и др., 2013; Xiong и др., 2015), Actinobacteria (Mendes и др., 2011; Xiong и др., 2015, 2017; Cha и др., 2016; van der Voort и др., 2016), Acidobacteria (Shen и др., 2015; van der Voort и др., 2016; Xiong и др., 2017), Firmicutes (Mendes и др., 2011; Shen и др., 2015; Xiong и др., 2015, 2017) и Proteobacteria (Mendes и др., 2011; Michelsen и др., 2015), вероятно, вовлечены в подавление этих болезней растений. Более того, Bacillus был описан как важные бактерии, способствующие росту растений (Fan и др., 2018). Тяжесть почвенных болезней растений, таких как вызываемые Rhizoctonia solani (Postma и др., 2008), Verticillium albo-atrum (Postma и др., 2008), Phytophthora infestans (Wang и др., 2001) и Fusarium (Champeil и др., 2004; Lenc и др., 2015), в органических хозяйствах часто была ниже, чем в традиционных хозяйствах. В органических хозяйствах большое количество органического удобрения может способствовать развитию супрессивности болезней, часто называемой общей супрессивностью, путем неспецифической активации активности аборигенных почвенных микроорганизмов (Qiu и др., 2012; Wang и др., 2015; Liu и др., 2018). Таким образом, считалось, что общая супрессивность обусловлена не столько несколькими таксономическими группами, сколько специфической супрессивной почвой (Weller и др., 2002). Однако долгосрочное применение органических удобрений может вызывать глубокие изменения в почвенных микробных сообществах, что подтверждено как полевыми, так и тепличными экспериментами (Marschner и др., 2003; Ndubuisi-Nnaji и др., 2011; Hartmann и др., 2014; Luo и др., 2015), и некоторые изменения микробных сообществ были связаны с подавлением болезней (Zhang и др., 2017). Между тем, изменения физико-химических свойств почвы, таких как содержание органического вещества и питательных веществ, сопровождались сдвигами в почвенных микробиомах (Fernandez и др., 2016; Li и др., 2017; Pronk и др., 2017). Изменения физико-химических свойств, вероятно, формируют биогеохимические интерфейсы в почве, где обитают микроорганизмы; таким образом, такие изменения могут также влиять на активность и судьбу как фитопатогенов (Strunnikova и др., 2015), так и других микроорганизмов (Pronk и др., 2017). В целом, до сих пор неясно, в какой степени изменения почвенного микробиома после долгосрочного органического земледелия могут способствовать супрессивности почвенных болезней.
Долгосрочный тепличный эксперимент был заложен в 2002 году на экспериментальной станции Цюйчжоу в провинции Хэбэй, Китай. Этот эксперимент включал органическую, интегрированную и традиционную системы земледелия для оценки супрессивности почвы к фитофторозу перца. Все три системы земледелия находились в одинаковых условиях севооборота, орошения и обработки почвы, но различались по управлению удобрениями и защитой растений. Различия как в микробной структуре, так и в физико-химических свойствах (Han и др., 2017) были обнаружены между этими различными системами земледелия. Заболеваемость ложной мучнистой росой, листовой плесенью, фитофторозом, мучнистой росой и бактериальной угловатой пятнистостью листьев на томате (Solanum lycopersicum L.) или огурце (Cucumis sativus L.) часто была ниже в органической системе земледелия, чем в традиционной (Yang и др., 2009a,b). Этот долгосрочный эксперимент предоставил возможность изучить механизмы, лежащие в основе влияния органической почвы на здоровье растений, которые зависят от почвенных микробиомов, сформированных долгосрочным органическим земледелием.
Здесь мы провели вегетационный эксперимент в контролируемых условиях для оценки влияния почвенного микробиома из органической, интегрированной и традиционной систем земледелия на подавление фитофтороза перца — серьезного заболевания, вызываемого Phytophthora capsici, которое ответственно за мировые потери более 100 миллионов долларов США только на перце (Barchenger и др., 2017). Чтобы смягчить потенциальное влияние физико-химических свойств почвы, растения перца выращивали в смеси песка и почвы в соотношении 5:1,2 (об.:об.). Ризосферные микробиомы при различных обработках изучались перекрестно с помощью культурально-зависимого анализа и высокопроизводительного секвенирования. В этом исследовании наши конкретные вопросы были следующими: (1) подавляет ли и в какой степени почвенный микробиом, сформированный долгосрочным органическим земледелием, фитофтороз перца; (2) влияют ли различные обработки инокуляцией почвенным микробиомом на формирование ризосферного микробного сообщества перца; и (3) какие таксоны ассоциированы с подавлением фитофтороза перца в почве, управляемой органически?
Материалы и методы
Долгосрочный эксперимент по органическому земледелию, отбор проб и биоанализ
Образцы почвы были отобраны из долгосрочного тепличного эксперимента, заложенного в 2002 году на экспериментальной станции Цюйчжоу (36°52′ с.ш., 115°01′ в.д.), Хэбэй, Китай. Обработки включали органическую, интегрированную и традиционную системы земледелия. Подробности о сельскохозяйственном управлении в каждом случае были описаны Han и др. (2017). Вкратце, все системы земледелия следовали одинаковым схемам севооборота, орошения и обработки почвы. Органическая система земледелия характеризовалась внесением компоста (165 т га⁻¹ год⁻¹) из куриного и коровьего навоза, а также биологическими и физическими методами защиты растений (желтые клейкие ловушки и противомоскитные сетки для борьбы с вредителями; Kocide, содержащий гидроксид меди, механическое удаление больных растений и гребневая посадка для борьбы с болезнями растений; механическая/мульчирующая обработка для подавления сорняков). Традиционное земледелие следовало местному стилю ведения тепличного овощеводства и использовало химические удобрения (мочевина, суперфосфат и хлорид калия), пестициды (диметоморф, лямбда-цигалотрин, имидаклоприд и карбендазим) и 46,8 т га⁻¹ год⁻¹ куриного и коровьего навоза. Наконец, интегрированная система земледелия использовала половину количества химических удобрений и пестицидов, используемых в традиционной системе, и половину количества органических удобрений, используемых в органической системе. Для каждой системы земледелия в июне 2017 года было отобрано в общей сложности 75 кернов почвы (диаметром 2 см) из верхнего слоя почвы (глубина 1–20 см). Все образцы почвы были просеяны через сито с ячейками 2 мм для удаления камней и растительных остатков. Просеянные образцы хранились при 4°C до проведения вегетационного эксперимента (Ding и др., 2010).
Биоанализ проводился следующим образом: семена перца "Cayenne" (C. annuum L.) (Zhong liang xin) высевались в горшки (диаметром 12 см и высотой 15 см), содержащие смесь 500 г стерилизованного песка и 120 г почвы, и выращивались в течение 28 дней в климатической камере (Hangzhou Lvbo Instrument Co., Ltd., LB-1000D-LED) при 30°C, 95% относительной влажности и 12-часовом световом периоде (15000 лк). Все растения поливались дважды стандартным раствором Хогланда. Почва использовалась как инокулят микробиома. Все горшки были полностью рандомизированы внутри климатической камеры. По шесть сеянцев на горшок сохранялось до 20 дней после посева; восемь из 20 горшков были случайным образом отобраны для заражения P. capsici. Зооспоры P. capsici культивировали и готовили, как описано ранее (Bi и др., 2014). 6 мл суспензии зооспор (приблизительно 5 × 10⁵ зооспор/мл) вносили вблизи корня каждого сеянца. Заболеваемость оценивали через 7 дней после инокуляции. Параллельно проводили четыре отбора проб ризосферы неинокулированных сеянцев на 27, 31, 35 и 38 день после посева (соответственно T1, T2, T3 и T4). Отбирали три горшка, и все шесть сеянцев в каждом горшке использовали как повторность для каждой обработки и отбора проб. Отбор ризосферных образцов проводили следующим образом: растения перца осторожно извлекали из горшков и встряхивали, чтобы освободить корни от как можно большего количества почвы, и только те частицы песка или почвы, которые сильно прилипали к корням, оставались прикрепленными к корням. Затем корни энергично промывали 0,85% раствором NaCl. После удаления корневых систем смесь центрифугировали при 6000 × g в течение пяти минут. Осадок хранили при -20°C для экстракции тотальной ДНК сообщества. Ризосферные образцы растений, не имеющих симптомов болезни через 9 дней после инокуляции P. capsici, собирали отдельно (Bi и др., 2014). Часть ризосферных образцов от здоровых растений перца смешивали с равным объемом 40% глицерина и немедленно замораживали при -80°C.
Анализ ампликонов бактериального гена 16S рРНК методом высокопроизводительного секвенирования
Тотальную ДНК микробного сообщества из ризосферных образцов экстрагировали с использованием набора FastDNA spin Kit для почвы (MP, Biomedicals, Санта-Ана, Карлсбад, Калифорния, США) в соответствии с инструкциями производителя. Анализ высокопроизводительного секвенирования проводился на платформе Hiseq2500. Вкратце, универсальные праймеры 515F (5′-GTGCCAGCMGCCGCGGTAA-3′) и 909R (5′-CCCCGYCAATTCMTTTRAGT-3′) с уникальным 12-нуклеотидным штрих-кодом (Caporaso и др., 2010) использовались для амплификации фрагментов гена 16S рРНК. Все прочтения последовательностей были обрезаны и распределены по каждому образцу на основе штрих-кодов. Высококачественные последовательности (длина > 300 п.н., без неоднозначного основания "N" и средний показатель качества оснований > 30) использовались для последующего анализа. Формирование таксономических OTU выполнялось, как описано ранее, с использованием набора общедоступных баз данных и программного обеспечения (Schloss и др., 2009; Caporaso и др., 2010; Ding и др., 2012; Cole и др., 2014; Yilmaz и др., 2014; Rognes и др., 2016). Сравнение структуры сообществ, идентификация таксонов со значительно различающейся относительной численностью и сетевой анализ выполнялись, как описано ранее (Kropf и др., 2004; Hothorn и др., 2017). Сеть совместной встречаемости была построена с использованием рангового коэффициента корреляции Спирмена (cor > 0.6, p < 0.01) (Barberán и др., 2012). Эта сеть была проанализирована с использованием программного обеспечения gephi (версия 0.91) (Bastian и др., 2009). Структура сообщества была визуализирована с помощью анализа главных координат (PCoA) на основе попарного расстояния Брея-Кертиса, рассчитанного по относительной численности различных OTU. Все статистические анализы и построение графиков выполнялись с использованием программного обеспечения R 3.1.21, и эти инструменты были реализованы в экземпляре galaxy в соответствии с описанием команды разработчиков galaxy. Все последовательности были отправлены в NCBI SRA (PRJNA497732).
Выделение, скрининг, BOX-PCR и анализ секвенирования 16S рРНК in vitro антагонистов P. capsici
Серийные разведения ризосферных образцов от растений перца, свободных от симптомов болезни, высевали на среду R2A (Beijing Land Bridge Technology Co., Ltd.) с добавлением 100 мг/л циклогексимида с последующей инкубацией при 30°C в течение 2–3 дней. Все колонии с разной морфологией были отобраны для дальнейшего анализа. In vitro антагонисты против P. capsici были отобраны в соответствии с Yang и др. (2012). Геномную ДНК антагонистов экстрагировали с использованием набора для экстракции бактериальной геномной ДНК (Beijing Biomed Co., Ltd.). BOX-PCR-анализ проводили в соответствии с Adesina и др. (2007) с использованием праймеров BOX-A1R (5′-CTACGGCAAGGCGACGCTGACG-3′) (Louws и др., 1994). Цифровые изображения далее анализировались с помощью программного пакета GelCompar II (Applied Maths, Кортрейк, Бельгия, Китай) для распределения бактериальных изолятов по различным BOX-PCR-паттернам, когда сходство профилей было менее 80%. Для каждого BOX-PCR-паттерна один изолят был случайным образом отобран для анализа секвенирования по Сэнгеру гена 16S рРНК, амплифицированного праймерами 27F (5′-AGAGTTTGATCATGGCTCAG-3′) и 1492R (5′-TACGGTTACCTGTTACGACTT-3′) (Lane, 1991). Филогенетическое дерево было построено с использованием программного обеспечения Mega 6.0 (Tamura и др., 2013). BOX-PCR использовался для изучения бактерий одного вида, у которых гены 16S рРНК высоко сходны. Здесь субпоследовательности между 515F и 909R были выбраны для идентификации уникальных филотипов с помощью программного обеспечения VSEARCH (Rognes и др., 2016). Эти уникальные филотипы были сопоставлены с библиотекой последовательностей 16S рРНК с минимальным идентичностью последовательности 99% с использованием автономного анализа BLASTN.
Эксперимент по биологическому контролю в тепличных условиях
Восемнадцать антагонистов Bacillus (11 Bacillus methylotrophicus, три Bacillus licheniformis, два Bacillus aerophilus, один Bacillus cereus и один Bacillus subtilis), выделенных от здоровых растений перца, выращенных в смеси с почвой из органической системы земледелия, были отобраны для создания микробного консорциума, способность которого улучшать супрессивность почвы из интегрированной системы земледелия к фитофторозу перца была оценена. Наиболее высокая заболеваемость наблюдалась в почве из интегрированной системы земледелия в двух предыдущих биоанализах. Споры Bacillus готовили в соответствии с Paidhungat и др. (2000). Когда сеянцы перца достигли 20-дневного возраста, вносили восемь миллилитров суспензии, содержащей споры Bacillus (приблизительно 2 × 10⁸ спор/мл). Заражение P. capsici и оценка заболеваемости проводились в соответствии с вышеприведенным описанием.
Результаты
Фитофтороз перца был менее тяжелым у растений, выращенных в органической почве
Два независимых биоанализа с почвами из органической, интегрированной и традиционной систем земледелия были проведены для оценки их супрессивности к фитофторозу перца, вызываемому P. capsici. Оба эксперимента показали, что почва из органической системы земледелия была более супрессивной, чем почва из интегрированной или традиционной системы (Рисунок 1A,B). В первом биоанализе процент заболеваемости составил 56,3% в органической почве, что было на 41,3 и 34,2% ниже, чем в интегрированной или традиционной системе соответственно (Рисунок 1A). Кроме того, во втором биоанализе процент заболеваемости составил 54,2% в органической почве, 87,5% в почве из интегрированной системы и 81,3% в почве из традиционной системы (Рисунок 1B).
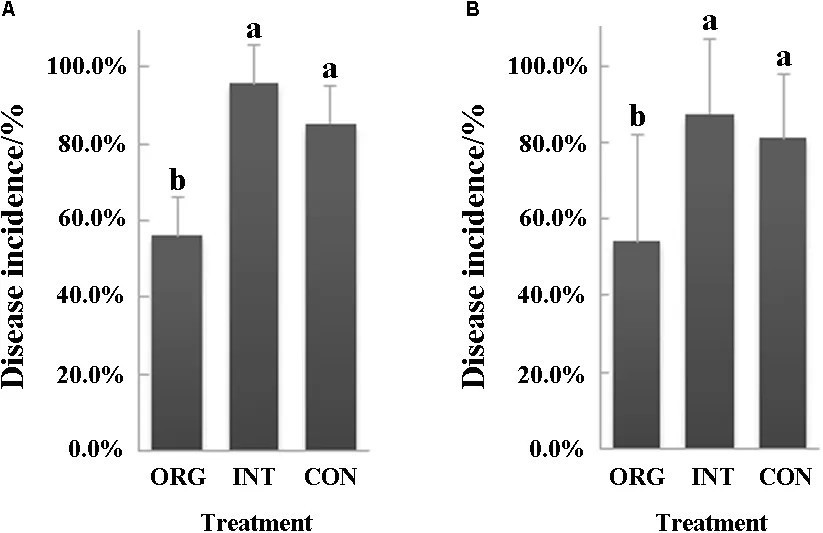
РИСУНОК 1 Процент (среднее ± стандартное отклонение, N = 8) сеянцев перца с симптомами фитофтороза (P. capsici) в почвах из органической (ORG), интегрированной (INT) и традиционной (CON) систем земледелия (p < 0.05, множественный диапазонный тест Дункана) в первом (A) и втором (B) биоанализе. Значимые различия обозначены разными буквами.
Бета-разнообразие ризосферных микробных сообществ различалось между обработками почвой
Всего было получено 656 105 последовательностей гена 16S рРНК из 36 ризосферных образцов, отобранных в четыре срока. Здесь мы исключили 60 322 последовательности, относящиеся к Chloroplast или Cyanobacteria, из последующего анализа. Более подробная информация о секвенировании, такая как количество прочтений, покрытие и неклассифицированные таксоны, показана в Дополнительной таблице S1. Для всех образцов доминировали Proteobacteria (46%), за которыми следовали Bacteroidetes (10,7%), Firmicutes (10,2%), Acidobacteria (8,1%) и Actinobacteria (6,1%) (Рисунок 2A). PCoA на основе расстояния Брея-Кертиса показала, что ризосферное бактериальное сообщество различалось между обработками (Рисунок 2B), при этом большее колебание было обнаружено в почве из традиционной системы (Рисунок 2B). Дисперсионный анализ показал, что обработка и срок отбора проб объясняли 22% и 10% общего изменения ризосферного микробного сообщества соответственно. Пермутационный анализ также подтвердил, что структура сообщества значительно различалась между обработками (Дополнительная таблица S2). Индекс богатства Chao1 в ризосферном бактериальном сообществе был немного выше в обработке органической почвой по сравнению с почвой из традиционной системы (Рисунок 2C). Выравненность Пиелу была немного ниже в обработке органической почвой по сравнению с почвой из интегрированной системы (Рисунок 2D).
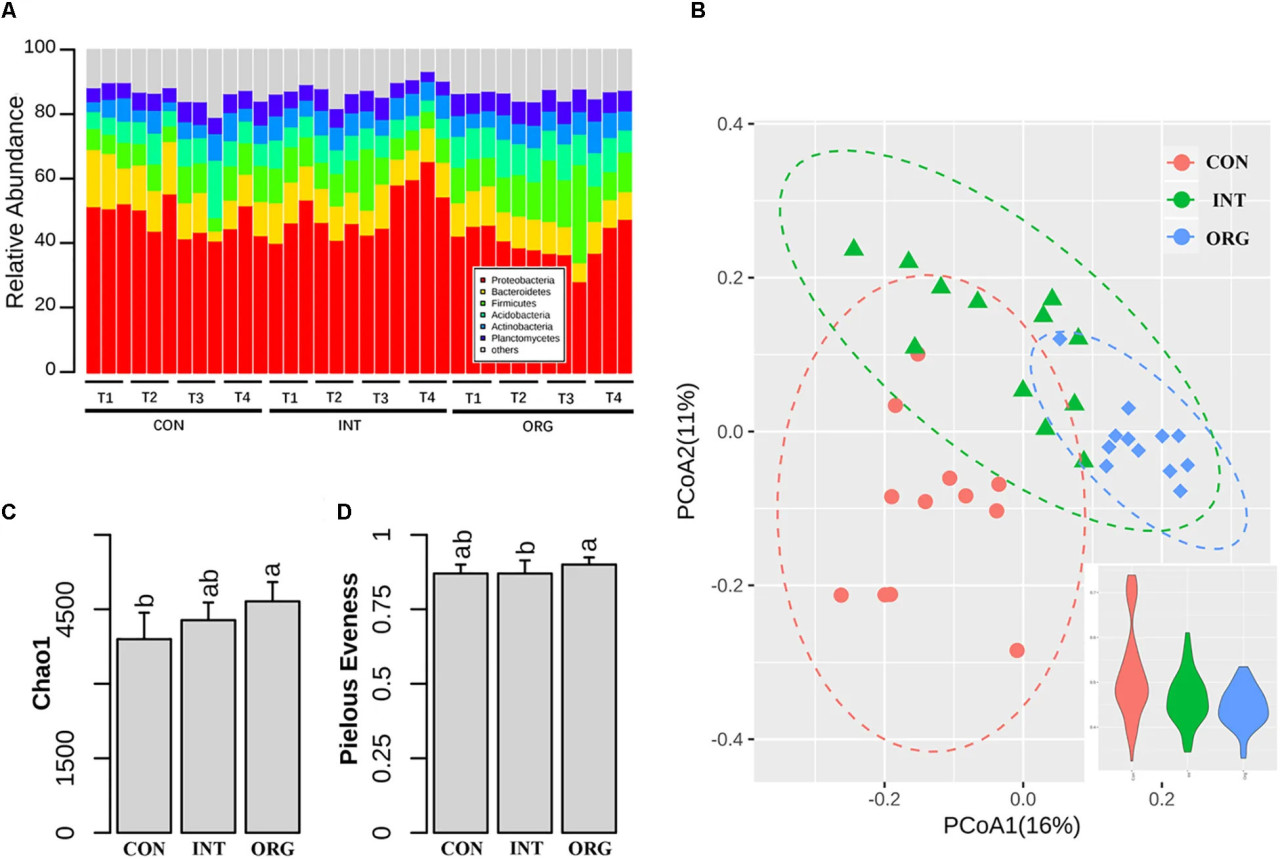
РИСУНОК 2 Относительная численность различных филумов (A), PCoA-анализ микробного сообщества и вариация внутри обработок (B), индекс видового богатства (C) и индекс выравненности (D) в ризосфере сеянцев перца в почвах из органической (ORG), интегрированной (INT) и традиционной (CON) систем земледелия в течение четырех периодов отбора проб, т.е. T1 (27 дней после посева), T2 (31 день после посева), T3 (35 дней после посева) и T4 (38 дней после посева). Значимые различия обозначены разными буквами.
Таксоны, ассоциированные с подавлением фитофтороза перца
Роды, которые значимо коррелировали (коэффициент корреляции Спирмена > 0,6 и p < 0,001) друг с другом, были подвергнуты сетевому анализу. В общей сложности 25 коррелирующих родов образовали семь микробных узлов (Рисунок 3A). Относительная численность пяти узлов различалась между обработками (Рисунок 3B). Узел, состоящий из Bacillus, Sporosarcina, Hyphomicrobium, Gaiella, Pirellula и Blastopirellula, был значительно более обильным в обработке органической почвой по сравнению с двумя другими обработками, в отличие от узла, включающего Rhizobium, Sphingobium, Pseudoxanthomonas и Dyadobacter (Рисунок 3B). Два других узла (один состоял из Aquabacterium и Noviherbaspirillum, а другой включал Luteimonas и Lysobacter) были значительно более обильными в обработке почвой из интегрированной системы, в отличие от узла, состоящего из Acidobacteria Gp6, Gemmatimonas и WPS-2_genera_incertae_sedis (Рисунок 3B). Множественные сравнения были выполнены для выявления таксонов со значительно различающейся относительной численностью между обработками (Рисунок 3C). Одиннадцать родов (Bacillus, Sporosarcina, Gaiella, Blastopirellula, Rhizobium, Sphingobium, Pseudoxanthomonas, Dyadobacter, Luteimonas, Noviherbaspirillum и Acidobacteria Gp6) показали сходные закономерности с соответствующими микробными узлами (Рисунок 3C). Кроме того, наименьшая относительная численность других родов, относящихся к Proteobacteria, таких как Rheinheimera, Pseudomonas, Ensifer и Pseudoxanthomonas, была обнаружена для обработки органической почвой, в отличие от Microvirga (Рисунок 3C). Две другие подгруппы Acidobacteria (Gp5 и Gp22), а также Ignavibacterium, Zavarzinella и Levilinea были более обильны в обработке органической почвой, в отличие от Acidobacteria Gp4 (Рисунок 3C). Наибольшая численность Flavobacterium часто обнаруживалась в обработке почвой из традиционной системы, за исключением T4 (Рисунок 3C).
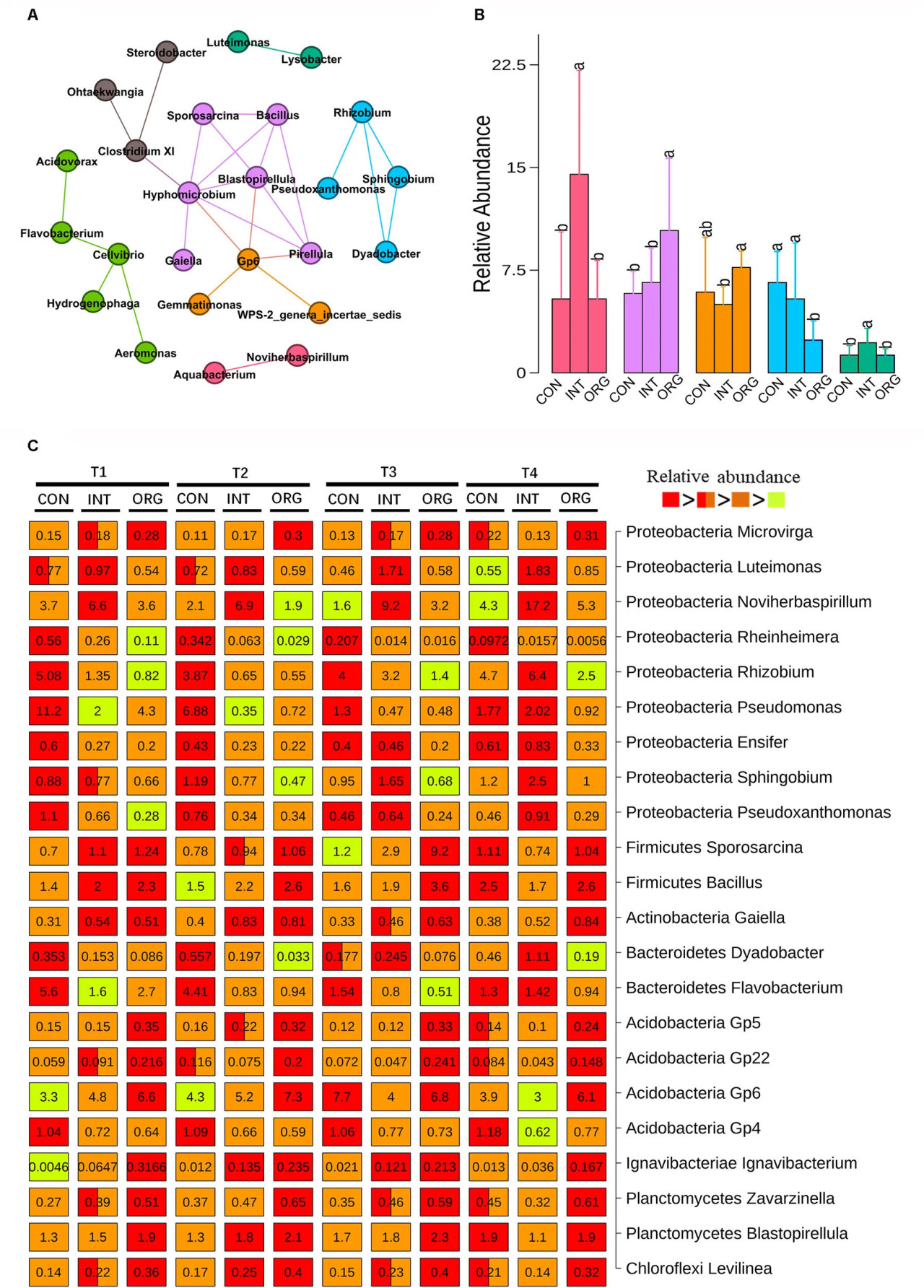
РИСУНОК 3 Микробные узлы (A) и их относительная численность (B) в ризосфере сеянцев перца в почвах из органической (ORG), интегрированной (INT) и традиционной (CON) систем земледелия на стадиях T1 (27 дней после посева), T2 (31 день после посева), T3 (35 дней после посева) и T4 (38 дней после посева), согласно анализу сети совместной встречаемости. Каждый цвет представляет собой отдельный микробный узел, а связи указывают на значимость (p < 0.001), коэффициент ранговой корреляции Спирмена > 0.6, совместную встречаемость между таксонами. (C) Роды со значительно различающейся относительной численностью между обработками в четыре срока отбора проб. Числа на рамках показывают относительную численность, выраженную в процентах. Значимое различие обозначено разным цветом. Рамка с двумя цветами указывает на отсутствие значимого различия с другой обработкой, содержащей один из двух цветов. Значимые различия обозначены разными буквами.
In vitro антагонисты P. capsici в ризосфере здоровых растений перца
Ризосферные образцы от здоровых растений были проанализированы культурально-зависимым подходом. Всего было получено 1215 бактериальных изолятов. Среди них 151 изолят показал in vitro антагонистическую активность против P. capsici. BOX-PCR-анализ далее распределил эти антагонисты по 49 паттернам (Рисунок 4A и Дополнительная таблица S3), при этом только пять паттернов были общими для различных обработок (Рисунок 4B). Большинство паттернов были специфически обнаружены в каждой обработке (Рисунок 4B). Ампликоны 16S рРНК репрезентативных изолятов для каждого BOX-PCR-паттерна были подвергнуты секвенированию по Сэнгеру. В совокупности эти in vitro антагонисты были распределены по 18 родам, причем большинство из них относились к следующим таксонам: Bacillus (76 изолятов, 50,3%), Exiguobacterium (22 изолята, 14,6%), Beijerinckia (13 изолятов, 8,6%), Stenotrophomonas (6,6%), Arthrobacter (4,6%) и Pseudomonas (3,3%) (Рисунок 4C). Филогенетический анализ дополнительно показал, что гены 16S рРНК изолятов из 26 BOX-PCR-паттернов были высоко сходны с B. subtilis (AJ276351), B. methylotrophicus (EU194897), B. thuringiensis (D16281), B. cereus (AE016877), B. licheniformis (CP000002), B. aerophilus (AJ831844) и B. firmus (D16268) (Рисунок 4A). Изоляты одного BOX-PCR-паттерна показали высокое сходство с Pseudomonas geniculate (AB021404) (Рисунок 4A). Изоляты семи BOX-PCR-паттернов были сходны со штаммами Actinobacteria — филумом, известным как содержащий бактерии, продуцирующие антибиотики (Рисунок 4A). В целом, эти результаты показали, что в ризосфере здоровых растений перца существует богатое разнообразие in vitro антагонистов, преимущественно относящихся к Bacillus, Pseudomonas и Actinobacteria.
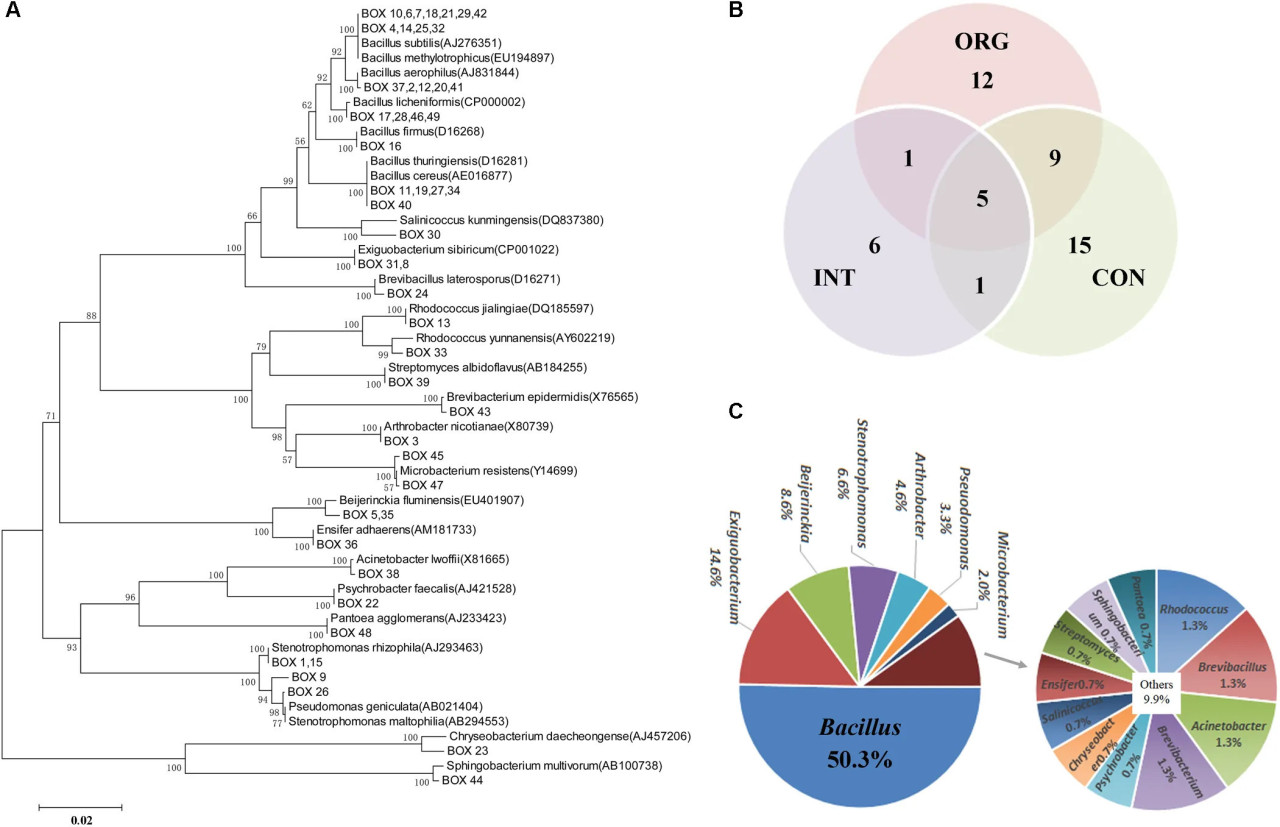
РИСУНОК 4 (A) Филогенетический анализ гена 16S рРНК антагонистов в каждом BOX-PCR-паттерне, число на узлах = (значение бутстрапа/1000) × 100; (B) Диаграмма Венна, показывающая количество in vitro антагонистов с уникальным BOX-PCR-паттерном среди обработок почвами из органической (ORG), интегрированной (INT) и традиционной (CON) систем земледелия. (C) Процент in vitro антагонистов, относящихся к различным родам.
Обогащение in vitro антагонистов, относящихся к Bacillus, в ризосферном микробном сообществе
Чтобы оценить распространенность этих антагонистов в ризосфере, гены 16S рРНК антагонистов были сопоставлены с библиотеками последовательностей 16S рРНК. Двадцать пять уникальных филотипов были извлечены из субпоследовательностей (между 515F и 909R) гена 16S рРНК репрезентативных изолятов 49 BOX-PCR-паттернов. Все филотипы были сопоставлены с библиотеками последовательностей 16S рРНК. В общей сложности 19 923 прочтения в библиотеках последовательностей 16S рРНК были сопоставлены, что составляло от 2,5 до 8,7% от общего числа прочтений в каждой библиотеке. Средняя относительная численность прочтений, сопоставленных с in vitro антагонистами, составляла всего 2,0% в обработке органической почвой, что было значительно ниже, чем в обработке из интегрированной или традиционной системы (Рисунок 5A). Сходные тенденции наблюдались для антагонистов, относящихся к Rhizobium и Ensifer (Рисунок 5B,C). Интересно, что относительная численность Bacillus была значительно выше в почве из органической системы (Рисунок 5D). Дальнейший анализ показал, что два филотипа (B. cereus и B. firmus) были значительно более обильны в обработке органической почвой, чем в почве из интегрированной или традиционной системы (Рисунок 5E). Филотип, сходный с B. cereus (AE016877), соответствовал четырем BOX-PCR-паттернам (11, 19, 27 и 34) (Рисунок 4A). Другой филотип, сходный с B. firmus (D16268), соответствовал только BOX-PCR-паттерну 16 (Рисунок 4A). Поскольку наименьшая заболеваемость наблюдалась в обработке почвой из органической системы, эти результаты показали, что антагонисты, относящиеся к Bacillus, возможно, участвовали в подавлении фитофтороза перца в данном случае. Микробный консорциум, состоящий из 18 антагонистов Bacillus, был оценен на его способность улучшать супрессивность почвы из интегрированной системы к фитофторозу перца. В двух описанных выше экспериментах наиболее высокая заболеваемость была обнаружена в почве из интегрированной системы. Здесь уровень заболеваемости при обработке почвы консорциумом Bacillus составил только 38,9%, что было значительно ниже, чем в обработке только почвой из интегрированной системы (97,2%) (Рисунок 5F). Эти результаты показали, что эти антагонисты Bacillus эффективно улучшали способность почвы из интегрированной системы подавлять фитофтороз перца.
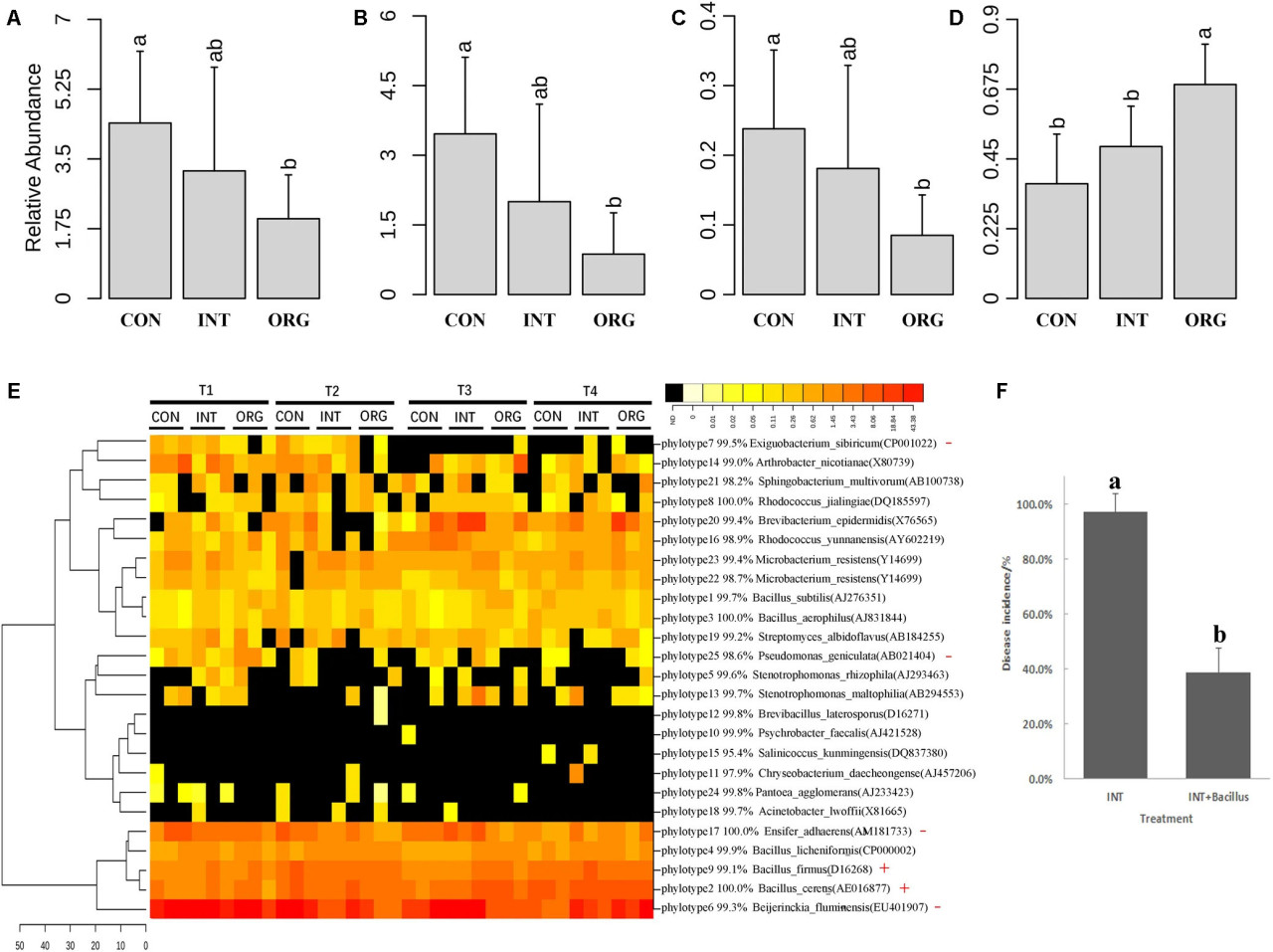
РИСУНОК 5 Средняя относительная численность прочтений, сопоставленных с in vitro антагонистами (A) и с антагонистами, относящимися к Rhizobium (B); Ensifer (C); и Bacillus (D). (E) Тепловая карта анализа in vitro антагонистов, принадлежащих к каждому филотипу. Филотип со значительно более высокой или более низкой относительной численностью в обработке органической почвой обозначен красным крестом (+) или минусом (-) соответственно. (F) Процент (среднее ± стандартное отклонение, N = 6) сеянцев перца с симптомами фитофтороза (P. capsici) в почвах из интегрированной системы (INT) отдельно и инокулированных консорциумом Bacillus (INT + Bacillus) (p < 0.05, множественный диапазонный тест Дункана). Значимые различия обозначены разными буквами.
Обсуждение
Это исследование было основано на долгосрочном тепличном эксперименте, в котором несколько схем управления сельским хозяйством, включая севооборот, обработку почвы и орошение, были одинаковыми для всех трех систем с 2002 года. Наиболее существенное различие между тремя системами заключается в удобрении, борьбе с вредителями и болезнями растений. Считается, что этот сравнительно долгосрочный эксперимент ослабляет неожиданные эффекты, такие как исходные различия между почвами (Schreiter и др., 2014; Schlemper и др., 2017), растениями (Weinert и др., 2011) и стратегиями управления (Hartmann и др., 2014). По сравнению с традиционной системой, тяжесть болезней была ниже в органической системе (Yang и др., 2009a,b). Таким образом, этот сравнительный эксперимент может предоставить возможность изучить механизмы влияния почвенных микроорганизмов на здоровье растений, что является необходимым условием для конструирования почвенного микробиома для подавления почвенных болезней растений. Здесь мы использовали дополнительные методы, включая вегетационные эксперименты, высокопроизводительное секвенирование и культурально-зависимый анализ для изучения микробного разнообразия, ассоциированного с подавлением почвой фитофтороза перца. Наше исследование предоставляет новое понимание механизмов подавления болезней почвой, сформированной долгосрочным органическим земледелием.
Сельскохозяйственное управление и подавление почвенных болезней
Почвенный микробиом в органической системе земледелия показал большую способность подавлять фитофтороз перца. Сходные результаты наблюдались и для других почвенных болезней в органических хозяйствах (Wang и др., 2001; Champeil и др., 2004; Postma и др., 2008; Lenc и др., 2015). В целом, природные почвы могут подавлять почвенные болезни растений, что подтверждается наблюдением, что растения, выращенные в стерилизованных почвах, более восприимчивы к патогенам, чем растения, растущие в нестерилизованных почвах (Thuerig и др., 2009; Mendes и др., 2011). Такая супрессивность почвы была связана с активностью аборигенных почвенных микроорганизмов. В органических системах земледелия несколько практик управления сельским хозяйством, таких как использование органических удобрений (Sharma и др., 2012; Wang и др., 2015), совместное выращивание культур (Lithourgidis и др., 2011; Boudreau, 2013) и севооборот (Ball и др., 2005), применялись для сохранения благополучия агроэкосистем, и некоторые из этих процедур действительно усиливали способность почвенного микробиома подавлять патогены растений (van Bruggen и Finckh, 2016). Среди них использование компоста способствовало подавлению нескольких болезней растений, вызываемых Pythium ultimum, Rhizoctonia solani, Verticillium dahlia, Fusarium oxysporum, Ralstonia solanacearum и P. capsici (Noble и Coventry, 2010; Sang и др., 2010; Liu и др., 2016). В настоящем исследовании большое количество (приблизительно 165 т га⁻¹) компоста вносилось ежегодно в органическую систему с 2002 года. Подавление фитофторозных болезней компостом хорошо установлено (Chae и др., 2006; Sang и др., 2010; Hadar и Papadopoulou, 2012). Предполагалось, что механизм, благодаря которому компост оказывает столь заметное влияние на подавление болезней, является многомерным: добавление питательных веществ, антибиотики, антагонисты и изменение физико-химических свойств почвы (Noble и Coventry, 2010). Здесь влияние физико-химических свойств почвы было смягчено путем выращивания перца в смеси, состоящей в основном из песка, за исключением небольшой фракции почвы, используемой в качестве инокулята микробиома. Такая процедура может позволить нам различить влияние почвенного микробиома на здоровье растений. Тем не менее, стоит отметить, что супрессивность почвы к болезням при долгосрочном органическом земледелии является высоко сложной, и в силу своих внутренних ограничений эта упрощенная процедура не позволяет полностью понять сложные взаимодействия между физико-химическими свойствами почвы, растениями и микробиомом. Как физико-химические свойства почвы, так и растения, вероятно, влияют на структуру и функцию почвенного микробного сообщества (Babin и др., 2012; Ding и др., 2013; Pronk и др., 2017). Тем не менее, представленные здесь результаты показывают, что изменения в почвенном микробном сообществе, связанные с долгосрочным органическим земледелием, вероятно, способствовали супрессивности почвы к фитофторозу перца, вызываемому P. capsici.
Bacillus и подавление фитофтороза
В последнее время супрессивность почвы к почвенным болезням была связана с коллективными изменениями в почвенном или ризосферном микробном сообществе (Yim и др., 2015; Cha и др., 2016; Xiong и др., 2017; Gouda и др., 2018), которые чрезвычайно сложны и часто содержат одновременно многие тысячи различных OTU. Анализ бактериального разнообразия с помощью технологий высокопроизводительного секвенирования (Yim и др., 2015) или ДНК-микрочипов (Mendes и др., 2011) позволяет провести углубленный анализ этих микробных сообществ. Здесь такие роды, как Bacillus, Acidobacteria (Gp22, Gp16, GP6) и Ignavibacterium, были более обильны в ризосфере перца, обработанного органической почвой. Среди них Bacillus, вероятно, был связан с подавлением фитофтороза перца. Культурально-зависимые исследования предоставили доказательства того, что большинство (50%) in vitro антагонистов из ризосферы перца, свободного от симптомов болезней, относились к Bacillus, и их численность была выше в обработке органической почвой, чем в других изученных обработках. Другим доказательством является то, что микробный консорциум из 18 антагонистов Bacillus, выделенных от растений перца, свободных от каких-либо симптомов болезней, был способен усиливать способность почвы из интегрированной системы подавлять фитофтороз перца. Штаммы Bacillus, такие как B. subtilis Bs 8B-1 (Khabbaz и др., 2015), B. licheniformis ATCC 14580 (Bibi и др., 2018) и B. cereus B1301 (Yang и др., 2012), были способны подавлять фитофтороз как в тепличных, так и в полевых условиях. Ранее некоторые практики управления сельским хозяйством, связанные с органическим земледелием, такие как внесение органических материалов, были способны усиливать супрессивность почвы против нескольких болезней растений (van Bruggen и др., 2016). Это явление часто называют общей супрессивностью, при которой считается, что активность аборигенных почвенных микроорганизмов запускается неспецифически (van Bruggen и Finckh, 2016). В совокупности наши результаты показывают, что специфическое обогащение Bacillus в ризосфере перца способствовало подавлению фитофтороза перца в почве, подвергнутой долгосрочному органическому земледелию.
Предполагалось, что представители Acidobacteria являются K-стратегами, которые могут медленно реагировать на изменения окружающей среды (Fierer и др., 2007). Ignavibacterium был предложен как участвующий в деградации органического вещества (Bleyen и др., 2018), которое было значительно выше в почве при органическом земледелии. Таким образом, возможно, что высокая численность Acidobacteria (Gp22, Gp16, GP6) и Ignavibacterium может быть связана с долгосрочным использованием компоста, который был extensively разложен микроорганизмами. Таким образом, роли этих родов в подавлении фитофтороза перца требуют дальнейшего изучения. In vitro антагонисты, относящиеся к Alpha- и Gamma-Proteobacteria, были связаны со специфической супрессивностью в почвах (Mendes и др., 2011; Cha и др., 2016). Здесь их численность была ниже в обработке органической почвой, чем в почве из интегрированной или традиционной системы. Этот результат показывает, что эти бактерии в меньшей степени способствовали подавлению фитофтороза в данном эксперименте. Однако дальнейшие исследования, такие как метатранскриптомные или метапротеомные исследования, могут пролить больше света на функции различных микробных таксонов в ризосфере. Общая супрессивность отличается от специфической супрессивности в нескольких аспектах, таких как спектр и сила подавления болезней, проводимость или развитие подавления болезней (van Bruggen и Finckh, 2016). Таким образом, возможно, что общая и специфическая супрессивность связаны с различными микроорганизмами. Однако это все еще исследование отдельного случая, и полученные знания необходимо валидировать в других долгосрочных экспериментах для получения всестороннего понимания роли почвенного микробиома в здоровье растений, поскольку он в основном формируется каждой конкретной экологической нишей (Bahram и др., 2018).
Заключение
Почвенный микробиом в органической системе земледелия был более эффективен в подавлении фитофтороза перца, чем в интегрированной или традиционной системе, и приводил к иному микробному сообществу в ризосфере сеянцев перца. Анализы in vitro антагонистов, ризосферных микробных сообществ и микробных консорциумов подчеркивают роль Bacillus в подавлении фитофтороза.
Дополнительный материал
Дополнительный материал к этой статье можно найти онлайн по адресу: https://www.frontiersin.org/articles/10.3389/fmicb.2019.00342/full#supplementary-material
Сноски
1. ^ http://www.r-project.org/
2. ^ www.freebioinfo.org
3. ^ https://galaxyproject.org/
Список литературы
1. Adesina M. F., Lembke A., Costa R., Speksnijder A., Smalla K. (2007). Screening of bacterial isolates from various European soils for in vitro antagonistic activity towards Rhizoctonia solani and Fusarium oxysporum: site-dependent composition and diversity revealed. Soil Biol. Biochem. 39, 2818–2828. doi: 10.1016/j.soilbio.2007.06.004. CrossRef. Google Scholar.
2. Babin D., Ding G., Pronk G. J., Heister K., Kögel-Knabner I., Smalla K. (2012). Metal oxides, clay minerals and charcoal determine the composition of microbial communities in matured artificial soils and their response to phenanthrene. FEMS Microbiol. Ecol. 86, 3–14. doi: 10.1111/1574-6941.12058. CrossRef. Google Scholar.
3. Bahram M., Hildebrand F., Forslund S. K., Anderson J. L., Soudzilovskaia N. A., Bodegom P. M., et al. (2018). Structure and function of the global topsoil microbiome. Nature 560, 233–243. doi: 10.1038/s41586-018-0386-6. CrossRef. Google Scholar.
4. Ball B. C., Bingham I., Rees R. M., Watson C. A., Litterick A. (2005). The role of crop rotations in determining soil structure and crop growth conditions. Can. J. Soil Sci. 85, 557–577. doi: 10.4141/S04-078. CrossRef. Google Scholar.
5. Barberán A., Bates S. T., Casamayor E. O., Fierer N. (2012). Using network analysis to explore co-occurrence patterns in soil microbial communities. ISME J. 6, 343–351. doi: 10.1038/ismej.2011.119. CrossRef. Google Scholar.
6. Barchenger D. W., Lamour K. H., Sheu Z. M., Shrestha S., Kumar S., Lin S., et al. (2017). Intra- and intergenomic variation of ploidy and clonality characterize Phytophthora capsici on Capsicum sp. in Taiwan. Mycol. Prog. 16, 955–963. doi: 10.1007/s11557-017-1330-0. CrossRef. Google Scholar.
7. Bastian M., Heymann S., Jacomy M. (2009). "Gephi: An open source software for exploring and manipulating networks," in Proceedings of the International AAAI conference on weblogs and social media, San Jose, CA. Google Scholar.
8. Bi Y., Hu J., Cui X., Shao J., Lu X., Meng Q., et al. (2014). Sexual reproduction increases the possibility that Phytophthora capsici will develop resistance to dimethomorph in China. Plant Pathol. 63, 1365–1373. doi: 10.1111/ppa.12220. CrossRef. Google Scholar.
9. Bibi F., Strobel G. A., Naseer M. I., Yasir M., Khalaf Al-Ghamdi A. A., Azhar E. I. (2018). Microbial flora associated with the halophyte--salsola imbricate and its biotechnical potential. Front. Microbiol. 9:65. doi: 10.3389/fmicb.2018.00065. CrossRef. Google Scholar.
10. Bleyen N., Hendrix K., Moors H., Durce D., Vasile M., Valcke E. (2018). Biodegradability of dissolved organic matter in boom clay pore water under nitrate-reducing conditions: effect of additional C and P sources. Appl. Geochem. 92, 45–58. doi: 10.1016/j.apgeochem.2018.02.005. CrossRef. Google Scholar.
11. Boudreau M. A. (2013). Diseases in intercropping systems. Ann. Rev. Phytopathol. 51, 499–519. doi: 10.1146/annurev-phyto-082712-102246. CrossRef. Google Scholar.
12. Caporaso J. G., Kuczynski J., Stombaugh J., Bittinger K., Bushman F. D., Costello E. K., et al. (2010). QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 7, 335–336. doi: 10.1038/nmeth.f.303. CrossRef. Google Scholar.
13. Cha J. Y., Han S., Hong H. J., Cho H., Kim D., Kwon Y., et al. (2016). Microbial and biochemical basis of a Fusarium wilt-suppressive soil. ISME J. 10, 119–129. doi: 10.1038/ismej.2015.95. CrossRef. Google Scholar.
14. Chae D. H., De Jin R., Hwangbo H., Kim Y. W., Kim Y. C., Park R. D., et al. (2006). Control of late blight (Phytophthora capsici) in pepper plant with a compost containing multitude of chitinase-producing bacteria. Biocontrol 51, 339–351. doi: 10.1007/s10526-005-2934-x. CrossRef. Google Scholar.
15. Champeil A., Fourbet J. F., Doré T., Rossignol L. (2004). Influence of cropping system on Fusarium head blight and mycotoxin levels in winter wheat. Crop Prot. 23, 531–537. doi: 10.1016/j.cropro.2003.10.011. CrossRef. Google Scholar.
16. Cole J. R., Wang Q., Fish J. A., Chai B., McGarrell B. M., Sun Y., et al. (2014). Ribosomal database project: data and tools for high throughput rRNA analysis. Nucleic Acids Res. 42, 633–642. doi: 10.1093/nar/gkt1244. CrossRef. Google Scholar.
17. Ding G., Heuer H., Smalla K. (2012). Dynamics of bacterial communities in two unpolluted soils after spiking with phenanthrene: soil type specific and common responders. Front. Microbiol. 3:290. doi: 10.3389/fmicb.2012.00290. CrossRef. Google Scholar.
18. Ding G., Heuer H., Zuhlke S., Spiteller M., Pronk G. J., Heister K., et al. (2010). Soil type-dependent responses to phenanthrene as revealed by determining the diversity and abundance of polycyclic aromatic hydrocarbon ring-hydroxylating dioxygenase genes by using a novel PCR detection system. Appl. Environ. Microbiol. 76, 4765–4771. doi: 10.1128/AEM.00047-10. CrossRef. Google Scholar.
19. Ding G., Pronk G. J., Babin D., Heuer H., Heister K., Kögel-Knabner I., et al. (2013). Mineral composition and charcoal determine the bacterial community structure in artificial soils. FEMS Microbiol. Ecol. 86, 15–25. doi: 10.1111/1574-6941.12070. CrossRef. Google Scholar.
20. Fan B., Wang C., Song X., Ding X., Wu L., Wu H., et al. (2018). Bacillus velezensis FZB42 in 2018: the gram-positive model strain for plant growth promotion and biocontrol. Front. Microbiol. 9:2491. doi: 10.3389/fmicb.2018.02491. CrossRef. Google Scholar.
21. Fernandez A. L., Sheaffer C. C., Wyse D. L., Staley C., Gould T. J., Sadowsky M. J. (2016). Associations between soil bacterial community structure and nutrient cycling functions in long-term organic farm soils following cover crop and organic fertilizer amendment. Sci. Total Environ. 566-567, 949–959. doi: 10.1016/j.scitotenv.2016.05.073. CrossRef. Google Scholar.
22. Fierer N., Bradford M. A., Jackson R. B. (2007). Toward an ecological classification of soil bacteria. Ecology 88, 1354–1364. doi: 10.1890/05-1839. CrossRef. Google Scholar.
23. Gómez Expósito R., de Bruijn I., Postma J., Raaijmakers J. M. (2017). Current insights into the role of rhizosphere bacteria in disease suppressive soils. Front. Microbiol. 8:2529. doi: 10.3389/fmicb.2017.02529. CrossRef. Google Scholar.
24. Gouda S., Kerry R. G., Das G., Paramithiotis S., Shin H. S., Patra J. K. (2018). Revitalization of plant growth promoting rhizobacteria for sustainable development in agriculture. Microbiol. Res. 206, 131–140. doi: 10.1016/j.micres.2017.08.016. CrossRef. Google Scholar.
25. Hadar Y., Papadopoulou K. K. (2012). Suppressive composts: microbial ecology links between abiotic environments and healthy plants. Ann. Rev. Phytopathol. 50, 133–153. doi: 10.1146/annurev-phyto-081211-172914. CrossRef. Google Scholar.
26. Han H., Teng Y., Yang H., Li J. (2017). Effects of long-term use of compost on N₂O and CO₂ fluxes in greenhouse vegetable systems. Compost Sci. Uti. 25(Suppl. 1), S61–S69. doi: 10.1080/1065657X.2016.1238786. CrossRef. Google Scholar.
27. Hartmann M., Frey B., Mayer J., Mader P., Widmer F. (2014). Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 9, 1177–1194. doi: 10.1038/ismej.2014.210. CrossRef. Google Scholar.
28. Hothorn T., Bretz F., Westfall P. (2017). Simultaneous inference in general parametric models. Biomed. J. 50, 346–363. Google Scholar.
29. Khabbaz S. E., Zhang L., Cáceres L. A., Sumarah M., Wang A., Abbasi P. A. (2015). Characterisation of antagonistic Bacillus and Pseudomonas strains for biocontrol potential and suppression of damping-off and root rot diseases. Ann. Appl. Biol. 166, 456–471. doi: 10.1111/aab.12196. CrossRef. Google Scholar.
30. Kropf S., Läuter J., Eszlinger M., Krohn K., Paschke R. (2004). Nonparametric multiple test procedures with data-driven order of hypotheses and with weighted hypotheses. J. Stat. Plan. Inference 125, 31–47. doi: 10.1016/j.jspi.2003.07.021. CrossRef. Google Scholar.
31. Lane D. J. (1991). "16S/23S rRNA Sequencing," in Nucleic Acid Techniques in Bacterial Systematics, eds Stackebrandt E., Goodfellow M. (New York, NY: Wiley). Google Scholar.
32. Lenc L., Kwaśna H., Sadowski C., Grabowski A. (2015). Microbiota in wheat roots, rhizosphere and soil in crops grown in organic and other production systems. J. Phytopathol. 163, 245–263. doi: 10.1111/jph.12313. CrossRef. Google Scholar.
33. Li F., Chen L., Zhang J., Yin J., Huang S. (2017). Bacterial community structure after long-term organic and inorganic fertilization reveals important associations between soil nutrients and specific taxa involved in nutrient transformations. Front. Microbiol. 8:187. doi: 10.3389/fmicb.2017.00187. CrossRef. Google Scholar.
34. Lithourgidis A. S., Dordas C. A., Damalas C. A., Vlachostergios D. N. (2011). Annual intercrops: an alternative pathway for sustainable agriculture. Aust. J. Crop Sci. 5, 396–410. Google Scholar.
35. Liu H., Xiong W., Zhang R., Hang X., Wang D., Li R., et al. (2018). Continuous application of different organic additives can suppress tomato disease by inducing the healthy rhizospheric microbiota through alterations to the bulk soil microflora. Plant Soil. 423, 229–240. doi: 10.1007/s11104-017-3504-6. CrossRef. Google Scholar.
36. Liu L., Sun C., He X., Liu X., Wu H., Liu M., et al. (2016). The secondary compost products enhances soil suppressive capacity against bacterial wilt of tomato caused by Ralstonia solanacearum. Eur. J. Soil Biol. 75, 70–78. doi: 10.1016/j.ejsobi.2016.04.005. CrossRef. Google Scholar.
37. Louws F. J., Fulbright D. W., Stephens C. T., De Bruijn F. J. (1994). Specific genomic fingerprints of phytopathogenic Xanthomonas and Pseudomonas pathovars and strains generated with repetitive sequences and PCR. Appl. Environ. Microbiol. 60, 2286–2295. Google Scholar.
38. Luo P., Han X., Wang Y., Han M., Shi H., Liu N., et al. (2015). Influence of long-term fertilization on soil microbial biomass, dehydrogenase activity, and bacterial and fungal community structure in a brown soil of northeast China. Ann. Microbiol. 65, 533–542. doi: 10.1007/s13213-014-0889-9. CrossRef. Google Scholar.
39. Marschner P., Kandeler E., Marschner B. (2003). Structure and function of the soil microbial community in a long-term fertilizer experiment. Soil Biol. Biochem. 35, 453–461. doi: 10.1007/s00248-008-9372-0. CrossRef. Google Scholar.
40. Mendes R., Kruijt M., de Bruijn I., Dekkers E., van der Voort M., Schneider J. H. M., et al. (2011). Deciphering the rhizosphere microbiome for disease-suppressive bacteria. Science 332, 1097–1100. doi: 10.1126/science.1203980. CrossRef. Google Scholar.
41. Michelsen C. F., Watrous J., Glaring M. A., Kersten R., Koyama N., Dorrestein P. C., et al. (2015). Nonribosomal peptides, key biocontrol components for Pseudomonas fluorescens In5, isolated from a Greenlandic suppressive soil. mBio 6:e00079. doi: 10.1128/mBio.00079-15. CrossRef. Google Scholar.
42. Ndubuisi-Nnaji U. U., Adegoke A. A., Ogbu H. I., Ezenobi N. O., Okoh A. I. (2011). Effect of long-term organic fertilizer application on soil microbial dynamics. Afr. J. Biotechnol. 10, 556–559. doi: 10.5897/ajb10.699. CrossRef. Google Scholar.
43. Noble R., Coventry E. (2010). Suppression of soil-borne plant diseases with composts: a review. Biocontrol Sci. Technol. 15, 3–20. doi: 10.1080/09583150400015904. CrossRef. Google Scholar.
44. Paidhungat M., Setlow B., Driks A., Setlow P. (2000). Characterization of spores of Bacillus subtilis which lack dipicolinic acid. J. Bacteriol. 182, 5505–5512. doi: 10.1128/JB.182.19.5505-5512.2000. CrossRef. Google Scholar.
45. Postma J., Schilder M. T., Bloem J., van Leeuwen-Haagsma W. K. (2008). Soilsuppressiveness and functional diversity of the soil microflora in organic farming systems. Soil Biol. Biochem. 40, 2394–2406. doi: 10.1016/j.soilbio.2008.05.023. CrossRef. Google Scholar.
46. Pronk G. J., Heister J., Vogel C., Babin D., Bachmann J., Ding G., et al. (2017). Interaction of minerals, organic matter, and microorganisms during biogeochemical interface formation as shown by a series of artificial soil experiments. Biol. Fertil. Soils 53, 9–22. doi: 10.1007/s00374-016-1161-1. CrossRef. Google Scholar.
47. Qiu M., Zhang R., Xue C., Zhang S., Li S., Zhang N., et al. (2012). Application of bio-organic fertilizer can control Fusarium wilt of cucumber plants by regulating microbial community of rhizosphere soil. Biol. Fertil. Soils 48, 807–816. doi: 10.1007/s00374-012-0675-4. CrossRef. Google Scholar.
48. Rognes T., Flouri T., Nichols B., Quince C., Mahe F. (2016). VSEARCH: a versatile open source tool for metagenomics. PeerJ 4:e2584. doi: 10.7717/peerj.2584. CrossRef. Google Scholar.
49. Sang M. K., Kim J. G., Kim K. D. (2010). Biocontrol activity and induction of systemic resistance in pepper by compost water extracts against Phytophthora capsici. Phytopathology 100, 774–783. doi: 10.1094/PHYTO-100-8-0774. CrossRef. Google Scholar.
50. Schlemper T. R., Leite M. F. A., Lucheta A. R., Shimels M., Bouwmeester H. J., van Veen J. A., et al. (2017). Rhizobacterial community structure differences among sorghum cultivars in different growth stages and soils. FEMS Microbiol. Ecol. 93, 1–11. doi: 10.1093/femsec/fix096. CrossRef. Google Scholar.
51. Schloss P. D., Westcott S. L., Ryabin T., Hall J. R., Hartmann M., Hollister E. B., et al. (2009). Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 75, 7537–7541. doi: 10.1128/AEM.01541-09. CrossRef. Google Scholar.
52. Schreiter S., Ding G., Heuer H., Neumann G., Sandmann M., Grosch R., et al. (2014). Effect of the soil type on the microbiome in the rhizosphere of field-grown lettuce. Front. Microbiol. 5:144. doi: 10.3389/fmicb.2014.00144. CrossRef. Google Scholar.
53. Sharma K., Bruns C., Butz A. F., Finckh M. R. (2012). Effects of fertilizers and plant strengtheners on the susceptibility of tomatoes to single and mixed isolates of Phytophthora infestans. Eur. J. Plant Pathol. 133, 739–751. doi: 10.1007/s10658-012-9954-z. CrossRef. Google Scholar.
54. Shen Z., Ruan Y., Chao X., Zhang J., Li R., Shen Q. (2015). Rhizosphere microbial community manipulated by 2 years of consecutive biofertilizer application associated with banana Fusarium wilt disease suppression. Biol. Fertil. Soils 51, 553–562. doi: 10.1007/s00374-015-1002-7. CrossRef. Google Scholar.
55. Strunnikova O. K., Vishnevskaya N. A., Ruchiy A. S., Shakhnazarova V. Y., Vorobyov N. A., Chebotar V. K. (2015). The influence of soils with different textures on development, colonization capacity and interactions between Fusarium culmorum and Pseudomonas fluorescens in soil and on barley roots. Plant Soil. 389, 131–144. doi: 10.1007/s11104-014-2351-y. CrossRef. Google Scholar.
56. Tamura K., Stecher G., Peterson D., Filipski A., Kumar S. (2013). MEGA6: molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 30, 2725–2729. doi: 10.1093/molbev/mst197. CrossRef. Google Scholar.
57. Thuerig B., Fließbach A., Berger N., Fuchs J. G., Kraus N., Mahlberg N., et al. (2009). Re-establishment of suppressiveness to soil- and air-borne diseases by re-inoculation of soil microbial communities. Soil Biol. Biochem. 41, 2153–2161. doi: 10.1016/j.soilbio.2009.07.028. CrossRef. Google Scholar.
58. van Bruggen A. H. C., Finckh M. R. (2016). Plant diseases and management approaches in organic farming systems. Ann. Rev. Phytopathol. 54, 25–54. doi: 10.1146/annurev-phyto-080615-100123. CrossRef. Google Scholar.
59. van Bruggen A. H. C., Gamliel A., Finckhc M. R. (2016). Plant disease management in organic farming systems. Pest Manag. Sci. 72, 30–44. doi: 10.1002/ps.4145. CrossRef. Google Scholar.
60. van der Voort M., Kempenaar M., van Driel M., Raaijmakers J. M., Mendes R. (2016). Impact of soil heat on reassembly of bacterial communities in the rhizosphere microbiome and plant disease suppression. Ecol. Lett. 19, 375–382. doi: 10.1111/ele.12567. CrossRef. Google Scholar.
61. Wang L., Lu X., Yuan H., Wang B., Shen Q. (2015). Application of bio-organic fertilizer to control tomato Fusarium wilting by manipulating soil microbial communities and development. Commun. Soil Sci. Plant Anal. 46, 2311–2322. doi: 10.1080/00103624.2015.1081694. CrossRef. Google Scholar.
62. Wang R., Xu H., Mridha M. A. U. (2001). Phytophthora resistance of organically-fertilized tomato plants. J. Crop Prod. 3, 77–84. doi: 10.1300/J144v03n01_07. CrossRef. Google Scholar.
63. Weinert N., Piceno Y., Ding G., Meincke R., Heuer H., Berg G., et al. (2011). Phylo-Chip hybridization uncovered an enormous bacterial diversity in the rhizosphere of different potato cultivars: many common and few cultivar-dependent taxa. FEMS Microbiol. Ecol. 75, 497–506. doi: 10.1111/j.1574-6941.2010.01025.x. CrossRef. Google Scholar.
64. Weller D. M., Raaijmakers J. M., Gardener B. B., Thomashow L. S. (2002). Microbial populations responsible for specific soil suppressiveness to plant pathogens. Ann. Rev. Phytopathol. 40, 309–348. doi: 10.1146/annurev.phyto.40.030402.110010. CrossRef. Google Scholar.
65. Xiong W., Li R., Ren Y., Liu C., Zhao Q., Wu H., et al. (2017). Distinct roles for soil fungal and bacterial communities associated with the suppression of vanilla Fusarium wilt disease. Soil Biol. Biochem. 107, 198–207. doi: 10.1016/j.soilbio.2017.01.010. CrossRef. Google Scholar.
66. Xiong W., Zhao Q., Zhao J., Xun W., Li R., Zhang R., et al. (2015). Different continuous cropping spans significantly affect microbial community membership and structure in a vanilla-grown soil as revealed by deep pyrosequencing. Microb. Ecol. 70, 209–218. doi: 10.1007/s00248-014-0516-0. CrossRef. Google Scholar.
67. Yang H., Fan J., Ge Z., Shen Z., Lv R., Li J. (2009a). Main diseases and control effects of organic, integrated and conventional cultivation patterns of greenhouse tomato. Chin. J. Eco Agric. 17, 933–937. doi: 10.3724/SP.J.1011.2009.00933. CrossRef. Google Scholar.
68. Yang H., Fan J., Liang L., Meng Y., Zhang S., Li J. (2009b). Studies on the main diseases and control effects under organic, integrated and conventional cultivation patterns of cucumber in greenhouse. Acia Agric. Boreali-sjnica 24, 240–245. Google Scholar.
69. Yang M., Xu L., Xue Q., Yang J., Xu Q., Liu H., et al. (2012). Screening potential bacterial biocontrol agents towards Phytophthora capsici in pepper. Eur. J. Plant Pathol. 134, 811–820. doi: 10.1007/s10658-012-0057-7. CrossRef. Google Scholar.
70. Yilmaz P., Yarza P., Gerken J., Pruesse E., Quast C., et al. (2014). The SILVA and "All-species Living Tree Project (LTP)" taxonomic frameworks. Nucleic Acids Res. 42, 643–648. doi: 10.1093/nar/gkt1209. CrossRef. Google Scholar.
71. Yim B., Winkelmann T., Ding G., Smalla K. (2015). Different bacterial communities in heat and gamma irradiation treated replant disease soils revealed by 16S rRNA gene analysis--contribution to improved aboveground apple plant growth? Front. Microbiol. 6:1224. doi: 10.3389/fmicb.2015.01224. CrossRef. Google Scholar.
72. Yin C., Hulbert S. H., Schroeder K. L., Mavrodi O., Mavrodi D., Dhingra A., et al. (2013). Role of bacterial communities in the natural suppression of Rhizoctonia solani bare patch disease of wheat (Triticum aestivum L.). Appl. Environ. Microbiol. 79, 7428–7438. doi: 10.1128/AEM.01610-13. CrossRef. Google Scholar.
73. Zhang N., Pan R., Shen Y., Yuan J., Wang L., Luo X., et al. (2017). Development of a novel bio-organic fertilizer for plant growth promotion and suppression of rhizome rot in ginger. Biol. Control 114, 97–105. doi: 10.1016/j.biocontrol.2017.08.001. CrossRef. Google Scholar.

Li H, Cai X, Gong J, Xu T, Ding G and Li J (2019) Long-Term Organic Farming Manipulated Rhizospheric Microbiome and Bacillus Antagonism Against Pepper Blight (Phytophthora capsici). Front. Microbiol. 10:342. doi: 10.3389/fmicb.2019.00342
Перевод статьи «Long-Term Organic Farming Manipulated Rhizospheric Microbiome andBacillusAntagonism Against Pepper Blight (Phytophthora capsici)» авторов Li H, Cai X, Gong J, Xu T, Ding G and Li J., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык
Фото: wikipedia

















































































































Комментарии (0)