Защита от болезней и ускорение роста: влияние пробиотиков на здоровье тихоокеанской белой креветки
Тихоокеанская белая креветка (Penaeus vannamei) является одним из наиболее продуктивных и экономически важных видов в мире. Однако развитие и непрерывное расширение масштабов разведения привели к увеличению риска возникновения заболеваний в креветочном хозяйстве. Применение пробиотиков как эффективного метода борьбы с болезнями в аквакультуре широко рассматривается. В креветководстве уже используется несколько пробиотиков, и было показано, что они приносят пользу здоровью хозяина.
Аннотация
Для расширения спектра бактериальных видов, используемых в качестве пробиотиков в креветководстве, в данном исследовании мы поставили цель выяснить влияние диетических пробиотиков (Clostridium butyricum I9 (I9), Clostridium butyricum G15 (G15) или Paraclostridium bifermentans X13) на рост, иммунный ответ и кишечный микробиом белой креветки. Креветок кормили рационами, содержащими либо фосфатно-солевой буфер (PBS), I9 (10⁷ КОЕ/г корма), G15 (10⁷ КОЕ/г корма) или X13 (10⁷ КОЕ/г корма) в течение 30 дней, после чего проводили заражение Vibrio parahaemolyticus (Vp). Результаты показали, что выживаемость, прирост массы тела и удельная скорость роста креветок в группах I9, X13 и G15 значительно увеличились по сравнению с группой PBS. Добавление пробиотиков повышало содержание короткоцепочечных жирных кислот и эффективно поддерживало нормальную морфологию и структуру кишечника и гепатопанкреаса. Группы I9, X13 и G15 показали положительные изменения в разнообразии и обилии кишечных бактерий. У креветок в группах I9, X13 и G15 наблюдалась значительная регуляция вверх экспрессии генов CTL, SOD, proPO, Crustin, PEN2-4 и ALF1-3. Кроме того, диетические пробиотики значительно повышали выживаемость, поддерживали структуру кишечника, стимулировали активность ферментов SOD, AKP, ACP и T-AOC, а также снижали уровень MDA у креветок после заражения Vp. В заключение, диетическая добавка I9, G15 или X13 улучшала рост, иммунитет и устойчивость к болезням тихоокеанской белой креветки, что обеспечивает научную основу для креветководства.
1 Введение
Ракообразные являются вторым по величине подтипом в мире. Тихоокеанская белая креветка (Penaeus vannamei) принадлежит к типу членистоногих, классу ракообразных, отряду десятиногих и является одним из трех наиболее продуктивных и экономически важных видов креветок в мире (Cahú et al., 2012; Cornejo-Granados et al., 2017; Liu et al., 2017; Jin et al., 2018; Tran et al., 2023). С быстрым развитием аквакультуры высокоплотное культивирование приводит к риску возникновения заболеваний в интенсивной аквакультуре, что стало одним из важнейших факторов, ограничивающих повышение продуктивности аквакультуры (Kumar et al., 2014). В креветководстве заболевания, вызываемые бактериями (такими как Vibrio parahaemolyticus-Vp) и вирусами (вирус синдрома белых пятен-WSSV), оказывают негативное влияние на развитие креветководства (Aziz et al., 2012; Chang et al., 2012; Nimrat et al., 2012). Антибиотики часто применяются для минимизации неблагоприятных последствий заболеваний в креветководстве. Однако длительное использование может приводить к возникновению устойчивости к антибиотикам и вызывать загрязнение воды в системах аквакультуры (Mo et al., 2015; Okocha et al., 2018). Поэтому поиск экологически чистых методов выращивания важен для борьбы с болезнями и защиты окружающей среды в индустрии креветководства.
Предыдущие исследования показали, что активные ингредиенты из растений, химические вещества и пробиотики в качестве добавок в корма для культивируемых водных животных могут улучшать состояние здоровья и предотвращать заболевания (Pérez-Sánchez et al., 2018). В последние годы пробиотики стали стратегией выращивания без антибиотиков для предотвращения и снижения риска заболеваний в аквакультуре. Добавление Bacillus tequilensis AP BFT3 в рацион тихоокеанской белой креветки (P. vannamei) значительно повышало экспрессию генов Litv1, тропомиозина и миозина, а также улучшало показатели роста (Panigrahi et al., 2022). Диетическая добавка Rhodobacter sphaeroides значительно способствовала улучшению ростовых показателей и относительного обилия полезной микробиоты (включая Paracoccus и Sulfitobacter) в кишечнике тихоокеанской белой креветки (Song et al., 2024). Другое исследование показало, что пробиотики Lactobacillus reuteri и Pediococcus acidilactici могут увеличивать прирост массы тела, удельную скорость роста и выживаемость, а также снижать коэффициент конверсии корма у тихоокеанской белой креветки после 8 недель кормления (Wu et al., 2022). Результаты этих исследований показали, что пробиотики в качестве кормовых добавок оказывают благотворное влияние на улучшение состояния здоровья и профилактику заболеваний в креветководстве.
У водных животных производство короткоцепочечных жирных кислот (особенно масляной кислоты) бактериями, продуцирующими бутират, в результате ферментации углеводов способствует регенерации и восстановлению кишечного эпителия, а также регуляции здоровой кишечной микроэкологической среды (Duan et al., 2017; Tran et al., 2020a). Добавление бутират-продуцирующих бактерий в корма может регулировать кишечную микробиоту, ингибировать рост патогенных бактерий и улучшать иммунитет хозяина и устойчивость хозяина к патогенным бактериям (Tran et al., 2020b). Среди бутират-продуцирующих бактерий Clostridium butyricum является наиболее распространенной бактерией, используемой в аквакультуре. В предыдущем исследовании сообщалось, что тихоокеанские белые креветки, получавшие корм с добавлением C. butyricum в течение 42 дней, показали увеличение массы тела, скорости роста, выживаемости и устойчивости к инфекции Vp (Li et al., 2019). Также было обнаружено, что применение C. butyricum CG30 может стимулировать щелочную фосфатазу (AKP), снижать экспрессию фенолоксидазы (proPO), α-2-макроглобулина (A2M) и антилипополисахаридного фактора (ALF) и повышать выживаемость тихоокеанской белой креветки после заражения V. alginolyticus (Wang et al., 2023). Таким образом, C. butyricum является превосходным классом пробиотиков, которые, как было показано, играют важную роль у водных животных, например, в регуляции кишечного микроэкологического баланса и улучшении здоровья кишечника (Liang et al., 2023).
Чтобы диверсифицировать потенциальные источники, используемые в креветководстве, в этом исследовании мы оценили влияние трех бактерий: C. butyricum I9, C. butyricum G15 и Paraclostridium bifermentans X13 на показатели роста, микробиоту кишечника, антиоксидантную способность, иммунный ответ и устойчивость к болезням у тихоокеанской белой креветки. Эти бактериальные штаммы I9, G15 и X13 были ранее выделены из кишечника белого морского окуня (Atractoscion nobilis) и показали положительный вклад в рост и здоровье белого морского окуня (неопубликованные данные). Результаты этого исследования предоставляют важную научную основу для потенциального применения этих бактерий в качестве пробиотиков в креветководстве.
2 Материалы и методы
2.1 Штамм бактерий, источник креветок, условия культивирования
Пробиотические бактерии I9, G15 и X13, использованные в этом исследовании, были ранее выделены из желудочно-кишечного тракта белого морского окуня и хранятся в лаборатории болезней и иммунитета крабов-плавунцов Шаньтоуского университета (Шаньтоу, Китай). Бактерии культивировали отдельно в среде PY при 28°C при постоянном встряхивании при 120 об/мин в течение 24 ч (Liang et al., 2023). После 24 ч инкубации бактериальный раствор центрифугировали при 6000 об/мин в течение 10 мин для сбора бактериальных клеток, которые трижды промывали стерильным PBS-буфером и ресуспендировали в стерильном PBS-буфере. Конечную концентрацию биомассы доводили до 1 × 10⁷ КОЕ/мл.
Шестьсот здоровых ювенильных креветок (начальная масса тела: 8,1 ± 0,2 г; начальная длина тела: 11,5 ± 0,4 см) культивировали на экспериментальной базе Хайкан лаборатории Fujian Dabeinong Huayou Aquatic Technology Group Co. Ltd. (Фуцзянь, Китай) в системе с фильтрованной аэрируемой морской водой (температура воды: 28 ± 2°C, pH: 7,2 ± 0,4, растворенный кислород: >6 мг/л, аммиачный азот: <0,2 мг/л, нитритный азот: <0,05 мг/л) в течение 5 дней. Плотность посадки составляла 50 креветок/аквариум (300 л воды). В период акклиматизации креветок кормили коммерческим гранулированным кормом (≥43% сырого протеина, ≤15% сырой золы, ≥5,0% сырого жира, ≤5,0% сырой клетчатки, ≥2,6% лизина, ≤11% влаги и 0,9–2,0% общего фосфора), приобретенным у Fujian Dabeinong Huayou Aquatic Technology Group Co., Ltd. (Фуцзянь, Китай). Культуральную воду заменяли дважды в день (каждая замена составляла около 20%). Остатки корма и фекалии удаляли с помощью сифона одновременно с заменой воды.
2.2 Эксперимент по кормлению
Креветок случайным образом разделили на четыре группы: PBS (креветки получали базовый рацион с добавлением стерильного PBS), I9 (креветки получали базовый рацион с добавлением I9), G15 (креветки получали базовый рацион с добавлением G15) и X13 (креветки получали базовый рацион с добавлением X13). Базовый корм непосредственно опрыскивали PBS или бактериальным раствором (1 × 10⁷ КОЕ/г корма), помещали в сушильный шкаф при 40°C на 30 мин и хранили в холодильнике при 4°C. Креветок кормили дважды в день (в 6:00 и 15:00) в течение 30 дней. В период культивирования воду заменяли дважды в день, удаляя остатки корма и фекалии. Регистрировали смертность креветок.
2.3 Параметры роста
Выживаемость, прирост массы, прирост длины и удельную скорость роста креветок оценивали в конце эксперимента по культивированию (30 дней), и каждый параметр роста рассчитывали следующим образом:

Выживаемость (SR,%) = (Конечное количество креветок)/(Начальное количество креветок) × 100
Удельная скорость роста (SGR,%/день) = [(ln Конечная масса тела – ln Начальная масса тела)/t] × 100%
Темп прироста массы (WGR,%) = (Конечная масса тела – Начальная масса тела)/(Начальная масса тела) × 100
Прирост длины (%) = [(Конечная длина тела – Начальная длина тела)/Начальная длина тела] × 100%
2.4 Сбор и обработка образцов
В конце эксперимента по культивированию были собраны ткани, включая гемолимфу, кишечник, гепатопанкреас и содержимое кишечника креветок, немедленно помещены в жидкий азот для быстрой заморозки и затем хранились при –80°C для дальнейшего использования. Ткани кишечника и гепатопанкреаса собирали и фиксировали в 4% параформальдегиде для гистопатологического наблюдения.
Гепатопанкреас собирали и гомогенизировали в стерильном физиологическом растворе (в соотношении 1:9, м/о). Супернатант (10%) получали центрифугированием при 2500 об/мин в течение 10 мин и использовали для анализа активности ферментов. Активность ферментов, включая супероксиддисмутазу (SOD), аланинаминотрансферазу (ALT), аспартатаминотрансферазу (AST), общую антиоксидантную способность (T-AOC), каталазу (CAT), щелочную фосфатазу (AKP), кислую фосфатазу (ACP), α-амилазу (AMS) и малоновый диальдегид (MDA), определяли с использованием коммерческих наборов (Nanjing Jiancheng Institute of Bioengineering, Нанкин, Китай) в соответствии с инструкциями производителя.
Содержимое кишечника тщательно смешивали со стерильным физиологическим раствором в соотношении 1:9 (м/о), и смесь центрифугировали при 12 000 об/мин в течение 10 мин. Супернатант собирали, доводили pH до 2–3 (с использованием серной кислоты), фильтровали через мембранный фильтр с размером пор 0,22 мкм и использовали для обнаружения КЦЖК. Содержание КЦЖК определяли с использованием газового хроматографа Agilent GC6890N Network (Agilent Technologies, Санта-Клара, Калифорния, США), как ранее описано Liang et al. (2023).
2.5 Выделение РНК и ОТ-ПЦР в реальном времени
Кишечник промывали стерильным PBS-буфером для удаления содержимого, и общую РНК экстрагировали из кишечника с использованием коммерческого набора для экстракции РНК (Shanghai Feijie Biotechnology Co., Ltd. Шанхай, Китай) в соответствии с инструкциями производителя. Качество и количество общей РНК оценивали и затем использовали в качестве матрицы для синтеза кДНК с использованием набора для обратной транскрипции (Tiangen, Пекин, Китай). Концентрацию кДНК доводили до 200 нг/мкл и использовали для ОТ-ПЦР в реальном времени.
Реакции ОТ-ПЦР в реальном времени проводили в трех повторностях в смеси объемом 20 мкл, содержащей 1 мкл (каждого из прямого и обратного праймеров), 1 мкл кДНК, 7 мкл ddH₂O и 10 мкл 2 × ПЦР-смеси. Условия реакции включали три этапа: сначала 95°C в течение 30 с, затем 95°C в течение 5 с в течение 40 циклов. Наконец, 60°C в течение 20 с и анализ кривых плавления (от 65°C до 95°C). Для анализа экспрессии C-тип лектина (CTL), супероксиддисмутазы (SOD), пропофенолоксидазы (proPO) и антимикробных пептидов (АМП, включая Crustin, пенеидин-PEN2-4 и антилипополисахаридные факторы-ALF1-3) использовали Premix Ex Taq (Probe qPCR) (TaKaRa, Далянь, Китай). Ген β-актина использовали в качестве референтного гена. Относительную экспрессию этих генов нормализовали по экспрессии гена β-актина и анализировали с помощью алгоритма 2⁻ΔΔCt.
2.6 Выделение ДНК, амплификация ПЦР и секвенирование 16S рДНК
Содержимое кишечника использовали для выделения ДНК с использованием набора QIAamp Fast DNA Stool Mini Kit (Qiagen, Германия) в соответствии с инструкциями производителя. Для каждого образца ДНК выделяли в трех повторностях и затем объединяли. Качество выделенной ДНК проверяли с помощью электрофореза в 1,0% агарозном геле, а концентрацию измеряли с использованием NanoDrop One (Thermo Scientific, США). Образцы ДНК хранили при –20°C до дальнейшего анализа. Продукты ПЦР были отправлены в коммерческую компанию (Novogene Biotech Co., Ltd., Пекин, Китай) для секвенирования.
Риды каждого образца собирали с использованием FLASH (Version 1.2.11)¹ (Magoè и Salzberg, 2011) для получения исходных данных по тегам (Raw Tags). Последовательности обратных праймеров подбирали, а оставшиеся последовательности вырезали с использованием программного обеспечения Cutadapt. Затем Raw Tags фильтровали для получения высококачественных данных с использованием программного обеспечения fastp (Version 0.23.1) для получения Clean Tags (Bokulich et al., 2013). Clean Tags сравнивали с базой данных аннотаций видов и удаляли химерные последовательности для получения Effective Tags (Edgar et al., 2011). Effective Tags кластеризовали с использованием алгоритма Uparse (Uparse v7.0.1001)² (Edgar, 2013), последовательности кластеризовали с 97% идентичностью (Identity) в ОТЕ (Операционные таксономические единицы), после чего ОТЕ аннотировали для получения классификации на уровне видов. Наконец, данные каждого образца гомогенизировали и анализировали на альфа-разнообразие и бета-разнообразие на основе ОТЕ.
Для оценки богатства сообщества и размера выборки проводили анализ альфа-разнообразия (включая индексы Chao1, ACE, Shannon и Simpson) с использованием программного обеспечения Qiime (Version 1.9.1). Кривые разрежения строили с использованием программного обеспечения R (Version 4.0.3). Для оценки разнообразия сообщества проводили анализ бета-разнообразия на основе взвешенных и невзвешенных расстояний с использованием программного обеспечения Qiime. Строили тепловые карты, показывающие расстояния Unifrac между образцами. Деревья кластеризации строили для эволюционной классификации с использованием функции UPGMA.tre. Анализ главных компонент (PCA) и анализ главных координат (PCoA) выполняли с использованием ade4 и ggplot в программном обеспечении R для уменьшения размерности исходных переменных и получения и визуализации главных координат из данных. Для выявления дифференциации структуры сообщества проводили LEfSe-анализ и строили графики с использованием vegan и ggplot2 в программном обеспечении R.
2.7 Эксперимент по заражению Vibrio parahaemolyticus
В конце эксперимента по кормлению креветок в группах разделили на PBS, PBS + Vp (Vp), I9 + Vp, X13 + Vp и G15 + Vp. Объем стерильного PBS или 200 мкл Vp (1 × 10⁶ КОЕ/мл) вводили в соответствующие экспериментальные группы. Условия культивирования поддерживали такими же, как в эксперименте по культивированию, и регистрировали выживаемость креветок через 3 дня после заражения Vp. По окончании заражения у креветок в каждой группе собирали ткани (содержимое кишечника, кишечник и гепатопанкреас). Содержимое кишечника использовали для обнаружения КЦЖК. Кишечник и гепатопанкреас помещали в 4% полиметанольный тканевой фиксатор для наблюдения морфологии и структуры тканей с помощью окрашивания гематоксилином и эозином (H&E). Гепатопанкреас гомогенизировали в стерильном физиологическом растворе (в соотношении 1:9, м/о). Супернатант (10%), полученный центрифугированием при 2500 об/мин в течение 10 мин, использовали для анализа активности ферментов. Активность ферментов, включая SOD, ALT, AST, T-AOC, CAT, AKP, ACP, AMS и MDA, определяли с использованием наборов производства Nanjing Jiancheng Institute of Bioengineering (Нанкин, Китай).
2.8 Статистический анализ
Данные в результатах были представлены тремя повторностями. Все данные выражены как среднее ± стандартное отклонение. Сравнительный анализ различий между различными экспериментальными группами проводили с использованием дисперсионного анализа (ANOVA) с последующим критерием Тьюки HSD. Звездочки указывают на наличие статистически значимых различий (P < 0,05; P < 0,01).
3 Результаты
3.1 Ростовые показатели
Ростовые показатели креветок, получавших корма с добавлением I9, G15 или X13, показаны на Рисунке 1. Результаты показали, что добавление I9, G15 и X13 значительно увеличивало выживаемость креветок на 69%, 69,2% и 72% (P < 0,05) соответственно по сравнению с PBS (62,50%) (Рисунок 1A). Удельная скорость роста креветок в группах I9 (69,04%), X13 (70,01%) и G15 (64,49%) была значительно выше по сравнению с PBS (62,02%) (Рисунок 1B). Более того, темп прироста массы креветок в группах I9 (185,51%), X13 (184,31%) и G15 (156,17%) был значительно выше по сравнению с PBS (145,85%) (Рисунок 1C). Прирост длины креветок в группах I9 (36,76%), X13 (33,56%) и G15 (31,96%) был значительно выше, чем в PBS (11,01%) (Рисунок 1D).
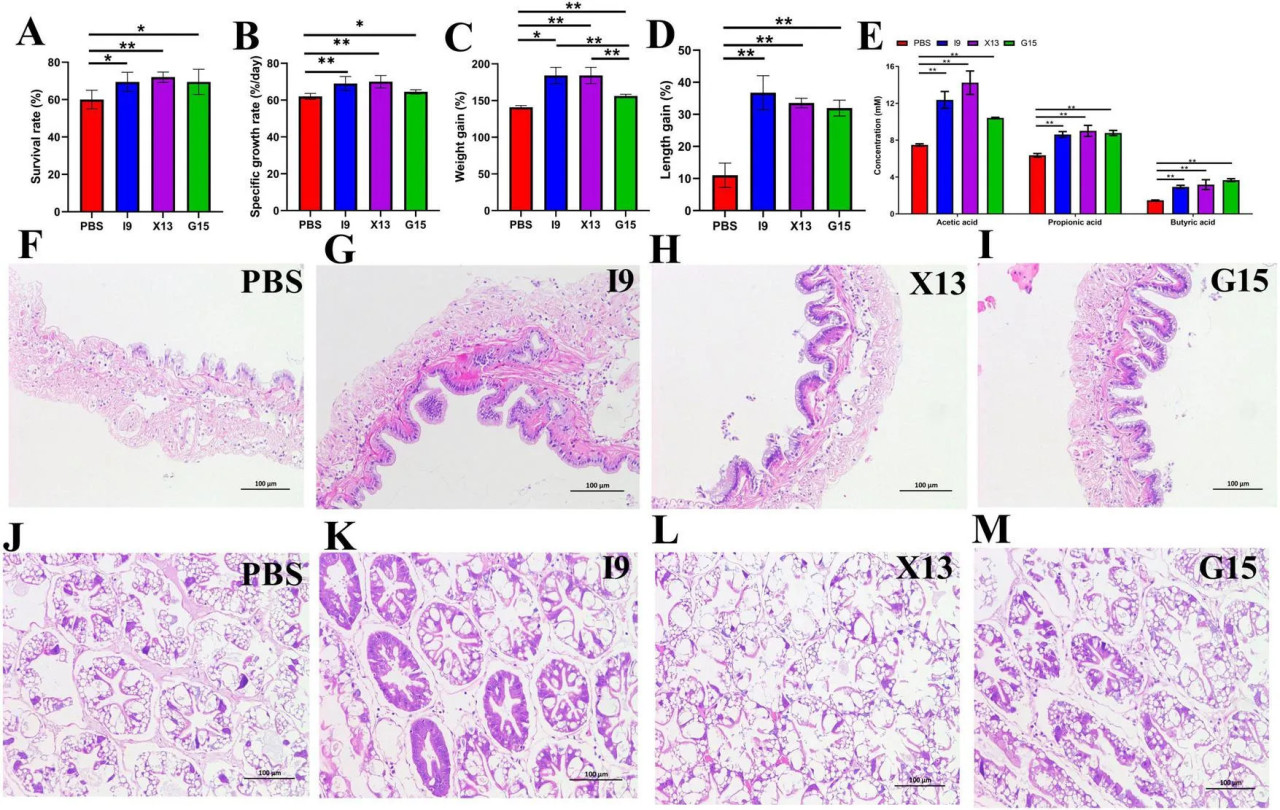
РИСУНОК 1 Ростовые показатели креветок в различных экспериментальных группах. (A–D) Изменения выживаемости, удельной скорости роста, темпа прироста массы и темпа прироста длины креветок. (E) Изменения содержания короткоцепочечных жирных кислот (КЦЖК) в кишечнике. (F–I) Структура кишечника. (J–M) Структура гепатопанкреаса. Сравнительный анализ различий между различными экспериментальными группами проводили с использованием дисперсионного анализа (ANOVA) с последующим критерием Тьюки HSD. Звездочки указывают на наличие статистически значимых различий (P < 0,05; P < 0,01).
3.2 Обнаружение КЦЖК и наблюдение структуры кишечника
Результаты газохроматографического анализа показали, что добавление пробиотиков значительно увеличивало содержание уксусной кислоты (I9: 12,37%, G15: 10,41% и X13: 14,23%), пропионовой кислоты (I9: 8,61%, G15: 8,78% и X13: 9,01%) и масляной кислоты (I9: 2,93%, G15: 3,17% и X13: 3,66%). Среди них креветки, получавшие X13, показали самое высокое общее количество КЦЖК (26,41%), за ними следовали I9 (23,92%) и G15 (22,85%) (Рисунок 1E).
Гистологический анализ показал, что кишечные ворсинки были длиннее у креветок, получавших пробиотические рационы, чем у креветок в группе PBS (Рисунки 1F–I). В гепатопанкреасе не было вакуолей; печеночные микрофибриллы были более плотно расположены в группах I9, G15 и X13 по сравнению с PBS (Рисунки 1J–M). Результаты показали, что добавление пробиотиков эффективно поддерживает здоровую морфологию тканей кишечника и гепатопанкреаса.
3.3 Активность ферментов и иммунный ответ
После эксперимента по кормлению гепатопанкреас креветок использовали для определения активности SOD, ALT, AST, T-AOC, CAT, AKP, ACP, AMS и MDA. Результаты показали, что добавление пробиотиков значительно повышало активность ферментов, включая SOD, ALT, AST, T-AOC, CAT, ACP и AMS, но значительно снижало содержание MDA (P < 0,05) (Рисунки 2A–I). Активность AKP была значительно выше в группе I9, тогда как активность ACP и AMS была значительно выше в группе X13 по сравнению с PBS (P < 0,05) (Рисунки 2A–I). Эти результаты позволяют предположить, что пробиотики могут улучшать антиоксидантную способность креветок.
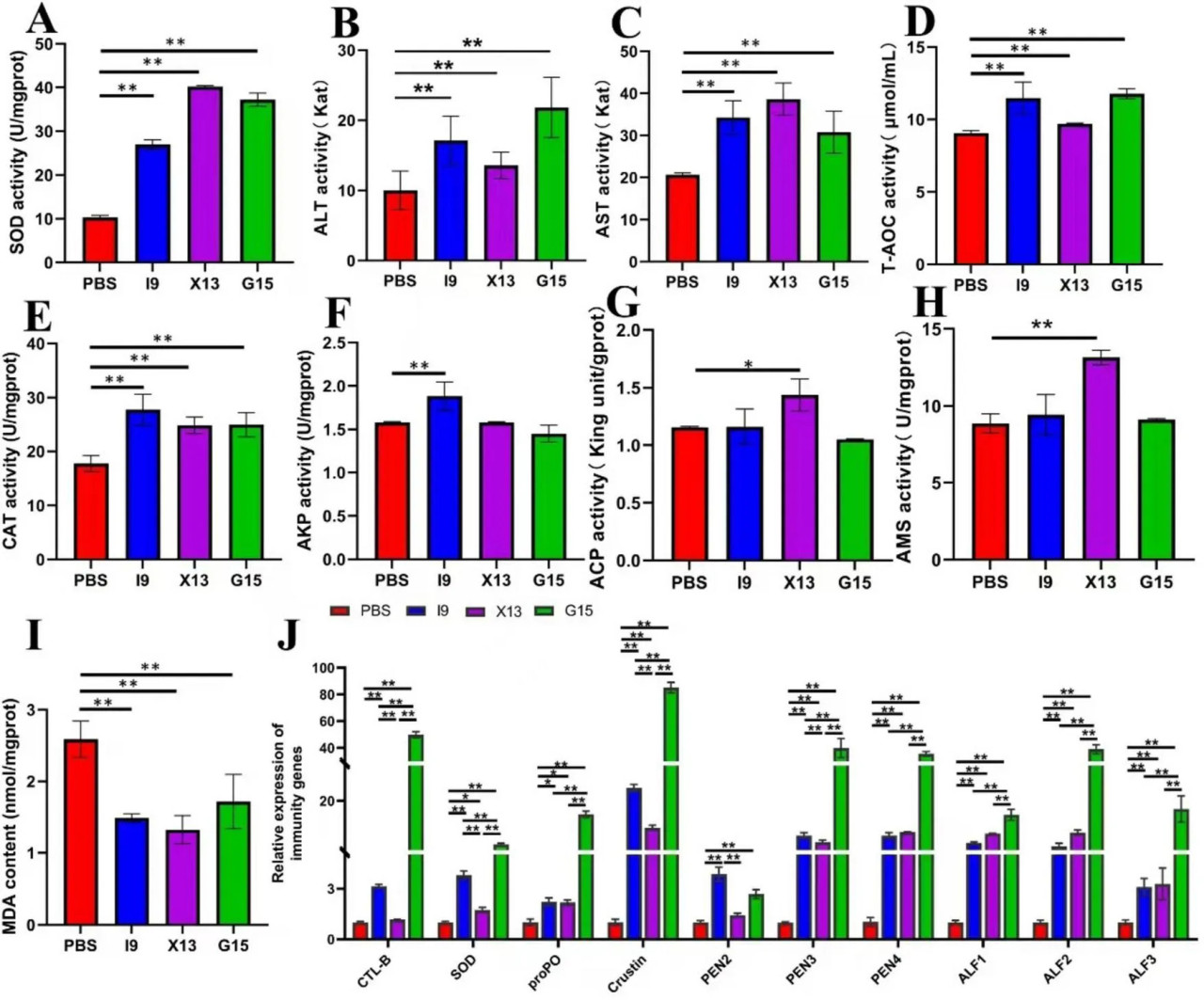
РИСУНОК 2 Изменения иммунной функции креветок в различных экспериментальных группах. (A–I) Представляют активность ферментов (SOD, ALT, AST, T-AOC, CAT, AKP, ACP, AMS и MDA) в гепатопанкреасе различных экспериментальных групп. (J) Экспрессия иммуносвязанных генов (CTL, SOD, proPO, Crustin, PEN2-4 и ALF1-3) в ткани кишечника. Сравнительный анализ различий между различными экспериментальными группами проводили с использованием дисперсионного анализа (ANOVA) с последующим критерием Тьюки HSD. Звездочки указывают на наличие статистически значимых различий (P < 0,05; P < 0,01).
Для оценки экспрессии иммуносвязанных генов в кишечнике креветок после кормления пробиотиками использовали ОТ-ПЦР в реальном времени. Результаты показали, что гены CTL, SOD, proPO, Crustin, PEN2, PEN3, PEN4, ALF1, ALF2 и ALF3 были значительно активированы в группах I9, X13 и G15 по сравнению с PBS (P < 0,05). Среди этих генов, за исключением PEN2, который имел самую высокую экспрессию в I9, остальные гены имели самую высокую экспрессию в G15 (Рисунок 2J). Эти результаты указывают на то, что добавление пробиотиков значительно улучшало иммунный ответ креветок.
3.4 Изменения в кишечной микробиоте
Изменения в кишечной микробиоте креветок после кормления в течение одного месяца рационами с добавлением PBS, I9, G15 или X13 были исследованы с использованием анализа секвенирования 16S рРНК. Кривые насыщения и разрежения показывают насыщение образцов на основе анализа высокопроизводительного секвенирования (Рисунок 3A). Показатель Good coverage был выше 99% для всех образцов, что указывает на то, что бактериальное сообщество в каждом образце было в значительной степени определено до глубины насыщения для секвенирования. Кривые рангового обилия различают обилие между образцами (Рисунок 3B). Всего из всех образцов было получено 1 593 224 высококачественных рида, и необработанные данные были загружены в Genbank под номером доступа PRJNA1143973. Количество ридов варьировалось от 64 477 до 117 644 (в PBS), от 102 586 до 105 085 (в I9), от 102 265 до 106 169 (в G15) и от 70 636 до 105 421 (в X13). Было обнаружено 289 общих операционных таксономических единиц (ОТЕ) между группами, и 216, 77, 449 и 121 уникальных ОТЕ были найдены в PBS, I9, G15 и X13 соответственно (Рисунок 3C).
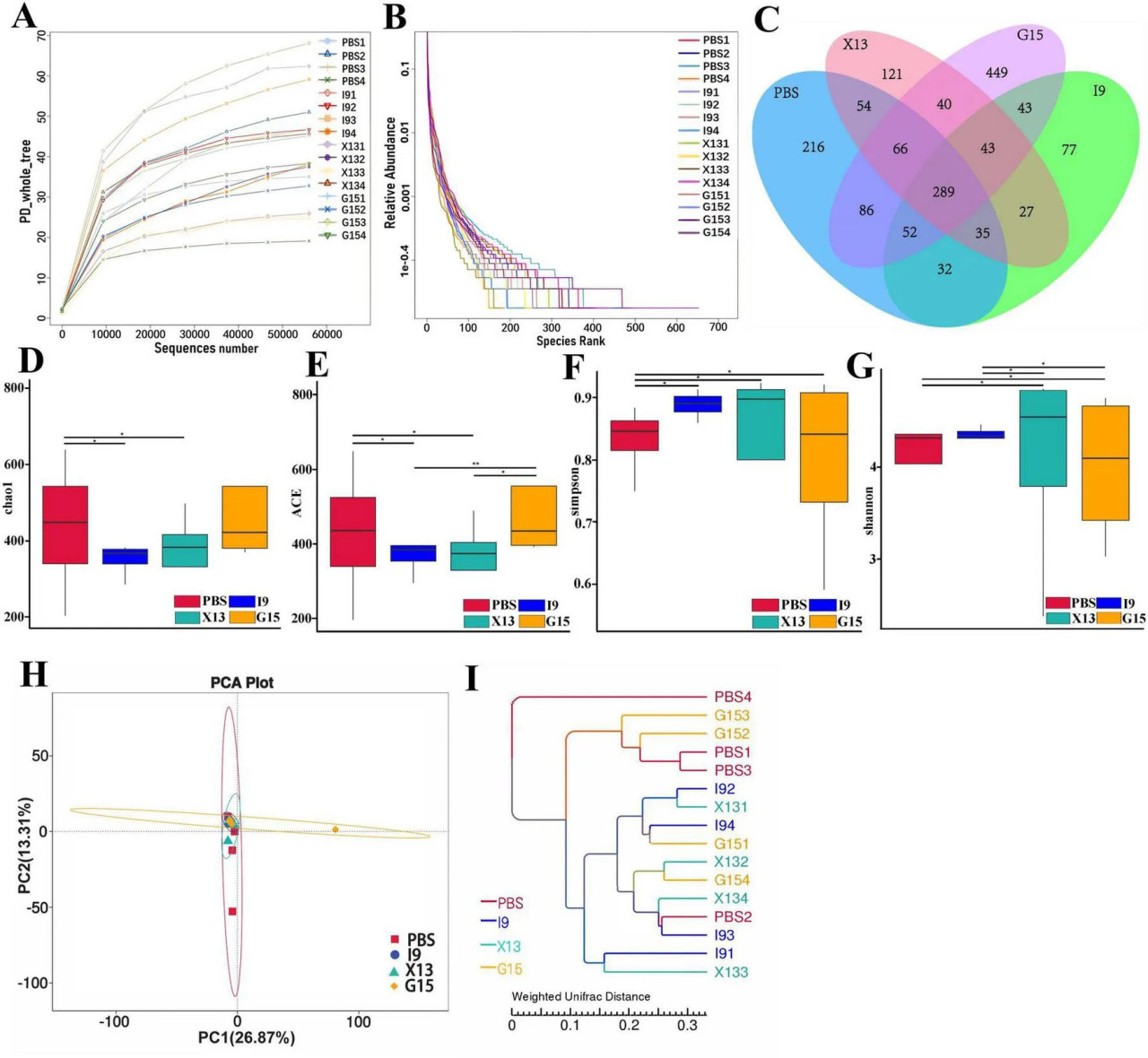
РИСУНОК 3 Изменения разнообразия кишечной микробиоты в различных экспериментальных группах. (A,B) Кривые разрежения, показывающие обилие кишечной микробиоты в каждой группе. (C) Количество общих и уникальных ОТЕ между группами. (D–G) Разнообразие и богатство кишечной микробиоты (на основе индексов Chao1, ACE, Simpson и Shannon). (H) Сходство структуры сообщества между группами выражено с использованием метода PCoA на основе взвешенных расстояний UniFrac. (I) Структура дерева кластеризации методом невзвешенного попарного группирования со средним арифметическим (UPGMA) использовалась для представления распределения кластеров между образцами. Сравнительный анализ различий между различными экспериментальными группами проводили с использованием дисперсионного анализа (ANOVA) с последующим критерием Тьюки HSD. Звездочки указывают на наличие статистически значимых различий (P < 0,05; P < 0,01).
Что касается альфа-разнообразия, результаты показали, что индексы Shannon и Simpson в группах I9, G15 и X13 были значительно выше, чем в PBS. Напротив, по сравнению с PBS, индексы ACE и Chao1 имели снижение в группах I9 и G15 (Рисунки 3D–G). Анализ бета-разнообразия показал различия в бактериальных сообществах в PBS по сравнению с другими группами. Анализы PCoA и WPGMA показали, что бактериальное сообщество в PBS было отделено от других (Рисунки 3H, I).
Были исследованы изменения в составе кишечной микробиоты креветок, получавших разные рационы. Обилие 10 наиболее распространенных бактериальных типов в группах показано на Рисунке 4A. Наблюдалось увеличение относительного обилия Proteobacteria, Bacteroidota, Actinobacteriota и Verrucomicrobiota (в I9), Firmicutes, Cyanobacteria и Verrucomicrobiota (в G15), а также Proteobacteria и Actinobacteriota (в X13) по сравнению с PBS (P < 0,05). Одновременно относительное обилие Firmicutes и Fusobacteriota (в I9), Firmicutes, Bacteroidota и Fusobacteriota (в X13), а также Actinobacteriota и Fusobacteriota (в G15) показало тенденцию к снижению по сравнению с PBS.
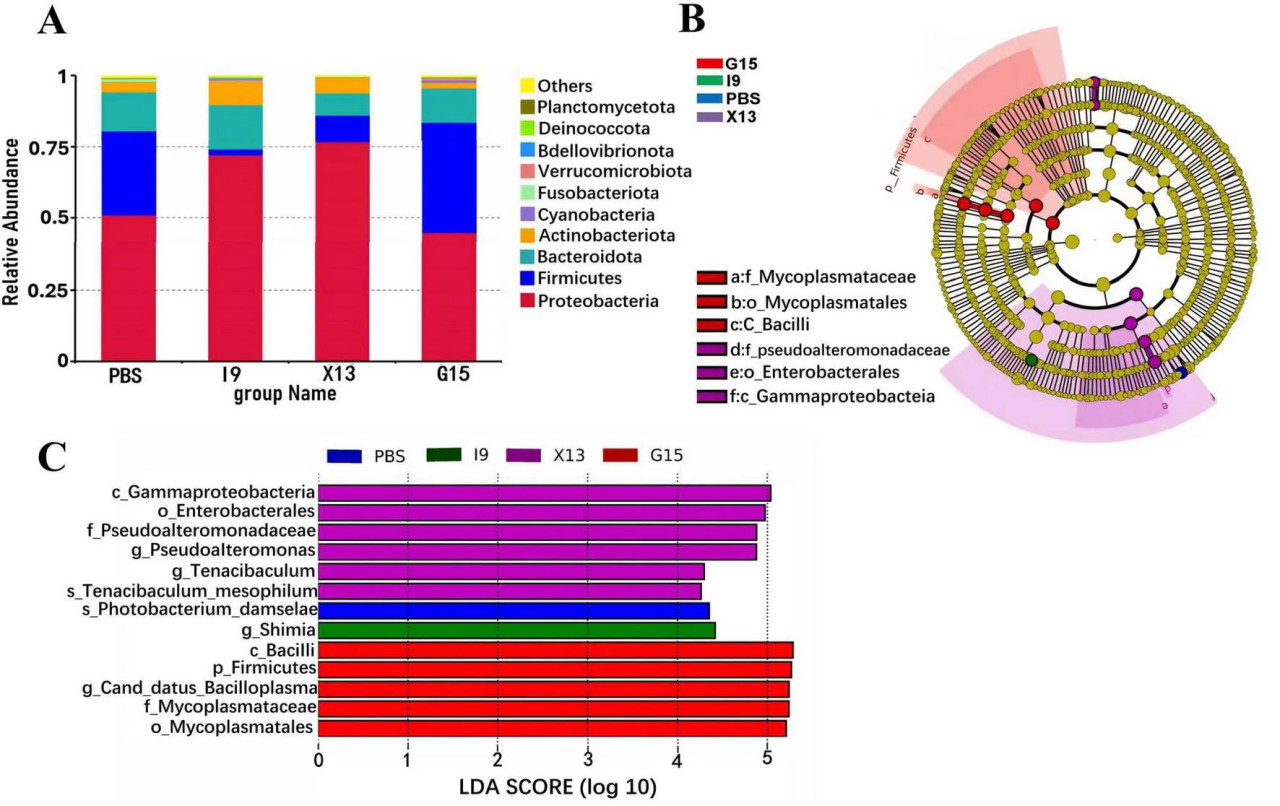
РИСУНОК 4 Состав микробного сообщества кишечника в различных экспериментальных группах. (A) Относительное обилие кишечной микробиоты на уровне типа. (B) LEfSe-анализ филогенетического распределения микробных таксонов. (C) Гистограмма распределения значений LDA, указывающих на значительное обилие микробиоты (LDA > 3,0).
LEfSe (LDA > 3,0) использовали для изучения участия специфических бактериальных таксонов в различных группах (Рисунки 4B, C). Бактериальные таксоны Gammaproteobacteria и Enterobacterales были значительно обогащены в X13, тогда как Firmicutes, Bacilli, Candidatus Bacilloplasma, Mycoplasmataceae и Mycoplasmateles были в G15 по сравнению с PBS.
Функциональный профиль кишечной микробиоты креветок в группах PBS, I9, G15 и X13 был получен с использованием PICRUSt-анализа. Результаты PCA показали разделение в микробных функциях в G15 по сравнению с PBS (Рисунок 5A). Распределение общих и уникальных генов в различных экспериментальных группах показано на диаграммах Венна (Рисунок 5B). Рисунок 5B показывает в общей сложности 5466 общих генов во всех группах и 87, 6, 135 и 19 уникальных генов в PBS, I9, G15 и X13 соответственно. Различия в функциях микробиоты между группами были проанализированы и показаны на Рисунке 5C. Добавление I9 и X13 значительно усиливало пути, связанные с метаболизмом (включая биодеградацию липидов и ксенобиотиков, углеводов, терпеноидов и поликетидов, а также биосинтез), систему кровообращения, мембранную систему и нервную систему. Добавление G15 увеличивало рост и гибель клеток, энергетический метаболизм и трансляцию, но снижало сердечно-сосудистую и дыхательную функции, трансляцию и сердечно-сосудистые заболевания.
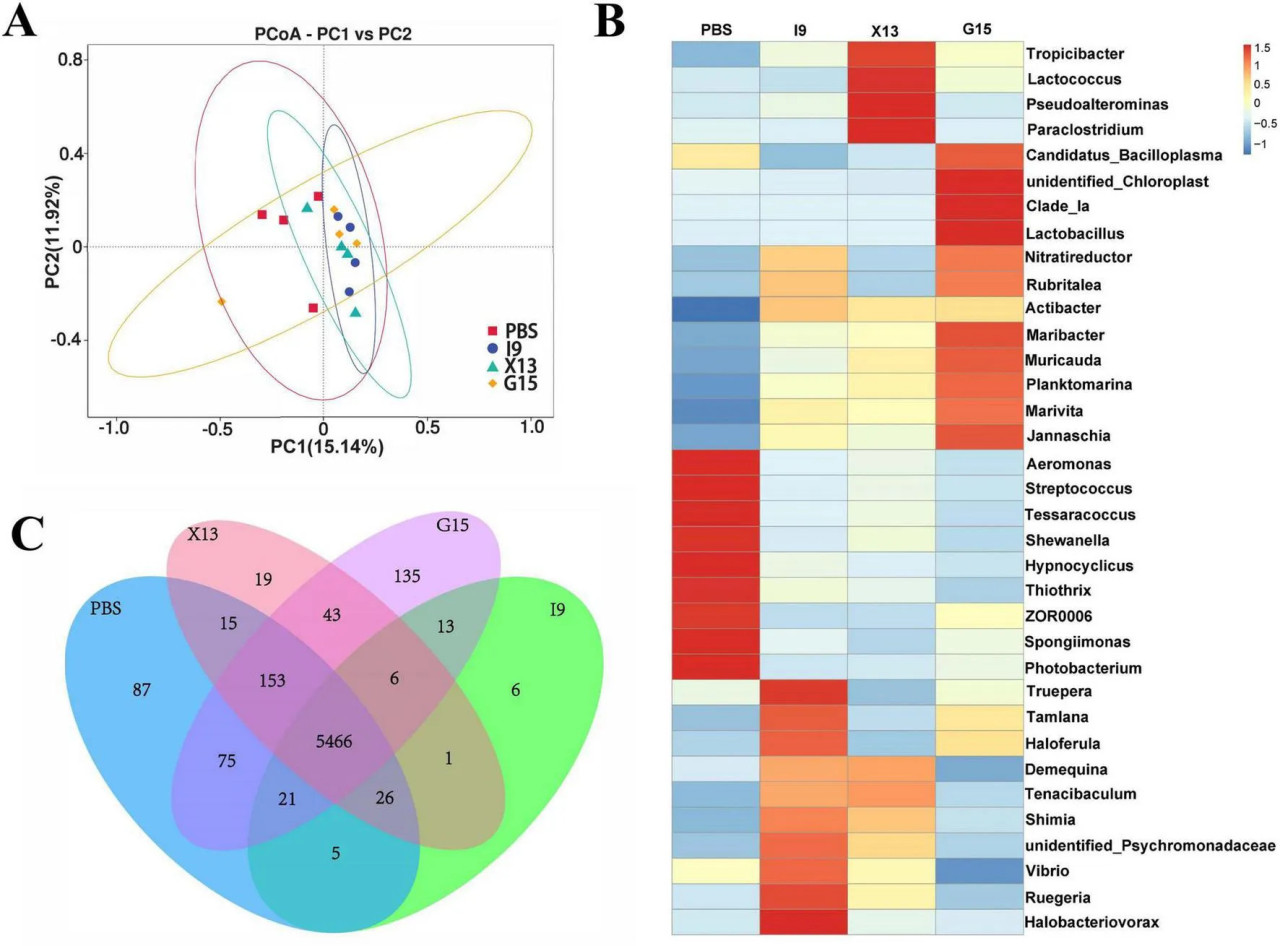
РИСУНОК 5 Функциональный профиль кишечной микробиоты в различных экспериментальных группах. (A) Прогнозирование функции кишечной микробиоты с помощью PICRUSt. График PCA-редукции размерности, показывающий функцию кишечной микробиоты. (B) Количество генов между группами представлено диаграммой Венна. (C) Относительное обилие и сигнальные пути кишечной микробиоты в экспериментальных группах были проанализированы (на уровне 2).
3.5 Выживаемость и иммунный ответ креветок после заражения Vp
По окончании эксперимента по кормлению креветок в каждой группе подвергали заражению Vp для оценки роли пробиотических бактерий в повышении устойчивости креветок к инфекции Vp. Результаты показали, что после стимуляции Vp выживаемость креветок в группах I9 + Vp (55,00%), G15 + Vp (50,83%) и X13 + Vp (49,17%) была значительно выше, чем в положительном контроле (Vp, 33,33%) (Рисунок 6A). Кроме того, наше исследование показало, что содержание КЦЖК (уксусной, пропионовой и масляной кислот) было значительно выше в группах I9 + Vp (26,53%), G15 + Vp (23,69%) и X13 + Vp (24,40%) по сравнению с положительным контролем (Vp, 10,66%) (Рисунок 6B). Гистопатологически ткани кишечника креветок в группе Vp показали уменьшение количества и длины кишечных ворсинок (Рисунки 6C–G). Ткани гепатопанкреаса группы Vp показали множество вакуолей, тогда как ткани кишечника и гепатопанкреаса креветок в группах I9 + Vp, G15 + Vp и X13 + Vp показали нормальную гистологическую структуру (Рисунки 6H–L).
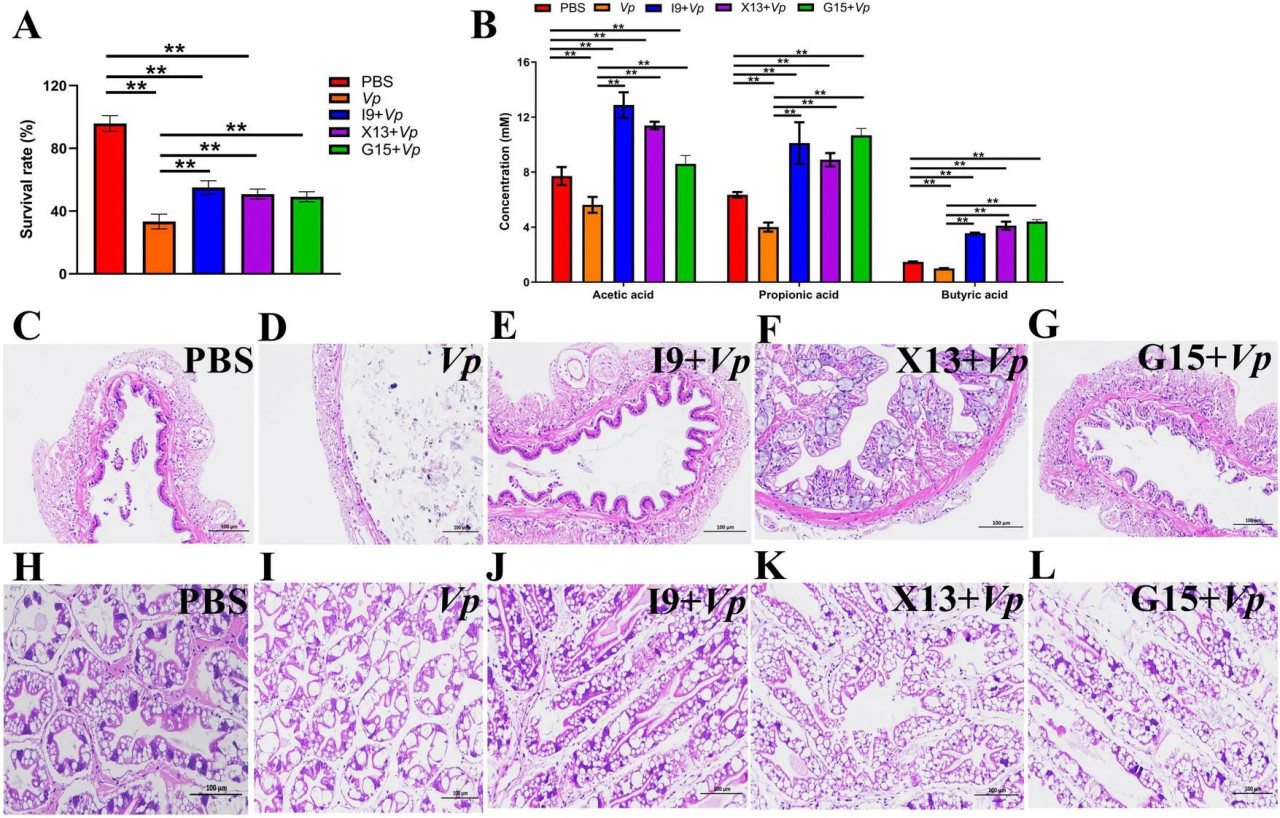
РИСУНОК 6 Изменения выживаемости, продукции КЦЖК и гистологии тканей креветок в различных экспериментальных группах после заражения Vp. (A) Выживаемость. (B) Продукция КЦЖК в содержимом кишечника. (C–G) Структура кишечника. (H–L) Структура гепатопанкреаса. Сравнительный анализ различий между различными экспериментальными группами проводили с использованием дисперсионного анализа (ANOVA) с последующим критерием Тьюки HSD. Звездочки указывают на наличие статистически значимых различий (P < 0,05; P < 0,01).
Активность ферментов, включая SOD, ALT, AST, T-AOC, CAT, AKP и ACP, была значительно повышена, но активность AMS и MDA была значительно снижена в группах I9 + Vp, G15 + Vp и X13 + Vp по сравнению с положительным контролем (Vp) (Рисунки 7A–I). Кроме того, экспрессия иммуносвязанных генов, включая CTL, SOD, proPO, Crustin, PEN2-4 и ALF1-3, была значительно повышена в группах I9 + Vp, G15 + Vp и X13 + Vp после заражения Vp по сравнению с PBS (Рисунок 7J). Эти результаты указали на то, что добавление пробиотиков может улучшать иммунитет и устойчивость к болезням креветок против инфекции Vp.
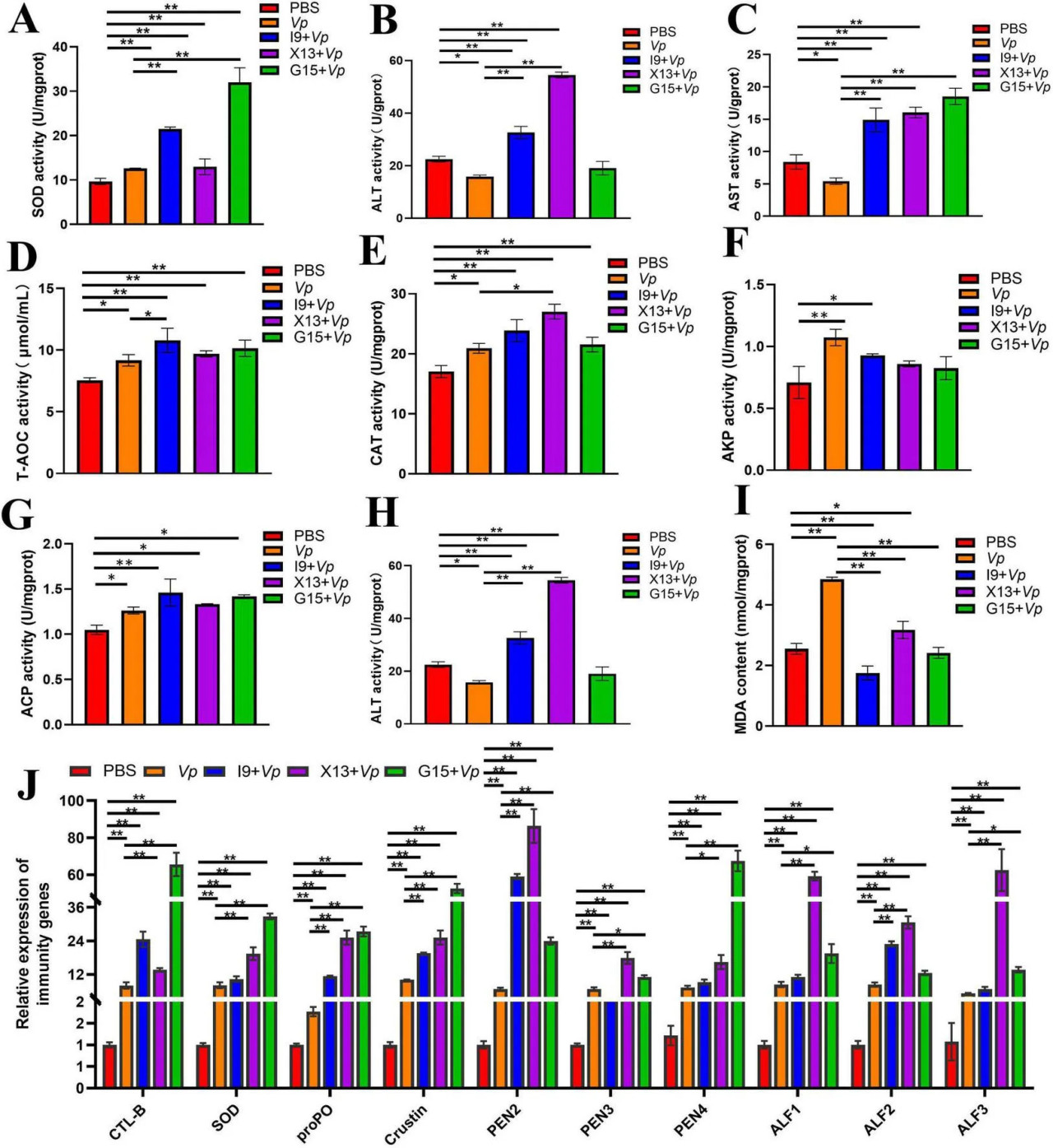
РИСУНОК 7 Изменения иммунной функции креветок в различных экспериментальных группах после заражения Vp. (A–I) Активность ферментов (SOD, ALT, AST, T-AOC, CAT, AKP, ACP, AMS и MDA) в гепатопанкреасе. (J) Экспрессия иммуносвязанных генов (CTL, SOD, proPO, Crustin, PEN2-4 и ALF1-3) в ткани кишечника. Сравнительный анализ различий между различными экспериментальными группами проводили с использованием дисперсионного анализа (ANOVA) с последующим критерием Тьюки HSD. Звездочки указывают на наличие статистически значимых различий (P < 0,05; P < 0,01).
4 Обсуждение
Гомеостаз в кишечнике является одним из важнейших факторов, влияющих на рост и иммунитет креветок (Hou et al., 2018). Исследования показали, что Vp и WSSV являются основными факторами, разрушающими микробный гомеостаз и приводящими к массовой гибели креветок в интенсивной аквакультуре (Zhang et al., 2016). В креветководстве существует множество способов борьбы с проникновением патогенных бактерий, таких как иммунная регуляция, лечение антибиотиками, хорошее управление качеством воды, рациональное управление кормлением, генетическая селекция, а также регулярный мониторинг и профилактика. Среди них пробиотики для защиты хозяина от заражения бактериальными патогенами стали новым способом профилактики болезней креветок в последние годы (Xia et al., 2014). Сообщалось, что добавление пробиотиков в корма не только способствует росту креветок, но и улучшает их антиоксидантную способность и иммунитет путем модуляции обилия кишечной микробиоты (Imaizumi et al., 2021).
В этом исследовании кормление I9, G15 или X13 значительно повышало выживаемость, удельную скорость роста, темп прироста массы и темп прироста длины креветок. В предыдущем исследовании сообщалось, что добавление C. butyricum CB1-3 в рационы может приносить пользу здоровью и модулировать кишечную микробиоту ювенильных тихоокеанских белых креветок в условиях выращивания (Zhang et al., 2023). Кормление C. butyricum CB в течение 56 дней увеличивало содержание общей антиоксидантной способности (T-AOC), активность лизоцима (LSZ) и относительную экспрессию генов Toll и иммунодефицита (Imd), улучшало ростовые показатели L. vannamei (Duan et al., 2017), что показало эффекты, сходные с результатами данного исследования. Здесь продукция КЦЖК, таких как уксусная, пропионовая и масляная кислоты, в кишечнике креветок также значительно увеличивалась после кормления пробиотиками, что было сходно с предыдущим сообщением о том, что пробиотики человеческого происхождения Lactobacilli paracasei D3-5 и Enterococci raffinosus D24-1 могут увеличивать содержание КЦЖК путем модуляции кишечной микробиоты у людей и мышей, тем самым улучшая дисбиоз кишечной микробиоты (Nagpal et al., 2018). Эти результаты позволили предположить, что добавление пробиотиков может значительно увеличивать продукцию КЦЖК в кишечнике хозяина, эффективно улучшая рост креветок. В этом исследовании кормление пробиотиками I9, G15 или X13 было эффективным для поддержания нормальной структуры кишечника, что в основном подтверждалось количеством видимых кишечных ворсинок и отчетливых складок. Это сходно с исследованием, в котором кормление C. butyricum CBG01 в течение 42 дней значительно увеличивало высоту кишечных ворсинок и толщину стенки кишечника у тихоокеанской белой креветки (Li et al., 2019).
Более того, рационы с добавлением I9, G15 или X13 способствовали повышению активности SOD, ALT, AST, T-AOC, CAT, ACP и AMS, но снижали содержание MDA в гепатопанкреасе креветок. Это указывало на то, что добавление пробиотиков влияет на активность ферментов и улучшает иммунные показатели, дополнительно способствуя росту хозяина. Это согласуется с результатами предыдущего исследования, в котором добавление C. butyricum G13 приводило к значительному увеличению активности SOD, CAT, AKP и ACP у крабов-плавунцов (Liang et al., 2023). Между тем, повышение экспрессии иммуносвязанных генов у креветок, получавших обогащенные пробиотиками рационы, здесь указало на то, что эти бактерии оказывают благотворное влияние на иммунную систему креветок. Наши выводы подтверждаются наблюдениями в предыдущем исследовании, в котором экспрессия CAT, LYS, proPO и SOD значительно увеличивалась в гепатопанкреасе крабов-плавунцов, получавших Pediococcus pentosaceus G11 (Yang et al., 2019). Диетическая добавка Bacillus subtilis DCU или Bacillus shortus BP значительно стимулировала экспрессию CAT и proPO, тем самым дополнительно способствуя иммунным показателям и ростовому статусу крабов-плавунцов (Wu et al., 2014). Таким образом, результаты этого исследования показали положительные эффекты I9, G15 и X13 в стимуляции антиоксидантной способности и иммунного ответа креветок.
Баланс кишечной микробиоты оказывает важное влияние на рост, развитие, переваривание и усвоение питательных веществ, а также иммунную функцию животных (MacDonald et al., 2011). Кишечный гомеостаз связан с воспалительным ответом, который влияет на предотвращение колонизации патогенами (Maloy и Powrie, 2011). Это исследование показало, что кишечная микробиота тесно связана с гомеостазом организма; патогены напрямую повреждают барьерную функцию кишечника, приводя к дисбиозу кишечной микробиоты. Xie et al. (2019) указали, что диетическая добавка 1000 или 2000 мг/кг пробиотиков значительно увеличивала богатство микробного разнообразия кишечника. Это исследование показало, что индексы Chao1, ACE, Shannon и Simpson были увеличены в группах I9, G15 и X13, и что микробное сообщество в этих группах (особенно G15) отличалось от PBS. Результаты указали на то, что добавление пробиотических бактерий эффективно улучшало разнообразие кишечной микробиоты, что, вероятно, поддерживает иммунитет креветок.
Добавление I9, G15 и X13 показало значительные различия в составе кишечной микробиоты по сравнению с PBS, со значительным увеличением обилия Proteobacteria и Firmicutes, что позволяет предположить, что пробиотики модулируют состав кишечной микробиоты. Это сходно с предыдущим исследованием, в котором применение C. butyricum изменяет структуру кишечной микробиоты с увеличением относительного обилия Proteobacteria и Firmicutes у большеротого окуня (Micropterus salmoides) (Meng et al., 2023). Другое исследование показало, что обилие Bacteroidetes и Firmicutes увеличивалось при кормлении тиляпии диетической добавкой C. butyricum CB2. Добавка корма C. butyricum C2 способствовала росту Bifidobacterium bifidum, но ингибировала рост Escherichia coli и Puccinia в средней и задней кишке горбыля Мии (Miichthys miiuy) (Song и Wu, 2007). Кроме того, в настоящем исследовании добавление I9 и X13 значительно усиливало метаболические процессы биодеградации липидов и ксенобиотиков, аминосахаров и терпеноидов. Это сходно с результатами предыдущего исследования, в котором диетическая добавка C. butyricum увеличивала кишечные уровни лизофосфатидилхолина и снижала связанный с этим пуриновый метаболизм у китайского мохнаторукого краба (Eriocheir sinensis) (Gao et al., 2023). В совокупности добавление пробиотиков стимулировало рост полезных бактерий и дополнительно влияло на рост и иммунный ответ креветок.
Инфекция Vp является одним из инфекционных заболеваний в креветководстве (Proespraiwong et al., 2023). Были исследованы эффекты I9, G15 или X13 на изменения физиологических и иммунных реакций креветок после заражения Vp. Результаты показали, что выживаемость креветок значительно улучшалась после заражения Vp в группах с добавлением пробиотиков. Это сходно с предыдущим исследованием, в котором C. butyricum CB30 значительно улучшал способность к выживанию и поддержанию нормальной структуры кишечника тихоокеанской белой креветки после заражения V. alginolyticus (Wang et al., 2023). Кроме того, добавление фагов (A3S и Vpms1) снижало смертность тихоокеанской белой креветки, инфицированной Vp (Lomelí-Ortega и Martínez-Díaz, 2014). В этом текущем исследовании кишечные ворсинки креветок показали значительное отслоение и разделение после заражения Vp, но оно сохранялось в нормальной структуре при кормлении I9, G15 или X13. Это сходно с результатами предыдущего исследования, в котором кормление C. butyricum G13 поддерживало нормальную структуру кишечника крабов-плавунцов после заражения Vp (Liang et al., 2023). Кроме того, добавление Alcaligenes sp. AFG22 показало значительное увеличение длины, ширины и площади ворсинок, что полезно для поддержания кишечной микробиоты и стимулирования усвоения питательных веществ у ювенильной Tor tambroides (Asaduzzaman et al., 2018). Продукция КЦЖК, особенно масляной кислоты, была значительно выше в группах I9 + Vp, G15 + Vp и X13 + Vp, чем в группе Vp. Это согласуется с выводами предыдущего исследования, в котором C. butyricum CB1-3 может повышать ростовые показатели, перевариваемость, КЦЖК в кишечнике, иммунный ответ и устойчивость к аммиачному стрессу у креветок (L. vannamei) (Duan et al., 2017). Кроме того, было обнаружено повышение активности ферментов (включая SOD, ALT, AST, T-AOC, CAT, AKP и ACP) и снижение AMS и MDA в группах I9 + Vp, G15 + Vp и X13 + Vp. Это согласуется с предыдущим исследованием на L. vannamei, в котором диетическая добавка Enterobacter cloacae E3 и Lactobacillus lactis L3 повышала активность SOD, PPO, ACP, POD, CAT и LZM и улучшала иммунные показатели креветок (Zuo et al., 2019). Известно, что T-AOC, T-SOD и CAT являются важными антиоксидантными ферментами, образующими первый уровень защиты путем предотвращения образования свободных радикалов (Wołonciej et al., 2016). AKP является ключевым компонентом защиты от патогенов и окислительного стресса (Callewaert и Michiels, 2010). Повышенная активность этих ферментов способствовала антиоксидантной способности и иммунитету тихоокеанских белых креветок в ответ на инфекцию Vp. В этом исследовании мы также обнаружили, что экспрессия иммуносвязанных генов (включая CTL, SOD, proPO, Crustin и PEN4) была значительно повышена в группах I9 + Vp, G15 + Vp и X13 + Vp. Это указало на то, что I9, G15 и X13 стимулируют иммунный ответ для поддержания здоровья кишечника и защиты хозяина от инфекции. Эти выводы были сходны со случаем тихоокеанской белой креветки, получавшей корм с добавлением Aspergillus niger (Zhang et al., 2023). Таким образом, результаты этого исследования продемонстрировали положительные эффекты I9, G15 и X13 на рост, кишечную микробиоту, иммунный ответ и устойчивость к болезням тихоокеанской белой креветки.
5 Заключение
Это исследование показало, что бактерии I9, G15 и X13 обладают пробиотическими свойствами в культивировании тихоокеанской белой креветки. Добавление этих бактерий может эффективно способствовать росту, изменять разнообразие и структуру кишечной микробиоты, с увеличением обилия полезной микробиоты и продукции КЦЖК в кишечнике, поддерживать нормальную структуру кишечника и усиливать антиоксидантную активность, иммунный ответ и устойчивость к болезням (против Vp) у креветок. В совокупности наши результаты указывают на то, что I9, G15 и X13 могут использоваться в качестве кормовых добавок для стимулирования роста и профилактики заболеваний в креветководстве.
Сноски
1. http://ccb.jhu.edu/software/FLASH/
2. http://www.drive5.com/uparse/
Ссылки
1. Asaduzzaman M., Iehata S., Akter S., Kader M. A., Ghosh S. K., Khan M. N. A., et al. (2018). Effects of host gut-derived probiotic bacteria on gut morphology, microbiota composition and volatile short chain fatty acids production of Malaysian Mahseer Tor tambroides. Aquacult. Rep. 9, 53–61. doi: 10.1016/j.aqrep.2017.12.003. CrossRef. Google Scholar.
2. Aziz R. K., Zhou J., Fang W., Yang X., Zhou S., Hu L., et al. (2012). A nonluminescent and highly virulent Vibrio harveyi strain is associated with "Bacterial White Tail Disease" of Litopenaeus vannamei Shrimp. PLoS One 7:e0029961. doi: 10.1371/journal.pone.0029961. CrossRef. Google Scholar.
3. Bokulich N. A., Subramanian S., Faith J. J., Gevers D., Gordon J. I., Knight R., et al. (2013). Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 10, 57–59. doi: 10.1038/nmeth.2276. CrossRef. Google Scholar.
4. Cahú T. B., Santos S. D., Mendes A., Córdula C. R., Chavante S. F., Carvalho L. B., et al. (2012). Recovery of protein, chitin, carotenoids and glycosaminoglycans from Pacific white shrimp (Litopenaeus vannamei) processing waste. Process Biochem. 47, 570–577. doi: 10.1016/j.procbio.2011.12.012. CrossRef. Google Scholar.
5. Callewaert L., Michiels C. W. (2010). Lysozymes in the animal kingdom. J. Biosci. 35, 127–160. doi: 10.1007/s12038-010-0015-5. CrossRef. Google Scholar.
6. Chang Y. P., Liu C. H., Wu C. C., Chiang C. M., Lian J. L., Hsieh S. L. (2012). Dietary administration of zingerone to enhance growth, non-specific immune response, and resistance to Vibrio alginolyticus in Pacific white shrimp (Litopenaeus vannamei) juveniles. Fish Shellfish Immunol. 32, 284–290. doi: 10.1016/j.fsi.2011.11.017. CrossRef. Google Scholar.
7. Cornejo-Granados F., Lopez-Zavala A. A., Gallardo-Becerra L., Mendoza-Vargas A., Sánchez F., Vichido R., et al. (2017). Microbiome of Pacific whiteleg shrimp reveals differential bacterial community composition between Wild, Aquacultured and AHPND/EMS outbreak conditions. Sci. Rep. 7:11783. doi: 10.1038/s41598-017-11805-w. CrossRef. Google Scholar.
8. Duan Y., Zhang Y., Dong H., Wang Y., Zheng X., Zhang J. (2017). Effect of dietary Clostridium butyricum on growth, intestine health status and resistance to ammonia stress in Pacific white shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 65, 25–33. doi: 10.1016/j.fsi.2017.03.048. CrossRef. Google Scholar.
9. Edgar R. C. (2013). UPARSE: highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 10, 996–998. doi: 10.1038/nmeth.2604. CrossRef. Google Scholar.
10. Edgar R. C., Haas B. J., Clemente J. C., Quince C., Knight R. (2011). UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 27, 2194–2200. doi: 10.1093/bioinformatics/btr381. CrossRef. Google Scholar.
11. Gao X., Liu X., Wang Y., Wang T., Fang D., Hu K. (2023). Effects of Clostridium butyricum on intestinal microflora and metabolism of Eriocheir sinensis. Int. J. Mol. Sci. 24:13784. doi: 10.3390/ijms241813784. CrossRef. Google Scholar.
12. Hou D., Huang Z., Zeng S., Liu J., Wei D., Deng X., et al. (2018). Intestinal bacterial signatures of white feces syndrome in shrimp. Appl. Microbiol. Biotechnol. 102, 3701–3709. doi: 10.1007/s00253-018-8855-2. CrossRef. Google Scholar.
13. Imaizumi K., Tinwongger S., Kondo H., Hirono I. (2021). Analysis of microbiota in the stomach and midgut of two penaeid shrimps during probiotic feeding. Sci. Rep. 11:9936. doi: 10.1038/s41598-021-89415-w. CrossRef. Google Scholar.
14. Jin M., Xiong J., Zhou Q. C., Yuan Y., Wang X. X., Sun P. J. F., et al. (2018). Dietary yeast hydrolysate and brewer's yeast supplementation could enhance growth performance, innate immunity capacity and ammonia nitrogen stress resistance ability of Pacific white shrimp (Litopenaeus vannamei). Fish Shellfish Immunol. 82, 121–129. doi: 10.1016/j.fsi.2018.08.020. CrossRef. Google Scholar.
15. Kumar B. K., Deekshit V. K., Raj J. R. M., Rai P., Shivanagowda B. M., Karunasagar I., et al. (2014). Diversity of Vibrio parahaemolyticus associated with disease outbreak among cultured Litopenaeus vannamei (Pacific white shrimp) in India. Aquaculture 433, 247–251. doi: 10.1016/j.aquaculture.2014.06.016. CrossRef. Google Scholar.
16. Li H., Tian X., Zhao K., Jiang W., Dong S. (2019). Effect of Clostridium butyricum in different forms on growth performance, disease resistance, expression of genes involved in immune responses and mTOR signaling pathway of Litopenaeus vannamai. Fish Shellfish Immunol. 87, 13–21. doi: 10.1016/j.fsi.2018.12.069. CrossRef. Google Scholar.
17. Liang H., Tran N. T., Deng T., Li J., Lei Y., Bakky M. A. H., et al. (2023). Identification and characterization of a potential probiotic, Clostridium butyricum G13, isolated from the intestine of the mud crab (Scylla paramamosain). Microbiol. Spectr. 11:e01317-23. doi: 10.1128/spectrum.01317-23. CrossRef. Google Scholar.
18. Liu G., Zhu S., Liu D., Guo X., Ye Z. (2017). Effects of stocking density of the white shrimp Litopenaeus vannamei (Boone) on immunities, antioxidant status, and resistance against Vibrio harveyi in a biofloc system. Fish Shellfish Immunol. 67, 19–26. doi: 10.1016/j.fsi.2017.05.038. CrossRef. Google Scholar.
19. Lomelí-Ortega O. C., Martínez-Díaz F. S. (2014). Phage therapy against Vibrio parahaemolyticus infection in the whiteleg shrimp (Litopenaeus vannamei) larvae. Aquaculture 2014, 208–211. doi: 10.1016/j.aquaculture.2014.08.018. CrossRef. Google Scholar.
20. MacDonald T. T., Monteleone I., Fantini M. C., Monteleone G. (2011). Regulation of homeostasis and inflammation in the intestine. Gastroenterology 140, 1768–1775. doi: 10.1053/j.gastro.2011.02.047. CrossRef. Google Scholar.
21. Magoè T., Salzberg S. L. (2011). FLASH: fast length adjustment of short reads to improve genome assemblies. Bioinformatics 27, 2957–2963. doi: 10.1093/bioinformatics/btr507. CrossRef. Google Scholar.
22. Maloy K. J., Powrie F. (2011). Intestinal homeostasis and its breakdown in inflammatory bowel disease. Nature 474, 298–306. doi: 10.1038/nature10208. CrossRef. Google Scholar.
23. Meng X., Cai H., Li H., You F., Jiang A., Hu W., et al. (2023). Clostridium butyricum-fermented Chinese herbal medicine enhances the immunity by modulating the intestinal microflora of largemouth bass (Micropterus salmoides). Aquaculture 562:738768. doi: 10.1016/j.aquaculture.2022.738768. CrossRef. Google Scholar.
24. Mo W. Y., Chen Z., Leung H. M., Leung A. O. W. (2015). Application of veterinary antibiotics in China's aquaculture industry and their potential human health risks. Environ. Sci. Pollut. Res. 24, 8978–8989. doi: 10.1007/s11356-015-5607-z. CrossRef. Google Scholar.
25. Nagpal R., Wang S., Ahmadi S., Hayes J., Gagliano J., Subashchandrabose S., et al. (2018). Human-origin probiotic cocktail increases short-chain fatty acid production via modulation of mice and human gut microbiome. Sci. Rep. 8:12649. doi: 10.1038/s41598-018-30114-4. CrossRef. Google Scholar.
26. Nimrat S., Suksawat S., Boonthai T., Vuthiphandchai V. J. (2012). Potential Bacillus probiotics enhance bacterial numbers, water quality and growth during early development of white shrimp (Litopenaeus vannamei). Vet. Microbiol. 159, 443–450. doi: 10.1016/j.vetmic.2012.04.029. CrossRef. Google Scholar.
27. Okocha R. C., Olatoye I. O., Adedeji O. B. (2018). Food safety impacts of antimicrobial use and their residues in aquaculture. Public Health Rev. 39:21. doi: 10.1186/s40985-018-0099-2. CrossRef. Google Scholar.
28. Panigrahi A., Esakkiraj P., Saranya C., Das R. R., Sundaram M., Sudheer N. S., et al. (2022). A biofloc-based aquaculture system bio-augmented with probiotic bacteria Bacillus tequilensis AP BFT3 improves culture environment, production performances, and proteomic changes in Penaeus vannamei. Probiotics Antimicrob. Proteins 14, 277–287. doi: 10.1007/s12602-022-09926-4. CrossRef. Google Scholar.
29. Pérez-Sánchez T., Mora-Sánchez B., Balcázar J. L. (2018). Biological approaches for disease control in aquaculture: Advantages, limitations and challenges. Trends Microbiol. 26, 896–903. doi: 10.1016/j.tim.2018.05.002. CrossRef. Google Scholar.
30. Proespraiwong P., Mavichak R., Imaizumi K., Hirono I., Unajak S. (2023). Evaluation of Bacillus spp. as potent probiotics with reduction in AHPND-related mortality and facilitating growth performance of pacific white shrimp (Litopenaeus vannamei) farms. Microorganisms 11:2176. doi: 10.3390/microorganisms11092176. CrossRef. Google Scholar.
31. Song Z. F., Wu T. X. (2007). Studies on the effect of Clostridium butyricum C2 strain on the intestinal flora of miiuy croaker (Miichthys miiuy) (in Chinese). J. Hydroecol. 04, 91–93. doi: 10.15928/j.1674-3075.2007.04.035. CrossRef. Google Scholar.
32. Song Z., Li K., Li K. (2024). Acute effects of the environmental probiotics Rhodobacter sphaeroides on intestinal bacteria and transcriptome in shrimp Penaeus vannamei. Fish Shellfish Immunol. 145:109316. doi: 10.1016/j.fsi.2023.109316. CrossRef. Google Scholar.
33. Tran N. T., Li Z., Ma H., Zhang Y., Zheng H., Gong Y., et al. (2020b). Clostridium butyricum: a promising probiotic confers positive health benefits in aquatic animals. Aquaculture 12, 2573–2589. doi: 10.1111/raq.12459. CrossRef. Google Scholar.
34. Tran N. T., Li Z., Wang S., Zheng H., Aweya J., Wen X., et al. (2020a). Progress and perspectives of short-chain fatty acids in aquaculture. Aquaculture 12, 283–298. doi: 10.1111/raq.12317. CrossRef. Google Scholar.
35. Tran N. T., Liang H., Li J., Deng T., Zhang M., Li S. (2023). Health benefits of butyrate and its producing bacterium, Clostridium butyricum, on aquatic animals. Fish Shellfish Immunol. Rep. 4:100088. doi: 10.1016/j.fsirep.2023.100088. CrossRef. Google Scholar.
36. Wang C., Li P. F., Hu D. G., Wang H. (2023). Effect of Clostridium butyricum on intestinal microbiota and resistance to Vibrio alginolyticus of Penaeus vannamei. Fish Shellfish Immunol. 138:108790. doi: 10.1016/j.fsi.2023.108790. CrossRef. Google Scholar.
37. Wołonciej M., Milewska E., Roszkowska-Jakimiec W. (2016). Trace elements as an activator of antioxidant enzymes. Postepy Hig. Med. Dosw. 70, 1483–1498. doi: 10.5604/17322693.1229074. CrossRef. Google Scholar.
38. Wu H. J., Sun L. B., Li C. B., Li Z. Z., Zhang Z., Wen X. B., et al. (2014). Enhancement of the immune response and protection against Vibrio parahaemolyticus by indigenous probiotic Bacillus strains in mud crab (Scylla paramamosain). Fish Shellfish Immunol. 41, 156–162. doi: 10.1016/j.fsi.2014.08.027. CrossRef. Google Scholar.
39. Wu Y. S., Chu Y. T., Chen Y. Y., Chang C. S., Lee B. H., Nan F. H. (2022). Effects of dietary Lactobacillus reuteri and Pediococcus acidilactici on the cultured water qualities, the growth and non-specific immune responses of Penaeus vannamei. Fish Shellfish Immunol. 127, 176–186. doi: 10.1016/j.fsi.2022.06.004. CrossRef. Google Scholar.
40. Xia Z., Zhu M., Zhang Y. (2014). Effects of the probiotic Arthrobacter sp. CW9 on the survival and immune status of white shrimp (Penaeus vannamei). Lett. Appl. Microbiol. 58, 60–64. doi: 10.1111/lam.12156. CrossRef. Google Scholar.
41. Xie J., Liu Q., Liao S., Fang H., Peng Y., Xie S., et al. (2019). Effects of dietary mixed probiotics on growth, non-specific immunity, intestinal morphology and microbiota of juvenile pacific white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol. 90, 456–465. doi: 10.1016/j.fsi.2019.04.301. CrossRef. Google Scholar.
42. Yang Q., Lv Y., Zhang M., Gong Y., Li Z., Tran N. T., et al. (2019). Lactic acid bacteria, Enterococcus faecalis Y17 and Pediococcus pentosaceus G11, improved growth performance, and immunity of mud crab (Scylla paramamosain). Fish Shellfish Immunol. 93, 135–143. doi: 10.1016/j.fsi.2019.07.050. CrossRef. Google Scholar.
43. Zhang M., Liang H., Lei Y., Zhang Y., Tan Z., Chen W., et al. (2023). Aspergillus niger confers health benefits and modulates the gut microbiota of juvenile Pacific white shrimp (Penaeus vannamei) under farming conditions. Front. Mar. Sci. 10:3389. doi: 10.3389/fmars.2023.1211993. CrossRef. Google Scholar.
44. Zhang X., Song X., Huang J. (2016). Impact of Vibrio parahaemolyticus and white spot syndrome virus (WSSV) co-infection on survival of penaeid shrimp Litopenaeus vannamei. Chin. J. Oceanol. Limnol. 34, 1278–1286. doi: 10.1007/s00343-016-5165-3. CrossRef. Google Scholar.
45. Zuo Z. H., Shang B. J., Shao Y. C., Li W. Y., Sun J. S. (2019). Screening of intestinal probiotics and the effects of feeding probiotics on the growth, immune, digestive enzyme activity and intestinal flora of Litopenaeus vannamei. Fish Shellfish Immunol. 86, 160–168. doi: 10.1016/j.fsi.2018.11.003. CrossRef. Google Scholar.

Yang W, Liang H, Chen R, Du Z, Deng T, Zheng Y, Song Y, Duan Y, Lin J, Bakky MAH, Tran NT, Zhang M and Li S (2024) Effects of dietary probiotic (Clostridium butyricum I9, C. butyricum G15, or Paraclostridium bifermentans X13) on growth, digestive enzyme activities, immunity, and intestinal microbiota of Pacific white shrimp (Penaeus vannamei). Front. Microbiol. 15:1479446. doi: 10.3389/fmicb.2024.1479446
Перевод статьи «Effects of dietary probiotic (Clostridium butyricumI9,C. butyricumG15, orParaclostridium bifermentansX13) on growth, digestive enzyme activities, immunity, and intestinal microbiota of Pacific white shrimp (Penaeus vannamei)» авторов Yang W, Liang H, Chen R, Du Z, Deng T, Zheng Y, Song Y, Duan Y, Lin J, Bakky MAH, Tran NT, Zhang M and Li S., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык
Фото: wikipedia

















































































































Комментарии (0)