Как полезные микробы помогают озимой ржи выжить зимой и дать хороший урожай
Полезный эффект от применения микробного консорциума на растения сильно зависит от почвенных условий, которые, в свою очередь, определяются агротехническими приёмами. Закрепление микробных инокулянтов в ризосфере является обязательным условием для успешного взаимодействия между растениями и микроорганизмами. В данном исследовании изучалось, закрепляется ли консорциум полезных микроорганизмов в ризосфере озимой культуры в течение вегетационного периода, включая зимний сезон. Кроме того, мы стремились глубже понять его влияние на продуктивность растений в условиях различных систем земледелия.
Аннотация
Растения озимой ржи, выращенные в ходе долгосрочного полевого эксперимента в условиях традиционного или органического земледелия, были инокулированы осенью после появления всходов микробным консорциумом, содержащим Pseudomonas sp. (RU47), Bacillus atrophaeus (ABi03) и Trichoderma harzianum (OMG16). Плотность микробных инокулянтов в ризосфере и прикорневой почве оценивали количественно осенью и следующей весной. Кроме того, с помощью междисциплинарного подхода было исследовано влияние консорциума на продуктивность растений и формирование бактериальных сообществ в ризосфере.
Высев на селективные среды показал высокую плотность колонизации ризосферы и прикорневой почвы озимой ржи отдельными микроорганизмами из состава консорциума на протяжении всего раннего этапа жизненного цикла растений. Секвенирование ампликонов гена 16S рРНК показало, что система земледелия оказывала влияние преимущественно на бактериальные сообщества ризосферы как осенью, так и весной. Однако инокуляция микробным консорциумом также изменяла состав бактериальных сообществ в каждый момент отбора проб, особенно в начале нового вегетационного сезона весной.
Инокуляция озимой ржи микробным консорциумом значительно улучшала питательный статус растений и их продуктивность, особенно в условиях органического земледелия. Резюмируя, можно сказать, что микробный консорциум, внесённый осенью, показал достаточную эффективность на протяжении всего периода вегетационного покоя и способствовал улучшению продуктивности растений, что указывает на потенциальную применимость микробных решений в органическом сельском хозяйстве, где доступность питательных веществ ограничена.
Введение
Почвенные микроорганизмы играют центральную роль практически во всех почвенных процессах, поскольку они не только участвуют в почвообразовании (Watteau и Villemin, 2018), но и оказывают значительное влияние на важные экосистемные функции, такие как круговорот питательных веществ и продуктивность растений (Latz и др., 2016; Lori и др., 2017). Поэтому управление почвенным микробным сообществом имеет важное значение для поддержания здоровья и продуктивности почвы. Микробное разнообразие положительно связано с устойчивостью микробных сообществ и плодородием почвы (van Elsas и др., 2012; Usero и др., 2021). Сельскохозяйственные культуры получают выгоду от разнообразных почвенных микробных сообществ, поскольку они рекрутируют и обогащают полезные микроорганизмы (БМ) с помощью корневых экссудатов в ризосфере (Berg и Smalla, 2009; Liu и др., 2021). БМ могут стимулировать рост растений через различные механизмы, такие как модуляция уровней фитогормонов (Zubair и др., 2019) и повышение доступности питательных веществ (Jacoby и др., 2017). Кроме того, полезные ризосферные микроорганизмы могут запускать индуцированную системную устойчивость и защищать растение от почвенных и надземных патогенов (Wei и др., 2020; Vlot и др., 2021). Действительно, растительные патогены могут контролироваться БМ либо напрямую, за счет продукции литических ферментов и биоактивных соединений (Fira и др., 2018; Ali и др., 2020), либо косвенно, за счет конкуренции за ресурсы в одной и той же экологической нише, такой как ризосфера (Rai и др., 2016).
Инокуляция микроорганизмов с известными полезными для растений функциями представляет собой практический подход, способный модулировать состав аборигенных микробных сообществ, особенно в ризосфере (Deng и др., 2019; Berg и др., 2021). Хотя многие БМ показывают многообещающие функции в отношении роста и защиты растений в контролируемых условиях in vitro и in vivo (Mendes и др., 2013; Mazzola и Freilich, 2017), их эффективность в полевых условиях часто ограничена из-за недостаточной колонизации ризосферы хозяина и/или неблагоприятных условий (Parnell и др., 2016; Batista и Singh, 2021). Для успешной колонизации ризосферы инокулированные БМ должны преодолеть барьер аборигенного почвенного микробного сообщества, адаптироваться к меняющимся условиям окружающей среды и взаимодействовать с растением-хозяином, используя главным образом корневые экссудаты в качестве хемоаттрактанта и субстрата (Finkel и др., 2017; Berg и др., 2021). Различные стадии развития растения и меняющиеся условия окружающей среды изменяют характер корневых экссудатов и, соответственно, состав микробного сообщества в ризосфере (Chaparro и др., 2014; Windisch и др., 2021), что может влиять на ризосферную компетентность БМ. Таким образом, даже если инокулированные БМ успешно закрепляются в ризосфере на ранних стадиях развития растения, их сохранение в течение длительного периода не гарантируется. Инокуляция многоштаммовыми консорциумами, содержащими два или более БМ, показала повышенную эффективность по сравнению с отдельными видами БМ (Sun и др., 2022). Консорциумы БМ, включающие членов с разнообразными функциями, могут занимать разные экологические ниши, что делает их более устойчивыми к изменчивым условиям среды (Bradáčová и др., 2019; Tosi и др., 2020). Кроме того, взаимодействие между членами консорциума повышает их экологическую приспособленность, а взаимодополняющие полезные функции создают синергию, способствующую их взаимодействию с растением-хозяином (Moradtalab и др., 2020; Pascale и др., 2020).
Различные агротехнические практики, такие как традиционное и органическое земледелие, сильно влияют на структуру микробных сообществ (Francioli и др., 2016; Windisch и др., 2021). Chowdhury и др. (2019) показали, что традиционное земледелие с использованием синтетических пестицидов и минеральных удобрений по-разному формирует состав ризосферных микробных сообществ по сравнению с органическим земледелием, что было связано с различным состоянием здоровья растений. В настоящее время существует ограниченное понимание того, как агротехническая практика влияет на эффективность консорциума полезных микроорганизмов (БМк). Например, растения могут получать больше выгоды от повышенной биодоступности питательных веществ благодаря БМк в бедных питательными веществами почвах (Eltlbany и др., 2019), тогда как в почвах с высоким уровнем питательных веществ ризосферная колонизация инокулированным БМк может быть снижена (Lopes и др., 2021).
Основной целью данного исследования была оценка ризосферной компетентности инокулированных членов БМк у озимой ржи (Secale cereale cv. Conduct) в течение вегетационного периода в зависимости от различных агротехнических практик. Озимые зерновые культуры подвергаются экстремальным изменениям окружающей среды в течение вегетационного периода, что ставит вопрос о том, могут ли члены БМк, инокулированные осенью, закрепиться в ризосфере озимой ржи и поддерживать высокую плотность колонизации в течение зимнего периода. Кроме того, влияние применяемого БМк на продуктивность растений может определяться физико-химическими свойствами почвы и зависящими от агротехнической практики почвенными микробными сообществами. В этом контексте мы выдвинули гипотезу о том, что (i) ранняя инокуляция озимой ржи обеспечивает достаточную колонизацию каждого члена БМк на ранней стадии развития растения, поддерживая его сохранение в ризосфере на протяжении всего вегетационного периода; (ii) применение БМк формирует состав ризосферного бактериального сообщества в зависимости от агротехнической практики и, таким образом, по-разному влияет на продуктивность растений.
Для проверки наших гипотез мы сначала охарактеризовали in vitro признаки, способствующие росту растений, и способность каждого члена БМк ингибировать рост почвенных патогенов. Затем мы использовали долгосрочный полевой эксперимент для оценки ризосферной компетентности и эффективности тестируемого БМк на продуктивность растений в полевых условиях при различных агротехнических практиках (органическое и традиционное земледелие). Растения озимой ржи инокулировали вскоре после появления всходов, а колонизацию БМк и продуктивность растений оценивали осенью и весной. Динамику бактериального сообщества на протяжении вегетационного покоя характеризовали с помощью секвенирования ампликонов гена 16S рРНК.
Материалы и методы
Микробный консорциум, использованный в исследовании
Микробные члены консорциума [Pseudomonas sp. (RU47; коллекция штаммов Института Юлиуса Кюна, Брауншвейг, Германия), Bacillus atrophaeus (ABi03; штамм предоставлен ABiTEP, Берлин, Германия) и Trichoderma harzianum (OMG16; коллекция штаммов Университета прикладных наук Анхальт, Бернбург, Германия)] были отобраны, поскольку каждый штамм показал полезные эффекты на продуктивность растений в предыдущих экспериментах и испытаниях (Schreiter и др., 2014; Mpanga и др., 2018; Schreiter и др., 2018; Mpanga и др., 2019; Moradtalab и др., 2020; Hafiz и др., 2022).
Характеристика членов консорциума in vitro
Для более полной характеристики признаков, полезных для растений, связанных с каждым членом консорциума (Pseudomonas sp. RU47, B. atrophaeus ABi03 и T. harzianum OMG16), была проведена серия тестов in vitro для оценки их антагонистических способностей и способности к стимуляции роста растений. Для каждого штамма были протестированы модуляция гормонов роста растений за счет продукции индол-3-уксусной кислоты (ИУК) и активности 1-аминоциклопропан-1-карбоксилат (АЦК) деаминазы (Koo и др., 2010), а также повышение доступности питательных веществ за счет продукции сидерофоров (Schwyn и Neilands, 1987). Кроме того, для отдельных штаммов был проанализирован растворимость калиевого полевого шпата (KAlSi3O8) (Breitkreuz и др., 2021), фосфата кальция (Ca3(PO4)2) (Nautiyal, 1999), оксида цинка (ZnO) (Mumtaz и др., 2017), диоксида марганца (MnO2) (Sanket и др., 2017) и продукция аммиака (Cappuccino и Sherman, 2014). Для анализов солюбилизации питательных веществ среда была модифицирована для OMG16 с использованием агара Ваксмана (Huber и др., 1987).
Образование целлюлазы (Berg и др., 2005), хитиназы (Berg и др., 2001), β-1,3-глюканазы (Weinert и др., 2010), протеазы (Berg и др., 2005) и цианистого водорода (HCN) (Donate-Correa и др., 2005) было протестировано в качестве антагонистических функций. Тесты in vitro на секрецию ферментов с антагонистическими функциями не были выполнимы для OMG16 из-за несовместимости культивирования. Модифицированный метод двойной культуры по Lin и Ho (2021) был использован для оценки способности каждого штамма ингибировать рост патогенов in vitro. Для двух бактериальных штаммов RU47 и ABi03 20 мкл бактериальной суспензии (105 колониеобразующих единиц (КОЕ) мл⁻¹) наносили по центральной линии на чашку с питательным агаром I (Sifin diagnostics, Берлин, Германия). Свежевыращенные диски мицелия (ø 6 мм) Fusarium culmorum (изолят F247), F. graminearum (изолят FG66) и Rhizoctonia solani изолят AG3 (изолят Ben3) помещали на расстоянии 2,5 см по обе стороны от линии. Для грибного штамма OMG16 предварительно выращенные диски мицелия OMG16 и патогена помещали на противоположные стороны чашки с картофельно-декстрозным агаром (PDA, Carl Roth, Карлсруэ, Германия). Анализы двойной культуры инкубировали в темноте при 25°C в течение 10 дней. Антагонистические свойства, т.е. образование зоны ингибирования, отдельных штаммов БМк проверяли каждые два дня (Дополнительный рисунок 1).
Описание полевого участка
Эксперимент проводился с 2020 по 2021 год на экспериментальном поле с делянками в условиях традиционного и органического земледелия в рамках долгосрочного полевого испытания в Тюрове (Thy_ABS; 52°15' с.ш., 13°14' в.д., 44 м над уровнем моря), которое было заложено в 2005 году на Сельскохозяйственном исследовательском институте Университета им. Гумбольдта в Берлине. Палево-бурая земля (лювисоли) из песка над глубоким суглинком с низким содержанием гумуса является преобладающим типом почвы (83% песка, 14% ила и только 3% глины) на этом участке. Среднегодовая температура (1981-2020) составляет 9,2°C со среднегодовым количеством осадков 509,8 мм.
В традиционном земледелии севооборот включал озимый рапс (Brassica napus ssp. napus), озимую пшеницу (Triticum aestivum) и озимую рожь (Secale cereale), а в органическом земледелии — люцерну (Medicago sativa), люцерну, картофель (Solanum tuberosum), тритикале (Triticale), горох полевой (Pisum sativum ssp. arvense), яровой ячмень (Hordeum vulgare) и озимую рожь (Secale cereale). В традиционной практике земледелия использовалось минеральное удобрение в соответствии с сельскохозяйственными стандартами региона (N = 120 кг га⁻¹, P = 21 кг га⁻¹, K = 120 кг га⁻¹, Mg = 12 кг га⁻¹, S = 14 кг га⁻¹). Азот вносили в виде кальциево-аммиачной селитры (КАС), а калий, магний и серу — в равных соотношениях Patentkali® (K + S Minerals and Agriculture, Кассель, Германия) и Korn-Kali® (K + S Minerals and Agriculture). Для фосфорного удобрения использовали тройной суперфосфат (46% P2O5). Перед выращиванием озимой ржи и тритикале в органическом земледелии в почву вносили крупный рогатый скот (15 000 кг га⁻¹) с добавлением люцерны и гороха полевого. Защита растений проводилась в соответствии с сельскохозяйственными стандартами традиционного возделывания [гербициды: 2,0 л га⁻¹ Picona (BASF, Людвигсхафен, Германия), 0,25 л га⁻¹ Cadou SC (Bayer, Леверкузен, Германия); фунгициды: 2,0 л га⁻¹ Ceriax (BASF)]. В органическом земледелии пестициды не использовались, а появляющиеся сорняки удаляли с помощью механического культиватора. Перед посевом озимой ржи (норма высева: 200 семян на м² в традиционном земледелии и 300 семян на м² в органическом земледелии) проводилась обработка почвы, включая лущение стерни дисковой бороной, вспашку на глубину около 20-25 см и подготовку посевного ложа с помощью возвратно-поступательной бороны.
Приготовление микробного консорциума и его полевое применение
Штамм T. harzianum OMG16 культивировали на среде PDA (Carl Roth), дополненной 100 мг L⁻¹ пенициллина (Carl Roth), 50 мг L⁻¹ стрептомицина сульфата (Sigma-Aldrich, Сент-Луис, США) и 10 мг L⁻¹ тетрациклина гидрохлорида (AppliChem, Дармштадт, Германия). Для приготовления полевого инокулюма зерна проса (Panicum miliaceum) замачивали на ночь в водопроводной воде. Затем зерна тщательно промывали холодной водопроводной водой. Чистые зерна (по 500 г на мешок) переносили в мешки, предназначенные для культивирования грибов (sun bag; Sigma-Aldrich), и автоклавировали в течение трех последующих дней. Диск со свежевыращенным мицелием OMG16 добавляли к зернам и инкубировали в течение 30 дней в темноте при комнатной температуре. После того как OMG16 полностью колонизировал зерна проса, инокулюм лиофилизировали и измельчали. Непосредственно перед посевом озимой ржи порошок (100 мг м⁻²) вносили в почву на глубину 20 см. Лиофилизированный порошок зерна проса без OMG16 использовали в качестве контроля.
Штаммы B. atrophaeus ABi03 и Pseudomonas sp. RU47 использовали в виде спонтанных рифампицин-устойчивых мутантов, предоставленных в виде готового инокулюма компанией ABiTEP (Schreiter и др., 2018). Через две недели после появления всходов озимой ржи каждую делянку поливали 4 л бактериальной суспензии ABi03 и RU47, смешанной с водопроводной водой (по 7,5 x 10⁷ КОЕ мл⁻¹). Контрольные делянки поливали водопроводной водой. Каждая обработка включала четыре повторности (1,5 x 2 м), расположенные в рандомизированном блоковом дизайне для обеих агротехнических практик.
Отбор образцов и проверка ризосферной компетентности
Для отбора образцов учитывались только растения из внутренней части делянки (0,5 x 1 м) каждой повторности, отбор проводился осенью через семь недель после инокуляции (WPI) на стадии EC 21 и весной следующего года (22 WPI) на стадии EC 25-29. Всего отбирали 20 растений озимой ржи на делянку и обрабатывали как один образец на повторность обработки. После извлечения растений побеги отделяли от корней. Для получения прикорневой почвы отряхивали почвенную фракцию, слабо прилегающую к корневой системе. Корни, побеги и прикорневую почву немедленно хранили при 4°C. После определения сырой массы побегов побеги высушивали лиофилизацией и измеряли сухую массу побегов (СМП).
Корни с оставшейся плотно прилегающей почвой промывали стерильной водопроводной водой и нарезали на кусочки по 1 см для отбора ризосферных образцов. После объединения фрагментов корней пять граммов корневого материала переносили в стерильный мешок для гомогенизатора, содержащий 15 мл стерильного 0,3% NaCl. Ризосферные образцы получали с помощью гомогенизатора Stomacher 400 Circulator (Seward Ltd., Уортинг, Великобритания) в течение 60 с при 300 об/мин. Супернатант переносили в пробирку Falcon, а этапы гомогенизации повторяли дважды. Объединенный супернатант использовали для определения количества бактериальных членов БМк путем немедленного высева серийных разведений на чашки с питательным агаром I (Sifin diagnostics), содержащим 75 мкг мл⁻¹ рифампицина (Serva Electrophoresis, Гейдельберг, Германия) и 100 мкг мл⁻¹ циклогексимида (Serva Electrophoresis), и инкубировали в темноте в течение 48 ч при 28°C. Учитывая различную морфологию колоний ABi03 и RU47, подсчитывали КОЕ и рассчитывали КОЕ на грамм сухой массы корней, использованных для гомогенизации. Оставшийся супернатант гомогенизатора далее обрабатывали, как описано Schreiter и др. (2014), для анализа бактериального ризосферного сообщества на основе экстракции тотальной ДНК сообщества.
Прикорневую почву использовали для определения количества OMG16. Для этого 5 г прикорневой почвы экстрагировали в общем объеме 50 мл NaCl (0,3%) путем встряхивания в течение 30 мин при комнатной температуре на орбитальном шейкере (HSM-10; Hettich, Тутлинген, Германия) при 70 об/мин. Затем супернатант немедленно высевали на селективную среду для триходермы, как предложено Williams и др. (2003), содержащую 250 мг L⁻¹ хлорамфеникола (Carl Roth), 90 мг L⁻¹ стрептомицина (Sigma-Aldrich), 200 мг L⁻¹ квинтозена (Sigma-Aldrich), 926 мг L⁻¹ пропамокарба (ProPlant; Arysta LifeScience, Париж, Франция) и 150 мг L⁻¹ розового бенгала (AppliChem). После инкубации OMG16 при 28°C в темноте в течение 10 дней количественно определяли КОЕ на грамм сухой массы почвы. Сухую массу почвы получали путем высушивания пяти граммов свежей почвы при 110°C до достижения постоянной массы.
Колосья озимой ржи со всей делянки отбирали при полной спелости. После подсчета колосьев зерно обмолачивали и определяли массу. Урожайность с гектара экстраполировали для лучшего сравнения с литературными данными.
Анализ питательных веществ
Анализ питательных веществ в растительном материале проводили в соответствии с сертифицированными протоколами Ассоциации немецких сельскохозяйственных аналитических и исследовательских институтов, VDLUFA, Германия (Verband Deutscher Landwirtschaftlicher Untersuchungs- und Forschungsanstalten, 2011). 200-500 мг сухого растительного материала растворяли в 5 мл HNO3 (65%) и 3 мл H2O2 (30%) с помощью микроволновой минерализации при 210°C в течение 25 мин. Экстракт фильтровали и доводили до конечного объема 100 мл. Концентрации K, P, Mg, S, Ca, Mn, Cu и Zn определяли с помощью масс-спектрометрии с индуктивно связанной плазмой (ICP-OES, Thermo Fisher Scientific, Драйайх, Германия); общее содержание C и N определяли с помощью элементного анализатора (Elementary Vario El cube; Лангензельбольд, Германия). Содержание макро- и микроэлементов в прикорневой почве анализировали в соответствии с сертифицированными протоколами VDLUFA компанией AGROLAB (Лайнефельде-Ворбис, Германия).
Экстракция ДНК и высокопроизводительное секвенирование ампликонов гена 16S рРНК
Тотальную ДНК сообщества экстрагировали из суммарных ризосферных осадков с использованием системы бисерного гомогенизатора FastPrep-24 и набора FastDNA Spin Kit for Soil (MP Biomedicals, Санта-Ана, США) в соответствии с рекомендациями производителя. Образцы очищали с помощью набора GeneClean Spin Kit (MP Biomedicals). Создание библиотек и секвенирование гена 16S рРНК было выполнено компанией Novogene (Кембридж, Великобритания) на платформе NovaSeq PE250 с использованием праймеров Uni341F (5'-CCTAYGGGRBGCASCAG-3') и Uni806R (5'- GGACTACNNGGGTATCTAAT-3') (Sundberg и др., 2013). Праймеры и адаптеры удаляли с помощью программного обеспечения Cutadapt (Martin, 2011). Парноконцевые прочтения обрабатывали с использованием конвейера DADA2 (Callahan и др., 2016). Для таксономической идентификации полученных вариантов последовательностей ампликонов (ASV) репрезентативные последовательности классифицировали до максимально низкого таксономического уровня в рабочем процессе Galaxy (Cock и др., 2013) с порогом e-value 0,001 и порогом идентичности процентов 80% по базе данных SILVA 138.1 SSU Ref NR99 (v.138.1, Quast и др., 2012; v138.1). Последовательности, идентифицированные как пластиды (хлоропласты или митохондрии), и последовательности с менее чем пятью прочтениями были удалены. В нашем наборе данных не было ASV, классифицированных как архейные таксоны. Кривые разрежения были построены для оценки покрытия прочтений, и глубина секвенирования достаточная покрывала разнообразие каждого образца. Данные ампликонных последовательностей депонированы в Sequence Read Archive (https://www.ncbi.nlm.nih.gov/sra) под номером BioProject PRJNA975889.
Анализ данных
Статистический анализ роста озимой ржи, урожайности, концентрации питательных веществ и обилия БМк выполняли с использованием R (v.4.2.2, R Core Team, 2022). Основные эффекты и эффекты взаимодействия между долгосрочными агротехническими практиками и использованием БМк анализировали с помощью двухфакторного дисперсионного анализа (two-way ANOVA) для СМП и питательных веществ побегов. Различия в обилии БМк в ризосфере и прикорневой почве проверяли с помощью двухфакторного дисперсионного анализа, анализируя основные эффекты и эффекты взаимодействия времени отбора проб и различных долгосрочных агротехнических практик. Все анализы визуально проверяли на гомоскедастичность и нормальное распределение остатков с использованием теста Дурбина-Уотсона из пакета "car" (v.3.1.1, Fox и Weisberg, 2019) и нормальных Q-Q графиков. Если предположения дисперсионного анализа не выполнялись, данные трансформировали с использованием пакета R "rcompanion" (v.2.4.26, Mangiafico, 2023) в соответствии с подходом лестницы степеней Тьюки. Статистическую оценку проводили с использованием алгоритма теста Сидака и попарного сравнения в сочетании с компактным отображением букв из пакетов "emmeans" (v.1.8.3, Lenth, 2022), "multcomp" (v.1.4, Hothorn и др., 2008) и "multcompView" (v.0.1.8, Graves и др., 2019). Данные визуализировали с использованием пакетов "ggplot2" (v.3.4.1, Wickham, 2016) и "ggpattern" (v.1.0.1, FC и Davis, 2022). Сравнения с p-значениями ниже 0,05 считали статистически значимыми.
Вся обработка данных и статистические анализы для данных ASV выполнялись с использованием пакетов R "tidyverse" (v.1.3.1, Wickham и др., 2019), "ggplot2", "dplyr" (v.1.1.1, Wickham и др., 2023), "tibble" (v.3.1.8, Müller и Wickham, 2022), "reshape2" (v.1.4.4, Wickham, 2007), "vegan" (v.2.6.1, Oksanen и др., 2022) и "phyloseq" (v.3.16, McMurdie и Holmes, 2013). Для учета неравномерной глубины секвенирования мы многократно выполняли 1000 разрежений до наименьшего числа последовательностей в наборе данных (49 206). Средние обилия ASV рассчитывали на основе 1000 разрежений. Эта процедура позволяет представить все наблюдаемые последовательности в равной пропорциональности и учитывает случайную вариацию, вносимую разрежением (Cameron и др., 2021). Расстояние Брея-Кертиса использовали для оценки β-разнообразия путем нормализации разреженных наборов данных к относительной обильности (%) и применения log10-трансформации с добавлением псевдосчета. PERMANOVA-тесты применяли для оценки того, как агротехническая практика и инокуляция БМк влияли на состав бактериального сообщества. Попарный PERMANOVA-тест с коррекцией Бенджамини-Хохберга выполняли после объединения двух обработок в четыре группы с использованием пакета "pairwiseadonis" (v.0.4, Martinez, 2022). Значительные различия в α-разнообразии (богатство ASV, индекс Симпсона, индекс Шеннона и выравненность) оценивали с помощью двухфакторных дисперсионных тестов. Если данные не соответствовали критериям нормальности на основе теста Шапиро, выполняли непараметрический дисперсионный анализ с выравниванием рангов (пакет "ARTool", v.0.11.1, Kay и др., 2021).
Для исследования влияния типа агротехнической практики и инокуляции БМк на обильность ASV выполняли ANCOM-BC2 (v2.1.2, Lin и Peddada, 2020) в качестве теста на дифференциальную обильность с коррекцией p-значений методом Бенджамини-Хохберга. После теста на дифференциальную обильность мы выполнили множественные логистические регрессии с коррекцией Бенджамини-Хохберга в качестве пост-тестов. Логистическая регрессия была выбрана из-за ее прогностической силы, аналогичной алгоритмам машинного обучения (например, случайный лес), и ее внутренней интерпретируемости (Topçuoğlu и др., 2020). Мы использовали уравнение логистической регрессии для прогнозирования вероятности возникновения обработки (т.е. Organic-BMc, Organic-Control, Conventional-BMc или Conventional-Control) на основе относительной обильности ASV. Следовательно, тесты логистической регрессии позволили нам оценить, реагировали ли дифференциально обильные ASV на конкретную обработку (ASV-респондеры). Для каждой конкретной обработки положительный или отрицательный коэффициент модели указывал на более высокое или более низкое относительное обилье соответственно. ASV были назначены как положительные или отрицательные респондеры для каждой обработки на основе коэффициента модели.
Для оценки того, была ли биомасса ржи связана с более высокими долями инокулированных бактериальных штаммов по сравнению с остальными членами бактериального сообщества в различных обработках, последовательности ASV-респондеров, классифицированные как Pseudomonas spp. или Bacillus spp., были выровнены по созданным здесь базам данных. Для Pseudomonas spp. мы создали базу данных, содержащую последовательности гена 16S рРНК из шести копий гена 16S рРНК инокулянта RU47 (Kuzmanović и др., 2018). Для штамма ABi03 мы выполнили клонирование и секвенирование полной области 16S рРНК с использованием праймеров U8-27 и R1494-1514 (Heuer и др., 2009), где мы идентифицировали шесть различных генов 16S рРНК. Кроме того, мы извлекли последовательности Pseudomonas spp. или Bacillus spp. из базы данных GenBank 16S рРНК, ограниченной коллекцией типовых штаммов (Federhen, 2015). Область V3-V4 гена 16S рРНК, используемую для секвенирования ампликонов Illumina, извлекали из последовательностей базы данных с использованием программного обеспечения Cutadapt (Martin, 2011). Попарное выравнивание выполняли для Bacillus spp. и Pseudomonas spp., используя ASV и созданные базы данных с пакетом "msa" (v.1.30.1, Bodenhofer и др., 2015). Матрицу расстояний выравнивания рассчитывали с помощью пакета "seqinr" (v.4.2.23, Charif и Lobry, 2007). Филогенетическое расстояние рассчитывали с помощью алгоритма присоединения соседей (пакет "ape", v.5.7, Paradis и Schliep, 2019). Филогенетическое расстояние визуализировали как анализ главных координат.
Результаты
Признаки членов микробного консорциума in vitro
In vitro отдельные члены использованного микробного консорциума были протестированы на антагонистические и стимулирующие рост функции (Таблица 1). ABi03 показал активность целлюлазы, хитиназы, β-1,3-глюканазы и протеазы, но не продуцировал HCN. В анализах двойной культуры ABi03 подавлял рост всех тестируемых патогенов, ингибируя рост мицелия (Дополнительный рисунок 1). Несмотря на неспособность RU47 ингибировать рост F. culmorum или F. graminearum, примечательно, что колонии RU47 не были переращены этими патогенами. Кроме того, мы подтвердили незначительное ингибирование роста R. solani RU47, а также продукцию протеазы и HCN. OMG16 также подавлял рост F. culmorum и F. graminearum, распространяясь быстрее, чем патоген на PDA. Литической деградации мицелия в пограничной зоне между OMG16 и тестируемыми патогенами не наблюдалось (Дополнительный рисунок 1).
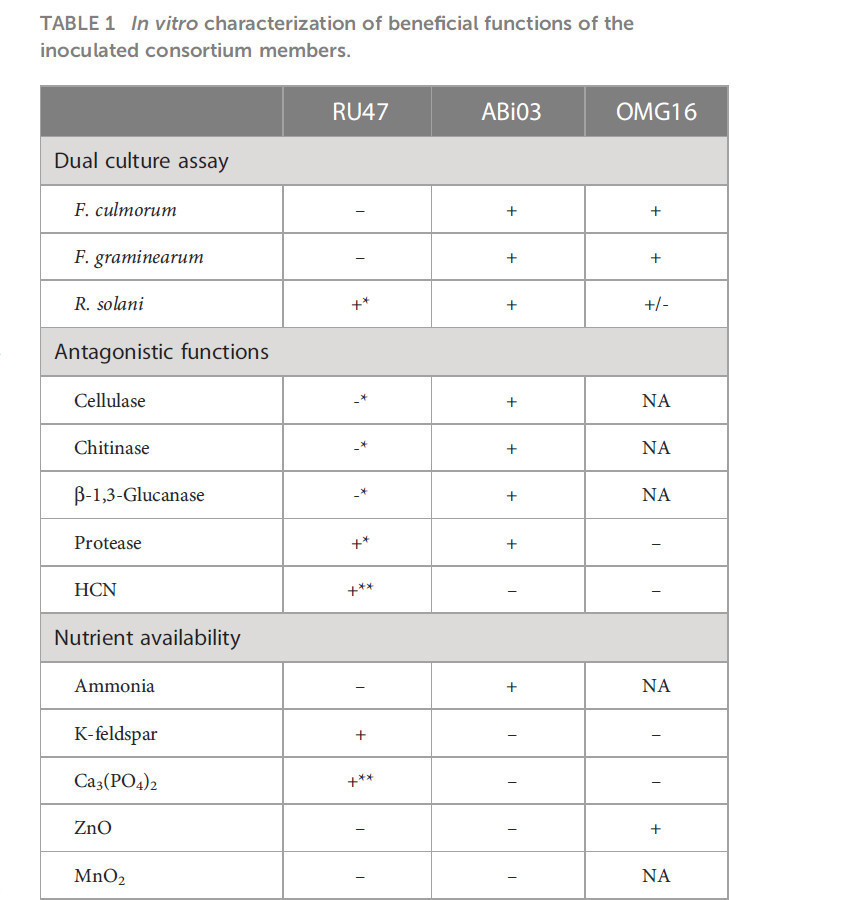
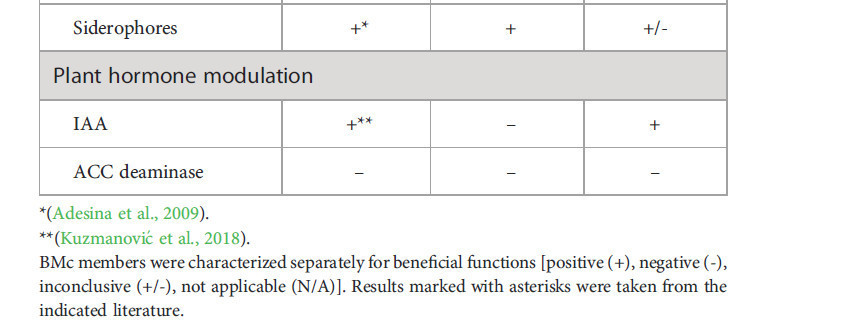
Таблица 1 Характеристика полезных функций инокулированных членов консорциума in vitro.
RU47 растворял калиевый полевой шпат и Ca3(PO4)2 in vitro в анализах доступности питательных веществ и показал продукцию сидерофоров, а также ИУК (Таблица 1). Для ABi03 были обнаружены аммиак и сидерофоры, но не наблюдалось увеличения растворимости тестируемых питательных веществ или модуляции гормонов растений. Кроме того, для OMG16 были показаны продукция ИУК, низкая продукция сидерофоров и растворимость ZnO.
Плотность членов консорциума в ризосфере и почве
Для оценки сохранения отдельных БМ в ризосфере и прикорневой почве озимой ржи в полевых условиях мы оценивали их плотность после инокуляции с помощью двух последующих отборов проб осенью и весной следующего года. Оба бактериальных штамма ABi03 и RU47 достаточно колонизировали ризосферу озимой ржи с плотностью более шести Log10 КОЕ на грамм сухой массы корней через семь недель после инокуляции при обеих агротехнических практиках (Рисунок 1). Грибной штамм OMG16 также показывал достаточную плотность в прикорневой почве — более пяти Log10 КОЕ на грамм сухой массы почвы. Различий в плотности отдельных БМ в соответствующей среде обитания в зависимости от агротехнической практики не наблюдалось. При втором отборе проб весной (22 недели после инокуляции) не было обнаружено изменений в плотности ABi03 в ризосфере озимой ржи независимо от агротехнической практики (Рисунок 1, Дополнительная таблица 1). Плотность RU47 снизилась примерно до пяти Log10 КОЕ на грамм сухой массы корней при обеих агротехнических практиках по сравнению с первым отбором осенью. Растения, выращенные в условиях органического земледелия, показали значительно более низкую плотность OMG16 в прикорневой почве по сравнению с первым отбором осенью. Две различные агротехнические практики не оказывали существенного влияния на способность отдельных БМ закрепляться в ризосфере ржи или прикорневой почве в оба срока отбора.
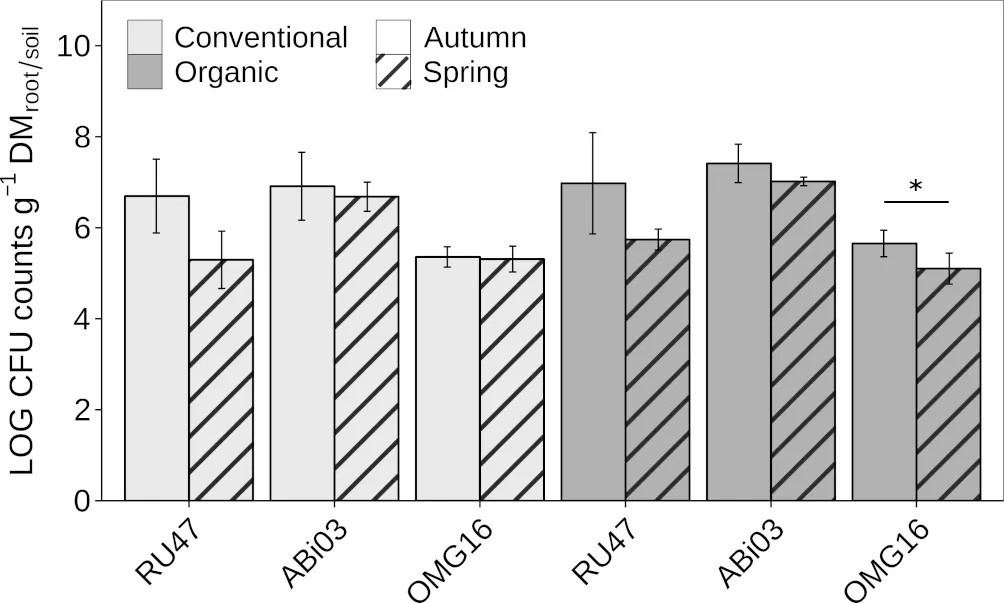
Рисунок 1 Обильность полезных микроорганизмов у озимой ржи (cv. Conduct), возделываемой при различных долгосрочных агротехнических практиках (традиционное и органическое земледелие), осенью и весной того же вегетационного периода. Столбцы представляют средние значения ± стандартное отклонение четырех повторностей. Двухфакторный дисперсионный анализ был проведен для проверки влияния агротехнической практики и времени отбора проб на ризосферную компетентность каждого члена консорциума (Дополнительная таблица 1). Звездочка указывает на значительное различие КОЕ между сроками отбора для каждого члена консорциума отдельно (∗p ≤ 0,05).
Продуктивность растений
Средняя СМП растений озимой ржи при первом отборе (EC 21) осенью составляла от 0,07 до 0,10 г на растение, тогда как средняя СМП при втором отборе (EC 25-29) весной была между 0,26 и 0,48 г на растение (Рисунок 2A, Дополнительная таблица 2A). Контрольные растения без инокуляции БМк не показывали значительных различий в СМП в ответ на агротехническую практику (традиционное и органическое) в оба срока отбора. Растения озимой ржи, возделываемые в органическом земледелии и обработанные БМк, показали значительное увеличение СМП по сравнению с растениями без применения БМк осенью (7 недель после инокуляции, +20%) и особенно весной (22 недели после инокуляции, +45%). В отличие от органического земледелия, обработка БМк не влияла на СМП растений, выращенных в условиях традиционного земледелия, в оба срока отбора. Двухфакторный дисперсионный анализ дополнительно выявил значительное взаимодействие между агротехнической практикой и инокуляцией БМк в отношении СМП в оба срока отбора (Дополнительная таблица 2B), что указывает на зависимые от агротехнической практики эффекты БМк на СМП. При уборке урожая не было значительных различий в урожайности между органическим и традиционным земледелием, хотя органически выращенные растения имели немного более низкую массу зерна с гектара (Рисунок 2B). Обработка БМк не оказывала существенного влияния на урожайность ни в традиционном, ни в органическом земледелии. Однако наблюдалась небольшая тенденция к повышению урожайности с единицы площади в органическом земледелии после обработки БМк.
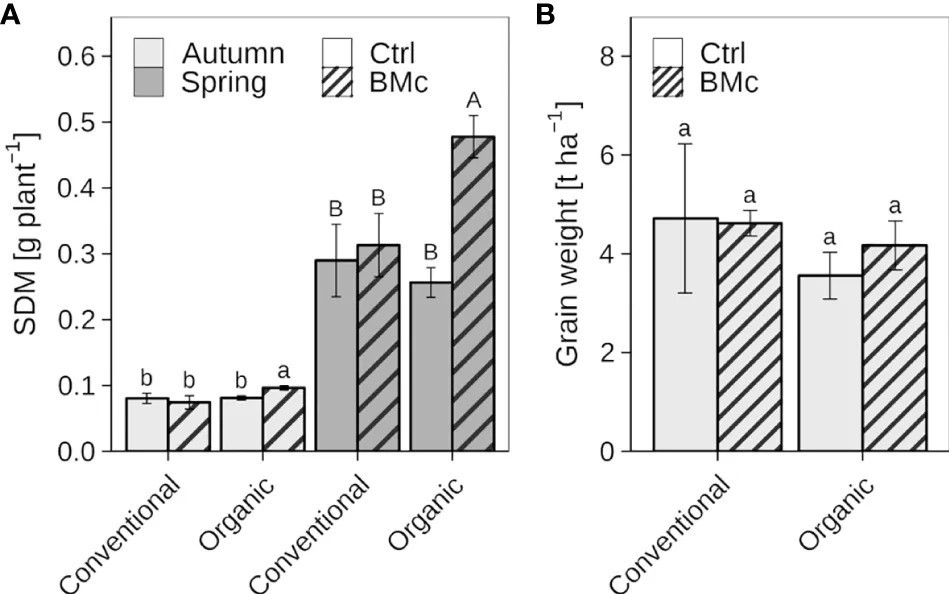
Рисунок 2 Влияние консорциума полезных микроорганизмов (БМк) на сухую массу побегов (СМП) осенью и весной (A) и урожайность (B) озимой ржи (cv. Conduct), возделываемой в условиях традиционного и органического долгосрочного земледелия. Столбцы представляют средние значения ± стандартное отклонение четырех повторностей (Дополнительная таблица 2). Разные буквы указывают на значительные различия, рассчитанные с помощью теста Сидака (p ≤ 0,05). Тип буквы указывает на отдельный статистический анализ (строчные: осень, заглавные: весна).
Для определения того, был ли питательный статус озимой ржи связан с агротехнической практикой и инокуляцией БМк, концентрации нескольких микро- и макроэлементов были измерены в растениях, выращенных на всех исследуемых делянках, в оба срока отбора (Таблица 2). При первом отборе осенью большинство питательных веществ были на одном уровне для обеих агротехнических практик, независимо от инокуляции БМк. Двухфакторный дисперсионный анализ показал общий значительный эффект (p ≤ 0,021) инокуляции БМк на концентрацию азота (Дополнительная таблица 3), которая немного повышалась при инокуляции БМк для обеих агротехнических практик. Однако попарное сравнение не подтвердило значительной разницы между четырьмя обработками. В отличие от осеннего отбора, при втором отборе весной следующего года концентрации почти всех питательных веществ, за исключением углерода (C), меди (Cu), кальция (Ca) и марганца (Mn), были снижены в условиях органического земледелия (Таблица 2). На основании двухфакторного дисперсионного анализа, обработка БМк показала значительный эффект на большинство питательных веществ в обеих агротехнических практиках, за исключением цинка (Zn) (Дополнительная таблица 3). Однако попарное сравнение всех четырех обработок показало, что обработка БМк значительно повышала только концентрацию фосфора (P) в органически выращенных растениях и концентрацию серы (S) в традиционно выращенных растениях по сравнению с контролем (Таблица 2). Свойства почвы не изменялись под влиянием обработки БМк, но в основном определялись агротехнической практикой (Дополнительная таблица 4).
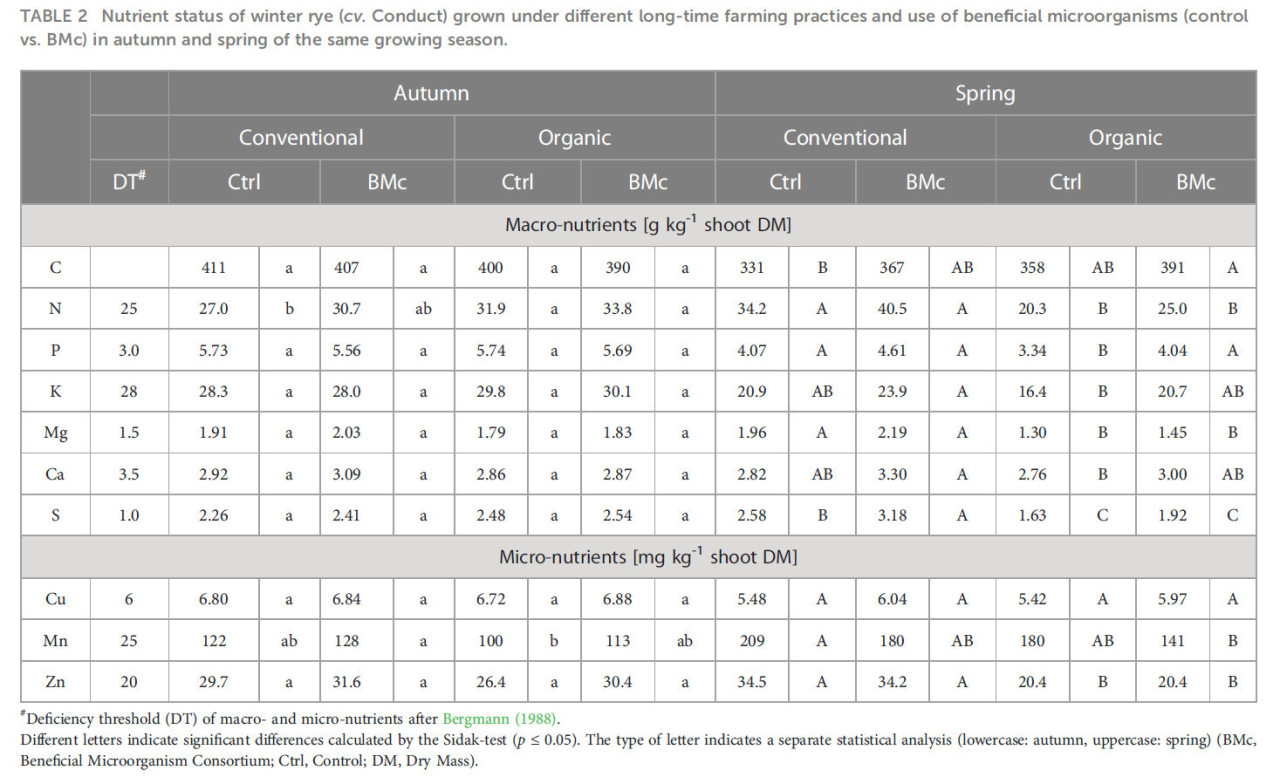
Таблица 2 Питательный статус озимой ржи (cv. Conduct), выращенной при различных долгосрочных агротехнических практиках и использовании полезных микроорганизмов (контроль и БМк) осенью и весной того же вегетационного периода.
Бактериальное сообщество в ризосфере
Для выяснения того, влияли ли выбор агротехнической практики и инокуляция БМк на состав бактериальных сообществ ризосферы озимой ржи, мы выполнили секвенирование ампликонов гена 16S рРНК. Агротехническая практика сильно влияла на состав ризосферных бактериальных сообществ осенью (PERMANOVA, R² = 0,29, p < 0,001, Рисунок 3A) и весной (PERMANOVA, R² = 0,23, p < 0,001, Рисунок 3B). Аналогично, инокуляция БМк значительно влияла на состав бактериального сообщества ризосферы в оба срока отбора (PERMANOVA, R² = 0,10-0,13, p < 0,001, Рисунок 3). Однако мы не обнаружили эффекта взаимодействия между выбором агротехнической практики и инокуляцией БМк (PERMANOVA, R² = 0,04-0,05). Объединение двух переменных и выполнение попарных PERMANOVA-тестов показало, что все комбинации обработок значимо различались при осеннем отборе (Таблица 3). Аналогично, большинство групп значимо различались при весеннем отборе, за исключением комбинации Organic-BMc и Organic-Control (Таблица 3). В то время как и агротехническая практика, и инокуляция БМк влияли на состав бактериального сообщества, они не оказывали последовательного влияния на α-разнообразие. В частности, агротехническая практика значительно влияла на богатство ASV, индекс Шеннона и выравненность весной, но не осенью (Дополнительный рисунок 2, Дополнительная таблица 5). Независимо от времени отбора, инокуляция БМк не влияла на бактериальное α-разнообразие, и эффекта взаимодействия между двумя экспериментальными факторами не обнаружено (Дополнительный рисунок 2).
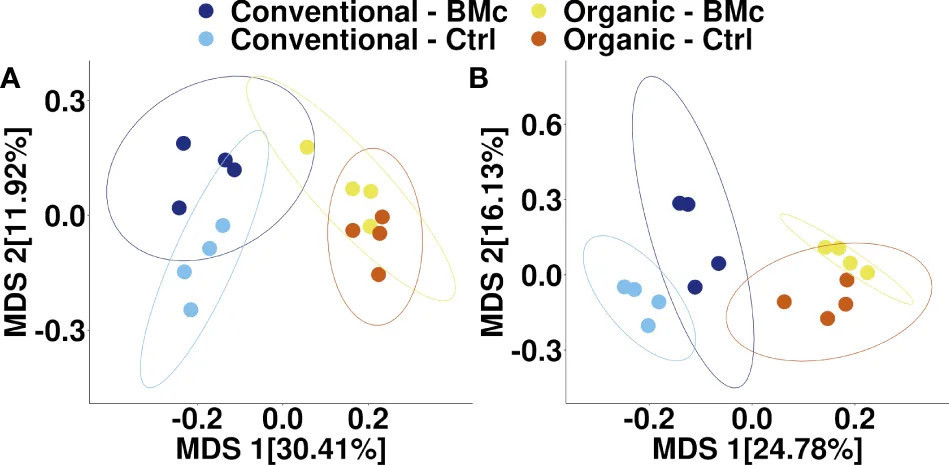
Рисунок 3 Многомерное шкалирование (MDS) состава бактериального сообщества, ассоциированного с ризосферой озимой ржи, при различных агротехнических практиках и обработке инокуляцией БМк, отобранных осенью (A) и весной (B). Эллипсы представляют 95% доверительный интервал. (БМк, Консорциум полезных микроорганизмов; Ctrl, Контроль).
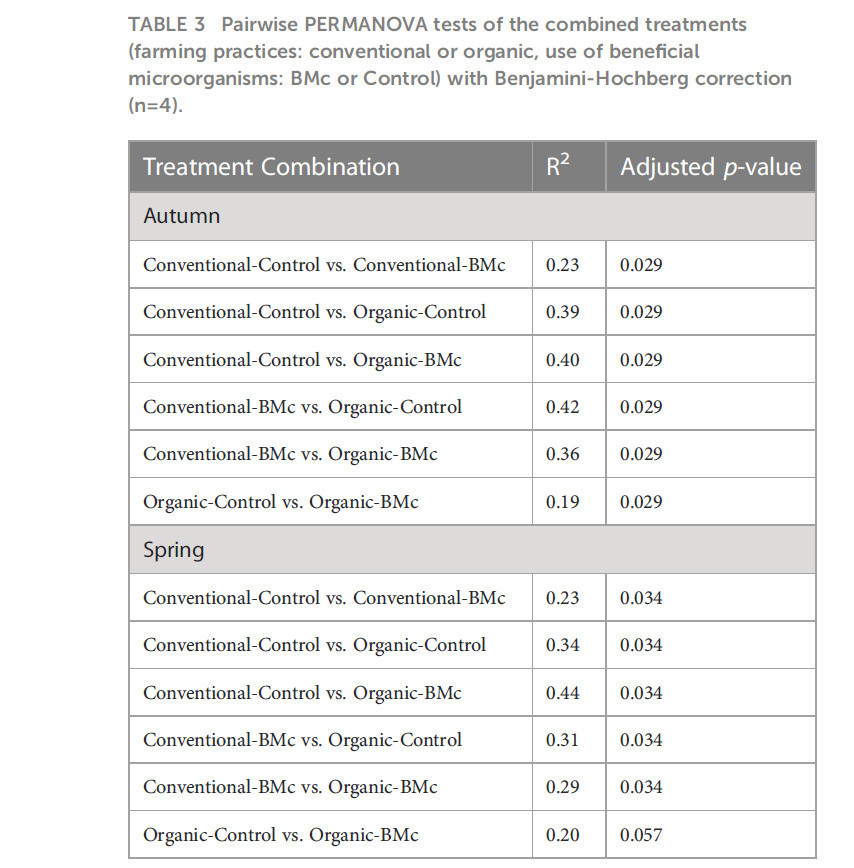
Таблица 3 Попарные PERMANOVA-тесты комбинированных обработок (агротехнические практики: традиционное или органическое, использование полезных микроорганизмов: БМк или Контроль) с коррекцией Бенджамини-Хохберга (n=4).
Мы стремились оценить ASV, на которые влиял либо выбор агротехнической практики, либо инокуляция БМк. В общей сложности 371 ASV осенью и 273 ASV весной значимо различались между двумя агротехническими практиками (Дополнительная таблица 6, ANCOM-BC2). Из этих дифференциально обильных ASV 38 ASV осенью и 16 ASV весной были репрезентативными для исследованных бактериальных сообществ, так как показывали относительную обильность выше 0,5% (Рисунок 4). Доминирующими дискриминантными ASV с наибольшей дифференциальной обильностью в зависимости от агротехнической практики были ASV, таксономически связанные с группой Rhizobium (ASV44), Sphingobacterium spp. (ASV418), Sphingomonas spp. (ASV17, ASV42, ASV53 и ASV69) и Pedobacter spp. (ASV14, ASV40, ASV104, ASV140 и ASV145). Два доминирующих ASV, принадлежащих к Pedobacter spp. (ASV14 и ASV40), и один доминирующий ASV, принадлежащий к Rhizobium spp. (ASV44), различались между агротехническими практиками независимо от времени отбора (Дополнительная таблица 6). В частности, относительная обильность ASV44 (Rhizobium spp.) увеличивалась, в то время как относительная обильность ASV40 (Pedobacter spp.) снижалась при органической агротехнической практике (Рисунок 4). Напротив, ASV14 (Pedobacter spp.) показывал противоположные тенденции между двумя отборами в органическом земледелии: увеличение обильности осенью и снижение весной. Инокуляция БМк изменяла относительную обильность 21 ASV осенью и 14 ASV весной (Дополнительная таблица 6), причем пять ASV осенью и три ASV весной составляли более 0,5% относительной обильности (Рисунок 4). Два ASV, классифицированных как Luteibacter spp. (ASV48 и ASV61), стабильно встречались в более высокой относительной обильности в образцах, инокулированных БМк. Другие ASV, которые увеличивались из-за инокуляции БМк, принадлежали к Mucilaginibacter spp. (ASV37) и Pedobacter spp. (ASV78, ASV73). Кроме того, один доминирующий ASV, принадлежащий к Pedobacter spp. (ASV31), был негативно затронут инокуляцией БМк при весеннем отборе.
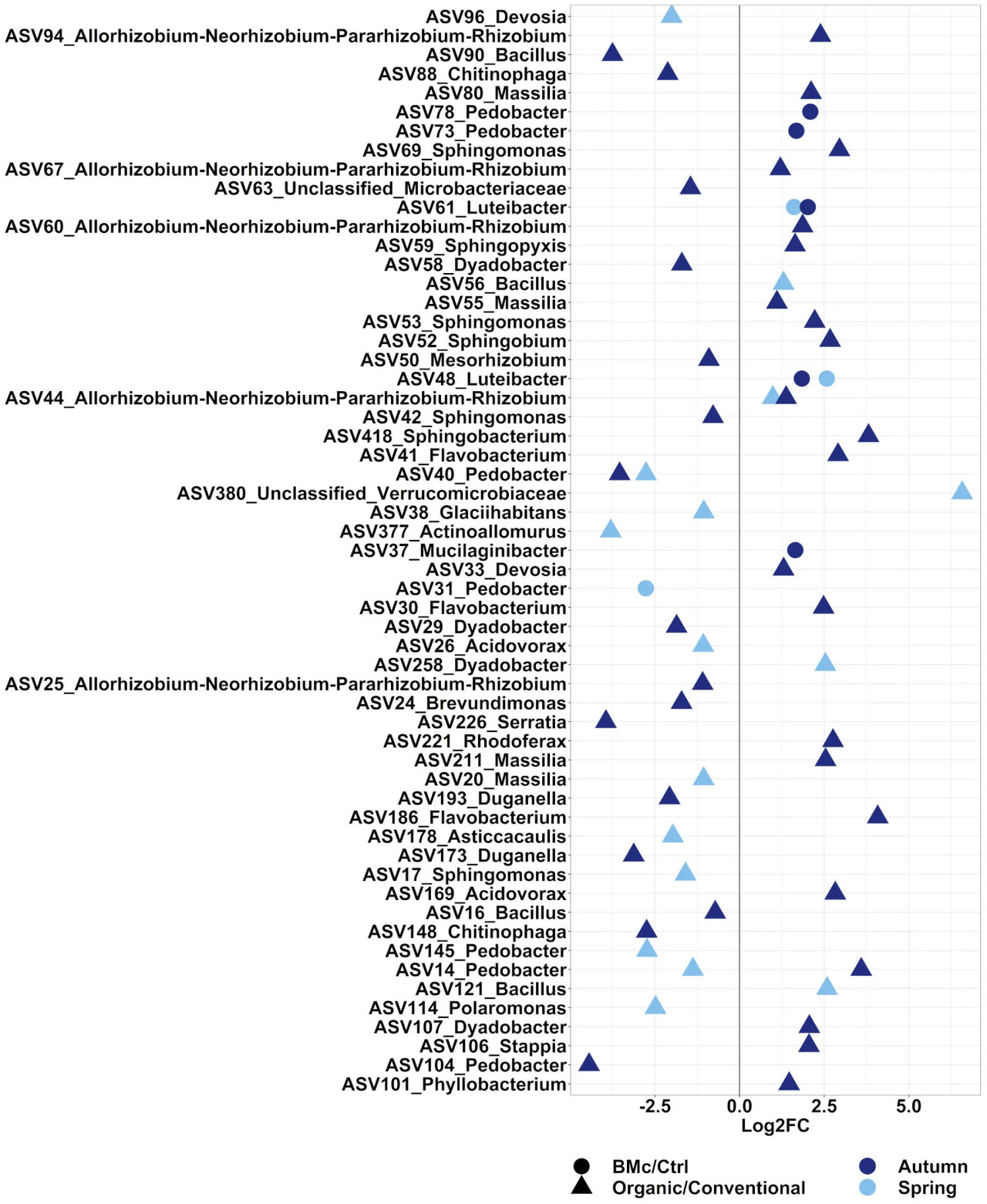
Рисунок 4 Log2-кратное изменение (FC) ASV, которые значительно различались и встречались с относительной обильностью выше 0,5%. Сравнения проводились между агротехническими практиками (Органическое и Традиционное) или инокуляцией БМк (БМк и Ctrl) в осеннюю или весеннюю кампанию отбора проб в течение роста растений озимой ржи. Тест на дифференциальную обильность выполнялся на разреженных наборах данных с помощью ANCOM-BC2 с коррекцией Бенджамини-Хохберга (скорректированное p-значение < 0,05).
Затем мы стремились выяснить ASV, которые значительно различались между каждой комбинацией обработок. В общей сложности 304 и 236 ASV были значительно различались между комбинациями обработок в осеннем и весеннем отборах соответственно (Дополнительная таблица 7), причем 43 ASV значительно различались независимо от времени отбора (Рисунок 4, Дополнительная таблица 8). Модели логистической регрессии выявили 277 (осенний отбор) и 210 (весенний отбор) ASV в качестве репрезентативных предикторов для каждой конкретной комбинации обработок (Дополнительная таблица 9). Из этих ASV относительная обильность 30 и 46 ASV увеличивалась в образцах, инокулированных БМк, в условиях органического удобрения осенью и весной соответственно (Дополнительная таблица 9), где была обнаружена значительно более высокая СМП (Рисунок 2A). В отличие от этого, в органическом земледелии инокуляция БМк приводила к снижению относительной обильности 57 ASV при осеннем отборе и 19 ASV при весеннем отборе (Дополнительная таблица 9). Кроме того, три ASV стабильно реагировали на инокуляцию БМк в органической обработке в каждую кампанию отбора (Дополнительная таблица 10).
Из этих дифференциально обильных ASV между четырьмя комбинациями обработок 28 и 12 ASV превышали относительную обильность 0,5% хотя бы в одном образце в осеннем (Рисунок 5A) и весеннем отборах соответственно (Рисунок 5B). Из этих доминирующих дискриминантных ASV 13 ASV ассоциировались положительно или отрицательно с инокуляцией БМк в органической обработке. Отрицательные ASV-респондеры были отнесены к Chitinophaga spp. (ASV148 и ASV88), Duganella spp. (ASV173 и ASV193), Brevundimonas spp. (ASV24), Microbacteriaceae spp. (ASV63), Sphingomonas spp. (ASV17) и Actinoallomurus spp. (ASV377), в то время как положительные респондеры были отнесены к Sphingobium spp. (ASV52), Bacillus spp. (ASV121), Pedobacter spp. (ASV95) и Luteibacter spp. (ASV61; только весна). Наиболее примечательно, что ASV88 (Chitinophaga sp.) был единственным наиболее обильным, значительно различающимся ASV, который отрицательно реагировал на инокуляцию БМк в органической обработке в оба срока отбора (Рисунок 5). Некоторые из этих ASV также ассоциировались с другими обработками, особенно в контроле традиционного земледелия. Например, ASV88 (Chitinophaga spp.) был обогащен в контрольных образцах в условиях традиционного земледелия и истощался при инокуляции БМк в органической обработке в оба срока отбора. Однако при весеннем отборе ASV61 (Luteibacter spp.) положительно реагировал на инокуляцию БМк в органической обработке и отрицательно — на контроль в традиционном земледелии (Рисунок 5).
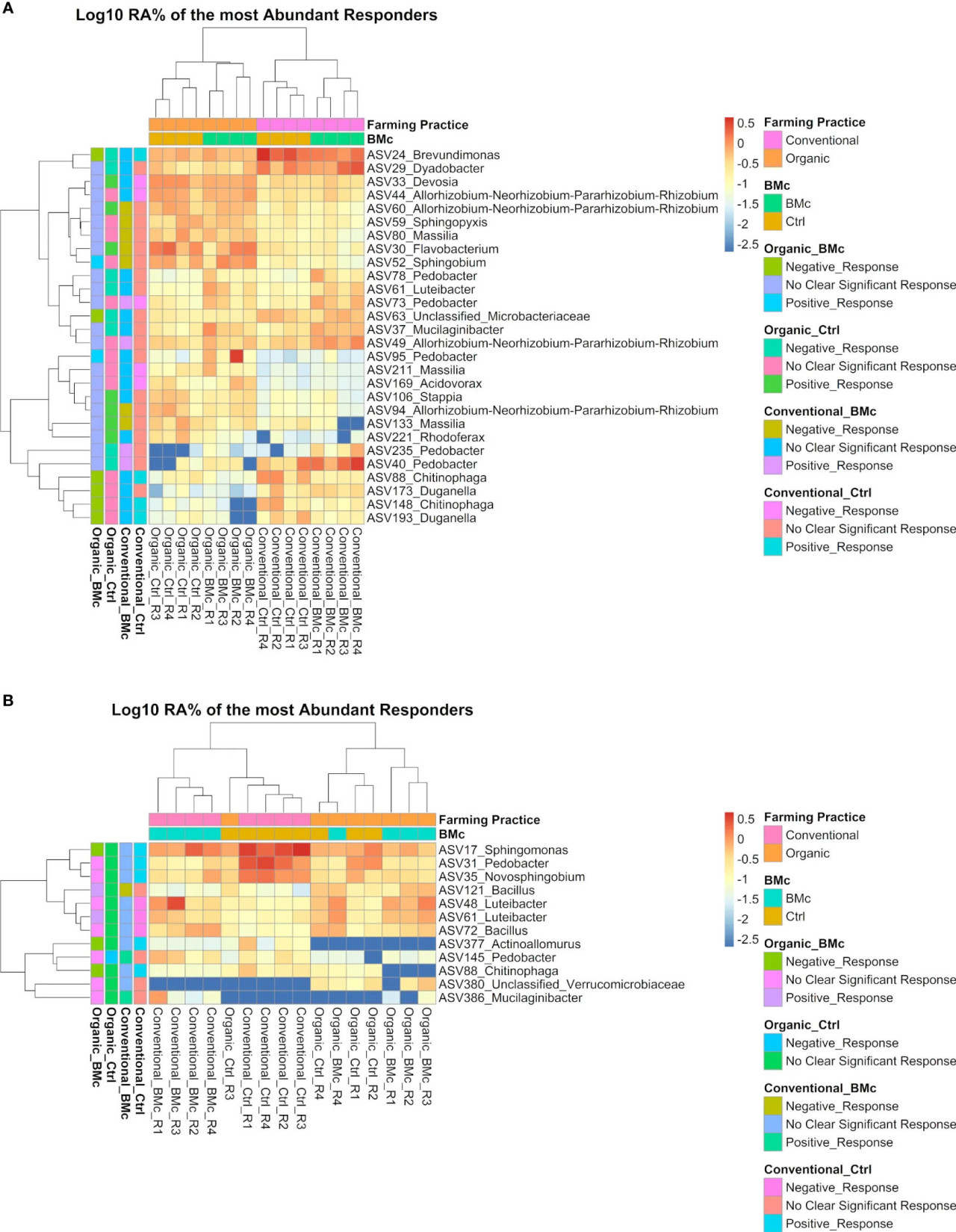
Рисунок 5 Тепловые карты с log10-преобразованным средним относительным обилием дифференциально обильных ASV в осеннем (A) и весеннем (B) отборе. Представленные ASV встречались с относительной обильностью выше 0,5% хотя бы в одном образце. Значительно различающиеся ASV были идентифицированы с помощью ANCOM-BC2 (скорректированное p-значение по Бенджамини-Хохбергу < 0,05, n=4). Цветная легенда (слева) показывает, реагировали ли эти ASV положительно или отрицательно на конкретную комбинацию обработок (Organic-Ctrl, Organic-BMc, Conventional-Ctrl, Conventional-BMc). ASV-респондеры были идентифицированы с помощью логистической регрессии (скорректированное p-значение по Бенджамини-Хохбергу < 0,05). RA, Относительная обильность.
Наконец, мы выполнили анализ филогенетической кластеризации для оценки того, были ли ASV-респондеры на инокуляцию БМк в органической обработке филогенетически связаны с инокулированными штаммами БМ (Рисунок 6). ASV-респондеры, классифицированные как Bacillus spp., кластеризовались ближе к другим Bacillus spp., чем к генам 16S рРНК B. atrophaeus ABi03 (Рисунок 6A). В отличие от этого, два из трех ASV, классифицированных как Pseudomonas spp. (ASV1123 и ASV1576), кластеризовались вместе с шестью копиями гена 16S рРНК Pseudomonas sp. RU47 (Рисунок 6B). Эти два ASV положительно ассоциировались с инокуляцией БМк в органической обработке при осеннем отборе, но не при весеннем (Дополнительный рисунок 3).
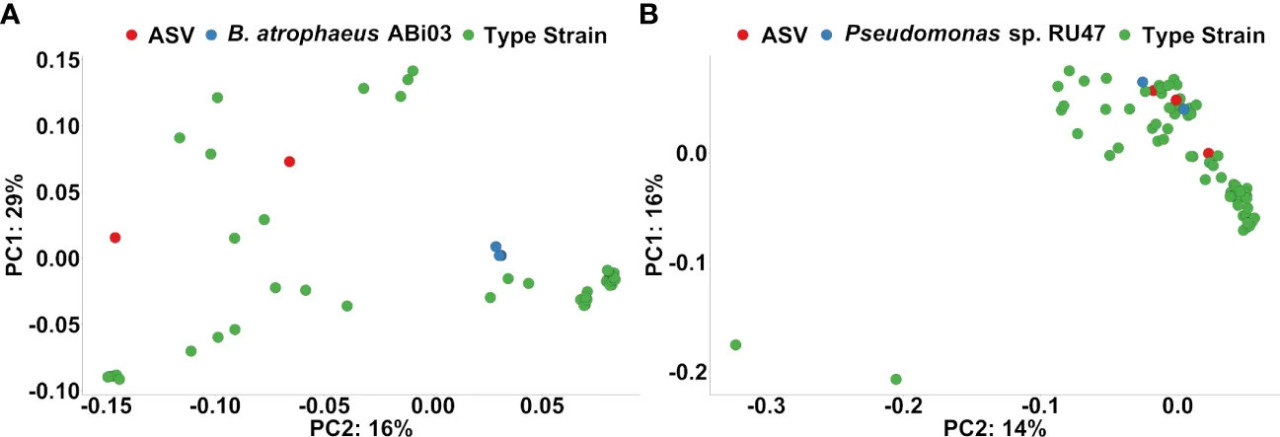
Рисунок 6 (A) Графики ординации на основе филогенетического расстояния после попарного локального выравнивания последовательности гена 16S рРНК из ASV-респондеров на Organic-BMc, классифицированных как Bacillus spp., генов 16S рРНК Bacillus atrophaeus ABi03 и Bacillus spp. из коллекции типовых штаммов Genbank (Красный: ASV-респондеры, Синий: Bacillus atrophaeus ABi03, Зеленый: типовые штаммы Bacillus spp.). График не показывает указаний на то, что эти ASV-респондеры, классифицированные как Bacillus spp., могут принадлежать к Bacillus ABi03. (B) Графики ординации на основе филогенетического расстояния после попарного локального выравнивания последовательности гена 16S рРНК из ASV-респондеров на Organic-BMc, классифицированных как Pseudomonas spp., штамма Pseudomonas sp. RU47 (шесть генов 16S рРНК) и Pseudomonas spp. из коллекции типовых штаммов Genbank (Красный: ASV-респондеры, Синий: Pseudomonas sp. RU47, Зеленый: типовые штаммы Pseudomonas spp.). График указывает на то, что два ASV-респондера (ASV1123 и ASV1576) вероятно принадлежат к штамму-инокулянту Pseudomonas sp. RU47, поскольку они кластеризуются с генами 16S рРНК штамма-инокулянта Pseudomonas sp. RU47.
Обсуждение
Успешное закрепление микробного консорциума на протяжении зимнего покоя
В этом исследовании мы выдвинули гипотезу, что каждый член консорциума колонизирует ризосферу растений озимой ржи и поддерживает высокую плотность в течение зимнего покоя независимо от агротехнических практик (органическое и традиционное земледелие). Три члена консорциума были отобраны на основе ранее наблюдаемых положительных эффектов на продуктивность растений. Штамм Pseudomonas sp. RU47 усиливал рост томатов и кукурузы в бедных фосфором почвах (Eltlbany и др., 2019) и очень эффективно подавлял корневую гниль (Rhizoctonia solani) у салата в тепличных и полевых условиях, хотя в анализах in vitro не было обнаружено сильных антагонистических свойств (Adesina и др., 2009; Schreiter и др., 2018). Мы предположили, что эффект подавления болезней был в основном основан на продукции HCN (Kuzmanović и др., 2018). В отличие от этого, штамм Bacillus atrophaeus ABi03 показывал различные антагонистические функции in vitro. Стимуляция роста растений также наблюдалась в консорциумах, содержащих штамм Trichoderma harzianum OMG16, у томатов (Mpanga и др., 2018) и кукурузы (Mpanga и др., 2019; Moradtalab и др., 2020). Кроме того, OMG16 снижал корневую инфекцию Verticillium longisporum у рапса (Hafiz и др., 2022). В этом исследовании штаммы с различными механизмами действия применялись в виде консорциума в полевых условиях. Несколько исследований показали, что использование консорциума из двух или более микроорганизмов может оказывать более сильное и устойчивое влияние на здоровье и продуктивность растений, чем отдельный полезный микроорганизм (Sharma и др., 2020; Hafiz и др., 2022). Аддитивные или синергетические эффекты могут частично объясняться разделением местообитаний, таких как ризосфера, прикорневая почва и корневая кора, а также различными полезными функциями, такими как улучшение доступности питательных веществ, продукция фитогормонов, антагонистические функции против патогенов растений и смягчение (а)биотического стресса (Jha и Saraf, 2012; De Vrieze и др., 2018; Gu и др., 2020; Santoyo и др., 2021).
Долгосрочное сосуществование между членами БМк в ризосфере или прикорневой почве имеет решающее значение для обеспечения полезных эффектов на растения в короткие и длительные периоды. Через семь недель после инокуляции (осенний отбор) все члены консорциума колонизировали корни озимой ржи с плотностью, сходной с предыдущими исследованиями (Schreiter и др., 2018; Jamil и др., 2021), что указывает на то, что совместное применение полезных микроорганизмов в виде консорциума не ухудшало их ризосферную компетентность. Бактериальные штаммы ABi03 и RU47 обнаруживались главным образом в ризосфере и не ингибировали друг друга, как наблюдалось в двойной культуре (данные не показаны), тогда как OMG16 преимущественно обитает в прикорневой почве, но также ранее сообщалось как эндофит у рапса (Hafiz и др., 2022). Из-за этого пространственного разделения потенциальные антагонистические функции, продемонстрированные in vitro для RU47 и ABi03 (Таблица 1), вероятно, не могут действовать против OMG16. Помимо прямой конкуренции, закрепление применяемых микроорганизмов сильно зависит от состава и структуры аборигенного микробного сообщества, как было показано в предыдущих исследованиях, которые связывали низкую ризосферную компетентность применяемых микроорганизмов с высоким микробным разнообразием, которое действует как барьер против внешних захватчиков (Schierstaedt и др., 2020; Mawarda и др., 2022). Успешное закрепление инокулированных микроорганизмов в ризосфере связывали с быстрым использованием питательных веществ, образованием биопленки, а также антагонистическими способностями (Adam и др., 2016; Kumawat и др., 2019). Продемонстрированные in vitro антагонистические функции всех членов консорциума могут способствовать их избирательной оккупации ниш в ризосфере, тем самым способствуя их интеграции в микробное сообщество (Berg и др., 2021).
Хотя наши результаты (Рисунок 3) подтверждают, что агротехническая практика определяет состав микробных сообществ в почве и ризосфере (Schrama и др., 2018; Babin и др., 2019; Bziuk и др., 2021; Fernandez-Gnecco и др., 2022), ризосферная компетентность БМк не пострадала, что указывает на надежную компетентность членов консорциума в различных условиях окружающей среды. Поскольку лишь несколько ASV-респондеров стабильно различались между агротехническими практиками в оба срока отбора, мы предполагаем, что формирование ризосферной микробиоты было стохастическим процессом, на который влияли условия окружающей среды и стадии развития растений. Постоянная зависимая от времени стохастическая сборка бактериальных сообществ в ризосфере могла способствовать высокой ризосферной компетентности членов консорциума независимо от агротехнических практик. Весной плотность колонизации RU47 в ризосфере снизилась в целом в 10 раз, тогда как плотности ABi03 и OMG16 (в прикорневой почве) остались почти такими же по сравнению с осенним отбором. Более высокая устойчивость ABi03 и OMG16 может быть объяснена их способностью к спорообразованию, что позволяет им лучше выживать в неблагоприятных зимних условиях по сравнению с RU47 (Fernández-Sandoval и др., 2012; Sella и др., 2014). Тем не менее, все три микроорганизма достаточно колонизировали ризосферу озимой ржи даже через 22 недели после инокуляции. Это указывает на сильную положительную обратную связь между растением и членами консорциума, поскольку сохранение высокой плотности колонизации зависит, с одной стороны, от профиля корневых экссудатов растения, а с другой — от хемотаксиса каждого члена консорциума к этим метаболитам и их использования в качестве субстрата (Malgioglio и др., 2022).
Консорциум улучшает продуктивность растений и усвоение питательных веществ в условиях органического земледелия
Рост побегов озимой ржи стимулировался инокуляцией БМк осенью (+20%), но особенно во время отрастания после зимы (+45%) исключительно в условиях органического земледелия. Эти результаты согласуются с высокой ризосферной компетентностью, показанной исследованными инокулянтами, которая оставалась неизменной даже в неблагоприятных зимних условиях. Более того, в условиях долгосрочной органической агротехнической практики инокуляция БМк ассоциировалась с обогащением бактериальных таксонов с документированными свойствами стимуляции роста растений. Следовательно, органическая агротехническая практика может поддерживать почвенные биологические процессы, которые обеспечивают более сложные таксономические и функциональные сообщества (Bender и др., 2016). Многочисленные исследования стабильно сообщают о положительных эффектах на полезные взаимодействия растений с микроорганизмами при использовании удобрений на основе навоза вместе с инокулянтами, такими как Bacillus, Pseudomonas и Trichoderma (Thonar и др., 2017; Mpanga и др., 2018; Bradáčová и др., 2019). Полезное взаимодействие растений с микроорганизмами в условиях органического земледелия может улучшить углеродное снабжение быстрорастущих копиотрофных инокулянтов, а также аборигенных микроорганизмов, стимулирующих рост растений, тем самым способствуя закреплению полезного микробного сообщества. Этот эффект может иметь особое значение на легких песчаных почвах с низким содержанием органического углерода, которые наблюдались в этом исследовании даже после длительной органической агротехнической практики (Дополнительная таблица 4). Соответственно, мета-анализ показал, что отзывчивость на микробные инокулянты снижается с увеличением содержания органического углерода в почве (Schütz и др., 2018). Кроме того, высокая доступность N и P, характерная для удобрений на основе навоза, может служить стартовым удобрением для растения-хозяина, облегчая рост корней и закрепление микробных инокулянтов в ризосфере (Bittman и др., 2006; Chekanai и др., 2018).
Полезные эффекты инокулянтов и их влияние на ризосферный микробиом могут частично объясняться улучшенным питанием растений. Осенью значительных различий в концентрациях питательных веществ в побегах между обработками изначально не наблюдалось, что отражало сопоставимый питательный статус (Таблица 2). Однако дисперсионный анализ показал общее влияние инокуляции БМк на концентрацию N в побегах (Дополнительная таблица 3). Дефицит питательных веществ был очевиден только для Ca (Bergmann, 1988) и мог быть связан с низким уровнем Ca на легких песчаных почвах полевого участка (Дополнительная таблица 4). В отличие от этого, во время отрастания после зимы критические концентрации питательных веществ были зафиксированы, особенно в условиях органического удобрения, включая N, P, K, Mg, Ca, Cu и Zn. Инокуляция БМк значительно улучшала питательный статус по P, который является критическим питательным веществом во многих органических системах земледелия (Cooper и др., 2018). Однако сходные тенденции также наблюдались для всех остальных питательных веществ, за исключением Zn и Mn (Таблица 2). Улучшенный питательный статус растений благодаря инокуляции БМк также подтверждается дисперсионным анализом, который показывает общий эффект инокуляции БМк на концентрацию этих питательных веществ независимо от агротехнической практики (Дополнительная таблица 3). В результате концентрации питательных веществ у растений, инокулированных БМк, достигали или даже превышали пороги достаточности, особенно для N и P. Эти результаты свидетельствуют об общем положительном эффекте БМк на усвоение питательных веществ, возможно, опосредованном стимуляцией роста корней, что хорошо документировано для штаммов-инокулянтов (Mpanga и др., 2018; Eltlbany и др., 2019; Moradtalab и др., 2020). Однако улучшение усвоения питательных веществ растениями за счет прямой мобилизации питательных веществ БМк является менее вероятным сценарием, хотя тесты in vitro действительно обнаружили солюбилизацию P и K. Анализ пулов питательных веществ в прикорневой почве не показал никаких изменений после инокуляции БМк, что свидетельствует об отсутствии специфических эффектов на солюбилизацию доступных почвенных питательных веществ весной (Дополнительная таблица 4). Согласно современной литературе, P-солюбилизирующие микроорганизмы вносят вклад в питание растения-хозяина главным образом через их долгосрочное влияние на круговорот питательных веществ, а не через прямое растворение питательных веществ (Raymond и др., 2021). Интересно, что сходные тенденции улучшения питательного статуса после инокуляции БМк также были показаны для растений, выращенных в условиях традиционного земледелия. Однако по сравнению с органическим земледелием традиционная агротехническая практика в целом приводила к более высокому питательному статусу растений, достигая уровней достаточности для N, P и Mg даже у неинокулированных растений. Это может объяснять отсутствие дополнительных ростовых ответов на инокуляцию БМк в условиях традиционного земледелия.
Хотя значительных различий между обработками не было, итоговые данные по урожайности (Рисунок 2B) соответствовали эффектам на биомассу побегов во время весеннего вегетативного роста (Рисунок 2A). Урожайность зерна в условиях долгосрочного традиционного земледелия была немного выше средней урожайности ржи за 2015-2020 гг. (4,1 т га⁻¹) в округе Бранденбург (Германия). Органическая агротехническая практика имела тенденцию к снижению урожайности примерно на 22%, что сходно со средними потерями урожайности, определенными мета-исследованиями (de Ponti и др., 2012; Seufert и др., 2012). Хотя это и не значимо, наши результаты указывают на то, что это снижение урожайности при органическом земледелии может быть частично компенсировано применением БМк.
Зависящая от инокуляции модуляция бактериального сообщества ризосферы сохраняется на протяжении зимнего покоя
Мы предположили, что применение БМк будет влиять на состав бактериального сообщества в ризосфере в зависимости от агротехнической практики. Следовательно, мы ожидали, что эти вариации будут оказывать различное влияние на продуктивность озимой ржи. Изменение состава бактериального сообщества в ризосфере озимой ржи было обнаружено в обработках с инокуляцией БМк, независимо от агротехнических практик. Этот эффект сохранялся до весны, что указывает на продолжительное влияние БМк (Рисунок 3). Сходные результаты были получены в исследовании Deng и др. (2019), где инокуляция БМк влияла на состав бактериального сообщества корней земляники в различные моменты времени, независимо от агротехнических практик. Это указывает на то, что повторное применение инокулянтов может не требоваться после того, как их эффекты установлены. Однако для выяснения того, сохраняются ли эффекты БМк на ризосферную микробиоту и продуктивность растений на протяжении всего вегетационного периода в полевых условиях, необходимы дальнейшие эксперименты с временными рядами.
В дополнение к инокуляции БМк мы наблюдали влияние агротехнических практик на почвенную микробиоту, сходное с нашими предыдущими исследованиями (Bziuk и др., 2021; Windisch и др., 2021; Fernandez-Gnecco и др., 2022; Sommermann и др., 2022). Однако впервые мы сообщаем, что этот эффект сохраняется на различных стадиях развития растений. В то время как и агротехническая практика, и инокуляция БМк значительно влияли на состав бактериального сообщества, лишь немногие ASV демонстрировали стабильные закономерности дифференциальной обильности с течением времени (Рисунок 4). Это указывает: a) на то, что рекрутирование микроорганизмов в ризосфере различалось на разных стадиях развития растений, и b) на то, что инокуляция БМк и агротехнические практики влияли на это рекрутирование. Сходное влияние агротехнических практик на рекрутирование микроорганизмов в ризосфере было ранее продемонстрировано (Windisch и др., 2021; Sommermann и др., 2022). В итоге, влияние инокуляции БМк на ризосферную микробиоту сохранялось на протяжении зимнего покоя, что указывает на специфические стратегии, такие как спорообразование членами консорциума для выживания в зимний период. Кроме того, изменения в корневых экссудатах весной могли дополнительно активировать дремлющие БМ (Windisch и др., 2017).
Согласно экологическим теориям взаимодействия растений и микроорганизмов, мутуалистический отбор полезных для растений микроорганизмов часто возникает в условиях ограничения питательных веществ (Sánchez-Cañizares и др., 2017). Поскольку химические удобрения обеспечивают высокую доступность питательных веществ для растений и ризосферной микробиоты, они могут вмешиваться в процессы отбора растений в пользу полезных микроорганизмов (Sánchez-Cañizares и др., 2017). Интересно, что мы наблюдали повышенную ростовую активность растений, инокулированных БМк, только в условиях органического земледелия, где доступность питательных веществ была низкой (Дополнительная таблица 4). Мы полагаем, что этот аддитивный эффект может быть связан с более высоким давлением отбора как на растения, так и на почвенные микроорганизмы из-за ограничения питательных веществ, в отличие от традиционного земледелия с высокой доступностью питательных веществ. Однако данные КОЕ показали, что агротехнические практики незначительно изменяли плотность применяемого БМк в почве (Рисунок 1). При этом тест на дифференциальную обильность показал обогащение ASV, тесно связанных с Pseudomonas spp. или Bacillus spp., которые могут соответствовать нашим инокулянтам БМк. Применяя филогенетическую кластеризацию, мы идентифицировали два ASV Pseudomonas, тесно связанных с Pseudomonas sp. RU47 (Рисунок 6). Более высокие доли (относительная обильность) Pseudomonas sp. RU47 в условиях органического земледелия и инокуляции БМк могут указывать на меньшую конкуренцию со стороны других бактериальных таксонов, населяющих ризосферу в органическом земледелии. Эффект взаимодействия между агротехническими практиками и инокуляцией БМк на профили бактериального сообщества ризосферы был слабым. Таким образом, две агротехнические практики лишь незначительно влияли на способность БМк модифицировать бактериальное сообщество ризосферы. Тем не менее, мы попытались идентифицировать ASV, ассоциированные с инокуляцией БМк в условиях органического земледелия, чтобы определить, какая часть бактериальных таксонов связана с инокулированным БМк. Было обнаружено несколько ASV-респондеров, но большинство из них имели относительную обильность ниже 0,5%.
Среди доминантных респондеров (>0,5% относительной обильности) несколько ASV, классифицированных как известные потенциально полезные для растений бактерии, увеличивали свою относительную обильность в ризосфере озимой ржи (Рисунок 5). Например, члены рода Luteibacter (которые относятся к γ-протеобактериям) известны своими свойствами стимуляции роста растений (Guglielmetti и др., 2013; Hoffman и др., 2013) и были обогащены в ризосфере органической обработки. Интересно, что Luteibacter spp. могут действовать как симбионты грибных эндофитов растений и запускать продукцию ИУК, которая стимулирует рост растений. Аналогично, один ASV, классифицированный как Pedobacter spp. (ASV95), род, включающий несколько полезных для растений видов, увеличивал относительную обильность благодаря инокуляции БМк в условиях органического земледелия (Morais и др., 2019). Хотя комбинация инокуляции БМк и органического земледелия может не оказывать существенного влияния на общий профиль бактериального сообщества, она увеличивала относительную обильность бактериальных таксонов, которые потенциально действуют как полезные для растений микроорганизмы, стимулирующие рост растений. Однако для подтверждения того, что микробные респондеры, вызванные инокуляцией БМк, также способствовали увеличению роста растений, необходимы дальнейшие полевые исследования.
Среди доминантных респондеров со снижающейся относительной обильностью из-за инокуляции БМк, в основном в органическом земледелии, мы идентифицировали ASV, классифицированные как Chitinophaga sp. или Duganella sp. Эти ASV ранее были связаны с возникновением заболеваний растений (Carrión и др., 2019; Li и др., 2021). Органическая агротехническая практика и комбинация БМк, вероятно, снижали бактериальные таксоны, ассоциированные с болезнями растений и стрессовыми условиями. В отличие от этого, некоторые из этих ASV имели более высокую относительную обильность в условиях традиционной агротехнической практики без инокуляции БМк, что указывает на то, что высокий уровень поступления питательных веществ через химические удобрения мог привести к накоплению бактериальных таксонов, ассоциированных с болезнями растений. Альтернативно, растения часто рекрутируют несколько близкородственных бактерий в ответ на патоген (например, Duganella spp.) (Haack и др., 2016), поэтому прямая ассоциация этих таксонов с патогенностью растений невозможна, тем более что фенотипа заболевания растений не обнаружено.
Заключение
Это исследование проиллюстрировало эффекты консорциума полезных микроорганизмов (БМк) на продуктивность озимой ржи в зависимости от различных агротехнических стратегий, т.е. традиционного и органического земледелия. Основной целью нашего исследования была оценка способности членов консорциума выживать в достаточной плотности в ризосфере/прикорневой почве озимой ржи как предпосылки для успешного взаимодействия растений и микроорганизмов. Наши результаты показали, что члены консорциума сохранялись в ризосфере ржи на протяжении вегетационного периода, поддерживая высокую плотность даже после зимнего покоя независимо от агротехнической практики. Как и ожидалось, БМк оказывал положительное влияние на продуктивность ржи, увеличивая сухую биомассу побегов и питательный статус растений, особенно при органическом удобрении. Эти результаты дополнительно подтвердили протестированные in vitro ростостимулирующие свойства, которые характеризовали микробных членов, инокулированных в ризосферу озимой ржи в полевых условиях. Более того, инокулированный БМк оказывал значительное влияние на динамику бактериального сообщества, поскольку после инокуляции как в органических, так и в традиционных обработках наблюдались значительные сдвиги в бактериальной сборке ризосферы, и такие сдвиги в сообществе также обнаруживались на разных стадиях развития растений. Интересно, что в условиях органического земледелия мы наблюдали, что в ризосфере ржи, обработанной БМк, несколько бактериальных таксонов, ранее ассоциированных с болезнями растений, были истощены, в то время как несколько предполагаемых полезных для растений бактериальных таксонов были обогащены по своей относительной обильности, что дополнительно подчеркивает положительные эффекты БМк на здоровье растений путем модуляции ризосферного микробиома ржи. Примечательно, что хотя наш полевой эксперимент показал положительное влияние БМк на продуктивность ржи, особенно при органическом удобрении, положительное влияние на урожайность ржи было лишь незначительным. Тем не менее, наши результаты указывают на то, что инокуляция БМк может иметь потенциал для компенсации потерь урожайности, вызванных дефицитом питательных веществ в органической агротехнической практике. Кроме того, обработка БМк может способствовать лучшей стабильности урожайности за счет улучшения питательного статуса растений и стимулирования полезной микробиоты, что представляет собой устойчивый подход, особенно в сочетании с органическим земледелием. Наше исследование подчеркивает важность проведения дополнительных полевых экспериментов для понимания эффективности микробных инокулянтов в различных системах земледелия, способствуя разработке эффективных микробных решений для устойчивого сельского хозяйства.
Дополнительный материал
Дополнительный материал к этой статье можно найти онлайн по адресу: https://www.frontiersin.org/articles/10.3389/fpls.2023.1232288/full#supplementary-material
Ссылки
1. Adam E., Groenenboom A. E., Kurm V., Rajewska M., Schmidt R., Tyc O., et al. (2016). Controlling the microbiome: microhabitat adjustments for successful biocontrol strategies in soil and human gut. Front. Microbiol. 7. doi: 10.3389/fmicb.2016.01079. CrossRef. Google Scholar.
2. Adesina M. F., Grosch R., Lembke A., Vatchev T. D., Smalla K. (2009). In vitro antagonists of Rhizoctonia solani tested on lettuce: rhizosphere competence, biocontrol efficiency and rhizosphere microbial community response. FEMS Microbiol. Ecol. 69, 62–74. doi: 10.1111/j.1574-6941.2009.00685.x. CrossRef. Google Scholar.
3. Ali S., Hameed S., Shahid M., Iqbal M., Lazarovits G., Imran A. (2020). Functional characterization of potential PGPR exhibiting broad-spectrum antifungal activity. Microbiol. Res. 232, 126389. doi: 10.1016/j.micres.2019.126389. CrossRef. Google Scholar.
4. Babin D., Deubel A., Jacquiod S., Sørensen S. J., Geistlinger J., Grosch R., et al. (2019). Impact of long-term agricultural management practices on soil prokaryotic communities. Soil Biol. Biochem. 129, 17–28. doi: 10.1016/j.soilbio.2018.11.002. CrossRef. Google Scholar.
5. Batista B. D., Singh B. K. (2021). Realities and hopes in the application of microbial tools in agriculture. Microb. Biotechnol. 14, 1258–1268. doi: 10.1111/1751-7915.13866. CrossRef. Google Scholar.
6. Bender S. F., Wagg C., van der Heijden M. G. A. (2016). An underground revolution: biodiversity and soil ecological engineering for agricultural sustainability. Trends Ecol. Evol. 31, 440–452. doi: 10.1016/j.tree.2016.02.016. CrossRef. Google Scholar.
7. Berg G., Fritze A., Roskot N., Smalla K. (2001). Evaluation of potential biocontrol rhizobacteria from different host plants of Verticillium dahliae Kleb. J. Appl. Microbiol. 91, 963–971. doi: 10.1046/j.1365-2672.2001.01462.x. CrossRef. Google Scholar.
8. Berg G., Krechel A., Ditz M., Sikora R. A., Ulrich A., Hallmann J. (2005). Endophytic and ectophytic potato-associated bacterial communities differ in structure and antagonistic function against plant pathogenic fungi. FEMS Microbiol. Ecol. 51, 215–229. doi: 10.1016/j.femsec.2004.08.006. CrossRef. Google Scholar.
9. Berg G., Kusstatscher P., Abdelfattah A., Cernava T., Smalla K. (2021). Microbiome modulation—Toward a better understanding of plant microbiome response to microbial inoculants. Front. Microbiol. 12. doi: 10.3389/fmicb.2021.650610. CrossRef. Google Scholar.
10. Berg G., Smalla K. (2009). Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microbiol. Ecol. 68, 1–13. doi: 10.1111/j.1574-6941.2009.00654.x. CrossRef. Google Scholar.
11. Bergmann W. (Ed.) (1988). Ernährungsstörungen bei Kulturpflanzen. Stuttgart: G. Fischer. Google Scholar.
12. Bittman S., Kowalenko C. G., Hunt D. E., Forge T. A., Wu X. (2006). Starter phosphorus and broadcast nutrients on corn with contrasting colonization by mycorrhizae. Agron. J. 98, 394–401. doi: 10.2134/agronj2005.0093. CrossRef. Google Scholar.
13. Bodenhofer U., Bonatesta E., Horejš-Kainrath C., Hochreiter S. (2015). msa: an R package for multiple sequence alignment. Bioinformatics 31, btv494. doi: 10.1093/bioinformatics/btv494. CrossRef. Google Scholar.
14. Bradáčová K., Florea A., Bar-Tal A., Minz D., Yermiyahu U., Shawahna R., et al. (2019). Microbial consortia versus single-strain inoculants: an advantage in PGPM-assisted tomato production? Agronomy 9, 105. doi: 10.3390/agronomy9020105. CrossRef. Google Scholar.
15. Breitkreuz C., Reitz T., Schulz E., Tarkka M. (2021). Drought and plant community composition affect the metabolic and genotypic diversity of pseudomonas strains in grassland soils. Microorganisms 9, 1677. doi: 10.3390/microorganisms9081677. CrossRef. Google Scholar.
16. Bziuk N., Maccario L., Douchkov D., Lueck S., Babin D., Sørensen S. J., et al. (2021). Tillage shapes the soil and rhizosphere microbiome of barley—but not its susceptibility towards Blumeria graminis f. sp. hordei. FEMS Microbiol. Ecol. 97, fiab018. doi: 10.1093/femsec/fiab018. CrossRef. Google Scholar.
17. Callahan B. J., McMurdie P. J., Rosen M. J., Han A. W., Johnson A. J. A., Holmes S. P. (2016). DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 13, 581–583. doi: 10.1038/nmeth.3869. CrossRef. Google Scholar.
18. Cameron E. S., Schmidt P. J., Tremblay B. J.-M., Emelko M. B., Müller K. M. (2021). Enhancing diversity analysis by repeatedly rarefying next generation sequencing data describing microbial communities. Sci. Rep. 11, 22302. doi: 10.1038/s41598-021-01636-1. CrossRef. Google Scholar.
19. Cappuccino J. G., Sherman N. (2014). Microbiology: A Laboratory Manual. 10th ed. Boston: Pearson. Google Scholar.
20. Carrión V. J., Perez-Jaramillo J., Cordovez V., Tracanna V., de Hollander M., Ruiz-Buck D., et al. (2019). Pathogen-induced activation of disease-suppressive functions in the endophytic root microbiome. Science 366, 606–612. doi: 10.1126/science.aaw9285. CrossRef. Google Scholar.
21. Chaparro J. M., Badri D. V., Vivanco J. M. (2014). Rhizosphere microbiome assemblage is affected by plant development. ISME J. 8, 790–803. doi: 10.1038/ismej.2013.196. CrossRef. Google Scholar.
22. Charif D., Lobry J. R. (2007). "SeqinR 1.0-2: A contributed package to the R project for statistical computing devoted to biological sequences retrieval and analysis," in Structural Approaches to Sequence Evolution Biological and Medical Physics, Biomedical Engineering. Eds. Bastolla U., Porto M., Roman H. E., Vendruscolo M. Berlin, Heidelberg: Springer Berlin Heidelberg, 207–232. doi: 10.1007/978-3-540-35306-5_10. CrossRef. Google Scholar.
23. Chekanai V., Chikowo R., Vanlauwe B. (2018). Response of common bean (Phaseolus vulgaris L.) to nitrogen, phosphorus and rhizobia inoculation across variable soils in Zimbabwe. Agric. Ecosyst. Environ. 266, 167–173. doi: 10.1016/j.agee.2018.08.010. CrossRef. Google Scholar.
24. Chowdhury S. P., Babin D., Sandmann M., Jacquiod S., Sommermann L., Sørensen S. J., et al. (2019). Effect of long-term organic and mineral fertilization strategies on rhizosphere microbiota assemblage and performance of lettuce. Environ. Microbiol. 21, 2426–2439. doi: 10.1111/1462-2920.14631. CrossRef. Google Scholar.
25. Cock P. J. A., Grüning B. A., Paszkiewicz K., Pritchard L. (2013). Galaxy tools and workflows for sequence analysis with applications in molecular plant pathology. PeerJ 1, e167. doi: 10.7717/peerj.167. CrossRef. Google Scholar.
26. Cooper J., Reed E. Y., Hörtenhuber S., Lindenthal T., Løes A.-K., Mäder P., et al. (2018). Phosphorus availability on many organically managed farms in Europe. Nutr. Cycling Agroecosyst. 110, 227–239. doi: 10.1007/s10705-017-9894-2. CrossRef. Google Scholar.
27. Deng S., Wipf H. M.-L., Pierroz G., Raab T. K., Khanna R., Coleman-Derr D. (2019). A plant growth-promoting microbial soil amendment dynamically alters the strawberry root bacterial microbiome. Sci. Rep. 9, 17677. doi: 10.1038/s41598-019-53623-2. CrossRef. Google Scholar.
28. de Ponti T., Rijk B., van Ittersum M. K. (2012). The crop yield gap between organic and conventional agriculture. Agric. Syst. 108, 1–9. doi: 10.1016/j.agsy.2011.12.004. CrossRef. Google Scholar.
29. De Vrieze M., Germanier F., Vuille N., Weisskopf L. (2018). Combining different potato-associated pseudomonas strains for improved biocontrol of phytophthora infestans. Front. Microbiol. 9. doi: 10.3389/fmicb.2018.02573. CrossRef. Google Scholar.
30. Donate-Correa J., León-Barrios M., Pérez-Galdona R. (2005). Screening for plant growth-promoting rhizobacteria in Chamaecytisus proliferus (tagasaste), a forage tree-shrub legume endemic to the Canary Islands. Plant Soil 266, 261–272. doi: 10.1007/s11104-005-0754-5. CrossRef. Google Scholar.
31. Eltlbany N., Baklawa M., Ding G.-C., Nassal D., Weber N., Kandeler E., et al. (2019). Enhanced tomato plant growth in soil under reduced P supply through microbial inoculants and microbiome shifts. FEMS Microbiol. Ecol. 95, fiz124. doi: 10.1093/femsec/fiz124. CrossRef. Google Scholar.
32. FC M., Davis T. (2022). ggpattern: "ggplot2" Pattern geoms. Available at: https://CRAN.R-project.org/package=ggpattern. Google Scholar.
33. Federhen S. (2015). Type material in the NCBI taxonomy database. Nucleic Acids Res. 43, D1086–D1098. doi: 10.1093/nar/gku1127. CrossRef. Google Scholar.
34. Fernandez-Gnecco G., Covacevich F., Consolo V. F., Behr J. H., Sommermann L., Moradtalab N., et al. (2022). Effect of long-term agricultural management on the soil microbiota influenced by the time of soil sampling. Front. Soil Sci. 2. doi: 10.3389/fsoil.2022.837508. CrossRef. Google Scholar.
35. Fernández-Sandoval M. T., Ortiz-García M., Galindo E., Serrano-Carreón L. (2012). Cellular damage during drying and storage of Trichoderma harzianum spores. Process Biochem. 47, 186–194. doi: 10.1016/j.procbio.2011.10.006. CrossRef. Google Scholar.
36. Finkel O. M., Castrillo G., Herrera Paredes S., Salas González I., Dangl J. L. (2017). Understanding and exploiting plant beneficial microbes. Curr. Opin. Plant Biol. 38, 155–163. doi: 10.1016/j.pbi.2017.04.018. CrossRef. Google Scholar.
37. Fira D., Dimkić I., Berić T., Lozo J., Stanković S. (2018). Biological control of plant pathogens by Bacillus species. J. Biotechnol. 285, 44–55. doi: 10.1016/j.jbiotec.2018.07.044. CrossRef. Google Scholar.
38. Fox J., Weisberg S. (2019). An R companion to applied regression. 3rd ed. Thousand Oaks, CA: Sage. Available at: https://socialsciences.mcmaster.ca/jfox/Books/Companion/. Google Scholar.
39. Francioli D., Schulz E., Lentendu G., Wubet T., Buscot F., Reitz T. (2016). Mineral vs. Organic amendments: microbial community structure, activity and abundance of agriculturally relevant microbes are driven by long-term fertilization strategies. Front. Microbiol. 7. doi: 10.3389/fmicb.2016.01446. CrossRef. Google Scholar.
40. Graves S., Piepho H.-P., Selzer L. (2019). multcompView: visualizations of paired comparisons. Available at: https://CRAN.R-project.org/package=multcompView. Google Scholar.
41. Gu S., Yang T., Shao Z., Wang T., Cao K., Jousset A., et al. (2020). Siderophore-mediated interactions determine the disease suppressiveness of microbial consortia. mSystems 5, e00811–e00819. doi: 10.1128/mSystems.00811-19. CrossRef. Google Scholar.
42. Guglielmetti S., Basilico R., Taverniti V., Arioli S., Piagnani C., Bernacchi A. (2013). Luteibacter rhizovicinus MIMR1 promotes root development in barley (Hordeum vulgare L.) under laboratory conditions. World J. Microbiol. Biotechnol. 29, 2025–2032. doi: 10.1007/s11274-013-1365-6. CrossRef. Google Scholar.
43. Haack F. S., Poehlein A., Kröger C., Voigt C. A., Piepenbring M., Bode H. B., et al. (2016). Molecular Keys to the Janthinobacterium and Duganella spp. Interaction with the Plant Pathogen Fusarium graminearum. Front. Microbiol. 7, 7. doi: 10.3389/fmicb.2016.01668. CrossRef. Google Scholar.
44. Hafiz F. B., Moradtalab N., Goertz S., Rietz S., Dietel K., Rozhon W., et al. (2022). Synergistic effects of a root-endophytic trichoderma fungus and bacillus on early root colonization and defense activation against verticillium longisporum in rapeseed. MPMI 35, MPMI–11-21-0274-R. doi: 10.1094/MPMI-11-21-0274-R. CrossRef. Google Scholar.
45. Heuer H., Kopmann C., Binh C. T. T., Top E. M., Smalla K. (2009). Spreading antibiotic resistance through spread manure: characteristics of a novel plasmid type with low %G+C content. Environ. Microbiol. 11, 937–949. doi: 10.1111/j.1462-2920.2008.01819.x. CrossRef. Google Scholar.
46. Hoffman M. T., Gunatilaka M. K., Wijeratne K., Gunatilaka L., Arnold A. E. (2013). Endohyphal bacterium enhances production of indole-3-acetic acid by a foliar fungal endophyte. PloS One 8, e73132. doi: 10.1371/journal.pone.0073132. CrossRef. Google Scholar.
47. Hothorn T., Bretz F., Westfall P. (2008). Simultaneous inference in general parametric models. Biom. J. 50, 346–363. doi: 10.1002/bimj.200810425. CrossRef. Google Scholar.
48. Huber J., Bochow H., Junge H. (1987). Selektion und biotechnische Herstellung von Kulturlösungen mikrobieller Antagonisten zur Unterdrückung phytopathogener Bodenpilze. J. Basic Microbiol. 27, 497–503. doi: 10.1002/jobm.3620270907. CrossRef. Google Scholar.
49. Jacoby R., Peukert M., Succurro A., Koprivova A., Kopriva S. (2017). The role of soil microorganisms in plant mineral nutrition—Current knowledge and future directions. Front. Plant Sci. 8. doi: 10.3389/fpls.2017.01617. CrossRef. Google Scholar.
50. Jamil A., Musheer N., Ashraf S. (2021). Antagonistic potential of Trichoderma harzianum and Azadirachta indica against Fusarium oxysporum f. sp. capsici for the management of chilli wilt. J. Plant Dis. Prot. 128, 161–172. doi: 10.1007/s41348-020-00383-1. CrossRef. Google Scholar.
51. Jha C. K., Saraf M. (2012). Evaluation of multispecies plant-growth-promoting consortia for the growth promotion of jatropha curcas L. J. Plant Growth Regul. 31, 588–598. doi: 10.1007/s00344-012-9269-5. CrossRef. Google Scholar.
52. Kay M., Elkin L. A., Higgins J. J., Wobbrock J. O. (2021). mjskay/ARTool: ARTool 0.11.0. Zenodo. doi: 10.5281/ZENODO.594511. CrossRef. Google Scholar.
53. Koo S.-Y., Hong S. H., Ryu H. W., Cho K. (2010). Plant growth-promoting trait of rhizobacteria isolated from soil contaminated with petroleum and heavy metals. J. Microbiol. Biotechnol. 20, 587–593. doi: 10.4014/jmb.0907.07017. CrossRef. Google Scholar.
54. Kumawat K. C., Sharma P., Sirari A., Singh I., Gill B. S., Singh U., et al. (2019). Synergism of Pseudomonas aeruginosa (LSE-2) nodule endophyte with Bradyrhizobium sp. (LSBR-3) for improving plant growth, nutrient acquisition and soil health in soybean. World J. Microbiol. Biotechnol. 35, 35. doi: 10.1007/s11274-019-2622-0. CrossRef. Google Scholar.
55. Kuzmanović N., Eltlbany N., Ding G., Baklawa M., Min L., Wei L., et al. (2018). Analysis of the genome sequence of plant beneficial strain Pseudomonas sp. RU47. J. Biotechnol. 281, 281. doi: 10.1016/j.jbiotec.2018.07.023. CrossRef. Google Scholar.
56. Latz E., Eisenhauer N., Rall B. C., Scheu S., Jousset A. (2016). Unravelling linkages between plant community composition and the pathogen-suppressive potential of soils. Sci. Rep. 6, 23584. doi: 10.1038/srep23584. CrossRef. Google Scholar.
57. Lenth R. (2022). emmeans: Estimated Marginal Means, aka Least-Squares Means. Available at: https://CRAN.R-project.org/package=emmeans. Google Scholar.
58. Li H., Song F., Wu X., Deng C., Xu Q., Peng S., et al. (2021). Microbiome and metagenome analysis reveals huanglongbing affects the abundance of citrus rhizosphere bacteria associated with resistance and energy metabolism. Horticulturae 7, 151. doi: 10.3390/horticulturae7060151. CrossRef. Google Scholar.
59. Lin C.-P., Ho Y.-C. (2021). Beneficial microbes and basal fertilization in antagonism of banana fusarium wilt. Agronomy 11, 2043. doi: 10.3390/agronomy11102043. CrossRef. Google Scholar.
60. Lin H., Peddada S. D. (2020). Analysis of compositions of microbiomes with bias correction. Nat. Commun. 11, 3514. doi: 10.1038/s41467-020-17041-7. CrossRef. Google Scholar.
61. Liu H., Li J., Carvalhais L. C., Percy C. D., Prakash Verma J., Schenk P. M., et al. (2021). Evidence for the plant recruitment of beneficial microbes to suppress soil-borne pathogens. New Phytol. 229, 2873–2885. doi: 10.1111/nph.17057. CrossRef. Google Scholar.
62. Lopes M. J., dos S., Dias-Filho M. B., Gurgel E. S. C. (2021). Successful plant growth-promoting microbes: inoculation methods and abiotic factors. Front. Sustain. Food Syst. 5. doi: 10.3389/fsufs.2021.606454. CrossRef. Google Scholar.
63. Lori M., Symnaczik S., Mäder P., De Deyn G., Gattinger A. (2017). Organic farming enhances soil microbial abundance and activity—A meta-analysis and meta-regression. PloS One 12, e0180442. doi: 10.1371/journal.pone.0180442. CrossRef. Google Scholar.
64. Malgioglio G., Rizzo G. F., Nigro S., Lefebvre du Prey V., Herforth-Rahmé J., Catara V., et al. (2022). Plant-microbe interaction in sustainable agriculture: the factors that may influence the efficacy of PGPM application. Sustainability 14, 2253. doi: 10.3390/su14042253. CrossRef. Google Scholar.
65. Mangiafico S. S. (2023). rcompanion: Functions to Support Extension Education Program Evaluation. Available at: https://CRAN.R-project.org/package=rcompanion/. Google Scholar.
66. Martin M. (2011). Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 17, 10. doi: 10.14806/ej.17.1.200. CrossRef. Google Scholar.
67. Martinez A. P. (2022). pairwiseAdonis: Pairwise Multilevel Comparison Using Adonis. Available at: https://github.com/pmartinezarbizu/pairwiseAdonis. Google Scholar.
68. Mawarda P. C., Lakke S. L., Dirk van Elsas J., Salles J. F. (2022). Temporal dynamics of the soil bacterial community following Bacillus invasion. iScience 25, 104185. doi: 10.1016/j.isci.2022.104185. CrossRef. Google Scholar.
69. Mazzola M., Freilich S. (2017). Prospects for biological soilborne disease control: application of indigenous versus synthetic microbiomes. Phytopathology® 107, 256–263. doi: 10.1094/PHYTO-09-16-0330-RVW. CrossRef. Google Scholar.
70. McMurdie P. J., Holmes S. (2013). phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PloS One 8, e61217. doi: 10.1371/journal.pone.0061217. CrossRef. Google Scholar.
71. Mendes R., Garbeva P., Raaijmakers J. M. (2013). The rhizosphere microbiome: significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 37, 634–663. doi: 10.1111/1574-6976.12028. CrossRef. Google Scholar.
72. Moradtalab N., Ahmed A., Geistlinger J., Walker F., Höglinger B., Ludewig U., et al. (2020). Synergisms of microbial consortia, N forms, and micronutrients alleviate oxidative damage and stimulate hormonal cold stress adaptations in maize. Front. Plant Sci. 11. doi: 10.3389/fpls.2020.00396. CrossRef. Google Scholar.
73. Morais M. C., Mucha Â., Ferreira H., Gonçalves B., Bacelar E., Marques G. (2019). Comparative study of plant growth-promoting bacteria on the physiology, growth and fruit quality of strawberry. J. Sci. Food Agric. 99, 5341–5349. doi: 10.1002/jsfa.9773. CrossRef. Google Scholar.
74. Mpanga I., Dapaah H., Geistlinger J., Ludewig U., Neumann G. (2018). Soil type-dependent interactions of P-solubilizing microorganisms with organic and inorganic fertilizers mediate plant growth promotion in tomato. Agronomy 8, 213. doi: 10.3390/agronomy8100213. CrossRef. Google Scholar.
75. Mpanga I., Nkebiwe P., Kuhlmann M., Cozzolino V., Piccolo A., Geistlinger J., et al. (2019). The form of N supply determines plant growth promotion by P-solubilizing microorganisms in maize. Microorganisms 7, 38. doi: 10.3390/microorganisms7020038. CrossRef. Google Scholar.
76. Müller M., Wickham H. (2022). tibble: Simple Data Frames. Available at: https://CRAN.R-project.org/package=tibble. Google Scholar.
77. Mumtaz M. Z., Ahmad M., Jamil M., Hussain T. (2017). Zinc solubilizing Bacillus spp. potential candidates for biofortification in maize. Microbiol. Res. 202, 51–60. doi: 10.1016/j.micres.2017.06.001. CrossRef. Google Scholar.
78. Nautiyal C. S. (1999). An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 170, 265–270. doi: 10.1111/j.1574-6968.1999.tb13383.x. CrossRef. Google Scholar.
79. Oksanen J., Simpson G. L., Blanchet F. G., Kindt R., Legendre P., Minchin P. R., et al. (2022). vegan: Community Ecology Package. Available at: https://CRAN.R-project.org/package=vegan. Google Scholar.
80. Paradis E., Schliep K. (2019). ape 5.0: an environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 35, 526–528. doi: 10.1093/bioinformatics/bty633. CrossRef. Google Scholar.
81. Parnell J. J., Berka R., Young H. A., Sturino J. M., Kang Y., Barnhart D. M., et al. (2016). From the lab to the farm: an industrial perspective of plant beneficial microorganisms. Front. Plant Sci. 7. doi: 10.3389/fpls.2016.01110. CrossRef. Google Scholar.
82. Pascale A., Proietti S., Pantelides I. S., Stringlis I. A. (2020). Modulation of the root microbiome by plant molecules: the basis for targeted disease suppression and plant growth promotion. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.01741. CrossRef. Google Scholar.
83. Quast C., Pruesse E., Yilmaz P., Gerken J., Schweer T., Yarza P., et al. (2012). The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 41, D590–D596. doi: 10.1093/nar/gks1219. CrossRef. Google Scholar.
84. Rai S., Kashyap P. L., Kumar S., Srivastava A. K., Ramteke P. W. (2016). Identification, characterization and phylogenetic analysis of antifungal Trichoderma from tomato rhizosphere. SpringerPlus 5, 1939. doi: 10.1186/s40064-016-3657-4. CrossRef. Google Scholar.
85. Raymond N. S., Gómez-Muñoz B., van der Bom F. J. T., Nybroe O., Jensen L. S., Müller-Stöver D. S., et al. (2021). Phosphate-solubilising microorganisms for improved crop productivity: a critical assessment. New Phytol. 229, 1268–1277. doi: 10.1111/nph.16924. CrossRef. Google Scholar.
86. R Core Team (2022). R: A Language and Environment for Statistical Computing. Available at: https://www.R-project.org/. Google Scholar.
87. Sánchez-Cañizares C., Jorrín B., Poole P. S., Tkacz A. (2017). Understanding the holobiont: the interdependence of plants and their microbiome. Curr. Opin. Microbiol. 38, 188–196. doi: 10.1016/j.mib.2017.07.001. CrossRef. Google Scholar.
88. Sanket A. S., Ghosh S., Sahoo R., Nayak S., Das A. P. (2017). Molecular identification of acidophilic manganese (Mn)-solubilizing bacteria from mining effluents and their application in mineral beneficiation. Geomicrobiol. J. 34, 71–80. doi: 10.1080/01490451.2016.1141340. CrossRef. Google Scholar.
89. Santoyo G., Guzmán-Guzmán P., Parra-Cota F. I., de los Santos-Villalobos S., del Orozco-Mosqueda, Ma. C., Glick B. R. (2021). Plant growth stimulation by microbial consortia. Agronomy 11, 219. doi: 10.3390/agronomy11020219. CrossRef. Google Scholar.
90. Schierstaedt J., Jechalke S., Nesme J., Neuhaus K., Sørensen S. J., Grosch R., et al. (2020). Salmonella persistence in soil depends on reciprocal interactions with indigenous microorganisms. Environ. Microbiol. 22, 2639–2652. doi: 10.1111/1462-2920.14972. CrossRef. Google Scholar.
91. Schrama M., de Haan J. J., Kroonen M., Verstegen H., van der Putten W. H. (2018). Crop yield gap and stability in organic and conventional farming systems. Agric. Ecosyst. Environ. 256, 123–130. doi: 10.1016/j.agee.2017.12.023. CrossRef. Google Scholar.
92. Schreiter S., Babin D., Smalla K., Grosch R. (2018). Rhizosphere Competence and Biocontrol Effect of Pseudomonas sp. RU47 Independent from Plant Species and Soil Type at the Field Scale. Front. Microbiol. 9. doi: 10.3389/fmicb.2018.00097. CrossRef. Google Scholar.
93. Schreiter S., Sandmann M., Smalla K., Grosch R. (2014). Soil type dependent rhizosphere competence and biocontrol of two bacterial inoculant strains and their effects on the rhizosphere microbial community of field-grown lettuce. PloS One 9, e103726. doi: 10.1371/journal.pone.0103726. CrossRef. Google Scholar.
94. Schütz L., Gattinger A., Meier M., Müller A., Boller T., Mäder P., et al. (2018). Improving crop yield and nutrient use efficiency via biofertilization—A global meta-analysis. Front. Plant Sci. 8. doi: 10.3389/fpls.2017.02204. CrossRef. Google Scholar.
95. Schwyn B., Neilands J. B. (1987). Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 160, 47–56. doi: 10.1016/0003-2697(87)90612-9. CrossRef. Google Scholar.
96. Sella S. R. B. R., Vandenberghe L. P. S., Soccol C. R. (2014). Life cycle and spore resistance of spore-forming Bacillus atrophaeus. Microbiol. Res. 169, 931–939. doi: 10.1016/j.micres.2014.05.001. CrossRef. Google Scholar.
97. Seufert V., Ramankutty N., Foley J. A. (2012). Comparing the yields of organic and conventional agriculture. Nature 485, 229–232. doi: 10.1038/nature11069. CrossRef. Google Scholar.
98. Sharma S., Compant S., Ballhausen M.-B., Ruppel S., Franken P. (2020). The interaction between Rhizoglomus irregulare and hyphae attached phosphate solubilizing bacteria increases plant biomass of Solanum lycopersicum. Microbiol. Res. 240, 126556. doi: 10.1016/j.micres.2020.126556. CrossRef. Google Scholar.
99. Sommermann L., Babin D., Behr J. H., Chowdhury S. P., Sandmann M., Windisch S., et al. (2022). Long-term fertilization strategy impacts Rhizoctonia solani--microbe interactions in soil and rhizosphere and defense responses in lettuce. Microorganisms 10, 1717. doi: 10.3390/microorganisms10091717. CrossRef. Google Scholar.
100. Sun X., Xu Z., Xie J., Hesselberg-Thomsen V., Tan T., Zheng D., et al. (2022). Bacillus velezensis stimulates resident rhizosphere Pseudomonas stutzeri for plant health through metabolic interactions. ISME J. 16, 774–787. doi: 10.1038/s41396-021-01125-3. CrossRef. Google Scholar.
101. Sundberg C., Al-Soud W. A., Larsson M., Alm E., Yekta S. S., Svensson B. H., et al. (2013). 454 pyrosequencing analyses of bacterial and archaeal richness in 21 full-scale biogas digesters. FEMS Microbiol. Ecol. 85, 612–626. doi: 10.1111/1574-6941.12148. CrossRef. Google Scholar.
102. Thonar C., Lekfeldt J. D. S., Cozzolino V., Kundel D., Kulhánek M., Mosimann C., et al. (2017). Potential of three microbial bio-effectors to promote maize growth and nutrient acquisition from alternative phosphorous fertilizers in contrasting soils. Chem. Biol. Technol. Agric. 4, 7. doi: 10.1186/s40538-017-0088-6. CrossRef. Google Scholar.
103. Topçuoğlu B. D., Lesniak N. A., Ruffin M. T., Wiens J., Schloss P. D. (2020). A framework for effective application of machine learning to microbiome-based classification problems. mBio 11, e00434–e00420. doi: 10.1128/mBio.00434-20. CrossRef. Google Scholar.
104. Tosi M., Mitter E. K., Gaiero J., Dunfield K. (2020). It takes three to tango: the importance of microbes, host plant, and soil management to elucidate manipulation strategies for the plant microbiome. Can. J. Microbiol. 66, 413–433. doi: 10.1139/cjm-2020-0085. CrossRef. Google Scholar.
105. Usero F. M., Armas C., Morillo J. A., Gallardo M., Thompson R. B., Pugnaire F. I. (2021). Effects of soil microbial communities associated to different soil fertilization practices on tomato growth in intensive greenhouse agriculture. Appl. Soil Ecol. 162, 103896. doi: 10.1016/j.apsoil.2021.103896. CrossRef. Google Scholar.
106. van Elsas J. D., Chiurazzi M., Mallon C. A., Elhottovā D., Krištůfek V., Salles J. F. (2012). Microbial diversity determines the invasion of soil by a bacterial pathogen. Proc. Natl. Acad. Sci. U.S.A. 109, 1159–1164. doi: 10.1073/pnas.1109326109. CrossRef. Google Scholar.
107. Verband Deutscher Landwirtschaftlicher Untersuchungs- und Forschungsanstalten (2011). Umweltanalytik. 4th ed. Ed. Janßen E. Darmstadt: VDLUFA-Verlag. Google Scholar.
108. Vlot A. C., Sales J. H., Lenk M., Bauer K., Brambilla A., Sommer A., et al. (2021). Systemic propagation of immunity in plants. New Phytol. 229, 1234–1250. doi: 10.1111/nph.16953. CrossRef. Google Scholar.
109. Watteau F., Villemin G. (2018). Soil microstructures examined through transmission electron microscopy reveal soil-microorganisms interactions. Front. Environ. Sci. 6. doi: 10.3389/fenvs.2018.00106. CrossRef. Google Scholar.
110. Wei Z., Friman V.-P., Pommier T., Geisen S., Jousset A., Shen Q. (2020). Rhizosphere immunity: targeting the underground for sustainable plant health management. Front. Agr. Sci. Eng. 7, 317. doi: 10.15302/J-FASE-2020346. CrossRef. Google Scholar.
111. Weinert N., Meincke R., Gottwald C., Heuer H., Schloter M., Berg G., et al. (2010). Bacterial diversity on the surface of potato tubers in soil and the influence of the plant genotype. FEMS Microbiol. Ecol. 74, 114–123. doi: 10.1111/j.1574-6941.2010.00936.x. CrossRef. Google Scholar.
112. Wickham H. (2007). Reshaping data with the reshape package. J. Stat. Software 21, 1–20. doi: 10.18637/jss.v021.i12. CrossRef. Google Scholar.
113. Wickham H. (2016). ggplot2: Elegant Graphics for Data Analysis. 2nd ed. Cham: Springer International Publishing: Imprint: Springer. doi: 10.1007/978-3-319-24277-4. CrossRef. Google Scholar.
114. Wickham H., Averick M., Bryan J., Chang W., McGowan L., François R., et al. (2019). Welcome to the tidyverse. JOSS 4, 1686. doi: 10.21105/joss.01686. CrossRef. Google Scholar.
115. Wickham H., François R., Henry L., Müller K., Vaughan D. (2023). dplyr: A Grammar of Data Manipulation. Available at: https://dplyr.tidyverse.org. Google Scholar.
116. Williams J., Clarkson J. M., Mills P. R., Cooper R. M. (2003). A Selective Medium for Quantitative Reisolation of Trichoderma harzianum from Agaricus bisporus Compost. Appl. Environ. Microbiol. 69, 4190–4191. doi: 10.1128/AEM.69.7.4190-4191.2003. CrossRef. Google Scholar.
117. Windisch S., Bott S., Ohler M.-A., Mock H.-P., Lippmann R., Grosch R., et al. (2017). Rhizoctonia solani and bacterial inoculants stimulate root exudation of antifungal compounds in lettuce in a soil-type specific manner. Agronomy 7, 44. doi: 10.3390/agronomy7020044. CrossRef. Google Scholar.
118. Windisch S., Sommermann L., Babin D., Chowdhury S. P., Grosch R., Moradtalab N., et al. (2021). Impact of long-term organic and mineral fertilization on rhizosphere metabolites, root--microbial interactions and plant health of lettuce. Front. Microbiol. 11. doi: 10.3389/fmicb.2020.597745. CrossRef. Google Scholar.
119. Zubair M., Hanif A., Farzand A., Sheikh T. M. M., Khan A. R., Suleman M., et al. (2019). Genetic screening and expression analysis of psychrophilic bacillus spp. Reveal their potential to alleviate cold stress and modulate phytohormones in wheat. Microorganisms 7, 337. doi: 10.3390/microorganisms7090337. CrossRef. Google Scholar.

Behr JH, Kampouris ID, Babin D, Sommermann L, Francioli D, Kuhl-Nagel T, Chowdhury SP, Geistlinger J, Smalla K, Neumann G and Grosch R (2023) Beneficial microbial consortium improves winter rye performance by modulating bacterial communities in the rhizosphere and enhancing plant nutrient acquisition. Front. Plant Sci. 14:1232288. doi: 10.3389/fpls.2023.1232288
Перевод статьи «Beneficial microbial consortium improves winter rye performance by modulating bacterial communities in the rhizosphere and enhancing plant nutrient acquisition» авторов Behr JH, Kampouris ID, Babin D, Sommermann L, Francioli D, Kuhl-Nagel T, Chowdhury SP, Geistlinger J, Smalla K, Neumann G and Grosch R., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык
Фото: Designed by Freepik

















































































































Комментарии (0)