Что показал анализ ДНК: скрытые патогены в системах гидропоники и аквапоники
Гидропонное и аквапонное земледелие становятся всё более популярными как решение проблемы глобальной продовольственной безопасности. Растения в гидропонных системах выращиваются без почвы в контролируемых условиях и считаются менее опасными с точки зрения безопасности пищевых продуктов по сравнению с традиционным полевым земледелием. Однако гидропоника и аквапоника могут иметь совершенно иные источники микробиологических рисков для безопасности пищевых продуктов, которые остаются недостаточно изученными.
Аннотация
В данном исследовании мы изучили микробиомы, микробиологические опасности и потенциальные пути передачи бактерий внутри двух коммерческих гидропонных и аквапонных фермерских систем, используя секвенирование 16S-ITS-23S рРНК и опрос по практике обеспечения безопасности гидропонных продуктов питания. Микробиом гидропонной фермерской системы был проанализирован на образцах свежей продукции, питательного раствора, инструментов и работников ферм. Proteobacteria, Actinobacteria, Cyanobacteria, Bacteroidetes и Firmicutes были основными компонентами гидропонных/аквапонных фермерских систем, при этом Pseudomonas был наиболее распространённым родом в образцах свежей продукции. Мы также выявили наличие множества бактерий порчи и потенциальных патогенов человека, растений и рыб на подвидовом уровне. Бактерии порчи Pseudomonas spp. и Clostridium spp. были обильны, соответственно, на гидропонной ферме микрозелени и на аквапонной ферме салата. Кроме того, мы продемонстрировали картирование прочтений последовательностей 16S-ITS-23S рРНК Escherichia coli (длиной около 2500 п.н.) с базами данных рРНК малой или большой субъединиц и базами данных целых геномов для подтверждения патогенности и показали потенциал использования секвенирования 16S-ITS-23S рРНК для идентификации патогенов. С помощью SourceTracker и перекрывающихся вариантов ампликонных последовательностей мы предсказали двунаправленный путь передачи между растениями и окружающей средой и построили карту передачи бактерий, которая может быть использована в будущих планах контроля рисков для безопасности пищевых продуктов.
Введение
Гидропонные системы возделывания (HCS) выращивают растения в смеси воды и питательных растворов в замкнутой и контролируемой среде (Riggio и др., 2019). Аквапоника представляет собой форму HCS, в которой вода из рыбоводного бассейна используется в качестве источника питания для растений. По сравнению с традиционными фермами на почве, HCS не ограничена климатом или местоположением, лучше использует вертикальное пространство, экономит примерно 90% оросительной воды и может круглогодично снабжать фруктами и овощами окружающие сообщества (Pegasus Agriculture Group, 2017). HCS рассматривается как перспективное решение для обеспечения продовольствием растущего населения планеты. Стоимость мирового рынка гидропоники в 2016 году оценивалась в 6,63 миллиарда долларов, а к 2025 году прогнозируется её рост до 12,1 миллиарда долларов (FMR, 2019).
Считается, что гидропонные системы возделывания имеют меньше проблем с безопасностью пищевых продуктов, поскольку они исключают обычные микробные контаминанты полей, поступающие из почвы, поверхностных вод, диких и сельскохозяйственных животных и вредителей. Они также свободны от химических загрязнителей, таких как пестициды и почвенные удобрения (Center for Disease Control and Prevention [CDC], 2019; Morgan, 2019). Однако имеется несколько зарегистрированных вспышек пищевых заболеваний и отзывов продукции, связанных со свежей продукцией, выращенной в HCS (Center for Disease Control and Prevention [CDC], 2014, 2016, 2020). Правила внедрения, разработанные Управлением по санитарному надзору за качеством пищевых продуктов и медикаментов США в ответ на Закон о модернизации безопасности пищевых продуктов, требуют от фермеров соблюдения планов безопасности пищевых продуктов и анализа опасностей, а также периодического проведения аудитов безопасности пищевых продуктов (U.S. Food and Drug Administration, 2011). Однако мелкие, местные фермы и фермы прямого сбыта, такие как большинство гидропонных/аквапонных ферм, были исключены из этих правил (Deering, 2018). Таким образом, существует настоятельная необходимость в анализе опасностей и рисков для безопасности пищевых продуктов на этих фермах (Colorado Department of Public Health and Environment, 2012; Deering, 2018).
Ороско и Итурриага (2008) обнаружили Salmonella и Escherichia coli на гидропонной томатной ферме и связали это с наводнением и проникновением диких птиц в помещение. Лопес-Гальвес и др. (2014) сообщили об обнаружении Salmonella spp., Listeria spp. и E. coli на гидропонной ферме, орошаемой очищенной и поверхностной водой. Также велись дискуссии о том, что патогены, которые могут присутствовать в фекалиях рыб, могут загрязнять свежую продукцию, выращиваемую в аквапонных системах (Bergšpica и др., 2020; Sawyer и др., 2020). Однако предыдущие разрозненные исследования лишь сообщали о случаях выявления патогенов в HCS, и не предпринималось попыток систематически проанализировать источник и передачу опасностей.
Микробиомы в экосистеме могут дать представление о потенциальных источниках загрязнения и путях передачи (De Filippis и др., 2021). Секвенирование 16S рРНК с методом классификации операционных таксономических единиц (OTU) в качестве основного подхода (Westcott and Schloss, 2015; Kopylova и др., 2016; Ranjan и др., 2016) использовалось для характеристики микробиомов, связанных с сельскохозяйственными и пищевыми системами, включая почву, поверхности свежей продукции, кухонную среду и образцы рук человека (Fierer и др., 2008; Bibby и др., 2010; Flores и др., 2013; Leff and Fierer, 2013). Полноразмерное секвенирование 16S рРНК позволяет различать бактерии на видовом уровне, однако оно недостаточно для идентификации опасностей на подвидовом уровне, например, для различения тесно связанных патогенных и непатогенных E. coli (Srinivasan и др., 2015; Miao и др., 2017; Callahan и др., 2019; Deguenon и др., 2019; Numberger и др., 2019). Для выявления патогенных опасностей исследователи обычно нацеливаются на распространённые патогенные бактерии или специфические гены вирулентности с использованием селективного культивирования или мультиплексной ПЦР в реальном времени; однако добавление праймеров на множество целевых генов является дорогостоящим, и целевые патогены могут не присутствовать в тестируемых образцах (Lopez-Galvez и др., 2014; Liu и др., 2019; Dankwa и др., 2020; Zhong и др., 2020).
Для повышения разрешающей способности и достоверности таксономической классификации был предложен новый подход к секвенированию ампликонов, при котором секвенируются полноразмерная 16S рРНК, внутренний транскрибируемый спейсер (ITS) и частичная 23S рРНК (16S-ITS-23S рРНК), что даёт длинный прочт длиной около 2500 п.н. (Martijn и др., 2019; Graf и др., 2021; Kinoshita и др., 2021). Этот метод секвенирования может выполняться на системах PacBio по стоимости, аналогичной полноразмерному секвенированию 16S рРНК. Также была разработана новая концепция вариантов ампликонных последовательностей (ASV), заменившая кластеризацию OTU и повысившая разрешающую способность классификации (Callahan и др., 2016, 2017). Этот подход уже использовался для различения тесно связанных штаммов Klebsiella, E. coli и Enterobacter в фекалиях младенцев (Graf и др., 2021).
С другой стороны, опрос по практике обеспечения безопасности пищевых продуктов использовался как инструмент оценки рисков для выявления потенциальных опасностей на ферме (Soon и др., 2013; Ilic и др., 2017). Исследователи пытались объединить данные опросов и отбора проб из разных исследований для оценки микробной безопасности пищевых продуктов и рисков для общественного здоровья (Allende и др., 2017; Barragán и др., 2021). Таким образом, сочетание опроса с отбором проб микробиома может обеспечить лучшее понимание количественных микробных измерений для разработки стратегий снижения рисков для безопасности пищевых продуктов.
Это исследование было проведено с целью изучения микробиомов растений и окружающей среды в гидропонных экосистемах, а также влияния методов ведения хозяйства на микробиомы с использованием комбинации опроса по практике безопасности пищевых продуктов и высокопроизводительного анализа микробиома на основе секвенирования 16S-ITS-23S. Мы сравнили количество аэробных бактерий, а также состав и разнообразие микробиомов коммерческих гидропонных/аквапонных ферм с лабораторными контрольными системами. Кроме того, мы провели скрининг микробиомов на наличие микробных опасностей, включая патогены растений, человека и рыб, а также микроорганизмы порчи, и предложили потенциальные пути передачи бактерий внутри HCS. Вместе с результатами опроса мы продемонстрировали влияние методов ведения хозяйства на микробиомы окружающей среды и растений, а также предоставили индивидуальные или целевые стратегии улучшения для ферм.
Материалы и методы
Дизайн исследования и набор участников
Набор участников и отбор проб проводились с марта по август 2020 года. Поскольку план опроса и отбора проб включал участие людей, было получено одобрение Институционального наблюдательного совета (IRB) для исследований с участием человека от Университета Иллинойса, Управления по защите субъектов исследований (протокол IRB: 20653).
Всего в Иллинойсе было идентифицировано 12 гидропонных ферм с помощью поиска на Google Maps и в сельскохозяйственных информационных бюллетенях, с фермами связались по телефону или электронной почте. В этом исследовании приняли участие две фермы: вертикальная гидропонная ферма по выращиванию микрозелени (ферма H) и аквапонная ферма, выращивающая салат и тилапию совместно (ферма A). Схема гидропонной системы и процедуры эксплуатации представлены на дополнительном рисунке 1. На ферме A семена высевали в стартовые лотки для рассады до развития всходов, затем переносили в систему затопления-дренажа до формирования розетки молодого салата, после чего розетки переносили в систему глубоководного культивирования (DWC) для созревания. Вода поступала из бассейнов с тилапией, сначала подавалась в систему DWC, затем в систему затопления-дренажа и стартовые лотки для рассады. В системе DWC выращивали три сорта салата в порядке: HoneyCrisp, Green Oak Leaf и GreenCrisp от входа к выходу. На ферме H использовались две раздельные системы затопления-дренажа и пятиуровневые вертикальные конструкции. В первой системе выращивали пять видов микрозелени, включая гранат, редис, брокколи, кинзу и брюссельскую капусту. Во второй системе выращивали только микрозелень капусты кале.
План отбора проб
У двух ферм было запрошено согласие на план отбора проб, посещение фермы и опрос. Образцы свежей продукции были собраны с обеих ферм. Образцы отбирались непосредственно с лотков для выращивания с использованием стерильных перчаток и ножниц и взвешивались в стерильные пакеты для образцов (Whirl-Pak, Madison, WI, США). На ферме H отбирали образцы микрозелени редиса, граната, брокколи, брюссельской капусты, кинзы и капусты кале путём случайного сбора с 10 различных лотков для выращивания до общего веса 25 г каждый. Мы также выращивали микрозелень брокколи, редиса и капусты кале гидропонно в лабораторной системе (L1) в качестве контроля. На ферме A случайным образом отбирали три сорта салата на стадии полного созревания (по три кочана каждого сорта) из системы DWC. Для сравнения были приобретены четыре аналогичных гидропонно выращенных сорта салата в продуктовом магазине в Шампейне, Иллинойс. Мы выращивали салат ромэн и дуболистный салат в системе затопления-дренажа и в меньшей гидропонной системе DWC в лаборатории (L2) в качестве контроля.
Образцы питательного раствора (по 250 мл) отбирались с обеих ферм непосредственно в стерильные пакеты для образцов. На ферме H мы отобрали по два образца питательного раствора из каждой системы затопления-дренажа в резервуарном баке и в трубопроводной системе под лотками с отобранной микрозеленью соответственно. Три образца питательного раствора были взяты из системы DWC фермы A у входа воды из рыбоводного бассейна, в средней точке системы возле корней салата и в конце системы возле корней салата.
Мазки с поверхностей окружающей среды были получены с согласованных объектов с использованием стерильных ватных тампонов. Тампоны предварительно замачивали в пептонно-солевой воде на 1 минуту, и участки размером 3×3 см тщательно протирались всей поверхностью ватного тампона. Три образца мазков были получены на ферме H: с рук работника, подошв обуви и санитарно-обработанных лотков для выращивания. Три образца мазков были получены на ферме A: мазки с лотков для выращивания из системы DWC (trayM), системы затопления-дренажа (trayB) и мазок с подошвы обуви работника фермы.
Все собранные образцы были доставлены в микробиологическую лабораторию Университета Иллинойса в портативном холодильнике в тот же день. Образцы, собранные для анализа микробиома, перечислены в дополнительной таблице 1.
Дизайн опроса
Был разработан опрос из пяти разделов, посвящённых текущим методам обеспечения безопасности пищевых продуктов на ферме, включая здоровье и гигиену работников, осведомлённость о безопасности пищевых продуктов, обработку оросительной воды, санитарию оборудования и процедуры обращения с продукцией (дополнительная таблица 2). Вопросы были собраны из доступных опросов по безопасности пищевых продуктов на фермах с изменениями, отражающими производственные практики гидропонных систем (Harrison и др., 2013; Ilic и др., 2017; FamilyFarmed, 2019). Для обеспечения конфиденциальности каждой ферме был присвоен код для связи с данными отбора проб. Результаты опроса были преобразованы в числовые баллы для расчёта общих баллов по каждому разделу.
Определение микробной нагрузки
Определение количества аэробных микроорганизмов проводилось для образцов свежей продукции и питательного раствора. Образцы свежей продукции массой 25 г смешивали с 225 мл фосфатно-солевого буфера в стерильных фильтруемых пакетах для образцов. После гомогенизации в течение 2 минут в стомачере и серийного разведения, по 100 мкл фильтратов образцов свежей продукции или питательного раствора высевали на агар для подсчёта колоний (BD Difco, Franklin Lakes, NJ, США). Планшеты инкубировали при 37 ± 2°C в течение 24 часов и подсчитывали колониеобразующие единицы (КОЕ). Оставшиеся фильтраты и образцы питательного раствора фильтровали с использованием вакуумных фильтров с порами 0,2 мкм объёмом 250 мл (Foxx Life Sciences, Pittsburgh, PA, США) для сбора микроорганизмов. Образцы мазков и фильтры хранились при –20°C не более 2 недель до молекулярного анализа.
Выделение ДНК и секвенирование гена 16S-ITS-23S рРНК
Для профилирования микробиома бактериальная ДНК была экстрагирована из мазков и фильтров с использованием набора DNeasy PowerSoil Pro Kit (QIAGEN, Hilden, Германия) в соответствии с инструкциями производителя. Положительный контроль с 12 известными бактериальными штаммами экстрагировался вместе с обычными образцами. Качество ДНК оценивали с помощью гель-электрофореза, а концентрацию определяли с использованием набора Qubit dsDNA HS Assay Kit и флуориметра Qubit 2.0 (Invitrogen, Thermo Fisher Scientific, Орегон, США). Экстрагированные образцы ДНК хранились при –20°C до дальнейшего анализа.
Для секвенирования ампликона, охватывающего полноразмерные гены 16S, ITS и частичной 23S рРНК, использовался набор Wave StrainID Kit (Shoreline Biome, Farmington, CT, США). Процедуры ранее были описаны Графом и др. (2021). Библиотеки ампликонов были созданы с использованием набора SMRTbell express template prep kit 2.0 (каталожный номер 100-938-900; Pacific Biosciences, Menlo Park, CA, США) в соответствии с инструкциями производителя. Библиотека была секвенирована на 1 SMRTBell 8M на системе PacBio Sequel II (Pacific Biosciences) с использованием режима циркулярного консенсусного (CCS) секвенирования в Центре биотехнологий имени Роя Дж. Карвера Университета Иллинойса, Урбана, Иллинойс, США. Циркулярные консенсусные прочтения (ccs) определялись с минимальной прогнозируемой точностью 0,999 и минимальным количеством проходов, установленным на три, в программном пакете SMRT Link 5.1 (Pacific Biosciences). Всего было получено 826 974 прочтений ccs со средней длиной прочтения 2421 п.н. с использованием настроек по умолчанию.
Обработка последовательностей и таксономическое присвоение
Прочтения ccs были дополнительно обработаны с использованием SBanalyzer 2.4 (Shoreline Biome) в соответствии с рабочим процессом, описанным Графом и др. (2021). Вкратце, все прочтения были отсортированы в файлы FASTQ по образцам без обрезания и классифицированы путём картирования на базу данных Athena (Shoreline Biome). Таксономическое присвоение выполнялось при идентичности >97% для штаммового уровня и >95% для видового уровня по сравнению с эталонной последовательностью 16S-ITS-23S рРНК в базе данных Athena. После демультиплексирования прочтения ccs были дополнительно обработаны с помощью DADA2 (версия 1.9.1) для получения ампликонов с однонуклеотидным разрешением (Callahan и др., 2019). ASV были присвоены восемь таксономических уровней: царство, тип, класс, отряд, семейство, род, вид и подвид. Таксономический идентификатор и соответствующее количество прочтений для всех образцов были созданы в итоге, что дало в общей сложности 1 939 ASV. Кроме того, ASV-контаминанты были обнаружены и удалены с помощью R-пакета "decontam" с использованием выявления контаминантов на основе распространённости с пороговым значением 0,5 (Davis и др., 2018). После дезаминирования осталось 1 096 ASV.
Анализ микробного сообщества
Анализ микробного сообщества проводился с использованием пакетов "Bioconductor", "microbiome", "phyloseq" и "vegan" и визуализировался с помощью ggplot2 в R (Gentleman и др., 2004; McMurdie and Holmes, 2013; Oksanen и др., 2020; Wickham; Lahti and Shetty). Набор данных был композиционно трансформирован для визуализации составов на уровне типов и 50 наиболее распространённых ASV родов. Нетрансформированный набор данных использовался для анализа разнообразия сообществ. Были рассчитаны индексы альфа-разнообразия "Chao1", "Shannon" и "Simpson". Нормальность распределения отдельных индексов альфа-разнообразия проверялась с помощью теста Шапиро–Уилка. Данные оценивались на однородность дисперсии с использованием теста Ливена. Статистическая значимость альфа-разнообразия оценивалась с использованием дисперсионного анализа повторных измерений с последующим тестом Тьюки. Альфа-разнообразия также сравнивались по типам образцов или местам отбора с использованием парного t-теста с процедурой Бенджамини–Хохберга для снижения частоты ложных обнаружений. Бета-разнообразие визуализировалось с помощью графиков анализа главных координат (PCoA) на основе взвешенных и невзвешенных филогенетических расстояний UniFrac, а также на основе подсчётов расстояния Брея–Кёртиса. Анализ перестановочной многомерной дисперсии (PERMANOVA) проводился с использованием функции "adonis" для измерения различий в индексах бета-разнообразия по месту или типу отбора (Anderson, 2017).
Идентификация патогенов и видов порчи
Наличие патогенов во всех образцах проверялось по базе данных патогенов NCBI (NCBI, 2021). Был проведён скрининг видов бактерий порчи, ассоциированных со свежей продукцией (Kaczmarek и др., 2019). ASV порчи были сгруппированы по виду или роду. Прочтения были композиционно преобразованы в процентные значения и визуализированы в виде тепловых карт с использованием функции "pheatmap"¹.
ASV1628 была отмечена как "E. coli unclassified" и содержит 38 прочтений последовательностей. Мы использовали её в качестве примера для дальнейшего изучения потенциальной патогенности. 38 исходных последовательностей были картированы на базы данных SILVA с помощью EMBL-EBI/ENA, GTDB, RDP, LTP таксономии, используя сервисы SILVA Alignment, Classification, and Tree Services (ACT) для малой (16S) и большой (23S) субъединиц соответственно (Leinonen и др., 2011; Pruesse и др., 2012; Quast и др., 2013; Cole и др., 2014; Yilmaz и др., 2014; Chaumeil и др., 2020). 38 исходных последовательностей также были картированы на полно-геномные последовательности бактерий в коллекции нуклеотидов NCBI (nt) с использованием Basic Local Alignment Search Tool (BLASTn; Zhang и др., 2000; Morgulis и др., 2008). Для каждой исходной последовательности были извлечены выровненные последовательности 10 наиболее близкородственных штаммов с наибольшей идентичностью и построены быстрые минимальные эволюционные деревья (Rzhetsky and Nei, 1993). Близкородственные патогенные штаммы были отобраны и выровнены с исходными последовательностями с использованием ClustalW в Jalview 2 (Версия 2.11.1.4; Larkin и др., 2007; Waterhouse и др., 2009). После выравнивания расстояния Джукса–Кантора между исходными последовательностями и извлечёнными последовательностями были рассчитаны с использованием MegaX с параметром гамма = 1 (Jukes and Cantor, 1969; Stecher и др., 2020). Потенциальными патогенами считаются те, которые находятся в пределах расстояния Джукса–Кантора 0,03 от известной последовательности патогена (Ibekwe и др., 2013).
SourceTracker и общие варианты ампликонных последовательностей
Отслеживание источника микроорганизмов проводилось с использованием R-пакета SourceTracker (версия 1.0.1) с параметрами по умолчанию (Knights и др., 2011). Глубина секвенирования 1000 последовательностей/образец была выбрана в соответствии с Цвирзицом и др. (2020), которые показали, что 1000 достаточно для получения сопоставимого результата с наборами данных с большей глубиной секвенирования. Образцы питательного раствора и окружающей среды были назначены источниками, а образцы свежей продукции – приемниками. Результаты визуализированы с помощью диаграммы Санкея². Общие ASV между образцами из разных систем визуализированы с помощью диаграммы Венна³.
Результаты
Количество бактериальных клеток из разных мест и типов образцов
Общая микробная нагрузка была определена с помощью подсчёта аэробных колоний. Микрозелень с гидропонной фермы (ферма H) и лабораторной системы (L1) показала сходную бактериальную нагрузку в диапазоне от 7,3 до 8,6 log КОЕ/г (Рисунок 1A). Образцы салата с аквапонной фермы (ферма A) имели значительно более низкие показатели бактерий (3,7–4,0 log КОЕ/г), чем образцы салата из продуктового магазина (G, Рисунок 1B).
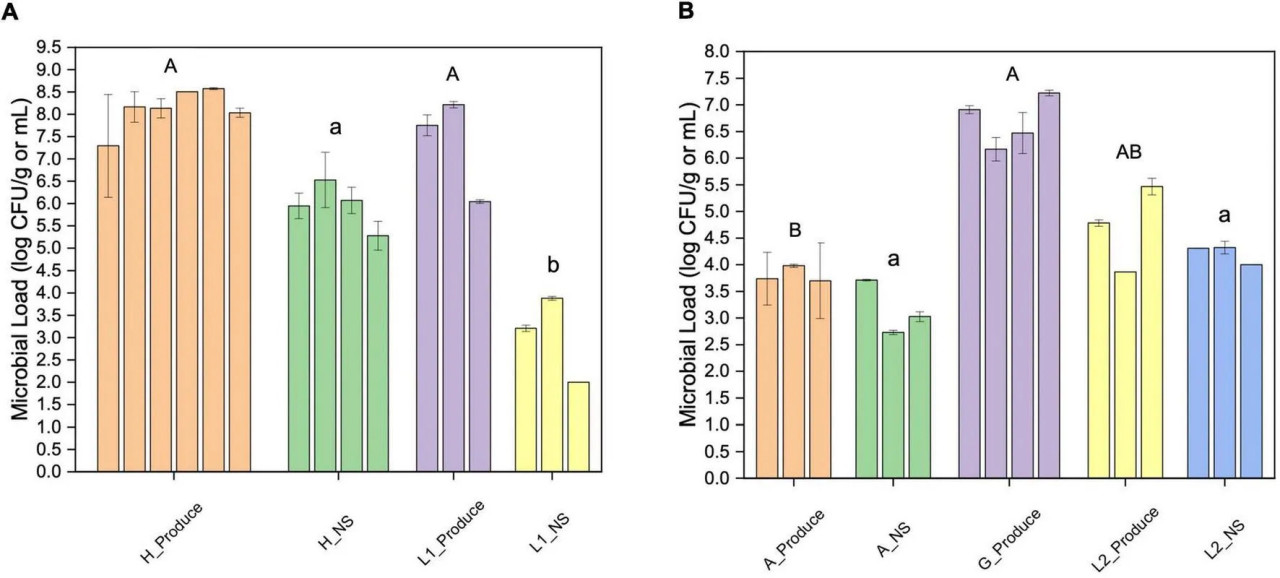
РИСУНОК 1 Подсчёт аэробных колоний свежей продукции и их питательных растворов из разных мест отбора. (A) Микрозелень (Продукция) и питательный раствор (NS) с фермы H и лабораторной системы 1 (L1); (B) салат (Продукция) и питательный раствор (NS) с фермы A, из продуктового магазина (G) и лабораторной системы 2 (L2). Планка погрешности показывает стандартное отклонение (SD). Разные прописные или строчные буквы указывают на значительные различия между группами образцов (p < 0,05).
Питательный раствор фермы H показал значительно более высокую бактериальную нагрузку, чем у L1. Бактериальная нагрузка питательного раствора фермы A была значительно ниже, чем у лабораторных гидропонных систем (L2). В водной системе фермы A наибольшее количество бактерий было обнаружено на входе из рыбоводного бассейна (3,7 log КОЕ/мл).
Опрос по практике обеспечения безопасности пищевых продуктов
Осведомлённость и практика обеспечения безопасности пищевых продуктов на двух фермах были оценены с помощью опроса, баллы представлены в дополнительной таблице 3. Ферма H внедряла в целом более качественные методы обеспечения безопасности пищевых продуктов, чем ферма A, и получила более высокие баллы за санитарию оборудования и окружающей среды, а также за осведомлённость о безопасности пищевых продуктов, например, применение дезинфицирующих средств и использование средств индивидуальной защиты. Обе фермы уделяли минимальное внимание обработке циркулирующей воды, что может привести к накоплению отходов и микробных опасностей в системе. При обращении со свежей продукцией ферма A использовала водопроводную воду для ополаскивания семян перед началом проращивания, в то время как ферма H не применяла никакой обработки семян.
Структура микробного сообщества и взаимосвязи
Состав микробиома и роды 50 наиболее распространённых ASV представлены на Рисунке 2 и в дополнительной таблице 4. Proteobacteria была наиболее распространённой группой в системах микрозелени (Рисунок 2A). Pseudomonas была доминирующим родом на ферме H, и P. alcaligenes (ASV1746), P. fluorescens (ASV1756 и ASV1758) и P. lutea (ASV1767) входили в число 50 наиболее распространённых ASV. Actinobacteria и Firmicutes устойчивы к санитарной обработке (Ramlal и др., 2021). Они присутствовали в пустой среде для выращивания микрозелени, в санитарно-обработанных лотках и на обуви работников. Cyanobacteria являются распространёнными фотосинтезирующими бактериями в сточных водах и известны как "сине-зелёные водоросли". Они часто наблюдались в системах с салатом (ферма A и L2) в виде зелёных отложений на лотках для выращивания и возле корней растений. В числе 50 наиболее распространённых ASV были идентифицированы пять ASV цианобактерий. Чрезмерный рост цианобактерий приводит к образованию опасных токсинов, истощению кислорода и гибели водных животных (Ezenarro и др., 2021).
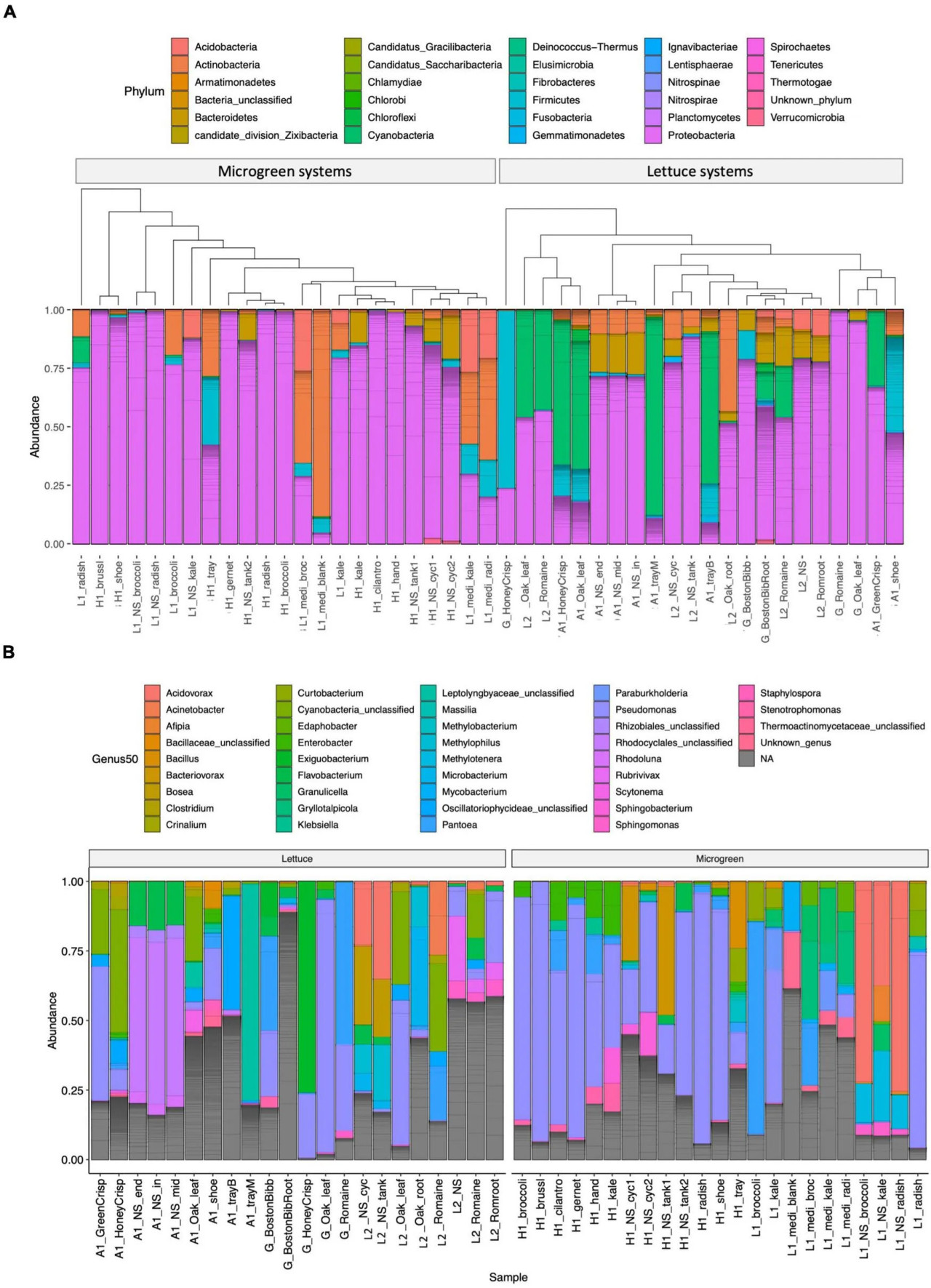
РИСУНОК 2 Состав микробиома. (A) Состав на уровне типов: образцы сгруппированы по системам микрозелени (слева) или салата (справа). Образцы кластеризованы по сходству микробиома с использованием филогенетических древовидных структур. (B) Классификация на уровне родов для 50 наиболее распространённых ASV, разделённых по типу системы: салат (слева) или микрозелень (справа). Отдельные ASV разделены серой линией на столбчатых диаграммах.
Микробиомы из одного типа и места отбора располагались близко на дереве (Рисунок 2A). Образцы сходных типов из разных мест также кластеризовались близко друг к другу, например, образец салата из продуктового магазина и L2, а также микрозелень капусты кале с фермы H и L1.
Разнообразие микробиома в зависимости от места и типа образца
Разнообразие микробиома было изучено с использованием нескольких индексов альфа-разнообразия (Рисунок 3A) и бета-разнообразия (Рисунок 3B). Микрозелень с ферм H и L1 имела сходное альфа-разнообразие сообществ. Однако питательные растворы с фермы H имели более высокое богатство сообщества, чем у L1. Питательный раствор фермы H использовался для нескольких видов микрозелени, что потенциально увеличивало богатство сообщества. Образцы салата и образцы окружающей среды (мазки с обуви и лотков) с фермы A показали наибольшее богатство сообщества среди всех образцов (Рисунок 3A).
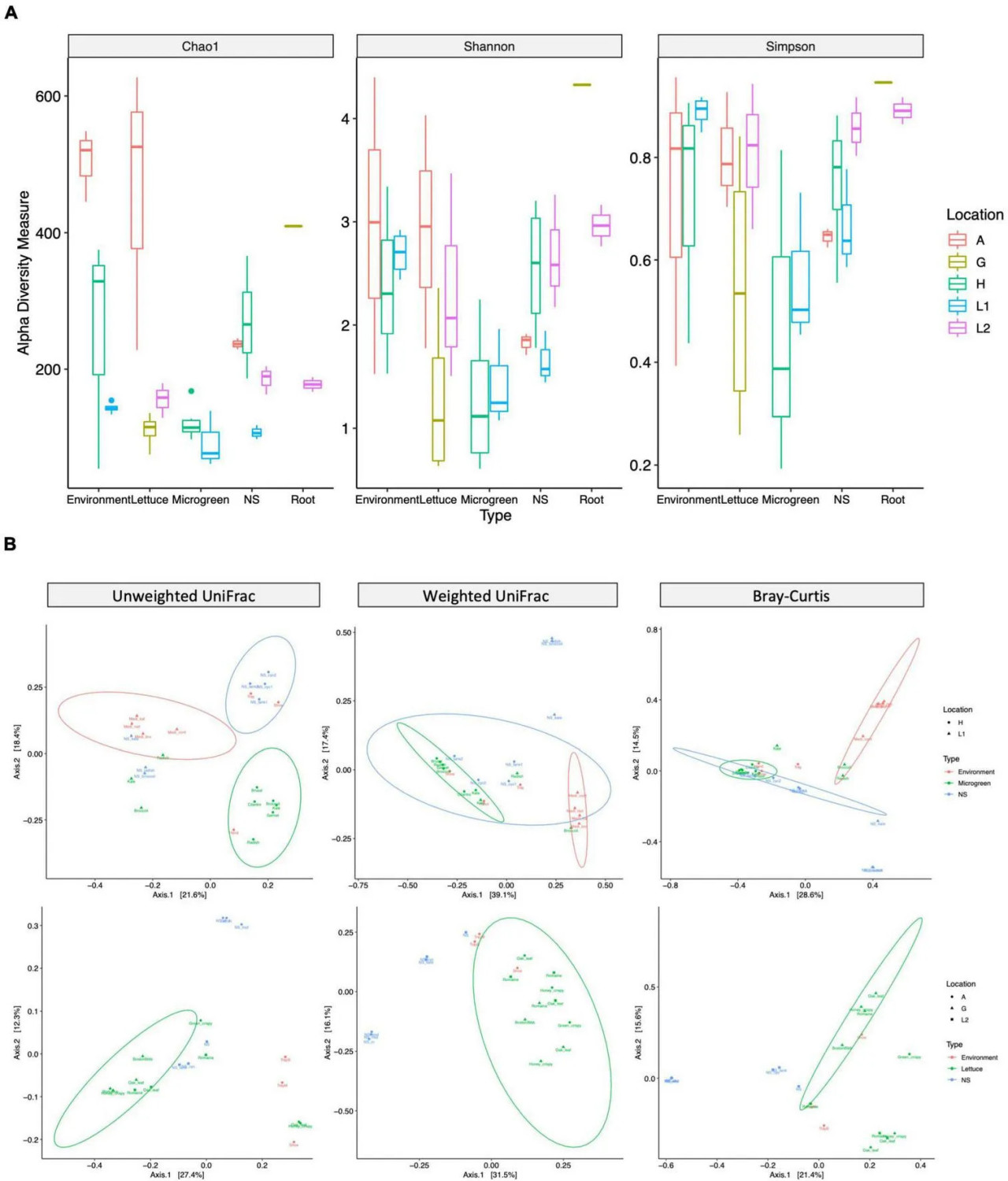
РИСУНОК 3 Измерения альфа- и бета-разнообразия бактериальных сообществ. (A) Альфа-разнообразие: богатство и равномерность сообщества для разных типов образцов. Индексы альфа-разнообразия: chao1 измеряет богатство сообщества; Shannon и Simpson измеряют богатство и равномерность сообщества. Прямоугольник представляет первый и третий квартили, центральная линия представляет медиану, а усы — значения от минимума до максимума. (B) Бета-разнообразие: различия в составе микробиома между типами образцов. Образцы сгруппированы по системам микрозелени (верхний ряд, H и L1) или салата (нижний ряд, A, G и L2). Расстояния: невзвешенное UniFrac (слева, присутствие таксонов), взвешенное UniFrac (середина, присутствие и относительное обилие таксонов) и Брей–Кёртис (справа, присутствие и относительное обилие таксонов) визуализированы с помощью графиков PCoA. Эллипсы указывают 95% доверительные области для кластеров по нормальному распределению.
Тест PERMANOVA выявил значительные композиционные различия между разными типами образцов и разными местами отбора (дополнительная таблица 5). Согласно невзвешенному расстоянию UniFrac, микрозелень с фермы H имела сходные ASV с рукой работника, в то время как её питательные растворы имели сходные ASV с обувью и лотками; два образца салата с фермы A имели сходные ASV с обувью работника, а не с другим салатом (Рисунок 3B слева). Образцы микрозелени и салата из разных мест кластеризовались вместе на графике взвешенного UniFrac, что указывает на то, что микробиомы растений из разных мест отбора имели общие филогенетические паттерны (Рисунок 3B середина).
Идентификация патогенов и организмов порчи
Мы провели скрининг микробиомов на наличие патогенов растений, человека и рыб, а также организмов порчи (Рисунок 4). Патогены с нулевой толерантностью E. coli O157:H7, Salmonella spp. и Listeria monocytogenes не были обнаружены ни на одной ферме. Не классифицированная E. coli (ASV1628) присутствовала на обуви работника фермы A (Рисунок 4A). E. coli используется как индикатор фекального загрязнения в полевом земледелии (Kaczmarek и др., 2019). Согласно опросу, ферма A имела открытую ферму, примыкающую к теплице, и работники не меняли обувь между фермами. Хотя у входа была установлена раковина для дезинфекции, это не устранило риск занесения контаминантов из открытой среды. Pseudomonas aeruginosa является патогеном человека и была обильна в микрозелени капусты кале и брокколи на ферме H, в салате L2 и на обуви работника фермы A. Патоген растений P. syringae был идентифицирован в низком количестве на обуви работника фермы H. P. syringae, как известно, вызывает бактериальную пятнистость листьев на нескольких видах крестоцветной микрозелени (UMass Extension, 2018). P. putida является полезным организмом для стимуляции роста растений, но также и оппортунистическим патогеном человека, и она была обильна в питательном растворе фермы H. Патоген рыб Aeromonas hydrophila (ASV1571 и ASV1572) был идентифицирован в низком количестве в салате GreenCrisp на ферме A, но не в других образцах (Рисунок 4). Если вода циркулирует между системой растений и рыбоводным бассейном без предварительной обработки, существует потенциальный риск передачи патогенов через воду рыбам.
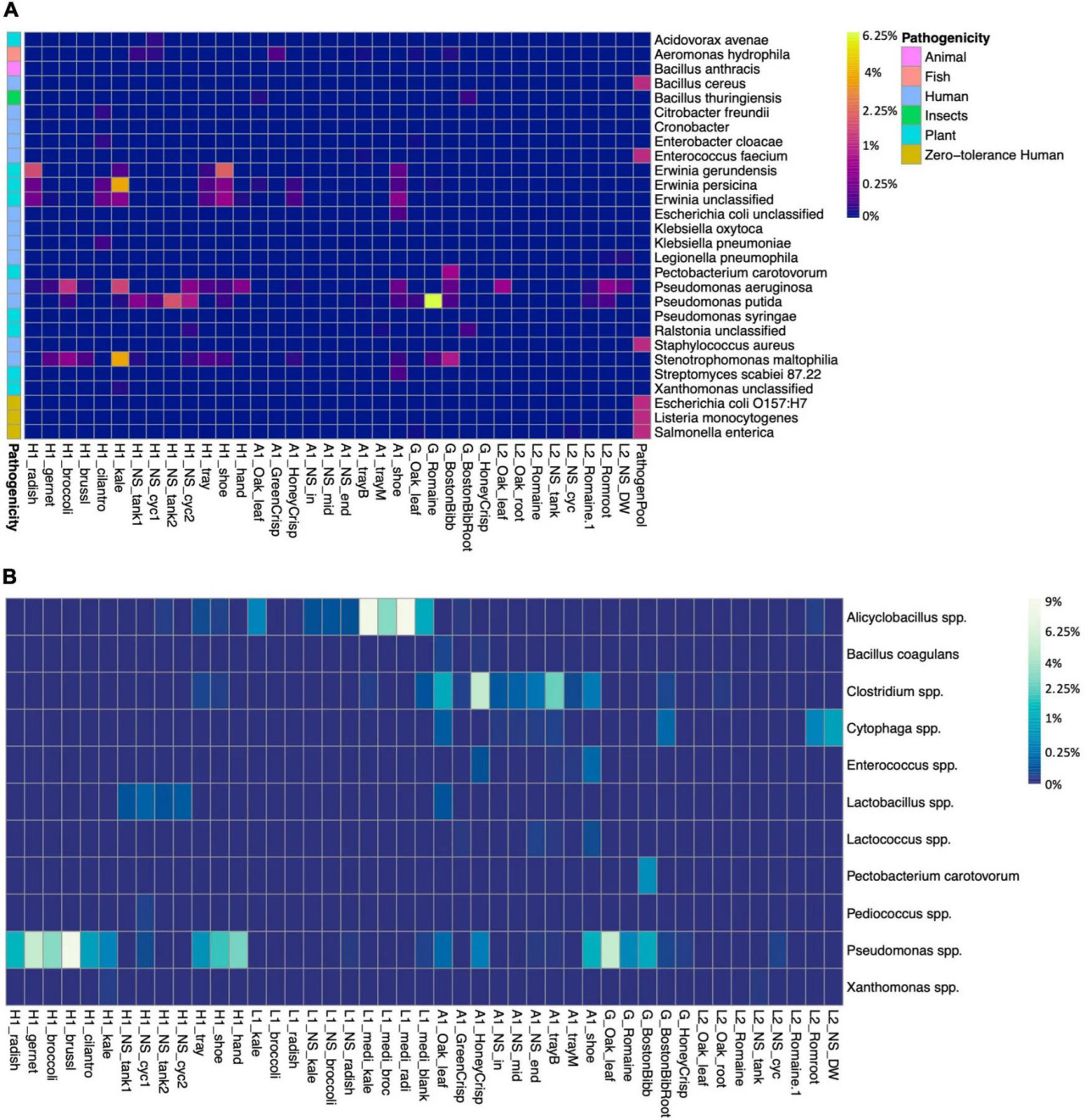
РИСУНОК 4 Наличие патогенов и представляющих интерес бактерий, а также видов порчи в образцах. (A) Наличие патогенов и представляющих интерес бактерий. Цветные блоки слева указывают на потенциальную патогенность для животных, рыб, человека, насекомых или растений. Образцы L1 не содержат ни одного из перечисленных штаммов, поэтому не показаны на графике. (B) Наличие видов бактерий порчи в образцах, ASV, связанные с порчей, сгруппированы на видовом уровне. Цветовая шкала тепловой карты представлена в масштабе квадратного корня.
Бактерии порчи ухудшают качество продуктов и создают неприятный запах, вкус и текстуру. Основную порчу свежей продукции вызывают молочнокислые бактерии, и эта группа включает Lactobacillus, Leuconostoc, Pedicoccus, Lactococcus и Enterococcus (Kaczmarek и др., 2019). Lactobacillus, Lactococcus и Enterococcus были обнаружены в нескольких образцах фермы A и в питательном растворе фермы H (Рисунок 4B). Pseudomonas spp. были обильны в микрозелени фермы H и на санитарно-обработанных лотках, в образцах обуви и салата фермы A, а также в салате G, включая виды порчи P. fluorescens, P. viridiflava и P. tolaasii. На ферме H было две системы, и согласно опросу, питательный раствор системы 1 (H1_NS_tank1 и H1_NS_cyc1) циркулировал почти месяц, в то время как питательный раствор системы 2 (H1_NS_tank2 и H1_NS_cyc2) был недавно заменён. Pseudomonas порчи были относительно менее обильны в питательном растворе системы 2 и микрозелени капусты кале по сравнению с системой 1 и другими пятью видами микрозелени. Род Clostridium был идентифицирован в образцах фермы A и включал несколько видов порчи. Однако вызывающие вспышки пищевых заболеваний виды C. botulinum и C. perfringens не были обнаружены. На обуви работника фермы A сохранялось множество бактерий порчи, что снова указывает на недостаточную санитарию.
Картирование последовательностей вариантов ампликонов Escherichia coli по нескольким базам данных
Патогенность ASV1628 ("E. coli_unclassified") была неопределённой. Поэтому мы дополнительно картировали 38 последовательностей из ASV1628 по нескольким базам данных последовательностей рРНК (дополнительная таблица 6). Базы данных большой субъединицы (23S) (средняя идентичность 93,2%) классифицировали все 38 последовательностей как Escherichia–Shigella с 84 соседними штаммами, имеющими идентичность >97%, и 9 соседними штаммами E. coli, выровненными на 100% с частичной последовательностью 23S исходного образца. Более высокая идентичность была достигнута с использованием баз данных малой субъединицы (16S) (средняя идентичность 99,1%) со 130 соседними штаммами, имеющими идентичность >97%, и некоторые исходные последовательности были классифицированы как Enterobacter, Pantoea или Salmonella. Нам не удалось повысить таксономическое разрешение с помощью баз данных рРНК малой или большой субъединицы.
Для дальнейшего подтверждения патогенности мы картировали 38 исходных последовательностей по данным полных геномов в NCBI и рассчитали расстояние Джукса–Кантора между исходными последовательностями и выбранными патогенными штаммами (дополнительный рисунок 3). Расстояние Джукса–Кантора измеряет сходство на видовом уровне, и расстояние < 0,03 использовалось для классификации потенциальных патогенов (Schloss and Handelsman, 2006; Bibby и др., 2010; Chidamba and Korsten, 2015). Среди 38 исходных последовательностей 9 последовательностей могли происходить от нескольких серотипов патогенных E. coli, Shigella flexneri, Klebsiella oxytoca или Klebsiella pneumoniae (Таблица 1). Одна последовательность (E. coli_85263602) была тесно связана с несколькими штаммами E. coli O157:H7.
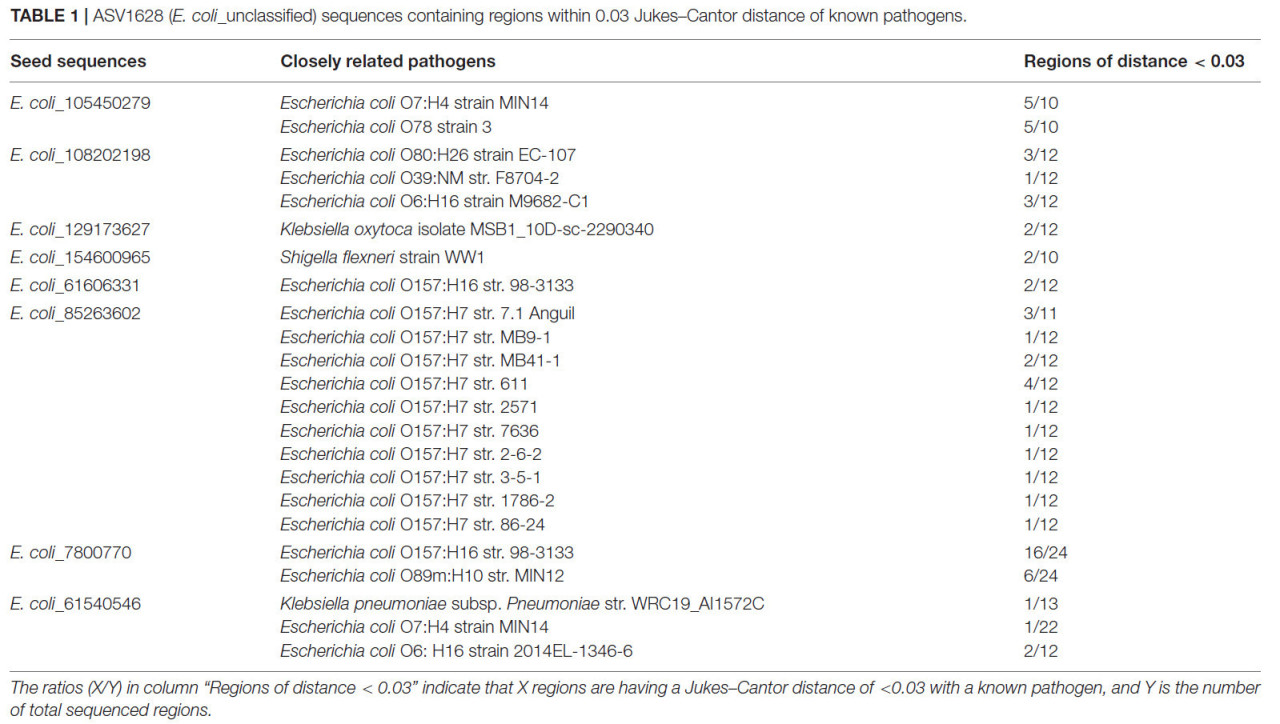
ТАБЛИЦА 1 Последовательности ASV1628 (E. coli_unclassified), содержащие участки в пределах расстояния Джукса–Кантора 0,03 от известных патогенов.
Отношения (X/Y) в колонке "Участки с расстоянием < 0,03" означают, что X участков имеют расстояние Джукса–Кантора <0,03 с известным патогеном, а Y — общее число секвенированных участков.
Идентификация путей передачи бактерий
Путь передачи бактерий в гидропонных/аквапонных системах был исследован с использованием SourceTracker и визуализирован с помощью диаграмм Санкея (Рисунок 5). Образцы окружающей среды были назначены источниками для проверки на наличие бактериальных сообществ из образцов свежей продукции. На ферме H (Рисунок 5A слева) обувь и руки работника вносили 21,1–89,7% и 1,5–55,1% микробиома микрозелени соответственно. На ферме A (Рисунок 5B слева) обувь работника и лоток для выращивания рассады вносили 8,6–49,8% и 29,9–66,6% микробиома салата соответственно. От 15,2 до 32,0% ASV в образцах салата не были общими ни с одним из образцов окружающей среды.
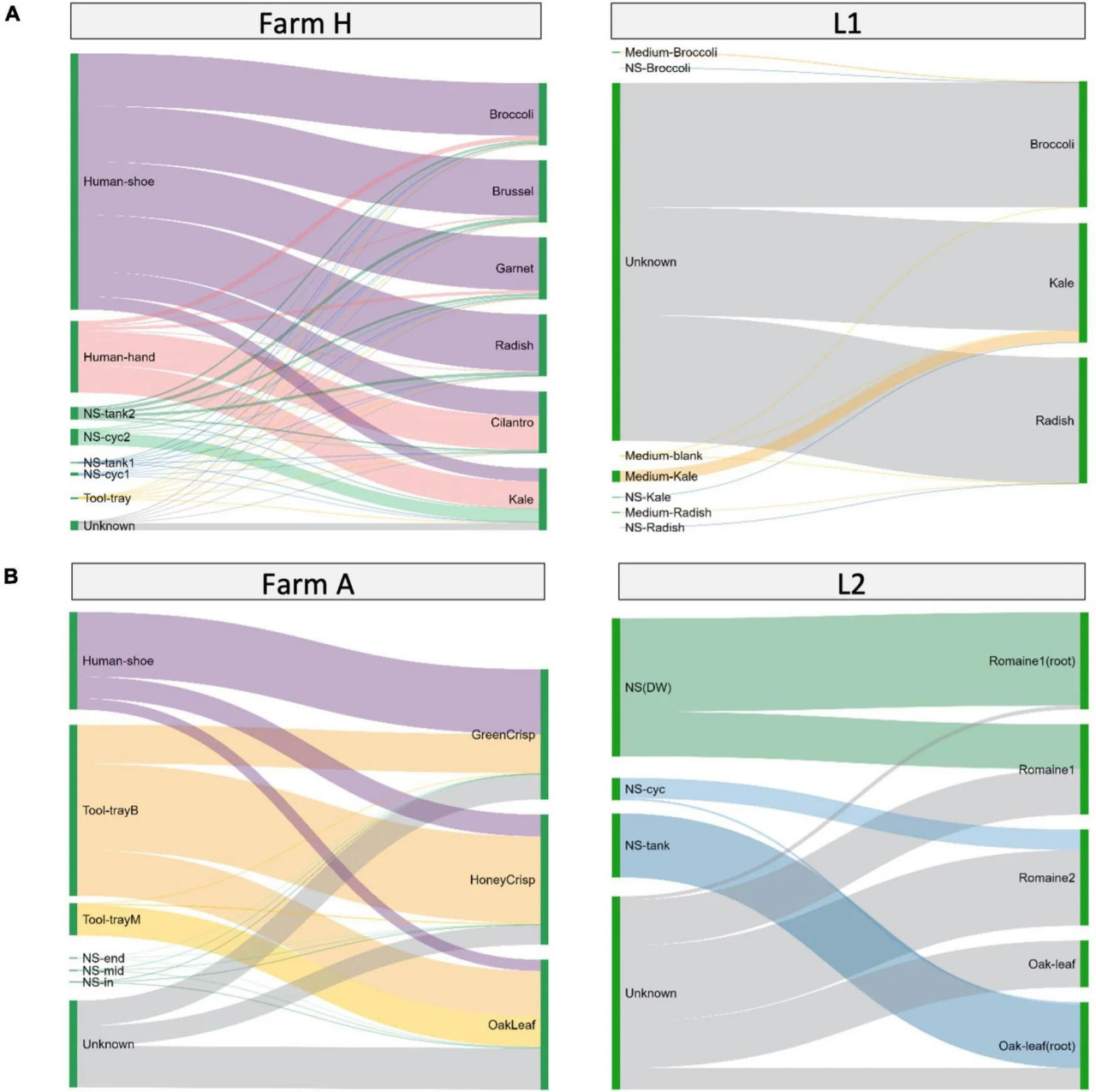
РИСУНОК 5 Доли источников окружающей среды для гидропонно выращенной микрозелени или салата. Образцы сгруппированы по типу системы: (A) микрозелень или (B) салат. Доли источников окружающей среды для образцов свежей продукции, оценённые с помощью SourceTracker и визуализированные в виде диаграмм Санкея. Образцы источников окружающей среды представлены слева, а образцы свежей продукции являются "приёмниками" и представлены справа. Ширина линии отдельных потоков указывает на процентное содержание микроорганизмов, передаваемых от источника к приёмнику. Доли вклада от разных источников в сумме составляют 100% для образца-приёмника (правая сторона). Высота отдельных столбцов (левая сторона) образцов-источников представляет сумму долей для каждого образца-приёмника.
При рассмотрении лабораторных систем микрозелень L1 и салат L2 имели 85,24–99,47% и 47–78% ASV из неизвестных источников (Рисунки 5A, B справа). Для лучшего понимания источника бактерий мы отобрали среду для выращивания микрозелени отдельно в L1, а также корень и съедобную часть в L2. Среда для выращивания или питательный раствор вносили небольшое количество ASV в микрозелень. Резервуарные баки питательного раствора разделяли большие доли (66–96%) ASV с корнем салата и меньшие доли (0–46%) с листом салата. ASV из неизвестных источников могут быть частью фонового микробиома растений.
Чтобы лучше рассмотреть фоновый микробиом растений, мы построили диаграммы Венна для исследования перекрытий микробиома (Рисунок 6 и дополнительная таблица 7). Микрозелень фермы H разделяла 53 ASV с микрозеленью L1 и 27 уникальных ASV с окружающей средой системы L1, в то время как микрозелень L1 разделяла 39 уникальных ASV с окружающей средой фермы H (Рисунок 6A). В системах салата было 141 и 193 перекрывающихся ASV среди всех салатов (G, ферма A и L2) и между фермой A и L2 (Рисунок 6B). Кроме того, салат L2 разделял 29 уникальных ASV с окружающей средой фермы A, и 46 уникальных ASV из салата фермы A также появились в питательном растворе L2. Этот результат согласуется с наблюдением в бета-разнообразии, что сходные образцы свежей продукции из разных систем имели общие паттерны микробиома. Общие ASV между образцами салата, а также микрозелени, в основном принадлежат к родам Pseudomonas и Pantoea. Сообщалось, что Pseudomonas и Pantoea являются частью фонового микробиома листа салата (Rastogi и др., 2012). Образцы салата также разделяли ASV из Cyanobacteria, которые связаны с водной средой.
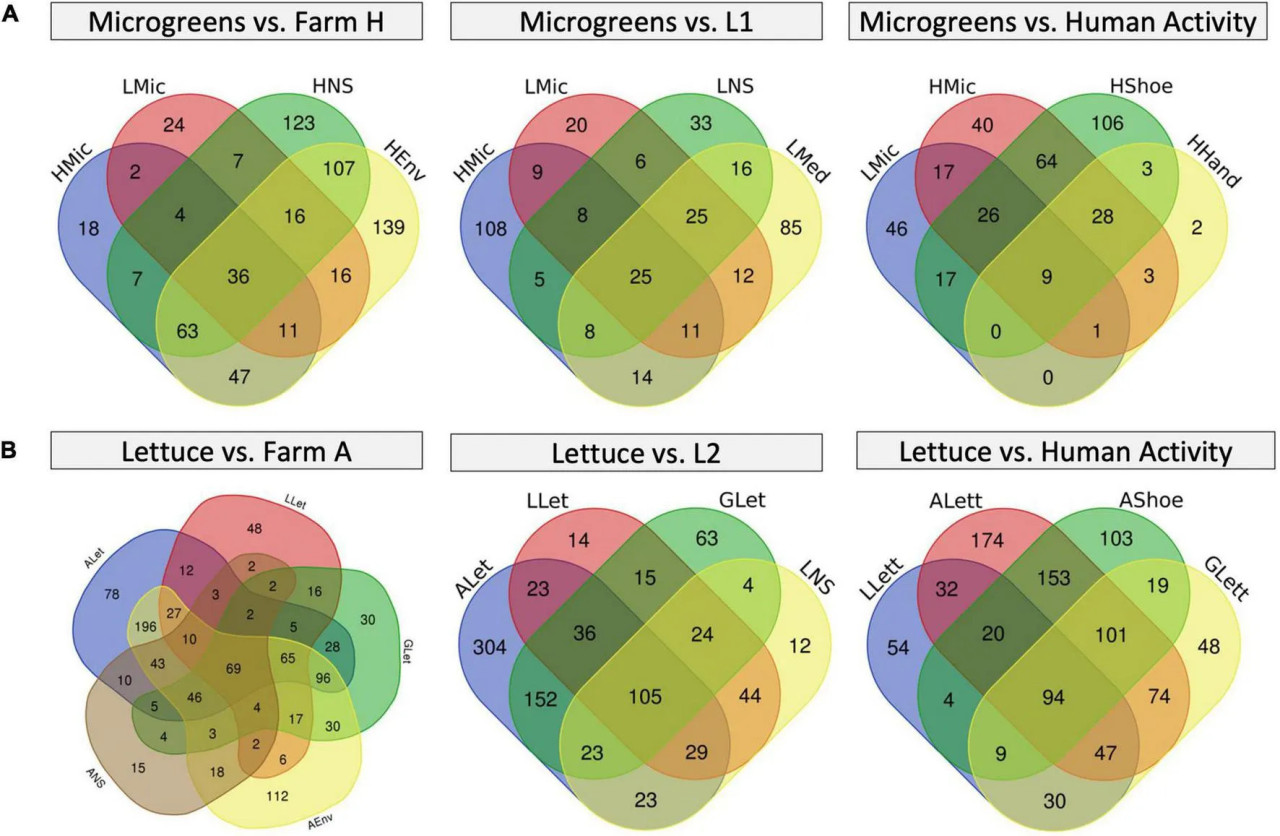
РИСУНОК 6 Диаграммы Венна перекрытия микробиома между различными типами образцов. (A) Перекрытие микрозелени с окружающей средой фермы H (слева), с окружающей средой системы L1 (середина) и с обувью и руками работника фермы H (справа). (B) Перекрытие салата с окружающей средой фермы A (слева), с системой L2 (середина) и с обувью работника фермы A (справа). Числа в каждой области представляют количество уникальных ASV в этой области.
Таким образом, мы дополнительно предложили двунаправленный путь передачи бактерий внутри HCS (Рисунок 7). При хорошей личной гигиене основным источником бактерий будет свежая продукция. При плохой личной гигиене контаминанты попадают в систему через деятельность человека. Инструменты, вода и работники являются переносчиками бактерий, а плохая санитария объектов приводит к перекрёстному загрязнению и накоплению опасностей.
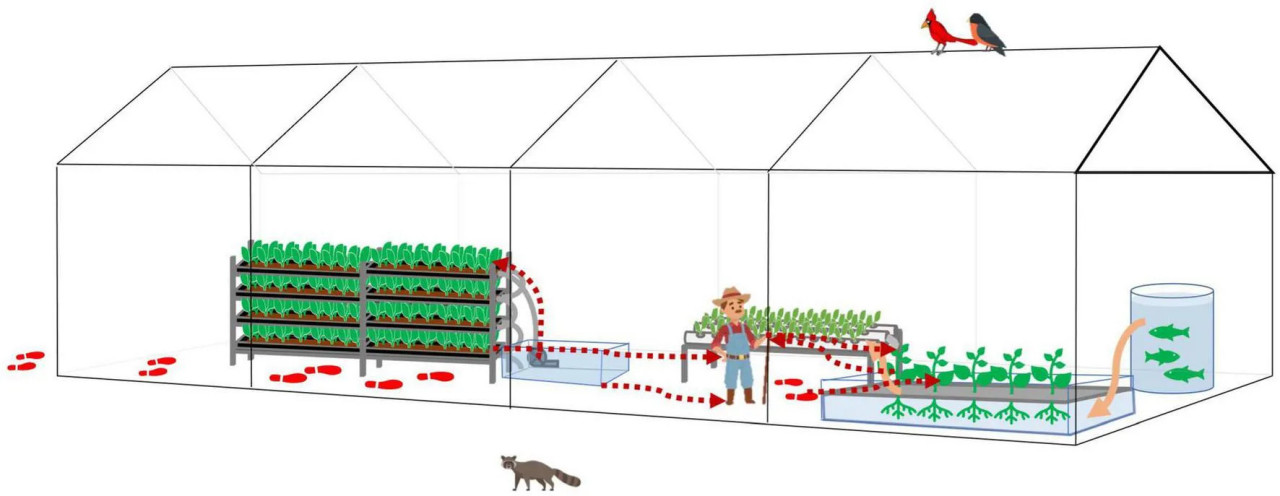
РИСУНОК 7 Передача бактерий под влиянием деятельности человека в типичной тепличной гидропонной/аквапонной экосистеме. Передача бактерий следует по красным пунктирным линиям. Следы указывают на потенциальные контаминанты, заносимые деятельностью человека и передаваемые внутри теплицы. Поток воды следует по оранжевым сплошным линиям или трубам, соединяющим водные объекты.
Обсуждение
Секвенирование 16S рРНК, как золотой стандарт для профилирования микробиома, использовалось в системах производства продуктов питания для оценки эффективности санитарии и сдвигов микробиома (Gu и др., 2019, 2020). Однако секвенирование 16S рРНК обеспечивает недостаточное разрешение для различения близкородственных видов (UCD Centre for Food Safety и др., 2018; Zwirzitz и др., 2020). Сообщалось, что секвенирование 16S-ITS-23S рРНК вместе с таксономическим присвоением ASV обеспечивает более высокое таксономическое разрешение (Martijn и др., 2019; Graf и др., 2021). В этом исследовании были объединены посещения объектов, опрос по практике безопасности пищевых продуктов, традиционный подсчёт колоний, секвенирование 16S-ITS-23S рРНК и множественные биоинформатические инструменты для изучения микробиомов, микробных опасностей и передачи бактерий в гидропонных/аквапонных экосистемах. Результат дал нам картину микробных сообществ и статуса риска гидропонной/аквапонной экосистемы.
Мы исследовали два типа коммерческих фермерских систем (гидропонную и аквапонную) и сравнили микробные нагрузки и микробиомы с лабораторными контрольными гидропонными системами. Подсчёт аэробных колоний измеряет общую микробную нагрузку циркулирующей воды и свежей продукции. В литературе количество аэробных бактерий для микрозелени и гидропонно выращенного салата обычно составляет примерно 7,5 и 4 log КОЕ/г соответственно, и наши образцы находились в аналогичном диапазоне (Lee и др., 2009; Chandra и др., 2012; Dankwa и др., 2020, 2021). Салат из продуктового магазина имел более высокую микробную нагрузку, возможно, из-за процедур после сбора урожая (упаковка, хранение, транспортировка и другие), которые могут вносить бактерии и способствовать их росту (Gil и др., 2015; Dankwa и др., 2020). Салат фермы A имел более высокое богатство сообщества, чем другие образцы салата, возможно, из-за плохой гигиенической практики, например, работники не меняли обувь и одежду между крытой и открытой фермами. Мазок с обуви работника фермы A имел наибольшее богатство микробиома и разделял множество уникальных ASV с салатом фермы A, включая ASV833 (Thermoactinomycetaceae_unclassified), ASV830 (Staphylospora_unclassified) и ASV744 (Parageobacillus_unclassified). Эти ASV вместе с ASV1628 (E. coli_unclassified), вероятно, происходили из внешней среды теплицы (Рисунок 6B и дополнительная таблица 7). На основании этих результатов мы предложили ферме A инвестировать в средства индивидуальной защиты, такие как смена обуви при работе внутри теплицы.
В гидропонике питательный раствор циркулирует внутри системы и связывает каждое растение, что может представлять значительный риск перекрёстного загрязнения свежей продукции (Gil и др., 2015). Питательный раствор фермы H показал более высокую микробную нагрузку, чем другие системы, возможно, из-за относительно длительного времени пребывания (1 месяц). Pseudomonas spp. были обильны на ферме H. Ферма H использовала защитную одежду перед входом в теплицу, что ограничивало внешний источник бактерий. Сообщалось, что P. aeruginosa и P. putida являются фоновой микрофлорой микрозелени, которая может присутствовать в семенах (Bergšpica и др., 2020). Однако P. fluorescens (ASV1821) была обильна на ферме H и является одной из основных Pseudomonas порчи, вызывающих мягкую гниль и порчу сочных тканей овощей (Liao, 2006). Микрозелень собирают, срезая стебель над корнем, и она сильно скоропортящаяся в зависимости от вида (Berba and Uchanski, 2012). Таким образом, мы предложили ферме H включить этап санитарной обработки семян в стандартную операционную процедуру, улучшить метод санитарии инструментов и чаще промывать систему для снижения накопления микробных опасностей и потенциального улучшения срока годности продукции микрозелени.
Картирование на базу данных Athena привело к множеству неклассифицированных ASV. Вероятно, это связано с ограниченным охватом базы данных (Graf и др., 2021). Например, на обуви работника фермы A 38 прочтений последовательностей с идентичностью >95% были сгруппированы в ASV1628 (E. coli_unclassified). В базе данных Athena содержится 187 штаммов E. coli, что является лишь минимальной частью ныне идентифицированных штаммов E. coli. Для повышения разрешения классификации и подтверждения патогенности мы картировали 38 исходных последовательностей по нескольким базам данных. При использовании баз данных 16S рРНК некоторые последовательности были картированы на Pantoea, Enterobacter cloacae complex или Salmonella. Это связано с тем, что виды бактерий могут иметь общие участки последовательности 16S рРНК, что приводит к низкому таксономическому разрешению. Путём картирования последовательности 16S-ITS-23S на полные бактериальные геномы в коллекции нуклеотидов NCBI и использования критерия расстояния Джукса–Кантора <0,03 мы смогли дополнительно повысить разрешение и идентифицировать потенциальные патогены (Таблица 1). Однако мы не можем определить истинный патоген, используя только расстояние Джукса–Кантора (Bibby и др., 2010). Для дальнейшего подтверждения патогенности рекомендуется применять другие методы секвенирования, такие как WGS, для вызывающих беспокойство образцов.
Это исследование демонстрирует, что высокопроизводительное секвенирование 16S-ITS-23S рРНК может выявить ценную информацию о микробиоме гидропонных/аквапонных систем и повысить таксономическое разрешение для идентификации микробных опасностей. Мы локализовали множество ASV до конкретных источников и провели скрининг на потенциальные патогены или организмы порчи в проанализированных системах. Наши результаты внесут вклад в понимание экологии гидропонных/аквапонных систем и управления рисками. Кроме того, методы, использованные в этом исследовании, могут быть применены к другим сельскохозяйственным/пищевым производственным системам в качестве инструмента оценки рисков, а также углубить наши знания о взаимосвязях микробиомов в экосистемах. Продолжающееся совершенствование стратегий секвенирования длинных прочтений всего оперона рРНК и расширение охвата бактериальных генетических баз данных дополнительно повысят пропускную способность и таксономическое разрешение и могут предоставить большой потенциал для их внедрения в качестве экономически эффективного инструмента для идентификации микробных опасностей.
Дополнительный материал
Дополнительный материал к этой статье можно найти онлайн по адресу: https://www.frontiersin.org/articles/10.3389/fmicb.2022.879260/full#supplementary-material
Сноски
¹ https://rdrr.io/cran/pheatmap/
² https://sankeymatic.com/build/
³ https://bioinformatics.psb.ugent.be/webtools/Venn/
Ссылки
1. Allende A., Castro-Ibáñez I., Lindqvist R., Gil M. I., Uyttendaele M., Jacxsens L. (2017). Quantitative contamination assessment of Escherichia coli in baby spinach primary production in Spain: effects of weather conditions and agricultural practices. Int. J. Food Microbiol. 257, 238–246. doi: 10.1016/j.ijfoodmicro.2017.06.027. CrossRef. Google Scholar.
2. Anderson M. J. (2017). "Permutational Multivariate Analysis of Variance (PERMANOVA)," in Wiley StatsRef: Statistics Reference Online, eds Balakrishnan N., Colton T., Everitt B., Piegorsch W., Ruggeri F., Teugels J. L. (Atlanta: American Cancer Society), 1–15. doi: 10.1002/9781118445112.stat07841. CrossRef. Google Scholar.
3. Barragán J. L. M., Cuesta L. D. I., Susa M. S. R. (2021). Quantitative microbial risk assessment to estimate the public health risk from exposure to enterotoxigenic E. coli in drinking water in the rural area of Villapinzon, Colombia. Microb. Risk Anal. 18:100173. doi: 10.1016/j.mran.2021.100173. CrossRef. Google Scholar.
4. Berba K. J., Uchanski M. E. (2012). Postharvest Physiology of Microgreens. J. Young Investig. 24, 1–5. doi: 10.3390/molecules26113247. CrossRef. Google Scholar.
5. Bergšpica I., Ozola A., Miltiòa E., Alksne L., Meistere I., Cibrovska A., et al (2020). Occurrence of Pathogenic and Potentially Pathogenic Bacteria in Microgreens, Sprouts, and Sprouted Seeds on Retail Market in Riga, Latvia. Foodborne Pathog. Dis. 17, 420–428. doi: 10.1089/fpd.2019.2733. CrossRef. Google Scholar.
6. Bibby K., Viau E., Peccia J. (2010). Pyrosequencing of the 16S rRNA gene to reveal bacterial pathogen diversity in biosolids. Water Res. 44, 4252–4260. doi: 10.1016/j.watres.2010.05.039. CrossRef. Google Scholar.
7. Callahan B. J., McMurdie P. J., Holmes S. P. (2017). Exact sequence variants should replace operational taxonomic units in marker-gene data analysis. ISME J. 11, 2639–2643. doi: 10.1038/ismej.2017.119. CrossRef. Google Scholar.
8. Callahan B. J., McMurdie P. J., Rosen M. J., Han A. W., Johnson A. J. A., Holmes S. P. (2016). DADA2: high-resolution sample inference from Illumina amplicon data. Nat. Methods 13, 581–583. doi: 10.1038/nmeth.3869. CrossRef. Google Scholar.
9. Callahan B. J., Wong J., Heiner C., Oh S., Theriot C. M., Gulati A. S., et al (2019). High-throughput amplicon sequencing of the full-length 16S rRNA gene with single-nucleotide resolution. Nucleic Acids Res. 47:e103. doi: 10.1093/nar/gkz569. CrossRef. Google Scholar.
10. Center for Disease Control and Prevention [CDC] (2014). Multistate Outbreak of Shiga toxin-producing Escherichia coli O121 Infections Linked to Raw Clover Sprouts (Final Update). Atlanta: Centers for Disease Control and Prevention. Google Scholar.
11. Center for Disease Control and Prevention [CDC] (2016). Multistate Outbreak of Shiga toxin-producing Escherichia coli O157 Infections Linked to Alfalfa Sprouts Produced by Jack & The Green Sprouts (Final Update). Atlanta: Centers for Disease Control and Prevention. Google Scholar.
12. Center for Disease Control and Prevention [CDC] (2019). Outbreak of E. coli Infections Linked to Romaine Lettuce. Atlanta: Centers for Disease Control and Prevention. Google Scholar.
13. Center for Disease Control and Prevention [CDC] (2020). Outbreak of E. coli Infections Linked to Clover Sprouts. Atlanta: Centers for Disease Control and Prevention. Google Scholar.
14. Chandra D., Kim J. G., Kim Y. P. (2012). Changes in microbial population and quality of microgreens treated with different sanitizers and packaging films. Hortic. Environ. Biotechnol. 53, 32–40. doi: 10.1007/s13580-012-0075-6. CrossRef. Google Scholar.
15. Chaumeil P.-A., Mussig A. J., Hugenholtz P., Parks D. H. (2020). GTDB-Tk: a toolkit to classify genomes with the Genome Taxonomy Database. Bioinformatics 36, 1925–1927. doi: 10.1093/bioinformatics/btz848. CrossRef. Google Scholar.
16. Chidamba L., Korsten L. (2015). Pyrosequencing analysis of roof-harvested rainwater and river water used for domestic purposes in Luthengele village in the Eastern Cape Province of South Africa. Environ. Monit. Assess. 187:41. doi: 10.1007/s10661-014-4237-0. CrossRef. Google Scholar.
17. Cole J. R., Wang Q., Fish J. A., Chai B., McGarrell D. M., Sun Y., et al (2014). Ribosomal Database Project: data and tools for high throughput rRNA analysis. Nucleic Acids Res. 42, D633–D642. doi: 10.1093/nar/gkt1244. CrossRef. Google Scholar.
18. Colorado Department of Public Health and Environment (2012). A Farm Food Safety Toolkit: A Nationwide Survey. Denver, CO: Colorado Department of Public Health and Environment. Google Scholar.
19. Dankwa A. S., Machado R. M., Perry J. J. (2020). Sources of food contamination in a closed hydroponic system. Lett. Appl. Microbiol. 70, 55–62. doi: 10.1111/lam.13243. CrossRef. Google Scholar.
20. Dankwa A. S., Machado R. M., Perry J. J. (2021). Sanitizer efficacy in reducing microbial load on commercially grown hydroponic lettuce. J. Sci. Food Agric. 101, 1403–1410. doi: 10.1002/jsfa.10753. CrossRef. Google Scholar.
21. Davis N. M., Proctor D. M., Holmes S. P., Relman D. A., Callahan B. J. (2018). Simple statistical identification and removal of contaminant sequences in marker-gene and metagenomics data. Microbiome 6:226. doi: 10.1186/s40168-018-0605-2. CrossRef. Google Scholar.
22. De Filippis F., Valentino V., Alvarez-Ordóñez A., Cotter P. D., Ercolini D. (2021). Environmental microbiome mapping as a strategy to improve quality and safety in the food industry. Curr. Opin. Food Sci. 38, 168–176. doi: 10.1016/j.cofs.2020.11.012. CrossRef. Google Scholar.
23. Deering A. (2018). Guidelines for Food Safety in Hydroponic Production. Available Online at: https://www.purdue.edu/hla/sites/cea/wp-content/uploads/sites/15/2018/10/AmandaDeering9-5-18.pdf (accessed March 20, 2019). Google Scholar.
24. Deguenon J. M., Travanty N., Zhu J., Carr A., Denning S., Reiskind M. H., et al (2019). Exogenous and endogenous microbiomes of wild-caught Phormia regina (Diptera: Calliphoridae) flies from a suburban farm by 16S rRNA gene sequencing. Sci. Rep. 9:20365. doi: 10.1038/s41598-019-56733-z. CrossRef. Google Scholar.
25. Ezenarro J. J., Ackerman T. N., Pelissier P., Combot D., Labbé L., Muñoz-Berbel X., et al (2021). Integrated Photonic System for Early Warning of Cyanobacterial Blooms in Aquaponics. Anal. Chem. 93, 722–730. doi: 10.1021/acs.analchem.0c00935. CrossRef. Google Scholar.
26. FamilyFarmed (2019). Typical Audit Questions. On Farm Food Safety. Available online at: http://onfarmfoodsafety.org/typical-audit-questions/ (accessed April 22, 2019). Google Scholar.
27. Fierer N., Hamady M., Lauber C. L., Knight R. (2008). The influence of sex, handedness, and washing on the diversity of hand surface bacteria. Proc. Natl. Acad. Sci. U. S. A. 105, 17994–17999. doi: 10.1073/pnas.0807920105. CrossRef. Google Scholar.
28. Flores G. E., Bates S. T., Caporaso J. G., Lauber C. L., Leff J. W., Knight R., et al (2013). Diversity, distribution and sources of bacteria in residential kitchens. Environ. Microbiol. 15, 588–596. doi: 10.1111/1462-2920.12036. CrossRef. Google Scholar.
29. FMR (2019). Hydroponics Market: Modernistic Cultivation Methodologies Adopted to Prevent the Contamination of Vegetables to Gain Perennial Popularity: Global Industry Analysis 2013-2016 and Opportunity Assessment 2017-2025. Valley Cottage, NY: FMR. Google Scholar.
30. Gentleman R. C., Carey V. J., Bates D. M., Bolstad B., Dettling M., Dudoit S., et al (2004). Bioconductor: open software development for computational biology and bioinformatics. Genome Biol. 5:R80. doi: 10.1186/gb-2004-5-10-r80. CrossRef. Google Scholar.
31. Gil M. I., Selma M. V., Suslow T., Jacxsens L., Uyttendaele M., Allende A. (2015). Pre- and Postharvest Preventive Measures and Intervention Strategies to Control Microbial Food Safety Hazards of Fresh Leafy Vegetables. Crit. Rev. Food Sci. Nutr. 55, 453–468. doi: 10.1080/10408398.2012.657808. CrossRef. Google Scholar.
32. Graf J., Ledala N., Caimano M. J., Jackson E., Gratalo D., Fasulo D., et al (2021). High-Resolution Differentiation of Enteric Bacteria in Premature Infant Fecal Microbiomes Using a Novel rRNA Amplicon. mBio 12, e03656–20. doi: 10.1128/mBio.03656-20. CrossRef. Google Scholar.
33. Gu G., Ottesen A., Bolten S., Luo Y., Rideout S., Nou X. (2020). Microbiome convergence following sanitizer treatment and identification of sanitizer resistant species from spinach and lettuce rinse water. Int. J. Food Microbiol. 318:108458. doi: 10.1016/j.ijfoodmicro.2019.108458. CrossRef. Google Scholar.
34. Gu G., Ottesen A., Bolten S., Wang L., Luo Y., Rideout S., et al (2019). Impact of routine sanitation on the microbiomes in a fresh produce processing facility. Int. J. Food Microbiol. 294, 31–41. doi: 10.1016/j.ijfoodmicro.2019.02.002. CrossRef. Google Scholar.
35. Harrison J. A., Gaskin J. W., Harrison M. A., Cannon J. L., Boyer R. R., Zehnder G. W. (2013). Survey of Food Safety Practices on Small to Medium-Sized Farms and in Farmers Markets. J. Food Prot. 76, 1989–1993. doi: 10.4315/0362-028X.JFP-13-158. CrossRef. Google Scholar.
36. Ibekwe A. M., Leddy M., Murinda S. E. (2013). Potential Human Pathogenic Bacteria in a Mixed Urban Watershed as Revealed by Pyrosequencing. PLoS One 8:e79490. doi: 10.1371/journal.pone.0079490. CrossRef. Google Scholar.
37. Ilic S., LeJeune J., Lewis Ivey M. L., Miller S. (2017). Delphi expert elicitation to prioritize food safety management practices in greenhouse production of tomatoes in the United States. Food Control 78, 108–115. doi: 10.1016/j.foodcont.2017.02.018. CrossRef. Google Scholar.
38. Jukes T. H., Cantor C. R. (1969). "Evolution of protein molecules," in Mammalian protein mnet.abolism, III, ed. Munro H. N. (New York: Academic Press), 21–132. doi: 10.1016/b978-1-4832-3211-9.50009-7. CrossRef. Google Scholar.
39. Kaczmarek M., Avery S. V., Singleton I. (2019). "Chapter Two - Microbes associated with fresh produce: Sources, types and methods to reduce spoilage and contamination," in Advances in Applied Microbiology, eds Gadd G. M., Sariaslani S. (Cambridge, Massachusetts: Academic Press), 29–82. doi: 10.1016/bs.aambs.2019.02.001. CrossRef. Google Scholar.
40. Kinoshita Y., Niwa H., Uchida-Fujii E., Nukada T. (2021). Establishment and assessment of an amplicon sequencing method targeting the 16S-ITS-23S rRNA operon for analysis of the equine gut microbiome. Sci. Rep. 11:11884. doi: 10.1038/s41598-021-91425-7. CrossRef. Google Scholar.
41. Knights D., Kuczynski J., Charlson E. S., Zaneveld J., Mozer M. C., Collman R. G., et al (2011). Bayesian community-wide culture-independent microbial source tracking. Nat. Methods 8, 761–763. doi: 10.1038/nmeth.1650. CrossRef. Google Scholar.
42. Kopylova E., Navas-Molina J. A., Mercier C., Xu Z. Z., Mahé F., He Y., et al (2016). Open-Source Sequence Clustering Methods Improve the State Of the Art. mSystems 1, e00003–15. doi: 10.1128/mSystems.00003-15. CrossRef. Google Scholar.
43. Lahti L., Shetty S. A. (2017). Introduction to the Microbiome R Package. Available online at: https://microbiome.github.io/tutorials/ (accessed November 8, 2021). Google Scholar.
44. Larkin M. A., Blackshields G., Brown N. P., Chenna R., McGettigan P. A., McWilliam H., et al (2007). Clustal W and Clustal X version 2.0. Bioinformatics 23, 2947–2948. doi: 10.1093/bioinformatics/btm404. CrossRef. Google Scholar.
45. Lee J., Kim J., Park S. (2009). Effects of chlorine wash on the quality and microbial population of "Tah Tasai" Chinese cabbage (Brassica campestris var. narinosa) microgreen. Korean J. Hortic. Sci. Technol. 27, 625–630. Google Scholar.
46. Leff J. W., Fierer N. (2013). Bacterial Communities Associated with the Surfaces of Fresh Fruits and Vegetables. PLoS One 8:e59310. doi: 10.1371/journal.pone.0059310. CrossRef. Google Scholar.
47. Leinonen R., Akhtar R., Birney E., Bower L., Cerdeno-Tárraga A., Cheng Y., et al (2011). The European Nucleotide Archive. Nucleic Acids Res. 39, D28–D31. doi: 10.1093/nar/gkq967. CrossRef. Google Scholar.
48. Liao C. H. (2006). "Bacterial soft rot," in Microbiology of fruits and vegetables, eds Sapers G. M., Gorney J. R., Yousef A. E. (Boca Raton: CRC Press), 117–134. doi: 10.1201/9781420038934.sec2. CrossRef. Google Scholar.
49. Liu Y., Cao Y., Wang T., Dong Q., Li J., Niu C. (2019). Detection of 12 Common Food-Borne Bacterial Pathogens by TaqMan Real-Time PCR Using a Single Set of Reaction Conditions. Front. Microbiol. 10:222. doi: 10.3389/fmicb.2019.00222. CrossRef. Google Scholar.
50. Lopez-Galvez F., Allende A., Pedrero-Salcedo F., Alarcon J. J., Gil M. I. (2014). Safety assessment of greenhouse hydroponic tomatoes irrigated with reclaimed and surface water. Int. J. Food Microbiol. 191, 97–102. doi: 10.1016/j.ijfoodmicro.2014.09.004. CrossRef. Google Scholar.
51. Martijn J., Lind A. E., Schön M. E., Spiertz I., Juzokaite L., Bunikis I., et al (2019). Confident phylogenetic identification of uncultured prokaryotes through long read amplicon sequencing of the 16S-ITS-23S rRNA operon. Environ. Microbiol. 21, 2485–2498. doi: 10.1111/1462-2920.14636. CrossRef. Google Scholar.
52. McMurdie P. J., Holmes S. (2013). phyloseq: an R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS One 8:e61217. doi: 10.1371/journal.pone.0061217. CrossRef. Google Scholar.
53. Miao J., Han N., Qiang Y., Zhang T., Li X., Zhang W. (2017). 16SPIP: a comprehensive analysis pipeline for rapid pathogen detection in clinical samples based on 16S metagenomic sequencing. BMC Bioinformatics 18:568. doi: 10.1186/s12859-017-1975-3. CrossRef. Google Scholar.
54. Morgan L. (2019). Organic and Hydroponic Food Safety. Nanaimo, British Columbia: Maximum Yield. Google Scholar.
55. Morgulis A., Coulouris G., Raytselis Y., Madden T. L., Agarwala R., Schäffer A. A. (2008). Database indexing for production MegaBLAST searches. Bioinformatics 24, 1757–1764. doi: 10.1093/bioinformatics/btn322. CrossRef. Google Scholar.
56. NCBI (2021). Pathogen Detection -- NCBI. Available online at: https://www.ncbi.nlm.nih.gov/pathogens (accessed November 8, 2021). Google Scholar.
57. Numberger D., Ganzert L., Zoccarato L., Mühldorfer K., Sauer S., Grossart H.-P., et al (2019). Characterization of bacterial communities in wastewater with enhanced taxonomic resolution by full-length 16S rRNA sequencing. Sci. Rep. 9:9673. doi: 10.1038/s41598-019-46015-z. CrossRef. Google Scholar.
58. Oksanen J., Blanchet F. G., Friendly M., Kindt R., Legendre P., McGlinn D., et al (2020). vegan: Community Ecology Package. Available Online at: https://CRAN.R-project.org/package=vegan (accessed Nov 7, 2021). Google Scholar.
59. Orozco R. L., Iturriaga M. H. (2008). Animal and Environmental Impact on the Presence and Distribution of Salmonella and Escherichia coli in Hydroponic Tomato Greenhouses. J. Food Prot. 71, 676–683. doi: 10.4315/0362-028x-71.4.676. CrossRef. Google Scholar.
60. Pacific Bioscience (2021). Pacific Biosciences SMRT§Tools Reference Guide. Available online at: https://www.pacb.com/wp-content/uploads/SMRT_Tools_Reference_Guide_v10.1.pdf (accessed November 7, 2021). Google Scholar.
61. Pegasus Agriculture Group (2017). Hydroponic Food Produce Tackles Rising Concerns Over Food Safety Issues - Pegasus Agriculture. New York: Cision PR Newswire. Google Scholar.
62. Pruesse E., Peplies J., Glöckner F. O. (2012). SINA: accurate high-throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics 28, 1823–1829. doi: 10.1093/bioinformatics/bts252. CrossRef. Google Scholar.
63. Quast C., Pruesse E., Yilmaz P., Gerken J., Schweer T., Yarza P., et al (2013). The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 41, D590–D596. doi: 10.1093/nar/gks1219. CrossRef. Google Scholar.
64. Ramlal P. S., Lin J., Buckley C. A., Stenström T. A., Amoah I. D., Okpeku M., et al (2021). 16S rRNA-based metagenomic profiling of microbes on contact surfaces within shared sanitation facilities. Ecol. Genet. Genomics 21:100095. doi: 10.1016/j.egg.2021.100095. CrossRef. Google Scholar.
65. Ranjan R., Rani A., Metwally A., McGee H. S., Perkins D. L. (2016). Analysis of the microbiome: advantages of whole genome shotgun versus 16S amplicon sequencing. Biochem. Biophys. Res. Commun. 469, 967–977. doi: 10.1016/j.bbrc.2015.12.083. CrossRef. Google Scholar.
66. Rastogi G., Sbodio A., Tech J. J., Suslow T. V., Coaker G. L., Leveau J. H. J. (2012). Leaf microbiota in an agroecosystem: spatiotemporal variation in bacterial community composition on field-grown lettuce. ISME J. 6, 1812–1822. doi: 10.1038/ismej.2012.32. CrossRef. Google Scholar.
67. Riggio G. M., Jones S. L., Gibson K. E. (2019). Risk of Human Pathogen Internalization in Leafy Vegetables During Lab-Scale Hydroponic Cultivation. Horticulturae 5:25. doi: 10.3390/horticulturae5010025. CrossRef. Google Scholar.
68. Rzhetsky A., Nei M. (1993). Theoretical foundation of the minimum-evolution method of phylogenetic inference. Mol. Biol. Evol. 10, 1073–1095. doi: 10.1093/oxfordjournals.molbev.a040056. CrossRef. Google Scholar.
69. Sawyer T., Savidov N., Pate G., Laberge M. (2020). Food Safety and E. coli in Aquaponic and Hydroponic Systems. Euless, Texas: Urban Ag News. Google Scholar.
70. Schloss P. D., Handelsman J. (2006). Toward a Census of Bacteria in Soil. PLoS Comput. Biol. 2:e92. doi: 10.1371/journal.pcbi.0020092. CrossRef. Google Scholar.
71. Soon J. M., Davies W. P., Chadd S. A., Baines R. N. (2013). Field application of farm-food safety risk assessment (FRAMp) tool for small and medium fresh produce farms. Food Chem. 136, 1603–1609. doi: 10.1016/j.foodchem.2012.01.029. CrossRef. Google Scholar.
72. Srinivasan R., Karaoz U., Volegova M., MacKichan J., Kato-Maeda M., Miller S., et al (2015). Use of 16S rRNA Gene for Identification of a Broad Range of Clinically Relevant Bacterial Pathogens. PLoS One 10:e0117617. doi: 10.1371/journal.pone.0117617. CrossRef. Google Scholar.
73. Stecher G., Tamura K., Kumar S. (2020). Molecular Evolutionary Genetics Analysis (MEGA) for macOS. Mol. Biol. Evol. 37, 1237–1239. doi: 10.1093/molbev/msz312. CrossRef. Google Scholar.
74. U.S. Food and Drug Administration (2011). Food Safety Modernization Act (FSMA). Silver Spring: U.S. Food and Drug Administration. Google Scholar.
75. UCD Centre for Food Safety, Dublin I., Van Hoorde K., Butler F. (2018). Use of next-generation sequencing in microbial risk assessment. EFSA J. 16:e16086. doi: 10.2903/j.efsa.2018.e16086. CrossRef. Google Scholar.
76. UMass Extension (2018). Impatiens -- Leaf spot (Pseudomonas syringae). Center for Agriculture, Food, and the Environment. Massachusetts: University of Massachusetts Amherst. Google Scholar.
77. Waterhouse A. M., Procter J. B., Martin D. M. A., Clamp M., Barton G. J. (2009). Jalview Version 2---a multiple sequence alignment editor and analysis workbench. Bioinformatics 25, 1189–1191. doi: 10.1093/bioinformatics/btp033. CrossRef. Google Scholar.
78. Westcott S. L., Schloss P. D. (2015). De novo clustering methods outperform reference-based methods for assigning 16S rRNA gene sequences to operational taxonomic units. PeerJ 3:e1487. doi: 10.7717/peerj.1487. CrossRef. Google Scholar.
79. Wickham H. (2021). ggplot2: Elegant Graphics for Data Analysis, 3rd Edn. Available online at: https://ggplot2-book.org/ (accessed November 7, 2021). Google Scholar.
80. Yilmaz P., Parfrey L. W., Yarza P., Gerken J., Pruesse E., Quast C., et al (2014). The SILVA and "All-species Living Tree Project (LTP)" taxonomic frameworks. Nucleic Acids Res. 42, D643–D648. doi: 10.1093/nar/gkt1209. CrossRef. Google Scholar.
81. Zhang Z., Schwartz S., Wagner L., Miller W. (2000). A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 7, 203–214. doi: 10.1089/10665270050081478. CrossRef. Google Scholar.
82. Zhong Y., Wang Y., Zhao T., He X., Ke Y., Liu W., et al (2020). Multiplex real-time SYBR Green I PCR assays for simultaneous detection of 15 common enteric pathogens in stool samples. Mol. Cell. Probes 53:101619. doi: 10.1016/j.mcp.2020.101619. CrossRef. Google Scholar.
83. Zwirzitz B., Wetzels S. U., Dixon E. D., Stessl B., Zaiser A., Rabanser I., et al (2020). The sources and transmission routes of microbial populations throughout a meat processing facility. NPJ Biofilms Microbiomes 6, 1–12. doi: 10.1038/s41522-020-0136-z. CrossRef. Google Scholar.

Dong M and Feng H (2022) Microbial Community Analysis and Food Safety Practice Survey-Based Hazard Identification and Risk Assessment for Controlled Environment Hydroponic/Aquaponic Farming Systems. Front. Microbiol. 13:879260. doi: 10.3389/fmicb.2022.879260
Перевод статьи «Microbial Community Analysis and Food Safety Practice Survey-Based Hazard Identification and Risk Assessment for Controlled Environment Hydroponic/Aquaponic Farming Systems» авторов Dong M and Feng H., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык
Фото: Designed by Freepik



















































































































Комментарии (0)