Микробиота филлосферы сахарного тростника: сравнительный анализ органического, переходного и традиционного земледелия
Перевод обычных ферм на органические системы для улучшения здоровья экосистемы является актуальной тенденцией последних десятилетий, однако до сих пор мало изучено, в какой степени и каким образом этот процесс влияет на таксономическое разнообразие и функциональную способность надземных микроорганизмов. В связи с этим данное исследование было проведено с целью изучения влияния различных систем земледелия (органической, переходной и традиционной) на структуру и функцию микробного сообщества филлосферы сахарного тростника с использованием подхода дробовик-метагеномики.
Аннотация
Сравнительный метагеномный анализ показал, что методы ведения сельского хозяйства сильно влияют на таксономическое и функциональное разнообразие, а также на взаимодействия (коокурренцию) микробов филлосферы. В органическом земледелии наблюдалась сложная микробная сеть с наибольшей связностью, что указывает на высокую устойчивость её микробного сообщества к динамическим экологическим стрессорам. Органическое земледелие также характеризовалось наличием рода Streptomyces в качестве потенциального ключевого (эдификаторного) вида и бактерий, стимулирующих рост растений, в качестве микробных сигнатур, включая Mesorhizobium loti, Bradyrhizobium sp. SG09, Lactobacillus plantarum и Bacillus cellulosilyticus. Интересно, что многочисленные виды, разлагающие токсичные соединения, были особенно обогащены при переходном земледелии, что может указывать на их важную роль в процессе трансформации от традиционного к органическому хозяйствованию. Более того, традиционная практика снижала содержание генов, связанных с подвижностью клеток и энергетическим метаболизмом микробов филлосферы, что могло негативно способствовать снижению микробного разнообразия в этой среде обитания. В совокупности наши результаты продемонстрировали реакцию микробиоты филлосферы сахарного тростника на конкретные методы управления сельским хозяйством, которые играют жизненно важную роль в устойчивом производстве сахарного тростника.
Введение
За последние десятилетия традиционная практика ведения сельского хозяйства, которая в основном полагается на агрохимикаты, значительно увеличила мировое производство сельскохозяйственной продукции на душу населения (Pretty, 2008). Однако эти усилия не только повышают производственные затраты, но и вызывают серьезные проблемы для окружающей среды, включая загрязнение и деградацию почв, выбросы удобрений и пестицидов, утрату биоразнообразия, а также ряд негативных последствий для здоровья человека (Rivera et al., 2017). Таким образом, необходимы альтернативные методы для улучшения сельскохозяйственных показателей или снижения зависимости от агрохимикатов и уравновешивания продуктивности с устойчивостью.
Перевод традиционных ферм на системы органического управления, известный как переходное земледелие, потенциально может стать решением для улучшения биоразнообразия и устойчивого увеличения производства продуктов питания (Gonthier et al., 2014). Однако остается вопросом, сколько времени требуется для восстановления утраченного биоразнообразия и функциональности, вызванных неустойчивыми методами ведения хозяйства. Органическое земледелие — это один из видов сельскохозяйственной практики, направленный на снижение потребности в химических веществах для улучшения общего здоровья экосистемы. Оно основано на использовании животного навоза, зеленых удобрений, биологической борьбе с вредителями, а также некоторых методах, включая севооборот и многокультурье, для повышения питательных веществ почвы и плодородия культур (Pimentel et al., 2005). По сравнению с традиционным земледелием, органическое земледелие гораздо ближе к естественным экосистемам, поскольку оно меньше зависит от агрохимикатов и в большей степени опирается на системы рециклинга питательных веществ. Следовательно, было показано, что такое хозяйствование более эффективно снижает негативное воздействие сельскохозяйственной практики на окружающую среду и способствует сохранению биоразнообразия (Bengtsson et al., 2005). Предыдущие исследования продемонстрировали, что органическое земледелие может способствовать более высокому разнообразию подземных микробных сообществ (например, в почве и ризосфере) по сравнению с традиционным земледелием (Lupatini et al., 2017; Hartman et al., 2018). Другое исследование также показало, что органическое удобрение улучшает количество генов микробного азотного (N) цикла, которые связаны с доступностью азота в почве (Morugán-Coronado et al., 2019).
Филлосфера — это надземная часть растительных органов, в которой доминируют листья, и она плотно заселена многочисленными микробными таксонами, такими как бактерии, грибы и дрожжи (Müller и Ruppel, 2014). Известно, что микробное сообщество филлосферы участвует в круговороте питательных веществ, например, углерода (Knief et al., 2012) и азота (Fürnkranz et al., 2008), а также в деградации загрязнителей воздуха (Ali et al., 2012). Считается, что микроорганизмы, обитающие в этой среде, играют важную роль в здоровье растений и продуктивности экосистем (Laforest-Lapointe et al., 2017). Например, микробные колонизаторы могут стимулировать рост своего хозяина за счет производства фитогормонов (Taghavi et al., 2009), защиты от патогенов листьев (Ritpitakphong et al., 2016) и повышения устойчивости к стрессам (Redman et al., 2002). Кроме того, как открытая система, среда филлосферы может легко подвергаться инвазии экзогенных микроорганизмов, передающихся через атмосферу, почву и насекомых (Vorholt, 2012; Maignien et al., 2014), включая фитопатогены. Таким образом, поддержание гомеостаза микробиоты филлосферы будет критически важным для выживания растения-хозяина (Chen et al., 2020). Учитывая все эти особенности, использование полезной микробиоты филлосферы в качестве новых альтернатив биоудобрениям, биопестицидам или биостимуляторам представляется перспективной целью для улучшения фитосанитарного состояния и устойчивости окружающей среды, что ведет к устойчивому сельскому хозяйству для обеспечения постоянно растущего мирового населения. Однако всестороннее понимание разнообразия и функциональности микробиоты филлосферы в ответ на переход в сельском хозяйстве все еще требует выяснения и будет важным шагом для достижения этих целей.
Сахарный тростник (Saccharum officinarum L.) — это C4-растение, которое имеет примерно на 50% более высокую эффективность фотосинтеза, чем C3-растения (Kajala et al., 2011). Это также одна из наиболее экономически важных культур, используемых в качестве основного источника для производства сахара и биоэтанола (Waclawovsky et al., 2010). Это растение выращивается в тропических и субтропических странах, причем Бразилия является крупнейшим мировым производителем, за ней следуют Индия, Китай и Таиланд (FAO, 2018). Однако для повышения урожайности сахарного тростника обычно применяются высокие дозы синтетических азотных удобрений, а также пестициды для защиты растений от инфекций фитопатогенов (Robinson et al., 2011), что приводит к экологическому ущербу. Кроме того, предыдущие исследования микробиома сахарного тростника были в основном сосредоточены на подземных частях (например, почве, ризосфере и корнях) с использованием ампликонного секвенирования, такого как области штрихкодирования 16S рРНК и ITS, которые ограничены таксономической классификацией на уровне рода и подвержены искажениям в процессе амплификации ПЦР. Более того, важная роль микробного сообщества филлосферы изучена меньше, чем других ниш, особенно в связи с методами управления сельским хозяйством, несмотря на их потенциальную пользу для продуктивности сельского хозяйства.
Предыдущие исследования продемонстрировали, что органическое земледелие способствует более высокому видовому богатству грибного микробиома филлосферы пшеницы по сравнению с традиционным земледелием (Karlsson et al., 2017). Более того, различные агрохимические воздействия в органических и традиционных хозяйствах могут прямо и/или косвенно влиять на микробное сообщество филлосферы. Karlsson et al. (2014) показали, что фунгициды негативно влияют на состав грибного сообщества на листьях пшеницы. Perazzolli et al. (2014) сообщили, что богатство и разнообразие бактериальных и грибных сообществ листьев винограда изменялись под действием пенконазольного фунгицида и агента биологической борьбы (Lysobacter capsici AZ78). Аналогично, Cernava et al. (2019) показали, что методы борьбы с вредителями и патогенами, такие как обработка пестицидами и биологическими средствами, формируют микробную популяцию листьев чая. Кроме того, Xiang et al. (2020) предположили, что функциональные признаки микроорганизмов филлосферы более чувствительны к землепользованию и антропогенному воздействию по сравнению с микробиотой почвы. Таким образом, мы выдвинули гипотезу о том, что различные методы ведения сельского хозяйства влияют на таксономическое разнообразие и функциональную способность надземного микробного сообщества и способствуют формированию различных микробных профилей с уникальными сигнатурами.
Чтобы глубже понять, как таксономическое разнообразие и функциональная способность микробного сообщества филлосферы формируются под влиянием методов ведения сельского хозяйства, мы провели дробовик-метагеномное секвенирование листьев сахарного тростника, собранных в органическом хозяйстве, где в качестве удобрений применялись биокомпост и свиной навоз, и в традиционном хозяйстве, которое долгое время обрабатывалось агрохимикатами. Мы также включили переходное хозяйство, которое было переведено на органическую систему за 2 года до сбора наших образцов, чтобы наблюдать, как микробы филлосферы участвуют в этом процессе трансформации. Мы стремились наблюдать структурные изменения микробиоты филлосферы в зависимости от типов сельскохозяйственного управления и выявить полезные микроорганизмы, которые потенциально могут быть использованы в качестве биостимуляторов для замены агрохимикатов с целью перехода от традиционных методов к органическим системам для более устойчивого ведения сельского хозяйства.
Материалы и методы
Район исследования, сбор и хранение образцов
В этом исследовании листья сахарного тростника (возраст растений: 10 месяцев; фаза созревания и дозревания) были отобраны в апреле 2019 года в трех хозяйствах с различными методами ведения сельского хозяйства в Таиланде. Эти места отбора проб включали органическое хозяйство (OP) в Rai Sukphoang, район Chom Bueng, провинция Ratchaburi (13°35′42.5″N 99°34′58.0″E); переходное хозяйство (TP) в Rai Sarot, Rang Bua, район Chom Bueng, провинция Ratchaburi (13°37′52.8″N 99°33′10.7″E); и традиционное хозяйство (CP) в Rai Pramote, район Photharam, провинция Ratchaburi (13°44′34.2″N 99°54′16.6″E) (Supplementary Table 1 и Supplementary Figure 1). Всего было собрано девять образцов сахарного тростника сорта Khon Kaen 3 (KK3) по три повтора (три разных побега сахарного тростника с одного поля) на каждое хозяйство. Согласно системе нумерации листьев Kuijper (Kuijper, 1915), были выбраны листья с номерами от +1 до +4, при этом листовое влагалище и кончик листа были удалены с помощью поверхностно-стерилизованных ножниц. Все образцы затем помещали в стерильный пластиковый пакет с застежкой-молнией и хранили в прохладе во время полевого сбора, транспортировали при 4°C, а затем замораживали при -80°C до использования для последующих процессов.
Выделение ДНК
Образцы листьев сахарного тростника измельчали с помощью ступок и пестиков в жидком азоте для ускорения процесса измельчения. Микробную ДНК филлосферы из 0,2 г листового порошка экстрагировали с использованием набора innuPREP Plant DNA Kit (AnalytikJena AG, Берлин, Германия) в соответствии с инструкциями производителя. ДНК элюировали в буфере Tris-EDTA (TE), а затем хранили при -80°C для последующих этапов. Образцы тотальной геномной ДНК были качественно и количественно оценены с помощью гель-электрофореза и спектрофотометра NanoDrop (Thermo Scientific, Wilmington, DE, Соединенные Штаты) соответственно. Полученные образцы микробной ДНК филлосферы включали как эпифитную, так и эндофитную микробную ДНК.
Приготовление библиотек и секвенирование
Образцы микробной ДНК филлосферы были отправлены на секвенирование в службу дробовик-метагеномного секвенирования ZymoBIOMICS® (Zymo Research Corp., Irvine, CA, Соединенные Штаты) при содействии S. M. Chemical Supplies Co., Ltd., Таиланд. Кратко, библиотека метагеномного секвенирования была приготовлена из до 100 нг геномной ДНК с использованием набора Nextera® DNA Flex Library Prep Kit (Illumina, San Diego, CA, Соединенные Штаты) в соответствии с инструкциями производителя. Библиотека была помечена внутренними двухиндексными 8-основными штрихкодами и адаптерами Nextera® (Illumina, San Diego, CA, Соединенные Штаты), затем количественно оценена с использованием TapeStation® (Agilent Technologies, Santa Clara, CA, Соединенные Штаты) и объединена в равных пропорциях. Полученный пул ДНК был измерен с помощью количественной ПЦР (кПЦР). Кроме того, отрицательный контроль (т.е. контроль приготовления пустой библиотеки) использовался в процессе приготовления метагеномной библиотеки. Наконец, 11 метагеномных библиотек, включая образец-имитатор (ZymoBIOMICSTM Microbial Community DNA Standard II, номер по каталогу D6311), были секвенированы на платформе Illumina HiSeq 1500 с парноконцевым секвенированием (2 × 100 п.н.).
Метагеномный анализ
Контроль качества и удаление ДНК хозяина
Для максимизации качества наборов данных были выполнены следующие шаги. Сначала последовательности чтений были проверены на качество с использованием FastQC версии 0.11.9 (Andrews, 2010) и MultiQC версии 1.8 (Ewels et al., 2016). Кроме того, оставшиеся адаптеры и низкое качество чтений с показателем Phred менее 15 были обрезаны, а короткие чтения менее 50 п.н. были отброшены с использованием Trimmomatic версии 0.39 (Bolger et al., 2014) со следующими параметрами: NexteraPE-PE.fa:2:30:10:2:keepBothReads, SLIDINGWINDOW 4:15 и MINLEN: 50. Высококачественные чтения затем были выровнены по общедоступному геному сахарного тростника (Saccharum spontaneum cultivar AP85-441: номер доступа GenBank: GCA_003544965.1, загружено по состоянию на 24 марта 2020 г.) с использованием Bowtie2 версии 2.3.5.1 (Langmead и Salzberg, 2012) для идентификации и фильтрации последовательностей-загрязнителей хозяина.
Таксономическое профилирование и анализ разнообразия
Последовательности, не принадлежащие хозяину, были таксономически классифицированы с использованием Kraken2 версии 2.0.8-beta (Wood et al., 2019) против пользовательской базы данных Kraken2, состоящей из архейных, бактериальных, грибных, плазмидных и вирусных последовательностей (загружено из NCBI RefSeq по состоянию на 15 августа 2020 г.). Bracken версии 2.5.0 (Lu et al., 2017) дополнительно использовался для переоценки микробной численности на видовом уровне. Таблица микробной численности, полученная в результате таксономического профилирования метагенома с помощью Kraken2 + Bracken, использовалась в качестве входных данных для анализа разнообразия. Они выполнялись на программе R версии 3.6.2 (R Core Team, 2019) с пакетами vegan версии 2.5-6 (Oksanen et al., 2019), ggplot2 версии 3.3.0 (Wickham, 2016) и phyloseq версии 1.28.0 (McMurdie и Holmes, 2013). Кратко, однофакторный дисперсионный анализ (ANOVA) был вычислен на основе индекса Шеннона для определения видового разнообразия и индекса Пиелу для количественной оценки видовой выравненности в каждом образце (α-разнообразие). Кроме того, различия в общих микробных профилях между методами ведения хозяйства (β-разнообразие) были оценены с помощью неметрического анализа главных координат (PCoA) с использованием расстояния Брея-Кертиса. Статистическая значимость β-разнообразия была затем определена с помощью теста PERMANOVA с 999 перестановками, после чего следовал тест на многомерную однородность групповых дисперсий (BETADISPER) для проверки того, являются ли исследуемые группы гомогенно распределенными по отношению к их микробным таксонам. Географическое положение (район) также было включено в эти статистические оценки β-разнообразия как потенциальный смешивающий фактор. Кроме того, адекватность выборки была количественно оценена путем расчета индекса покрытия Гуда и построения графика α-разрежения.
Функциональная аннотация метагенома
Функциональный анализ метагенома был проведен с помощью поиска по сходству последовательностей с использованием SqueezeMeta версии 1.2.0 с функцией "sqm_reads.pl" (Tamames и Puente-Sánchez, 2019). Кратко, последовательности чтений были функционально классифицированы по базе данных ортологии Kyoto Encyclopedia of Genes and Genomes (KEGG) (KO), загруженной по состоянию на 18 августа 2020 г. (Kanehisa и Goto, 2000; Kanehisa et al., 2016), с использованием алгоритма DIAMOND (blastx) (Buchfink et al., 2015) с параметрами по умолчанию (максимальное E-значение = 1e-03 и минимальное процентное совпадение = 50). Наконец, функция fun3 в программе SqueezeMeta была использована для функциональных назначений. Кроме того, только последовательности, которые ранее были идентифицированы как археи, бактерии, грибы и вирусы, были сохранены для функционального анализа. Таким образом, другие последовательности, которые не могли быть идентифицированы ни по таксономии, ни по функции, были отброшены.
Сгенерированная таблица численности KO впоследствии была предварительно обработана в соответствии с рабочим процессом предыдущего исследования (Yurgel et al., 2019). Сначала матрица количеств была нормализована с помощью метода Reads Per Kilobase per Genome equivalent (RPKG) с использованием MicrobeCensus версии 1.1.0 (Nayfach и Pollard, 2015). Затем MinPath (Ye и Doak, 2009) и функция "pathway_pipeline.py" в PICRUSt2 версии 2.3.0-b (Douglas et al., 2020) были использованы для отображения нормализованной таблицы KO на пути KEGG с настройкой "--no_regroup". Предсказанные пути были дополнительно вручную сгруппированы в уровни иерархии KEGG BRITE, называемые категорией KEGG и путем KEGG. Кроме того, генное разнообразие на уровне KO было оценено с использованием индекса Шеннона. Функциональное β-разнообразие было рассчитано путем проецирования данных на график неметрического PCoA с использованием несходства Брея-Кертиса, после чего следовали тесты PERMANOVA с 999 перестановками и BETADISPER, в которых методы ведения хозяйства и районы были основными и смешивающими эффектами соответственно. Кроме того, однофакторный дисперсионный анализ (ANOVA) был использован для статистической проверки того, влияют ли методы ведения хозяйства на метаболические процессы на уровне категории KEGG и пути KEGG.
Идентификация основных признаков и анализ дифференциальной численности
Основные таксономические и функциональные метагеномы были количественно оценены с использованием функции "core_members" в пакете R microbiome версии 1.6.0 (Lahti и Shetty, 2019) со следующими критериями: detection = 0.001 и prevalence = 95%. Кроме того, дифференциальная численность таксономических и функциональных метагеномов среди методов ведения хозяйства была статистически проверена с использованием метода линейного дискриминантного анализа (LDA) effect size (LEfSe), реализованного в LEfSe версии 1.0.8 (Segata et al., 2011). LEfSe использует непараметрический ранговый тест Крускала-Уоллиса для обнаружения любых признаков с дифференциальной численностью между группами. Биологическая согласованность затем проверяется с помощью набора попарных тестов с использованием непарного теста ранговых сумм Уилкоксона. Наконец, размер эффекта значительно различающихся признаков оценивается с помощью теста LDA. В этом исследовании параметры LEfSe, такие как оценка LDA и значение p Уилкоксона, были установлены на 2,0 и 0,05 соответственно. Результаты были затем визуализированы в виде столбчатых графиков с использованием пакета R ggplot2 версии 3.3.0 (Wickham, 2016).
Построение микробной сети совместного присутствия
Микробная сеть совместного присутствия была построена на основе алгоритма Sparse Correlations for Compositional data (SparCC) (Friedman и Alm, 2012) с использованием FastSpar версии 0.0.10 (Watts et al., 2018). Матрица микробной численности, в которой строки и столбцы представляют таксоны и образцы соответственно, была использована в качестве входных данных и была предварительно обработана следующим образом. Сначала данные были разделены на три подмножества в зависимости от их происхождения (органическое, переходное и традиционное). Микробные виды (признаки), присутствующие менее чем в 2 из 3 образцов и с численностью менее 11, были удалены из таблицы количеств. Чтобы избежать систематических ошибок сравнения, вызванных неравным количеством признаков после этапа фильтрации, для анализа сети было случайным образом отобрано по 500 признаков из каждого отфильтрованного набора данных. Эмпирические коэффициенты корреляции SparCC (значение p) были рассчитаны с помощью метода бутстрапа с 1000 итераций, и затем были построены три сети совместного присутствия, специфичных для каждого метода ведения хозяйства. Cytoscape версии 3.8.0 (Shannon et al., 2003) была использована для визуализации сетей со следующими критериями: сильная корреляция (|r| > 0,6) и значение p < 0,05. Кроме того, алгоритм Гирвана-Ньюмана был выполнен для поиска модулей сети с использованием приложения Cytoscape clusterMaker2 версии 1.3.1 (Morris et al., 2011), в то время как другие топологические характеристики сети были исследованы с использованием доступных функций в пакете R igraph версии 1.2.4.2 (Csardi и Nepusz, 2006). Микробные таксоны (узлы) со степенью > 10 и промежуточной центральностью < 0,1 считались узловыми видами, как объяснено в предыдущем исследовании (Berry и Widder, 2014). Атрибуты узлов каждой сети были количественно оценены с помощью метода бутстрапа с 10 000 итераций и впоследствии сравнены с помощью двухвыборочного теста Колмогорова-Смирнова с использованием функции "ks.test" в пакете stats R.
Результаты
Общая информация о наборе метагеномных данных
Всего с помощью платформы Illumina было получено 118 535 073 необработанных последовательности чтений из 9 метагеномных библиотек дробовик-секвенирования, в диапазоне от 9 892 628 до 29 216 274 чтений на образец. Средняя длина парноконцевых чтений составляла 100 п.н. со средним показателем Phred выше 30. После фильтрации качества, т.е. удаления контаминации хозяина и обрезки по качеству оснований, было отброшено около 93% чтений, осталось 8 176 818 высококачественных последовательностей для последующего анализа (Supplementary Table 2). Кроме того, около 250 000 чтений могли быть таксономически идентифицированы как принадлежащие к архейным, бактериальным, грибным и вирусным видам.
Таксономический состав микробиоты филлосферы сахарного тростника
Таксономическое профилирование метагенома на основе чтений с помощью Kraken2 с последующей переоценкой микробной численности с использованием Bracken выявило в общей сложности 6 572 микробных таксона во всех образцах. В частности, было обнаружено 5 488 бактериальных видов (OP: 5 255; TP: 4 968; и CP: 4 945), 331 грибной вид (OP: 330; TP: 330; и CP: 331), 287 архейных видов (OP: 272; TP: 252; и CP: 251) и 466 вирусных видов (OP: 319; TP: 218; и CP: 189). На уровне доменов бактерии (70,6%) были наиболее распространенными микробными колонизаторами, населяющими среду филлосферы сахарного тростника, за которыми следовали эукариотические, т.е. грибные (27%), архейные (1,8%) и вирусные (0,6%) сообщества (Figure 1A). На уровне типа Proteobacteria, Ascomycota, Actinobacteria, Firmicutes, Bacteroidetes, Basidiomycota, Cyanobacteria, Mucoromycota, Euryarchaeota и Tenericutes были десятью наиболее распространенными микробными типами (Figure 1B).
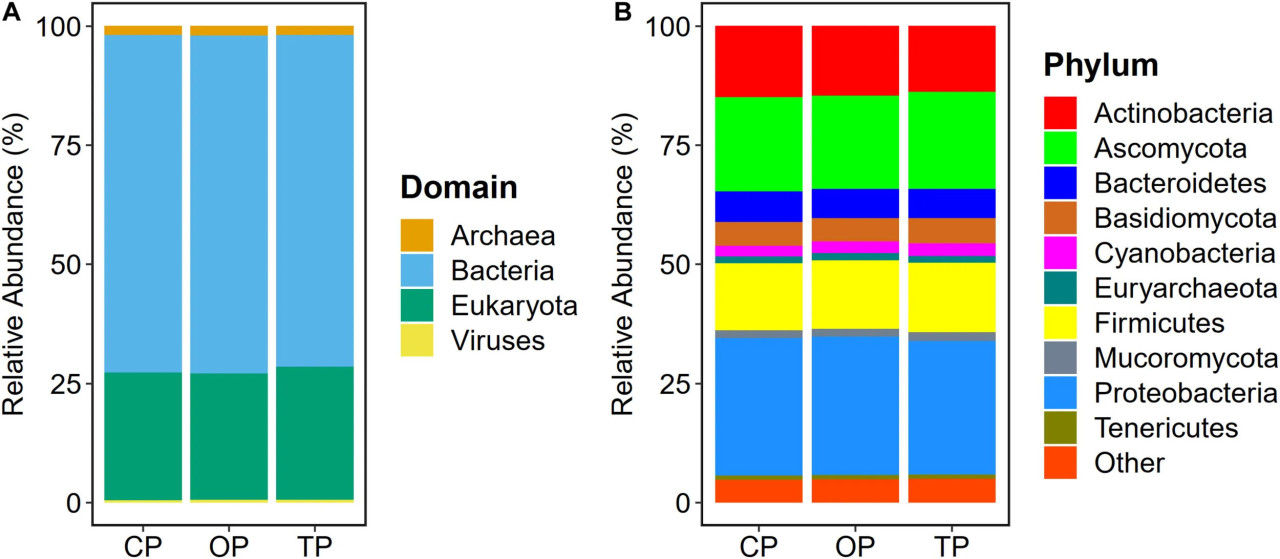
FIGURE 1 Таксономический состав микробиоты филлосферы сахарного тростника. Относительная численность на уровне (A) доменов и (B) типов в традиционном (CP), органическом (OP) и переходном (TP) методах ведения хозяйства.
Функциональные потенциалы микробиоты филлосферы сахарного тростника
Всего во всех образцах было обнаружено 3 797 ортологов KEGG (KO), состоящих из 185 путей KEGG и 36 категорий KEGG. Генные функции, связанные с метаболизмом, были наиболее распространенными, составляя 69,4% от всех аннотированных последовательностей (Supplementary Figure 2). В частности, метаболизм аминокислот (12,1%), энергетический метаболизм (11%), метаболизм углеводов (9,4%), биосинтез и метаболизм гликанов (8,1%), биосинтез других вторичных метаболитов (7,3%), метаболизм терпеноидов и поликетидов (5,3%), метаболизм липидов (4,8%) и метаболизм кофакторов и витаминов (4,8%) были среди основных функциональных категорий KEGG (Supplementary Figure 3). На уровне пути KEGG путь деградации других гликанов (ko00511) был преобладающим, за ним следовали фотосинтез (ko00195), биосинтез валина, лейцина и изолейцина (ko00290), биосинтез фенилпропаноидов (ko00940), биосинтез аминоацил-тРНК (ko00970), окислительное фосфорилирование (ko00190) и биосинтез ансамицинов (ko01051), дополняя доминирующие пути с численностью выше 2% (Supplementary Figure 4).
Методы ведения хозяйства значительно индуцируют различные таксономические и функциональные β-разнообразия
Перед анализом таксономического разнообразия количество чтений было разрежено до 19 000 для каждого образца, чтобы избежать систематических ошибок сравнения, вызванных неравным размером библиотек (Supplementary Figure 5). После этого процесса средний показатель покрытия Гуда составил 95% (Supplementary Table 3), что указывает на то, что размер выборки все еще был достаточен для последующего анализа. Видовое разнообразие было самым высоким в органическом земледелии и постепенно снижалось в переходном и традиционном хозяйствах соответственно (Figure 2A), в то время как индекс выравненности показал противоположную тенденцию, будучи самым низким в органическом земледелии (Figure 2B). Аналогично, функциональное разнообразие на уровне генов с использованием индекса Шеннона также показало, что органическое земледелие обладало наибольшим микробным функциональным разнообразием по сравнению с двумя другими методами (Figure 2C). Однако статистической значимости для оценки α-разнообразия не наблюдалось.
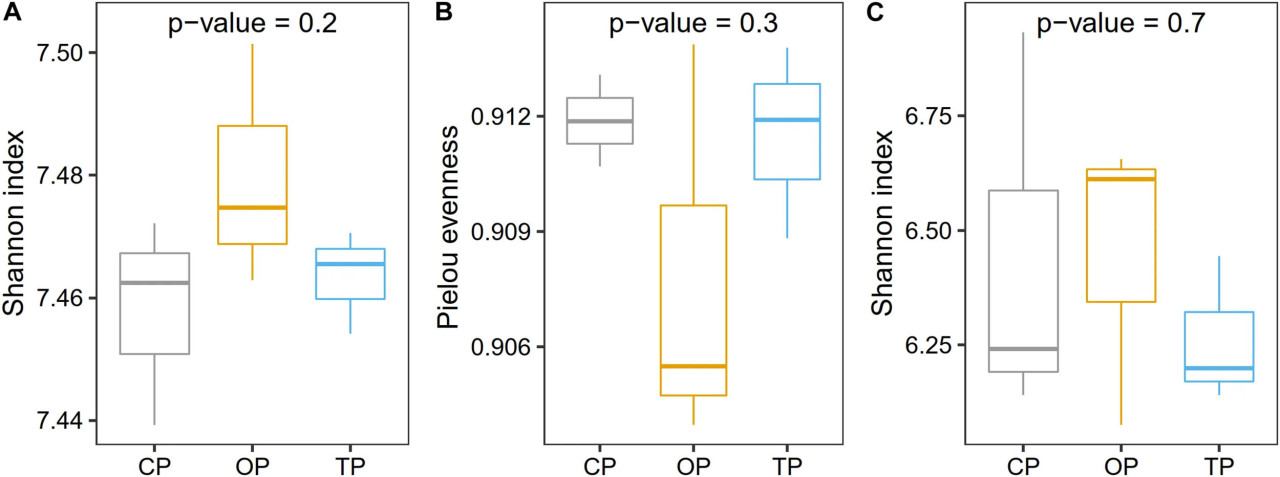
FIGURE 2 Влияние методов ведения хозяйства на таксономическое α-разнообразие, определенное с использованием (A) индекса Шеннона и (B) выравненности Пиелу, и (C) микробное генное разнообразие, оцененное с помощью индекса Шеннона. Статистический анализ проведен с помощью однофакторного дисперсионного анализа (ANOVA).
β-Разнообразие было оценено путем построения неметрического PCoA и выполнения теста PERMANOVA на основе несходства Брея-Кертиса на уровне видов и KO для таксономии и функционального потенциала соответственно. Графики PCoA показали, что микробные сообщества и их генные функции четко разделены вдоль первой главной координаты в соответствии с типами методов ведения хозяйства (Figure 3). Результаты PERMANOVA дополнительно подтвердили наблюдаемые различия между тремя микробными средами обитания (R2 = 0,27, p < 0,01 и R2 = 0,28, p < 0,05 для таксономического и функционального разнообразия соответственно) (Supplementary Table 4). Однако значимый результат теста BETADISPER в таксономическом профиле (p < 0,01), но не в функциональном потенциале (p > 0,05), показал, что различия в таксономическом β-разнообразии могут также незначительно зависеть от внутригрупповой дисперсии (Supplementary Table 4). Анализ PERMANOVA также предоставил топ-50 наиболее влиятельных таксонов, которые потенциально способствуют дифференциации таксономического профиля в зависимости от методов ведения хозяйства (Supplementary Figure 6). Кроме того, мы также обнаружили значительное влияние района (географического положения) по результатам теста PERMANOVA как для таксономического β-разнообразия (R2 = 0,14, p < 0,05), так и для функционального β-разнообразия (R2 = 0,17, p < 0,05) (Supplementary Table 4).
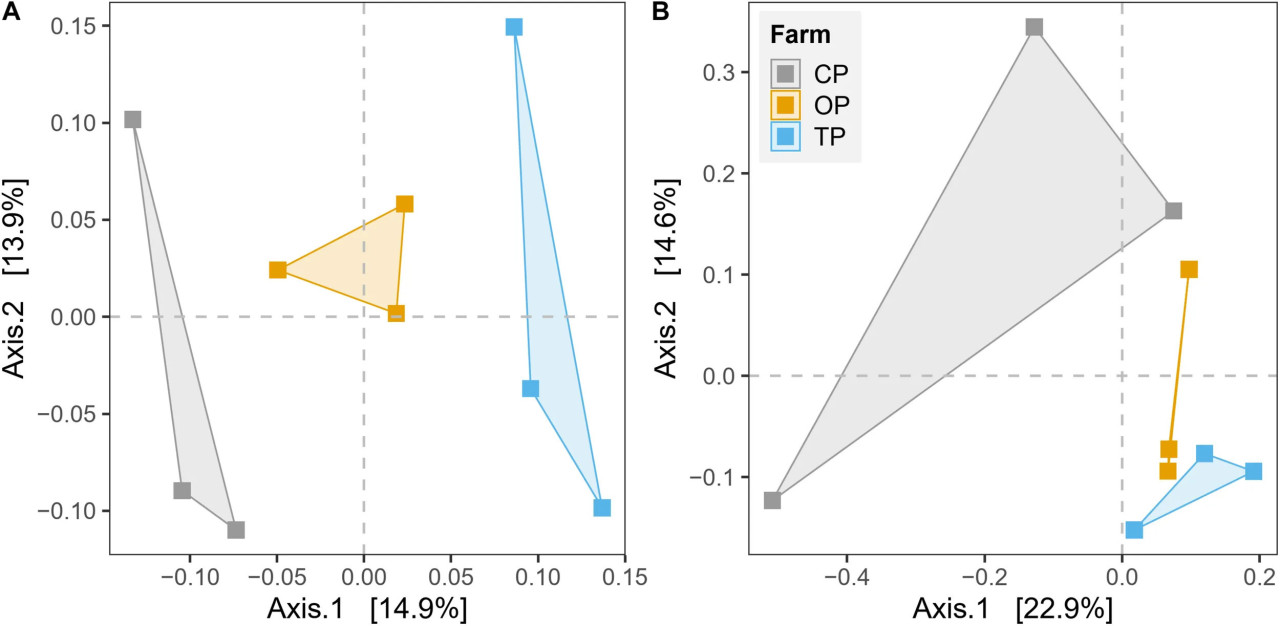
FIGURE 3 Неметрический анализ главных координат (PCoA) на основе расстояния Брея-Кертиса для оценки влияния методов ведения сельского хозяйства на (A) таксономическое разнообразие и (B) функцию генов.
Подвижность клеток и энергетический метаболизм значительно снижены при традиционном земледелии
Функциональный анализ метагенома на уровне категории KEGG и пути KEGG показал, что численность генов микробов, связанных с подвижностью клеток и энергетическим метаболизмом, значительно различалась в зависимости от методов ведения хозяйства. В частности, численность генов, отвечающих за подвижность клеток, была самой высокой в органическом земледелии (ANOVA, p < 0,05). Что касается энергетического метаболизма, то переходное земледелие было наиболее выраженным (ANOVA, p < 0,05), особенно путь фотосинтеза (ko00195) (ANOVA, p < 0,05). В отношении метаболизма аминокислот как переходное, так и органическое хозяйства показали несколько более высокие показатели, чем традиционное (ANOVA, p > 0,1). В частности, путь биосинтеза валина, лейцина и изолейцина (ko00290) был значительно преобладающим в органическом земледелии и самым низким в традиционном (ANOVA, p < 0,05). Кроме того, в традиционном хозяйстве наблюдалось слегка большее количество микробных генов, связанных с метаболизмом углеводов (ANOVA, p > 0,1), биодеградацией и метаболизмом ксенобиотиков (ANOVA, p < 0,1) и инфекционными заболеваниями: бактериальными (ANOVA, p > 0,1) (Figure 4 и Supplementary Figures 3, 4).
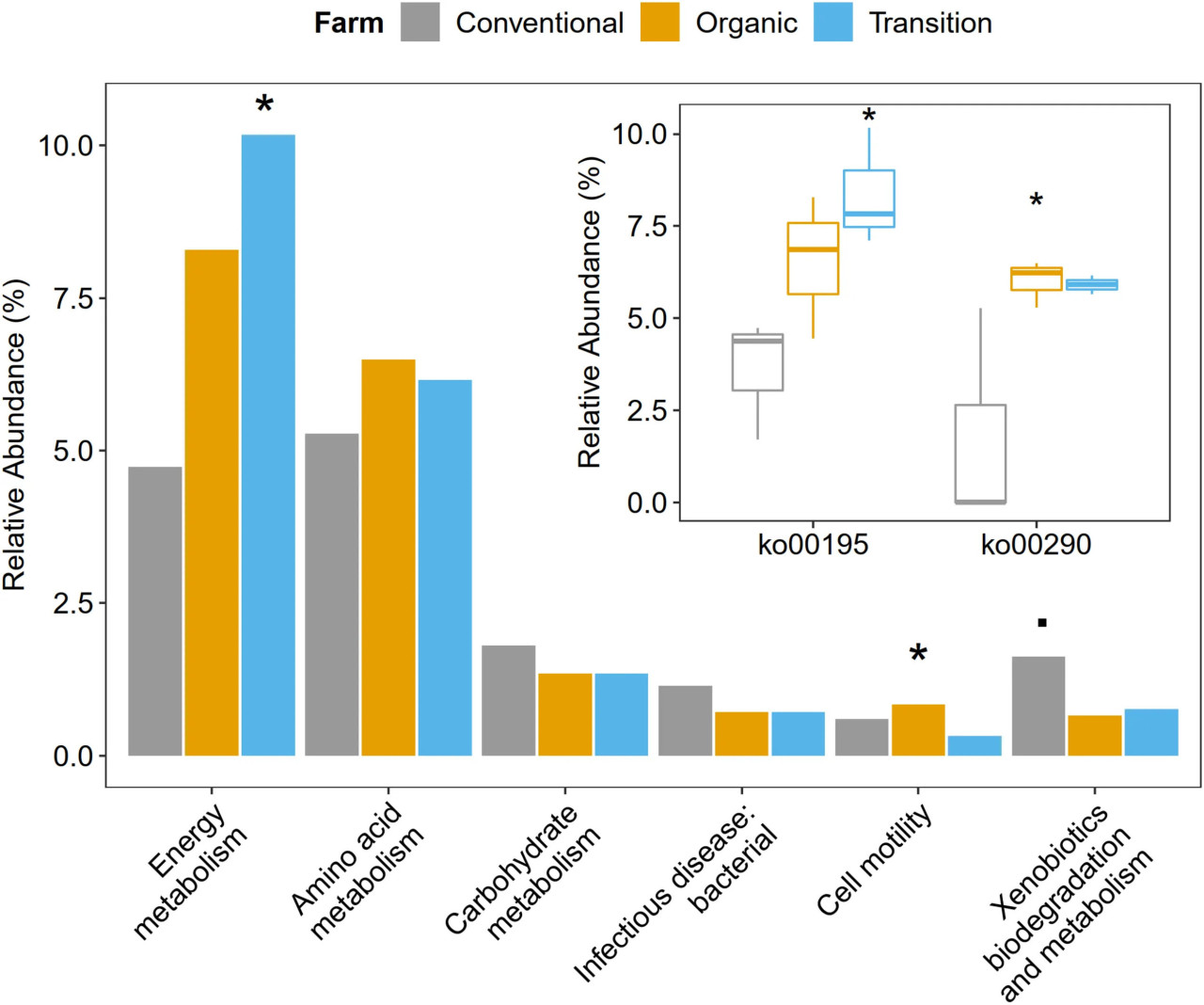
FIGURE 4 Влияние методов ведения хозяйства на метаболические процессы на втором и третьем уровнях иерархии KEGG. Статистический тест проведен с использованием однофакторного дисперсионного анализа (ANOVA), где значения p < 0,05 считались значимыми и обозначены символом (), в то время как символ (.) отражает значение p < 0,1.
Органическое земледелие содержит сложную микробную сеть филлосферы с Streptomyces в качестве основного узлового таксона
Анализ сети совместного присутствия показал, что органическое земледелие обладало наибольшей связностью сети (Figure 5), количеством ребер и плотностью сети (Supplementary Table 5). Аналогично, количество идентифицированных узловых видов было самым высоким в органическом земледелии (19 видов) по сравнению с 6 и 2 видами в переходном и традиционном хозяйствах соответственно (Supplementary Figure 7). Узловые виды органической сети были представлены 4 бактериальными видами, принадлежащими к роду Streptomyces, за которыми следовали азотфиксирующие бактерии [т.е. продуцирующая каротиноиды Calothrix sp. 336/3 (Kosourov et al., 2016), Paraburkholderia sp. CCGE1002 (Ormeño-Orrillo et al., 2012) и Azoarcus sp. DD4 (Deng et al., 2019)], деградирующая полисахариды Lacinutrix sp. 5H-3-7-4 (Klippel et al., 2011), деградирующая 1,4-диоксан Mycobacterium dioxanotrophicus (He et al., 2017), водород-продуцирующая Ethanoligenens harbinense (Li et al., 2019) и аммоний-окисляющая архея Candidatus Nitrosocosmicus franklandus (Lehtovirta-Morley et al., 2016). В переходной сети шесть узловых видов включали патогенный для грибов Janthinobacterium agaricidamnosum (Graupner et al., 2015), патогенный для человека Raoultella ornithinolytica (Morais et al., 2009), продуцирующую охратоксин A Aspergillus homomorphus (Varga et al., 2011), рибофлавин-продуцирующую Eremothecium gossypii (Ledesma-Amaro et al., 2015), Curtobacterium sp. csp3 и Micromonospora tulbaghiae, в то время как два бактериальных вида — деградирующая нефтяные углеводороды Pedobacter cryoconitis (Margesin et al., 2003) и Flavobacterium branchiophilum — были узловыми в традиционной сети.
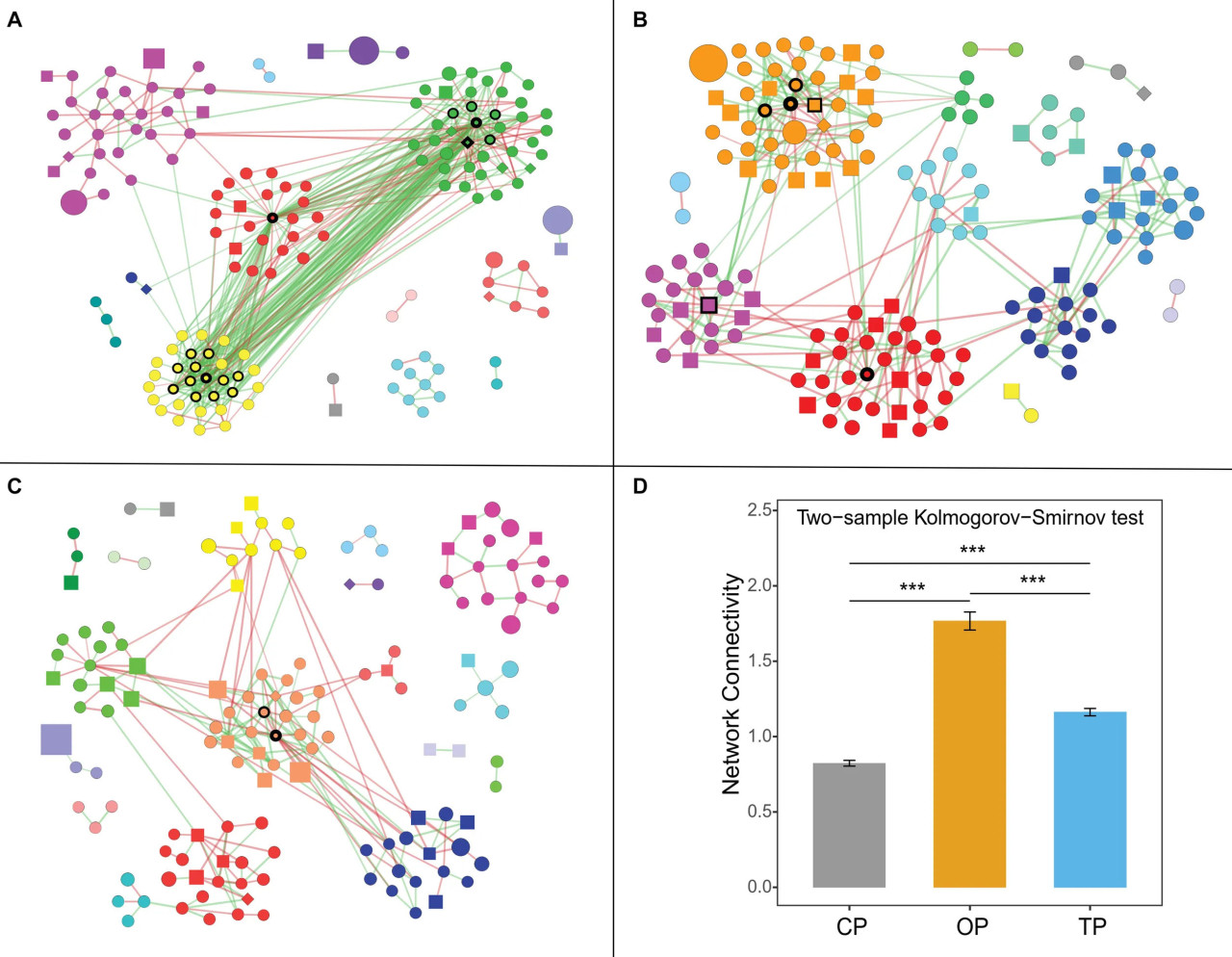
FIGURE 5 Сети совместного присутствия, специфичные для методов ведения хозяйства: (A) органическое, (B) переходное и (C) традиционное. Зеленый и красный цвета ребер представляют положительные и отрицательные взаимосвязи соответственно, в то время как цвет узлов демонстрирует модули сети. Форма вершин отражает микробные домены (т.е. круг: бактерии; квадрат: грибы; и ромб: археи), а размер узлов указывает на численность. Кроме того, узлы с черными границами определены как узловые таксоны, а ширина границы отражает показатель степени узла. Примечание: визуализированы только узлы, имеющие хотя бы одну связь. (D) Связность сети или степень узла представляет количество ребер, соединенных с узлом. Символ () обозначает статистическую значимость с p < 0,0001.
Полезные микроорганизмы были идентифицированы как основные виды, а методы ведения хозяйства индуцируют специфический микробный состав
Основные микробы были идентифицированы путем отбора наиболее распространенных (detection) и вездесущих (prevalence) таксонов во всех образцах. Мы наблюдали 28 бактерий и 32 гриба в качестве основных видов филлосферы сахарного тростника, что составляет 0,9% от общего числа наблюдаемых таксонов и 19,4% от общей относительной численности (Supplementary Figure 8). Эти основные бактериальные виды состояли из бактерий, стимулирующих рост растений [т.е. Bacillus cereus (Zhang et al., 2019), B. thuringiensis (Raddadi et al., 2008), Pseudomonas aeruginosa (Radhapriya et al., 2015), Rhodopseudomonas palustris (Xu et al., 2018) и Methylobacterium radiotolerans (Eevers et al., 2015)], продуцирующей антибиотики Streptomyces venezuelae (Kim et al., 2019), целлюлолитической Sorangium cellulosum (Wang et al., 2012), фотосинтезирующей Scytonema sp. NIES-4073 (Will et al., 2019), деградирующей катехол P. resinovorans (Nojiri et al., 2002), продуцирующей индолилуксусную кислоту (IAA) Acinetobacter baumannii (Lin et al., 2018) и нескольких патогенов, связанных с животными и человеком. Основные грибные таксоны были представлены четырьмя видами Aspergillus, включая Aspergillus niger, A. neoniger, A. thermomutatus и A. heteromorphus, за которыми следовали два вида Fusarium, такие как Fusarium proliferatum и F. oxysporum. Как и в случае с основными бактериальными видами, несколько патогенных для человека грибов, а также фитопатогены также входили в состав основных видов филлосферы сахарного тростника.
Помимо идентификации общих видов филлосферы сахарного тростника, мы также исследовали, могут ли методы ведения хозяйства способствовать увеличению численности определенных таксонов в качестве их представителей (микробных сигнатур). В этом случае анализ LEfSe (оценка LDA > 2,0) был использован для статистического обнаружения любых микробных видов, численность которых значительно различалась между методами ведения хозяйства (таксоны, специфичные для метода ведения хозяйства). В отношении бактериального сообщества переходное земледелие имело наибольшее количество обогащенных таксонов (16 видов), в то время как органическое и традиционное хозяйства одинаково содержали по 7 уникальных бактериальных видов (Figure 6). Эти бактериальные виды включали бактерии, стимулирующие рост растений, деградаторы ксенобиотиков и патогены. Для домена архей было идентифицировано семь видов, включая три, связанных с традиционным земледелием, два с органическим и два с переходным (Figure 6). Кроме того, было обнаружено только два грибных таксона, по одному виду как в органическом, так и в переходном хозяйствах (Figure 6). Однако грибов, специфичных для традиционного земледелия, не наблюдалось.
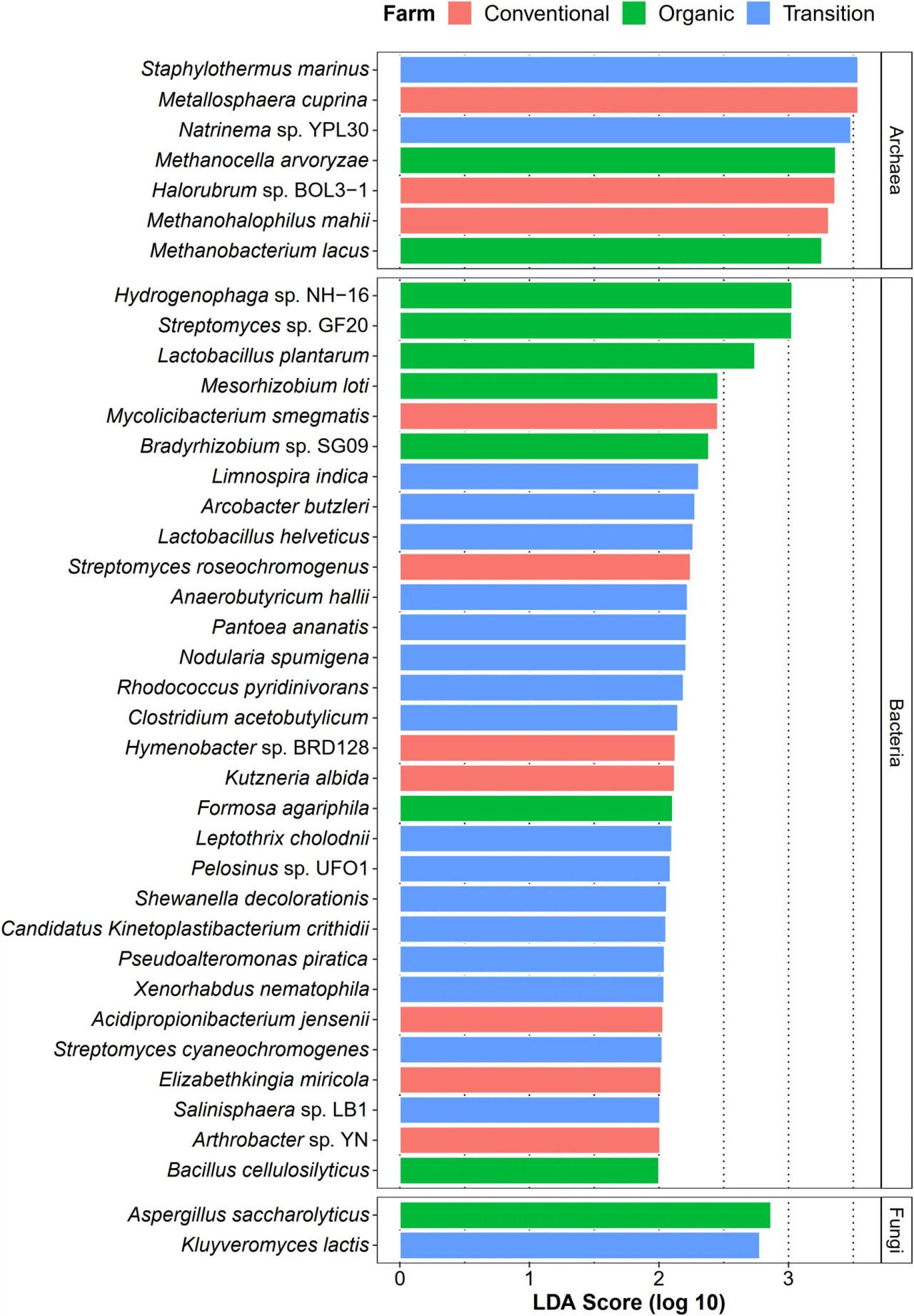
FIGURE 6 Анализ LEfSe таксономической численности среди органического (зеленый), переходного (синий) и традиционного (красный) методов ведения хозяйства на видовом уровне.
Высококонсервативные и уникальные функции генов микробов наблюдались в каждом методе ведения хозяйства
Аналогично, мы также заметили, что микробное сообщество филлосферы сахарного тростника имело общие функции во всех трех методах ведения хозяйства. В частности, 32 KO были идентифицированы как основные функции из всех образцов (Supplementary Figure 9). Среди них гены, связанные с окислительным фосфорилированием и фотосинтезом, были наиболее распространенными, включая альфа и бета субъединицы F-типа H+/Na+-транспортирующей АТФазы (K02111 и K02112), альфа субъединицу F-типа H+-транспортирующей АТФазы (K02132), субъединицу 1 цитохром с-оксидазы (K02256), апо-белок A1 и A2 P700 хлорофилл a фотосистемы I (K02689 и K02690), белок D1 реакционного центра P680 фотосистемы II (K02703), нитритредуктазу (NADH)-убихинон-оксидоредуктазу цепь 5 (K03883), NADH-дегидрогеназу (убихинон) железо-серный белок 2 (K03935), субъединицу 2 NAD(P)H-хинон-оксидоредуктазы (K05573) и большую субъединицу рибулозо-бисфосфат-карбоксилазы (K01601). Гены, участвующие в стрессовых реакциях, также входили в состав основных функций, такие как белок теплового шока 70 кДа 1/2/6/8 (K03283), молекулярный шаперон DnaJ (K03686), молекулярный шаперон HtpG (K04079) и проницаемый для кальция катионный канал, активируемый стрессом (K21989).
Что касается микробных функций, специфичных для методов ведения хозяйства, всего было идентифицировано 31 уникальных KO с помощью программы LEfSe с оценкой LDA > 2,0, включая 12 KO, связанных с органическим земледелием, 11 с традиционным и 8 с переходным (Supplementary Figure 10). На уровне пути KEGG путь бактериальной хемотаксиса (ko02030) и сигнальный путь hedgehog (ko04340) были уникально обогащены в органическом земледелии, в то время как три пути, специфичных для традиционного земледелия, включали амебиаз (ko05146), метаболизм азота (ko00910) и MAPK-сигнальный путь дрожжей (ko04011) (Figure 7). Однако в переходном земледелии не было обнаружено значимо обогащенных путей.
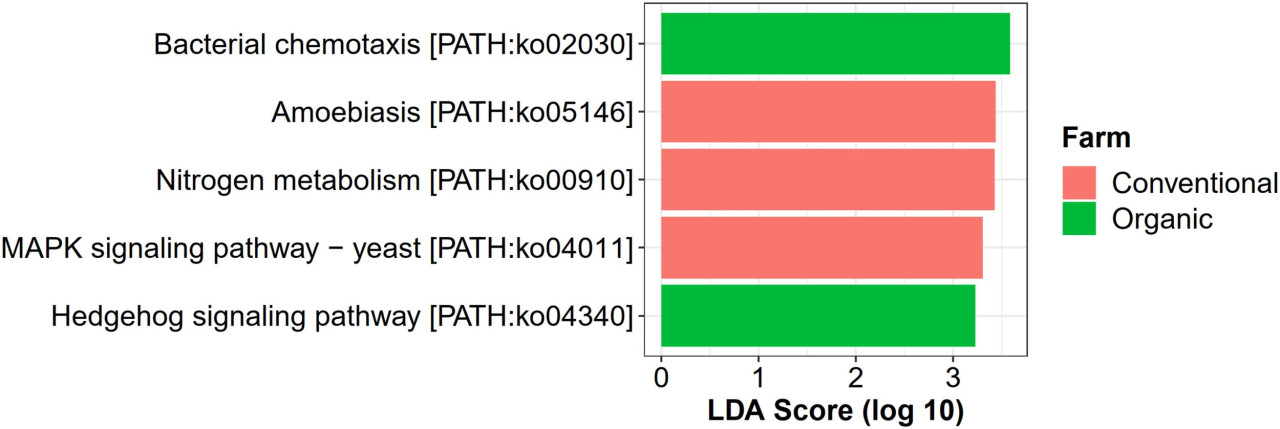
FIGURE 7 Анализ LEfSe функциональной численности на основе пути KEGG во всех методах ведения хозяйства.
Обсуждение
Филлосфера описывается как надземная часть растительных органов, в которой доминируют листья (Turner et al., 2013). Эта среда обитания плотно заселена до 1 × 10^8 бактериальных клеток на грамм свежего веса листьев (Remus-Emsermann et al., 2014), и предполагается, что эти ассоциированные микроорганизмы имеют большое значение для выживаемости растений-хозяев (Vorholt, 2012; Peñuelas и Terradas, 2014). В настоящем исследовании дробовик-метагеномное секвенирование было использовано для изучения влияния перехода в сельском хозяйстве на таксономическое разнообразие и функциональную способность микробного сообщества филлосферы сахарного тростника. Этот метод напрямую секвенирует всю геномную ДНК в образце, что обеспечивает минимально искаженную количественную оценку абсолютной численности микроорганизмов.
Таксономическое профилирование на основе чтений показало, что состав микробного сообщества филлосферы был в основном представлен бактериальными типами Proteobacteria, Actinobacteria и Firmicutes, а также грибными типами Ascomycota и Basidiomycota во всех трех методах ведения хозяйства. Преобладающие бактериальные и грибные типы, наблюдаемые в этой работе, были аналогичны ранее опубликованному исследованию сахарного тростника с использованием ампликонов 16S рРНК и ITS (Hamonts et al., 2018), а также на других видах растений, включая томат (Toju et al., 2019) и виноград (Perazzolli et al., 2014). Эти результаты подтверждают более ранние данные о высококонсервативных микробных сообществах филлосферы у разных видов растений-хозяев (Massoni et al., 2020).
Анализ разнообразия показал, что методы ведения хозяйства значительно влияют на таксономические и функциональные профили микробиоты филлосферы (β-разнообразие). Органическое земледелие было немного более разнообразным (индекс Шеннона), чем переходное и традиционное хозяйства, в то время как этот метод также имел самую низкую видовую выравненность (индекс Пиелу). Теоретически, более разнообразные микробные сообщества потенциально повышают устойчивость растения-хозяина к инвазии патогенов при поддержке полезных микробных симбионтов (Kinkel et al., 2011). Streptomyces sp. GF20 и Lactobacillus plantarum были идентифицированы как микробные сигнатуры органического земледелия с помощью LEfSe, и эти бактерии также входили в топ-100 наиболее распространенных таксонов во всех органических образцах. Это указывает на то, что эти виды могли способствовать снижению индекса выравненности за счет своего доминирования, что могло бы быть полезным для продуктивности хозяина. Примечательно, что аналогичная реакция наблюдалась для микробного сообщества почвы в предыдущем исследовании (Hartmann et al., 2014), что указывает на то, что надземные микроорганизмы реагируют на методы ведения сельского хозяйства так же, как и микроорганизмы, колонизирующие подземные части. Индекс разнообразия Шеннона для функциональных потенциалов также имел тенденцию к увеличению в органическом земледелии по сравнению с двумя другими методами. Однако статистической значимости для таксономического и функционального α-разнообразий обнаружено не было. Кроме того, было обнаружено, что район также оказывает значительное влияние как на таксономическое, так и на функциональное β-разнообразия. Хотя влияние метода ведения хозяйства было сильнее, чем влияние района, на основании значения p PERMANOVA, метод ведения хозяйства не был единственным фактором, определяющим структуру и функциональные потенциалы микробных сообществ филлосферы.
Считается, что основной микробиом тесно связан и эволюционировал вместе с наземными растениями в течение длительного времени (Yeoh et al., 2017), а его генные функции необходимы для роста и продуктивности растений (Lemanceau et al., 2017). Основные виды филлосферы сахарного тростника включали бактерии, стимулирующие рост растений, включая N2-фиксирующие, антибиотик-продуцирующие и IAA-продуцирующие виды, что подчеркивает важную роль микробов филлосферы в фитосанитарном состоянии растений-хозяев. Многие из них были культивированы in vitro (Raddadi et al., 2008; Radhapriya et al., 2015; Xu et al., 2018; Zhang et al., 2019). Присутствие аммоний-окисляющих, целлюлолитических и фотосинтезирующих бактерий в качестве основных членов микробов филлосферы сахарного тростника дополнительно указывает на их вклад в глобальные циклы азота и углерода, в то время как наличие бактерии, деградирующей катехол, свидетельствует о потенциале филоремедиации. Что касается грибов, представители Aspergillus и Fusarium были наиболее распространенными основными таксонами. Грибные виды Aspergillus изучались на предмет их свойств, стимулирующих рост растений, включая Aspergillus niger, который может растворять фосфор (P) (Mendes et al., 2015) и калий (K) (Lopes-Assad et al., 2010). Напротив, Fusarium является одним из разрушительных фитопатогенов, который может адаптироваться к широкому спектру условий окружающей среды и поражать различные экономически важные культуры по всему миру (Perincherry et al., 2019). Примечательно, что, связывая таксономию и функцию, мы обнаружили, что полезные гены, присущие упомянутым выше основным видам, обычно встречались во всех образцах. Однако некоторые из этих генов были более распространены в органическом и переходном хозяйствах, в то время как в традиционном хозяйстве их численность была снижена. В совокупности эти основные микроорганизмы подчеркивают важную роль микробных сообществ филлосферы в обеспечении роста и продуктивности растений, что ведет к улучшению здоровья всей экосистемы хозяина.
Аналогично предыдущему сообщению о неотропических деревьях (Lajoie et al., 2020), в этой работе также были обнаружены консервативные метаболические функции микробов филлосферы. В частности, метаболизм аминокислот и углеводов был обогащен во всех образцах, что указывает на то, что эти две функции имеют первостепенное значение для микробов филлосферы для доступа к питательным веществам в этой среде обитания (Müller et al., 2016), поскольку полисахариды и белки являются частью листовых экссудатов, выделяемых железистыми трихомами, которые также участвуют в растительно-микробных взаимодействиях (Schlechter et al., 2019). На уровне KO окислительное фосфорилирование, фотосинтез и стрессовые реакции представляли основные функции микробов филлосферы. Таким образом, неудивительно, что фотосинтетические гены были частью основных функций, поскольку многие основные микробные виды были фотосинтезирующими бактериями. Это также позволяет предположить, что микробы филлосферы могут использовать свет в качестве альтернативного источника энергии при дефиците питательных веществ (Vorholt, 2012; Stiefel et al., 2013). Кроме того, сильно динамичные и экстремальные условия среды филлосферы, включая воздействие УФ-излучения и высоких температур, которые меняются в течение всего дня, заставляют микробов филлосферы активировать ряд генов, связанных со стрессовыми реакциями, для успешной колонизации и процветания на поверхности листьев (Vorholt, 2012). В этом исследовании мы обнаружили многочисленные гены, кодирующие белки теплового шока, в качестве основных функций, что указывает на то, что эти гены могут играть критическую роль в адаптации и выживании микробов филлосферы в этой суровой среде обитания. Эти результаты согласуются с предыдущими исследованиями, которые сообщали о консервативных функциях, связанных с фототрофией и микробной адаптацией микробных сообществ филлосферы у различных типов растений (Finkel et al., 2016; Lambais et al., 2017; Flores-Núñez et al., 2020).
Затем был проведен анализ сети совместного присутствия для сравнения структуры и сложности микробных сообществ филлосферы в ответ на методы ведения хозяйства. Оценка топологии сети показала, что органическая сеть имела наибольшее количество ребер, плотность и связность, что может быть связано с высокой устойчивостью микробиоты филлосферы в этом хозяйстве к нарушениям или инвазии экзогенных микроорганизмов (Mendes et al., 2018; Banerjee et al., 2019; Liao et al., 2019). Более того, большая сложность и связность органической сети сопровождались количеством идентифицированных узловых видов, в то время как неустойчивое земледелие демонстрировало обратную картину. Эта закономерность была аналогична предыдущему исследованию микробиома корней пшеницы (Banerjee et al., 2019). Интересно, что три метода ведения хозяйства содержали совершенно разные узловые виды, что иллюстрирует, что переход в сельском хозяйстве меняет центральные таксоны в сообществе. Узловые виды органической сети были представлены Streptomyces spp., которые являются хорошо известными антибиотик-продуцирующими бактериями (Kong et al., 2019). Большинство ребер, связанных с узлами Streptomyces, были положительными (56 положительных ребер и 3 отрицательных ребра), включая меж- и внутривидовые связи. Это указывает на то, что Streptomyces могут сосуществовать и ингибировать рост других микроорганизмов, регулируя состав и структуру органической сети. Hu et al. (2020) предположили, что применение Streptomyces может способствовать росту томата, изменяя микробное сообщество ризосферы, и может положительно влиять на численность других полезных бактерий. Цианобактерия Calothrix sp. 336/3, которая способна фиксировать атмосферный углерод и азот (Kosourov et al., 2016), была наиболее связанным узлом в органической сети, за ней следовали деградирующая полисахариды Lacinutrix sp. 5H-3-7-4 (Klippel et al., 2011) и аммоний-окисляющая архея Candidatus Nitrosocosmicus franklandus (Lehtovirta-Morley et al., 2016). Благодаря своим способностям эти три узловых вида могут занимать более высокий трофический уровень и были положительно связаны с узлами Streptomyces, что объясняет важную роль Streptomyces, которые, следовательно, могут быть определены как потенциальные ключевые виды в органической сети. В переходной сети узловыми видами были бактериальные патогены грибов, растений и человека, в то время как традиционная сеть имела наименьшее количество узловых таксонов, включая деградирующую нефтяные углеводороды Pedobacter cryoconitis (Margesin et al., 2003) и патогенную Flavobacterium branchiophilum (Good et al., 2015). Короче говоря, идентифицированные ключевые виды микробов филлосферы могут играть важную экологическую роль в соответствующих методах ведения хозяйства, что открывает перспективы для будущих исследований. Однако необходима дальнейшая экспериментальная проверка биологических процессов, управляющих микробными сетями, например, с использованием синтетического сообщества.
Анализ дифференциальной численности показал, что традиционная практика индуцирует численность патогенных видов и микроорганизмов, способных переносить суровые условия окружающей среды, в то время как органическое земледелие характеризуется обогащением полезных для растения-хозяина видов. Таксоны, специфичные для традиционного земледелия, включали мобилизующую металлы Metallosphaera cuprina (Liu et al., 2011), устойчивую к экстремальным условиям среды Halorubrum sp. BOL3-1 (DasSarma et al., 2019) и патогенные для человека бактерии, такие как Elizabethkingia miricola (Zdziarski et al., 2017) и Mycolicibacterium smegmatis (Yamada et al., 2018). Таксоны, специфичные для органического земледелия, включали бактерии, стимулирующие рост растений, включая N2-фиксирующие бактерии [т.е. Mesorhizobium loti (Shimoda et al., 2016) и Bradyrhizobium sp. SG09 (Wasai-Hara et al., 2020)], молочнокислую бактерию L. plantarum, которая способна выступать в качестве агента биологической борьбы с фитопатогенами (Daranas et al., 2019), и целлюлазо-деградирующую Bacillus cellulosilyticus (Mead et al., 2013). Количество таксонов, специфичных для переходного хозяйства, было наибольшим, что позволяет предположить, что многие микроорганизмы могут выступать в качестве промежуточных агентов трансформации сельского хозяйства от традиционных методов к органическим системам. Эти виды были представлены в основном деградаторами токсичных соединений, включая деградирующую пиридин Rhodococcus pyridinivorans (Yoon et al., 2000), обесцвечивающую красители Shewanella decolorationis (Xu et al., 2005), восстанавливающую металлы Pelosinus sp. UFO1 (Brown et al., 2014), окисляющую Fe(II) Leptothrix cholodnii (Kunoh et al., 2016), а также фитопатогенную Pantoea ananatis (Stice et al., 2018) и энтеропатогенную Arcobacter butzleri (Prouzet-Mauléon et al., 2006). Одно из возможных объяснений заключается в том, что остатки примененных агрохимикатов в предыдущей практике переходного хозяйства остались в почве, были поглощены растениями и затем распределены по листьям (Myers et al., 2016). В ответ на это численность этих микроорганизмов увеличилась, что могло способствовать очистке этих токсичных веществ. Таким образом, эти полезные микробиомы сахарного тростника могут быть использованы в качестве перспективного подхода для ускорения перехода от традиционного земледелия к органическому управлению. Примечательно, что несколько видов, принадлежащих к роду Streptomyces, были обнаружены во всех методах ведения хозяйства с помощью анализа LEfSe, идентифицированы как основные виды во всех образцах и определены как потенциальные ключевые виды в органической сети. Это позволяет предположить, что Streptomyces spp. являются хорошо адаптированными таксонами в различных условиях и могут играть решающую роль в росте и продуктивности сахарного тростника (Sousa и Olivares, 2016; Wang et al., 2019).
Помимо формирования микробного состава, методы ведения хозяйства также значительно влияли на подвижность клеток и энергетический метаболизм микробов филлосферы. Подвижность клеток была преобладающей в органическом земледелии, особенно бактериальный хемотаксис, что позволяет предположить, что активное движение микробных видов в органическом земледелии было выше, чем в переходном и традиционном хозяйствах. Сигналинг хемотаксиса позволяет подвижным микроорганизмам филлосферы использовать химические градиенты для поиска источников питательных веществ, таких как листовые экссудаты, выделяемые растением-хозяином, или для ухода от вредных соединений (Scharf et al., 2016). Таким образом, ограниченность питательных веществ на поверхности листьев делает это свойство критически важным для эпифитов, чтобы выжить в этой олиготрофной среде обитания (Vorholt, 2012; Scharf et al., 2016). Энергетический метаболизм посредством фотосинтетического пути был самым высоким в переходном земледелии, за ним следовало органическое, и значительно снижался в традиционном земледелии, что позволяет предположить, что для преобразования загрязненных агрохимикатами земель в более здоровые сельскохозяйственные угодья требовалось больше энергии. Это также указывает на то, что применение агрохимикатов, особенно гербицидов, в традиционной практике снижает фотосинтетический потенциал микробных сообществ филлосферы в целом (Broser et al., 2011; Crouzet et al., 2019). Кроме того, удобрение в традиционном хозяйстве также может изменять состав листовых экссудатов (Barunawati et al., 2013), что может не благоприятствовать ассоциированным колонизаторам филлосферы. Кроме того, биосинтез валина, лейцина и изолейцина, также известных как аминокислоты с разветвленной цепью (BCAAs), также был снижен в традиционном земледелии. BCAAs являются глобальными регуляторами транскрипции у бактерий, которые важны для реагирования на доступность питательных веществ и метаболическое перепрограммирование при истощении питательных веществ (Kaiser и Heinrichs, 2018). В совокупности это может быть причиной снижения микробного разнообразия в традиционном земледелии, поскольку некоторые микробы филлосферы теряют эти важные способности для роста и выживания.
Традиционное земледелие, напротив, незначительно увеличивало численность генов биодеградации и метаболизма ксенобиотиков, в то время как органическое земледелие имело самый низкий показатель. Это подтверждает идею о том, что традиционное земледелие, основанное на использовании синтетических агрохимикатов (т.е. гербицидов и NPK-удобрений), создает токсичную среду для микроорганизмов, и в ответ увеличивается количество генов, деградирующих ксенобиотики (Liao et al., 2019). Это также повышает метаболизм азота у микробов филлосферы, включая глутаминсинтетазу (K01915) и большую субъединицу нитритредуктазы (NADH) (K00362), которые были идентифицированы как функции, специфичные для традиционного земледелия, с помощью LEfSe. Последняя, как сообщается, участвует косвенно в производстве оксида азота (NO) через процесс денитрификации (Härtig и Zumft, 1999; Ruiz et al., 2019). Интересно, что гены, связанные с патогенностью для человека, участвующие в пути амебиаза, были специфически обогащены в традиционном земледелии, что подчеркивает вредное влияние неустойчивых методов не только на окружающую среду, но и на здоровье человека.
Хотя было очевидно, что методы ведения хозяйства значительно влияют на таксономическое разнообразие и функциональную способность микробных сообществ филлосферы, наш экспериментальный план, в котором три повторных образца определенного типа хозяйства были собраны с одного поля, не позволял четко разделить эффекты методов ведения сельского хозяйства и площадей сельхозугодий. Поэтому в будущем необходимо изучение влияния географического положения при схожих методах ведения хозяйства, включая свойства почвы, а также экологические и сезонные условия, для выяснения важной роли микробных сообществ филлосферы в переходе к устойчивому сельскому хозяйству.
Заключение
Это исследование показало, что органическое земледелие способствует более высокому микробному разнообразию с более сложной и стабильной структурой сети. В рамках этой практики также было обогащено большее количество бактерий, стимулирующих рост растений, в качестве таксонов, специфичных для данного метода ведения хозяйства. В переходном земледелии преобладали многочисленные деградаторы токсичных соединений, которые могут вносить важный вклад в восстановление поврежденных агроэкосистем, таких как очистка от остатков агрохимикатов. Традиционное земледелие, имеющее самый низкий индекс разнообразия, увеличивало численность устойчивых к экстремальным условиям и патогенных для человека видов. Снижение микробного разнообразия в традиционном земледелии может быть связано с уменьшением подвижности клеток и энергетического метаболизма, которые необходимы для выживания и процветания микробов филлосферы в стрессовых условиях этой среды обитания, включая ограничение питательных веществ. В целом, наши результаты подчеркивают, что изменение методов ведения хозяйства индуцирует изменения в сообществе и различные функциональные профили микробного сообщества филлосферы, демонстрируя потенциальное использование микробиоты филлосферы в качестве сигнатур для улучшения агрохимического сельского хозяйства и перехода к более здоровым и устойчивым системам. Кроме того, наша работа послужила основой для будущих исследований реакции микробиоты филлосферы сахарного тростника на трансформацию сельскохозяйственных земель и перспектив их использования для повышения продуктивности сельскохозяйственных культур и глобального здоровья экосистем.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти онлайн по адресу: https://www.frontiersin.org/articles/10.3389/fmicb.2021.623799/full#supplementary-material
Дополнительный рисунок 1
Карта мест отбора проб, созданная с помощью Google maps.
Дополнительный рисунок 2
Относительная численность основных категорий KEGG, визуализированная с помощью гистограммы в трех различных методах ведения хозяйства.
Дополнительный рисунок 3
Относительная численность категорий KEGG от самой высокой (сверху) до самой низкой (снизу) в трех различных методах ведения хозяйства.
Дополнительный рисунок 4
Топ-25 относительных численностей путей KEGG в трех различных методах ведения хозяйства.
Дополнительный рисунок 5
Графики α-разрежения, построенные на основе численности микробных сообществ в традиционном, органическом и переходном методах ведения хозяйства, представленных красным, зеленым и синим цветами соответственно.
Дополнительный рисунок 6
Топ-50 наиболее влиятельных микробных видов на различия сообществ между методами ведения хозяйства. Значения по оси x основаны на коэффициенте теста PERMANOVA. Виды по оси y (микробные виды), выделенные зеленым цветом, являются микробными сигнатурами органического земледелия, идентифицированными с помощью LEfSe.
Дополнительный рисунок 7
Ключевые виды, наблюдаемые в трех сетях совместного присутствия, специфичных для методов ведения хозяйства. Таблица показывает степень узла каждого таксона. Строки представляют микробные виды, а столбцы — методы ведения хозяйства. Горизонтальные и вертикальные дендрограммы построены методом иерархической кластеризации.
Дополнительный рисунок 8
Основные микробы филлосферы сахарного тростника, отобранные в трех различных методах ведения хозяйства.
Дополнительный рисунок 9
Основные функции генов микробов филлосферы, ассоциированных с сахарным тростником, в органическом, переходном и традиционном методах ведения хозяйства.
Дополнительный рисунок 10
Тест LEfSe функциональной численности на основе ортологии KEGG (KO) в органическом, переходном и традиционном методах ведения хозяйства.
Дополнительная таблица 1
Описание трех методов ведения хозяйства на сахарном тростнике, отобранных в этом исследовании.
Дополнительная таблица 2
Количество чтений до и после процессов контроля качества.
Дополнительная таблица 3
Значения покрытия Гуда для всех образцов в трех методах ведения хозяйства.
Дополнительная таблица 4
Результаты анализа PERMANOVA и BETADISPER.
Дополнительная таблица 5
Атрибуты сети совместного присутствия в трех различных методах ведения хозяйства.

Khoiri AN, Cheevadhanarak S, Jirakkakul J, Dulsawat S, Prommeenate P, Tachaleat A, Kusonmano K, Wattanachaisaereekul S and Sutheeworapong S (2021) Comparative Metagenomics Reveals Microbial Signatures of Sugarcane Phyllosphere in Organic Management. Front. Microbiol. 12:623799. doi: 10.3389/fmicb.2021.623799
Перевод статьи «Comparative Metagenomics Reveals Microbial Signatures of Sugarcane Phyllosphere in Organic Management» авторов Khoiri AN, Cheevadhanarak S, Jirakkakul J, Dulsawat S, Prommeenate P, Tachaleat A, Kusonmano K, Wattanachaisaereekul S and Sutheeworapong S., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык
Фото: Designed by Freepik



















































































































Комментарии (0)