Куда пойдет вредитель? Оценка распространения Salurnis marginella из-за изменения климата
Salurnis marginella наносит ущерб сельскому и лесному хозяйству в различных регионах Азии. Однако, учитывая экологическую адаптивность этого вредителя и активную международную торговлю, в будущем он может проникнуть и в другие регионы. Поскольку ущерб местным сообществам от вредителей после инвазии становится трудно контролируемым, крайне важно разработать меры по минимизации потерь путем превентивного мониторинга и выявления зон повышенного риска, что может быть достигнуто с помощью прогнозов на основе моделей. Целью данного исследования была оценка потенциального распространения S. marginella путем разработки нескольких алгоритмов моделирования распределения видов (SDM).
Аннотация
В частности, мы разработали модель CLIMEX и три модели на основе машинного обучения (MaxEnt, случайный лес и многослойный перцептрон), объединили их для консервативной оценки встречаемости вредителя в текущих и будущих климатических условиях, а также наложили карту распространения растений-хозяев на климатически пригодные для S. marginella территории для выявления зон высокого риска, уязвимых для распространения и инвазии этого вредителя. Разработанная модель, демонстрирующая статистический показатель истинного мастерства (TSS) > 0.8, спрогнозировала потенциальное непрерывное распространение вида на юго-востоке США, в Южной Америке и Центральной Африке. В настоящее время это распространение охватывает примерно 9.53% мировой суши; однако модель предсказала, что эта область сократится до 6.85%. Возможные зоны распространения были выявлены в Азии и на юго-западе США с учетом ареала растений-хозяев. Это исследование предоставляет данные для активного мониторинга вредителя путем выявления территорий, где S. marginella может распространиться.
1. Введение
Инвазия и распространение чужеродных вредителей, а также изменение климата являются глобальными проблемами [1]. Salurnis marginella (Guérin-Méneville 1829) – это вредитель, который повреждает такие деревья, как Castanea crenata, Prunus verecunda, Prunus yedoensis, Quercus acutissima, Quercus serrata и Zanthoxylum schinifolium, высасывая их сок [2,3]. Этот вредитель обычно встречается в Азии, включая Корею, Китай, Малайзию и Индию, и имеет тенденцию к распространению в районы с относительно низкими зимними температурами, поскольку зимует на стадии яйца [2].
Из-за ограниченного ареала в изолированных районах и небольшого количества сообщений об ущербе по всему миру, S. marginella не привлекал к себе большого внимания, что привело к ограниченному количеству исследований по данной теме. Однако существуют опасения относительно этого вредителя, который имеет потенциал к быстрому распространению и нанесению значительного ущерба из-за широкого круга растений-хозяев. Эти опасения дополнительно усиливаются текущими социальными и экологическими изменениями, а способность этого вредителя выдерживать низкие температуры может позволить ему заселять различные регионы. Поэтому данный вид актуален для моделирования распределения видов (SDM), которое направлено на прогнозирование потенциальных местообитаний, предоставление данных для мониторинга и разработку стратегий борьбы с вредителями.
SDM является популярным инструментом для прогнозирования пространственного распределения вредителей в зависимости от переменных окружающей среды [4]. Таким образом, он может предоставить полезную информацию для разработки экономически эффективных стратегий мониторинга и борьбы с вредителями [5,6,7]. Например, риск распространения схожего вредителя Metcalfa pruinosa был изучен для целей борьбы с использованием SDM, что продемонстрировало эффективность этого подхода в определении потенциального ареала вида с ограниченным местообитанием, но высоким потенциалом распространения [8,9]. По сравнению с прошлыми SDM, которые ограничивались разработкой отдельных моделей, современные SDM были улучшены с помощью ансамблевого моделирования и включения множества переменных, полученных из высококачественных экологических данных, что привело к повышению производительности [10,11,12,13]. Таким образом, цель этого исследования состояла в том, чтобы определить потенциальное распространение с помощью различных моделей для эффективного мониторинга и предварительного контроля S. marginella.
Для оценки потенциального распространения S. marginella использовались четыре различные модели SDM: (1) CLIMEX, уникальная механистическая модель для оценки экологической ниши [14]; (2) MaxEnt, широко используемая коррелятивная модель, применяющая теорию максимальной энтропии для оценки физиологической ниши [15,16,17]; (3) случайный лес (RF), универсальный классификатор, основанный на ансамблевом машинном обучении [18]; и (4) многослойный перцептрон (MLP), нейронная сеть глубокого обучения, которая начала применяться для SDM [19,20,21]. Результаты были сравнены с точки зрения распределения и размера прогнозируемой области, а также производительности моделей, чтобы вывести оптимальную ансамблевую модель, которая показала наилучшее предсказание для S. marginella с помощью различных комбинаций моделей.
2. Материалы и методы
2.1. Получение данных о встречаемости Salurnis marginella
S. marginella обычно встречается в Азии, включая Корею, Китай и Малайзию [2,22]. В дополнение к предыдущим исследованиям, фактические записи о встречаемости были изучены через GBIF, который предоставляет геопривязанные места, где был обнаружен целевой вид [2,22,23]. Собранные данные были зарегистрированы с 2015 по 2023 год, всего 182 точки данных, из которых 122 точки данных о распространении, зарегистрированные с 2015 по 2021 год, были использованы для построения модели. Оставшиеся 60 точек данных о распространении за 2022 и 2023 годы были использованы для валидации и тестирования модели. Для минимизации ошибки выборки с помощью ArcGIS Pro версии 3.2.0 (ESRI, Redlands, CA, USA) была выполнена пространственная фильтрация с использованием буфера в 10 км [24]. В результате для использования в построении модели было окончательно отобрано 53 точки распространения S. marginella.
2.2. Экологические данные
Глобальные исторические данные о месячном климате (средняя минимальная температура, средняя максимальная температура и общее количество осадков) с 1992 по 2021 год с разрешением 10 минут (18.5 км на экваторе) были получены из WorldClim (https://www.worldclim.org/) [25,26] и преобразованы в формат данных, подходящий для CLIMEX (mm файл) [14]. Для модели на основе машинного обучения 19 биоклиматических переменных (bio1–bio19) с разрешением 10 минут за период 1992–2021 гг. были получены из WorldClim [25]. Аналогичным образом мы создали mm-файл для климатических сценариев SSP245 и SSP585 на 2050 год, в то время как биоклиматические переменные, соответствующие SSP, были получены напрямую. Климатические данные были преобразованы в модельные слои путем конвертации точек данных, записанных для каждого разрешения пикселя, в растровый формат с использованием ArcGIS Pro. Кроме того, поскольку растения-хозяева играют важную роль в определении ареалов распространения вредителей, мы включили распространение растений-хозяев в модель. Геопривязанные данные о распространении шести растений-хозяев (Castanea crenata, Prunus verecunda, Prunus yedoensis, Quercus acutissima, Quercus serrata и Zanthoxylum schinifolium), собранные до 2023 года, были получены из GBIF для оценки зон риска [27,28,29,30,31,32,33]. Область распространения растений-хозяев была визуализирована путем преобразования в область с использованием круга радиусом 50 км с центром в точке обнаружения шести растений-хозяев (Рисунок 1).
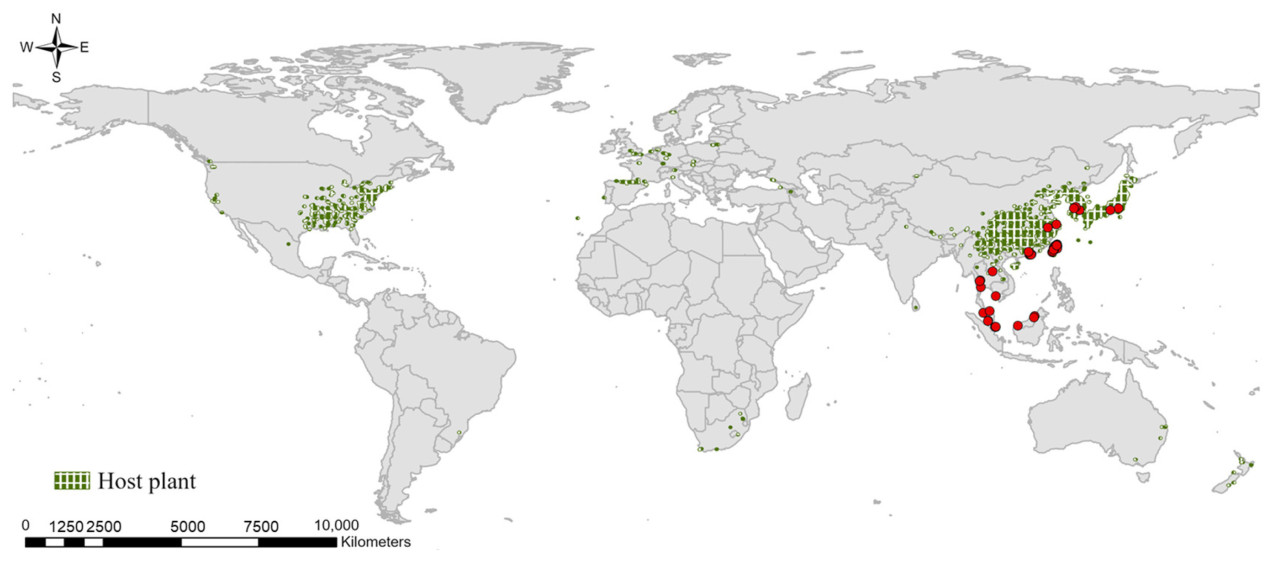
Рисунок 1. Глобальное распространение Salurnis marginella (красные точки) и растений-хозяев, визуализированное с помощью буфера радиусом 50 км вокруг зарегистрированных случаев встречаемости.
2.3. CLIMEX
CLIMEX (версия 4.0, Hearne software, Мельбурн, Австралия) – это механистический инструмент SDM, который оценивает климатическую пригодность территории для вида как функцию его биологии в ответ на региональные метеорологические данные [14]. В модели CLIMEX климатическая пригодность территории для вида количественно оценивается с помощью экологического индекса (EI) по шкале от 0 (непригодно) до 100 (оптимально пригодно) путем учета благоприятных и неблагоприятных климатических условий для роста и стресса вида, основанных на расчетных значениях параметров, отражающих биологические характеристики вида [34,35]. Критически важно оценить параметры, представляющие биологию вида, для работы модели CLIMEX, чтобы симуляция наилучшим образом соответствовала фактическому распределению на основе предыдущих исследований влияния климатических условий на популяции видов и климат в районах встречаемости [34,36,37].
В этом исследовании, из-за ограниченной доступности биологических данных для S. marginella, начальные параметры были получены из предыдущего исследования по моделированию CLIMEX для Metcalfa pruinosa, вида из того же семейства Flatidae, сходство с которым составляет 86% по данным NCBI BLAST [38]. Значения параметров по умолчанию из модели Metcalfa pruinosa были впоследствии скорректированы с использованием климатических данных, извлеченных из координат встречаемости S. marginella, чтобы значения EI соответствовали фактическим регионам распространения этого вредителя [9] (Таблица 1). Температурные параметры были подобраны с учетом климатического распределения Таиланда, где самая высокая среднемесячная максимальная температура среди регионов встречаемости, и Южной Кореи, где самая низкая среднемесячная минимальная температура. Кроме того, параметры, связанные с влажностью, были подобраны с учетом климатического распределения Малайзии, где самое высокое месячное количество осадков среди регионов встречаемости, и Таиланда, где самое низкое месячное количество осадков. Значения EI из модели CLIMEX были спроецированы на мировую карту с использованием ArcMap (версия 10.4.1; ESRI, Redlands, CA, USA) для визуализации потенциального распространения S. marginella. Разработанная модель дала значение EI > 1 для всех 74 координат фактического распространения S. marginella. Затем значения EI были классифицированы по следующим категориям: EI = 0 – непригодно; 0 < EI ≤ 10 – маргинально; 10 < EI ≤ 20 – умеренно; 20 < EI ≤ 30 – высоко; и EI > 30 – оптимально.
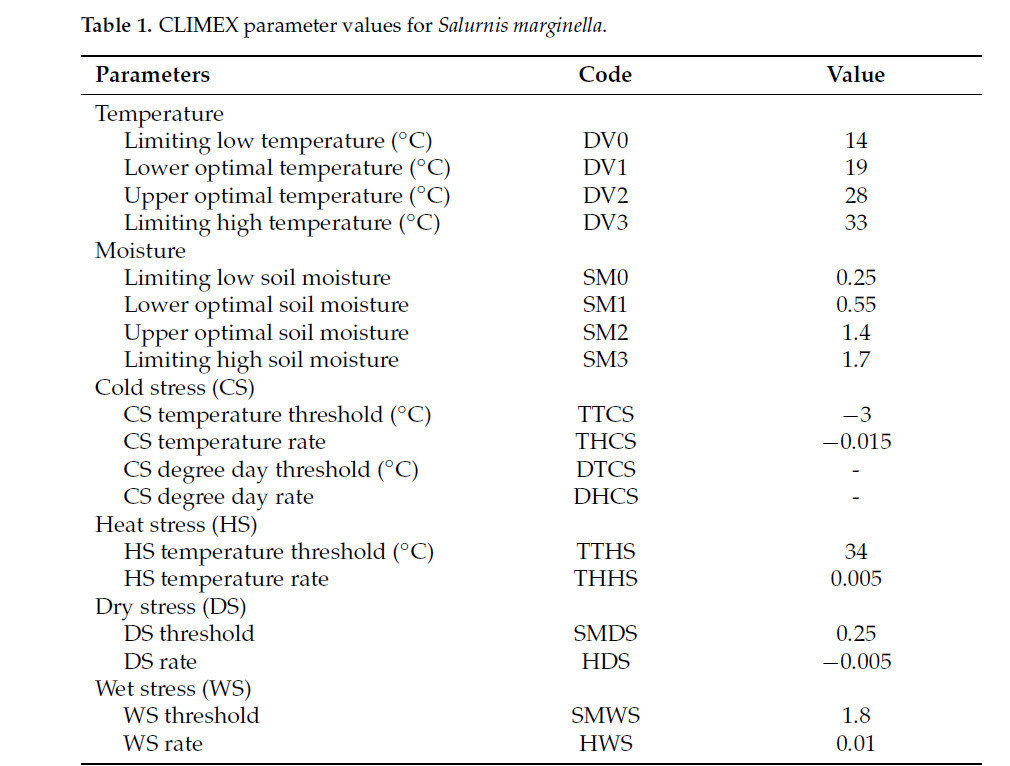
Таблица 1. Значения параметров CLIMEX для Salurnis marginella.
2.4. Модели на основе машинного обучения
Для выбора переменных, применяемых во всех коррелятивных моделях в этом исследовании, мы сначала рассмотрели биологические и экологические характеристики S. marginella. Поскольку предыдущих биологических/физиологических исследований S. marginella не проводилось, мы проанализировали климатические характеристики, извлеченные из районов распространения S. marginella, чтобы получить биоклиматические переменные для использования в модели. Максимальная температура составила 36.2 °C, что было определено по районам, где максимальная температура держалась на уровне 33 °C в течение 4 месяцев, в то время как самая холодная область была подтверждена в районе, где температура опускалась до -4.2 °C и оставалась ниже -3 °C в течение 2 месяцев. Для переменных, связанных с осадками, мы изучили самую влажную область, где максимальное количество осадков составляло 560 мм с устойчивым выпадением более 450 мм в течение 3 месяцев. Напротив, самая сухая область была там, где минимальное количество осадков было ниже 9 мм в течение 3 месяцев. Основываясь на этих условиях, мы сначала выбрали максимальную температуру самого теплого месяца (bio5), минимальную температуру самого холодного месяца (bio6), осадки самого влажного месяца (bio13) и осадки самого сухого месяца (bio14) среди биоклиматических переменных, которые были напрямую связаны с выживанием вида, а переменные с коэффициентом корреляции >0.7 были удалены [13,39]. В результате были отобраны семь биоклиматических переменных: bio2, 5, 6, 8, 13, 14 и 18 (Таблица 2).
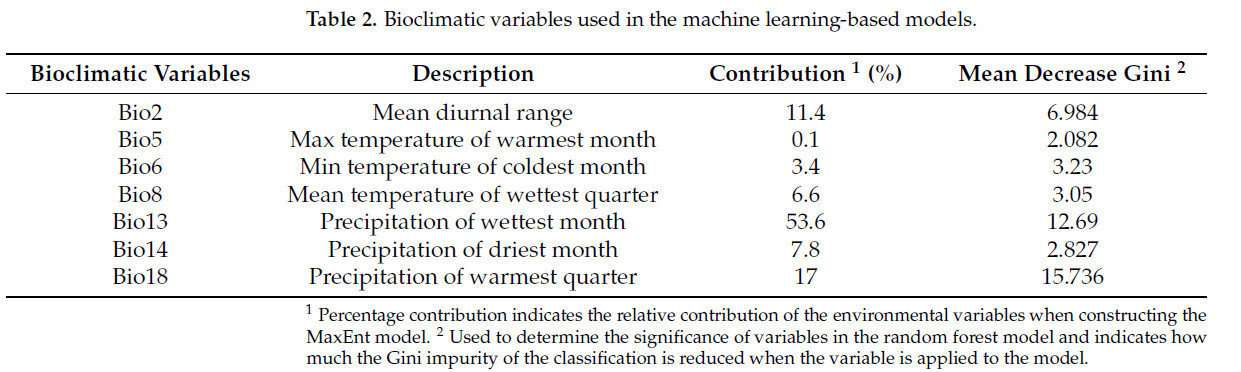
Таблица 2. Биоклиматические переменные, использованные в моделях на основе машинного обучения.
MaxEnt – это инструмент SDM на основе машинного обучения, который оценивает вероятность встречаемости вида на основе его максимальной энтропии [15]. Используя выбранные переменные модели, ENMeval использовался для определения оптимального класса признаков и множителя регуляризации (RM) для запуска модели MaxEnt [40]. Признаки модели представляют условия окружающей среды, которые влияют на прогнозы распределения видов, в то время как множитель регуляризации (RM) регулирует штраф за сложность модели, тем самым балансируя компромисс между подгонкой данных и предотвращением переобучения. Мы установили признаки модели на линейные, а множители регуляризации на 0.5, что привело к наименьшему дельта-скорректированному информационному критерию Акаике (AICc), и работали с 10 000 случайными фоновыми точками, используя 10-кратную перекрестную проверку. Модель была построена с использованием азиатских биоклиматических переменных, чтобы предотвратить переобучение, вызванное координатами распространения, сосредоточенными только в Азии. Затем построенная модель была применена к глобальным климатическим данным для прогнозирования потенциального глобального распространения.
RF – это алгоритм машинного обучения, широко используемый для классификации и регрессии, который применяет ансамблевое обучение, называемое бутстреп-агрегацией (бэггинг), к деревьям решений [18]. В этом исследовании RF использовался для классификации присутствия или отсутствия S. marginella. Набор данных для RF был создан путем объединения случайно выбранных данных о присутствии и отсутствии того же размера, что и данные о присутствии, в координатах по всему миру [41]. Набор данных был случайным образом разделен на 80% для обучения модели и 20% для валидации. Оптимальная структура RF была определена с использованием вышеупомянутой структуры RF. Количество деревьев (ntree) и количество переменных для каждого разбиения (mtry) были установлены на 3 и 500 путем итеративной настройки до достижения наименьшей оценки ошибки вне выборки (OOB error). Модель RF была построена с использованием пакета randomForest в программном обеспечении R версии 4.3.3 [42].
MLP – это базовая структура в искусственных нейронных сетях, и в последнее время она применяется для разработки модели распределения видов [21,43,44]. Модель MLP была создана с использованием того же набора данных, что и для RF, и тех же переменных, что и в MaxEnt, то есть были использованы семь переменных, а также равное количество наборов данных присутствия и отсутствия, разделенных на 80% и 20% для обучения и валидации модели соответственно. Мы систематически изменяли количество нейронов и скрытых слоев с размерами пакетов 16 и 100 эпох до достижения наименьших потерь без переобучения, чтобы определить оптимальную структуру MLP с вышеупомянутыми выбранными переменными. Оптимальная конфигурация состояла из семи скрытых слоев, каждый с функцией активации 'ReLU', в то время как финальный выходной слой, состоящий из двух нейронов, использовал функцию 'softmax' для расчета вероятностей присутствия и отсутствия вида. Конкретная структура была установлена как 14-20-40-20-2, что дало наименьшие потери 0.1319.
2.5. Оценка производительности
Точность, чувствительность, специфичность и истинный статистический навык (TSS) были рассчитаны в процессе разработки модели (валидация модели) для оценки производительности четырех моделей. Кроме того, мы использовали записи о распространении за 2022 и 2023 годы, которые выходили за пределы периода действия биоклиматических переменных, для тестирования производительности модели с теми же метриками (тест модели). TSS использовался для оценки модели, поскольку существует много критики относительно использования площади под ROC-кривой (AUC), предоставляемой результатами модели [45,46,47]. TSS – это значение, полученное путем сложения чувствительности и специфичности и вычитания 1. Оно находится в диапазоне от -1 до 1, и значение TSS, близкое к 1, свидетельствует о более высокой производительности.
2.6. Ансамблевое моделирование и проекция модели
Ансамблевое моделирование, которое объединяет результаты прогнозов двух или более моделей, снижает неопределенность прогнозов отдельных моделей [10,48]. Мы использовали метод взвешенного среднего, основанный на производительности каждой модели, для объединения результатов четырех различных типов алгоритмов моделирования следующим образом:
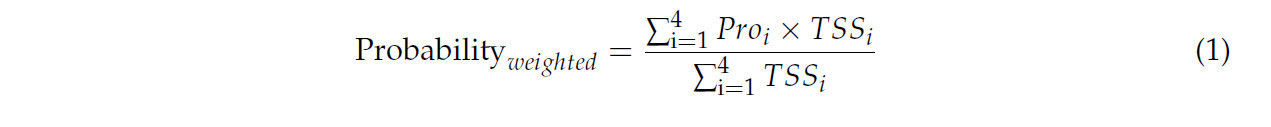
где 1, 2, 3 и 4 относятся к моделям CLIMEX, MaxEnt, RF и MLP соответственно.
Чтобы привести результаты модели к одному диапазону, EI из модели CLIMEX был разделен на 30, а значения выше 30 преобразованы в 1, нормализуя все выходные данные до диапазона от 0 до 1. Затем мы рассчитали взвешенное среднее на основе значения TSS, чтобы создать ансамблевый результат, пропорциональный производительности каждой модели. Среди метрик производительности модели мы выбрали TSS в качестве веса, потому что это практичная и широко используемая метрика [49]. Ансамблевый результат был спроецирован глобально путем извлечения областей консенсуса из каждого алгоритма и наложения их с распространением хозяев с использованием функции Fuzzy Overlay в ArcGIS Pro.
3. Результаты
3.1. Производительность модели
Все модели показали высокий TSS > 0.8, причем самый высокий TSS 0.906 был у модели MLP, в то время как модель RF показала самую высокую производительность (TSS = 0.981) с тестовым набором данных (Таблица 3). Напротив, самый низкий TSS наблюдался у модели MaxEnt как во время валидации, так и тестирования модели из-за ее низкой чувствительности. Несмотря на обычно более низкий TSS из-за низкой специфичности по сравнению с высокой чувствительностью, модель CLIMEX продемонстрировала общую высокую производительность [50]. Напротив, модель MaxEnt, которая обычно показывает высокую производительность в SDM, показала общую более низкую производительность. Универсальные классификаторы на основе машинного обучения, RF и MLP, показали очень высокую производительность, особенно с очень высокой чувствительностью. Хотя производительность модели варьировалась в зависимости от алгоритмов модели, все модели показали приемлемую производительность [51].
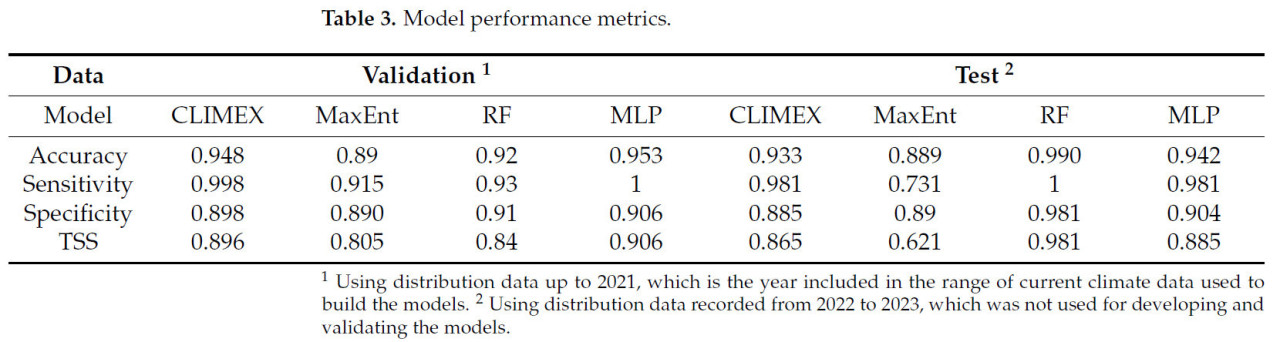
Таблица 3. Метрики производительности модели.
3.2. Глобальное потенциальное распространение S. marginella с использованием отдельных моделей
Все модели предсказали, что S. marginella потенциально будет распространен на юго-востоке США, в центральной Африке, прибрежных районах Австралии и Южной Америке, в дополнение к Азии, где были подтверждены фактические записи о встречаемости (Рисунок 2). За исключением модели MaxEnt, Европа была предсказана как имеющая подходящие климатические условия для возникновения вредителя. Прогнозируемая область потенциального распространения была самой широкой в модели CLIMEX из-за ее алгоритмической природы, которая находит области, соответствующие климатическим условиям для биологии вида. Напротив, самый узкий диапазон был предсказан с использованием модели MaxEnt, которая зависит только от данных о присутствии. Модель CLIMEX предсказала зоны риска в США более широко, чем другие модели, включая районы с растениями-хозяевами. RF и MLP предсказали, что вредитель, вероятно, будет распространен на самой большой территории в Азии, с высокой вероятностью встречаемости, включая Юго-Восточную Азию и Дальний Восток, где растения-хозяева были широко распространены.

Рисунок 2. Результаты прогноза глобального потенциального распространения Salurnis marginella с использованием (a) CLIMEX, (b) MaxEnt, (c) RF и (d) MLP с текущими климатическими данными.
При применении сценариев изменения климата было предсказано, что общее потенциальное распространение немного сместится к северу, но оно существенно не отличалось от модели в текущих климатических условиях (Рисунок 3 и Рисунок 4). Различия между моделями со сценариями SSP245 и SSP585 были незначительными. Потенциальное распространение, предсказанное с использованием модели CLIMEX, показало, что потенциальное распространение S. marginella в Южной Корее, Европе и США расширится к северу, но значительное снижение EI произойдет в Южной Америке и Африке. Ожидалось, что области потенциального распространения в Европе расширятся при использовании модели MaxEnt. Модель RF предсказала, что диапазон потенциального распространения в США, Южной Америке и Африке может уменьшиться к 2050 году, в то время как модель MLP предсказала, что потенциальное распространение в США и Европе сместится к северу, но уменьшится в Восточной Азии.
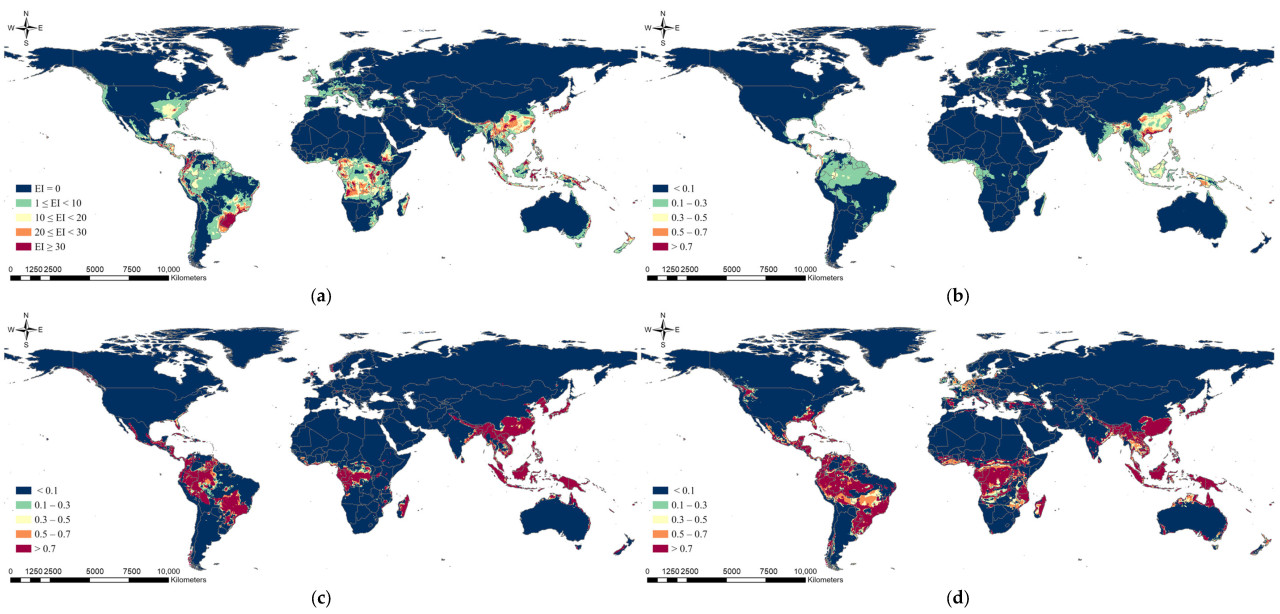
Рисунок 3. Результаты прогноза глобального потенциального распространения Salurnis marginella с использованием (a) CLIMEX, (b) MaxEnt, (c) RF и (d) MLP со сценарием изменения климата на 2050 год (SSP245).
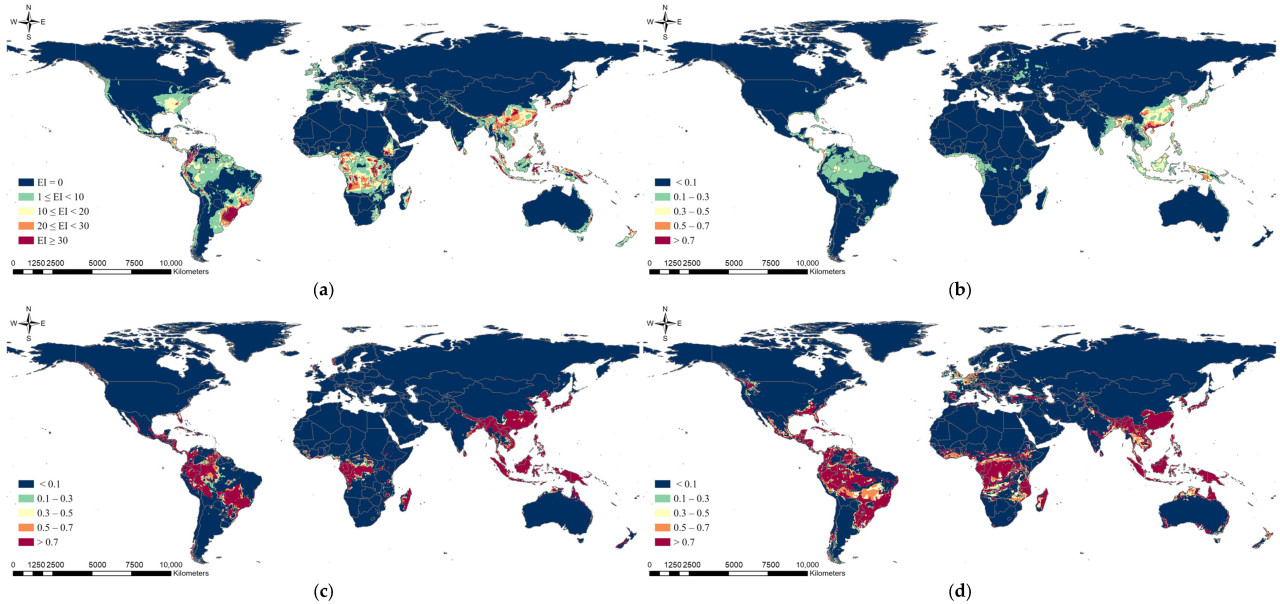
Рисунок 4. Результаты прогноза глобального потенциального распространения Salurnis marginella с использованием (a) CLIMEX, (b) MaxEnt, (c) RF и (d) MLP со сценарием изменения климата на 2050 год (SSP585).
3.3. Ансамблевый прогноз глобального потенциального распространения S. marginella
Ансамблевая модель в текущих климатических условиях показала, что потенциальное распространение S. marginella может охватывать Дальний Восток, Юго-Восточную Азию, Центральную Африку, юго-восток США, а также северо-восточное и западное побережье Южной Америки с высокой вероятностью встречаемости (Рисунок 5). В частности, зоны риска с вероятностью встречаемости > 0.7 были показаны на юге и западе Бразилии в Южной Америке, вокруг Конго в Африке, в южных регионах Дальнего Востока и во всей Юго-Восточной Азии. Потенциальное распространение немного сместится дальше на север в 2050 году при сценариях изменения климата, но общий диапазон распространения уменьшится (Таблица 4). Например, области с вероятностью встречаемости > 0.5 оценивались как 13.97% мира, тогда как в 2050 году они будут охватывать примерно 10%. Ожидалось уменьшение ареалов распространения в Южной Америке и Африке, в то время как зоны высокого риска возникновения вредителя на Дальнем Востоке и в Юго-Восточной Азии, согласно прогнозам, сохранятся. Мы извлекли вероятность области, где вероятность встречаемости по ансамблевой модели была > 0.3 как в текущем, так и в будущем климате, вместе с районами выращивания растений-хозяев, чтобы определить фактическую зону риска распространения. В результате Дальний Восток, включая Корею, Японию и Китай, где распространение уже подтверждено, был идентифицирован как наиболее опасный регион, подверженный ущербу растениям-хозяевам от S. marginella, с большими областями, имеющими вероятность встречаемости > 0.5, независимо от изменения климата (Рисунок 6). Юго-восточный регион США был оценен как область, где вредитель, вероятно, появится, с вероятностью более 0.3 и наличием растений-хозяев. Следовательно, существует потенциал для ущерба от распространения этого вредителя в случае его интродукции в будущем.
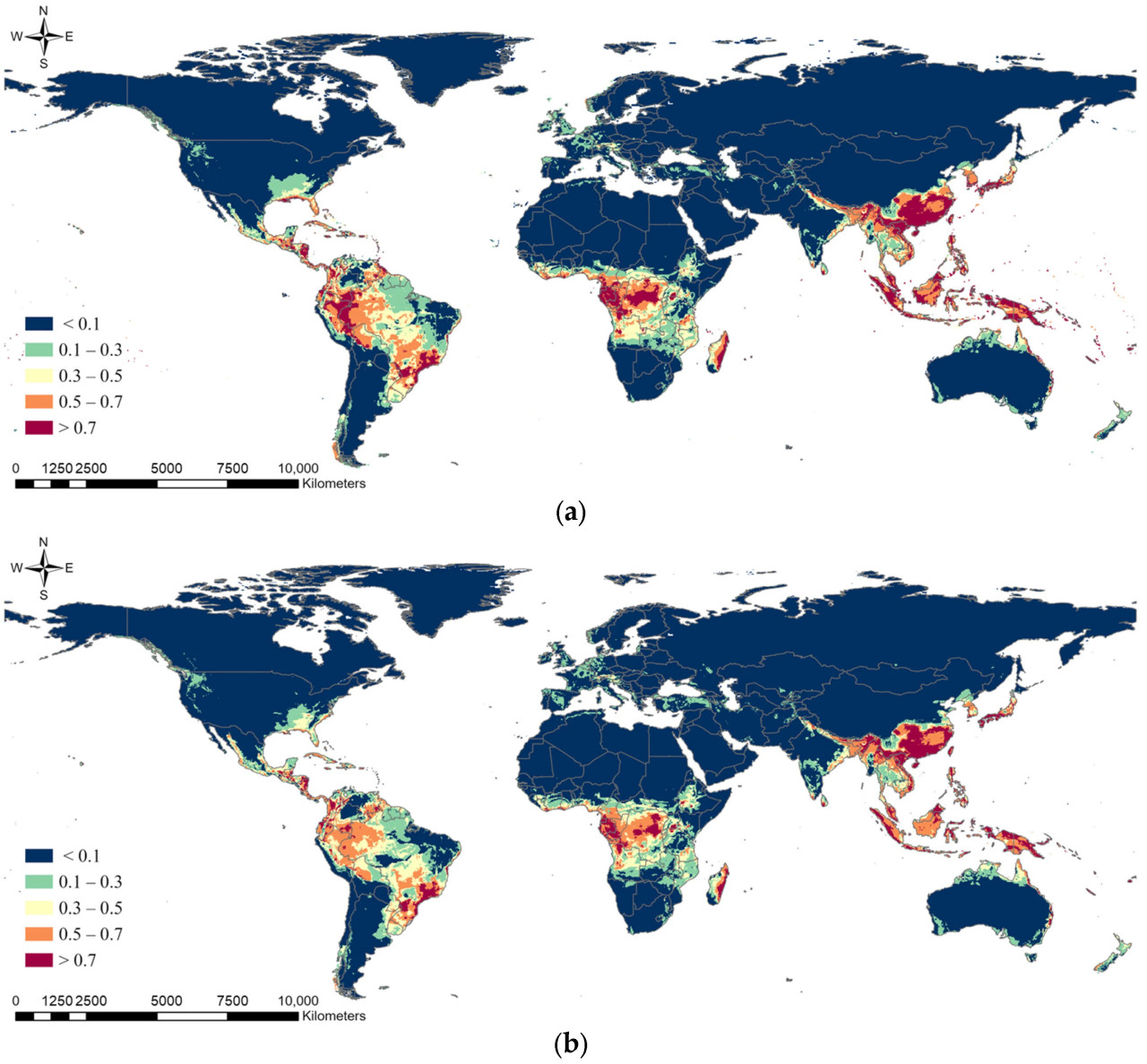
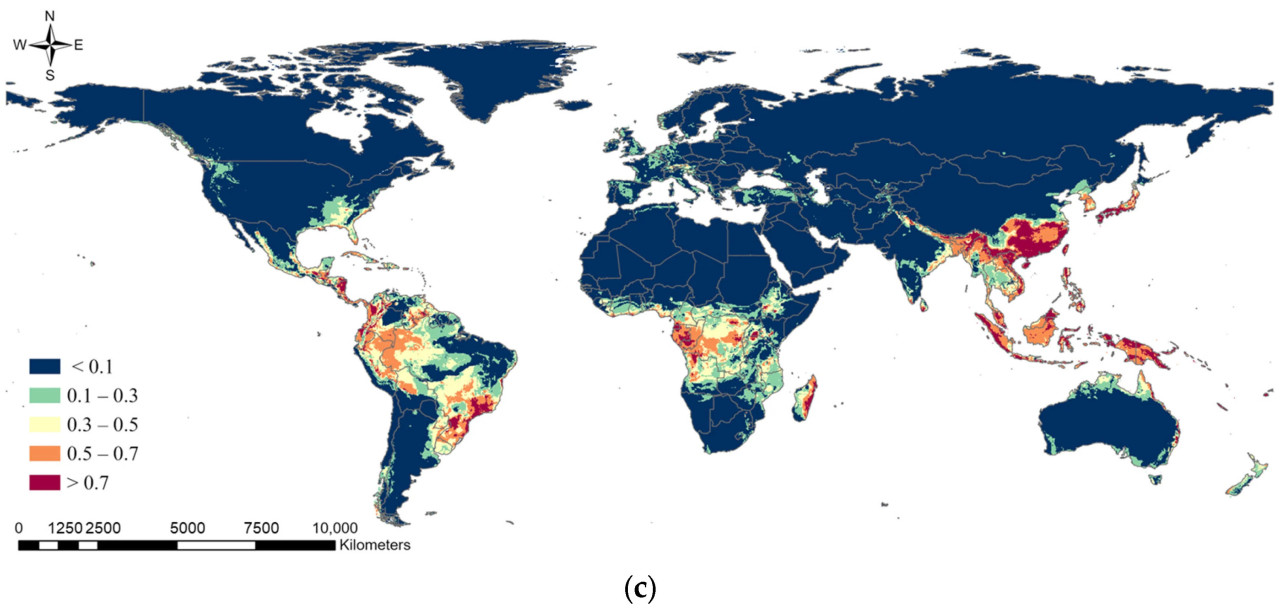
Рисунок 5. Результаты прогноза глобального потенциального распространения Salurnis marginella с использованием ансамблевой модели при (a) текущем климате, (b) будущем климате (SSP245) и (c) будущем климате (SSP585).
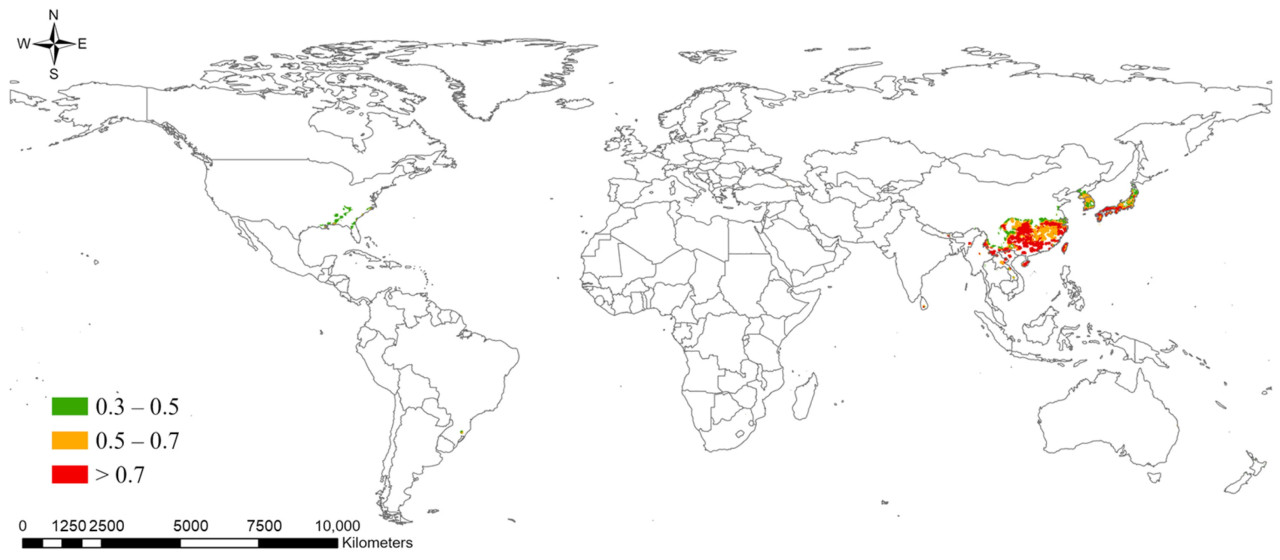
Рисунок 6. Регионы с высокой вероятностью встречаемости Salurnis marginella и присутствия растений-хозяев, где климатическая пригодность и кормовые потребности совпадают, что указывает на зоны постоянного риска при текущих и будущих климатических сценариях.
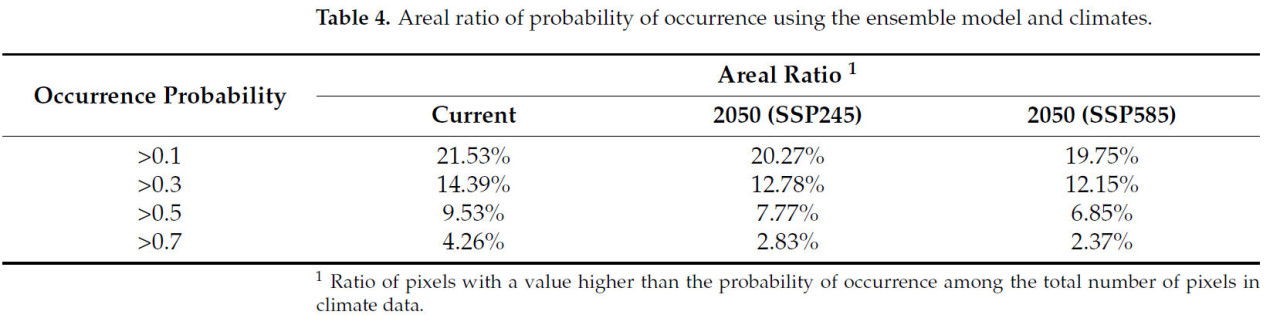
Таблица 4. Доля площади вероятности встречаемости с использованием ансамблевой модели и климатических условий.
4. Обсуждение
В этом исследовании мы спрогнозировали глобальное потенциальное распространение S. marginella в зависимости от изменения климата, используя множественные алгоритмы SDM, и разработали ансамблевую модель, учитывающую распространение хозяев. Из-за недавних открытий, в основном в Азии, данные, доступные для разработки модели, ограничены [2], что обуславливает необходимость использования множества моделей с разными алгоритмами для повышения надежности прогнозов [10,48].
CLIMEX показал самое широкое потенциальное распространение S. marginella, тогда как MaxEnt показал самую маленькую область вокруг фактического распространения. В отличие от моделей машинного обучения, обученных с использованием переменных окружающей среды в координатах встречаемости, модель CLIMEX идентифицирует области, где местный климат подходит для биологии вида, что приводит к относительно большим областям по сравнению с потенциальным распространением, предсказанным с использованием моделей машинного обучения [14,52]. В случае модели MaxEnt при построении модели использовалось 10 000 фоновых координат, в отличие от двух других моделей машинного обучения, которые использовали то же количество фоновых данных, что и данные о распространении [53]. Модели RF и MLP использовали то же количество данных об отсутствии, что и данные о присутствии, что привело к относительно большим прогнозным областям по сравнению с областью, предсказанной с использованием модели MaxEnt, где потенциальная область была более узко ограничена из-за большого количества фоновых точек. Это было продемонстрировано производительностью модели с использованием тестовых наборов данных, не использовавшихся для разработки и валидации модели, что в конечном итоге показало, что все модели продемонстрировали приемлемую предсказательную производительность. Модель MaxEnt показала самую низкую производительность, в отличие от типичных случаев, вероятно, из-за небольшого количества данных о встречаемости и их распределения в определенных регионах. Следовательно, относительно узкая предсказанная область MaxEnt могла привести к низкой производительности на тестовом наборе, что позволяет предположить, что модели MaxEnt требуется достаточное количество данных о встречаемости, чтобы показать надежную производительность [54,55]. Кроме того, модель MaxEnt была разработана с использованием фоновых точек, созданных только в Азии, чтобы избежать переобучения из-за регионально смещенных данных о распространении S. marginella, в отличие от двух других моделей машинного обучения. Напротив, модель RF показала отличную производительность по сравнению с другими моделями, показав примерно 7.6% увеличение на тестовых наборах данных по сравнению с данными валидации. Это означает, что RF была устойчива в прогнозировании распределения видов с недостаточным количеством данных о встречаемости [21]. Тем не менее, модели на основе машинного обучения, включая RF, сильно зависят от размера и структуры данных о встречаемости. В этом исследовании RF и MaxEnt имеют более узкий диапазон прогнозирования, сконцентрированный вокруг области распространения, по сравнению с двумя другими моделями. Это указывает на то, что при малом размере выборки точное прогнозирование областей, где вероятны встречи, может быть затруднено. Это также подчеркивает ценность использования нескольких моделей для смягчения неопределенности, вызванной зависимостью от входных данных, особенно при работе с ограниченным числом данных о встречаемости, путем использования разнообразных алгоритмов. Модель CLIMEX обычно показывала низкую специфичность и высокую чувствительность по сравнению с алгоритмами SDM на основе машинного обучения; однако в этом исследовании она показала относительно более высокую производительность, что может быть связано с количеством данных о встречаемости и структурой их распределения, сконцентрированного в конкретном регионе [50]. Следовательно, хотя S. marginella еще не распространился, вполне вероятно, что регионы с климатом, пригодным для обитания, будут использоваться в качестве данных об отсутствии, что приведет к более узкому потенциальному распространению и в целом более низкой вероятности встречаемости, чем у двух других моделей машинного обучения. Мы признаем, что использование параметров Metcalfa pruinosa для установки начальных значений в модели CLIMEX для S. marginella может внести некоторую погрешность, учитывая ограниченность биологических данных, доступных для S. marginella. Хотя эта проблема была решена путем корректировки параметров на основе климатических данных из фактических регионов распространения S. marginella и калибровки прогнозируемого распространения в соответствии с его наблюдаемым ареалом, дополнительные биологические данные были бы полезны для разработки видоспецифичной модели CLIMEX для S. marginella.
Было обнаружено, что осадки в сезон дождей влияют на потенциальное распространение S. marginella, о чем свидетельствует вклад переменных в модели MaxEnt и среднее уменьшение значения Gini в модели RF [56,57]. Благодаря анализу кривой отклика среди результатов модели MaxEnt, вероятность встречаемости была высокой при приблизительно 300 и 500 мм в самый влажный месяц, тогда как вероятность встречаемости достигала пика выше 600 мм в летний сезон. Это соответствует климатическим условиям Юго-Восточной Азии, где фактически встречается S. marginella, с относительно высоким количеством осадков, предполагая, что районы с аналогичными режимами осадков будут пригодны для распространения S. marginella, например, в северном регионе Бразилии [2]. Это было подтверждено в модели CLIMEX, в которой сухой и влажный стресс, связанные с осадками, оказались значительно выше в Юго-Восточной Азии, чем в других регионах. Более того, было замечено, что, несмотря на подходящие температуры, EI оставался нулевым из-за недостаточного количества осадков в определенных регионах, таких как провинция Шаньдун в Китае и район Караман в Турции. Это свидетельствует о том, что осадки действуют как лимитирующий фактор для развития и выживания вредителя, и этот вывод подтверждается согласованностью разработанных моделей. Среднесуточный диапазон температур был еще одним фактором, ограничивающим потенциальное распространение вредителя, предполагая, что краткосрочные колебания температуры ограничивают места обитания видов. Согласно кривой отклика в модели MaxEnt, вероятность встречаемости увеличивалась, когда суточный диапазон был близок к нулю, указывая на то, что небольшие колебания температуры могут быть благоприятны для этого вредителя. В совокупности районы, предпочитаемые S. marginella, соответствовали климату тропических лесов, тропических муссонов и влажному субтропическому климату согласно классификации климата Кёппена–Гейгера [58]. К сожалению, исследования биологических и экологических характеристик S. marginella ограничены; таким образом, предпочтительные климатические условия и связанные с ними регионы могут быть предложены с помощью вышеуказанного анализа. Например, даже если юго-восток США естественным образом недоступен для S. marginella, существует потенциал для инвазии через антропогенные пути, такие как торговля и туризм, поскольку были выявлены подходящий климат и распространение растений-хозяев.
Изменение климата оказало незначительное влияние на потенциальное распространение S. marginella, показав небольшое уменьшение пригодных районов. Мы изучили причины снижения климатической пригодности из-за изменения климата в экваториальных регионах, таких как Бразилия, Перу и Конго, которые показали наибольшие изменения. Текущий климат и климатический сценарий SSP585 в Конго были извлечены и сравнены. По сравнению с незначительными изменениями климатических условий, такими как уменьшение годового количества осадков на 40 мм и увеличение суточного диапазона температур на 0.2 °C, максимальная годовая температура увеличилась на 4.5 °C до 35.8 °C, температуры, при которой S. marginella не может выжить. Это свидетельствует о том, что по мере того, как региональные климаты становятся более различными из-за изменения климата, каждый регион подвергается воздействию по-разному, и количество климатически пригодных районов может уменьшиться. С другой стороны, этот результат может быть связан с характеристиками алгоритмов SDM. В случае CLIMEX изменения в областях потенциального распространения из-за изменения климата более выражены, чем в моделях машинного обучения [12]. Однако большое количество моделей машинного обучения могло привести к тому, что их результаты доминировали в ансамблевой модели, указывая на то, что общее влияние изменения климата не было значительным. Поэтому, по-видимому, уместно обсуждать распределение видов в регионах с меньшей климатической изменчивостью при изменении климата. Кроме того, различия в потенциальном распространении S. marginella не были значительными в зависимости от типа изменения климата (SSP245 и SSP585). Из-за природы сценариев SSP, где разрыв между сценариями увеличивается со временем, климатические различия между сценариями SSP245 и SSP585 не были значительными в 2050 году [59]. В результате сравнения средних значений климатических переменных между сценариями изменения климата, извлеченных из координат распространения S. marginella, было подтверждено очень незначительное изменение с разницей в среднегодовой температуре 0.35 °C и годовом количестве осадков 6 мм. В этом исследовании мы использовали новейший сценарий изменения климата, SSP. Среди различных сценариев SSP, использованных в исследовании, основанных на мерах по противодействию изменению климата, с учетом будущих социально-экономических изменений, SSP245 – это сценарий, предполагающий средний уровень смягчения последствий изменения климата и социально-экономического развития, а SSP585 – сценарий с высоким уровнем использования ископаемого топлива и бесконтрольного развития. Следовательно, мы стремились повысить надежность прогнозов, используя как средние, так и экстремальные сценарии. Однако будущие прогнозы, основанные на таких сценариях изменения климата, могли бы повысить надежность путем диверсификации результатов за счет использования различных типов сценариев.
5. Выводы
Области потенциального распространения S. marginella были оценены с использованием множественных и ансамблевых алгоритмов SDM, учитывающих изменение климата и распространение хозяев. Эти результаты свидетельствуют о том, что изменения среды обитания из-за изменения климата незначительны. Однако, учитывая растения-хозяева и климатические условия, существует потенциал для распространения вида на соседние территории, где он в настоящее время не встречается, или для инвазии в Северную Америку через искусственные пути. Из-за ограниченных и регионально смещенных данных о распространении S. marginella была использована ансамблевая модель, объединяющая CLIMEX и модели машинного обучения, чтобы компенсировать неопределенность, возникающую в процессе разработки модели. Этот подход может быть использован для моделирования видов с ограниченными биологическими или экологическими исследованиями и полевыми данными. Кроме того, используя области консенсуса, выявленные ансамблевой моделью, вместе с текущим распространением хозяев, мы смогли предложить районы, где вид потенциально мог бы акклиматизироваться, предоставляя ценную информацию для мониторинга и борьбы с вредителями. Хотя это исследование не включало потенциальные изменения в распространении растений-хозяев под влиянием изменения климата, будущие исследования, учитывающие изменения ареалов как вредителя, так и хозяев, могут привести к разработке более надежных моделей.
Ссылка
1. Skendžić, S.; Zovko, M.; Živković, I.P.; Lešić, V.; Lemić, D. The impact of climate change on agricultural insect pests. Insects 2021, 12, 440. [Google Scholar] [CrossRef]
2. Lee, H.; Bae, Y.-S.; Kim, D.E. First Record of the Salurnis marginella (Guérin-Méneville)(Hemiptera: Flatidae) in Korea. Korean J. Appl. Entomol. 2016, 55, 477–482. [Google Scholar] [CrossRef]
3. Choi, H.-S.; Jeong, S.Y.; Lee, K.H.; Jeong, J.S.; Park, J.S.; Jeong, N.R.; Kim, M.J.; Lee, W.; Kim, I. Population genetic analysis of Salurnis marginella (Hemiptera: Flatidae). Int. J. Ind. Entomol. 2021, 43, 67–77. [Google Scholar] [CrossRef]
4. Miller, J. Species distribution modeling. Geogr. Compass 2010, 4, 490–509. [Google Scholar] [CrossRef]
5. Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
6. Dicko, A.H.; Lancelot, R.; Seck, M.T.; Guerrini, L.; Sall, B.; Lo, M.; Vreysen, M.J.; Lefrançois, T.; Fonta, W.M.; Peck, S.L. Using species distribution models to optimize vector control in the framework of the tsetse eradication campaign in Senegal. Proc. Natl. Acad. Sci. USA 2014, 111, 10149–10154. [Google Scholar] [CrossRef] [PubMed]
7. Srivastava, V.; Lafond, V.; Griess, V.C. Species distribution models (SDM): Applications, benefits and challenges in invasive species management. CABI Rev. 2019, 14, 1–13. [Google Scholar] [CrossRef]
8. Strauss, G. Pest risk analysis of Metcalfa pruinosa in Austria. J. Pest Sci. 2010, 83, 381–390. [Google Scholar] [CrossRef]
9. Byeon, D.H.; Jung, J.M.; Jung, S.; Lee, W.H. Prediction of global geographic distribution of Metcalfa pruinosa using CLIMEX. Entomol. Res. 2018, 48, 99–107. [Google Scholar] [CrossRef]
10. Araújo, M.B.; New, M. Ensemble forecasting of species distributions. Trends Ecol. Evol. 2007, 22, 42–47. [Google Scholar] [CrossRef]
11. Shabani, F.; Kumar, L.; Ahmadi, M. A comparison of absolute performance of different correlative and mechanistic species distribution models in an independent area. Ecol. Evol. 2016, 6, 5973–5986. [Google Scholar] [CrossRef] [PubMed]
12. Byeon, D.H.; Kim, S.H.; Jung, J.M.; Jung, S.; Kim, K.H.; Lee, W.H. Climate-based ensemble modelling to evaluate the global distribution of Anoplophora glabripennis (Motschulsky). Agric. For. Entomol. 2021, 23, 569–583. [Google Scholar] [CrossRef]
13. Yoon, S.; Lee, W.-H. Methodological analysis of bioclimatic variable selection in species distribution modeling with application to agricultural pests (Metcalfa pruinosa and Spodoptera litura). Comput. Electron. Agric. 2021, 190, 106430. [Google Scholar] [CrossRef]
14. Kriticos, D.J.; Maywald, G.F.; Yonow, T.; Zurcher, E.J.; Herrmann, N.I.; Sutherst, R. Exploring the effects of climate on plants, animals and diseases. Climex Version 2015, 4, 184. [Google Scholar]
15. Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
16. Cutler, D.R.; Edwards, T.C., Jr.; Beard, K.H.; Cutler, A.; Hess, K.T.; Gibson, J.; Lawler, J.J. Random forests for classification in ecology. Ecology 2007, 88, 2783–2792. [Google Scholar] [CrossRef] [PubMed]
17. Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
18. Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
19. Park, Y.-S.; Lek, S. Artificial neural networks: Multilayer perceptron for ecological modeling. In Developments in Environmental Modelling; Elsevier: Amsterdam, The Netherlands, 2016; Volume 28, pp. 123–140. [Google Scholar]
20. Muñoz-Mas, R.; Martínez-Capel, F.; Alcaraz-Hernández, J.D.; Mouton, A.M. On species distribution modelling, spatial scales and environmental flow assessment with Multi–Layer Perceptron Ensembles: A case study on the redfin barbel (Barbus haasi; Mertens, 1925). Limnologica 2017, 62, 161–172. [Google Scholar] [CrossRef]
21. Lee, W.-H.; Song, J.-W.; Yoon, S.-H.; Jung, J.-M. Spatial evaluation of machine learning-based species distribution models for prediction of invasive ant species distribution. Appl. Sci. 2022, 12, 10260. [Google Scholar] [CrossRef]
22. Zhang, Y.-Z.; Hanula, J.L.; Sun, J.-H. Survey for potential insect biological control agents of Ligustrum sinense (Scrophulariales: Oleaceae) in China. Fla. Entomol. 2008, 91, 372–382. [Google Scholar] [CrossRef]
23. GBIF. GBIF.org. GBIF Occurrence. 2023. Available online: https://doi.org/10.15468/dl.wursy2 (accessed on 20 November 2023).
24. Brown, J.L. SDM toolbox: A python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
25. Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
26. Harris, I.; Osborn, T.J.; Jones, P.; Lister, D. Version 4 of the CRU TS monthly high-resolution gridded multivariate climate dataset. Sci. Data 2020, 7, 109. [Google Scholar] [CrossRef] [PubMed]
27. GBIF. GBIF.org. GBIF Occurrence. 2024. Available online: https://doi.org/10.15468/dl.ee9x3s (accessed on 4 May 2024).
28. GBIF. GBIF.org. GBIF Occurrence. 2024. Available online: https://doi.org/10.15468/dl.bpwa78 (accessed on 7 March 2024).
29. GBIF. GBIF.org. GBIF Occurrence. 2024. Available online: https://doi.org/10.15468/dl.8fzp89 (accessed on 8 March 2024).
30. GBIF. GBIF.org. GBIF Occurrence. 2024. Available online: https://doi.org/10.15468/dl.geb8av (accessed on 8 March 2024).
31. GBIF. GBIF.org. GBIF Occurrence. 2024. Available online: https://doi.org/10.15468/dl.5wnt4s (accessed on 8 March 2024).
32. GBIF. GBIF.org. GBIF Occurrence. 2024. Available online: https://doi.org/10.15468/dl.guywnc (accessed on 8 March 2024).
33. GBIF. GBIF.org. GBIF Occurrence. 2024. Available online: https://doi.org/10.15468/dl.tpp77j (accessed on 8 March 2024).
34. Shabani, F.; Kumar, L.; Taylor, S. Climate change impacts on the future distribution of date palms: A modeling exercise using CLIMEX. PLoS ONE 2012, 7, e48021. [Google Scholar] [CrossRef]
35. Jung, J.M.; Lee, W.H.; Jung, S. Insect distribution in response to climate change based on a model: Review of function and use of CLIMEX. Entomol. Res. 2016, 46, 223–235. [Google Scholar] [CrossRef]
36. Kumar, S.; Yee, W.L.; Neven, L.G. Mapping global potential risk of establishment of Rhagoletis pomonella (Diptera: Tephritidae) using MaxEnt and CLIMEX niche models. J. Econ. Entomol. 2016, 109, 2043–2053. [Google Scholar] [CrossRef]
37. Ramirez-Cabral, N.Y.; Kumar, L.; Shabani, F. Global alterations in areas of suitability for maize production from climate change and using a mechanistic species distribution model (CLIMEX). Sci. Rep. 2017, 7, 5910. [Google Scholar] [CrossRef]
38. Lee, H.J.; Kim, M.J.; Kim, I.R.; Kim, D.E. Salurnis marginella distribution and ecological characteristics, COI geneinformation. In Proceedings of the 2017 Korea Socieity of Applied Entomology Spring Meeting and International Symposium, Gyeongju, Republic of Korea, 26–28 April 2017. [Google Scholar]
39. Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
40. Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENM eval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for Maxent ecological niche models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef]
41. Barbet-Massin, M.; Jiguet, F.; Albert, C.H.; Thuiller, W. Selecting pseudo-absences for species distribution models: How, where and how many? Methods Ecol. Evol. 2012, 3, 327–338. [Google Scholar] [CrossRef]
42. R Core Team. R: A Language andEnvironment for Statistical Computing, R Foundation for Statistical Computing, Vienna, Austria. 2021. Available online: https://www.R-project.org/ (accessed on 8 March 2024).
43. Hinton, G.E. Connectionist learning procedures. In Machine Learning; Elsevier: Amsterdam, The Netherlands, 1990; pp. 555–610. [Google Scholar]
44. Watts, M.J.; Worner, S.P. Using artificial neural networks to determine the relative contribution of abiotic factors influencing the establishment of insect pest species. Ecol. Inform. 2008, 3, 64–74. [Google Scholar] [CrossRef]
45. Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
46. Lobo, J.M.; Jiménez-Valverde, A.; Real, R. AUC: A misleading measure of the performance of predictive distribution models. Glob. Ecol. Biogeogr. 2008, 17, 145–151. [Google Scholar] [CrossRef]
47. Peterson, A.T.; Papeş, M.; Soberón, J. Rethinking receiver operating characteristic analysis applications in ecological niche modeling. Ecol. Model. 2008, 213, 63–72. [Google Scholar] [CrossRef]
48. Grenouillet, G.; Buisson, L.; Casajus, N.; Lek, S. Ensemble modelling of species distribution: The effects of geographical and environmental ranges. Ecography 2011, 34, 9–17. [Google Scholar] [CrossRef]
49. Shabani, F.; Kumar, L.; Ahmadi, M. Assessing accuracy methods of species distribution models: AUC, specificity, sensitivity and the true skill statistic. Glob. J. Hum. -Soc. Sci. B Geogr. Geo-Sci. Environ. Sci. Disaster Manag. 2018, 18, 7–18. [Google Scholar]
50. Yoon, S.; Lee, W.-H. Application of true skill statistics as a practical method for quantitatively assessing CLIMEX performance. Ecol. Indic. 2023, 146, 109830. [Google Scholar] [CrossRef]
51. Tobeña, M.; Prieto, R.; Machete, M.; Silva, M.A. Modeling the potential distribution and richness of cetaceans in the Azores from fisheries observer program data. Front. Mar. Sci. 2016, 3, 202. [Google Scholar] [CrossRef]
52. Song, J.-W.; Jung, J.-M.; Nam, Y.; Jung, J.-K.; Jung, S.; Lee, W.-H. Spatial ensemble modeling for predicting the potential distribution of Lymantria dispar asiatica (Lepidoptera: Erebidae: Lymantriinae) in South Korea. Environ. Monit. Assess. 2022, 194, 889. [Google Scholar] [CrossRef] [PubMed]
53. Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
54. Feeley, K.J.; Silman, M.R. Keep collecting: Accurate species distribution modelling requires more collections than previously thought. Divers. Distrib. 2011, 17, 1132–1140. [Google Scholar] [CrossRef]
55. Wan, J.-Z.; Wang, C.-J.; Yu, F.-H. Effects of occurrence record number, environmental variable number, and spatial scales on MaxEnt distribution modelling for invasive plants. Biologia 2019, 74, 757–766. [Google Scholar] [CrossRef]
56. Calle, M.L.; Urrea, V. Stability of Random Forest importance measures. Brief. Bioinform. 2011, 12, 86–89. [Google Scholar] [CrossRef]
57. Cory, M.; Smith, M.J.; Silander, J.A., Jr. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 10. [Google Scholar] [CrossRef]
58. Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef]
59. Riahi, K.; Van Vuuren, D.P.; Kriegler, E.; Edmonds, J.; O’neill, B.C.; Fujimori, S.; Bauer, N.; Calvin, K.; Dellink, R.; Fricko, O. The Shared Socioeconomic Pathways and their energy, land use, and greenhouse gas emissions implications: An overview. Glob. Environ. Change 2017, 42, 153–168. [Google Scholar] [CrossRef]

Song J-W, Seo J, Lee W-H. Spatial Evaluation of Salurnis marginella Occurrence According to Climate Change Using Multiple Species Distribution Models. Agriculture. 2025; 15(3):297. https://doi.org/10.3390/agriculture15030297
Перевод статьи «Spatial Evaluation ofSalurnis marginellaOccurrence According to Climate Change Using Multiple Species Distribution Models» авторов Song J-W, Seo J, Lee W-H., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык

















































































































Комментарии (0)