Влияние климата на будущее спаржи в Китае
Спаржа (Asparagus officinalis L.) обладает высокой питательной ценностью и пользой для здоровья, однако отсутствие научно обоснованного и рационального планирования выращивания привело к снижению ее качества и урожайности. Для данного исследования были выбраны важные почвенные, климатические, антропогенные и топографические факторы окружающей среды, влияющие на распространение посевов спаржи.
Аннотация
Для оптимизации модели максимальной энтропии (MaxEnt) использовался пакет Kuenm в среде языка R (v4.2.1). С помощью корреляционного анализа Пирсона, оптимизированной модели MaxEnt и технологий пространственного анализа географической информации были определены основные экологические факторы, влияющие на пригодные местообитания для спаржи в Китае. Были смоделированы и спроецированы потенциальные закономерности распределения, миграция и изменения тенденций пригодности территорий Китая для выращивания спаржи в различных исторических и будущих климатических сценариях. Было установлено, что антропогенные факторы и климатические условия являются основными факторами окружающей среды, влияющими на распределение пригодных для выращивания спаржи зон в Китае, за которыми следуют почвенные и топографические факторы. Исторически сложившиеся пригодные местообитания занимали площадь 345,6 × 10⁵ км², что составляет 36% территории Китая. Прогнозируется, что в будущих климатических условиях эти ареалы значительно расширятся. Данное исследование создает основу для рационального планирования и устойчивого развития культуры спаржи.
1. Введение
Спаржа (Asparagus officinalis L.) является высокорентабельной и питательной овощной культурой, которая содержит больше белка и витаминов, чем обычные овощи. Она обладает противораковыми и иммуностимулирующими оздоровительными функциями и пользуется репутацией «Короля овощей» [1,2,3,4]. Китай является крупным производителем спаржи, занимая первое место в мире по площади посевов и объему производства, и играет ключевую роль в мировой торговле спаржей [5]. После почти 30 лет развития спаржевая индустрия Китая превратилась в новую отрасль, обладающую конкурентными преимуществами. Однако планирование посадок спаржи и интродукционное выращивание осуществляются вслепую и бессистемно из-за отсутствия руководства со стороны научной теории, что приводит к снижению урожайности и качества спаржи [6].
Географическое распространение видов находится под влиянием сочетания топографических, климатических, почвенных факторов и деятельности человека [7]. Глобальный климат потеплел с 20-го века. Согласно Шестому оценочному отчету Межправительственной группы экспертов по изменению климата (МГЭИК), глобальная приземная температура в период с 2001 по 2020 год повысилась на 0,99 °C по сравнению с 1850–1900 годами. Ожидается, что в 2021–2024 годах повышение температуры приблизится к 1,5 °C или превысит этот показатель [8]. Сельское хозяйство является одной из сфер, наиболее подверженных влиянию изменений климата, во многом из-за того, что тепло является источником энергии для сельскохозяйственных культур, а вода — их основной составляющей, и оба эти фактора влияют на их физиологические процессы [9]. Более высокие температуры приводят к увеличению содержания водяного пара в атмосфере, изменению характера осадков, увеличению частоты экстремальных погодных явлений и изменению структуры посевов сельскохозяйственных культур [10].
Изменение климата существенно изменяет географическое распространение видов: многие виды смещают свои ареалы на север или мигрируют на большие высоты из-за более выраженной тенденции повышения температуры на больших высотах по сравнению с более низкими [11,12]. Сравнение прошлого и современного распространения растительности в горах Монтсени в Испании показало, что холодные умеренные экосистемы постепенно вытеснялись средиземноморскими экосистемами из-за потепления климата, а буковые леса мигрировали на более высокие высоты, увеличив их примерно на 70 м [13]. Изменение климата вызвало значительный сдвиг к северу границ посевов перспективного однократного и двукратного посева риса в Китае [14]. Текстура почвы, химические и физические характеристики, питательные вещества и влажность влияют на рост корней сельскохозяйственных культур, а также на накопление, трансформацию и поглощение питательных веществ, таким образом воздействуя на процесс роста, развития, урожайности и качества культур и ограничивая ареал их возделывания и распространения [15]. Например, глинистая почва обладает высокой водоудерживающей способностью и плохой воздухопроницаемостью, что приводит к длительному задерживанию воды в почве, что может обеспечить хорошее водоснабжение растений, поэтому она подходит для влаголюбивых растений, в то время как песчаные почвы имеют хорошую водопроницаемость и подходят для выращивания засухолюбивых растений. Топография влияет на распределение гидротермических ресурсов, и основные экологические факторы, такие как водяной пар, тепло, свет и почва, закономерно изменяются с изменением высоты, уклона и экспозиции склона, оказывая значительное влияние на структуру распространения сельскохозяйственных культур [16]. Деятельность человека, включая выбор засухоустойчивых культур в зависимости от удаленности от источников воды и трансформацию земель в земли под застройку, также существенно влияет на структуру посевов [17].
Модели распространения видов (SDMs) количественно определяют связь между параметрами окружающей среды и распространением видов, отражая ограничивающие факторы расселения видов и адаптации к местообитаниям [18]. Они широко используются для моделирования и прогнозирования потенциально пригодных для сельскохозяйственных культур районов, структуры распространения и реакций на изменение климата [19]. В настоящее время обычно используемые SDMs включают генетический алгоритм для прогнозирования набора правил (GARP), модель биоклиматической оболочки (BIOCLIM), факторный анализ экологической ниши (ENFA) и модель максимальной энтропии (MaxEnt) [20]. Среди них модель MaxEnt является наиболее популярной и известной моделью распространения видов и обладает высокой точностью благодаря низким требованиям к размеру выборки, короткому времени выполнения, высокой точности моделирования и выдающейся предсказательной способности. Модель MaxEnt функционирует на основе принципа максимальной энтропии, который использует экологические факторы в качестве ограничений для определения вероятности максимальной энтропии и функциональной связи, установленной фактическими точками распространения видов, для прогнозирования вероятного распространения вида. Модели экологических ниш Domain, BIOCLIM, MaxEnt и GARP были использованы для моделирования распространения местообитаний различных видов в различных масштабах. Полученные результаты свидетельствуют о том, что модель MaxEnt обладала наилучшей прогностической способностью для видов с ограниченными данными о распространении и короткими географическими ареалами [21]. Elith и др. [22] установили, что модель MaxEnt показала наилучшие результаты при моделировании географического распространения 226 видов в шести регионах мира с использованием 16 SDMs.
MaxEnt была расширена от моделирования потенциальных местообитаний для исчезающих или экономически ценных видов и инвазивных видов до исследования сельскохозяйственных культур, например, изучения зон выращивания и оценки реакции на доминирующие экологические факторы риса, кукурузы, пшеницы и картофеля [23,24,25], что обеспечивает важную научную основу для сельскохозяйственного возделывания, сохранения и устойчивого развития. Модель MaxEnt была использована для моделирования и изучения влияния будущего изменения климата на зоны пригодности кукурузы в Кении. Было обнаружено, что будущее изменение климата приведет к увеличению непригодных для кукурузы районов на 1,9–3,9% и сокращению умеренно пригодных районов на 14,6–17,5%. Основными экологическими факторами, влияющими на распространение посевов кукурузы, были средняя температура самого влажного сезона, среднегодовая температура и годовая сумма осадков [26]. Прогнозный анализ потенциальных районов возделывания риса-ратуса и двуурожайного риса в Китае с помощью модели MaxEnt показал, что площадь, пригодная для возделывания двуурожайного риса, составляла 130 500 га, в то время как площадь, пригодная для выращивания риса-ратуса, составляла 56 400 га. Основными экологическими факторами, влияющими на пригодную площадь для риса-ратуса, являются средняя температура в самый теплый сезон, сумма температур ≥10 °C, годовая сумма осадков, высота над уровнем моря и годовая продолжительность солнечного сияния [27]. Модель MaxEnt использовалась в исследованиях прогнозирования распространения видов, но неоптимизированные результаты прогнозирования модели могут привести к серьезным ошибкам подгонки. Сложность модели, различные переменные окружающей среды и параметры влияют на производительность модели в разной степени. Исследования показывают, что оптимизация модели путем настройки параметров может значительно повысить точность моделирования модели MaxEnt и получить более точное распространение видов [13,28].
В настоящее время исследования пригодных для сельскохозяйственных культур районов на основе модели MaxEnt в основном сосредоточены на зерновых культурах, таких как рис, кукуруза и пшеница. Факторы влияния также в основном сосредоточены на воздействии климатических факторов, таких как свет, температура, вода и тепло. Нет исследований, которые бы в полной мере учитывали влияние природных экосистем и деятельности человека на распространение культуры спаржи [29,30]. Поэтому в данном исследовании в качестве объекта исследования была взята спаржа и учтено совокупное влияние топографии, почв, климата и деятельности человека; основные экологические факторы, влияющие на пригодное распределение спаржи, были определены с помощью модели MaxEnt. Дополнительно были спроецированы и проанализированы особенности пригодного распределения спаржи в Китае в текущих и различных будущих климатических сценариях. Исследование было проведено (1) для определения ключевых экологических факторов, влияющих на распределение пригодных местообитаний для спаржи в Китае; (2) для проверки и сравнения эффективности оптимизированной модели MaxEnt в прогнозировании пригодных местообитаний вида; (3) для прогнозирования и анализа лежащей в основе пространственной изменчивости и географического распределения спаржи в различных исторических и будущих климатических сценариях; и (4) для изучения траекторий изменения и миграции центров пригодного распределения спаржи в Китае в различные периоды времени.
2. Материалы и методы
2.1. Источники данных
Данные о распространении спаржи были собраны из Глобальной информационной системы по биоразнообразию (https://www.gbif.org (дата обращения: 9 мая 2024 г.)), Китайского виртуального гербария (https://www.cvh.ac.cn (дата обращения: 9 мая 2024 г.)) и Национальной платформы образцов (http://www.nsii.org.cn (дата обращения: 9 мая 2024 г.)). Когда в записях образцов отсутствовали подробные географические координаты, они определялись с помощью Google Карт, что в итоге дало 252 точки распространения спаржи [31]. Данные по точкам распространения были отфильтрованы с помощью пакета ENMTools в языке R (v4.2.1), чтобы гарантировать сохранение только одного пункта данных о распространении в пределах растра с точностью 2,5′ (~25 км²), избегая переобучения при работе модели и влияния на результаты прогнозирования [32]. После удаления ошибок и повторяющихся точек было окончательно определено 192 точки распространения спаржи (Рисунок 1).
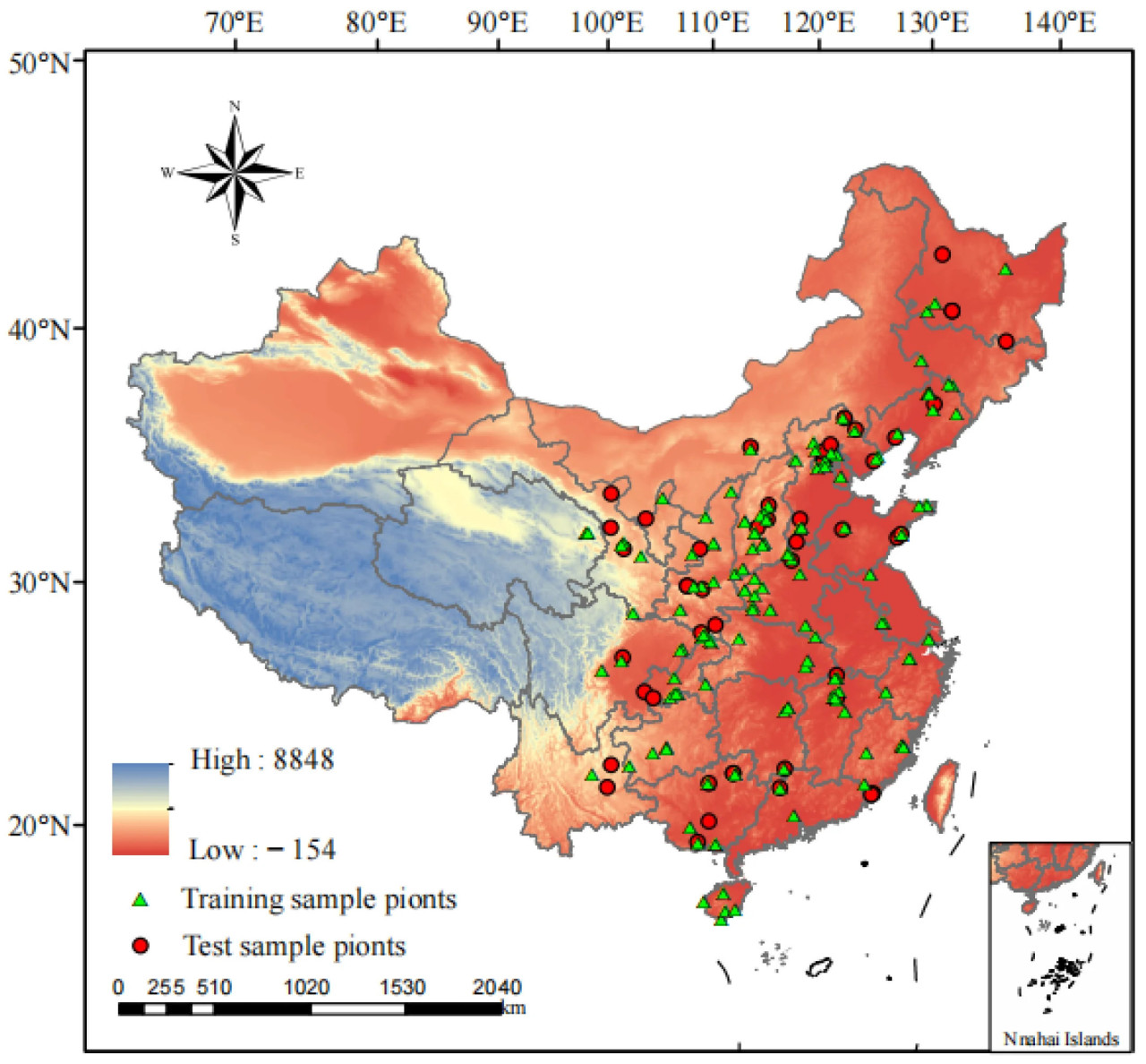
Рисунок 1. Точки распространения спаржи в Китае.
Путем обзора соответствующей литературы были предварительно отобраны и идентифицированы 25 экологических факторов, влияющих на распределение посевов спаржи (Таблица 1) [33]. Территориальные факторы (высота над уровнем моря и уклон) и факторы окружающей среды и климата были предоставлены Всемирной климатической базой данных (WorldClim, http://www.worldclim.org/ (дата обращения: 24 мая 2024 г.)). Данные о химических и физических характеристиках почвы были получены из Мировой почвенной базы данных (http://www.fao.org/ (дата обращения: 25 мая 2024 г.)). Сценарии будущего изменения климата включают четыре общих социально-экономических пути (SSPs), а именно SSP585, SSP370, SSP245 и SSP126, в которых используется климатическая модель BCC-CSM2-MR. Данные о физико-химических свойствах почвы включают pH почвы, содержание органического вещества и карбоната кальция. Антропогенным фактором является валовой внутренний продукт (ВВП) по данным Центра данных по ресурсам и окружающей среде Китайской академии наук (https://www.resdc.cn/ (дата обращения: 25 мая 2024 г.)). Из-за ограничений доступных данных влияние таких факторов, как ВВП, почва и рельеф, было всесторонне учтено только в исторические периоды. Кроме того, данное исследование было в основном направлено на влияние будущего изменения климата на пригодность выращивания спаржи. Поэтому было сделано предположение, что такие факторы, как ВВП, почва и топография, не изменятся в имитационном исследовании будущих периодов. ArcGIS 10.4 был применен для импорта всех переменных окружающей среды для кадрирования и повторной выборки с единым разрешением 2,5′ и выводом данных в формате ASCII.

Таблица 1. Климатические, почвенные, топографические факторы и факторы деятельности человека, влияющие на распределение посадок спаржи (предварительный отбор).
2.2. Тестирование корреляции и отбор переменных окружающей среды
Поскольку между переменными окружающей среды, влияющими на результаты работы модели, может существовать коллинеарность, чтобы предотвратить переобучение модели MaxEnt, выбранные переменные окружающей среды сначала необходимо было отсеять [22]. В модель MaxEnt были загружены выбранные 25 переменных окружающей среды для начальных вычислений, чтобы получить показатель вклада каждой переменной, а тесты корреляции Пирсона были проведены с использованием программного обеспечения ArcGIS (v10.4.1) и SPSS (v27.0.1) [34]. Результат моделирования MaxEnt сохраняет переменные окружающей среды с показателем вклада более 0,1%. Мультиколлинеарность присутствует, когда коэффициент корреляции Пирсона |r| между обеими переменными окружающей среды превышает 0,8. Переменная окружающей среды, которая вносит больший вклад, сохраняется, в то время как та, которая вносит меньший вклад, исключается. Если обе переменные окружающей среды имеют одинаковый показатель вклада, выбирается переменная, демонстрирующая меньшую мультиколлинеарность с другими переменными окружающей среды.
2.3. Модель MaxEnt
2.3.1. Оптимизация модели
Модель MaxEnt подвержена переобучению и систематической ошибке выборки. Способность модели к переносу эффективна только в условиях низких пороговых значений, а параметры по умолчанию могут влиять на точность прогнозирования модели. Сложность модели MaxEnt сильно коррелирует с ее классовыми признаками (FC) и параметрами регуляризационного множителя (RM) [35]. В этом исследовании для оптимизации параметров модели MaxEnt использовался R-пакет Kuenm [36]. В процессе оптимизации 0,1 был установлен в качестве начального значения для RM, и каждый раз оно увеличивалось на 0,1 вплоть до 4,0, что в общей сложности составило 40 контрольных удвоений частоты [37,38]. Оптимизация проводилась с помощью комбинаций пяти типов признаков: H — шарнирный; Q — квадратичный; L — линейный; T — пороговый; и P — произведенный [39]. Показатели пропусков (omission rates) наряду с информационным критерием Акаике (AICc) были использованы для оценки сложности и соответствия различных комбинаций параметров модели MaxEnt [40]. Оптимальным значением параметра была комбинация FC и RM, когда показатели пропусков были менее 5%, натуральный логарифм AICc был наименьшим, а значения дельты AICc равнялись 0 [41].
Поскольку площадь под кривой (AUC) не зависит от порога для оценки производительности модели, способность различных комбинаций параметров различать фоновые и тестовые точки была проверена с использованием AUC под рабочей характеристической кривой приемника (ROC) [42].
2.3.2. Моделирование и проверка точности модели
Данные о распространении спаржи и выбранные факторы окружающей среды (ВВП, Bio3, Bio5, Bio8, Bio11, Bio13, Bio14, Bio15, Bio19, CaCO3, Eleve, pH, OC, slope) были введены в программное обеспечение MaxEnt (v3.4.3) для построения модели. Модель была настроена на выполнение 10 итераций, при этом 75% данных были случайным образом выбраны в качестве обучающей выборки, а оставшиеся 25% — в качестве проверочной. AUC использовалась для определения точности прогнозирования модели MaxEnt, которая повышается по мере приближения значения AUC к 1. AUC ≥ 0,9, 0,8 ≤ AUC < 0,9, 0,7 ≤ AUC < 0,8, 0,6 ≤ AUC < 0,7 и 0,5 ≤ AUC < 0,6 указывают на отличный, улучшенный, средний, худший и неудовлетворительный прогноз модели соответственно [43].
2.3.3. Делимитация и анализ районов, пригодных для выращивания спаржи
Инструмент «Реклассификация» в ArcGIS был использован для классификации прогнозов модели MaxEnt на классы пригодности для посадки. Результаты прогнозирования модели MaxEnt отображались как непрерывная вероятность существования (p), которая варьируется от 0 до 1 в прогнозируемой области. Чем ближе значение p к 1, тем более вероятно, что присутствие вида указывает на более высокую степень пригодности для обитания. Принимая во внимание критерии классификации для оценки «вероятности» пригодности выживания видов, разработанные МГЭИК, в сочетании с климатическими условиями и условиями посадки в исследуемом районе, пригодность выращивания спаржи была разделена в соответствии со следующими критериями: p < 0,08, 0,08 ≤ p < 0,27, 0,27 ≤ p < 0,86 и p ≥ 0,86 для непригодных, пригодных, малопригодных, среднепригодных и высокопригодных районов выращивания соответственно. Инструмент SDMTools в программном обеспечении ArcGIS (v10.4.1) был использован для расчета тенденции изменения оптимальных зон спаржи под влиянием исторических и будущих изменений климата, а также для анализа изменения координат центроида и расстояния миграции наиболее пригодной зоны [44]. Блок-схема этого исследования представлена на Рисунке 2.
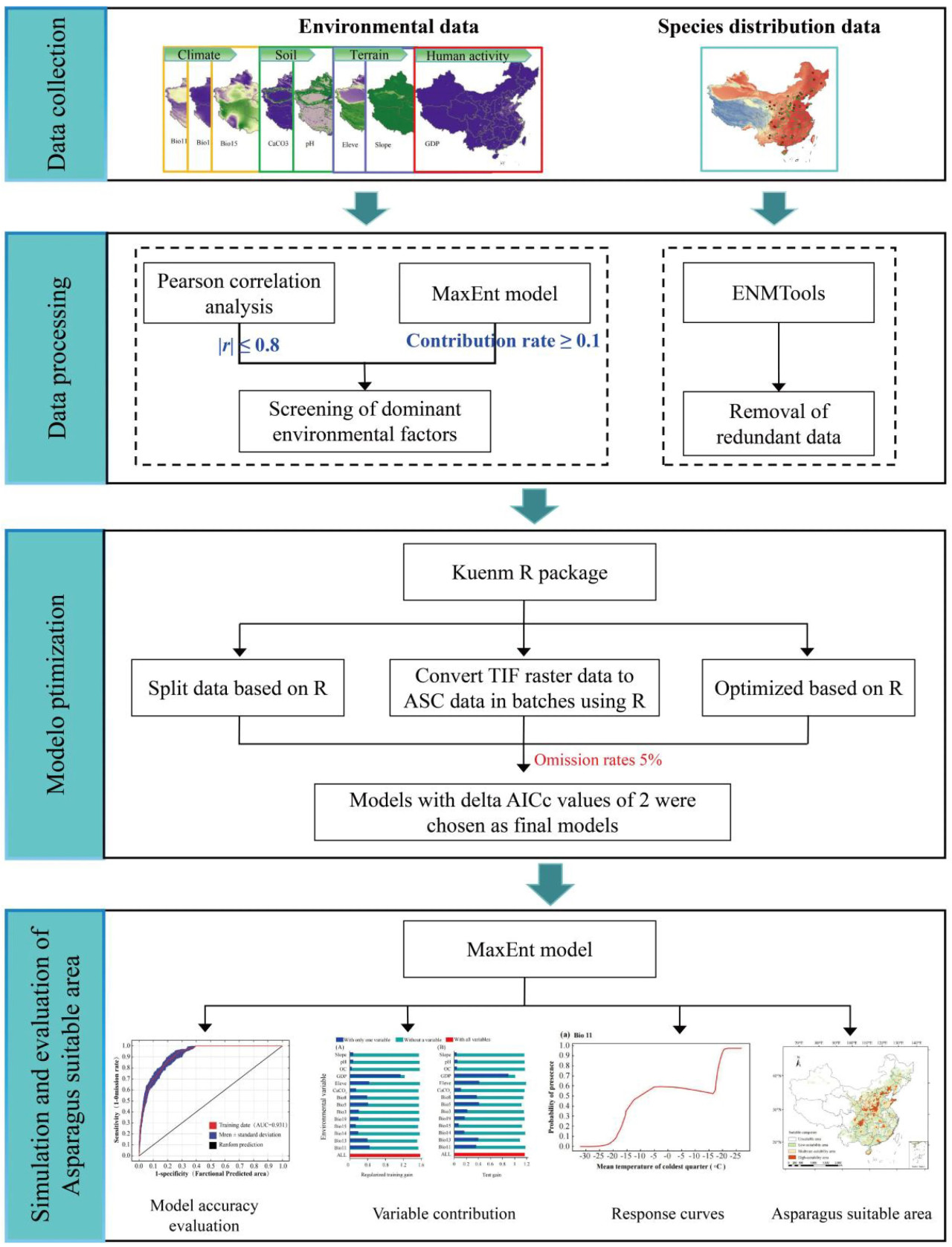
Рисунок 2. Методологическая структура данной работы.
3. Результаты
3.1. Оптимизация и оценка точности прогнозирования моделей MaxEnt
В этом исследовании с помощью пакета Kuenm было создано в общей сложности 1160 моделей распределения, из которых 8 соответствовали стандарту 5%-ного показателя пропусков (omission rate) (Рисунок 3). Для оптимальной модели ROC = 0, показатели пропусков = 0,04; AICc = 4968,31; и оптимизированные параметры модели RM = 0,4; и FC = FQH (Рисунок 3). Данные об окружающей среде вместе с отобранными данными о распространении спаржи были импортированы в модель MaxEnt, и оптимизированные параметры были повторены для 10 прогонов для перекрестной проверки. Согласно полученным результатам, среднее значение AUC модели MaxEnt составило 0,931, а его стандартное отклонение — 0,013, что указывает на отличную точность моделирования (Рисунок 4).
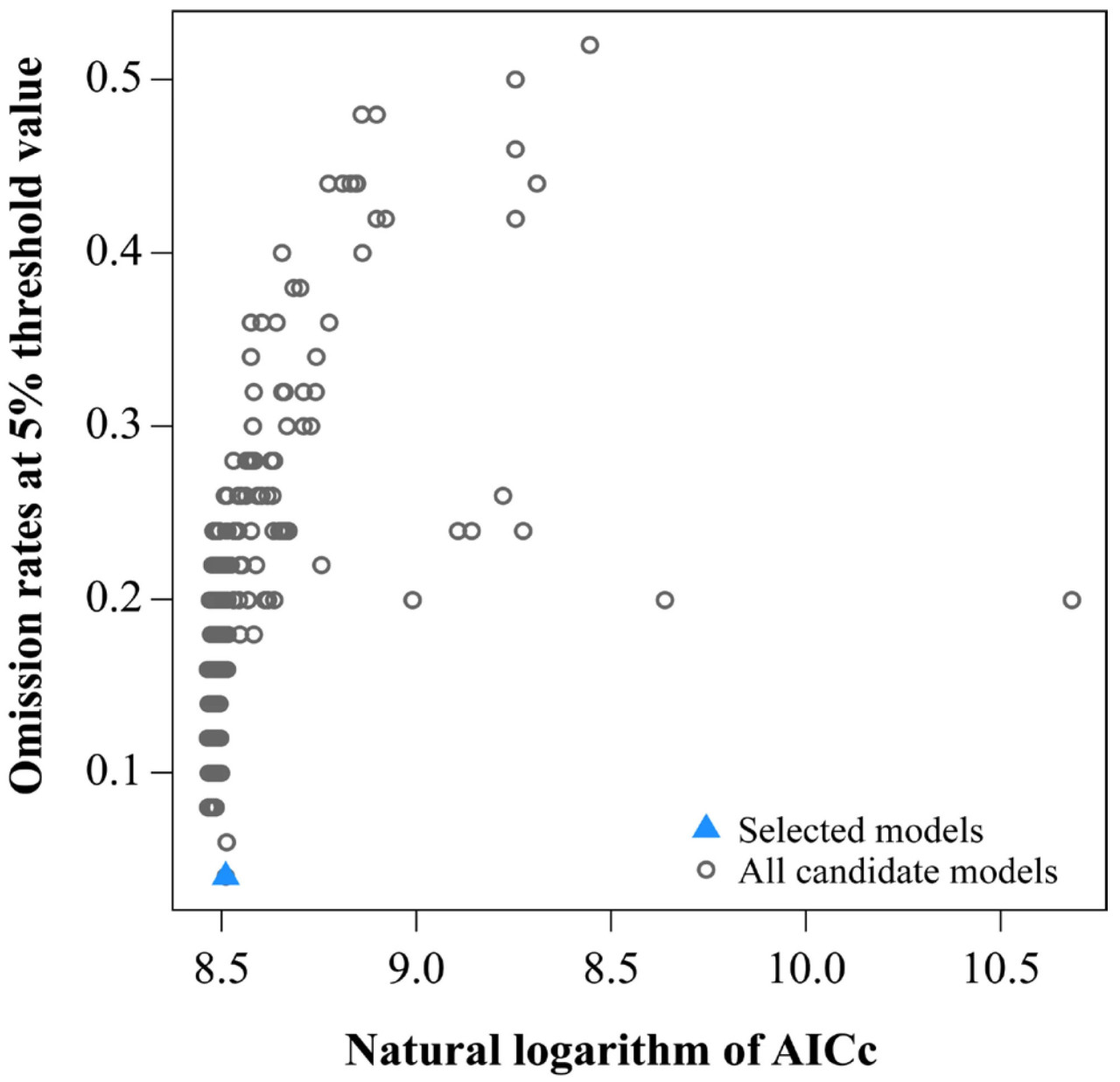
Рисунок 3. Значения AICc и показатели пропусков для всех моделей-кандидатов, незначимых моделей-кандидатов и выбранной «наилучшей» модели-кандидата для спаржи.
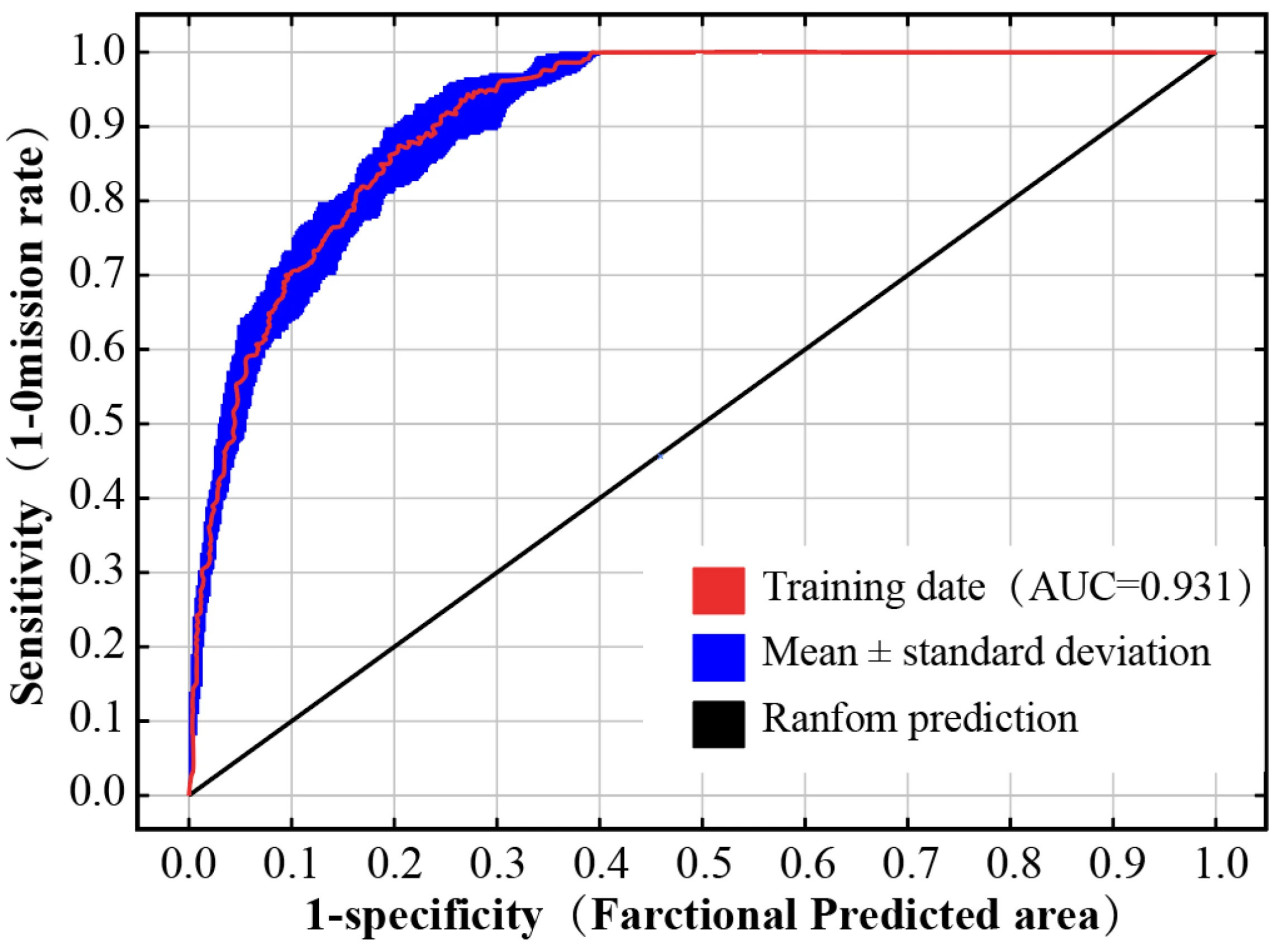
Рисунок 4. ROC-кривая для оценки распределения спаржи в условиях оптимальных параметров согласно модели MaxEnt.
3.2. Отбор основных переменных окружающей среды
Для определения показателя вклада каждого фактора в распределение культуры спаржи в Китае были выбраны первые 25 экологических факторов и импортированы в оптимизированную модель MaxEnt, после чего был проведен корреляционный анализ Пирсона. Были отобраны переменные окружающей среды с показателем вклада > 0,1% и коэффициентом корреляции < ±0,8. Всего было выбрано 14 ключевых переменных окружающей среды, влияющих на распределение культуры спаржи: средняя температура самой холодной четверти (Bio11), самой влажной четверти (Bio8), осадки самого влажного месяца (Bio13), самого сухого месяца (Bio14), соотношение суточных и годовых колебаний температуры (Bio3), среднее количество осадков самой холодной четверти (Bio19), максимальная температура самого жаркого месяца (Bio5), высота над уровнем моря (Eleve), содержание карбоната кальция в почве (CaCO3), pH почвы (pH) и содержание органического вещества в почве (OC) (Рисунок 5). По сравнению с другими переменными окружающей среды, ВВП (с показателем вклада 76,1%), Bio15 (4,5%), уклон (4,3%), Bio11 (4,0%), Bio13 (2,7%) и Bio14 (2,6%) внесли наибольший вклад в распределение культуры спаржи, с совокупным показателем вклада 94,2% (Таблица 2).
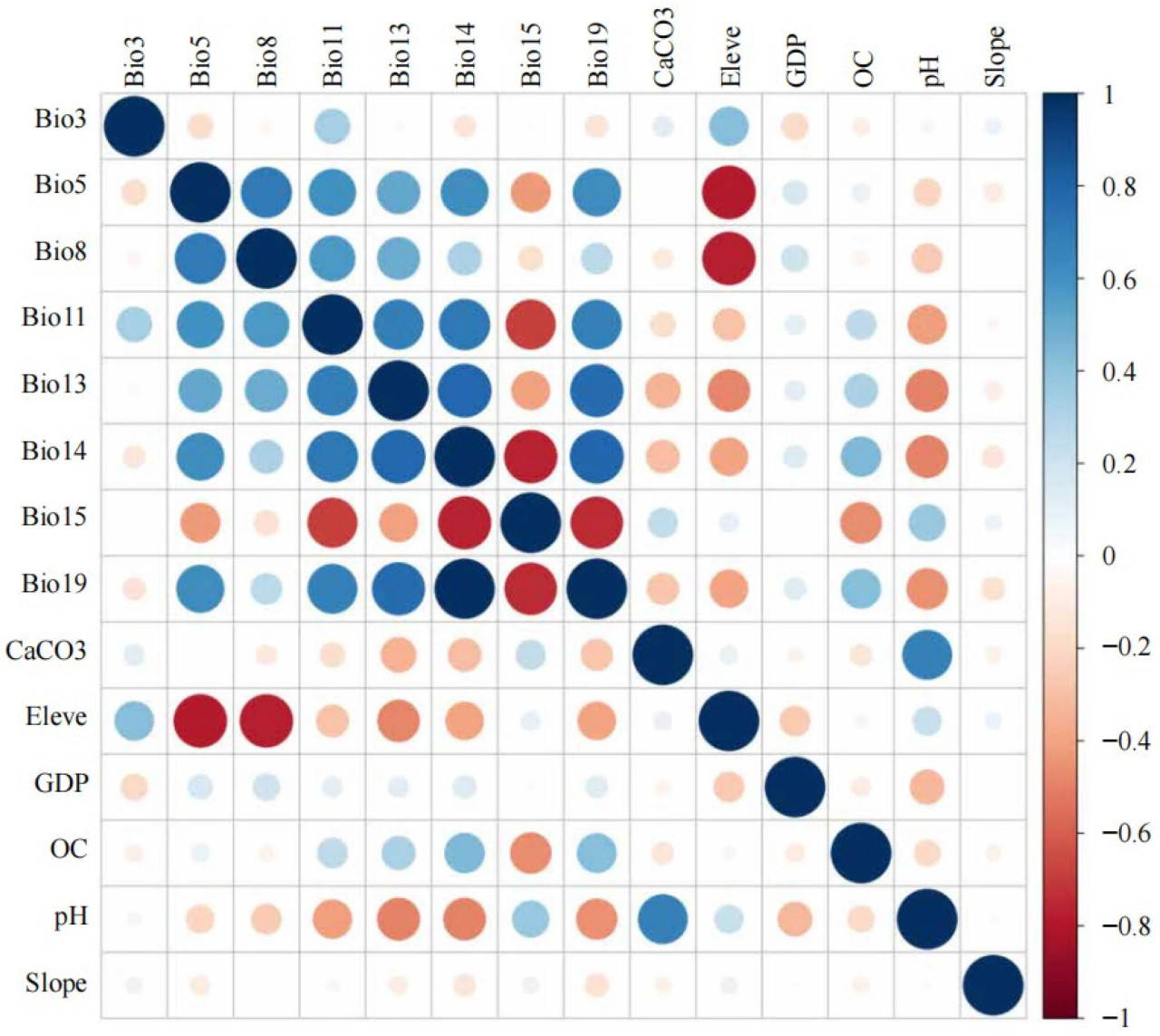
Рисунок 5. Корреляция первоначально отобранных переменных окружающей среды (названия факторов окружающей среды см. в Таблице 1).
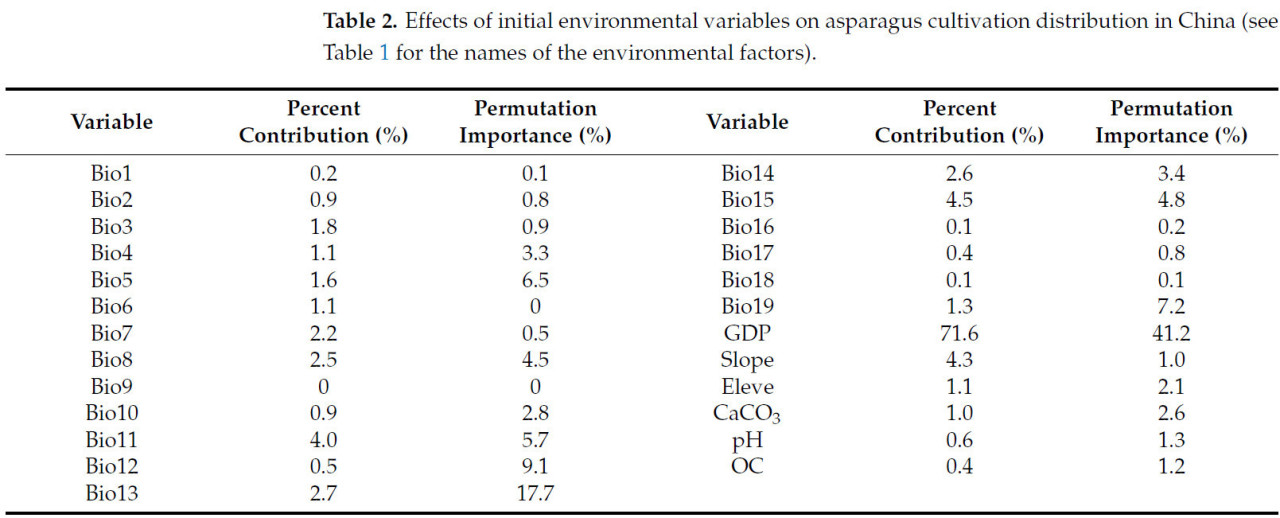
Таблица 2. Влияние исходных переменных окружающей среды на распределение культуры спаржи в Китае (названия факторов окружающей среды см. в Таблице 1).
3.3. Влияние ключевых факторов окружающей среды на распределение культуры спаржи
Согласно тесту значимости методом Jackknife, значимость переменных окружающей среды при наличии только одной переменной окружающей среды следующая: ВВП > Bio11 > Высота над уровнем моря > Bio13 > Bio5 > Bio8 > Bio14 > Bio19 > Bio3 > Bio15 > CaCO3 > уклон > pH > OC (Рисунок 6). Полученные результаты свидетельствуют о том, что Bio11 и ВВП были основными факторами окружающей среды, влияющими на географическое распределение спаржи.
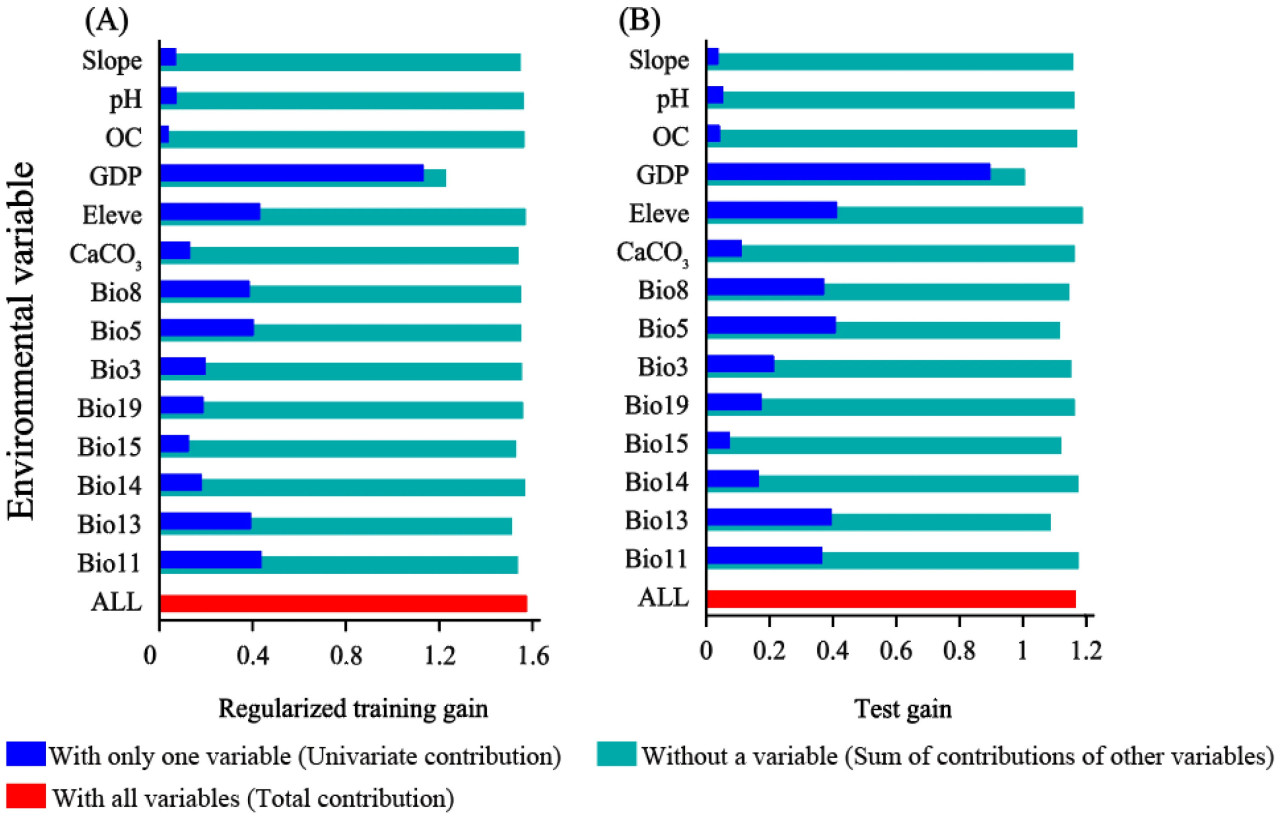
Рисунок 6. Тест Jackknife, использованный для установления влияния ключевых переменных на распределение спаржи: (A) регуляризованный обучающий выигрыш (regularized training gain) для спаржи; (B) тестовый выигрыш (test gain) для спаржи. Влияние переменных в наборе образцов было использовано для валидации модели.
На Рисунке 7 представлены кривые отклика шести ключевых факторов окружающей среды (ВВП, уклон, Bio11, Bio13, Bio14 и Bio15), которые значительно влияют на пригодность выращивания спаржи в Китае. По мере увеличения значений Bio13 и Bio14 вероятность пригодности для спаржи резко возрастала, а затем постепенно снижалась. В частности, вероятность пригодности для Bio13 достигла пика примерно 0,65 при количестве осадков около 200 мм, в то время как наибольшая пригодность для Bio14 наблюдалась при количестве осадков около 20 мм. Это указывает на то, что рост спаржи был наиболее благоприятным в определенном диапазоне осадков. Как чрезмерное, так и недостаточное количество осадков могло снизить ее потенциал роста. Прогнозируемая нелинейная зависимость подчеркивала потребность спаржи в воде и выделяла сложное влияние осадков на ее распределение. Вероятность пригодности спаржи сначала увеличивалась, а затем уменьшалась с ростом ВВП, но скорость увеличения и уменьшения была относительно низкой. Наибольшая вероятность пригодности спаржи составила 0,85 при ВВП 16700,9 млн юаней на км². Чем больше был уклон, тем ниже была вероятность пригодности спаржи, которая была ниже 0,5, когда уклон превышал 40°.
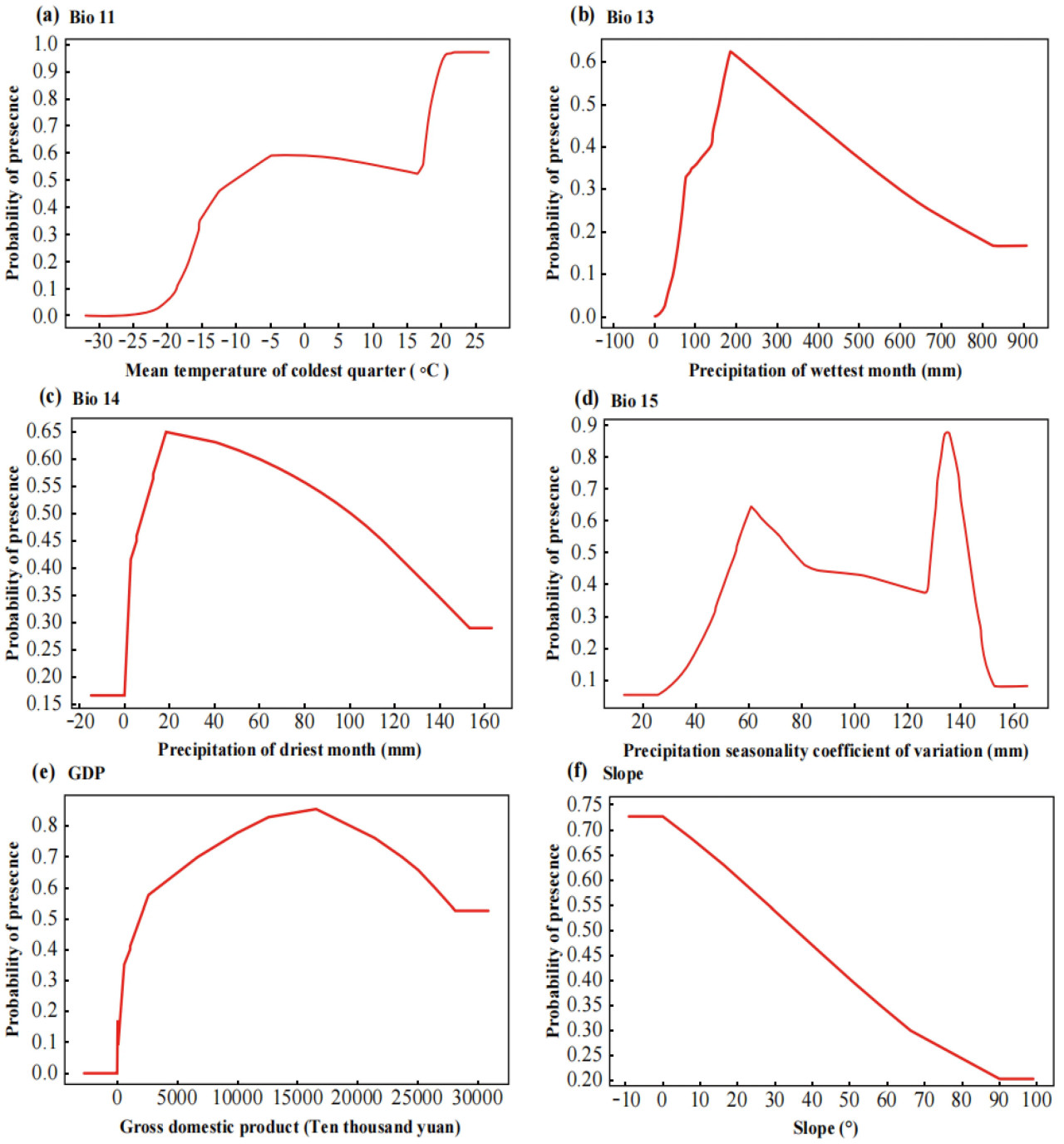
Рисунок 7. Кривые отклика пригодной для спаржи зоны на шесть основных влияющих переменных окружающей среды.
3.4. Пространственно-временные особенности распределения районов, пригодных для спаржи в Китае, в условиях исторических и будущих изменений климата
14 ключевых факторов окружающей среды были импортированы в модель MaxEnt для анализа распределения пригодных местообитаний для китайской спаржи в различных исторических и будущих климатических сценариях (SSP585, SSP370, SSP245 и SSP126). В исторические периоды общая пригодная площадь составляла 345,6 × 10⁵ км², что составляет 36% территории Китая, и была в основном сосредоточена в Восточном Китае. На этой территории наиболее пригодная площадь составляла 40,3 × 10⁵ км², в основном распределенная в провинциях Хэбэй, Фуцзянь, Шаньдун, Ляонин, Цзянсу, Хэнань, Шаньси, Шэньси, городах Тяньцзинь, Чунцин и провинции Сычуань. Среднепригодная площадь составляла 109,92 × 10⁵ км², в основном расположенная в центральных районах провинций Шаньдун, Хэнань и Шэньси в Китае. Малопригодная зона занимает площадь 192,51 × 10⁵ км², расположенную в основном в юго-западных, юго-восточных и северо-восточных регионах Китая (Рисунок 8j).
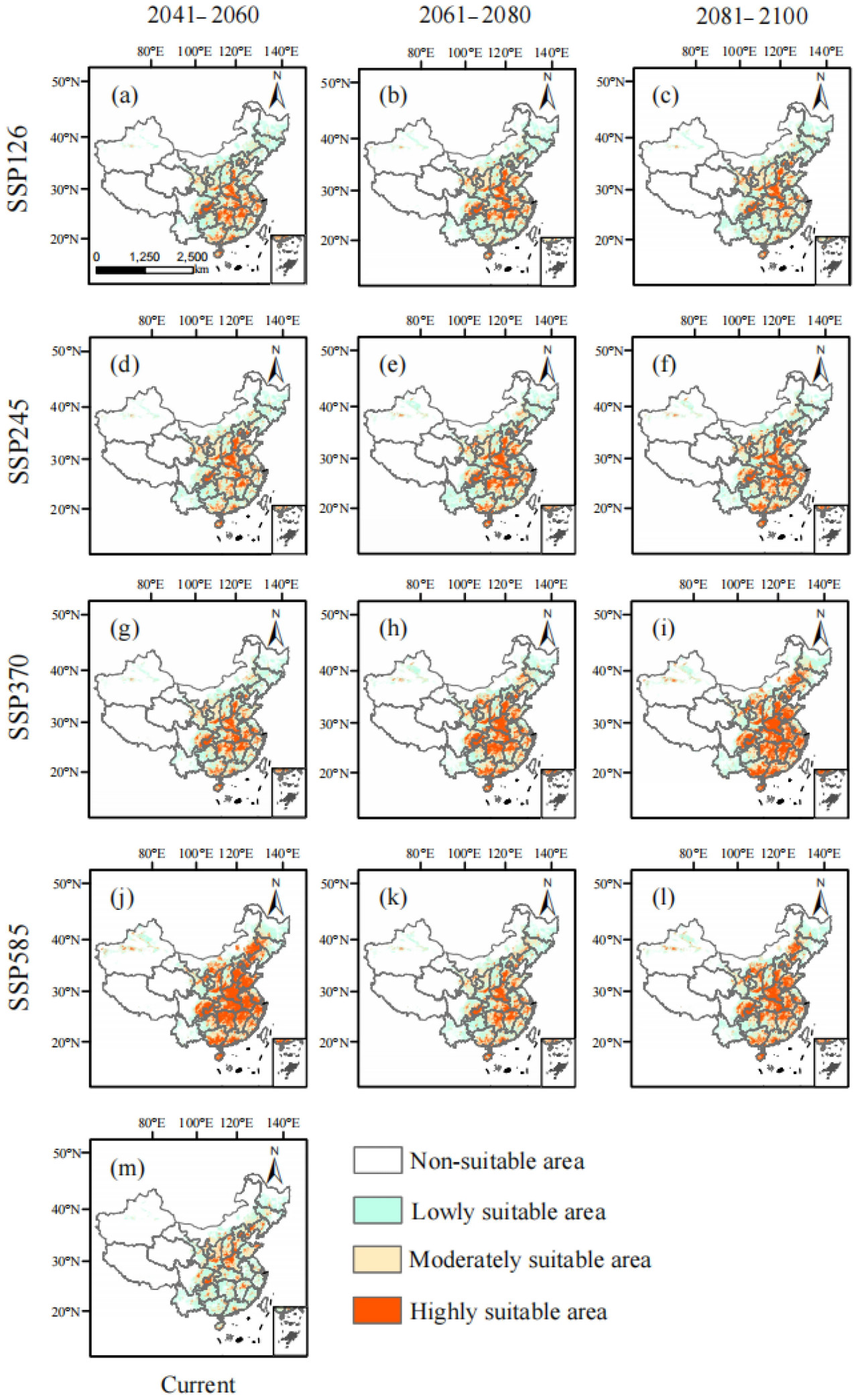
Рисунок 8. Районы пригодного распределения спаржи в Китае в различные исторические периоды (m) и будущие периоды и климатические сценарии ((a–c), (d–f), (g–i) и (j–l) относятся к периодам 2041–2060, 2061–2080 и 2081–2100 в сценариях SSP126, SSP245, SSP370 и SSP585).
Согласно модели MaxEnt, были спрогнозированы пригодные для выращивания спаржи районы в Китае при различных климатических сценариях на будущие периоды 2041–2060 (2050-е гг.), 2061–2080 (2070-е гг.) и 2081–2100 (2090-е гг.) (Рисунок 8a–l). Было спрогнозировано, что пригодная для спаржи площадь в 2050-х, 2070-х и 2090-х годах по сценарию SSP126 составит 373,52 × 10⁵, 379,62 × 10⁵ и 386,47 × 10⁵ км² соответственно. Площадь распределения ее пригодного местообитания была аналогична историческому периоду, но площадь увеличилась. По сравнению с историческим периодом, пригодная площадь увеличилась на 27,92 × 10⁵ км², 34,02 × 10⁵ км² и 40,87 × 10⁵ км² соответственно. Дополнительные пригодные районы были в основном распределены в провинциях Хубэй, Хунань, Чжэцзян и Цзянси. Пригодная для выращивания спаржи площадь составляла 384,15 × 10⁵ км², 400,11 × 10⁵ км² и 386,70 × 10⁵ км² в 2050-х, 2070-х и 2090-х годах по сценарию SSP245 соответственно, т.е. увеличение на 38,55 × 10⁵ км², 54,51 × 10⁵ км² и 41,60 × 10⁵ км² по сравнению с историческим периодом. Увеличенные пригодные районы произрастания были в основном распределены в Цзянси, Хубэе и Хунани. По сценарию SSP370 площадь пригодного местообитания для спаржи составляла 373,83 × 10⁵ км², 402,48 × 10⁵ км² и 438,68 × 10⁵ км² в будущие периоды. Она увеличилась на 28,23 × 10⁵ км², 56,88 × 10⁵ км² и 93,08 × 10⁵ км² по сравнению с историческим периодом, причем увеличенные пригодные районы были в основном распределены в Цзянси, Хубэе, Хунани, Чжэцзяне, Фуцзяни и Шанхае. Площадь пригодного местообитания для спаржи составляла 388,20 × 10⁵ км², 417,95 × 10⁵ км² и 437,52 × 10⁵ км² в будущий период по сценарию SSP585 соответственно. Это соответствует увеличению на 42,60 × 10⁵ км², 72,35 × 10⁵ км² и 91,92 × 10⁵ км² по сравнению с историческим периодом. Увеличенные пригодные районы были в основном распределены в провинциях Хунань, Цзилинь, Ляонин, Шанхай, Цзянси, Хубэй, Чжэцзян и Фуцзянь (Таблица 3).
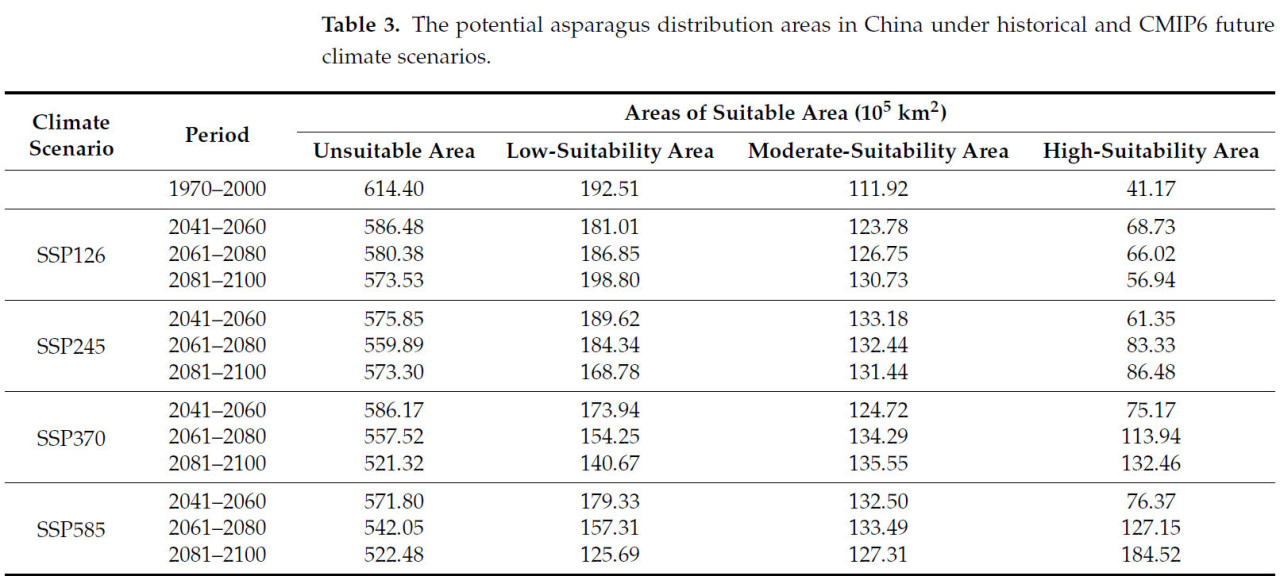
Таблица 3. Потенциальные районы распределения спаржи в Китае в условиях исторического и будущих климатических сценариев CMIP6.
Пригодные регионы для выращивания спаржи в Китае были в основном расположены в Цзилине, Ляонине, восточной Внутренней Монголии, южной Хэйлунцзян, южной Ганьсу, восточной Сычуани и Хайнани, а также в центральных и юго-восточных регионах в рамках сценариев SSP126, SSP245, SSP370 и SSP585. Также было несколько небольших разрозненных пятен, распределенных в Юньнани, Синьцзяне, Тибете и Цинхае (Рисунок 8). Тенденции изменения общей пригодной площади и высокопригодной площади от исторического периода к 2050-м годам, от 2050-х к 2070-м годам и от 2070-х к 2090-м годам демонстрируют последовательную картину, проявляя тенденцию к увеличению, затем уменьшению и снова увеличению. Изменения непригодных площадей уменьшились от настоящего времени к 2050-м годам, увеличились от 2050-х к 2070-м годам, а затем снова сократились от 2070-х к 2090-м годам (Таблица 4). Диапазон расширения был в основном расположен в северо-восточном и юго-западном Китае, а также в некоторых регионах Синьцзяна. Напротив, зона сокращения была в основном расположена в юго-западном Ляонине и северо-восточной Юньнани от наших дней до 2050-х годов при различных климатических сценариях. С 2050-х по 2070-е годы зона сокращения значительно уменьшилась по сравнению с периодом от исторического до 2050-х годов, и зона сокращения была в основном расположена в регионах Ляонин и Цзилинь. В период с 2070-х по 2090-е годы наблюдалось небольшое увеличение зоны сокращения, в основном сконцентрированное в Юньнани и восточных прибрежных регионах (Рисунок 9).
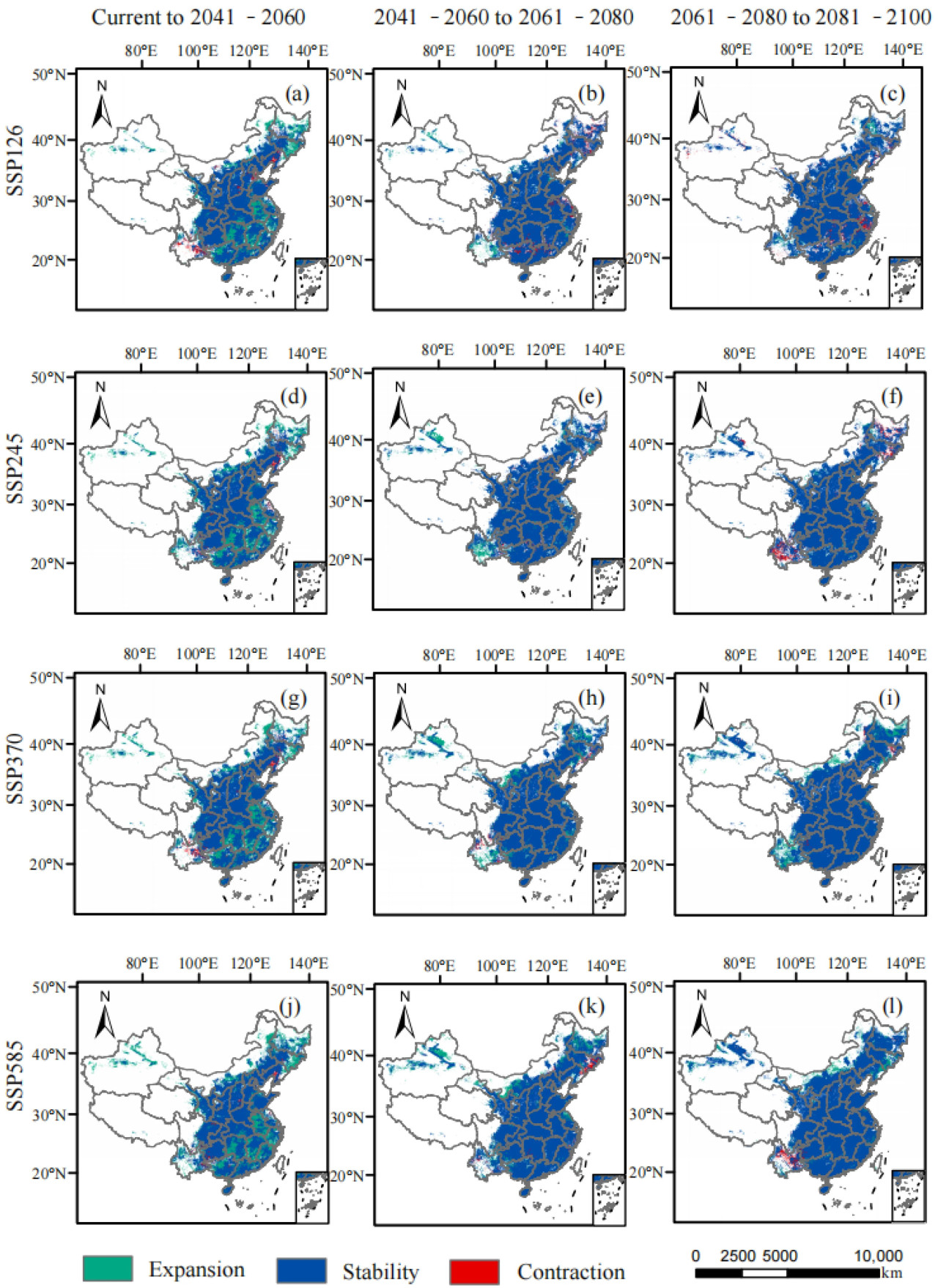
Рисунок 9. Изменения в структуре пригодной зоны спаржи в Китае в различные будущие периоды и при различных климатических сценариях ((a–c), (d–f), (g–i) и (j–l) соответствуют периодам от текущего до 2041–2060 гг., от 2041–2060 до 2061–2080 гг. и от 2061–2080 до 2081–2100 гг. в сценариях SSP126, SSP245, SSP370 и SSP585).
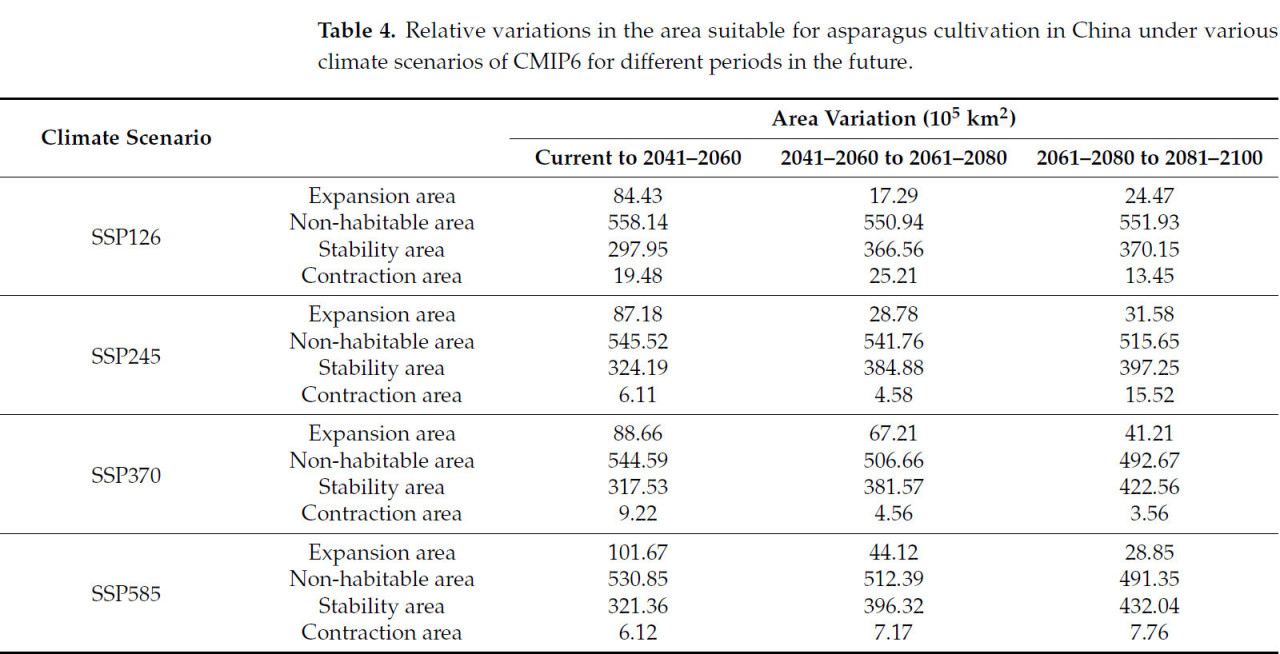
Таблица 4. Относительные изменения площади, пригодной для выращивания спаржи в Китае, при различных климатических сценариях CMIP6 для разных периодов в будущем.
3.5. Изменения центроида районов, пригодных для выращивания спаржи в Китае, в условиях будущего изменения климата
В этом исследовании был проанализирован путь миграции центроида районов, пригодных для спаржи, как в историческом, так и в различных будущих климатических сценариях. Было обнаружено, что центральная точка районов, пригодных для выращивания спаржи в Китае, имеет тенденцию к миграции на север в будущем. По мере усиления изменения климата в будущем миграция центроида пригодной для спаржи зоны проявляла тенденцию сначала приближаться, а затем удаляться, при этом расстояние миграции постепенно увеличивалось (Рисунок 10).
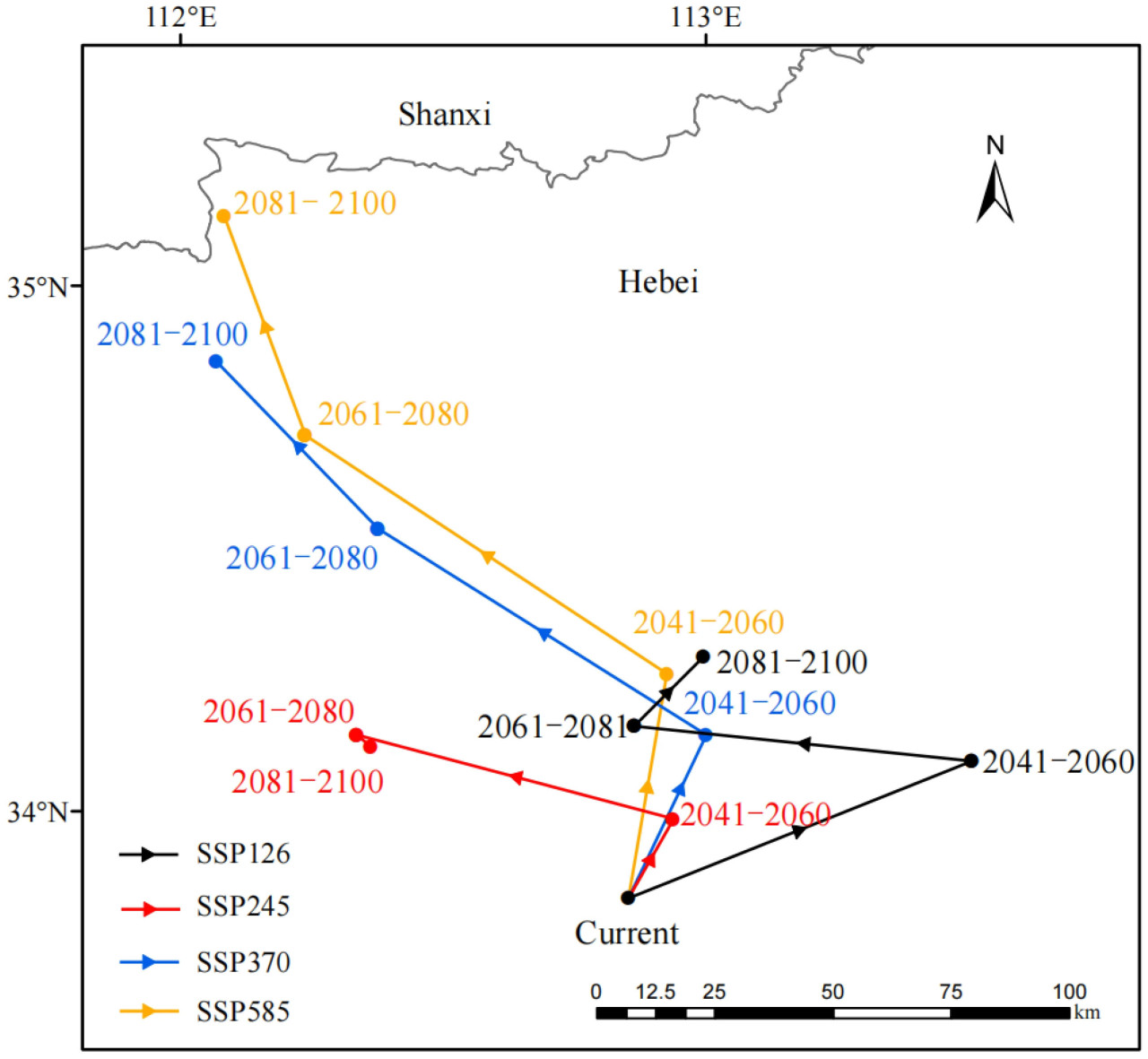
Рисунок 10. Будущие изменения в миграции центральной точки районов, пригодных для спаржи, при различных климатических сценариях.
Центральная точка района, пригодного для выращивания спаржи, находилась в центрально-западной части провинции Хэнань (112,85° в.д., 33,84° с.ш.) (Рисунок 10). Центроид пригодной для спаржи зоны в целом сместился на север по сценарию SSP126. Сначала он сместился на 67,82 км на северо-восток до города Сюйчан в провинции Хэнань (113,51° в.д., 34,11° с.ш.), затем на 60,04 км на северо-запад до города Пиндиншань в провинции Хэнань (112,86° в.д., 34,16° с.ш.); затем он сместился на 19,61 км на северо-восток до города Пиндиншань в провинции Хэнань (112,99° в.д., 34,30° с.ш.). Центроид спаржи в целом сместился на северо-запад по сценарию SSP245. Сначала он переместился на 18,62 км на северо-восток до города Пиндиншань в провинции Хэнань (112,94° в.д., 33,99° с.ш.), затем переместился на 58,02 км на северо-запад до города Лоян в провинции Хэнань (112,34° в.д., 34,15° с.ш.), а затем переместился на 2,89 км на юго-запад до города Лоян в провинции Хэнань (112,36° в.д., 34,13° с.ш.). По сценарию SSP370 центральная точка зоны пригодности спаржи также сместилась, в целом, на северо-запад. Сначала она переместилась на 37,12 км до города Пиндиншань в провинции Хэнань (113,00° в.д., 34,15° с.ш.) в северо-восточном направлении, затем переместилась на 71,51 км до города Лоян в провинции Хэнань (112,38° в.д., 34,54° с.ш.) в северо-западном направлении, а затем переместилась на 45,46 км до города Лоян в провинции Хэнань (112,07° в.д., 34,86° с.ш.) в северо-западном направлении. Центроид в целом сместился на северо-запад по сценарию SSP585. Сначала он переместился на 47,25 км в северо-восточном направлении до города Пиндиншань в провинции Хэнань (112,93° в.д., 34,26° с.ш.), затем переместился на 81,25 км в северо-западном направлении до города Лоян в провинции Хэнань (112,24° в.д., 34,72° с.ш.) и, наконец, переместился на 47,84 км в северо-западном направлении до города Цзиюань в провинции Хэнань (112,08° в.д., 35,13° с.ш.) (Таблица 5).
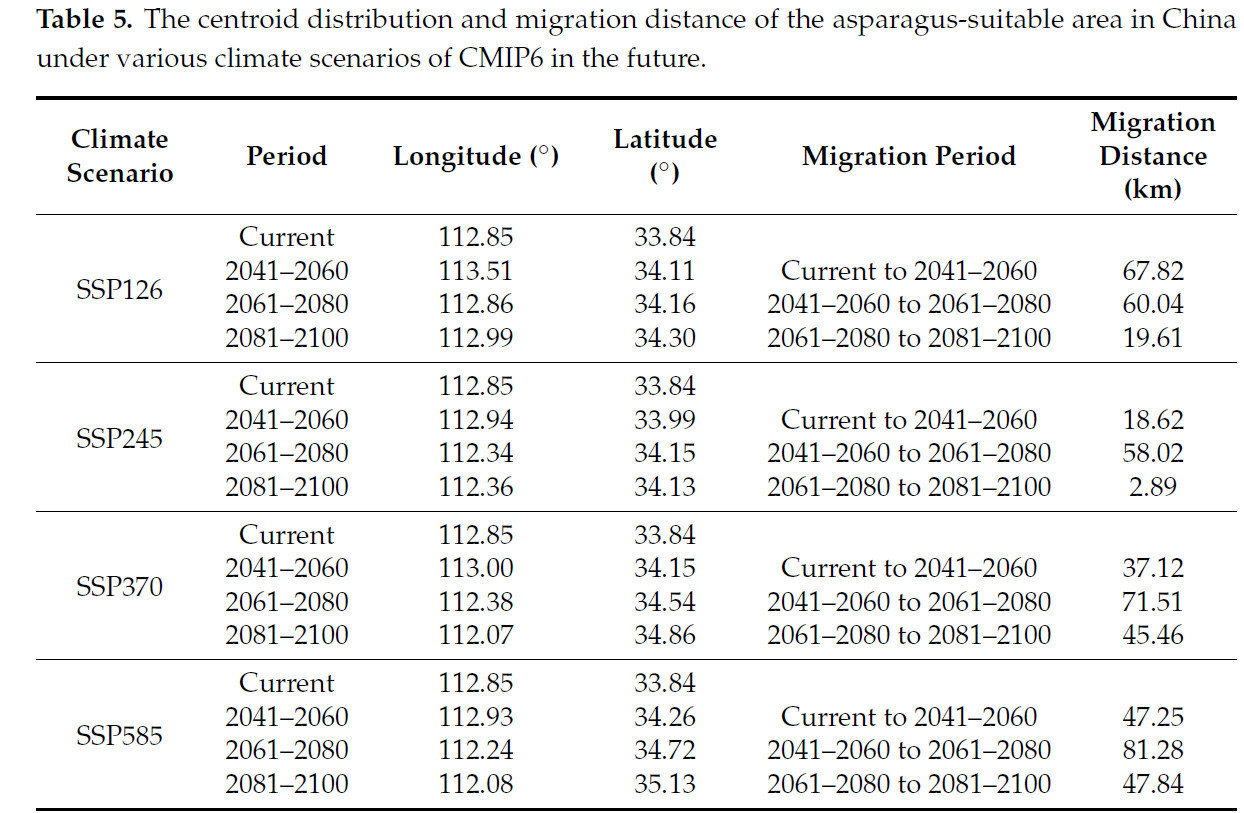
Таблица 5. Распределение центроида и расстояние миграции пригодной для спаржи зоны в Китае при различных будущих климатических сценариях CMIP6.
4. Обсуждение
Модель MaxEnt является желательной моделью для прогнозирования как фактического, так и потенциального распространения видов и все чаще используется в биологии инвазий [45,46], биологии сохранения и для определения влияния глобального изменения климата на распространение видов [47,48]. Хотя широкое использование исследований MaxEnt часто опирается на настройки по умолчанию, т.е. без обработки данных и параметризации модели, точность модели сильно зависит от информации о выборке видов, обработки переменных окружающей среды и настройки параметров [49]. Чтобы предотвратить влияние переобучения на точность модели, в этом исследовании данные о распространении спаржи были обработаны с помощью программного обеспечения ENMTools, а экологические данные — с помощью теста корреляции Пирсона. Для оптимизации модели MaxEnt использовался пакет Kuenm для языка R. Точность оптимизированной модели составила AUC = 0,931, что указывает на то, что она может точно прогнозировать потенциальные зоны пригодности спаржи в Китае.
Климат, рельеф, почва и деятельность человека являются основными факторами окружающей среды, определяющими распространение видов [50,51,52]. Это исследование показало, что осадки (Bio13, Bio14 и Bio19), температура (Bio3, Bio5, Bio8 и Bio11), уклон и ВВП являются основными факторами окружающей среды, влияющими на лежащее в основе географическое распределение спаржи, с совокупным вкладом 92,2%. ВВП отражает общий уровень социально-экономического развития, а также изменения рыночного спроса. С ростом ВВП увеличение инвестиций в сельское хозяйство, стимулирование потребительского спроса, а также прогресс и внедрение технологий будут побуждать фермеров корректировать структуру посевов сельскохозяйственных культур, исходя из экономической выгоды и рыночных перспектив, тем самым достигая более высокой стоимости продукции и прибыли [53]. В то же время такие факторы, как урбанизация, загрязнение окружающей среды и деградация экосистем, также будут оказывать давление на сельскохозяйственное производство и в дальнейшем влиять на распределение посевов сельскохозяйственных культур [54,55]. Спаржа обладает сильной адаптируемостью к температуре, будучи одновременно холодо- и жаростойкой; когда температура почвы повышается до 5 °C, спаржа прорастает; и она будет нормально расти при температуре около 10 °C. Оптимальный температурный диапазон для образования нежных побегов составляет от 15 °C до 17 °C. Низкие и высокие температуры могут ингибировать фотосинтез и влиять на развитие спаржи [56]. Спаржа впадает в летний покой, когда температура поднимается выше 30 °C. Надземные и подземные части впадают в состояние покоя для лучшей перезимовки, когда зимние температуры в северном регионе слишком низкие [57]. Хотя хорошо развитая корневая система спаржи может выдерживать засуху, она восприимчива к переувлажнению, и чрезмерное количество осадков может вызвать корневую гниль [58,59]. Пахотные земли на склонах играют ключевую роль в производстве продуктов питания, разнообразии экосистем и экономическом развитии. Уклон влиял на глубину почвы, почвенное дыхание и скорость использования питательных веществ, причем порог выживания спаржи был больше 0,5, когда уклоны земли были менее 40°. Wang и др. [60] количественно проанализировали пространственное распределение 12 основных культур на крутых склонах (>12%) в Европе и обнаружили, что сельское хозяйство на крутых склонах было уязвимо к засухе. Это исследование также показало, что более пологие склоны были более подходящими для выживания спаржи. Деятельность человека существенно влияет на глобальное изменение климата, и уровень экономического развития играет решающую роль в продуктивности сельского хозяйства. Среди антропоцентрических факторов ВВП оказывает наибольшее влияние на пригодность местообитания для спаржи, составляя 71,6%. Определив основные экологические факторы, влияющие на распространение спаржи, можно разработать рациональные стратегии зонирования для облегчения интродукции и выращивания спаржи, а также сохранения урожая.
Это исследование показало, что будущее изменение температуры привело к смещению центроида районов, пригодных для выращивания спаржи, на север, с максимальным расстоянием миграции 67,82 км. Сдвиг центроида свидетельствует о том, что пригодные для спаржи районы могут измениться из-за изменения климата, что побудит фермеров корректировать места посадки для адаптации к новым климатическим условиям. Чтобы обеспечить устойчивое развитие спаржи, важно будет оптимизировать управление водными ресурсами и ирригационные системы, а также корректировать производственные сезоны и методы землепользования. Пригодные для спаржи районы сместились в районы с меньшим количеством осадков. Это хорошо согласовывалось с характеристиками засухоустойчивости, но не с характеристиками устойчивости к переувлажнению спаржи [61]. Кроме того, зоны средней и высокой пригодности для спаржи в Китае демонстрировали растущую тенденцию, в то время как зоны непригодные и малопригодные проявляли тенденцию к сокращению в различных будущих климатических сценариях. С повышением температуры пригодная для спаржи площадь имеет тенденцию к увеличению, но этому расширению есть пределы. Наши результаты свидетельствуют о том, что прогнозируется увеличение площади, непригодной для спаржи в Китае, с 2081 по 2100 год по сценарию SSP245, вероятно, из-за повышения температуры, превышающего подходящий температурный диапазон роста для спаржи.
Основные точки распространения в этом исследовании были получены из информации о распространении образцов, которая указывает только на существование вида в конкретных местах. Эти данные не отражают точно все районы распространения или потенциально пригодные местообитания. Однако из-за ограниченности доступных баз данных фактическая область распространения спаржи может быть не полностью охвачена, и это исследование было в основном сосредоточено на Китае. Оптимизированная модель MaxEnt, использованная в этом исследовании, может быть применена в последующих исследованиях для прогнозирования глобального распространения культуры спаржи. Кроме того, это исследование опиралось исключительно на модель MaxEnt, и результаты могут быть неопределенными из-за влияния структуры этой модели. Будущие исследования могли бы включить несколько моделей распространения видов для анализа районов, пригодных для спаржи, тем самым повышая надежность их результатов.
5. Выводы
В этом исследовании учитывалось влияние почвы, климата, топографии и деятельности человека на распределение культуры спаржи. С помощью модели MaxEnt и инструментов ArcGIS были определены основные экологические параметры, влияющие на пригодное для спаржи распределение. Дополнительно это исследование спрогнозировало и проанализировало пространственно-временные изменения и вариации потенциальных пригодных для спаржи районов в Китае. Было обнаружено, что параметрически оптимизированная модель MaxEnt высокоэффективна в прогнозировании районов, пригодных для спаржи, с AUC 0,931. ВВП, уклон, Bio11, Bio13, Bio14 и Bio15 были основными факторами окружающей среды, влияющими на районы, пригодные для спаржи в Китае. Высокопригодные для спаржи районы были расположены в провинциях Хэбэй, Фуцзянь, Шаньдун, Ляонин, Цзянсу, Хэнань, Шаньси, Шэньси, городах Тяньцзинь, Чунцин и провинции Сычуань в Китае. Под влиянием будущего изменения климата ожидается расширение районов, пригодных для спаржи. Это расширение пригодных районов сосредоточено на юго-западе Китая, северо-востоке Китая и некоторых частях Синьцзяна. Дополнительно несколько зон расширения были обнаружены в провинциях Внутренняя Монголия, Цинхай и Ганьсу, в то время как зоны сокращения в основном были обнаружены на северо-востоке Юньнани и юго-западе провинции Ляонин. Примечательно, что, согласно прогнозам, центральные точки районов, пригодных для спаржи, сместятся на север из-за будущего изменения климата, но все они будут расположены в провинции Хэнань. Эти результаты могут дать ценную информацию для планирования выращивания спаржи, содействия развитию высококачественной и высокоурожайной спаржевой индустрии и эффективного решения проблем, связанных с изменением климата.
Ссылки
1. Dawid, C.; Hofmann, T. Identification of sensory-active phytochemicals in asparagus (Asparagus officinalis L.). J. Agric. Food Chem. 2012, 60, 11877–11888. [Google Scholar] [CrossRef] [PubMed]
2. He, C.Y.; Hsiang, T.; Wolyn, D.J. Induction of systemic disease resistance and pathogen defence responses in Asparagus officinalis inoculated with nonpathogenic strains of Fusarium oxysporum. Plant Pathol. 2002, 51, 225–230. [Google Scholar] [CrossRef]
3. Sharma, U.; Kumar, N.; Singh, B.; Munshi, R.K.; Bhalerao, S. Immunomodulatory active steroidal saponins from Asparagus racemosus. Med. Chem. Res. 2013, 22, 573–579. [Google Scholar] [CrossRef]
4. Słupski, J.; Korus, A.; Lisiewska, Z.; Kmiecik, W. Content of amino acids and the quality of protein in as-eaten green asparagus (Asparagus officinalis L.) products. Int. J. Food Sci. Technol. 2010, 45, 733–739. [Google Scholar] [CrossRef]
5. Chitrakar, B.; Zhang, M.; Adhikari, B. Asparagus (Asparagus officinalis): Processing effect on nutritional and phytochemical composition of spear and hard-stem byproducts. Trends Food Sci. Technol. 2019, 93, 1–11. [Google Scholar] [CrossRef]
6. Liu, D.; Yan, F.; Liu, C.; Chen, A.; Wu, J.; Yu, M.; Lyu, X. Develo EST-SSR Markers for Identifying and Evaluating Asparagus Germplasm Resources Based on Transcriptome Sequences. Horticulturae 2024, 10, 121. [Google Scholar] [CrossRef]
7. Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar]
8. Forster, P.M.; Smith, C.J.; Walsh, T.; Lamb, W.F.; Lamboll, R.; Hauser, M.; Ribes, A.; Rosen, D.; Gillett, N.; Palmer, M.D.; et al. Indicators of Global Climate Change 2022: Annual update of large-scale indicators of the state of the climate system and human influence. Earth Syst. Sci. Data 2023, 15, 2295–2327. [Google Scholar] [CrossRef]
9. Gregory, P.J.; Ingram, J.S.; Brklacich, M. Climate change and food security. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 2139–2148. [Google Scholar] [CrossRef]
10. Adhikari, D.; Barik, S.K.; Upadhaya, K. Habitat distribution modelling for reintroduction of Ilex khasiana Purk., a critically endangered tree species of northeastern India. Ecol. Eng. 2012, 40, 37–43. [Google Scholar] [CrossRef]
11. Chardon, N.I.; Cornwell, W.K.; Flint, L.E.; Flint, A.L.; Ackerly, D.D. Topographic, latitudinal and climatic distribution of Pinus coulteri: Geographic range limits are not at the edge of the climate envelope. Ecography 2015, 38, 590–601. [Google Scholar] [CrossRef]
12. Chen, I.C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid range shifts of species associated with high levels of climate warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef] [PubMed]
13. Bowen, A.K.; Stevens, M.H. Temperature, topography, soil characteristics, and NDVI drive habitat preferences of a shade-tolerant invasive grass. Ecol. Evol. 2020, 10, 10785–10797. [Google Scholar] [CrossRef] [PubMed]
14. Ling, X.; Zhang, Z.; Zhai, J.; Ye, S.; Huang, J. Advances in research on the effects of climate change on rice production in China. Acta Crop. Sin. 2019, 45, 323–334. [Google Scholar]
15. Unkovich, M.; McBeath, T.; Llewellyn, R.; Hall, J.; Gupta, V.V.; Macdonald, L.M. Challenges and opportunities for grain farming on sandy soils of semi-arid south and south-eastern Australia. Soil Res. 2020, 58, 323–334. [Google Scholar] [CrossRef]
16. Atanasso, J.A.; Mensah, S.; Salako, K.V.; Tohoun, R.J.; Kakaï, R.G.; Assogbadjo, A.E. Factors affecting survival of seedling of Afzelia africana, a threatened tropical timber species in West Africa. Trop. Ecol. 2021, 62, 443–452. [Google Scholar] [CrossRef]
17. Ong, P.-W.; Lin, Y.-P.; Chen, H.-W.; Lo, C.-Y.; Burlyaeva, M.; Noble, T.; Nair, R.M.; Schafleitner, R.; Vishnyakova, M.; Bishop-Von-Wettberg, E.; et al. Environment as a limiting factor of the historical global spread of mungbean. Elife 2023, 12, e85725. [Google Scholar] [CrossRef]
18. Higgins, S.I.; O’Hara, R.B.; Römermann, C. A niche for biology in species distribution models. J. Biogeogr. 2012, 39, 2091–2095. [Google Scholar] [CrossRef]
19. Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
20. Warren, D.L.; Seifert, S.N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef] [PubMed]
21. Hernandez, P.A.; Graham, C.H.; Master, L.L.; Albert, D.L. The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 2006, 29, 773–785. [Google Scholar] [CrossRef]
22. Elith, J.H.; Graham, C.P.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
23. Fitzgibbon, A.; Pisut, D.; Fleisher, D. Evaluation of Maximum Entropy (Maxent) machine learning model to assess relationships between climate and corn suitability. Land 2022, 11, 1382. [Google Scholar] [CrossRef]
24. Maguranyanga, C.; Murwira, A. Mapping maize, tobacco, and soybean fields in large-scale commercial farms of Zimbabwe based on multitemporal NDVI images in MAXENT. Can. J. Remote Sens. 2014, 40, 396–405. [Google Scholar] [CrossRef]
25. Zhao, C.; Zhang, F.; Huang, J.; Zhang, Q.; Lu, Y.; Cao, W. Prediction of the Climatically Suitable Areas of Rice in China Based on Optimized MaxEnt Model. Int. J. Plant Prod. 2024, 18, 549–561. [Google Scholar] [CrossRef]
26. Kogo, B.K.; Kumar, L.; Koech, R.; Kariyawasam, C.S. Modelling climate suitability for rainfed Maize cultivation in Kenya using a Maximum Entropy (MaxENT) approach. Agronomy 2019, 9, 727. [Google Scholar] [CrossRef]
27. Yu, X.; Tao, X.; Liao, J.; Liu, S.; Xu, L.; Yuan, S.; Zhang, Z.; Wang, F.; Deng, N.; Huang, J.; et al. Predicting potential cultivation region and paddy area for ratoon rice production in China using Maxent model. Field Crops Res. 2022, 275, 108372. [Google Scholar] [CrossRef]
28. Holder, A.M.; Markarian, A.; Doyle, J.M.; Olson, J.R. Predicting geographic distributions of fishes in remote stream networks using maximum entropy modeling and landscape characterizations. Ecol. Model. 2020, 433, 109231. [Google Scholar] [CrossRef]
29. Halvorsen, R.; Mazzoni, S.; Bryn, A.; Bakkestuen, V. Opportunities for improved distribution modelling practice via a strict maximum likelihood interpretation of MaxEnt. Ecography 2015, 38, 172–183. [Google Scholar] [CrossRef]
30. Montenegro, C.; Solitario, L.A.; Manglar, S.F.; Guinto, D.D. Niche modelling of endangered philippine birds using GARP and MAXENT. In Proceedings of the 2017 7th International Conference on Cloud Computing, Data Science & Engineering-Confluence, Noida, India, 12–13 January 2017; pp. 547–551. [Google Scholar]
31. Zhang, S.; Liu, X.; Li, R.; Wang, X.; Cheng, J.; Yang, Q.; Kong, H. AHP-GIS and MaxEnt for delineation of potential distribution of Arabica coffee plantation under future climate in Yunnan, China. Ecol. Indic. 2021, 132, 108339. [Google Scholar] [CrossRef]
32. Warren, D.L.; Glor, R.E.; Turelli, M. ENMTools: A toolbox for comparative studies of environmental niche models. Ecography 2010, 33, 607–611. [Google Scholar] [CrossRef]
33. Prasad, S.R.; Sharma, P. Species distribution modeling of asparagus (Asparagus officinalis) in India using MaxEnt model. Environ. Monit. Assess. 2020, 192, 518. [Google Scholar]
34. Yang, X.Q.; Kushwaha, S.P.S.; Saran, S.; Xu, J.; Roy, P.S. Maxent modeling for predicting the potential distribution of medicinal plant, Justicia adhatoda L. in Lesser Himalayan foothills. Ecol. Eng. 2013, 51, 83–87. [Google Scholar] [CrossRef]
35. Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
36. Cobos, M.E.; Peterson, A.T.; Barve, N.; Osorio-Olvera, L. kuenm: An R package for detailed development of ecological niche models using Maxent. PeerJ 2019, 7, e6281. [Google Scholar] [CrossRef]
37. Elith, J.; Leathwick, J.R.; Hastie, T. A working guide to boosted regression trees. J. Anim. Ecol. 2011, 77, 802–813. [Google Scholar] [CrossRef]
38. Yu, D.; Deng, L.; Acero, A. Using continuous features in the maximum entropy model. Pattern Recognit. Lett. 2009, 30, 1295–1300. [Google Scholar] [CrossRef]
39. Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
40. Anderson, R.P.; Lew, D.; Peterson, A.T. Evaluating predictive models of species’distributions: Criteria for selecting optimal models. Ecol. Model. 2003, 162, 211–232. [Google Scholar] [CrossRef]
41. Symonds, M.R.; Moussalli, A. A brief guide to model selection, multimodel inference and model averaging in behavioural ecology using Akaike’s information criterion. Behav. Ecol. Sociobiol. 2011, 65, 13–21. [Google Scholar] [CrossRef]
42. Zhou, Y.; Lu, X.; Zhang, G. Potentially differential impacts on niche overlap between Chinese endangered Zelkova schneideriana and its associated tree species under climate change. Front. Ecol. Evol. 2023, 11, 1218149. [Google Scholar] [CrossRef]
43. Jalaeian, M.; Golizadeh, A.; Sarafrazi, A.; Naimi, B. Inferring climatic controls of rice stem borers spatial distributions using maximum entropy modelling. J. Appl. Entomol. 2018, 142, 388–396. [Google Scholar] [CrossRef]
44. Zhang, K.; Yao, L.; Meng, J.; Tao, J. Maxent modeling for predicting the potential geographical distribution of two peony species under climate change. Sci. Total Environ. 2018, 634, 1326–1334. [Google Scholar] [CrossRef] [PubMed]
45. Morán Ordóñez, A.; Lahoz Monfort, J.J.; Elith, J.; Wintle, B.A. Evaluating 318 continental-scale species distribution models over a 60-year prediction horizon: What factors influence the reliability of predictions? Glob. Ecol. Biogeogr. 2017, 26, 371–384. [Google Scholar] [CrossRef]
46. Wan, J.Z.; Wang, C.J.; Yu, F.H. Effects of occurrence record number, environmental variable number, and spatial scales on MaxEnt distribution modelling for invasive plants. Biologia 2019, 74, 757–766. [Google Scholar] [CrossRef]
47. Searcy, C.A.; Shaffer, H.B. Do ecological niche models accurately identify climatic determinants of species ranges? Am. Nat. 2016, 187, 423–435. [Google Scholar] [CrossRef]
48. Sutton, G.F.; Martin, G.D. Testing MaxEnt model performance in a novel geographic region using an intentionally introduced insect. Ecol. Model. 2022, 473, 110139. [Google Scholar] [CrossRef]
49. Song, R.; Ma, Y.; Hu, Z.; Li, Y.; Li, M.; Wu, L.; Li, C.; Dao, E.; Fan, X.; Hao, Y.; et al. MaxEnt modeling of Dermacentor marginatus (Acari: Ixodidae) distribution in Xinjiang, China. J. Med. Entomol. 2020, 57, 1659–1667. [Google Scholar]
50. Cardoza-Martínez, G.F.; Becerra-López, J.L.; Esparza-Estrada, C.E.; Estrada-Rodríguez, J.L.; Czaja, A.; Ehsan, M.; Baltierra-Trejo, E.; Romero-Méndez, U. Shifts in Climatic Niche Occupation in Astrophytum Coahuilense (H. Möller) Kayser and Its Potential Distribution in Mexico. Sustainability 2019, 11, 1138. [Google Scholar] [CrossRef]
51. Davis, M.B.; Shaw, R.G. Range shifts and adaptive responses to Quaternary climate change. Science 2001, 292, 673–679. [Google Scholar] [CrossRef] [PubMed]
52. Pajunen, V.; Luoto, M.; Soininen, J. Climate is an important driver for stream diatom distributions. Glob. Ecol. Biogeogr. 2016, 25, 198–206. [Google Scholar] [CrossRef]
53. Smith, J.; Johnson, M.; Lee, H. Economic development and agricultural restructuring: A global perspective. Agric. Econ. Rev. 2015, 42, 67–82. [Google Scholar]
54. Zhang, Y.; Li, X. Impact of urbanization on agricultural production: A case study in China. J. Rural. Dev. 2018, 29, 145–160. [Google Scholar]
55. Zhou, T.; Liu, S.; Wang, F. Environmental degradation and its effect on crop distribution in China. Environ. Sci. Technol. 2020, 54, 5724–5733. [Google Scholar]
56. Radosavljevic, A.; Anderson, R.P. Making better Maxent models of species distributions: Complexity, overfitting and evaluation. J. Biogeogr. 2014, 41, 629–643. [Google Scholar] [CrossRef]
57. Short, W.; Wolyn, D.J. The role of drought stress on the acquisition of freezing tolerance in asparagus under controlled conditions. Can. J. Plant Sci. 2022, 102, 864–874. [Google Scholar] [CrossRef]
58. Drost, D.; Wilcox-Lee, D. Tillage alters root distribution in a mature asparagus planting. Sci. Hortic. 2000, 83, 187–204. [Google Scholar] [CrossRef]
59. Namaki, A.; Ghahremani, Z.; Aelaei, M.; Barzegar, T.; Ranjbar, M.E. The first report of drought tolerance assessment of Iranian asparagus. Gesunde Pflanz. 2022, 74, 141–149. [Google Scholar] [CrossRef]
60. Wang, W.; Straffelini, E.; Tarolli, P. 44% of steep slope cropland in Europe vulnerable to drought. Geogr. Sustain. 2024, 5, 89–95. [Google Scholar] [CrossRef]
61. Mantovani, D.; Rosati, A.; Perrone, D. Photosynthetic characterization and response to drought and temperature in wild asparagus (Asparagus acutifolius L.). HortScience 2019, 54, 1039–1043. [Google Scholar] [CrossRef]

Yang Q, Ji C, Li N, Lin H, Li M, Li H, Heng S, Liang J. Analysis of the Distribution Pattern of Asparagus in China Under Climate Change Based on a Parameter-Optimized MaxEnt Model. Agriculture. 2025; 15(3):320. https://doi.org/10.3390/agriculture15030320
Перевод статьи «Analysis of the Distribution Pattern of Asparagus in China Under Climate Change Based on a Parameter-Optimized MaxEnt Model» авторов Yang Q, Ji C, Li N, Lin H, Li M, Li H, Heng S, Liang J., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык
Фото: freepik



















































































































Комментарии (0)