Что скрывает почва манговых садов? Сравнение трёх систем земледелия
Деградация земель представляет собой серьёзную угрозу для экосистем. Долгосрочное применение традиционных методов ведения сельского хозяйства может приводить к сильной деградации почв и снижению урожайности культур, что создаёт проблемы как для местных, так и для мировых сообществ. Настоящее исследование было проведено с целью выяснить, как физико-химические свойства почвы и структура микробных сообществ реагируют на смену систем земледелия. В работу были включены садовые участки на склоновых землях при трёх подходах к ведению хозяйства: традиционном (CF), органическом (OF) и экологически безопасном (EFF).
Аннотация
Было установлено, что запасы углерода в поверхностном слое почвы (0–30 см) при использовании систем EFF и OF возросли в 3,6 и 5,1 раза соответственно. Применение EFF и OF также значительно увеличило содержание общего азота (на 0,33–0,46 г/кг), аммонийного азота (на 3,0–7,3 г/кг) и углерода микробной биомассы (на 0,56–1,04 г/кг), однако при этом привело к снижению pH как минимум на 0,6 единицы и уменьшению содержания доступного фосфора на 104–114 мг/кг. Использование фосфорсодержащих гербицидов и минеральных удобрений в системе CF, по-видимому, способствовало повышению уровня фосфора и нитратов в этих почвах.
Высокое обилие бактерий типов Acidobacteria и Actinobacteria в почвах при системах EFF и OF, вероятно, связано с дефицитом фосфора в этих условиях. В то же время обилие грибов в почве при органическом земледелии (OF) позволяет предположить, что потребность растений в доступном фосфоре стимулирует грибную минерализацию органических форм фосфора. В почвах при традиционном земледелии (CF), напротив, отмечено высокое обилие бактерий классов Gammaproteobacteria, Planctomycetes, Firmicutes и Nitrospirae, что, возможно, обусловлено регулярным внесением фосфорсодержащих гербицидов и удобрений с высоким содержанием общего азота.
Введение
Деградация земель является ключевой глобальной проблемой, требующей незамедлительного внимания. Устойчивые методы управления почвами могут смягчать последствия изменения климата (Lal, 2004) и эффективно решать проблемы, связанные с ростом численности населения мира и отсутствием продовольственной безопасности (Gnanavelrajah et al., 2008; Lal, 2018). Для устойчивого управления почвами необходимо сохранять качество почвы, в частности, путём увеличения запасов органического вещества и биоразнообразия с помощью различных агроэкологических практик, таких как внедрение систем органического земледелия (OF), снижение применения химических средств и повышение видового разнообразия растений за счёт севооборота или агролесоводства (Van Wesemael et al., 2010; Amelung et al., 2020).
Продовольственные потребности и дефицит земельных ресурсов приводят к интенсивному ведению хозяйства и чрезмерному использованию химических удобрений в почвах сельскохозяйственных угодий на Тайване. В условиях тропического и субтропического климата содержание почвенного органического углерода (SOC) в пахотных почвах Тайваня, как правило, ниже 20 г/кг (Huang, 1994). Для смягчения последствий изменения климата и восстановления агроэкосистем органическое земледелие (OF) и экологически безопасное земледелие (EFF) привлекли большое внимание исследователей, изучающих устойчивость и продуктивность сельскохозяйственных почв. Как известно, традиционное земледелие (CF) часто связано с чрезмерным применением химических удобрений и гербицидов, что ускоряет деградацию почв и создаёт экологические риски. Напротив, системы OF и EFF предполагают использование органических удобрений, которые повышают урожайность культур за счёт увеличения содержания органического вещества в почве (гумуса) и улучшения качества почвы. Это увеличение содержания органического вещества усиливает биологическую активность почвы, улучшает структуру почвы, инфильтрацию и удержание воды, а также накопление питательных веществ (Peltoniemi et al., 2021). Выяснение влияния методов управления на качество почвы и микробные сообщества должно стать ключевым шагом для смягчения деградации почв и решения проблем продовольственной безопасности в условиях изменения климата.
Помимо запасов SOC, бактериальные сообщества играют ключевую роль в почвенных экосистемах, поскольку они опосредуют многие почвенные функции и процессы, такие как разложение органического вещества, круговорот питательных веществ и азотфиксацию (Bender et al., 2016). Эффективность использования азота зависит от обильности почвенных бактерий, участвующих в азотфиксации и нитрификации (Levy-Booth et al., 2014). Разнообразие почвенных бактерий может служить биоиндикатором качества почвы (Pereira e Silva et al., 2013). В нескольких исследованиях было показано, что методы ведения сельского хозяйства изменяют состав, разнообразие, активность и биомассу почвенных микроорганизмов. Состав бактериального сообщества почвы различается в системах OF и CF; микробная активность, а также содержание углерода и азота в микробной биомассе выше в системе OF, чем в системе CF (Peltoniemi et al., 2021). Сообщается, что в системе OF повышается обильность почвенных бактерий, связанных с циклами углерода, азота, фосфора и серы (Zhang et al., 2019). Помимо наблюдаемых изменений в составе почвы, OF приводило к повышению урожайности; кроме того, OF положительно влияло на pH почвы, содержание кальция, фосфора и ферментативную активность (Durrer et al., 2021). Для понимания влияния методов сельскохозяйственного управления на почвенную экосистему необходимо проведение комплексных исследований изменений физико-химических свойств почвы и микробных сообществ при различных методах сельскохозяйственного управления.
Таким образом, данное исследование было направлено на: (1) моделирование и понимание пространственно-временных изменений в реакциях SOC и микробных сообществ на смену систем земледелия; (2) выяснение взаимосвязей между методами сельскохозяйственного управления, свойствами почвы, микробным разнообразием и запасами почвенного органического углерода; и (3) оценку механизмов, определяющих, как изменяется запас SOC в зависимости от свойств почвы и микробных сообществ под влиянием различных методов управления.
Материалы и методы
Район исследования
Данное исследование было проведено в восьми садах, где доминирующей плодовой культурой является манго (Mangifera indica). Эти сады расположены на аллювиальной пойме реки в районе Люгуй (23°00′ с.ш.; 120°39′ в.д.) в бассейне реки Лаонун, на юге Тайваня. Климат в районе исследования — тропический, со среднегодовым количеством осадков более 2000 мм и среднегодовой температурой 25°C. Почвы в исследуемых садах относятся к одному и тому же типу почв, который характеризуется слабокислой реакцией и низким плодородием. Почва может быть классифицирована как Typic Haplustept согласно Таксономии почв (Soil Survey Staff, 2016). Эта почва (Inceptisol) характеризуется супесчаным гранулометрическим составом, малой глубиной и слабой степенью выветрелости (Рисунок 1).
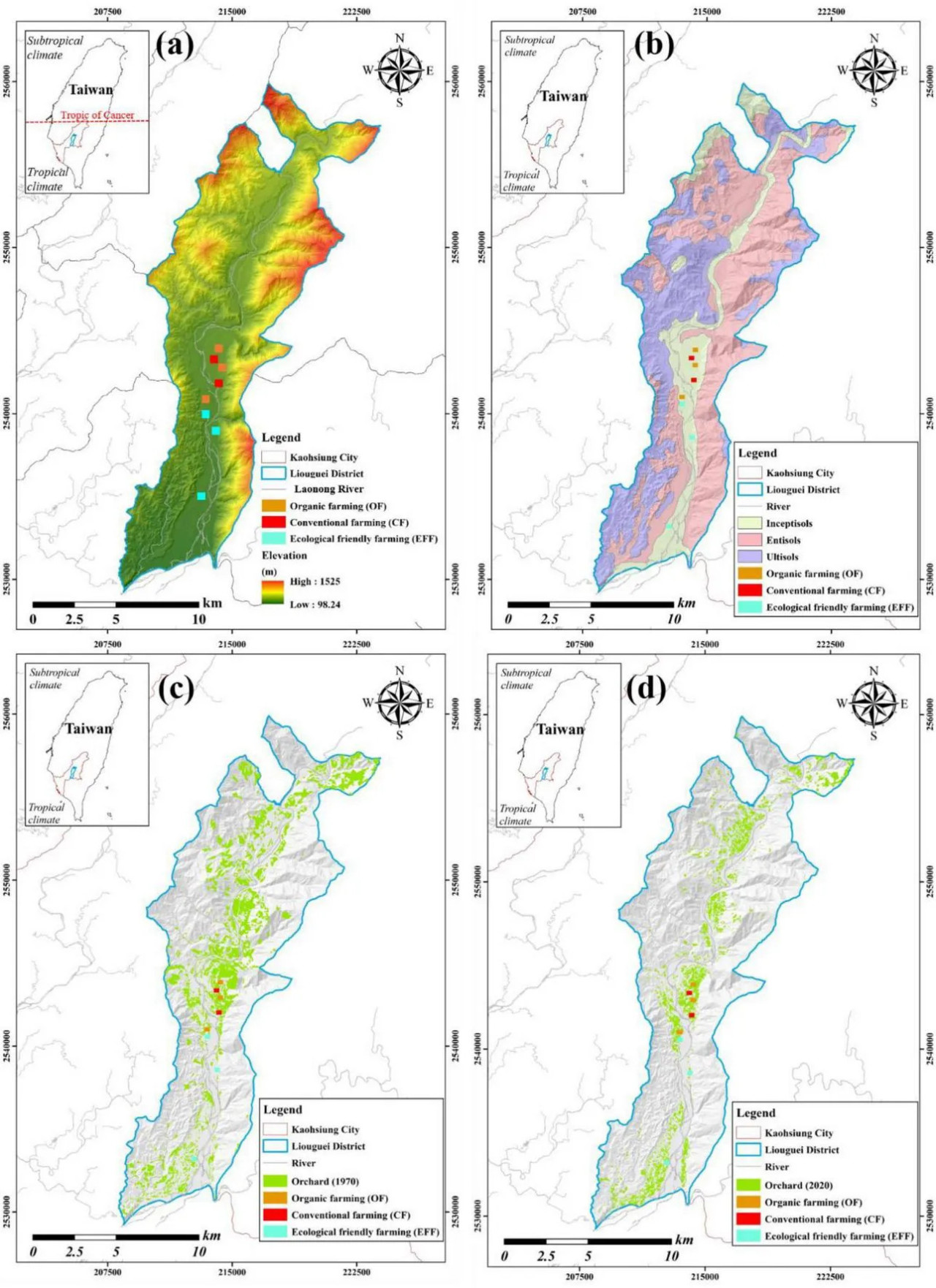
РИСУНОК 1 Участки исследования (сады) при различных методах сельскохозяйственного управления: (a) высота над уровнем моря и распределение исследуемых садов; (b) распределение почвенных таксонов в районе Люгуй; (c) землепользование (только сады показаны зелёным цветом) в 1970 г.; и (d) землепользование (только сады показаны зелёным цветом) в 2020 г.
С 1970 года на всех восьми исследуемых садах в данном исследовании последовательно применялась система традиционного земледелия (CF) (Рисунок 1). Однако три из этих садов были переведены на органическое земледелие (OF) с 2010 года, а ещё три — на экологически безопасное земледелие (EFF) с 2017 года. В садах при системе CF фермеры вносили два вида коммерческих химических удобрений. Одно удобрение (норма внесения 2 т/га/год) содержало 26% общего азота (TN), 13% оксида фосфора (P₂O₅) и 13% оксида калия (K₂O), тогда как другое (норма внесения 1,4 т/га/год) содержало 14% TN, 28% P₂O₅ и 14% K₂O. Кроме того, при CF дважды в год вносили гербицид (RS)-2-амино-4-(гидроксиметилфосфинил)масляная кислота (глюфосинат). В задернённых садах при системе OF в качестве органического удобрения вносили компост из соевого шрота (норма внесения 50 т/га/год). В задернённых садах при системе EFF вносили компосты из куриного помёта и соевого шрота (норма внесения 20 и 2 т/га/год соответственно). В системах OF и EFF гербициды не использовались.
Отбор почвенных образцов и анализ
Образцы почвы (около 2 кг) отбирали с глубины 0–15 см, 15–30 см и 30–50 см в каждом саду при различных методах управления в 2020 году. Участки для отбора проб в каждом саду выбирали случайным образом в пятикратной повторности. Образцы почвы отбирали с разных глубин каждого участка, высушивали на воздухе, осторожно измельчали и просеивали через сито с ячейкой 2 мм, после чего использовали для анализа физических и химических свойств.
Значения pH образцов почвы, отобранных в исследуемых садах, определяли в смеси с деионизированной водой [соотношение почва:вода 1:2,5 (об./об.)]. Для измерения pH использовали измеритель Horiba F-74 BW (Rhoades, 1982). Гранулометрический состав почвы определяли с использованием пипеточного метода (Gee et al., 1986). Емкость катионного обмена оценивали с использованием метода ацетата аммония (pH 7,0) (Sumner and Miller, 1996). Обменный калий экстрагировали 1 моль/л раствором ацетата аммония [соотношение 1:10 (об./об.) для образцов почвы; 1:20 (об./об.) для образцов биоугля] и определяли с помощью атомно-абсорбционной спектрометрии (Z-2300; Hitachi, Япония). Содержание SOC измеряли с использованием метода мокрого окисления (Nelson and Sommers, 1996). Содержание доступного фосфора измеряли с использованием теста Брея P-1 (Bray and Kurtz, 1945). Содержание аммонийного (NH₄⁺-N) и нитратного (NO₃⁻-N) азота измеряли методом паровой дистилляции с использованием оксида магния и сплава Деварда (Mulvaney, 1996).
Содержание запаса SOC рассчитывали следующим образом:
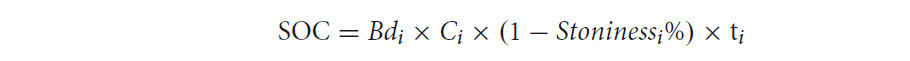
где Bd_i — плотность сложения почвы (мг/м³) в слое i, C_i — содержание углерода в почве (мг углерода на г почвы) в слое i, Stoniness_i% — процентное содержание обломков горных пород в почве с глубины > 2 мм, а t_i — толщина слоя i (глубина = 0–30 см). Процентное содержание каменистости оценивали в полевых условиях. В данном исследовании в каждом исследуемом саду с различными методами управления отбирали пять повторностей (около 200 г почвы на каждую повторность). Запас SOC (кг/м²) на глубинах 0–15 см, 15–30 см и 30–50 см, а также скорость секвестрации SOC (кг/м²/год) в поверхностном слое почвы (0–30 см) также были рассчитаны в этом исследовании для оценки преимуществ систем OF и EFF по сравнению с CF в отношении запаса SOC.
Моделирование переменных окружающей среды для прогнозирования запаса SOC
В этом исследовании использовались осадки, температура воздуха, высота над уровнем моря, крутизна склона, вегетационный индекс, содержание глины, плотность сложения, фактор эрозионной опасности осадков (Rm) и фактор эродируемости почвы (Km) в качестве предикторов SOC в выбранном районе исследования (район Люгуй) на юге Тайваня. Метеорологические данные, включая осадки и температуру воздуха за период 2010–2022 гг., были получены от Центрального метеорологического бюро Тайваня. Данные о высоте над уровнем моря и крутизне склона, полученные из цифровой модели рельефа (DEM), были загружены с платформы открытых данных правительства Тайваня. Нормализованный разностный вегетационный индекс (NDVI) был рассчитан по данным LANDSAT 8, полученным от USGS. Свойства почвы, такие как содержание глины и плотность сложения, были получены из базы данных почвенных обследований Тайваньского научно-исследовательского института сельского хозяйства (TARI). Карты Rm и Km были загружены из информационной системы по склоновым землям Бюро охраны почв и вод Тайваня.
Все модели были разработаны с использованием регрессии Cubist и выполнены с использованием статистического программного обеспечения R-4.3.1 для Windows с пакетом Cubist (Версия: 0.0.18) (Kuhn et al., 2023). Cubist — это мощный инструмент интеллектуального анализа данных для создания прогностических моделей на основе правил. Как правило, регрессионная модель Cubist обладает хорошей прогностической способностью и при этом легка для понимания и интерпретации (Minasny and McBratney, 2008). Этот инструмент интеллектуального анализа данных также может применять схему, подобную бустингу, называемую комитетами, где последовательно создаются итеративные деревья решений. Подробности процедуры бустинга можно найти в работе Quinlan (1992). Кроме того, модель Cubist также предоставляет информацию об использовании атрибутов (относительной важности) каждой переменной, что указывает на важность переменной в модели.
В R 4.0.5 для разделения набора данных на 70% обучающую выборку (калибровочную выборку) и оставшиеся 30% валидационной выборки использовался метод случайной выборки. Производительность модели оценивали с использованием среднеквадратичной ошибки (RMSE) и коэффициента детерминации (R²), рассчитанных по валидационной выборке. В этом исследовании набор данных о свойствах почвы был собран в рамках проекта детального почвенного обследования (база данных SSURGO Тайваня), проведённого TARI в период с 2010 по 2015 год; в исследовании использовались данные о 225 образцах верхнего слоя почвы (0–30 см).
Анализ фосфолипидных жирных кислот
Фосфолипидные жирные кислоты (PLFA) экстрагировали и анализировали в соответствии с методом, описанным Frostegård и соавт. (1993). PLFA экстрагировали с использованием однофазной смеси хлороформ–метанол–цитрат (1:2:0,8). Метиловые эфиры жирных кислот анализировали методом капиллярной газовой хроматографии и детектировали с помощью пламенно-ионизационного детектора на хроматографической системе Thermo Finnigan Trace, как описано Koza и Rezanka (1989). Номенклатура жирных кислот выполнялась в соответствии с описанием Frostegård и соавт. (1993). Суммарное содержание PLFA указывало на общую микробную биомассу. Следующие PLFA рассматривались как бактериальные: i15:0, a15:0, 15:0, i16:0, 16:1ω7c, 17:0, i17:0, cy17:0, 18:1ω7c и cy19:0. PLFA 16:1ω7c, cy17:0, 18:1ω7c и cy19:0 представляли грамотрицательные бактерии, тогда как PLFA i15:0, a15:0, i16:0 и i17:0 представляли грамположительные бактерии. PLFA 18:2ω6,9c, 16:1ω5c и 10Me18:0 представляли обычные грибы, везикулярно-арбускулярные микоризы и актиномицеты соответственно (Zogg et al., 1997; Zelles, 1999).
Выделение ДНК, ПЦР-амплификация и секвенирование ампликонов
Тотальную геномную ДНК экстрагировали из 0,25 г образцов почвы с использованием набора DNeasy PowerSoil Pro Kit (Qiagen, Хильден, Германия) в соответствии с инструкциями производителя. Очищенную ДНК количественно определяли с использованием набора Qubit dsDNA HS assay (ThermoFisher Scientific, Сан-Хосе, Калифорния, США), а качество оценивали с использованием системы Agilent 4200 TapeStation (Agilent Technologies, Пало-Альто, Калифорния, США). Для создания библиотеки ампликонов полноразмерной 16S рДНК использовали набор LoopSeq™ 16S Long Read Kit (Loop Genomics, Сан-Хосе, Калифорния, США). В частности, участки V1–V9 амплифицировали с использованием специфических праймеров (27F: 5′-AGAGTTTGATCMTGGCTCAG-3′, 1492R: 5′-TACCTTGTTACGACTT-3′) и секвенировали на платформе Illumina NovaSeq (Illumina, Сан-Диего, Калифорния, США) в режиме спаренных концов длиной 150 п.н. Облачная платформа, поддерживаемая Loop Genomics, обрабатывала короткие риды для сборки высококачественных консенсусных контигов. Следуя подходу Callahan и соавт. (2021), их дополнительно обрабатывали с использованием DADA2 (Callahan et al., 2016), где проводили обнаружение и коррекцию ошибок ампликонов, фильтрацию потенциальных ошибок оснований и удаление химерных последовательностей для получения вариантов последовательностей ампликонов (ASV). Подготовка образцов, конструирование библиотек, секвенирование и анализ данных были выполнены компанией Welgene Biotech Co., Ltd. (Тайвань, Китайская Республика). Индексы Шеннона и Симпсона рассчитывали с использованием пакета vegan в программе R v.4.3.0.
Анализ данных секвенирования
Классификацию таксонов проводили с использованием алгоритма наивного байесовского классификатора RDP (Wang et al., 2007) с минимальным порогом бутстреп-поддержки 50 при поиске в последовательностях 16S рибосомальной РНК NCBI (Национальный центр биотехнологической информации) по записям RefSeq BioProject 33175 и 33317 (Haft et al., 2018), дополненным базой данных RDP (Ribosomal Database Project) (Cole et al., 2014), всего было классифицировано 987 таксонов.
Статистический анализ
Данные, полученные из свежих образцов почвы, были скорректированы до сухой массы с учётом влажности почвы. Для сравнения между методами мы провели однофакторный дисперсионный анализ и множественный диапазонный тест Дункана. С использованием Canoco (версия 5.0) для Windows мы выполнили анализ избыточности (RDA) для оценки того, связаны ли микробные сообщества и ферментативная активность почвы со следующими факторами окружающей среды, оценёнными в нашем предыдущем исследовании (Lin et al., 2019): микробный биомассы углерод, микробный биомассы азот, SOC и TN. Статистические анализы, если не указано иное, выполнялись с использованием SPSS (версия 18.0; SPSS Inc., Чикаго, Иллинойс, США). Значение p < 0,05 считалось статистически значимым. RDA является многомерным (многооткликовым) аналогом регрессионного анализа.
Моделирование путевым анализом методом частичных наименьших квадратов (PLS-PM) было выполнено для выявления путей, через которые исследуемые переменные, включая различные методы управления, эрозию почвы, обильность бактерий, фосфор и азот, регулируют содержание запаса углерода в почве при различных методах сельскохозяйственного управления. Модель PLS-PM была построена с использованием функции «innerplot» пакета «plspm». Качество и производительность модели оценивали с помощью теста на соответствие. Пакет «ggplot2» в R использовался для построения RDA и создания графиков (Villanueva and Chen, 2019).
Результаты
Свойства почвы и запас почвенного углерода
Образцы почвы, отобранные в садах при системах OF и EFF, имели более низкое pH по сравнению с CK (Таблица 1). SOC и соотношение углерода к азоту были значительно выше в почве OF, чем в других почвах, что указывает на высокое поступление органического углерода в почву OF. Содержание TN было ниже в почве CF, чем в почвах OF и EFF. Однако значительной разницы между почвами OF и EFF не наблюдалось. Содержание общего и доступного фосфора было значительно выше в почве CF, чем в других почвах. В отличие от CF, системы OF и EFF приводили к низкому pH почвы из-за высокого содержания органического вещества. Микробный биомассы углерод отражает микробную активность почвы.
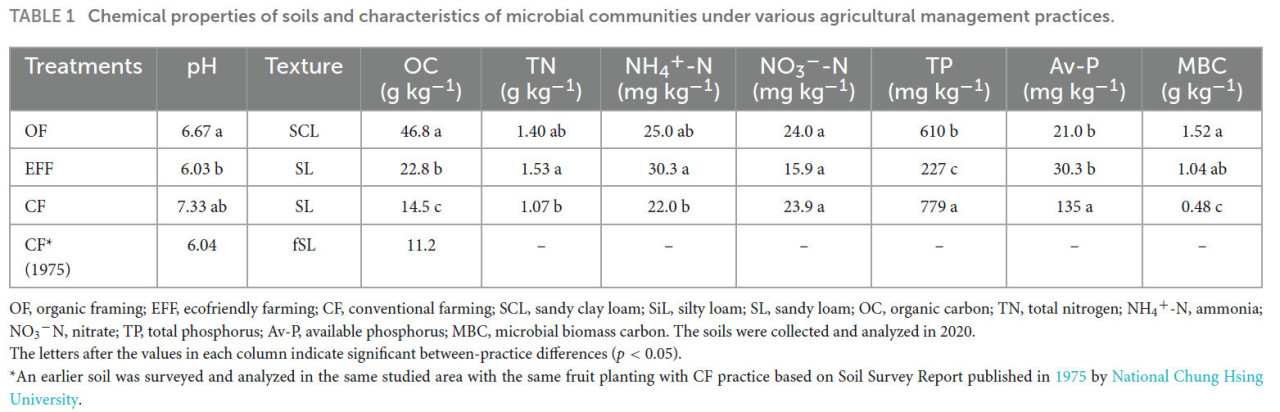
ТАБЛИЦА 1 Химические свойства почв и характеристики микробных сообществ при различных методах сельскохозяйственного управления.
OF — органическое земледелие; EFF — экологически безопасное земледелие; CF — традиционное земледелие; SCL — суглинисто-глинисто-песчаная; SiL — илисто-суглинистая; SL — супесчаная; OC — органический углерод; TN — общий азот; NH₄⁺-N — аммонийный азот; NO₃⁻-N — нитратный азот; TP — общий фосфор; Av-P — доступный фосфор; MBC — микробный биомассы углерод. Почвы были отобраны и проанализированы в 2020 году. Буквы после значений в каждом столбце указывают на значимые различия между методами (p < 0,05).
Более ранняя почва была обследована и проанализирована в том же районе исследования с теми же плодовыми культурами и практикой CF на основе отчёта о почвенном обследовании, опубликованного в 1975 году Национальным университетом Чжунсин.
Наши результаты показывают, что системы OF и EFF приводили к более высокому содержанию микробного биомассы углерода, чем CF. Что касается питательного статуса, содержание TN и NH₄⁺-N было выше в почве EFF, чем в других почвах, что свидетельствует о более высоком содержании органического азота в почве EFF. Содержание общего фосфора было выше в почве CF, чем в других почвах, возможно, из-за долгосрочного использования фермерами химических удобрений. Почва OF имела самый высокий запас SOC. Наши результаты показывают, что различные методы сельскохозяйственного управления оказывают различное влияние на секвестрацию углерода в почве и оценку качества почвы в высокопродуктивных плодоводческих районах, расположенных в мелководных горных регионах с тропическим климатом. Мы сравнили запас почвенного углерода и качество органического вещества между методами сельскохозяйственного управления. Внедрение диверсифицированных систем земледелия (агролесоводства), таких как OF и EFF, эффективно увеличило содержание запаса SOC. OF увеличило содержание запаса SOC примерно на 1,35 кг/м²/30 см и 1,65 кг/м²/50 см в год, тогда как EFF увеличило его примерно на 2,45 кг/м²/30 см и 3,40 кг/м²/50 см в год (Рисунок 2A). Результаты показали, что EFF значительно способствовало секвестрации углерода в почве, что привело к чистому увеличению запаса почвенного углерода на 6,0 кг/м² в год, что в 2,8 раза выше значения, полученного для почвы CF.
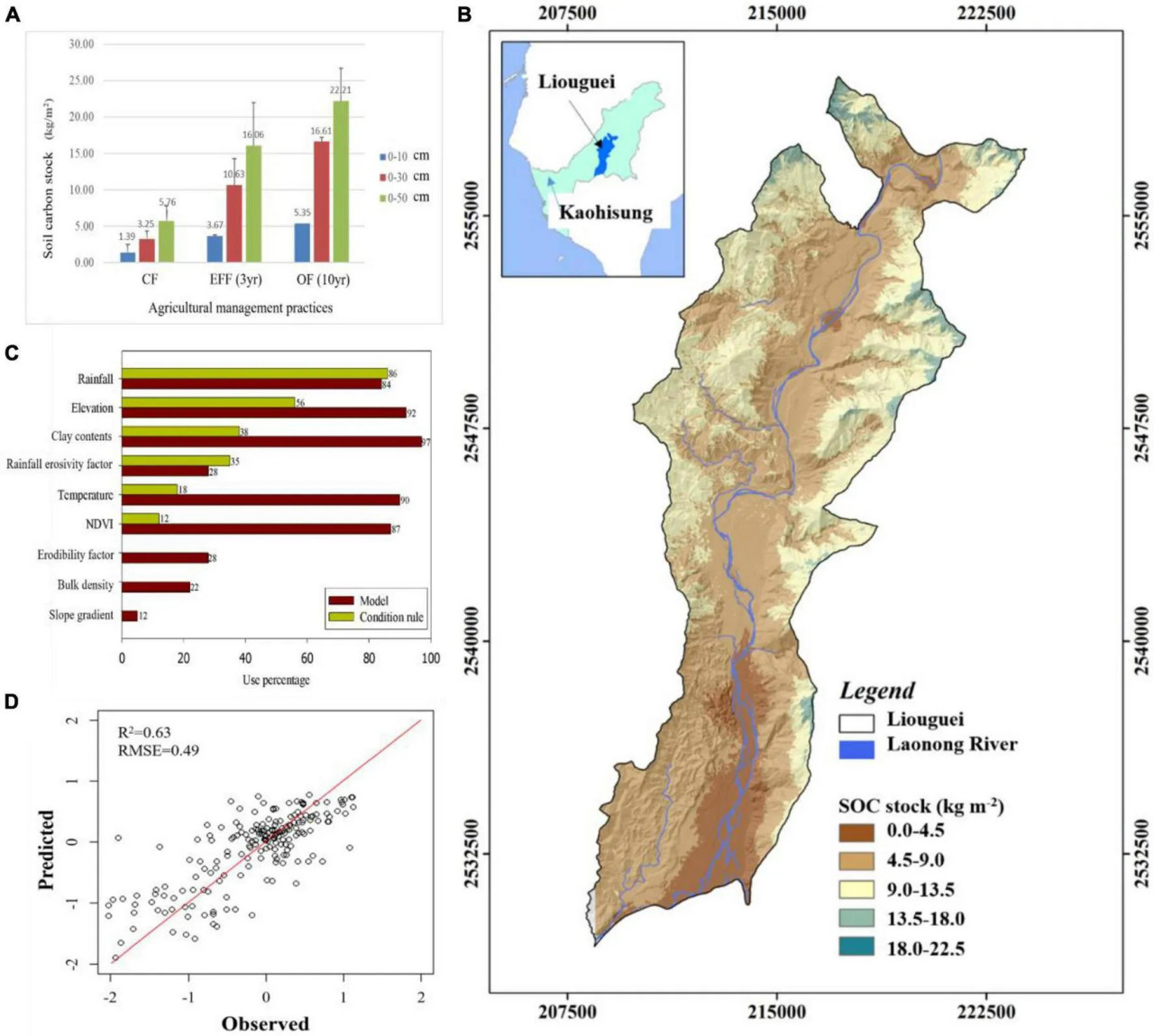
РИСУНОК 2 (A) Запас углерода (кг/м²) в образцах почвы, отобранных с разных глубин в садах при различных методах сельскохозяйственного управления; (B) относительная важность выбранных переменных окружающей среды; (C) измеренный запас SOC (кг/м²) в сравнении с прогнозируемым запасом SOC (кг/м²) по модели CUBIST (чёрная сплошная линия представляет линию 1:1, а синие точки — наборы данных для валидации модели); (D) прогнозируемое пространственное распределение запаса SOC в районе Люгуй в данном исследовании. CF — традиционное земледелие; EFF — экологически безопасное земледелие; OF — органическое земледелие.
Для дальнейшего понимания влияния типов землепользования и методов сельскохозяйственного управления на распределение запасов почвенного органического углерода в этом исследовании использовался подход множественной линейной регрессии на основе машинного обучения для прогнозирования и моделирования пространственного распределения запасов почвенного органического углерода с использованием переменных окружающей среды (Рисунки 2B–D). Наше моделирование показало хорошие результаты моделирования (r² = 0,63, RMSE = 0,49) для распределения запаса SOC, при этом осадки, высота над уровнем моря, содержание глины, Rm, температура и NDVI были наилучшими переменными. Результаты могут способствовать разработке оптимальных стратегий сельскохозяйственного управления для регионов с серьёзной деградацией почв на основе прогнозных результатов.
Биомаркеры PLFA
Фосфолипидные жирные кислоты служат индикаторами активной живой биомассы и отражают влияние методов управления. Общее содержание PLFA было наиболее высоким в микроорганизмах, собранных из почвы OF (Таблица 2). Содержание биомаркеров PLFA было значительно выше в почве OF, чем в других почвах. Аналогично закономерностям, наблюдаемым для общего содержания PLFA, содержание биомаркеров как грибов, так и везикулярно-арбускулярных микориз было наиболее высоким в почве OF и наиболее низким в почве EFF. Содержание биомаркеров PLFA для грамположительных и грамотрицательных бактерий было наиболее высоким в почве OF. Соотношение биомаркеров грамположительных к грамотрицательным бактериям значительно снижалось от почвы OF к почве CF. Значительных различий между методами в соотношении грибов к бактериям не наблюдалось.
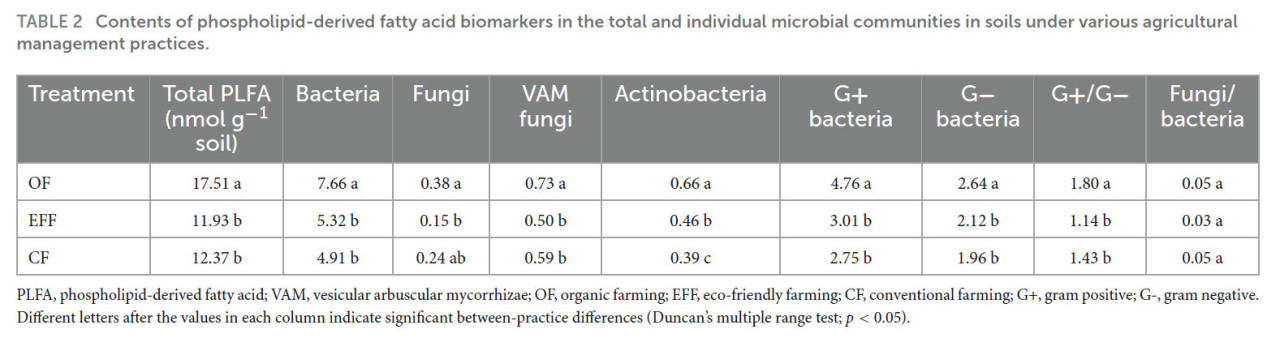
ТАБЛИЦА 2 Содержание биомаркеров фосфолипидных жирных кислот в общих и отдельных микробных сообществах в почвах при различных методах сельскохозяйственного управления.
PLFA — фосфолипидная жирная кислота; VAM — везикулярно-арбускулярные микоризы; OF — органическое земледелие; EFF — экологически безопасное земледелие; CF — традиционное земледелие; G+ — грамположительные; G- — грамотрицательные. Различные буквы после значений в каждом столбце указывают на значимые различия между методами (множественный диапазонный тест Дункана; p < 0,05).
Структура микробного сообщества почвы
В почвах при трёх методах сельскохозяйственного управления относительная обильность Acidobacteria (14,5–23,4%), Actinobacteria (11,0–21,2%) и Proteobacteria (25,9–35,2%) была наиболее высокой среди бактериальных сообществ (Рисунок 3). Однако относительная обильность других типов, таких как Chloroflexi, Firmicutes и Planctomycetes, была низкой, составляя < 9,0% от всех бактериальных сообществ (Рисунок 3). В группе Proteobacteria относительная обильность Alphaproteobacteria (примерно 14,9%) и Deltaproteobacteria (примерно 6,2%) была наиболее высокой, тогда как Betaproteobacteria и Gammaproteobacteria были наиболее низкими (каждая составляла < 5,9% от всех бактериальных сообществ; Рисунок 3).
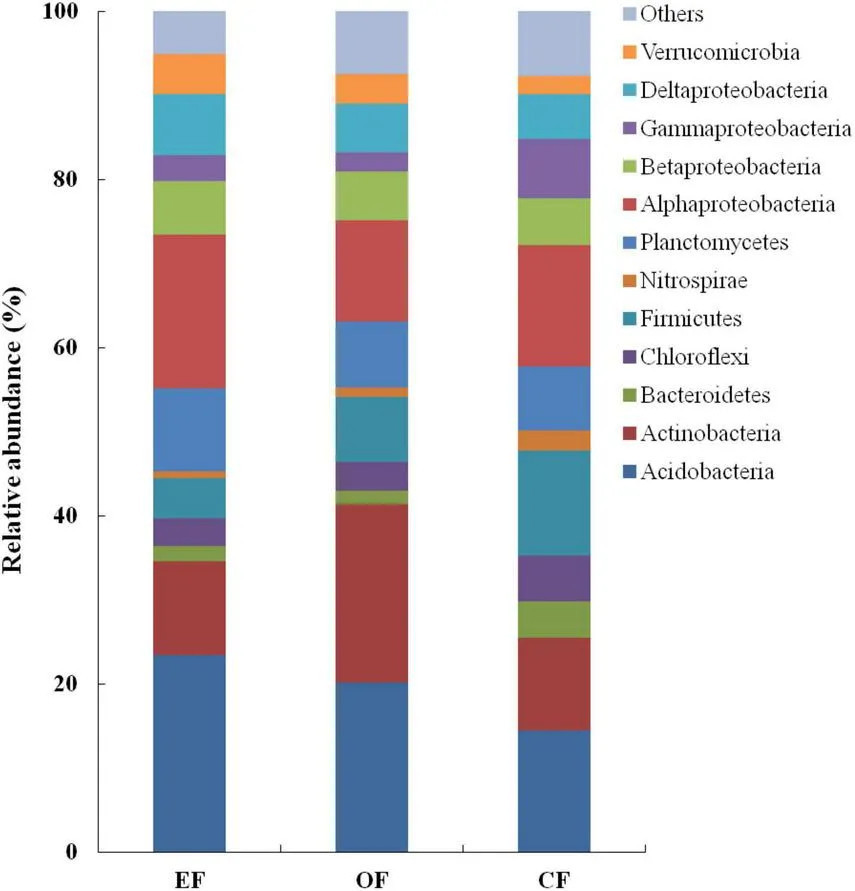
РИСУНОК 3 Относительная обильность бактериальных сообществ при различных методах сельскохозяйственного управления. CF — традиционное земледелие; OF — органическое земледелие; EF — экологически безопасное земледелие.
Несмотря на сходную обильность типов бактерий, мы отметили различия между тремя методами сельскохозяйственного управления в относительной обильности бактерий. В почве OF Actinobacteria составляли примерно 21,0% от всех бактериальных сообществ. Однако в почвах EFF и CF этот род бактерий составлял лишь примерно 11,0% от всех бактериальных сообществ. Proteobacteria составляли 35,0% от всех бактериальных сообществ в почве EFF, но только 26,0% от всех бактериальных сообществ в почве OF. В группе Proteobacteria относительная обильность Alphaproteobacteria, в частности представителей порядка Rhizobiales, была выше в почве EFF, чем в почвах OF и CF. Firmicutes составляли 12,5% от всех бактериальных сообществ в почве CF, но менее 8% от всех бактериальных сообществ в почвах EFF и OF.
Связь между микробными сообществами и свойствами почвы
Мы оценили связь между почвенными микробными сообществами и факторами окружающей среды. Анализ RDA микробных сообществ и факторов окружающей среды показал определённые закономерности (Рисунок 4). Содержание SOC, общего фосфора и TN положительно коррелировало с относительной обильностью почвенных микробных сообществ, но содержание доступного фосфора отрицательно коррелировало с этой обильностью. Таким образом, различные физико-химические свойства почвы и содержание SOC, азота и доступного фосфора влияли на развитие бактериальных и грибных сообществ в почвах при различных методах сельскохозяйственного управления.
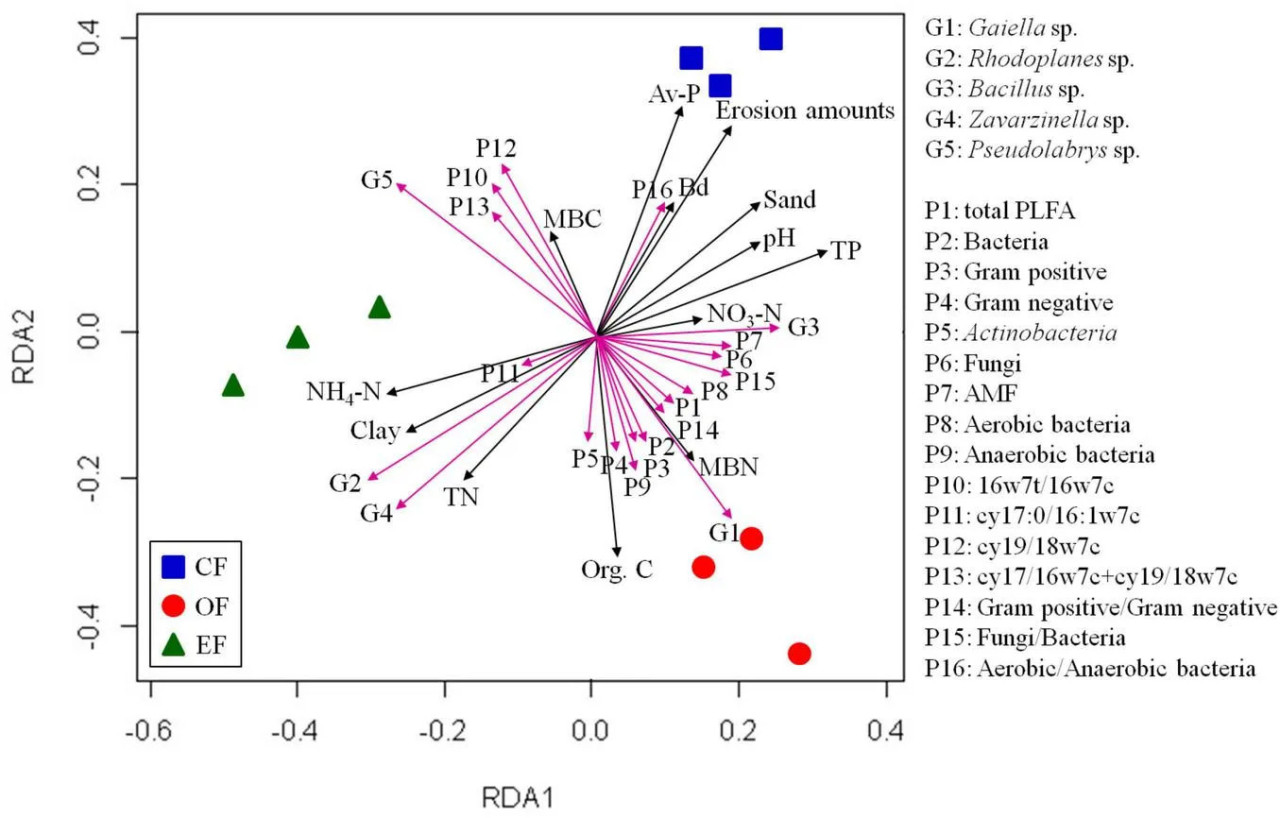
РИСУНОК 4 Результаты анализа избыточности свойств почвы и микробных сообществ при различных методах сельскохозяйственного управления. CF — традиционное земледелие; OF — органическое земледелие; EF — экологически безопасное земледелие.
RDA был проведён для выявления связей между микробными сообществами, содержанием PLFA и свойствами почвы. На основе результатов в почвах при различных методах сельскохозяйственного управления были идентифицированы три отдельных бактериальных кластера (Рисунок 4). Положительные корреляции наблюдались между микробными сообществами и содержанием органического углерода и микробного биомассы азота в почве OF, между микробными сообществами и содержанием TN и NH₄-N в почве EFF, а также между микробными сообществами и pH и содержанием доступного фосфора в почве CF (Рисунок 4). Кроме того, относительная обильность Gaiella и содержание PLFA положительно коррелировали с содержанием SOC и микробного биомассы азота, как и суммарное содержание PLFA и обильность грамположительных и грамотрицательных бактерий. Относительная обильность родов Rhodoplanes и Bacillus и содержание PLFA cy17:0/16:1w7c положительно коррелировали с содержанием TN и NH₄-N. Содержание PLFA в аэробных и анаэробных бактериях положительно коррелировало с pH и содержанием доступного фосфора (Рисунок 4).
Связи между методами управления, свойствами почвы, микробным разнообразием и содержанием запаса углерода в почве были исследованы с помощью анализа PLS-PM (Рисунок 5). Период после перехода от CF к OF или EFF был недостаточно длительным, чтобы значительно (p < 0,05) повлиять на все переменные (Рисунки 5A, B). Долгосрочное применение CF на исследуемых участках могло значительно повлиять на определённые переменные (Рисунок 5C). Все три метода управления улучшили запас углерода в почве. Однако прямое влияние (p < 0,05) оказывала только CF. Системы OF и EFF незначительно (p > 0,05) снижали эрозию почвы, что, в свою очередь, увеличивало содержание запаса SOC. Значительная отрицательная корреляция наблюдалась между запасом SOC и эрозией почвы (Рисунок 6). Все три метода управления положительно влияли на обильность бактерий; однако эффекты были незначительными (p > 0,05; Рисунок 5). Методы управления влияли на содержание азота и фосфора либо непосредственно, либо через влияние на обильность бактерий. Как показано на Рисунке 5, системы OF и EFF незначительно увеличивали (p > 0,05) содержание азота и фосфора, тогда как CF значительно (p > 0,05) снижала эти показатели. Что касается стандартизированных общих эффектов (Рисунок 5D), содержание запаса углерода в почвах CF, OF и EFF определялось в основном методами управления; методами управления, микробным разнообразием и азотным циклом; а также циклами азота и фосфора соответственно. Анализ PLS-PM объяснил 78, 72 и 80% общей дисперсии в содержании запаса углерода в почвах OF, EFF и CF соответственно (Рисунок 5).
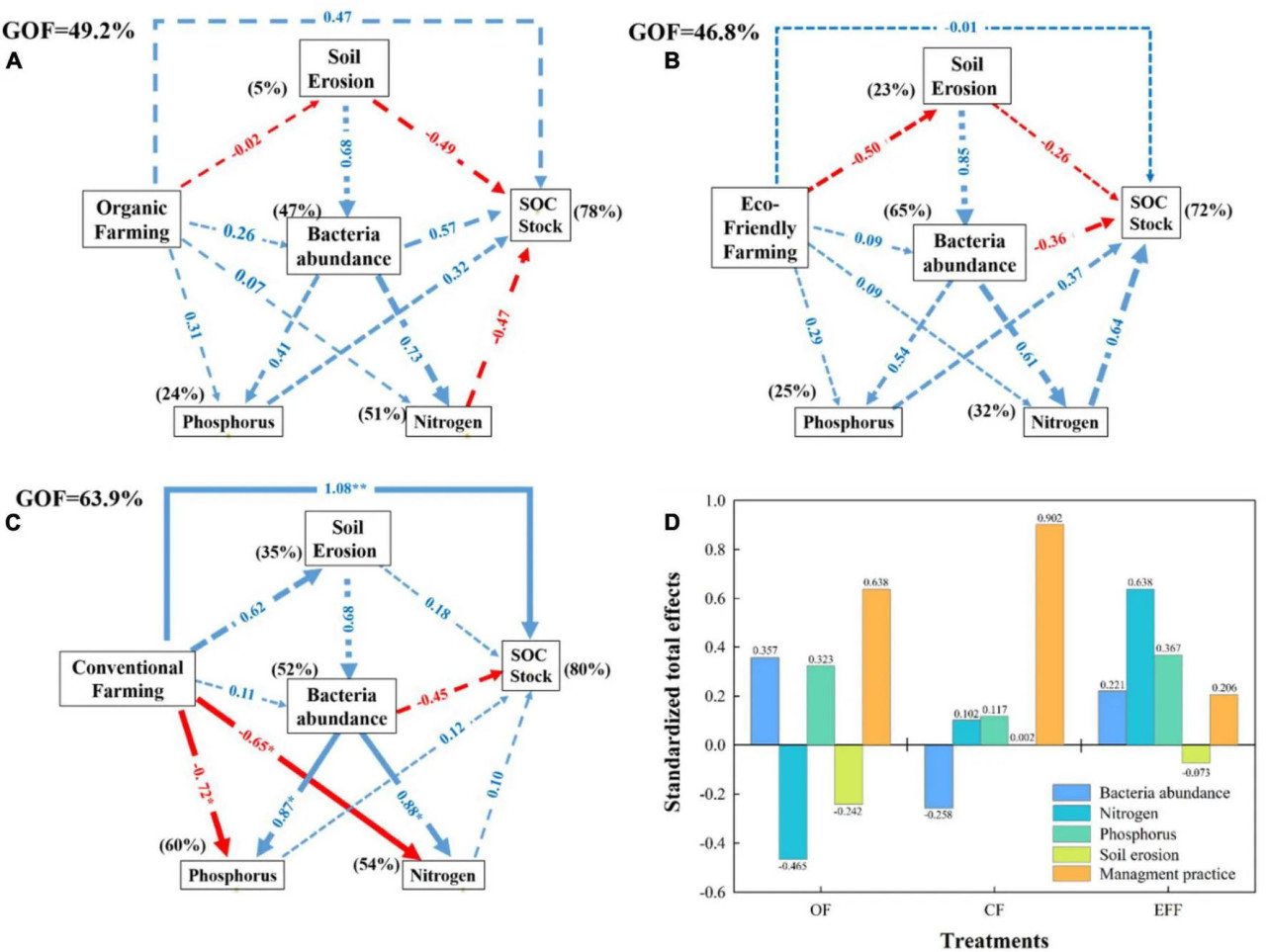
РИСУНОК 5 Анализ методом частичных наименьших квадратов (PLS-PM) взаимосвязей между методами управления: (A) органическое земледелие; (B) экологически безопасное земледелие; (C) традиционное земледелие; (D) стандартизированные общие эффекты для всех методов управления, свойств почвы, обильности бактерий и запаса углерода в почве. Стандартизированные общие эффекты обильности бактерий, азота, фосфора и эрозии почвы на запас углерода в почве. Методы управления включают объёмы внесения C, N и P. Эрозия почвы: Bd, глина, объёмы эрозии. Обильность бактерий: индекс Шеннона, общая PLFA, грибы, аэробные/анаэробные. Фосфор: общий P и доступный P. Азот: общий N, NH₄⁺-N, NO₃⁻-N. Запас SOC: NDVI, содержание SOC. Для оценки путевых коэффициентов было проведено 1000 бутстреп-итераций. Положительные и отрицательные эффекты обозначены синими и красными стрелками соответственно. Путевые коэффициенты, незначимо отличающиеся от нуля, показаны серыми пунктирными линиями; p < 0,05 и p < 0,01. Проценты над рамками представляют объяснительную силу переменных. Для оценки модели использовался показатель соответствия (goodness-of-fit).
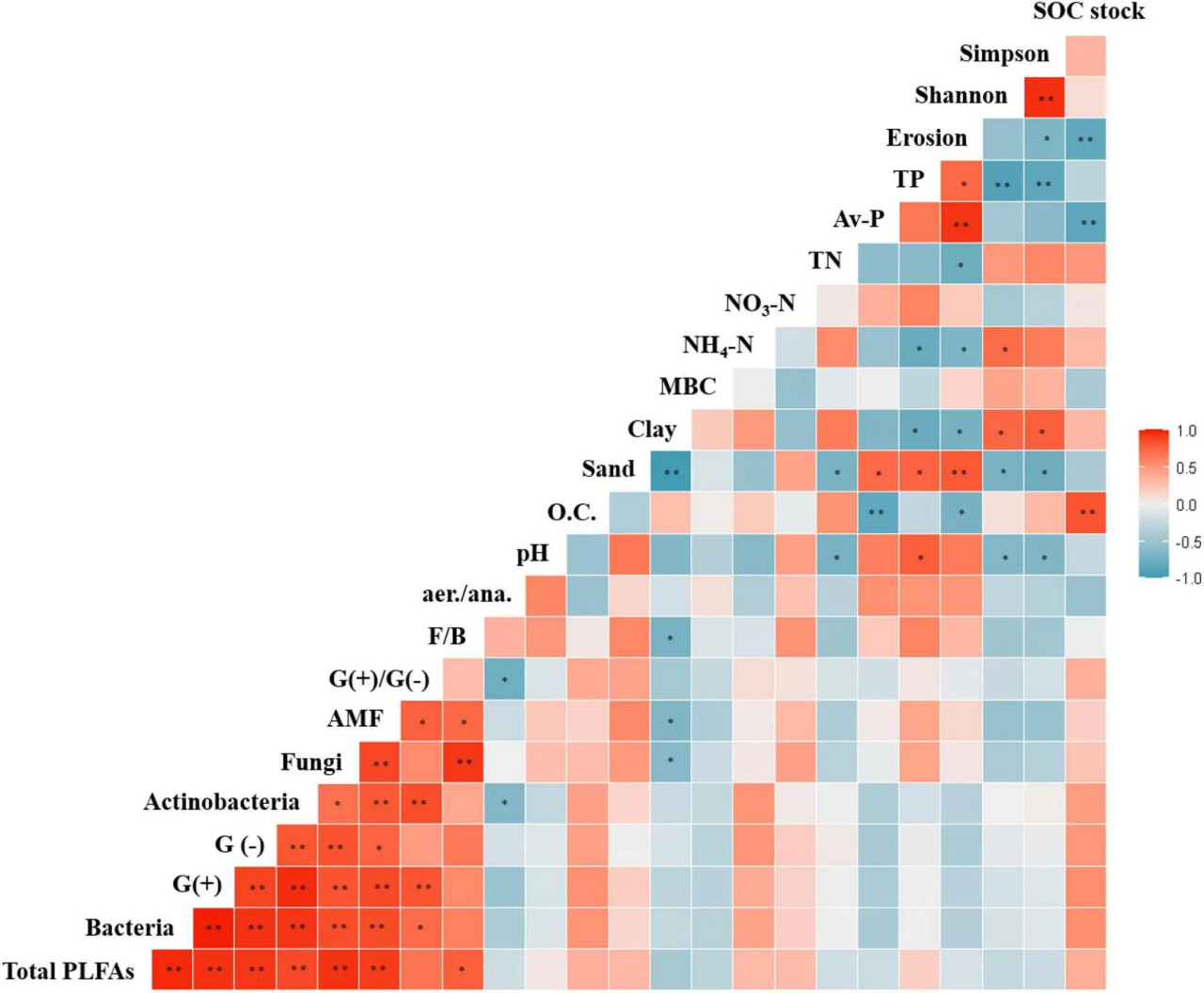
РИСУНОК 6 Корреляции Пирсона между свойствами почвы и методами сельскохозяйственного управления. p < 0,05 и p < 0,01.
Обсуждение
В настоящем исследовании бактериальные сообщества в почвах при трёх методах сельскохозяйственного управления были представлены преимущественно Acidobacteria, Actinobacteria и Proteobacteria. Было показано, что эти типы широко распространены в почвенных экосистемах при различных методах управления (Lupatini et al., 2017; Durrer et al., 2021). Относительная обильность Acidobacteria была ниже в почве CF, чем в почвах EFF и OF. Это может быть связано с отрицательной корреляцией между этим типом и pH почвы (Xiong et al., 2012), что также соответствовало результатам данного исследования (Рисунок 6). Rhizobiales были обильны в почве EFF. Этот порядок класса Alphaproteobacteria включает представителей, способных фиксировать азот, разлагать органические соединения и способствовать росту растений (Yarwood et al., 2009). Таким образом, эти бактерии могут помогать поддерживать почвенные функции и служить индикаторами влияния EFF на бактериальные сообщества почвы. Помимо относительной обильности Acidobacteria и Alphaproteobacteria, обильность Actinobacteria также различалась между тремя методами управления. Бактерии, относящиеся к типу Alphaproteobacteria, способны разлагать устойчивое органическое вещество (Zhou et al., 2016) и обильно присутствуют в почвах при органическом земледелии (Hartman et al., 2018).
Изменения физико-химических свойств почвы в ответ на различные методы сельскохозяйственного управления
В отличие от CF, системы OF и EFF значительно увеличили содержание SOC и TN, что указывает на то, что внесение органических и экологически безопасных удобрений может увеличить содержание SOC и TN. Более высокие объёмы внесения органических удобрений и биомассы трав в системе OF могли привести к более высокому содержанию органического углерода и азота в почве OF. Аналогичным образом, внесение более высокого количества биомассы трав в системе EFF могло увеличить содержание подстилки и гумусового слоя в поверхностной почве EFF, хотя фермеры садов применяли систему EFF с 2018 года. Это могло быть связано с внесением органических удобрений в системе OF или с обеспечением дополнительных питательных веществ в системе EFF, что могло увеличить содержание подстилки, тем самым улучшая плодородие почвы.
В отличие от OF и EFF, система CF предполагала использование гербицидов, которые изменяли структуру микробного сообщества почвы. Gammaproteobacteria, Planctomycetes, Firmicutes и Nitrospirae были обильны в почве CF; это могло быть связано с регулярным использованием фосфорсодержащих гербицидов (Giambò et al., 2021) и химических удобрений. Некоторые представители класса Gammaproteobacteria являются патогенами растений и животных; долгосрочное применение гербицидов в почве может способствовать росту фитопатогенов (Pileggi et al., 2020). Долгосрочный дефицит SOC может приводить к усилению роста Planctomycetes и Firmicutes, которые способны разлагать свежее органическое вещество и таким образом снижать стресс от дефицита питательных веществ (Liu et al., 2023). Влияние сублетальных доз фосфорсодержащего гербицида на микробиоту медоносных пчёл было описано Giambò и соавт. (2021); воздействие гербицида снижало обильность Planctomycetes и Firmicutes.
Изменения структуры микробного сообщества почвы в ответ на различные методы сельскохозяйственного управления
Растения могут регулировать содержание микробных PLFA в почве (Priha et al., 1999). Было показано, что изменения растительности, а не прямые изменения свойств почвы, изменяют структуру микробного сообщества почвы (Zornoza et al., 2009; Wang et al., 2022). Внесение гуано в почвы при системе OF, которое приводит к увеличению содержания азота, может повышать продуктивность трав и, таким образом, способствовать росту почвенных микроорганизмов (Wang et al., 2022).
Сообщалось, что суммарное содержание PLFA и относительная обильность бактериальных сообществ положительно коррелируют с содержанием SOC и TN (Shahbaz et al., 2020). Мы наблюдали высокое суммарное содержание PLFA и высокую относительную обильность грамположительных и грамотрицательных бактерий в почве OF; это может быть связано с высоким содержанием SOC и TN в почве OF. Хотя EFF увеличивало содержание SOC и TN, значительных различий в обильности почвенных микробных сообществ между EFF и CF не наблюдалось; это может быть связано с тем, что EFF не предполагало внесения дополнительного субстрата, что приводило к более низкой микробной биомассе в почве EFF по сравнению с почвой OF. Некоторые исследования показали, что грамотрицательные бактерии процветают в условиях богатого субстратами окружения; однако в условиях ограниченности ресурсов медленно растущие грамположительные бактерии были более обильны, чем грамотрицательные (Djukic et al., 2010; Fanin et al., 2019). Хотя в системе EFF не добавлялся дополнительный SOC, соотношение грамположительных к грамотрицательным бактериям было наиболее низким в почве EFF, что свидетельствует о том, что эта почва способствовала росту грамотрицательных бактерий.
Обильность грибов и везикулярно-арбускулярных микориз была наиболее высокой в почве OF. Значительных различий в соотношении грибов к бактериям между тремя методами не наблюдалось (Таблица 2). Несколько низкое соотношение грибов к бактериям в EFF может указывать на содержание и состав подстилки, поступающей в почву, поскольку грибы были преобладающими деструкторами устойчивых соединений в подстилке (Allison et al., 2005), а бактерии являются эффективными конкурентами за доступность лабильных субстратов (Djukic et al., 2010). Низкое соотношение грибов к бактериям в почве EFF также указывало на то, что экологически безопасные методы сельскохозяйственного управления могут способствовать быстрому круговороту углерода в почве. Быстрорастущие виды растений, особенно с сильно разветвлёнными тонкими корневыми системами, выделяют большие количества экссудатов в почву EFF (Personeni and Loiseau, 2004), что благоприятно для роста бактерий.
В данном исследовании RDA показал, что улучшение качества почвы с помощью методов ведения хозяйства было непосредственно связано с изменениями в структуре микробного сообщества почвы. Кроме того, OF положительно влияло на содержание как микробной биомассы почвы, так и SOC, что свидетельствует о том, что источники углерода и азота, присутствующие в почве OF, значительно увеличивали содержание микробной биомассы. Анализ PLS-PM показал, что тип питательных веществ различался между CF и двумя другими методами сельскохозяйственного управления, что может объяснять значительные различия в относительной обильности бактерий и содержании азота и фосфора между CF и OF или EFF. Долгосрочное применение химических удобрений в системе CF положительно влияло на циклы азота и фосфора, но отрицательно — на микробные характеристики (Рисунок 5). Tan и соавт. (2023) указали, что внесение навоза приводит к формированию более сложной сети совместного присутствия, чем внесение минеральных удобрений (Ling et al., 2016); эти результаты согласуются с нашими выводами (Рисунок 5). Вышеупомянутое явление может быть обусловлено различными питательными характеристиками навоза и минеральных удобрений. Минеральные удобрения высвобождают быстродействующие питательные вещества широкого спектра, что помогает снизить стресс от ограничения питательных веществ и межвидовую конкуренцию, тем самым благоприятствуя многим таксонам грибов (Sun et al., 2020). Напротив, навозы высвобождают питательные вещества узкого спектра, благоприятствуя росту только определённых видов грибов из-за их дифференциального использования углеродных субстратов (Hanson et al., 2000; Rinnan and Bååth, 2009). Таким образом, в отличие от минеральных удобрений, навозы оказывают существенное селективное давление на почвенные грибы.
Заключение
В нашем заключении, бактериальные сообщества тропических склоновых земель зависят от методов управления. Методы управления OF или EFF могут не только эффективно увеличивать микробные сообщества, но и способствовать ферментативной активности. Кроме того, запас почвенного органического углерода заметно увеличился как минимум в 3–5 раз при переходе от CF к EFF или OF. Круговорот питательных веществ также подвергается влиянию увеличения SOC в системах OF или EFF, при этом обильность углерода также означает ослабление углеродного лимитирования и дополнительно приводит к дефициту азота и фосфора в этих почвах. Однако в условиях сценариев изменения климата, включая деградацию почв и продовольственную безопасность, наши результаты предоставляют ценную информацию для демонстрации и сравнения изменений бактериальных сообществ почвы и качества почвы на возделываемых склоновых землях в условиях тропического климата.
Ссылки
1. Allison V. J., Miller R. M., Jastrow J. D., Matamala R., Zak D. R. (2005). Changes in soil microbial community structure in a tallgrass prairie chronosequence. Soil Sci. Soc. Am. J. 69, 1412–1421. doi: 10.2136/sssaj2004.0252. CrossRef. Google Scholar.
2. Amelung W., Bossio D., de Vries W., Kögel-Knabner I., Lehmann J., Amundson R., et al. (2020). Towards a global-scale soil climate mitigation strategy. Nat. Commun. 11:5427. doi: 10.1038/s41467-020-18887-7. CrossRef. Google Scholar.
3. Bender S. F., Wagg C., van der Heijden M. G. A. (2016). An underground revolution: Biodiversity and soil ecological engineering for agricultural sustainability. Trends Ecol. Evol. 31, 440–452. doi: 10.1016/j.tree.2016.02.016. CrossRef. Google Scholar.
4. Bray R. H., Kurtz L. T. (1945). Determination of total organic and available forms of phosphorus in soils. Soil Sci. 59, 39–45. doi: 10.1097/00010694-194501000-00006. CrossRef. Google Scholar.
5. Callahan B. J., Grinevich D., Thakur D., Balamotis M. A., Yehezkel T. B. (2021). Ultra-accurate microbial amplicon sequencing with synthetic long reads. Microbiome 9:130. doi: 10.1186/s40168-021-01072-3. CrossRef. Google Scholar.
6. Callahan B. J., McMurdie P. J., Rosen M. J., Han A. W., Johnson A. J. A., Holmes S. P. (2016). DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 13, 581–583. doi: 10.1038/nmeth.3869. CrossRef. Google Scholar.
7. Cole J. R., Wang Q., Fish J. A., Chai B., McGarrell D. M., Sun Y., et al. (2014). Ribosomal database project: Data and tools for high throughput rRNA analysis. Nucleic acids research 42, 633–642. doi: 10.1093/nar/gkt1244. CrossRef. Google Scholar.
8. Djukic I., Zehetner F., Mentler A., Gerzabek M. H. (2010). Microbial community composition and activity in different Alpine vegetation zones. Soil Biol. Biochem. 42, 155–161. doi: 10.1016/j.soilbio.2009.10.006. CrossRef. Google Scholar.
9. Durrer A., Gumiere T., Rumenos Guidetti Zagatto M., Petry Feiler H., Miranda Silva A. M., Henriques Longaresi R., et al. (2021). Organic farming practices change the soil bacteria community, improving soil quality and maize crop yields. PeerJ. 9:e11985. doi: 10.7717/peerj.11985. CrossRef. Google Scholar.
10. Fanin N., Kardol P., Farrell M., Nilsson M. C., Gundale M. J., Wardle D. A. (2019). The ratio of Gram-positive to Gram-negative bacterial PLFA markers as an indicator of carbon availability in organic soils. Soil Biol. Biochem. 128, 111–114. doi: 10.1016/j.soilbio.2018.10.010. CrossRef. Google Scholar.
11. Frostegård Å., Bååth E., Tunlio A. (1993). Shifts in the structure of soil microbial communities in limed forests as revealed by phospholipid fatty acid analysis. Soil Biol. Biochem. 25, 723–730. doi: 10.1016/0038-0717(93)90113-P. CrossRef. Google Scholar.
12. Gee G. W., Bauder J. W. (1986). "Particle-size analysis," in Methods of soil analysis, part 1. physical and mineralogical methods, agronomy monograph no. 9, 2nd Edn, ed. A. Klute (Madison, WI: American Society of Agronomy, Soil Science Society of America), 383–411.
13. Giambò F., Leone G. M., Gattuso G., Rizzo R., Cosentino A., Cinà D., et al. (2021). Genetic and epigenetic alterations induced by pesticide exposure: Integrated analysis of gene expression, microRNA expression, and DNA methylation datasets. Int. J. Environ. Res. Public Health 18:8697. doi: 10.3390/ijerph18168697. CrossRef. Google Scholar.
14. Gnanavelrajah N., Shrestha R. P., Schmidt-Vogt D., Samarakoon L. (2008). Carbon stock assessment and soil carbon management in agricultural land-uses in Thailand. Land Degrad. Dev. 19, 242–256. doi: 10.1002/ldr.838. CrossRef. Google Scholar.
15. Haft D. H., DiCuccio M., Badretdin A., Brover V., Chetvernin V., O'Neill K., et al. (2018). RefSeq: An update on prokaryotic genome annotation and curation. Nucleic Acids Res. 46, 851–860. doi: 10.1093/nar/gkx1068. CrossRef. Google Scholar.
16. Hanson P. J., Edwards N. T., Garten C. T., Andrew J. A. (2000). Separating root and soil microbial contributions to soil respiration: A review of methods and observations. Biogeochemistry 48, 115–146. doi: 10.1023/A:1006244819642. CrossRef. Google Scholar.
17. Hartman K., van der Heijden M. G. A., Wittwer R. A., Banerjee S., Walser J. C., Schlaeppi K. (2018). Cropping practices manipulate abundance patterns of root and soil microbiome members paving the way to smart farming. Microbiome 6:14. doi: 10.1186/s40168-017-0389-9. CrossRef. Google Scholar.
18. Huang S. N. (1994). Soil management for sustainable food production in Taiwan. Extension Bulletin 390. Taipei: Food and Fertilizer Technology Center.
19. Koza T., Rezanka M. W. T. (1989). Quantitative analysis of fatty acid methyl esters by capillary gas chromatography with flame-ionization detection: Quadrupole and sector mass spectrometer. Folia Microbiol. 34, 165–169. doi: 10.1007/BF02823697. CrossRef. Google Scholar.
20. Kuhn M., Weston S., Keefer C., Kuhn M. M. (2023). Package 'Cubist'. Austria: Innsbruck.
21. Lal R. (2004). Soil carbon sequestration impacts on global climate change and food security. Science 304, 1623–1627. doi: 10.1126/science.1097396. CrossRef. Google Scholar.
22. Lal R. (2018). Digging deeper: A holistic perspective of factors affecting soil organic carbon sequestration in agroecosystems. Glob. Change Biol. 24, 3285–3301. doi: 10.1111/gcb.14054. CrossRef. Google Scholar.
23. Levy-Booth D. J., Prescott C. E., Grayston S. J. (2014). Microbial functional genes involved in nitrogen fixation, nitrification and denitrification in forest ecosystems. Soil Biol. Biochem. 75, 11–25. doi: 10.1016/j.soilbio.2014.03.021. CrossRef. Google Scholar.
24. Lin Y. T., Lin Y. F., Tsai I. J., Chang E. H., Jien S. H., Lin Y. J., et al. (2019). Structure and diversity of soil bacterial communities in Offshore Islands. Sci. Rep. 9:4689. doi: 10.1038/s41598-019-41170-9. CrossRef. Google Scholar.
25. Ling N., Zhu C., Xue C., Chen H., Duan Y. H., Peng C., et al. (2016). Insight into how organic amendments can shape the soil microbiome in long-term field experiments as revealed by network analysis. Soil Biol. Biochem. 99, 137–149. doi: 10.1016/j.soilbio.2016.05.005. CrossRef. Google Scholar.
26. Liu X., Chen Q., Zhang H., Zhang J., Chen Y., Yao F., et al. (2023). Effects of exogenous organic matter addition on agricultural soil microbial communities and relevant enzyme activities in southern China. Sci. Rep. 13:8045. doi: 10.1038/s41598-023-33498-0. CrossRef. Google Scholar.
27. Lupatini M., Korthals G. W., de Hollander M., Janssens T. K. S., Kuramae E. E. (2017). Soil microbiome is more heterogeneous in organic than in conventional farming system. Front. Microbiol. 7:2064. doi: 10.3389/fmicb.2016.02064. CrossRef. Google Scholar.
28. Minasny B., McBratney A. B. (2008). Regression rules as a tool for predicting soil properties from infrared reflectance spectroscopy. Chemometr. Intell. Lab. Syst. 94, 72–79. doi: 10.1016/j.chemolab.2008.06.003. CrossRef. Google Scholar.
29. Mulvaney R. L. (1996). "Nitrogen-inorganic forms," in Methods of soil analysis: Soil science society of america book series 5 Part 3—chemical methods, eds. D. L. Sparks, A. L. Page, P. A. Helmke, and R. H. Loeppert (Madison, WI: ASA and SSSA), 1123–1184. doi: 10.2136/sssabookser5.3.c38. CrossRef. Google Scholar.
30. National Chung Hsing University (1975). Soil survey report in Kaohsiung county. Taichung: National Chung Hsing University.
31. Nelson D. W., Sommers L. E. (1996). "Total carbon, organic carbon, and organic matter," in Methods of soil analysis. part 3-chemical methods, eds. D. L. Sparks, A. L. Page, P. A. Helmke, R. H. Loeppert, P. N. Soltanpour, M. A. Tabatabai, et al. (Madison, WI: Soil Science Society of America. Inc), 961–1010.
32. Peltoniemi K., Velmala S., Fritze H., Lemola R., Pennanen T. (2021). Long-term impacts of organic and conventional farming on the soil microbiome in boreal arable soil. Eur. J. Soil Biol. 104:103314. doi: 10.1016/j.ejsobi.2021.103314. CrossRef. Google Scholar.
33. Pereira e Silva M. C., Semenov A. V., Schmitt H., van Elsas J. D., Salles J. F. (2013). Microbe-mediated processes as indicators to establish the normal operating range of soil functioning. Soil Biol. Biochem. 57, 995–1002. doi: 10.1016/j.soilbio.2012.10.002. CrossRef. Google Scholar.
34. Personeni E., Loiseau P. (2004). How does the nature of living and dead roots affect the residence time of carbon in the root litter continuum? Plant Soil 267, 129–141. doi: 10.1007/s11104-005-4656-3. CrossRef. Google Scholar.
35. Pileggi M., Pileggi S. A. V., Sadowsky M. J. (2020). Herbicide bioremediation: From strains to bacterial communities. Heliyon 6:e05767. doi: 10.1016/j.heliyon.2020.e05767. CrossRef. Google Scholar.
36. Priha O., Grayston S. J., Pennanen T., Smolander A. (1999). Microbial activities related to C and N cycling and microbial community structure in the rhizospheres of Pinus sylvestris, Picea abies and Betula pendula seedlings in an organic and mineral soil. FEMS Microbiol. Ecol. 30, 187–199. doi: 10.1111/j.1574-6941.1999.tb00647.x. CrossRef. Google Scholar.
37. Quinlan J. R. (1992). "Learning with continuous classes," in Proceedings of the 5th Australian joint conference on artificial intelligence, (Singapore).
38. Rhoades J. (1982). "Cation exchange capacity," in Methods of soil analysis. part 2. Chemical and microbiological properties, eds. A. L. Page, R. H. Miller, and D. R. Keeney (Madison, WI: American Society of Agronomy, Inc, Soil Science Society of America. Inc), 149–157.
39. Rinnan R., Bååth E. (2009). Differential utilization of carbon substrates by bacteria and fungi in tundra soil. Appl. Environ. Microbiol. 75, 3611–3620. doi: 10.1128/AEM.02865-08. CrossRef. Google Scholar.
40. Shahbaz M., Kätterer T., Thornton B., Börjesson G. (2020). Dynamics of fungal and bacterial groups and their carbon sources during the growing season of maize in a long-term experiment. Biol. Fertil. Soils 56, 759–770. doi: 10.1007/s00374-020-01454-z. CrossRef. Google Scholar.
41. Soil Survey Staff (2016). Natural Resources Conservation Service, United States Department of Agriculture. Web Soil Survey. Washington, D.C: Natural Resources Conservation Service.
42. Sumner M. E., Miller W. P. (1996). "Cation exchange capacity and exchange coefficients," in Methods of soil analysis part 3: Chemical methods, SSSA book series 5, ed. D. L. Sparks (Madison, WI: Soil Science Society of America), 1201–1230.
43. Sun X., Kosman E., Sharon O., Ezrati S., Sharon A. (2020). Significant host- and environment-dependent differentiation among highly sporadic fungal endophyte communities in cereal crops-related wild grasses. Environ. Microbiol. 22, 3357–3374. doi: 10.1111/1462-2920.15107. CrossRef. Google Scholar.
44. Tan Y., Wang J., He Y., Yu X., Chen S., Penttinen P., et al. (2023). Organic fertilizers shape soil microbial communities and increase soil amino acid metabolites content in a blueberry orchard. Soil Microbiol. 85, 232–246. doi: 10.1007/s00248-022-01960-7. CrossRef. Google Scholar.
45. Van Wesemael B., Paustian K., Meersmans J., Goidts E., Barancikova G., Easter M., et al. (2010). Agricultural management explains historic changes in regional soil carbon stocks. Proc. Natl. Acad. Sci. U.S.A. 107, 14926–14930. doi: 10.1073/pnas.1002592107. CrossRef. Google Scholar.
46. Villanueva R. A. M., Chen Z. J. (2019). ggplot2: Elegant graphics for data analysis (2nd ed.). Meas. Interdiscipl. Res. Perspect. 17, 160–167. doi: 10.1080/15366367.2019.1565254. CrossRef. Google Scholar.
47. Wang Q., Garrity G. M., Tiedje J. M., Cole J. R. (2007). Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 73, 5261–5267. doi: 10.1128/AEM.00062-07. CrossRef. Google Scholar.
48. Wang S., Mori T., Zou S., Zheng H., Heděnec P., Zhu Y., et al. (2022). Changes in vegetation types affect soil microbial communities in tropical islands of southern China. Glob. Ecol. Conserv. 37:e02162. doi: 10.1016/j.gecco.2022.e02162. CrossRef. Google Scholar.
49. Xiong J., Liu Y., Lin X., Zhang H., Zeng J., Hou J., et al. (2012). Geographic distance and pH drive bacterial distribution in alkaline lake sediments across Tibetan Plateau. Environ. Microbiol. 14, 2457–2466. doi: 10.1111/j.1462-2920.2012.02799.x. CrossRef. Google Scholar.
50. Yarwood S. A., Myrold D. D., Högberg M. N. (2009). Termination of belowground C allocation by trees alters soil fungal and bacterial communities in a boreal forest. FEMS Microbiol. Ecol. 70, 151–162. doi: 10.1111/j.1574-6941.2009.00733.x. CrossRef. Google Scholar.
51. Zelles L. (1999). Fatty acid patterns of phospholipids and lipopolysaccharides in the characterisation of microbial communities in soil: A review. Biol. Fertil. Soils 29, 111–129. doi: 10.1007/s003740050533. CrossRef. Google Scholar.
52. Zhang H., Zheng X., Bai N., Li S., Zhang J., Lv W. (2019). Responses of soil bacterial and fungal communities to organic and conventional farming systems in east China. J. Microbiol. Biotechnol. 29, 441–453. doi: 10.4014/jmb.1809.09007. CrossRef. Google Scholar.
53. Zhou G., Zhang J., Zhang C., Feng Y., Chen L., Yu Z., et al. (2016). Effects of changes in straw chemical properties and alkaline soils on bacterial communities engaged in straw decomposition at different temperatures. Sci. Rep. 6:22186. doi: 10.1038/srep22186. CrossRef. Google Scholar.
54. Zogg G. P., Zak D. R., Ringelberg D. B., MacDonald N. W., Pregitzer K. S., White D. C. (1997). Compositional and functional shifts in microbial communities due to soil warming. Soil Sci. Soc. Am. J. 61, 475–481. doi: 10.2136/sssaj1997.03615995006100020015x. CrossRef. Google Scholar.
55. Zornoza R., Guerrero C., Mataix-Solera J., Scow K. M., Arcenegui V., Mataix-Beneyto J. (2009). Changes in soil microbial community structure following the abandonment of agricultural terraces in mountainous areas of Eastern Spain. Appl. Soil Ecol. 42, 315–323. doi: 10.1016/j.apsoil.2009.05.011. CrossRef. Google Scholar.

Chen C-N, Liao C-S, Tzou Y-M, Lin Y-T, Chang E-H and Jien S-H (2024) Soil quality and microbial communities in subtropical slope lands under different agricultural management practices. Front. Microbiol. 14:1242217. doi: 10.3389/fmicb.2023.1242217
Перевод статьи «Soil quality and microbial communities in subtropical slope lands under different agricultural management practices» авторов Chen C-N, Liao C-S, Tzou Y-M, Lin Y-T, Chang E-H and Jien S-H, оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык
Фото: Designed by Freepik

















































































































Комментарии (0)