Как климат менял рацион древних жителей Перу: данные из Вичамы
Баланс между рыболовством и земледелием в развитии сложных обществ в древних Центральных Андах является предметом давних дискуссий. Несмотря на доступность морских ресурсов в одной из богатейших морских экосистем мира и крайнюю засушливость ландшафта, современные данные свидетельствуют о том, что земледелие было главной движущей силой ранней цивилизации Анд.
Аннотация
В этом исследовании мы применили анализ стабильных изотопов для реконструкции рациона питания 38 человек из двух слоёв заселения Вичамы — прибрежного поселения с монументальной архитектурой, расположенного в долине Уаура, — относящихся к периоду Раннего Форматива-1 (EF-1, 1800–1500 гг. до н. э.) и Позднему Промежуточному периоду (LIP, 1000–1300 гг. н. э.). Результаты показывают значительный вклад культурных растений С3-типа и морского белка в рацион в период EF-1, что указывает на первостепенное значение земледелия в то время, тогда как потребление морепродуктов возросло в период LIP, характеризующийся более высокой морской продуктивностью. Наши выводы проливают свет на сложное взаимодействие между климатическими изменениями и гегемонией прибрежных и внутриконтинентальных поселений в Центральных Андах.
Введение
Связь между системами жизнеобеспечения, климатическими колебаниями и социальной сложностью была подробно изучена и задокументирована для Центральных Анд. Исследования показали, что древние андские популяции развивали адаптивные стратегии, обусловленные экологическими ограничениями, с момента их прибытия в регион ~14 000 лет назад до начала позднего голоцена (Dillehay, 2017; León, 2013; Piperno and Dillehay, 2008). Свидетельства одомашнивания датируются 8000 г. до н. э., и имеющиеся данные свидетельствуют о том, что ирригационные технологии поддерживали выращивание растений уже к ~4500 г. до н. э. (Dillehay et al., 2005, 2007). С ~3000 г. до н. э. земледелие было важнейшей экономической деятельностью (Grobman et al., 2012; Yseki et al., 2023).
Увеличение размеров и распространения поселений, изменения в жилой архитектуре и пространственно-временное моделирование демографической динамики для перуанского побережья указывают на экспоненциальный рост населения между 2000 и 1 г. до н. э. (Goldberg et al., 2016). Этот процесс был связан со сложным взаимодействием между экологическими изменениями и расширением сельскохозяйственных режимов в регионе (Contreras, 2010; Goldberg et al., 2016; Wilson et al., 2022).
За последние десятилетия был получен большой объем археологической информации для Северо-Центрального побережья Перу (PNCC), эпицентра ранней сложности в Центральных Андах (Creamer et al., 2013; Haas and Creamer, 2006; Makowski, 2023; Shady et al., 2001; Shady, 2018). Хотя новаторские свидетельства предполагали, что морские ресурсы были основой начальной социальной сложности в этой крайне сложной среде (Moseley, 1975), новые данные о потреблении растений на прибрежных и среднедолинных монументальных памятниках убедительно свидетельствуют о том, что земледелие было системой жизнеобеспечения, поддерживавшей возникновение ранней цивилизации в регионе (например, цивилизация Караль) и последующее социополитическое развитие (Dillehay et al., 2007; Haas et al., 2013; Pezo-Lanfranco and Colonese, 2024; Shady, 2006a,b; Shoji et al., 2023; Tung et al., 2021).
В этом районе исследования показывают, что прибрежные и внутриконтинентальные общества были организованы вокруг симбиотических экономик земледелия и рыболовства и прибрежно-внутренних торговых сетей по крайней мере с начального формативного периода (3000–1800 гг. до н. э.), при этом прибрежные сообщества основывались на морских ресурсах с некоторым вкладом возделываемых культур, а внутриконтинентальные группы полагались на возделываемые растения, дополненные морскими продуктами (Pezo-Lanfranco et al., 2022; Shady, 2018; Yseki et al., 2023).
Однако на эту систему влияла изменчивость температуры поверхности моря (SST), динамика апвеллинга и интенсивность продуктивности в системе Гумбольдтова течения (HCS), а также Эль-Ниньо – Южное колебание (ENSO). Повышение SST приводит к снижению морской биомассы, тогда как в верхних долинах оползни усиливаются в результате интенсивных осадков, вызывая непредсказуемые последствия для сельскохозяйственного производства (Contreras, 2010). На северном и центральном побережье Перу влажные периоды связаны с преобладанием условий, подобных Эль-Ниньо, тогда как условия, подобные Ла-Нинья, могут ассоциироваться с сухим и холодным климатом (Salvatteci et al., 2014, 2019; Leclerc, 2023).
Хотя окончательные причины и сложные механизмы взаимодействия вовлеченных факторов (например, телесвязи ENSO, интенсивность циркуляции Уокера, меридиональное смещение внутритропической зоны конвергенции и Южно-Тихоокеанского субтропического антициклона) все еще обсуждаются (Mächtle and Eitel, 2013; Salvatteci et al., 2014, 2016; Scholz et al., 2014; Yseki et al., 2022), вариации SST, условий апвеллинга и интенсивности продуктивности в HCS, происходящие в разных временных масштабах (Salvatteci et al., 2016, 2019; Yseki et al., 2022), вероятно, оказали значительное влияние на доступность и выбор морских ресурсов, используемых древними прибрежными сообществами, и повлияли на подъем и упадок древних андских обществ в течение голоцена (Wilson et al., 2022, 2024). Эти эффекты проявлялись наряду с постоянными опасностями, такими как тектоническая активность и геоморфологические особенности (Orloff, 2022; Salvatteci et al., 2019; Sandweiss et al., 2009).
Экспортная продуктивность относится к доле органического углерода, производимого фотосинтезирующими организмами в фотической зоне, которая в конечном итоге откладывается на океанском дне; таким образом, она служит прокси для колебаний SST в океанографии (Herbert, 2003; Scholz et al., 2014; Wefer et al., 2015). Комплексное палеоокеанографическое исследование HCS на основе данных высокого разрешения (т.е. соотношения алкенонов и брома к титану [Br/Ti]) показало вековые изменения SST и, в целом, увеличение экспортируемой продуктивности с ~3000 по 1100 гг. до н. э. (Salvatteci et al., 2019).
Исследования показывают, что холодный период вдоль перуанского побережья между 1800 и 1200 гг. до н. э. привел к снижению SST и увеличению морской продуктивности (Salvatteci et al., 2016, 2019), что привело к изменениям в равновесии политической власти в пользу прибрежных популяций. Высокая морская продуктивность перуанского моря была зафиксирована между ~1700 и 1500 гг. до н. э. (Salvatteci et al., 2019), что совпадает с расцветом нескольких церемониальных центров вдоль прибрежной зоны, таких как Вичама (Shady et al., 2015) и Бандуррия (Chu, 2011), расположенных соответственно вдоль побережья северного и южного межбассейнов долины Уаура, наиболее продуктивной в PNCC. Однако степень, в которой изменения морской продуктивности могли привести к усилению зависимости от морских ресурсов в ущерб сельскохозяйственным продуктам, хорошо известным в регионе с предыдущих периодов, остается непроверенной. Для изучения этого вопроса мы интегрируем данные стабильных изотопов (δ13Cколлаген, δ15Nколлаген, δ13Cапатит) и байесовские модели смешения стабильных изотопов (BSIMM) человеческих индивидов из Вичамы — прибрежного памятника раннего формативного периода (EF) с монументальной архитектурой, «ключевого» для понимания хронологии изменений в питании и нюансов формативной экономики. С более широкой теоретической перспективы эта статья предоставляет ценные сведения о том, как климатические условия влияли на подъем и упадок сложных доисторических обществ (Kennett and Marwan, 2015).
Археологический контекст: Вичама
Вичама — это археологический памятник, расположенный в современном городе Вегета, в 130 км к северу от Лимы, в PNCC (UTM WGS84 N8779900/E212200). Памятник находится на правом берегу нижней части долины Уаура (межбассейн Уаура–Супе), в 1,5 км от Тихого океана и в 6,5 км к северо-западу от устья реки (Рисунок 1). Река Уаура является постоянным источником воды, протекающим ~158 км от своего истока в Андийском нагорье (т.е. лагуна Сура Сака — Кордильера-де-Раура) до моря, и служит естественным коридором, связывающим поперек побережье с важными социально-экономическими регионами, такими как бассейн Мантаро и восточные андийские низменности (например, Альто-Маранон, Альто-Уальяга).
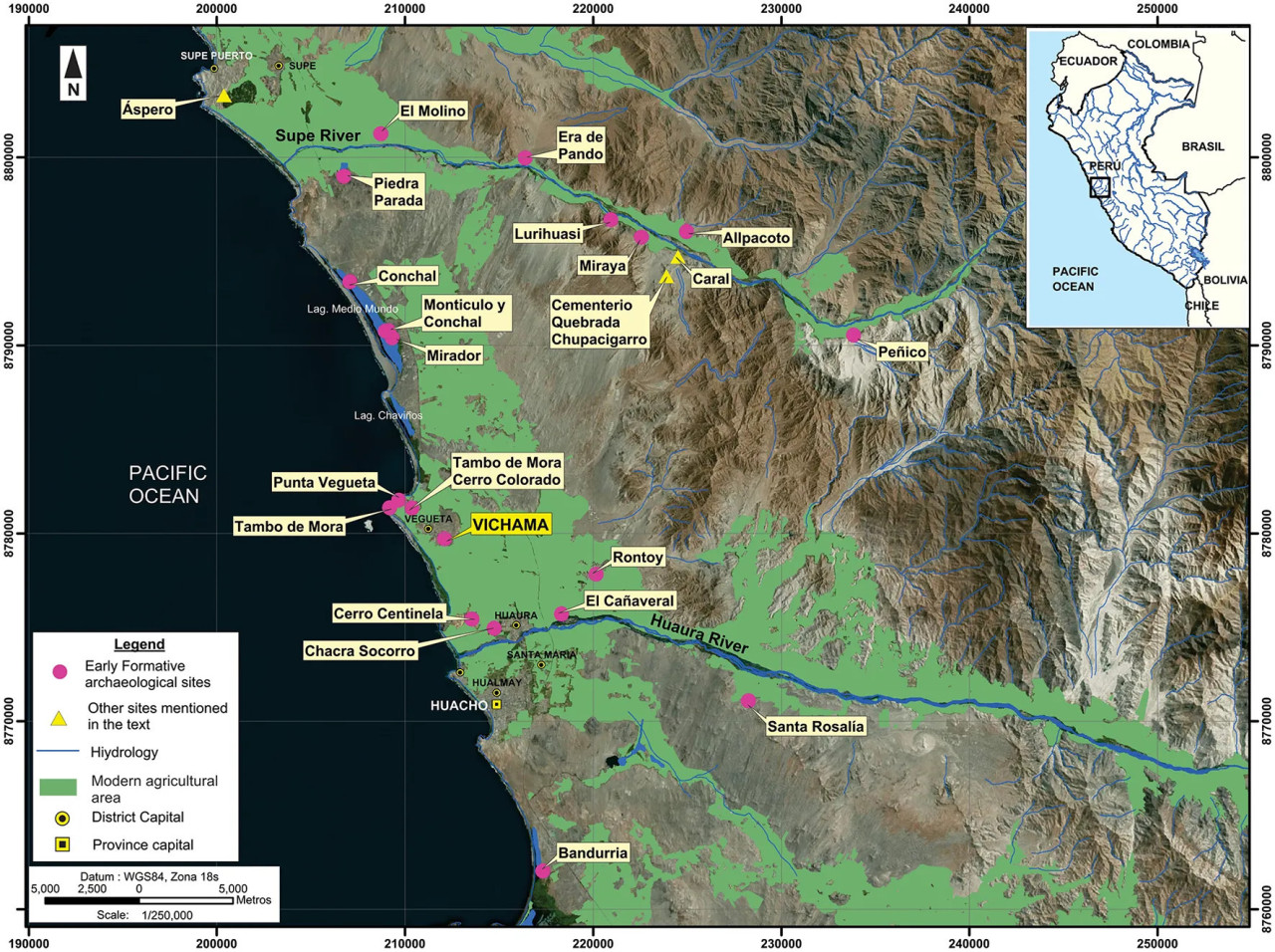
Рисунок 1 Расположение Вичамы в ранний формативный период (1800–1500 гг. до н. э.) и других крупных формативных поселений Северо-Центрального побережья Перу.
Хотя нижняя часть долины Уаура была объектом археологических разведок с начала XX века (Engel, 1987; Tello, 2004), Вичама не подвергалась систематическому изучению до 1970-х годов (Aguilar, 2006; Cárdenas, 1977). С 2007 года Вичама интенсивно раскапывается и консервируется проектом Караль (Zona Arqueológica Caral-ZAC) при поддержке перуанского правительства и местных учреждений (Shady et al., 2008, 2015).
Археологические свидетельства и радиоуглеродные даты указывают на то, что памятник был заселен в два хронологических периода: в период Раннего Форматива-1 (EF-1, 1800–1500 гг. до н. э.), когда монументальная архитектура процветала на протяжении шести архитектурных периодов (P1–P6), и в период Позднего Промежуточного периода (LIP, 1000–1300 гг. н. э.), когда памятник стал рыбацкой деревней с уменьшенным политическим влиянием в этом секторе долины в период высокой морской продуктивности (Рисунок 2).
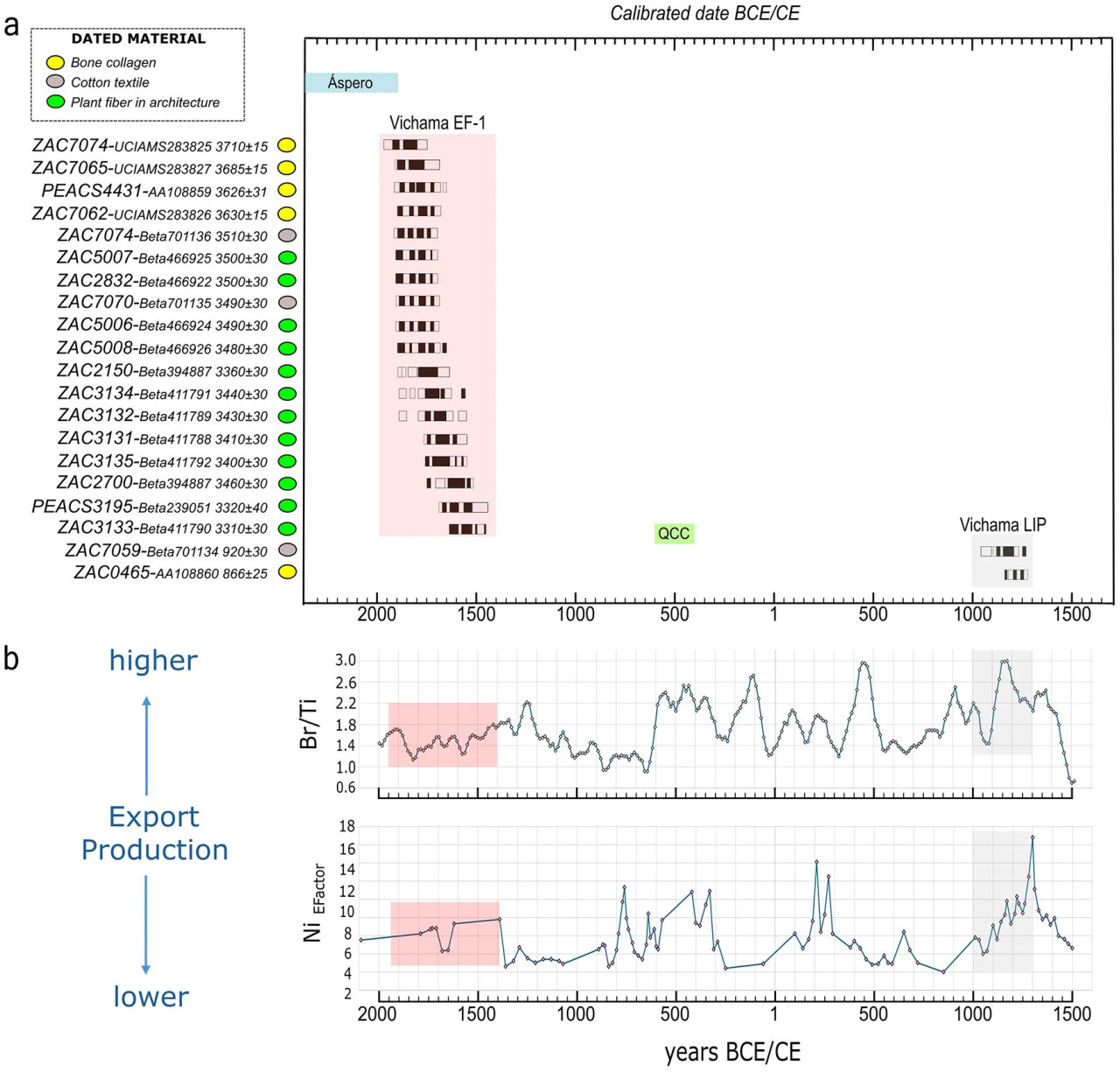
Рисунок 2 (a) Радиоуглеродные даты Вичамы и прокси экспортной продуктивности на перуанском побережье в периоды Раннего Форматива-1 (EF-1) и Позднего Промежуточного периода (LIP). См. также хронологию двух памятников, упомянутых в статье (т.е. Асперо и кладбище Кебрада-Чупасигарро — QCC). (b) Индексы экспортной продуктивности: записи соотношения брома к титану (Br/Ti, керн M135-004-3, широта 17° ю.ш.) и фактора обогащения аутигенным никелем (EFactor; керн G-10-GC-02, широта 14° ю.ш.). Соотношение брома к титану является полуколичественным методом оценки осадочного общего органического углерода, а никель (EFactor) указывает на поток оседающего органического вещества (Salvatteci et al., 2016). Цветные области представляют классификацию сравниваемых периодов (красным — EF-1 и серым — LIP). Исходные данные доступны по адресу: https://doi.pangaea.de/10.1594/PANGAEA.887109.
Вичама представляет собой многокомпонентный комплекс, включающий девять секторов, включая формативную монументальную архитектуру (Vi-01) и ракушечные кучи (Vi-02, Vi-03), а также архитектуру и кладбища более поздних периодов (Vi-04–Vi-09), разбросанные на 136 га в холмах Серро-Альконсильо, скалистой возвышенности, поднимающейся на 114 м над уровнем моря в центре пустынного оазиса площадью 305 га (Рисунок 3a).
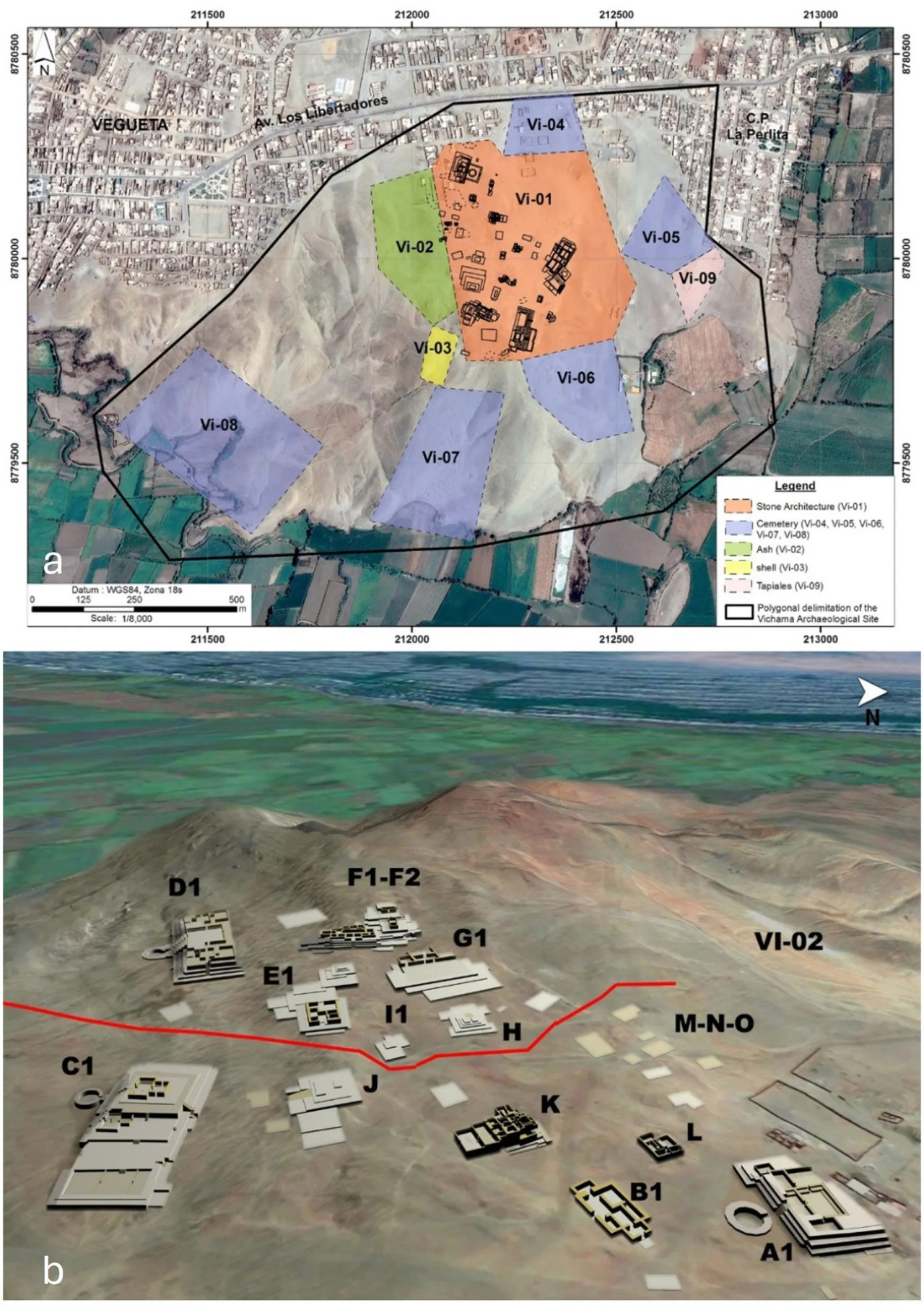

Рисунок 3 (a) Секторы археологического памятника Вичама. (b) Трехмерная реконструкция некоторых значимых монументальных зданий в верхней и нижней половинах памятника. Верхняя половина: Edificio Público Mayor (Сектор D1), Edificio Las Ofrendas (Сектор F), Edificio Público Residencial (Сектор E1), Edificio Público G1, Edificio Público Mediano (Сектор H), Edificio Público Menor (Сектор I1). Нижняя половина: Edificio Principal (Сектор A1), Edificio Residencial de Elite (Сектор B1), Edificio Público Los Depósitos (Сектор K), Edificio de las Chakanas (Сектор C1), Edificio Público Menor (Сектор L), Edificio Residencial de Elite (Сектор J), Sector Residencial (Сектор M-N-O), жилые и погребальные зоны (VI-02). См. дополнительный материал 2 для подробного расположения погребальных контекстов и индивидов, оцененных в этой работе.
В период EF-1 Вичама была частью формирующейся сложной политии в нижней части долины Уаура, которая, вероятно, состояла не менее чем из 20 поселений с монументальной архитектурой (Aguilar, 2006; Cárdenas, 1977). Возможно, Вичама представляла собой появление нового социополитического центра в регионе после упадка общества Караль. Ядерная зона в то время включала 29 зданий, включая общественные (например, ступенчатые платформы, круглые солнечные дворы, площади и стены с рельефами) и жилую архитектуру ортогонального плана, занимавшую 25 га (Shady et al., 2015). Эта территория разделена на две половины — верхнюю и нижнюю, следуя бипартитному пространственному делению, распространенному в андийских поселениях (Рисунок 3b). Церемониальные здания демонстрируют различные архитектурные особенности, включая каменные стены, расположенные горизонтальными рядами, архитектурные заполнения с использованием мега-шикрас и стены, украшенные рельефами высокого рельефа (Shady et al., 2008, 2015). Изменения в церемониальной архитектуре и инвентаре (т.е. полихромные скульптуры из необожженной глины и особая иконография) свидетельствуют об идеологических трансформациях.
Питание в Вичаме: современные данные
«Визуальный бассейн» Вичамы охватывает примерно 9,2 км² и включает разнообразные экосистемы, которые, возможно, использовались для жизнеобеспечения, включая скалистые и песчаные пляжи, болота, ломасы (туманные оазисы), сельскохозяйственные поля, реку и море. Побережье долины Уаура является одним из богатейших районов перуанского моря, на которое в настоящее время приходится ~20% от общего национального объема рыболовства (IMARPE, 2010, p. 64). Острова и островки (например, Анат, Асакан), расположенные менее чем в 1 км от берега, потенциально обеспечивали густые банки моллюсков, морских черепах и разнообразных морских птиц.
Значение рыболовства неоспоримо. Зооархеологический материал включает в общей сложности 91 вид (23 рыбы, 61 моллюск, 3 морских млекопитающих и 4 морские птицы), извлеченных из первичных контекстов (см. дополнительный материал 1: S1 — Морские виды).
Кроме того, эксплуатация прибрежных соляных месторождений (т.е. Пунта-де-Атауанка и Салинас-де-Уачо и др.) для производства сушеной соленой рыбы, вероятно, была важной экономической деятельностью в регионе в начальный формативный и EF периоды (Engel, 1987; Tello, 2004). Современные рыбаки Вегеты используют несколько традиционных методов, таких как редес-де-серко, дайвинг, чинчорро и арте-де-пинта, которые могли использоваться в прошлом для вылова широкого спектра зарегистрированных видов (IMARPE, 2010, p. 67).
Согласно последнему отчету (Yseki et al., 2024), археоботанический инвентарь съедобных растений в Вичаме содержит более 35 000 макроостатков, представляющих 17 таксонов, происходящих из 123 первичных контекстов (см. дополнительный материал 1: S1 — Виды съедобных растений). Ботанический материал включает фрукты (гуава, лукма, пакай, авокадо и чируэла-дель-фрайле), однолетние растения, такие как перец чили (например, Capsicum baccatum), бобы (т.е. лимская фасоль, обыкновенная фасоль и конские бобы), тыквы, а также корнеклубнеплоды (т.е. ачира, сладкий картофель и картофель). Судя по их распространенности, плодовые деревья, особенно гуава и лукма, а также перец чили, регулярно употреблялись и использовались в церемониальной деятельности. Аналогично, высокая распространенность в контекстах подношений остатков лимской фасоли, ачиры, сладкого картофеля и картофеля подчеркивает экономическую важность этих растений в Вичаме (Yseki et al., 2024).
Отсутствие макроостатков Zea mays (кукурузы) свидетельствует о том, что она, вероятно, не выращивалась, не перерабатывалась и не хранилась в Вичаме. Однако недавнее исследование сообщило о крахмальных зернах с потенциальными повреждениями, связанными с ферментацией, в зубном камне индивидов из Асперо (Yseki et al., 2023), при минимальном присутствии макроостатков кукурузы (Shady and Cáceda, 2008). Таким образом, потребление кукурузы в Вичаме не может быть исключено только на основе анализа макроостатков, поскольку кукуруза могла употребляться в виде напитка или муки (Yseki et al., 2024).
Морская фауна и некоторые различия между периодами свидетельствуют о колебаниях морских продуктивных условий, связанных с апвеллингом богатых питательными веществами морских вод. Например, в течение EF периода (1800–1200 гг. до н. э.) наблюдается общая тенденция к увеличению экспортируемой продуктивности, особенно в течение EF-1 (1800–1500 гг. до н. э.), с некоторыми колебаниями (т.е. более низкая продуктивность между 1800 и 1600 гг. до н. э., с самыми низкими значениями около 1700 гг. до н. э. и более высокой продуктивностью между 1600 и 1500 гг. до н. э.), которые должны были влиять на состав рациона, возможно, с более высоким потреблением морских продуктов; однако растительные материалы показывают, что земледелие продолжалось во время расцвета монументальной архитектуры.
Эти условия, однако, могли быть относительно иными по сравнению с наблюдаемыми в течение LIP (1000–1300 гг. н. э.), когда продуктивность достигла еще более высоких значений и усиленных условий апвеллинга, с максимумом около 1000–1100 гг. н. э. (Kennett and Marwan, 2015). В течение LIP археологические отложения изобилуют морскими остатками, такими как рыбьи кости и раковины, рыболовные сети и артефакты для морской эксплуатации.
Стабильные изотопы в реконструкции питания
Хотя остатки фауны и растений могут дать представление о сдвигах в хозяйственной практике, анализ стабильных изотопов является более эффективным для определения чистого потребления конкретных пищевых ресурсов на индивидуальном уровне. Анализ стабильных изотопов углерода и азота в коллагене (δ13Cколл и δ15N) и биоапатите (δ13Cап) из человеческих костей и зубов предоставляет ценную информацию о питании древних популяций (Schoeninger and Reitsema, 2023). δ13Cколл является прокси источников белка и трофического положения потребителей в пищевой сети, а также фотосинтетического пути потребляемых растений. δ13Cап, напротив, отражает всю совокупность углеводов, липидов и белков в питании, обеспечивая полезное средство для оценки энергетических составляющих рациона и фотосинтетических путей (Jim et al., 2004; Kellner and Schoeninger, 2007). На центральном побережье Анд растения C3, использующие фотосинтетический путь Кальвина–Бенсона, являются наиболее распространенными и обычно демонстрируют средние значения δ13C −25,7 ± 2,1‰. Напротив, среди растений C4, следующих фотосинтетический путь Хэтча–Слэка, есть только два потенциально съедобных одомашненных растения (кивича — Amaranthus sp. и кукуруза — Zea mays), с типичными значениями примерно −11,3 ± 1,4‰. Кроме того, растения с крассулоциевым метаболизмом кислот (CAM) имеют изотопные значения (−10,8 ± 0,2‰), которые перекрываются с C4 (Pezo-Lanfranco and Colonese, 2024).
Обычно потребители растений C4 показывают значительно более высокие значения δ13C (Kellner and Schoeninger, 2007). Однако в прибрежных зонах использование δ13C для отслеживания потребления кукурузы может быть затруднено из-за наличия морских ресурсов (т.е. рыбы и моллюсков), которые имеют перекрывающиеся значения δ13C с растениями C4, примерно −11,9 ± 2,1‰ (Pezo-Lanfranco and Colonese, 2024). С точки зрения значений δ15N, травоядные имеют тенденцию к значениям на 3–7‰ выше, чем потребляемые растения, в то время как плотоядные демонстрируют значения δ15N на 3–5‰ выше, чем их добыча. Поскольку морские источники имеют более высокие изотопные значения по сравнению с наземными источниками (Minagawa and Wada, 1984; Schoeninger and DeNiro, 1984), морская фауна показывает более высокие значения δ15N, чем наземная, что связано с трофическим фракционированием в сложных морских пищевых сетях (Chisholm et al., 1982; DeNiro and Epstein, 1981; Hedges and Reynard, 2007; Schoeninger, 2010; Schoeninger et al., 1983). Анализ стабильных изотопов особенно информативен в сочетании с байесовскими моделями смешения для получения количественных оценок состава рациона в виде распределений вероятностей (Cheung and Szpak, 2021; Fernandes et al., 2014, 2015; Phillips, 2012).
Гипотеза и ожидания
Используя комбинацию прокси стабильных изотопов из коллагена (δ13Cколл, δ15Nколл, n = 36) и апатита (δ13Cап, n = 26) 38 человеческих индивидов из Вичамы, а также оценки BSIMM калорийного вклада потенциальных источников пищи (т.е. морской и наземной фауны, а также растений C3 и C4; Fernandes et al., 2014, 2015), потребляемых 21 индивидом из Вичамы (см. Материалы и методы для подробностей реализации модели), мы стремимся выяснить основные источники энергии для двух периодов заселения Вичамы: EF-1 (1800–1500 гг. до н. э.) и LIP (1000–1300 гг. н. э.).
Мы проверили гипотезу о том, что изменения морской продуктивности влияли на степень включения морских ресурсов в андские системы жизнеобеспечения (Kennett and Marwan, 2015; Wilson et al., 2024). Согласно палеоклиматическим данным, повышенная морская продуктивность между 1800 и 1500 гг. до н. э. совпадала с расцветом Вичамы (Shady et al., 2015). Однако экономическая важность растений и рыбы и их относительный вклад в рацион неизвестны. Ожидается, что жители Вичамы, особенно в период EF-1, в большей степени полагались на морские продукты как на значительную часть своего рациона.
Для сравнения, LIP (1000–1300 гг. н. э.) был временем похолодания климата и снижения SST, когда морская продуктивность достигла своего пика (Kennett and Marwan, 2015). Поскольку LIP признан периодом более высокого развития сельского хозяйства в Центральных Андах, в условиях усиливающегося апвеллинга и морской продуктивности ожидается смешанный рацион с более высоким вкладом морских источников. Эта зависимость должна отражаться в палеодиетических реконструкциях.
Результаты
Реконструкция питания
Изотопные значения 38 человеческих индивидов из Вичамы были классифицированы по двум периодам заселения: EF период (1800–1500 гг. до н. э., n = 30) и LIP (1000–1300 гг. н. э., n = 8), использованные здесь для сравнительных целей (Таблица 1, дополнительный материал 1: S2 для контекстов индивидов и биоархеологических деталей и S3 для радиоуглеродных дат). Только часть из 59 отобранных элементов (36 костей и 23 зуба) дали неповрежденный коллаген с надежными значениями δ13Cколл и δ15N из костей (n = 21) или зубов (n = 15) в соответствии с принятыми критериями сохранности (DeNiro, 1985; Van Klinken, 1999). Исследования методом АТР-Фурье-ИК-спектроскопии (ATR-FTIR), проведенные на подвыборке из 10 индивидов, стабильно свидетельствуют о том, что диагенетические изменения не затронули неорганическую фракцию костей из контекстов Вичамы (France et al., 2020; см. дополнительный материал 1: S4 для данных ATR-FTIR). Таким образом, значения δ13Cап (n = 26) были включены в наш анализ для дополнения палеодиетической информации.
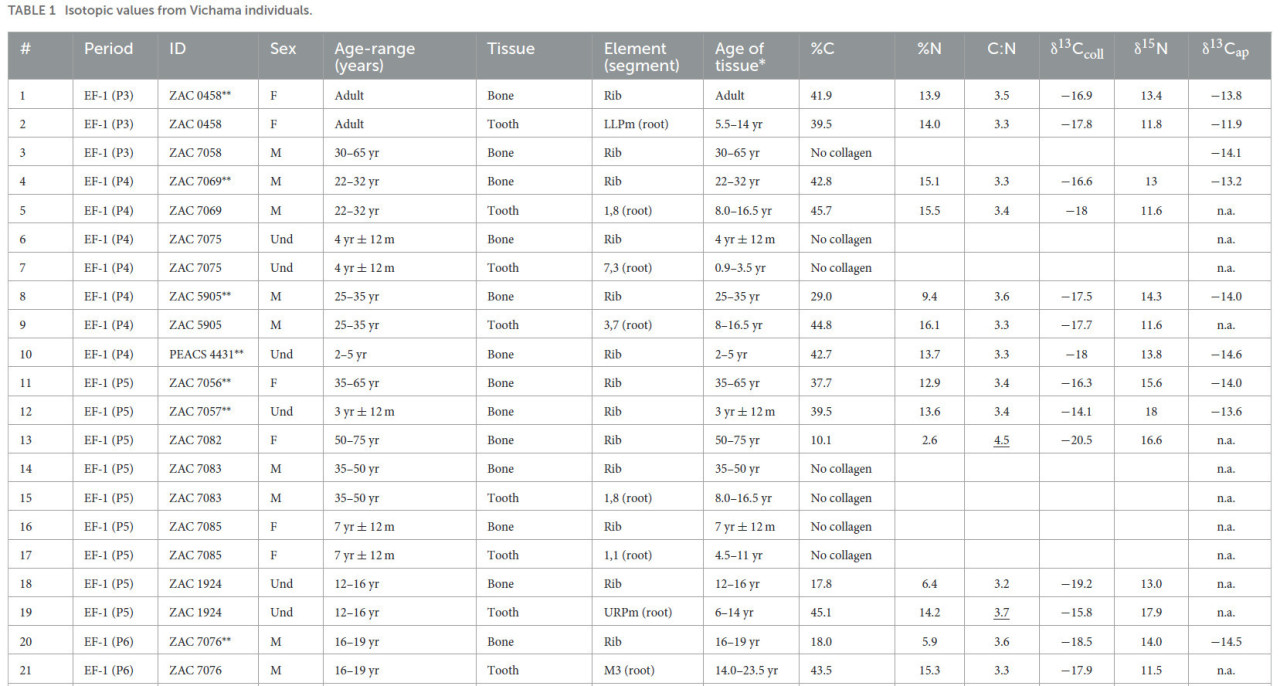
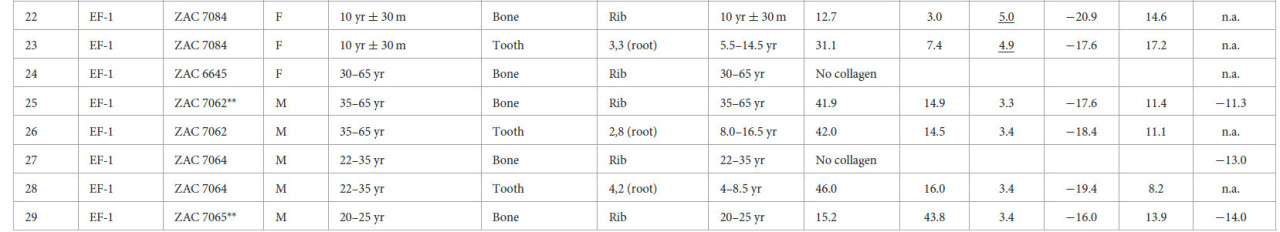
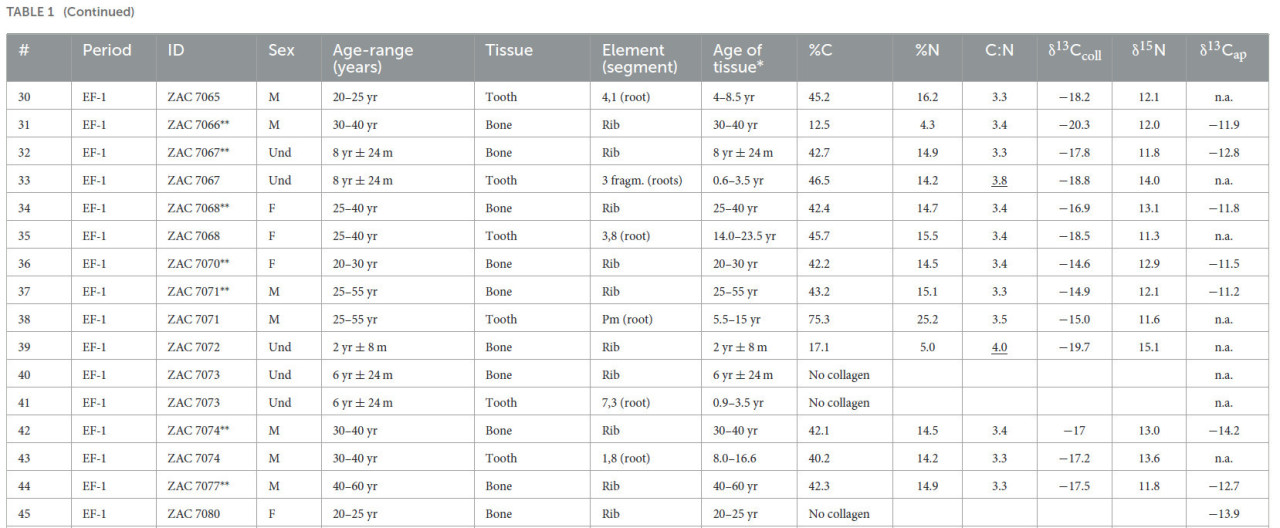
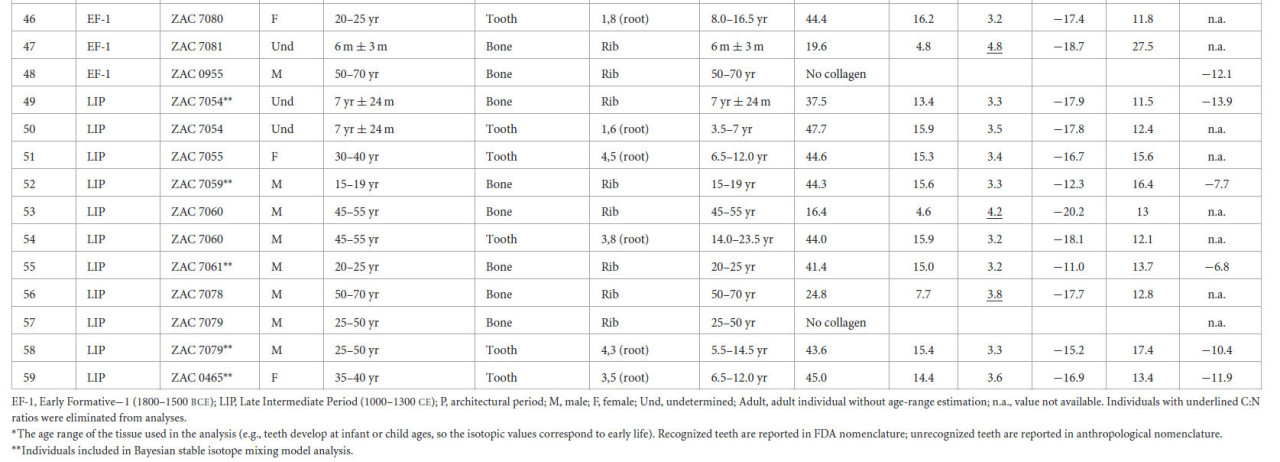
Таблица 1 Изотопные значения индивидов из Вичамы.
EF-1 — Ранний Форматив−1 (1800–1500 гг. до н. э.); LIP — Поздний Промежуточный период (1000–1300 гг. н. э.); P — архитектурный период; M — мужской; F — женский; Und — неопределенный; Adult — взрослый индивид без оценки возраста; n.a. — значение недоступно. Индивиды с подчеркнутыми соотношениями C:N были исключены из анализа.
Значения δ13Cколл из костей и дентина зубов варьировались от −20,3 до −11,0‰, значения δ15N — от +8,2 до +18,0‰, тогда как значения δ13Cап варьировались от −14,6 до −6,8‰ (дополнительный материал 1: S5).
Хотя вклад морского белка очевиден у некоторых мужских индивидов (Рисунок 4a), средние значения δ15N ниже, чем ожидалось для рыбаков, и свидетельствуют о потреблении морского белка низкого трофического уровня, возможно, мелкой рыбы и моллюсков (со средними значениями δ13Cколл = −12,3‰, σ = 0,5 и δ15N = +10,4‰, σ = 1,2) на протяжении всей последовательности. Значительных различий в значениях δ13Cколл, δ15N и δ13Cап между женщинами и мужчинами или между подростками и взрослыми для обоих периодов EF-1 и LIP не наблюдалось (дополнительный материал 1: S5). Значительное различие, обнаруженное между взрослыми и подростками EF-1, вероятно, является результатом эффекта отлучения у нескольких младенцев (т.е. PEACS4431 и ZAC7057) или различных источников белка (т.е. ZAC7067) у некоторых подростков.
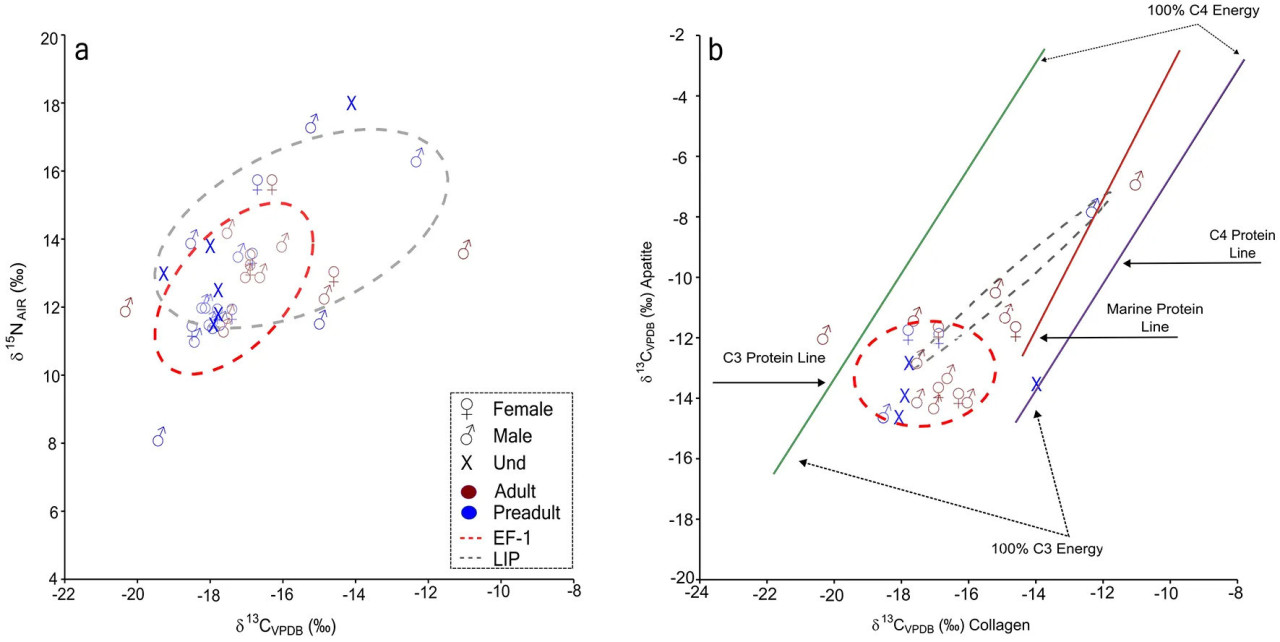
Рисунок 4 Диаграмма рассеяния маркеров стабильных изотопов у индивидов из Вичамы раннего формативного периода и позднего промежуточного периода и перекрывающиеся площади стандартных эллипсов (оцененные для 68,5% популяции) для сравнения. (a) Значения δ13Cколл и δ15N. (b) Значения δ13Cколл и δ13Cап в рабочем пространстве Kellner and Schoeninger (2007). V-PDB — Вена Пий Ди Билемнит; AIR — атмосферный воздух.
Межпопуляционные сравнения выявили значительное различие только для δ13Cап (Mann–Whitney U = 18,50, p = 0,027). В целом, δ13Cколл и δ13Cап указывают на рационы, в которых доминируют белки и источники энергии из экосистем C3 (Рисунок 4b; Kellner and Schoeninger, 2007). Тем не менее, индивиды из LIP демонстрировали гораздо более широкий диапазон значений δ13Cколл и δ15N, указывающий на относительно более высокое потребление морского белка, особенно у некоторых мужских индивидов (например, ZAC7061, ZAC7059), по сравнению с популяцией EF-1.
Сравнения источников белка фаз Вичамы EF-1 и LIP с другими популяциями из PNCC показывают тенденцию к потреблению диеты, промежуточной между диетой Асперо — наиболее репрезентативного памятника рыбаков в долине Супе в начальный формативный период (3000–1800 гг. до н. э.), основанной на растениях C3 и морском белке более высокого трофического уровня, и диетой более поздних земледельцев с кладбища Кебрада-Чупасигарро (средняя долина Супе, поздний формативный период, 500–400 гг. до н. э.), практически полностью основанной на растениях (Pezo-Lanfranco et al., 2022). Рационы LIP показывают большую площадь перекрытия с рационами Асперо, чем EP-1 (Рисунок 5).
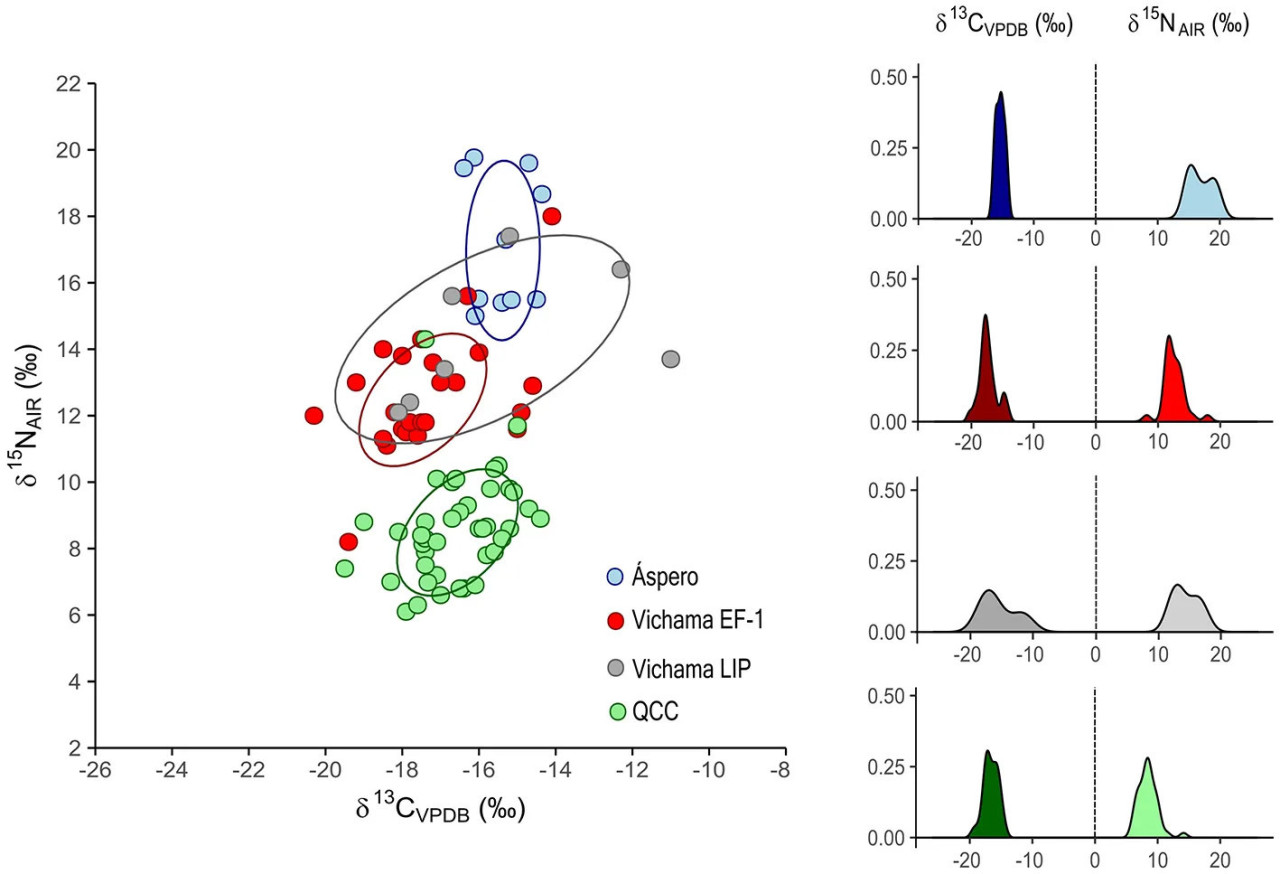
Рисунок 5 Диаграмма рассеяния δ13C и δ15N с площадями стандартных эллипсов (оцененными для 68,5% популяции) и графиками ядерной плотности значений δ13Cколл и δ15N индивидов из Вичамы и других формативных популяций Северо-Центрального побережья Перу. EF-1 — период Раннего Форматива−1; LIP — Поздний Промежуточный период; QCC — кладбище Кебрада-Чупасигарро.
Кроме того, мы оценили пропорции пищевых компонентов с помощью BSIMM для 21 индивида из Вичамы с полным набором изотопных данных (Таблица 2, Рисунок 6; см. также дополнительный материал 1: S6 для результатов модели). Эти результаты показывают, что большинство калорий поступало из растений C3 (вклад от 14% до 86%), за которыми следуют рыба и моллюски (8% и 58%), растения C4 (3%−19%) и наземная фауна (3%−11%).
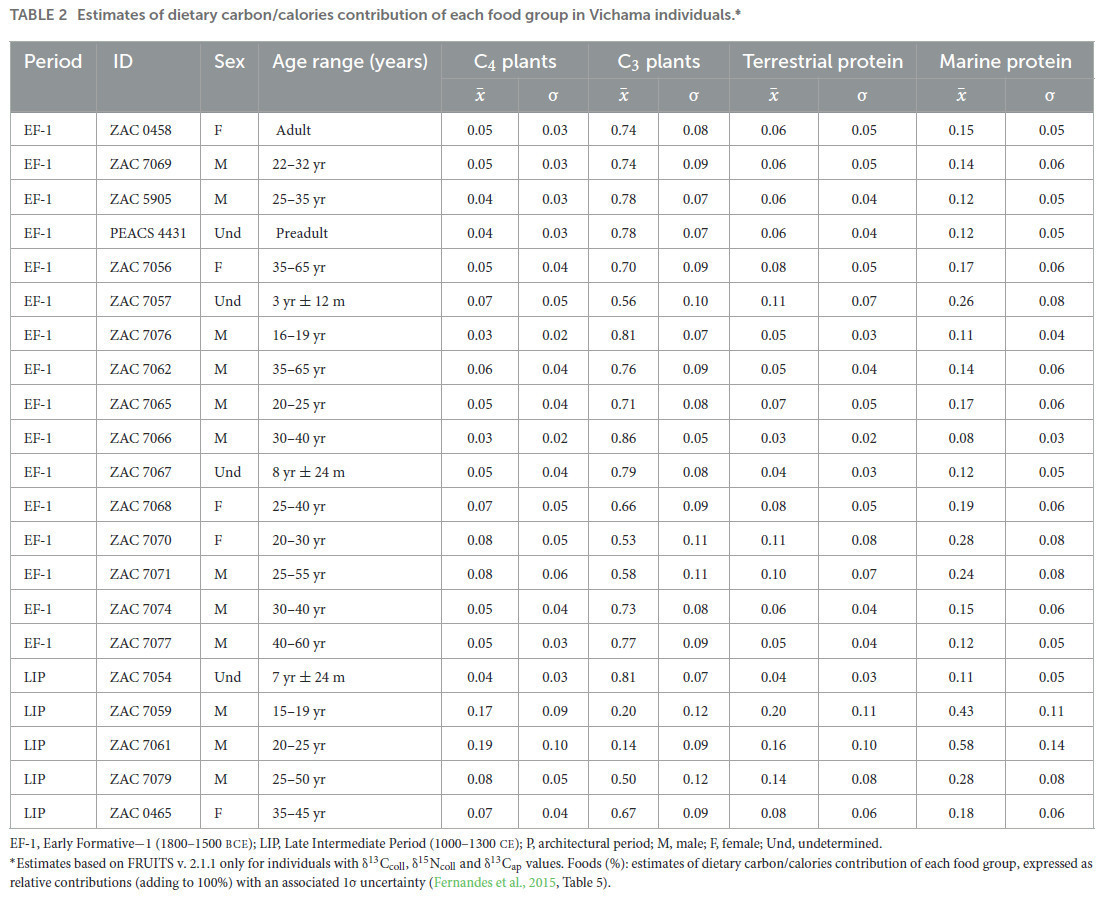
Таблица 2 Оценки вклада пищевого углерода/калорий каждой группы продуктов питания у индивидов из Вичамы.
Оценки основаны на FRUITS v. 2.1.1 только для индивидов со значениями δ13Cколл, δ15Nколл и δ13Cап. Продукты (%): оценки вклада пищевого углерода/калорий каждой группы продуктов, выраженные как относительные вклады (в сумме 100%) с соответствующей неопределенностью 1σ (Fernandes et al., 2015, Таблица 5).
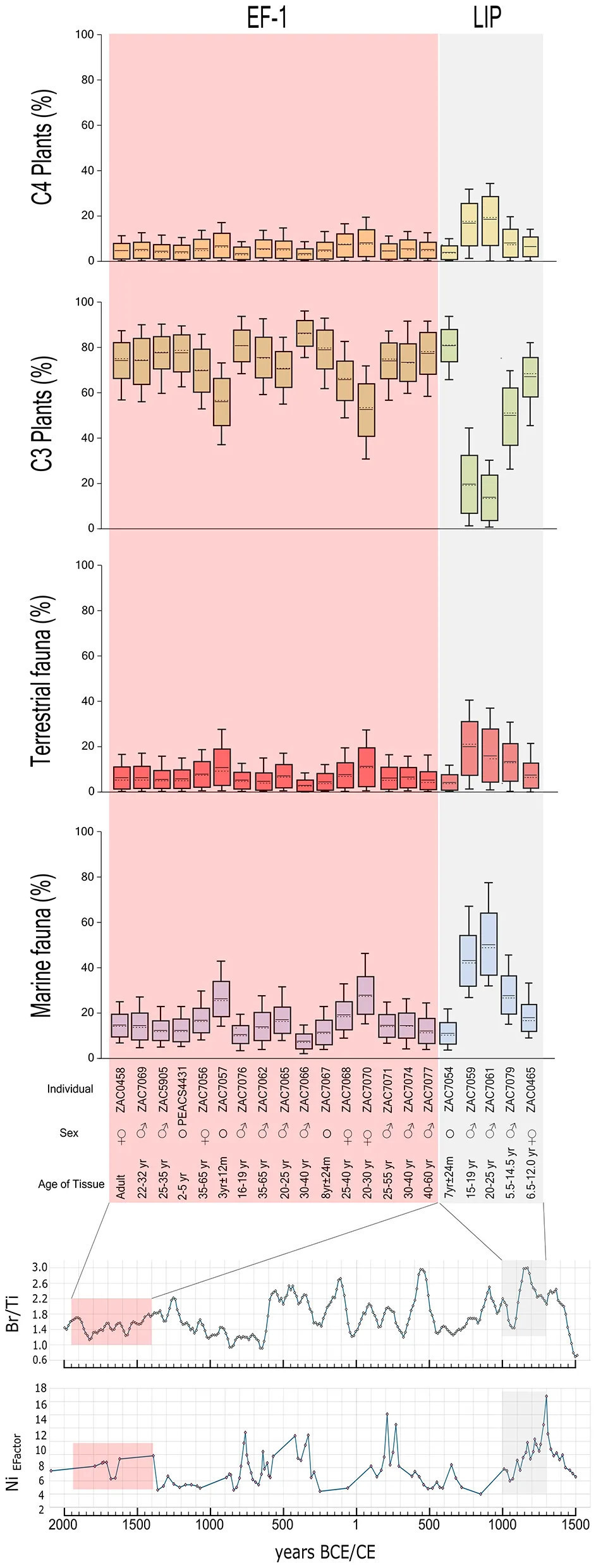
Рисунок 6 Оценки калорийности источников пищи у индивидов из Вичамы для периода Раннего Форматива−1 (EF-1, 1800–1500 гг. до н. э.) и Позднего Формативного периода (LIP, 1000–1300 гг. н. э.) и их соответствие маркерам экспортируемой продукции. Прямоугольники представляют 68% доверительный интервал, а усы — 95% доверительный интервал. Горизонтальная прерывистая линия указывает медиану, а сплошная линия — среднее значение. ♂ — мужской; ♀ — женский; ° — неопределенный пол.
Таким образом, большинство индивидов из периода EF-1 Вичамы показывают значения, совместимые с широким земледельческо-рыболовным рационом, с неожиданно высоким вкладом растений у людей, живущих у моря. Сравнение рационов между EF-1 и LIP свидетельствует о тенденции к увеличению потребления морского белка в более поздний период, меньшему количеству растений C3 и незначительному увеличению C4 и наземного белка. Дивергенция Кульбака–Лейблера составляет 0,141, что указывает на умеренное различие в средних распределениях вероятностей между индивидами EF-1 и LIP, что согласуется с изменениями в питании между периодами с некоторым перекрытием.
Обсуждение
Характер питания в Вичаме
Наше исследование свидетельствует о том, что две различные фазы заселения Вичамы были основаны на разных экономических системах. В период EF-1, датируемый 1800–1500 гг. до н. э., хотя некоторые индивиды значительно полагались на морские ресурсы для получения калорий (>25%), жизнеобеспечение было основано в первую очередь на возделывании растений C3. Растительные остатки, включая несколько высокоэнергетических культур, таких как клубнеплоды, бобы и фрукты, были найдены в основном в погребальных контекстах, связанных с общественными зданиями, однако наши результаты выявляют значимость этих растений на уровне домохозяйств за пределами ритуального использования.
Это сельскохозяйственное производство, вероятно, поддерживало политическое значение Вичамы в конце эры Караль. Возделывание было вероятным видом деятельности в Вичаме в период EF-1. Сельскохозяйственная площадь вблизи Вичамы простирается примерно на 970 га, а источники воды находятся менее чем в 2 км от Вичамы. В этой зоне уровень грунтовых вод составляет примерно 0,85 м глубины (INRENA, 2005).
Свидетельства ботанических макро- и микроостатков показывают потребление нескольких одомашненных культур в прибрежных поселениях в начальный формативный период (3000–1800 гг. до н. э.; Haas et al., 2013; Shady, 2006a,b,c; Vega-Centeno, 2005; Yseki et al., 2023, 2024; Zechenter, 1988). Например, растительные микроостатки из зубного камня человека в Асперо, в том же регионе, указывают на значительное потребление сладкого картофеля и умеренное потребление картофеля в начальный формативный период (Yseki et al., 2023), несмотря на их редкость в макроботаническом материале (Shady and Cáceda, 2008).
Расположение Вичамы подходит для выращивания кукурузы из-за обширных пахотных земель и доступности воды. Интересно, что изотопные данные, полученные в этой работе, предполагают незначительный вклад C4 (менее 10% в большинстве случаев) в период EF-1 и согласуются с отсутствием свидетельств кукурузы в археологическом материале. Почти полное отсутствие кукурузы в Вичаме контрастирует с ее присутствием в некоторых ранних памятниках из соседних долин PNCC (Haas et al., 2013; Shady, 2006b; Vega-Centeno, 2005; Zechenter, 1988). В Асперо крахмал кукурузы был обнаружен в зубном камне (Yseki et al., 2023), а в долине Пативилька пыльца и крахмалы кукурузы появляются уже 2500 кал. до н. э. (Haas et al., 2013). Присутствие амаранта может объяснять сигнал C4, обнаруженный в Вичаме, хотя макроостатки этого растения не были зарегистрированы на памятнике, в отличие от Караля, расположенного в 30 км к северо-востоку, где амарант был задокументирован в более ранний период (Shady, 2006c).
Примерно через 3000 лет, в период LIP (1300–1500 гг. н. э.), когда Вичама давно потеряла свое региональное политическое влияние, жители памятника в основном полагались на растения C3, дополненные более значительным потреблением морских ресурсов. Это повышенное внимание к морским организмам подтверждается наличием рыболовных снастей, таких как сети и рыболовные крючки из раковин мидий, а также обильными остатками рыбы и раковин, что подтверждает широкую рыболовную деятельность в период LIP. Хотя кукуруза могла потребляться в большей степени в период LIP, изотопная эквифинальность усложняет различие между растениями C4 и морскими источниками углерода. Два молодых мужских индивида из LIP (ZAC7059 и ZAC7061) демонстрируют значения потребления C4 примерно 20%; однако оба потребляли примерно 40%−60% калорий из морских источников и имеют слуховые экзостозы — остеологический маркер, связанный с повторяющейся водной активностью в холодных водах (Pezo-Lanfranco et al., 2009), что предполагает, что они были ныряльщиками или рыбаками, а не потребителями кукурузы. Напротив, единственный индивид из EF-1 со слуховыми экзостозами (ZAC5905, мужчина) показывает, вопреки ожиданиям, неожиданно низкую долю морского вклада в рацион и значения δ15N, совместимые с морскими ресурсами низшего трофического уровня.
Перекрытие между значениями углерода кукурузы и морских продуктов остается потенциальным источником неопределенности в изотопных анализах для этого региона. Тем не менее, если кукуруза и потреблялась в Вичаме в период EF-1 или LIP, она, вероятно, не составляла основного или значительного компонента питания, внося менее 30% от общего количества калорий (Ambrose and Norr, 1993).
В период EF-1 потребление наземных млекопитающих было эпизодическим, причем вклад этих источников белка увеличивался в LIP (20% от общего количества калорий), вероятно, из-за большей доступности верблюдовых в этот период (León, 2013). Фаунистический материал Вичамы включает только шесть таксонов: три вида птиц, синантропных грызунов, лягушек и верблюдовых, причем последние являются единственным потенциально съедобным одомашненным таксоном. Однако зооархеологические исследования межпериодного потребления все еще ожидаются. Ограниченная доступность одомашненных видов в Андах, таких как верблюдовые, морские свинки и собаки, а также некоторых диких видов из ломас и пустынных территорий, предполагает, что наземный белок оставался относительно второстепенным компонентом питания в формативный период, как наблюдалось и в других исследованиях (Pezo-Lanfranco et al., 2022; Pezo-Lanfranco and Colonese, 2024). Как потенциальное ограничение этого исследования, присутствие мелких видов рыб может имитировать сигнал дикого наземного белка из ломас, потенциально C4-белка (Coutts et al., 2011; Cadwallader et al., 2012).
Хотя археологические данные EP-1 и LIP подтверждают рыболовную деятельность в обоих периодах, наши результаты показали, что зависимость от морских продуктов была выше в LIP, который совпадает с периодом высокой морской продуктивности, по сравнению с EP-1. Однако из-за ограниченного размера выборки LIP эти интерпретации следует рассматривать как предварительные до получения дальнейших доказательств.
Наконец, хотя некоторые подростковые индивиды, включенные в BSIMM, теоретически могут сохранять азотные изотопные сигналы от грудного вскармливания и отлучения, что приводит к ошибочным интерпретациям, это маловероятно, поскольку проанализированные ткани (корни зубов) соответствуют детям старшего возраста, которые обычно потребляют взрослую диету. Среди индивидов LIP двое являются взрослыми и трое — детьми или подростками, с образцами тканей в возрасте от 5,5 до 14,5 лет. Предыдущие исследования особенностей отлучения в андских популяциях помещают завершение отлучения между 1,5 и 3 годами (см. Pezo-Lanfranco et al., 2020 для всестороннего обзора), что позволяет предположить, что изотопные значения в LIP не зависят от процесса отлучения. В случае EF-1 среди двух проанализированных младенцев (PEACS 4431 и ZAC7057) только ZAC7057 показывает повышенное значение азота, что, вероятно, указывает на продолжающееся потребление грудного молока и потенциально может имитировать высокое потребление морского белка в модели. Все остальные проанализированные индивиды являются взрослыми.
Диетические данные и климатические колебания в Вичаме
Данные изотопов, полученные в этом исследовании, предполагают два различных экономических режима, которые, вероятно, отражают изменения в политической организации Вичамы с течением времени. Системы производства продуктов питания в период EF-1, вероятно, координировались централизованными политическими институтами, направленными на создание излишков за счет интенсивного сельского хозяйства. Напротив, диверсифицированный рацион, наблюдаемый в период LIP, отражает отсутствие этих централизованных систем в Вичаме. Этот сдвиг подтверждает существование диверсифицированной экономики, ориентированной на мелкомасштабное земледелие и рыболовство для потребления в домохозяйствах, с меньшим акцентом на создание излишков. Эти экономические системы также могли формироваться климатическими колебаниями и преобладающими экологическими условиями.
Чередование влажных и сухих периодов, которое стимулировало и подавляло морскую продуктивность, рыболовную деятельность и, возможно, расширение и сокращение ломас и сельскохозяйственных угодий в регионе. Эти явления связаны со сложными океано-атмосферными взаимодействиями (Salvatteci et al., 2019), включая колебания Южно-Тихоокеанского антициклона, пассатов и прибрежных ветров с влиянием на морские течения (Salvatteci et al., 2016, 2019; Yseki et al., 2022).
Помимо диетической адаптации, эти изменчивые климатические условия также могли отражаться в социополитической устойчивости и идеологическом ответе на экологические ограничения, как на то указывает присутствие божеств, связанных с климатом, в иконографии Вичамы (например, божество лягушка-человек), что потенциально отражает идеологические адаптации к экологическим ограничениям (Shady et al., 2015).
Зависимость от морского белка в период EF-1 могла ограничиваться более низкой биомассой и морской продуктивностью вдоль перуанского побережья в то время. Между 2300 и 1300 гг. до н. э. SST показывали сниженную сезонность с более прохладным летом по сравнению с современными условиями (Carré et al., 2014; Loubere et al., 2013). Кроме того, многодесятилетние колебания апвеллинга и температуры поверхности моря между 1800 и 1500 гг. до н. э. предполагают, что период EF-1 в Вичаме разворачивался в условиях сильно изменчивых океанографических условий (Salvatteci et al., 2016, 2019), потенциально делая морские ресурсы менее предсказуемыми. Интересно, что этот период соответствует большей зависимости от растений C3.
Как утверждалось в предыдущих исследованиях (Caramanica et al., 2020; Contreras, 2010; Dillehay and Kolata, 2004; Lima et al., 2023; Wilson et al., 2024), эти колебания в сочетании с чередованиями ENSO могли вызывать оппортунистическую интенсификацию определенных продуктов (рыболовство/культуры), когда технологический уровень и социополитические условия позволяли это, порождая соответствующие рационы с наблюдаемыми морско-земледельческими колебаниями с течением времени. Теоретически, в условиях, подобных Эль-Ниньо, совпадение снижения морской продуктивности с более влажными условиями (дождями) вдоль побережья и цветением ломас, несмотря на оползни и катастрофы, приводило бы к повышению продуктивности земли в своего рода «земледелии с высоким риском». В сравнении, в условиях, подобных Ла-Нинья, похолодание морских температур, усиление апвеллинга и морской продуктивности, сопутствующее сухому климату (т.е. меньшему количеству осадков и сокращению ломас), поддерживало бы «земледелие с низким риском» и более высокую морскую продукцию, вероятно, с большим количеством морской фауны и большим количеством кукурузы в археологических данных, как мы можем наблюдать в период LIP.
Климатические условия в период EF-1 (1800–1500 гг. до н. э.) в Вичаме также включали относительно влажные периоды в высокогорьях и засушливые условия в прибрежных долинах, отмеченные большей сезонностью речных потоков, с увеличением осадков во влажный сезон и более низким стоком в сухой сезон (Leclerc, 2023; Salvatteci et al., 2019). Развитие ирригационных систем могло позволить интенсифицировать сельское хозяйство во влажный сезон, обеспечивая производство излишков для поддержания населения в более засушливые месяцы. И наоборот, в период LIP похолодание морских температур, усиление апвеллинга и повышение морской продуктивности в сочетании с более сухим климатом, отмеченным уменьшением осадков и сокращением растительности ломас, могли способствовать сдвигу к большей зависимости от эксплуатации морских ресурсов наряду с мелкомасштабным земледелием (Carré et al., 2014; Leclerc, 2023; Loubere et al., 2013; Salvatteci et al., 2019).
Это исследование предлагает новые сведения о том, как изменчивость климата формировала системы жизнеобеспечения в ранних андских обществах, и расширяет наше понимание траектории развития андской цивилизации. В этом смысле наша оценка моделей жизнеобеспечения в период EF-1 — период зарождающейся политической сложности — хорошо согласуется с идеей о том, что снижение морской продуктивности могло способствовать большей зависимости от наземных ресурсов и их интенсификации (Wilson et al., 2024). И наоборот, наши результаты частично подтверждают гипотезу о том, что стабильные климатические условия благоприятствуют развитию аграрных обществ, тогда как климатическая изменчивость способствует политической нестабильности, децентрализации и фрагментации (Kennett and Marwan, 2015). Оппортунизм в эксплуатации доступных ресурсов и устойчивость перед лицом климатических вызовов, по-видимому, были ключевыми моделями поведения этих обществ. Климатические колебания могли представлять собой сложную возможность для разработки адаптивных буферных стратегий, таких как интенсификация земледелия или других видов экономической деятельности. Это может совпадать с гегемонией прибрежных, внутриконтинентальных или высокогорных обществ в разные периоды.
Эти результаты должны быть уточнены и проверены в будущих исследованиях с учетом некоторых ограничений: (1) палеоокеанографические и климатические данные фрагментарны, неоднозначны и подлежат интерпретации и не обязательно могут соответствовать нашим выводам и (2) эквифинальность, то есть различные комбинации пищевых вкладов, которые могут давать одинаковые изотопные значения у потребителя, остается потенциальной проблемой для интерпретаций.
Материалы и методы
Индивиды и образцы
Это мультипрокси-исследование стабильных изотопов, предназначенное для количественной реконструкции состава рациона Вичамы. Наша цель — изучить экономику Вичамы для выяснения роли морских и растительных ресурсов в процессах социальной сложности в период EF.
Всего для анализа было отобрано 59 образцов (36 костей и 23 зуба) от 38 человеческих индивидов, извлеченных из погребальных контекстов Вичамы. Скелетные элементы, отобранные для изотопного анализа, были в основном ребрами и длинными костями, запястьями или черепными костями, когда ребра были недоступны. Зубы отбирались при наличии, а изотопные значения получали из корней, что позволяет проводить прямую сравнительную оценку образцов зубов и костей.
Пол и возраст индивидов определялись с использованием стандартизированных методов (Buikstra and Ubelaker, 1994). Для контроля потенциально искажающих факторов диет грудного вскармливания и отлучения для сравнений индивиды были классифицированы по двум категориям (т.е. подростки и взрослые) в соответствии с тканями (т.е. зубами или костями), из которых получены значения (AlQahtani et al., 2010).
Радиоуглеродные даты
Хронологическая основа этой работы основана на 8 прямых [датах AMS (ускорительная масс-спектрометрия) из коллагена индивидов и тканей из погребальных контекстов] и 12 косвенных радиоуглеродных датах из связанных материалов или стратиграфии (R. Shady, личное сообщение, июнь 2024). Радиоуглеродные даты Вичамы получены из AMS-установок трех различных лабораторий с использованием внутренних протоколов через коммерческие услуги. Даты индивидов были калиброваны (до н. э./н. э.) с помощью пакета Calib 8.1.0 (Stuiver and Reimer, 1993), с использованием смешанной морской калибровочной кривой SoHCal 20 (Heaton et al., 2020; Hogg et al., 2020) и локальной поправки на морской радиоуглеродный резервуар (ΔR = 200 ± 20), оцененной по парным образцам (т.е. костный коллаген и хлопковые ткани) из погребального контекста ZAC7074. Для индивидов с прямым датированием мы включили средний относительный вклад морского углерода в коллаген, рассчитанный с помощью FRUITS для каждой фазы. Калиброванные даты были округлены до 10 (см. дополнительный материал 1: S3). Индивиды, не имеющие прямого датирования, были отнесены к каждому периоду согласно археологическим методам (т.е. стратиграфия, погребальные паттерны, стили керамики и украшения тканей).
Лабораторные процедуры
Сначала образцы были механически очищены для удаления поверхностных загрязнений. Затем образцы были разделены для экстракции коллагена и апатита. Коллаген был экстрагирован в Институте экологических наук и технологий Автономного университета Барселоны (Испания).
Образцы были физически очищены и погружены в 0,6 M HCl при 4°C на несколько дней для деминерализации, затем промыты ультрачистой водой и погружены в 0,05 M гидроксид натрия (NaOH) на 20 минут при комнатной температуре. Циклы промывки NaOH повторялись по мере необходимости, обычно 1 или 2 раза, пока не прекращалось дальнейшее изменение цвета раствора. Образцы промывали трижды ультрачистой водой по 10 минут каждый раз для обеспечения полного удаления NaOH. Образцы желатинизировали в 0,001 M HCl (pH3) при 80°C в течение 48 часов. Супернатант, содержащий коллаген, фильтровали с использованием полиэтиленовых фильтров Ezee (9 мл, размер пор 60–90 мкм, Elkay Laboratories Ltd.), затем замораживали как минимум на 48 часов при −20°C и лиофилизировали. Анализ стабильных изотопов успешно экстрагированных образцов был проведен в SUERC, Ист-Килбрайд (Великобритания) с использованием масс-спектрометра Delta V Advantage с непрерывным потоком, сопряженного через ConfloIV с элементным анализатором IsoLink (Thermo Scientific, Бремен, Германия). Костный коллаген (примерно 0,7 мг) сжигали в присутствии кислорода в едином реакторе, содержащем оксид вольфрама и медную проволоку при 1020°C для получения азота (N2) и диоксида углерода (CO2). Ловушка из перхлората магния использовалась для удаления воды, образующейся в процессе сгорания, а газы разделялись в колонке газового хроматографа (GC), нагреваемой от 70°C до 240°C. Гелий использовался в качестве газа-носителя на протяжении всей процедуры. N2 и CO2 поступали в масс-спектрометр через устройство открытого расщепления внутри ConfloIV и анализировались относительно соответствующих эталонных газов. Эталонные материалы МАГАТЭ USGS40 (L-глутаминовая кислота, δ13C V-PDB = −26,39 ± 0,04‰, δ15N (AIR) = −4,52 ± 0,06‰) и USGS41a (L-глутаминовая кислота, δ13C (V-PDB) = 36,55 ± 0,08‰, δ15N (AIR) = 47,55 ± 0,15‰) использовались для нормализации значений δ15N и δ13C. Нормализация проверялась с использованием хорошо охарактеризованного стандарта рыбного желатина Elemental Microanalysis IRMS B2215 (δ13C (V-PDB) = −22,92 ± 0,10‰, δ15N (AIR) = 4,26 ± 0,12‰) и/или USGS88 (морской коллаген, δ13C (V-PDB) = −16,06 ± 0,07‰, δ15N (AIR) = 14,96 ± 0,14‰). Точность составляет ± 0,2‰ для δ15N и ± 0,1‰ для δ13C.
Для анализа биоапатита костные образцы измельчали в агатовой ступке с пестиком и обрабатывали в Лаборатории экологических изотопов Университета Аризоны (США). В этом случае костные образцы очищали в дистиллированной воде с помощью ультразвуковой ванны и измельчали в порошок с использованием ступки и пестика. Образцы весом ~10 мг каждый выдерживались в течение 24–72 часов в 2% растворе гипохлорита натрия для удаления органического материала. Затем образцы обрабатывали 0,1 M уксусной кислотой для удаления экзогенных карбонатов. Образцы также реагировали с обезвоженной фосфорной кислотой и серебром (серебро удаляет сернистые газы из выделяющегося CO2) при 70°C в системе подготовки проб KIEL-III. Значения δ13C полученного CO2 измерялись с использованием масс-спектрометра с отношением изотопов Finnigan MAT252. Калибровка изотопных отношений была основана на повторных измерениях NBS-19 и NBS-18 с точностью ±0,08‰ для δ13C (1σ).
В анализ были включены только индивиды с валидными изотопными значениями коллагена. Надежность коллагена была подтверждена в соответствии с общепринятыми критериями сохранности: соотношение C/N между 2,9 и 3,6 (DeNiro, 1985) и wt% C 26%−44% и wt% N 11%−16% (Van Klinken, 1999). Мы использовали подвыборку из 10 индивидов для оценки сохранности биоапатита (т.е. минералогии и кристалличности) с помощью ATR-FTIR.
ATR-FTIR анализ был проведен в Лаборатории молекулярной спектроскопии, Servei de Quimica — Автономный университет Барселоны с использованием компактного FTIR-спектрометра Bruker Alpha II, сопряженного с Eco-ATR. Спектры 1 мг костного порошка были собраны в режиме поглощения (A) с спектральным диапазоном от 400 до 4000 см⁻¹, для 100 сканирований и разрешением 8 см⁻¹. Генерация спектров и коррекция базовой линии выполнялись с помощью программного обеспечения OPUS 7.8. Для оценки диагенеза биоапатита мы оценили пять индексов (IRSF — Infrared Splitting Factor; C/P — Carbon/Phosphate; C/C — Carbon/Carbon; BPI — Type B Phosphate index; API — Type A Phosphate index) относительно стандартных значений для хорошо сохранившихся археологических образцов (France et al., 2020). См. дополнительный материал 1: S4 для исходных данных ATR-FTIR и индексов.
Изотопный анализ и модели BSIMM
Для выявления тенденций изотопных значений мы использовали «классические» диаграммы рассеяния изотопных значений (т.е. δ13Cколл против δ15N; δ13Cколл против δ13Cап) и площади стандартных эллипсов (rKin -Standard Ellipse Areas) (Albeke, 2017). После оценки нормального распределения с помощью теста Шапиро–Уилка (α = 0,05), дескриптивная статистика изотопных значений и сравнения по полу (т.е. мужчины, женщины и неопределенные) и широким возрастным категориям (т.е. подростки и взрослые) были выполнены с использованием тестов Крускала–Уоллиса (α = 0,05) и Манна–Уитни (α = 0,05) с помощью SPSS v.29 (Microsoft®). Анализ ядерной плотности и графики выполнялись с помощью функции geom_density() пакета ggplot2 в среде R.
Программное обеспечение BSIMM FRUITS (v. 2.1.1 Beta; Fernandes et al., 2014, 2015) использовалось для оценки пропорционального калорийного вклада различных источников пищи в рацион с использованием трех диетических прокси: δ13Cколл, δ15N и δ13Cап (см. дополнительный материал 2 для реализации модели FRUITS). В этом исследовании мы выполняли индивидуальные оценки. Данные потребителя для индивидов состоят из их изотопных значений δ13Cколл, δ15Nколл и δ13Cап. Для Вичамы как группы мы используем средние значения изотопов и стандартную ошибку среднего.
Мы загрузили нашу модель четырьмя потенциальными источниками пищи (группы продуктов) с соответствующим составом макронутриентов (фракции продуктов: общая, белки и энергия): наземная фауна, обеспечивающая белки и липиды; морская фауна, обеспечивающая белки и липиды; и растения C3 и C4, обеспечивающие углеводы и белки. Изотопные средние значения этих групп продуктов взяты из предыдущих диетических реконструкций для Центрального побережья Анд (Pezo-Lanfranco et al., 2022; см. дополнительный материал 1: S7).
На Центральном побережье Перу растения C3 (включая клубнеплоды, бобовые, однолетние и фрукты) имеют средние значения δ13C и δ15N соответственно −25,7 ± 2,1‰ и +5,3 ± 3,2‰. Растения C4 (ограниченные Zea mays в таких случаях) имеют средние значения δ13C и δ15N соответственно −11,3 ± 1,4‰ и +7,3 ± 3,0‰. Растения CAM имеют средние значения δ13C −10,8 ± 0,2‰, перекрывающиеся с C4, и δ15N +3,8 ± 0,4‰. Наземная фауна (например, олени, грызуны и верблюдовые) показывает средние значения δ13Cколл и δ15N в диапазоне от −17,5 ± 3,2‰ до +9,0 ± 3,1‰. Средние значения δ13C и δ15N для морской фауны (включая рыб, мелких рыб, таких как анчоусы и сардины, моллюсков, морских птиц и млекопитающих современного и археологического происхождения) составляют соответственно −11,9 ± 2,1‰ и +13,2 ± 3,0‰ (Pezo-Lanfranco and Colonese, 2024). Значения δ13C от современных образцов фауны были скорректированы на «эффект ископаемого топлива» путем корректировки значений на +1,5‰ (Marino and McElroy, 1991). Во избежание проблем с сохранностью (Szpak and Chiou, 2019) для питания модели использовались только значения современных растений.
Факторы изотопного фракционирования, полученные из экспериментальных исследований, были установлены на уровне +4,8 ± 0,5‰ между пищей и коллагеном, +10,1 ± 0,5‰ между пищей и апатитом, +5,5 ± 0,5‰ для δ15N от пищи к коллагену (Fernandes et al., 2014, 2015).
Взвешенные значения каждой фракции макронутриентов (липидов, углеводов и белков) следовали опубликованным параметрам (Fernandes et al., 2014, 2015; Pezo-Lanfranco et al., 2022; Pezo-Lanfranco and Colonese, 2024). Значения δ13Cап представляют общую углеродную смесь в рационе. Поэтому мы используем одно и то же общее значение для каждой группы продуктов. Общие значения δ13C для наземной и морской фауны были оценены как средневзвешенные значения δ13C липидов и белков (Fernandes et al., 2014, 2015). Для коллагена доля углерода белка и энергии, поступающая в общий коллаген, была установлена на уровне 74 ± 4% и 26% соответственно (Fernandes et al., 2012). Мы предположили, что азот поступает исключительно из белков (100%). Липиды и углеводы были добавлены в модель как «энергия». Для учета этого комбинаторного эффекта мы применили модель, зависимую от концентрации (Fernandes et al., 2014, 2015).
Следуя предыдущим реконструкциям с использованием FRUITS (Fernandes et al., 2014, 2015; Pezo-Lanfranco et al., 2022), изотопный состав каждой питательной фракции (белки, углеводы и липиды) был получен из средних значений δ13Cколл и δ15Nколл с использованием следующих факторов фракционирования: −2‰ (Δ13Cбелок-коллаген), −8‰ (Δ13Cлипиды-коллаген) и +2‰ (Δ15Nбелок-коллаген) для наземных млекопитающих и −1‰ (Δ13Cбелок-коллаген), −7‰ (Δ13Cлипиды-коллаген) и +2‰ (Δ15Nбелок-коллаген) для морских животных. Для растений смещения составили −2‰ (Δ13Cобщий-белок) и +0,5‰ (Δ13Cобщий-липиды), а для значения δ15N растительного белка было принято известное значение δ15N, зафиксированное для растения. Углеродный вес (концентрации) каждой пищевой фракции (белки и энергия) из каждой группы продуктов рассчитывался в соответствии с ее макронутриентным составом (Fernandes et al., 2014, 2015; Pezo-Lanfranco et al., 2022; см. подробности в дополнительном материале 2). Физиологический, консервативный и приемлемый диапазон потребления белка от 5% до 45% от общего количества калорий был загружен как «априорный» (Fernandes et al., 2014). Оценки моделей FRUITS отражают содержание углерода или эквивалентный калорийный вклад, выраженный как относительный вклад (в сумме 1 или 100%) с соответствующей неопределенностью 1σ (Fernandes et al., 2015).
Наконец, для оценки статистического различия между распределениями вероятностей четырех групп продуктов между периодами (EF-1 против LIP) был проведен тест дивергенции Кульбака–Лейблера (Lyman, 2008). Этот тест был выполнен с использованием пакета Entropy в среде R (дополнительный материал 2).
Дополнительный материал
Дополнительный материал к этой статье можно найти онлайн по адресу: https://www.frontiersin.org/articles/10.3389/fearc.2025.1611071/full#supplementary-material
Ссылки
1. Aguilar M. A. (2006). “Surgimiento de las sociedades complejas en los andes centrales,” in Una perspectiva desde el valle de Huaura, Perú. Licentiate Monography (Lima: Universidad Nacional Federico Villarreal). CrossRef. Google Scholar.
2. Albeke S. E. (2017). “rKIN: (kernel) isotope niche estimation,” in R Package Version 0.1. CrossRef. Google Scholar.
3. AlQahtani S. J., Hector M. P., Liversidge H. M. (2010). Brief communication: the London atlas of human tooth development and eruption. Am. J. Phys. Anthropol. 142, 481–490. doi: 10.1002/ajpa.21258. CrossRef. Google Scholar.
4. Ambrose S. H., Norr L. (1993). “Experimental evidence for the relationship of carbon isotope ratios of whole diet and dietary protein to those of bone collagen and carbonate,” in Prehistoric Human Bone: Archaeology at the Molecular Level (Berlin: Springer), 1–37. CrossRef. Google Scholar.
5. Buikstra J., Ubelaker D. (1994). Standards for Data Collection from Human Skeletal Remains. Fayetteville: Arkansas Archeological Survey Research Series, No.44. CrossRef. Google Scholar.
6. Cadwallader L., Beresford-Jones D. G., Whaley O., O’Connell T. (2012). The sign of maize? A reconsideration of what 13C values say about Paleodiet in the Andean Region. Hum. Ecol. 40, 487–509. doi: 10.1007/s10745-012-9509-0. CrossRef. Google Scholar.
7. Caramanica A., Huaman Mesia L., Morales C. R., Huckleberry G., Castillo L. J., Quilter J. (2020). El Niño resilience farming on the north coast of Peru. Proc. Natl. Acad. Sci. U.S.A. 117:24127. doi: 10.1073/pnas.2006519117. CrossRef. Google Scholar.
8. Cárdenas M. (1977). Informe Preliminar del Trabajo de Campo en el Valle de Huaura (Departamento de Lima). Lima: Instituto Riva Agüero, Seminario de Arqueología, PUCP. CrossRef. Google Scholar.
9. Carré M., Sachs J. P., Purca S., Schauer A. J., Braconnot P., Angeles-Falcón R., et al. (2014). Holocene history of ENSO variance and asymmetry in the eastern tropical Pacific. Science 345, 1045–1048. doi: 10.1126/science.1252220. CrossRef. Google Scholar.
10. Cheung C., Szpak P. (2021). Interpreting past human diets using stable isotope mixing models. J. Archaeol. Method Theory 28, 1106–1142. doi: 10.1007/s10816-020-09492-5. CrossRef. Google Scholar.
11. Chisholm B. S., Nelson D. E., Schwarcz H. P. (1982). Stable-carbon isotope ratios as a measure of marine versus terrestrial protein in ancient diets. Science 216, 1131–1132. doi: 10.1126/science.216.4550.1131. CrossRef. Google Scholar.
12. Chu A. (2011). Household organization and social inequality at Bandurria, a Late Preceramic village in Huaura, Peru (PhD. Dissertation). University of Pittsburgh, Pittsburgh, PA, United States. CrossRef. Google Scholar.
13. Contreras D. A. (2010). Landscape and environment: Insights from the Prehispanic Central Andes. J. Arch. Res. 18, 241–288. doi: 10.1007/s10814-010-9038-6. CrossRef. Google Scholar.
14. Coutts K., Chu A., Krigbaum J. (2011). Paleodiet in Late Preceramic Peru: Preliminary Isotopic data from Bandurria. J. Island Coast. Archeol. 6, 196–210. doi: 10.1080/15564894.2011.586086. CrossRef. Google Scholar.
15. Creamer W., Ruiz A., Perales M., Haas J. (2013). The Fortaleza Valley, Peru: Archaeological investigation of late archaic sites (3000–1800 BC). Field. Anthropol. 44, 1–108. doi: 10.3158/0071-4739-44.0.1. CrossRef. Google Scholar.
16. DeNiro M. J. (1985). Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to paleodietary reconstruction. Nature 317, 806–809. doi: 10.1038/317806a0. CrossRef. Google Scholar.
17. DeNiro M. J., Epstein S. (1981). Influence of diet on the distribution of nitrogen isotopes in animals. Geochim. Cosmochim. Acta 45, 341–351. doi: 10.1016/0016-7037(81)90244-1. CrossRef. Google Scholar.
18. Dillehay T. D. (2017). Where the Land Meets the Sea: Fourteen Millennia of Human History at Huaca Prieta, Peru. Austin: University of Texas Press. CrossRef. Google Scholar.
19. Dillehay T. D., Elling H., Rossen J. (2005). Preceramic irrigation canals in the Peruvian Andes. Proc. Natl. Acad. Sci. U.S.A. 102, 17241–17244. doi: 10.1073/pnas.0508583102. CrossRef. Google Scholar.
20. Dillehay T. D., Kolata A. L. (2004). Long-term human response to uncertain environmental conditions in the Andes. Proc. Natl. Acad. Sci. U.S.A. 101, 4325–4330. doi: 10.1073/pnas.0400538101. CrossRef. Google Scholar.
21. Dillehay T. D., Rossen J., Andres T. C., Williams D. E. (2007). Preceramic adoption of peanut, squash, and cotton in northern Peru. Science 316, 1890–1893. doi: 10.1126/science.1141395. CrossRef. Google Scholar.
22. Engel F. A. (1987). “De las Begonias al Maíz,” in Vida y Producción en el Perú Antiguo (Lima: Centro de Investigaciones de Zonas Áridas CIZA, Universidad Nacional Agraria La Molina). CrossRef. Google Scholar.
23. Fernandes R., Grootes P. M., Nadeau M. J., Nehlich O. (2015). Quantitative diet reconstruction of a neolithic population using a Bayesian mixing model (FRUITS): The case study of Ostorf (Germany). Am. J. Phys. Anthropol. 158, 325–340. doi: 10.1002/ajpa.22788. CrossRef. Google Scholar.
24. Fernandes R., Millard A. R., Brabec M., Nadeau M. J., Grootes P. M. (2014). Food reconstruction using isotopic transferred signals (FRUITS): a Bayesian model for diet reconstruction. PLoS ONE 9:e87436. doi: 10.1371/journal.pone.0087436. CrossRef. Google Scholar.
25. Fernandes R., Nadeau M. J., Grootes P. M. (2012). Macronutrient-based model for dietary carbon routing in bone collagen and bio-apatite. Archaeol. Anthropol. Sci. 4, 291–301. doi: 10.1007/s12520-012-0102-7. CrossRef. Google Scholar.
26. France C. A. M., Sugiyama N., Aguayo E. (2020). Establishing a preservation index for bone, dentin, and enamel bioapatite mineral using ATR-FTIR. J. Arch. Sci. Rep. 33, 102551. doi: 10.1016/j.jasrep.2020.102551. CrossRef. Google Scholar.
27. Goldberg A., Mychajliw A. M., Hadly E. A. (2016). Post-invasion demography of prehistoric humans in South America. Nature 532, 232–235. doi: 10.1038/nature17176. CrossRef. Google Scholar.
28. Grobman A., Bonavia D., Dillehay T. D., Piperno D. R., Iriarte J., Holst I. (2012). Preceramic maize from Paredones and Huaca Prieta Peru. Proc. Natl. Acad. Sci. U. S. A. 109, 1755–1759. doi: 10.1073/pnas.1120270109. CrossRef. Google Scholar.
29. Haas J., Creamer W. (2006). Crucible of Andean Civilization: The Peruvian coast from 3000 to 1800 BC. Curr. Anthropol. 47, 745–775. doi: 10.1086/506281. CrossRef. Google Scholar.
30. Haas J., Creamer W., Huamán L., Goldstein D., Reinhard K., Vergel C. (2013). Evidence for maize (Zea mays) in the Late Archaic (3000–1800 B.C.) in the Norte Chico region of Peru. Proc. Natl. Acad. Sci. U.S.A. 110, 4945–4949. doi: 10.1073/pnas.1219425110. CrossRef. Google Scholar.
31. Heaton T., Köhler P., Butzin M., Bard E., Reimer R., Austin W., et al. (2020). Marine20 - the marine radiocarbon age calibration curve (0–55,000 cal BP). Radiocarbon 62, 779–820. doi: 10.1017/RDC.2020.68. CrossRef. Google Scholar.
32. Hedges R. E. M., Reynard L. M. (2007). Nitrogen isotopes and the trophic level of humans in archaeology. J. Archaeol. Sci. 34, 1240–1251. doi: 10.1016/j.jas.2006.10.015. CrossRef. Google Scholar.
33. Herbert T. (2003). “Alkenone paleotemperature determinations,” in Treatise on Geochemistry: The Oceans and Marine Geochemistry, ed. H. Elderfeld (New York: Elsevier), 391–432. CrossRef. Google Scholar.
34. Hogg A., Heaton T., Hua Q., Palmer J., Turney C., Southon J., et al. (2020). SHCal20 southern hemisphere calibration, 0–55,000 years cal BP. Radiocarbon 62, 759–778. doi: 10.1017/RDC.2020.59. CrossRef. Google Scholar.
35. IMARPE (2010). Informe: Delimitación y caracterización de bancos naturales de invertebrados bentónicos comerciales y áreas de pesca artesanal en la región Lima entre Punta Litera-Playa Grande, Barranca y Huaura de la Region Lima. Lima: Instituto del Mar del Perú. CrossRef. Google Scholar.
36. INRENA (2005). Inventario y monitoreo de las aguas subterráneas en el valle Huaura: Anexos (Lima: Autoridad Nacional del Agua). CrossRef. Google Scholar.
37. Jim S., Ambrose S., Evershed R. P. (2004). Stable carbon isotopic evidence for differences in the dietary origin of bone cholesterol, collagen, and apatite: implications for their use in paleodietary reconstruction. Geochim. Cosmochim. Acta 68, 61–72. doi: 10.1016/S0016-7037(03)00216-3. CrossRef. Google Scholar.
38. Kellner C. M., Schoeninger M. J. (2007). A simple carbon isotope model for reconstructing human diet. Am. J. Phys. Anthropol. 133, 1112–1127. doi: 10.1002/ajpa.20618. CrossRef. Google Scholar.
39. Kennett D. J., Marwan N. (2015). Climatic volatility, agricultural uncertainty, and the formation, consolidation and breakdown of preindustrial agrarian states. Phylos. T. R. Soc. A 373:20140458. doi: 10.1098/rsta.2014.0458. CrossRef. Google Scholar.
40. Leclerc E. L. (2023). Coast and highland paleoclimate of the north-central Peruvian Andes and its influence on coastal water availability and cultural development, 6000 to 2000 cal BP. Quat. Sci. Rev. 314:108209. doi: 10.1016/j.quascirev.2023.108209. CrossRef. Google Scholar.
41. León E. (2013). 14,000 años de alimentación en el Perú. Lima: Universidad de San Martin de Porres. CrossRef. Google Scholar.
42. Lima M., Gayó E. M., Gurruchaga A., Estay S. A., Santoro C. M. (2023). 1000 years of population, warfare, and climate change in pre-Columbian societies of the Central Andes. PLoS ONE 18:e0278730. doi: 10.1371/journal.pone.0278730. CrossRef. Google Scholar.
43. Loubere P., Creamer W., Haas J. (2013). Evolution of the El Nino-Southern Oscillation in the late Holocene and insolation driven change in the tropical annual SST cycle. Glob. Planet. Change 100:129e144. doi: 10.1016/j.gloplacha.2012.10.007. CrossRef. Google Scholar.
44. Lyman R. L. (2008). Quantitative Paleozoology. Cambridge: Cambridge University Press. CrossRef. Google Scholar.
45. Mächtle B., Eitel B. (2013). Fragile landscapes, fragile civilizations - how climate determined societies in the pre-Columbian south Peruvian Andes. Catena 103, 62–73. doi: 10.1016/j.catena.2012.01.012. CrossRef. Google Scholar.
46. Makowski K. (2023). Pre-Hispanic andean urbanism and its 'anti-urban' peculiarities. J. Urban Archaeol. 8, 165–196. doi: 10.1484/J.JUA.5.135664. CrossRef. Google Scholar.
47. Marino B. D., McElroy M. B. (1991). Isotopic composition of atmospheric CO2 inferred from carbon in C4 plant cellulose. Nature 349, 127–131. doi: 10.1038/349127a0. CrossRef. Google Scholar.
48. Minagawa M., Wada E. (1984). Stepwise enrichment of 15N along food chains: further evidence and the relation between δ15N and animal age. Geochim. Cosmochim. Acta 48, 1135–1140. doi: 10.1016/0016-7037(84)90204-7. CrossRef. Google Scholar.
49. Moseley M. E. (1975). The Maritime Foundations of Andean Civilization. Menlo Park, CA: Cummings Publishing Company. CrossRef. Google Scholar.
50. Orloff C. R. (2022). Caral, South America's Oldest City (2600–1600 BC): ENSO environmental changes influencing the late archaic period site on the North Central Coast of Peru. Water 14:1403. doi: 10.3390/w14091403. CrossRef. Google Scholar.
51. Pezo-Lanfranco L., Colonese A. C. (2024). The role of farming and fishing in the rise of social complexity in the Central Andes: a stable isotope perspective. Sci. Rep. 14:4582. doi: 10.1038/s41598-024-55436-4. CrossRef. Google Scholar.
52. Pezo-Lanfranco L., Crispin A., Machacuay M., Novoa P., Shady R. (2020). Isotopic evidence of weaning behaviors from farming communities of the Peruvian North-Central Coast. Insights into the demography of the transition from the middle to the late formative periods (500-400 B.C.). Bioarchaeol. Int. 3–4, 191–216. doi: 10.5744/bi.2020.3019. CrossRef. Google Scholar.
53. Pezo-Lanfranco L., Machacuay M., Novoa P., Peralta R., Mayer E., Eggers S., et al. (2022). The diet at the onset of the Andean Civilization: New stable isotope data from Caral and Áspero, North-Central Coast of Peru. Am. J. Biol. Anthropol. 177, 402–424. doi: 10.1002/ajpa.24445. CrossRef. Google Scholar.
54. Pezo-Lanfranco L., Pezo-Lanfranco S., Eggers S. (2009). Exostosis auditiva como marcador osteológico de actividad acuática en poblaciones formativas de la Costa Norte del Perú. Paleopatología 6, 1–18. CrossRef. Google Scholar.
55. Phillips D. L. (2012). Converting isotope values to diet composition: the use of mixing models. J. Mammal. 93, 342–352. doi: 10.1644/11-MAMM-S-158.1. CrossRef. Google Scholar.
56. Piperno D. R., Dillehay T. D. (2008). Starch grains on human teeth reveal early broad crop diet in northern Peru. Proc. Natl. Acad. Sci. U.S.A. 105, 19622–19627. doi: 10.1073/pnas.0808752105. CrossRef. Google Scholar.
57. Salvatteci R., Gutiérrez D., Field D., Sifeddine A., Ortlieb L., Bouloubassi I., et al. (2014). The response of the Peruvian Upwelling Ecosystem to centennial-scale global change during the last two millennia. Clim. Past 10, 715–731. doi: 10.5194/cp-10-715-2014. CrossRef. Google Scholar.
58. Salvatteci R., Gutierrez D., Sifeddine A., Ortlieb L., Druffel E., Boussafir M., et al. (2016). Centennial to millennial-scale changes in oxygenation and productivity in the eastern tropical South Pacific during the last 25000 years. Quat. Sci. Rev. 131, 102–117. doi: 10.1016/j.quascirev.2015.10.044. CrossRef. Google Scholar.
59. Salvatteci R., Schneider R. R., Blanz T., Mollier-Vogel E. (2019). Deglacial to Holocene ocean temperatures in the humboldt current system as indicated by alkenone paleothermometry. Geophys. Res. Lett. 46, 281–292. doi: 10.1029/2018GL080634. CrossRef. Google Scholar.
60. Sandweiss D. H., Shady R., Moseley M., Keefer D. K., Orloff C. R. (2009). Environmental change and economic development in coastal Peru between 5,800 and 3,600 years ago. Proc. Natl. Acad. Sci. USA 106, 1359–1363. doi: 10.1073/pnas.0812645106. CrossRef. Google Scholar.
61. Schoeninger M. J. (2010). Diet reconstruction and ecology using stable isotope ratios, in A companion to Biological Anthropology (Hoboken, NJ: Willey-Blackwell), 445–464. CrossRef. Google Scholar.
62. Schoeninger M. J., DeNiro M. J. (1984). Nitrogen and carbon isotopic composition of bone collagen from marine and terrestrial animals. Geochim. Cosmochim. Acta 48, 625–639. doi: 10.1016/0016-7037(84)90091-7. CrossRef. Google Scholar.
63. Schoeninger M. J., DeNiro M. J., Tauber H. (1983). Stable nitrogen isotope ratios of bone collagen reflect marine and terrestrial components of prehistoric human diet. Science 220, 1381–1383. doi: 10.1126/science.6344217. CrossRef. Google Scholar.
64. Schoeninger M. J., Reitsema L. J. (2023). “Diet reconstruction and ecology,” in A Companion to Biological Anthropology, ed. C. S. Larsen (Malden: Willey-Blackwell), 510–526. CrossRef. Google Scholar.
65. Scholz F., McManus J., Mix A., Hensen C., Schneider R. (2014). The impact of ocean deoxygenation on the ocean's iron supply. Nat. Geosci. 7, 433–437. doi: 10.1038/ngeo2162. CrossRef. Google Scholar.
66. Shady R. (2006a). La civilización Caral: sistema social y manejo del territorio y sus recursos. Su trascendencia en el proceso cultural andino. Bol. Arqueol. PUCP 10, 59–89. doi: 10.18800/boletindearqueologiapucp.200601.004. CrossRef. Google Scholar.
67. Shady R. (2006b). “America's first city? The case of Late Archaic Caral,” in Andean Archaeology III, eds. D. W. Isbell, and H. Silverman (Boston: Springer), 28–66. CrossRef. Google Scholar.
68. Shady R. (2006c). “Caral-Supe and the North- Central area of Peru: The history of maize in the land where Civilization came into being,” in Histories of Maize: Multidisciplinary Approaches to the Prehistory, Linguistics, Biogeography, Domestication, and Evolution of Maize, eds. J. Staller, R. Tykot, and B. Benz (Cambridge, MA: Academic Press), 381–402. CrossRef. Google Scholar.
69. Shady R. (2018). El sistema social de Caral y su trascendencia: El manejo transversal del territorio, la complementariedad social y política y la interacción intercultural (Lima: Nayra Kunan Pacha, Revista de Arqueología Social, Proyecto Especial Arqueológico Caral-Supe), 19–90. CrossRef. Google Scholar.
70. Shady R., Cáceda D. (2008). Áspero, la ciudad pesquera de la civilización Caral - Supe, recuperamos su historia para vincularla con nuestro presente Proyecto Especial Arqueológico Caral – Supe. Lima: Instituto Nacional de Cultura. CrossRef. Google Scholar.
71. Shady R., Crispín A., Quispe E. (2008). “Vichama de Végueta comparte con Caral 5000 años de Identidad Cultural,” in Proyecto Especial Arqueológico Caral-Supe (Lima: Instituto Nacional de Cultura). CrossRef. Google Scholar.
72. Shady R., Haas J., Creamer W. (2001). Dating Caral, a Preceramic urban center in the Supe Valley on the central coast of Peru. Science 292, 723–726. doi: 10.1126/science.1059519. CrossRef. Google Scholar.
73. Shady R., Machacuay M., Quispe E., Novoa P., Leyva C. (2015). Vichama, historia social de la Civilización en Végueta. 3800 años de memoria colectiva de nuestros ancestros de Vichama ante el cambio climático. Lima: Biblioteca Nacional del Perú. CrossRef. Google Scholar.
74. Shoji K., Vásquez V. F., Rosales T. E. (2023). Starch grains on human teeth as evidence for 4000 BCE potato consumption at the Cruz Verde site, northern coast of Peru. J. Archaeol. Sci. Rep. 51:104152. doi: 10.1016/j.jasrep.2023.104152. CrossRef. Google Scholar.
75. Stuiver M., Reimer P. J. (1993). CALIB rev. 8. Radiocarbon 35, 215–230. doi: 10.1017/S0033822200013904. CrossRef. Google Scholar.
76. Szpak P., Chiou K. L. (2019). A comparison of nitrogen isotope compositions of charred and desiccated botanical remains from northern Peru. Veg. Hist. Archaeobot. 29, 527–538. doi: 10.1007/s00334-019-00761-2. CrossRef. Google Scholar.
77. Tello J. C. (2004). La Muralla de Santa. Arqueología y Sociedad 15, 11–32. doi: 10.15381/arqueolsoc.2004n15.e12731. CrossRef. Google Scholar.
78. Tung T. A., Dillehay T. D., Feranec R., DeSantis L. R. G. (2021). Early specialized maritime and maize economies on the north coast of Peru. Proc. Natl. Acad. Sci. U. S. A. 117, 32308–32319. doi: 10.1073/pnas.2009121117. CrossRef. Google Scholar.
79. Van Klinken G. J. (1999). Bone collagen quality indicators for palaeodietary and radiocarbon measurements. J. Arch. Sci. 26, 687–695. doi: 10.1006/jasc.1998.0385. CrossRef. Google Scholar.
80. Vega-Centeno R. (2005). Ritual and Architecture in a Context of Emergent Complexity: A Perspective from Cerro Lampay, a Late Archaic Site in the Central Andes (PhD. Dissertation). The University of Arizona, Tucson, AZ, United States. CrossRef. Google Scholar.
81. Wefer G., Fischer G., Iversen M. (2015). “Export production,” in Encyclopedia of Marine Geosciences, eds. J. Harff, M. Meschede, S. Petersen, and J. Thiede (Dordrecht: Springer). CrossRef. Google Scholar.
82. Wilson K. M., McCool W. C., Brewer S. C., Zamora-Wilson N., Schryver P. J., Lamson R. L. F., et al. (2022). Climate and demography drive 7000 years of dietary change in the Central Andes. Sci. Rep. 12:2026. doi: 10.1038/s41598-022-05774-y. CrossRef. Google Scholar.
83. Wilson K. M., McCool W. C., Coltrain J. B. (2024). Climate and oceanic condition changes influence subsistence economic adaptation through intensification on the Central Andean coasts. Quat. Int. 689, 79–92. doi: 10.1016/j.quaint.2023.07.002. CrossRef. Google Scholar.
84. Yseki M., Crispín A., Abad T., Prado A., Novoa P., Shady R. (2024). First comprehensive macroremains analysis of edible plants in Vichama site, Peru during the Early Formative Period 1 (1800-1500 BCE). Veg. Hist. Archaeobot. 34, 91–100. doi: 10.1007/s00334-024-01004-9. CrossRef. Google Scholar.
85. Yseki M., Pezo-Lanfranco L., Machacuay M., Novoa P., Shady R. (2023). Analysis of starch grains trapped in human dental calculus in Áspero, Peru during the Initial Formative Period (3000–1800 BCE). Sci. Rep. 13:14143. doi: 10.1038/s41598-023-41015-6. CrossRef. Google Scholar.
86. Yseki M., Turcq B., Caquineau S., Salvatteci R., Solis J., Skilbeck C. G., et al. (2022). Millennial variability of terrigenous transport to the central–southern Peruvian margin during the last deglaciation (18–13 kyr BP). Clim. Past 18, 2255–2269. doi: 10.5194/cp-18-2255-2022. CrossRef. Google Scholar.
87. Zechenter E. (1988). Subsistence strategies in the Supe Valley of the Peruvian Central Coast During the Complex Preceramic and Initial Periods (PhD. Dissertation). University of California, Los Angeles, Los Angeles, CA, United States. CrossRef. Google Scholar.

Pezo-Lanfranco L, Crispin A, Prado-Barragán A, Abad T, Machacuay M, Yseki M, Gorriti M, Miranda L, Apolín J, DiMuro A, Novoa P, Colonese AC and Shady R (2025) Refining dietary shifts linked to climate oscillations in the Central Andes: stable isotope evidence from Vichama (1800–1500 BCE). Front. Environ. Archaeol. 4:1611071. doi: 10.3389/fearc.2025.1611071
Перевод статьи «Refining dietary shifts linked to climate oscillations in the Central Andes: stable isotope evidence from Vichama (1800–1500BCE)» авторов Pezo-Lanfranco L, Crispin A, Prado-Barragán A, Abad T, Machacuay M, Yseki M, Gorriti M, Miranda L, Apolín J, DiMuro A, Novoa P, Colonese AC and Shady R., оригинал доступен по ссылке. Лицензия: CC BY. Изменения: переведено на русский язык
Фото: wikipedia


















































































































Комментарии (0)